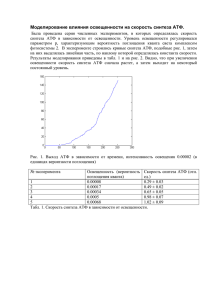
Белки – это высокомолекулярные азотсодержащие органические соединения, состоящие из остатков α-аминокислот, соединенных пептидной связью. В состав белков входит 20 аминокислот. O H2N CH C OH R HN COOH Пролин При построении белковой молекулы, аминокислоты объединяются друг с другом в длинные полипептидные цепи, образуя друг с другом пептидные связи. Пептидная связь настолько прочная, что для её гидролиза вне организма необходимы очень жесткие условия: нагревание при 105°C в течение суток в присутствии концентрированной HCl. В условиях организма существуют специфические ферменты – пептидазы, с помощью которых эти связи разрываются очень быстро. Основу белка составляет первичная структура. Первичная структура белка – это последовательность (порядок чередования) остатков α-аминокислот в полипептидной цепи. Разные белки имеют разную первичную структуру. Эта структура абсолютно уникальная для каждого белка! Цепь повторяющихся групп: называется: пептидный остов. –NH–CH–CO– Цепь повторяющихся групп: называется: пептидный остов. –NH–CH–CO– Формула пептида, содержащего пролин: O H2N CH C N O H C N CH COOH HC (CH2)4 H3C CH3 NH2 Лиз Про Вал Информация о первичной структуре белка лежит в ДНК: ДНК (гены) ↓ мРНК ↓ первичная стр-ра белка В условиях клетки первичная структура приобретает 3-х-мерную пространственную структуру, которая называется: конформация. В понятие конформация входит: вторичная и третичная структура белка. Вторичная структура белка – это пространственная структура, в формировании которой участвуют водородные связи между CO– и NH– группами пептидного остова. C O ... H : N 3 типа вторичной структуры: 1) α-спираль; 2) β-структура; 3) беспорядочный клубок. 1) α-спираль образуется за счет закручивания пептидного остова белка в спираль. В формировании α-спирали участвуют водородные связи между кислородом CO– группы одной аминокислоты и водородом NH– группы 4-ой от неё аминокислоты. Водородные связи ориентированы вдоль оси такой спирали: Водородные связи Радикалы аминокислот Пептидный остов На один виток спирали приходится 3,6 остатка аминокислот. Радикалы аминокислот располагаются снаружи спирали. Формирование α-спирали может быть нарушено: 1) В участках, содержащих пролин. Пролин не содержит атома H, способного образовывать водородную связь. В этом месте полипептидной цепи возникает изгиб или петля; 2) если последовательно расположены несколько одноименно заряженных радикалов, между которыми возникает электростатическое отталкивание; 3) участки с близкорасположенными объемными радикалами (триптофан, метионин, гистидин). 2) β-структура образуется между линейными областями одной полипептидной цепи или между разными полипептидными цепями: Параллельная β-структура: Антипараллельная β-структура: 3) Беспорядочный клубок – это области с нерегулярной вторичной структурой. Это петлеобразные и кольцеобразные стр-ры, с меньшей регулярностью укладки, чем α-спираль и βстр-ра. Они сильно отличаются в разных белках, но имеют одинаковую (или очень близкую) стр-ру для всех молекул одного и того же белка. Третичная структура белка – это трехмерная пространственная структура, в формировании которой участвуют межрадикальные взаимодействия. Свойства радикалов аминокислот по их полярности: I Гидрофильные (полярные) Незаряженные Анионные Катионные (–) (+) Радикалы, Радикалы, Радикалы, содержащие содержащие содержащие одну атом азота, COOH– из следующих группу: способный к полярных присоединению групп: –COOH → протона: COO– + H+ Пример: OH SH Сер; Тре; Тир; Асп; Глу Цис; Асн; Глн II Гидрофобные (неполярные) Радикалы не содержат ни одной полярной группы: NH2 + H+ → + NH3 Ала; Вал; Лиз; Арг; Гис Лей; Иле; Мет; Фен; Три; Про Гидрофильные и гидрофобные радикалы никогда не взаимодействуют друг с другом!! I. Гидрофильные (полярные) радикалы могут взаимодействовать только между собой, образуя 2 типа связей: а) водородная связь; б) ионная связь. а) Водородная связь образуется между 2-мя гидрофильными незаряженными радикалами или одним заряженным (чаще анионным) и одним незаряженным радикалом: ... H (Сер, тре, тир) O HO C NH2 (Сер, тре, тир) (Глн, Асн) - O HO HO ... O (Сер, тре, тир) ... ... HS OOC (Сер, тре, (Глу, Асп) (Цис) H (Сер, тре, тир) тир) б) Ионная связь образуется между разноименно заряженными радикалами: – Глу Асп и + Лиз Арг Гис Например: CH2 (Асп) COO + и H 3N (CH2)4 (Лиз) II. Гидрофобные (неполярные) радикалы могут взаимодействовать только между собой, образуя гидрофобную связь. Пример: CH3 CH2 Фен Ала Это всё были – слабые связи! Между радикалами цистеина может образовываться прочная ковалентная связь – дисульфидная: CH2 SH CH2 S HS Цис H 2C Цис S H 2C Цистин Дисульфидные связи имеют далеко не все белки. Такие связи имеют в основном секреторные белки, которые выделяются во внеклеточное пространство. К ним относятся, иммуноглобулины. например, инсулин и В водной среде третичная структура образовываться с гидрофобных связей. начинает Гидрофобные радикалы, «прячась» от воды, погружаются внутрь белковой молекулы, максимально сближаясь друг с другом и образуя между собой гидрофобные связи. Они составляют гидрофобное ядро белка. Гидрофильные радикалы стремятся располагаться снаружи, образуя с водой водородные связи, и составляют гидрофильную оболочку белка. За счет гидрофильной оболочки крупная молекула белка хорошо растворима в воде. Те гидрофильные радикалы, которые оказались внутри белковой молекулы, образуют друг с другом водородные и ионные связи. В белках, которые функционируют в гидрофобном окружении (белки мембран), обратное строение: внутри – гидрофильные радикалы, а снаружи – гидрофобные. Супервторичная структура белка Супервторичная структура белка – это сходные сочетания и взаиморасположение вторичных структур в белке. Это специфическое сочетание элементов вторичной структуры при формировании третичной стр-ры белка. В формировании супервторичной стр-ры могут принимать участие межрадикальные взаимодействия. Примеры: "α-спираль – поворот – α-спираль"; "лейциновая застежка молния", "цинковые пальцы", "β-бочонок" и др. Все связи, стабилизирующие вторичную и третичную структуру (кроме дисульфидных) – слабые. Они могут разрываться даже при комнатной t°. Поэтому, для белков характерно уникальное свойство: конформационная лабильность. Конформационная лабильность – это способность белков к небольшим изменениям конформации в результате разрыва одних и образования других слабых связей. Эта способность помогает белкам функционировать. Функции белков: 1. Структурная: белки соединительной ткани, фибрин тромбов; 2. Транспортная: гемоглобин – транспорт O2, трансферрин – транспорт Fe; 3. Ферментативная: белки-ферменты, катализирующие химич. реакции в организме; 4. Сократительная: актин, миозин; 5. Регуляторная: белковые гормоны, активаторы и ингибиторы других белков; 6. Защитная: белки-иммуноглобулины 7. Энергетическая Выполнение белком его функции обеспечивается специфическим участком, к которому присоединяется лиганд. Этот участок называется: активный центр. Лиганд – это вещество, которое взаимодействует с активным центром комплементарности. белка по принципу 1. Активный центр белка – это участок молекулы белка, сформированный на уровне третичной структуры (иногда, четвертичной), ответственный за связывание с лигандом. 2. Расположен в узком гидрофобном углублении (щели) поверхности молекулы белка. Комплементарность – это пространственное и химическое соответствие взаимодействующих поверхностей (активного центра белка и лиганда). ДНК (гены) ↓ мРНК ↓ первичная стр-ра белка ↓ конформация активный центр L →↓ функция Простые и сложные белки Белки Простые Имеют только белковую часть 1. Альбумины 2. Глобулины 3. Гистоны 4. Инсулин Сложные (Холопротеины) Имеют белковую и небелковую часть 1. Липопротеины 2. Гемопротеины 3. Фосфопротеины 4. Гликопротеины 5. Нуклеопротеины (липиды) (гем) (H3PO4) (углевод) (нукл. к-та) Небелковый компонент Олигомерные белки регуляции функций. и их способность к Белки Состоящие из одной полипептидной цепи Состоящие из нескольких полипептидных цепей Мономерные белки Олигомерные белки Протомер (субъединица) – это часть олигомерного белка, состоящая из одной полипептидной цепи. Причем, каждая полипептидная цепь такого белка имеет собственную первичную, вторичную и третичную структуру и приобретает собственную конформацию. Олигомерные белки – это белки, состоящие из нескольких протомеров. Олигомерные белки могут содержать от 2-х до нескольких тысяч субъединиц. Примеры: гемоглобин (4 субъединицы); аспартаттранскарбомоилаза (12 субъединиц); белок вируса табачной мозаики (2120 субъединиц). Четвертичная структура белка – это количество, способ расположения и пространственной укладки протомеров в олигомерном белке. Участки, с помощью которых протомеры объединяются между собой при сборке олигомерного белка, называются: контактные поверхности. Протомеры объединяются друг с другом по принципу комплементарности их контактных поверхностей. Связи между протомерами: водородные, ионные и гидрофобные. Способность олигомерных белков к регуляции функций Два родственных белка – гемопротеина: Миоглобин (Mb) Гемоглобин (Hb) Mb – белок красных мышц. Запасает O2 и отдает его только при очень интенсивной работе, т.к. имеет очень высокое сродство к O2. Hb – содержится в эритроцитах. Транспортирует O2 из легких в ткани. (Из тканей в легкие переносит CO2 и H+) Количество O2, отдаваемое в ткани, может регулироваться. I стр-ра: Одна полипептид. цепь – 153 аминокислоты. I стр-ра: α-цепь – 141 аминок-та β-цепь – 146 аминок-т. II стр-ра: 8α-спиралей (названия от A до H) ------------ // ------------ III стр-ра: глобула компактная IV стр-ра: отсутствует. ------------ // ------------ Олигомер: 2α-цепи и 2β-цепи. Строение активного центра Mb и Hb: 1) Активный центр Mb и Hb расположен в «кармане», образованными двумя спиралями F и E, имеющие гидрофобные радикалы аминокислот, за исключением 2 остатков гистидина F8 и E7. 2) В центре гема располагается ион Fe2+, связанный 4 связями с атомами азота пиррольных колец. 3) Fe гема связывается в активном центре ещё одной прочной связью с Гис F8 и выходит из плоскости гема. 4) O2 – гидрофобная молекула, легко проникает в активный центр и связывается с Fe гема по другую сторону его плоскости. 5) Гис E7 – создает оптимальные условия для связывания O2 и уменьшает сродство гема к CO. Строение активного центра Mb и Hb (схема из малинового учебника): Кривые диссоциации O2 для Mb и Hb: 1) Mb имеет очень большое сродство к O2. При ПД O2 – 40 мм рт. ст. в венозной крови Mb полностью насыщен O2. И даже при низком ПД O2 – (10-20 мм рт. ст.) Mb почти полностью насыщен O2. Только при тяжелой физической работе ПД O2 может снизиться до 5 мм рт. ст., и Mb способен легко отдавать O2. 2) Hb – насыщается в легких O2 на 100%, а в тканях отдает его в зависимости от ПД. В отдыхающих мышцах ПД O2 – 40-50 мм рт. ст. При этом Hb отдает тканям ~ 20% O2. В работающей мышце ПД O2 падает до 10 мм рт. ст. и Hb резко ↑ отдачу O2. Вывод: отдача O2 в ткани – регулируемая функция Hb. Это происходит благодаря кооперативным изменениям конформации протомеров. Кооперативные взаимодействия между протомерами характерны для всех олигомерных белков. Это проявляется в том, что изменение конформации одной из субъединиц олигомерного белка приводит к изменению конформации всех субъединиц, т.е. конформации всего белка. В легких: присоединение 1-й молекулы O2 к протомеру Hb меняет конформацию всего олигомерного белка и облегчает присоединение следующих молекул O2 к другим протомерам. Последняя молекула O2 присоединяется к Hb в 300 раз легче, чем первая. В тканях: отрыв 1-й молекулы O2 облегчает отщепление от Hb всех последующих молекул O2. Типы гемоглобина: Все типы гемоглобина являются тетрамерами и содержат 2α-цепи. 1) Гемоглобин A – 2α- и 2β-цепи. Составляет ~ 98% гемоглобина эритроцитов взрослого человека; 2) Гемоглобин A2 – 2α- и 2δ-цепи. Составляет ~ 2% гемоглобина взрослого человека; 3) Гемоглобин эмбриональный – 2α- и 2ε-цепи. Обнаруживается на ранних этапах развития плода; 4) Гемоглобин F – 2α- и 2γ-цепи. Приходит на смену раннему гемоглобину плода на 6-м месяце развития; На способность Hb отдавать O2 в тканях сильно влияет наличие регуляторов, к которым относятся H+, CO2 и 2,3-бифосфоглицерат (2,3-БФГ). Эффект Бора Основные функции гемоглобина: 1) транспорт O2 из легких в ткани; 2) транспорт CO2 и избытка H+ из тканей обратно в легкие. Эффект Бора – увеличение освобождения кислорода гемоглобином в зависимости от концентрации протонов. Влияние 2,3-БФГ на сродство Hb к O2. O O P O COO CH2 CH O O O P O O2,3-Бифосфоглицерат 2,3-БФГ – вещество, которое синтезируется в эритроцитах из промежуточного продукта гликолиза – 1,3-бифосфоглицерата. 2,3-БФГ в норме присутствует в эритроцитах ~ в той же концентрации, что и Hb. 2,3-БФГ присоединяется к Hb в специальном участке (центральной полости), расположенном между 4 протомерами. 2,3-БФГ может присоединяться только к дезоксигемоглобину. Олигомерная структура гемоглобина (схема из малинового учебника): В капиллярах легких, где высокое ПД O2 – Hb соединяется с O2 => происходит сужение полости между протомерами Hb, что приводит к вытеснению 2,3-БФГ из Hb. И Hb практически не связан с 2,3-БФГ. В капиллярах тканей, где ниже парциальное давление O2, 2,3-БФГ присоединяется к дезоксигемоглобину => ↓ его сродство к O2 => облегчается переход O2 в ткани. Hb(O2)4 + 2,3-БФГ Ткани Легкие Hb . 2,3-БФГ + 4O2 Концентрация 2,3-БФГ может увеличиваться при адаптации организма к гипоксии (высокогорье, обструктивная эмфизема легких). Это снижает сродство Hb к O2 и увеличивает количество O2, транспортируемого в ткани. 2,3-БФГ присоединяется к Hb не в том участке, к которому присоединяется O2, а в другом. Такой лиганд называется: аллостерический. Центр, к которому присоединяется аллостерический лиганд, называется: аллостерический центр. Аллостерические белки: Аллостерические белки: 1) Олигомерные белки, состоящие из 2-х или более субъединиц – регуляторной и каталитической. 2) Имеют 2 пространственно разделенных центра, расположенные в разных субъединицах: С – каталитическая субъединица. Содержит активный центр, к которому присоединяется – главный лиганд белка. R – регуляторная субъединица. Содержит регуляторный (аллостерический центр), к которому присоединяется – лигандэффектор. Эффекторы делятся на: активаторы (A) и ингибиторы (I). Они влияют на сродство АЦ белка к его главному лиганду. A – увеличивают сродство; I – уменьшают. Физико-химические свойства белков: 1) 2) 3) 4) 5) Форма молекул; Молекулярная масса; Суммарный заряд; Растворимость; Изоэлектрическая точка Растворимость белков зависит от суммарного заряда и общего количества полярных групп: 1. Растворимость белка тем больше, чем больше суммарный заряд. 2. Если у двух белков одинаковый суммарный заряд, то растворимость больше у того белка, у которого больше полярных групп. Также на растворимость белков влияет pH среды, присутствие солей в растворе и др. Изоэлектрическая точка белка (pI) – это такое значение pH, при котором суммарный заряд белка равен нулю. В изоэлектрической точке количество положительно и отрицательно заряженных групп белка одинаково. Если суммарный заряд белка в нейтральной среде то pI < 7. Если суммарный заряд белка в нейтральной среде то pI > 7. Если суммарный заряд белка в нейтральной среде = 0 то pI = 7. Ферменты – это биологические катализаторы белковой природы. Катализаторы – это вещества, принимающие участие в химических реакциях, ускоряя их, но сами в них не расходующиеся. Ферменты катализируют химические происходящие в организме. реакции, Ферменты катализируют превращение веществ, которые называются субстратами (S) в продукты (P): S E Отличие ферментов катализаторов: P от небиологических 1. Высокая эффективность действия (скорость ферментативных реакций в 106 – 1012 раз выше, чем соответствующих неферментативных реакций). 2. Высокая специфичность действия (фермент катализирует превращение одного конкретного субстрата, либо схожей группы субстратов). 3. Мягкие условия протекания ферментативных реакций (t ~ 37 °C, нормальное атмосферное давление, pH, близкий к нейтральному). 4. Способность к регуляции. Активный центр фермента: 1. Участок молекулы фермента, сформированный на уровне третичной структуры, ответственный за связывание с субстратом по принципу комплементарности и участвующий в катализе. 2. Расположен в узком гидрофобном углублении (щели) поверхности молекулы фермента. 3. Активный центр фермента в отличие активного центра белка имеет 2 участка: 1) Субстратсвязывающий участок; 2) Каталитический участок. от 4. В активный центр фермента часто входит участок или домен для связывания кофактора. Типы специфичности ферментов I. Субстратная II. Каталитическая (пути превращения) Абсолютная 1. Стереоспецифичность 3. Относительная (групповая) 2. I.1. Абсолютная субстратная специфичность – Фермент катализирует превращение только одного конкретного субстрата. Пример: I.2. Относительная (групповая) субстратная специфичность – Фермент катализирует однотипные превращения схожих по строению веществ. Пример: Фермент гидролиз жиров: Липаза катализирует – I.3. Стереоспецифичность катализирует превращение только стереоизомеров для данного вещества. Фермент одного из Пример: Фермент Фумараза катализирует присоединение воды только к фумарату, но не к малеиновой кислоте: II. Каталитическая специфичность (специфичность пути превращения) – фермент катализирует только одно превращение субстрата из всех возможных: E1 S A E2 E3 B C Пример: N CH2 CH COOH NH2 гистидиндекарбоксилаза N H гистидин гистидаза NH3 CO2 N CH2 CH2 N CH NH2 N H гистамин N H уроканиновая кислота CH COOH Этапы ферментативного катализа. I – образование фермент-субстратного комплекса: На этом этапе: а) субстрат приближается к активному центру фермента. б) происходит взаимное изменение конформации E и S, возникает строгая комплементарность между S и активным центром E (индуцированное соответствие). II. Дестабилизация связей в молекуле субстрата. III. Образование продуктов реакции и выход их из области активного центра фермента. (III этап часто делят на III и IV: III этап: образование продуктов реакции. IV этап: выход продуктов из области активного центра фермента.) Кинетика ферментативных реакций. I. Зависимость скорости реакции от концентрации субстрата: Vmax – это такая V ферментативной реакции, при которой достигается полное насыщение фермента субстратом, т.е. когда все активные центры фермента связаны с субстратом. KM – константа Михаэлиса: 1) KM численно равна концентрации субстрата, при которой скорость реакции равна ½ Vmax. 2) KM показывает сродство E к S. Чем меньше KM, тем больше сродство и наоборот. II. Зависимость скорости реакции от температуры: При нагревании: рвутся слабые связи в молекуле фермента (гидрофобные, ионные и водородные) => меняется конформация E => нарушается структура активного центра E => уменьшается активность E. III. Зависимость скорости реакции от pH среды: Оптимум pH – это такое значение pH, при котором E проявляет максимальную активность. Для каждого E характерен довольно интервал pH, при котором он активен: узкий При изменении pH среды в сторону от оптимума активность E уменьшается: Причины: изменяется ионизация ионогенных групп => рвутся ионные и некоторые водородные связи в молекуле фермента => меняется конформация E => нарушается структура активного центра E => уменьшается активность E. COO COO HO CH3 ... ... + H 3N HO HO H 3C + ион. связь + H вод. связь вод. связь гидрофобная связь COOH + H 3N COOH HO Кофакторы ферментов и их роль в катализе. Если фермент является сложным белком, то в его состав всегда входит небелковый компонент, который называется: Кофактор. Белковая часть Апофермент. такого фермента называется: А комплекс апофермента с кофактором получил название: Холофермент: Апофермент + Кофактор Холофермент Каталитическую активность в проявляет только Холофермент. этом случае Кофакторы Коферменты Ионы металлов Роль ионов металлов в катализе: 1) участвует в присоединении субстрата (Mg2+, Mn2+, Zn2+) 2) принимают участие в катализе (Zn2+, Cu2+, Fe2+) 3) принимают участие в стабилизации третичной и четвертичной структуры фермента (K+, Ca2+, Zn2+) 4) активаторы ферментов (Ca2+) Коферменты: 1) органические вещества, предшественниками которых, как правило, являются витамины; 2) принимают непосредственное участие в катализе, наряду с радикалами аминокислот. Некоторые коферменты непрочно связаны с апоферментом (NAD+, NADP+, HS-KoA, H4-фолат). Есть коферменты, прочно связанные с апоферментом, т.е. представляют с собой простетическую группу (гем, FAD, FMN, биотин). Примеры коферментов Тиаминдифосфат (ТДФ): Биотин (витамин H): Кофермент A: Классификация ферментов. В основе классификации ферментов лежит тип катализируемой ими реакции. Все ферменты делятся на 6 классов: I. Оксидоредуктазы – катализируют окислительно-восстановительные реакции. Основные подклассы: 1. Оксидазы 2. Гидроксилазы 3. Дегидрогеназы – катализируют реакции присоединения / отщепления атомов водорода (дегидрирования). По типу кофермента различают: 1) NAD+ и NADP+ – зависимые дегидрогеназы (предшественник: витамин PP); 2) FAD и FMN – зависимые дегидрогеназы (предшественник: витамин B2) Коферменты NAD+, NADP+, FAD и FMN выполняют роль акцепторов атомов водорода. + + Строение коферментов NAD и NADP : + + CO Активная часть коферментов NAD+ и NADP+: + N R CO Витамин PP: N NH2 NH2 1) Роль коферментов NAD+ и NADP+ в реакциях дегидрирования: H Д . H2 + + H CONH 2 CONH 2 + N N R R НАД+ (или НАДФ+) – окисленная форма Д Д H . H2 + NAD+ (или NADP+) + H + НАДH + H+ (или НАДФH + H+) – восстановленная форма Д + NADH + H+ (или NADPH + H+) Пример реакции с участием NAD+: COOH COOH HO C H + NAD+ CH2 COOH Малат Малатдегидрогеназа C O + NADH+H+ CH2 COOH Оксалоацетат Строение флавиновых коферментов (FMN и FAD): 2) Роль коферментов FAD и FMN в реакциях дегидрирования: O H3C Д . H2 + N H3C N NH N O Д + H3C H N H3C N R R . H2 + FAD (или FMN) NH N O H ФАДH2 (или ФМН H2) – восстановленная форма ФАД (или ФМН) – окисленная форма Д O Д + FADH2 (или FMN.H2) Пример реакции с участием FAD: COOH CH2 + E-FAD CH2 COOH Сукцинат Сукцинатдегидрогеназа COOH CH + CH COOH Фумарат E-FADH2 II. Трансферазы – катализируют реакции переноса функциональных групп с одной молекулы на другую. Основные подклассы: 1. Киназы – катализируют перенос фосфатной группы, донором которой является АТФ (реже ГТФ). Примеры: 1) H2C OH HC OH H2C OH + АТФ OH HC OH + АДФ H2C OPO3H2 Глицерол-3-фосфат Глицерол 2) H2C OH OPO 3H2 + АТФ протеин + АДФ фосфопротеин Кофакторы: ионы Mg2+ и Mn2+. Реакции, которые катализируют называются р-циями фосфорилирования. киназы, 2. Аминотрансферазы – катализируют перенос аминогруппы с аминокислоты на кетокислоту. Пример: Кофермент: витамина B6. Пиридоксальфосфат, 3. Метилтрансферазы группы. – перенос производное метильной 4. Ацилтрансферазы – перенос ацильной группы. III. Гидролазы – катализируют реакции разрыва связей в молекуле с участием воды, т.е. путем гидролиза. Основные подклассы: 1. Фосфатазы – катализируют гидролитическое отщепление фосфатной группы: OH OPO 3H2 + H 2O Фосфопротеин + H3PO4 Протеин Реакции, которые катализируют фосфатазы, называются р-циями дефосфорилирования. 2. Липазы – гидролиз связей в жирах: 3. Пептидазы 4. Гликозидазы 5. Эстеразы IV. Лиазы. Существует 2 разновидности лиаз: 1 разновидность: Лиазы, катализирующие реакции разрыва связей способом, отличным от гидролиза и окисления. Для этой разновидности подклассы: 1. лиаз существуют Декарбоксилазы – катализируют реакции отщепления карбоксильной группы в виде CO2. Пример: Кофермент – Пиридоксальфосфат (Производное витамина B6). 2. – катализируют реакции Дегидратазы отщепления воды от молекулы с образованием двойной связи. 2 разновидность: Лиазы, катализирующие реакции присоединения молекул по двойной связи. Подклассы: Гидратазы – катализирующие реакции присоединения молекулы воды по двойной связи. COOH CH + H 2O фумараза COOH OH HC CH CH2 COOH COOH Фумарат Малат V. Изомеразы – катализируют взаимопревращение изомеров. Подклассы: изомеразы, мутазы. Пример: CH2OPO3H2 CH2OPO3H2 O H H OH H H OH OH H O H OH Глюкозо - 6-фосфат Кофакторов нет. CH2OH OH OH H OH H Фруктозо - 6-фосфат VI. Лигазы. 2 разновидности: 1 разновидность: Синтетазы – катализируют реакции соединения 2-х молекул при участии энергии АТФ (реже ГТФ). Подклассы: Карбоксилазы – катализируют реакции присоединения карбоксильной группы к молекуле в виде CO2. Пример: Кофермент: биотин (витамин H). 2 разновидность: Синтазы – катализируют соединение молекул при участии энергии макроэргических связей субстрата. Активность фермента. В условиях избытка субстрата скорость ферментативной реакции прямо пропорциональна концентрации фермента. V [E] Концентрация E крайне мала (обычно для протекания р-ции с большой скоростью требуется буквально несколько десятков молекул E). Поэтому, на практике пользуются условными величинами, характеризующими активность E: 1 МЕ (международная единица активности) – это такое количество фермента, которое катализирует превращение 1 мкмоль субстрата за 1 мин или образование 1 мкмоль продукта за 1 мин при оптимальных условиях. Оптимальные условия индивидуальны для каждого E и зависят от температуры, pH среды, наличия Ko (если есть), отсутствие активаторов и ингибиторов. Для того, чтобы оценить количество молекул E среди других белков данной ткани, определяют удельную активность. Удельная активность – это отношение количества единиц активности фермента в образце ткани к массе белка (в мг) в этой ткани. По удельной активности часто судят об очистке E: чем меньше посторонних белков, тем выше Aуд. (В обязательную программу по Биохимии не входит): В 1973 году была активности – катал. принята новая единица Ингибиторы ферментов Ингибиторы – это вещества полностью или частично подавляющие активность ферментов. Типы ингибиторов Обратимые Необратимые Конкурентные Неконкурентные Специфические Неспецифические Обратимые ингибиторы присоединяются к ферменту слабыми связями и могут легко отделяться от фермента. Обратимые конкурентные ингибиторы: 1) являются субстрата; структурными аналогами 2) конкурируют с S за связывание с активным центром E; 3) При увеличении концентрации субстрата степень ингибирования уменьшается. А при избытке субстрата действие ингибитора на фермент практически полностью прекращается. Пример: FAD FAD.H2 COOH CH 2 CH 2 сукцинатдегидрогеназа COOH Сукцинат COOH CH 2 COOH сукцинатдегидрогеназа Малонат COOH CH CH COOH Фумарат Зависимость скорости реакции от концентрации S в присутствии и отсутствии обратимого конкурентного ингибитора: ! При обратимом конкурентном ингибировании возрастает KM, но, как правило, не меняется максимальная скорость р-ции. KM возрастает, т.к. как бы снижается сродство E к S, поскольку присоединению S к АЦ фермента мешает ингибитор. Vmax, как правило, не меняется, т.к. при большом избытке S происходит вытеснение I из АЦ фермента и может быть достигнута максимально возможная скорость реакции. Обратимые неконкурентные ингибиторы Присоединяются к ферменту в участке, расположенном вне области активного центра меняется конформация E меняется структура активного центра E уменьшается активность E. ! При обратимом неконкурентном ингибировании уменьшается Vmax, но, как правило, не меняется KM . Vmax уменьшается, т.к. кол-во активных молекул E, т.к. часть из них, связанных с I, на время полностью исключаются из катализа. У тех молекул E, которые не связаны ингибитором, сродство к S остается прежним. Поэтому, KM, как правило, меняться не будет. Хотя, в ряде случаев, KM может возрастать. с Необратимые ингибиторы – присоединяются к ферменту, образуя прочную ковалентную связь с его функциональными группами. Или вызывают необратимую денатурацию фермента. Необратимые специфические ингибиторы – присоединяются к строго определённой группе в области активного центра E, образуя с ней прочные ковалентные связи. Пример: Ацетилхолинэстераза ингибируется фосфорорганическими соединениями. Например: зарин и др. диизопропилфторфосфат (ДФФ), Необратимые неспецифические ингибиторы. Существует 2 разновидности: 1) ингибиторы, образующие прочную ковалентную связь с группами, расположенными в любом участке фермента (довольно часто: вне области активного центра). Они не специфичны к определенным ферментам. Пример: n-Хлормеркурибензоат присоединяется к свободным SH-группам любых ферментов. Его действие неспецифично. 2) ингибиторы, которые присоединяются к молекуле фермента вне области активного центра и образуют с ней плохо растворимые комплексы (H2S, соли тяжелых металлов). В больших дозах вызывают необратимую денатурацию фермента. Пример: Ацетат свинца (свинцовая примочка) – вызывает необратимую денатурацию многих ферментов (и других белков). Регуляция действия ферментов Метаболический путь – это цепь взаимосвязанных реакций, в ходе которой исходный субстрат S превращается в конечный продукт P за несколько стадий. Метаболический путь бывает: Линейный: E1 S E2 A B E3 C ....... En P Разветвленный: S E1 A E2 E3 B C E5 K E4 E6 D ....... L E7 En P1 M ....... Em Регуляция скорости реакций метаболического пути осуществляется путем регуляции действия определенных ферментов, которые называются регуляторными. Признаки регуляторных реакций: 1) самая медленная реакция пути; 2) необратимая или практически необратимая реакция; 3) реакции на разветвлении метаболического пути P2 Очень часто ингибитором регуляторного фермента является конечный продукт метаболического пути. S E1 A E2 B E3 C ....... En P Отрицательная обратная связь Регуляторные ферменты метаболического пути, как правило, являются аллостерическими. Существует 2 основных способа регуляции действия ферментов: 1) путем изменения количества фермента; 2) путем изменения активности фермента при неизменном количестве. 2 способ гораздо более быстрый. Способы регуляции активности ферментов. 1. Частичный протеолиз: Это регуляция на уровне первичной структуры белка. 2. Фосфорилирование и дефосфорилирование: - OH ... OOC АТФ I. АДФ взаи одно моотта име л нны кивани х зар е ядов OPO32 OO C - - II. Протеин H3PO4 H2O Фосфопротеин I. фосфорилирование – катализируют ферменты: протеинкиназы (A, C, G и др.); II. дефосфорилирование – ферменты: протеинфосфатазы. катализируют Это регуляция на уровне третичной структуры белка. Одни ферменты в ходе фосфорилирования активируются, другие, напротив, становятся неактивными. Примеры: Липаза – OH + АТФ Липаза – OPO3H2 (протеин) (фосфопротеин) неактивная активная Гликогенсинтаза –OH + АТФ Гликогенсинтаза –OPO3H2 (протеин) активная (фосфопротеин) неактивная 3. Аллостерическая регуляция: Аллостерические ферменты: 1) Олигомерные белки, состоящие из 2-х или более субъединиц – регуляторной (R) и каталитической (C). 2) Имеют 2 пространственно разделенных центра: активный центр, расположенный в C субъединице и аллостерический центр, расположенный в R субъединице. 3) К АЦ присоединяется S фермента. К аллостерическому – лиганд-эффектор. 4) Эффекторы делятся на: активаторы (A) и ингибиторы (I). Эффекторы влияют на сродство АЦ фермента к S. A – увеличивают сродство; I – уменьшают. Схема аллостерического ингибирования: Это регуляция белков-ферментов, имеющих четвертичную структуру. 4. Белок-белковые взаимодействия: Это обратимый механизм регуляции активности белков-ферментов, который сопровождается изменением их четвертичной структуры. Пример: регуляция каталитической активности фермента – протеинкиназы А ассоциацией/ диссоциацией протомеров. а) Активация протеинкиназы A: б) Инактивация протеинкиназы A: Фермент фосфодиэстераза катализирует гидролиз цАМФ до АМФ, что приводит к сборке тетрамерного комплекса протеинкиназы А, который неактивен. Ферменты в медицине. 1. Использование ферментов в качестве лекарств. 1) заместительная терапия – использование ферментов при их недостатке в организме Примеры: а) пепсин при резекции желудка, ахилии, гастритах; б) при дефиците ферментов поджелудочной железы принимают внутрь препараты фестал, энзистал, мезим-форте и др., содержащие эти ферменты. 2) комплексная терапия – применение ферментов в сочетании с другой терапией. Примеры: а) Протеолитические ферменты (трипсин, химотрипсин) для обработки гнойных ран. Они расщепляют белки погибших клеток, практически не воздействуя на живые клетки. Также эти ферменты используются для удаления сгустков крови или вязких секретов при воспалительных заболеваниях дыхательных путей. Трипсин и химотрипсин – это сериновые протеазы. Катализируют гидролиз пептидных связей в белках. В активный центр входит «триада» аминокислот: серин, гистидин и аспартат. ! Пепсин для обработки гнойных ран использовать нельзя, т.к. его оптимум pH = 1,5 – 2. При pH 6 происходит его денатурация. б) Ферменты нуклеазы (дезоксирибонуклеаза рибонуклеаза) – противовирусные средства. Например: лечение вирусного конъюнктивита. и в) Фермент гиалуронидаза, катализирующий расщепление гиалуроновой кислоты, используют подкожно и внутримышечно для рассасывания рубцов после ожогов и операций (гиалуроновая кислота образует сшивки в соединительной ткани). г) Ферменты стрептокиназа, урокиназа и др. широко применяются при тромбозах и тромбоэмболиях. д) Использование лейкозов. аспарагиназы для лечения Во многих лейкозных клетках отсутствует фермент, участвующий в синтезе аспарагина. Поэтому, они получают аспарагин только из крови. В кровь больных лейкозом внутривенно вводят фермента аспарагиназу снижается концентрация аспарагина в крови замедляется синтез белков в лейкозных клетках, что ведет к их гибели. 2. Использование ферментов аналитических реактивов. в качестве 1) глюкооксидаза – для количественного определения глюкозы в крови и моче. 2) уреаза – для определения содержания количества мочевины в крови и моче. 3) липаза – для определения количества жиров (триацилглицеролов) в крови. 4) холестеролоксидаза – для определения количества холестерола в крови. 3. Использование ферментов в диагностике заболеваний – энзимодиагностике. Энзимодиагностика – постановка или уточнение диагноза заболевания с помощью определения активности ферментов в крови или другой биологической жидкости. Требования к ферментам для использования в энзимодиагностике: их 1) Органо - и тканеспецифичность ферментов; 2) Выход ферментов в кровь или другую биологическую жидкость при повреждении органа; 3) Отсутствие или низкая активность ферментов в крови или другой биологической жидкости в норме; 4) Наличие специфичных изоферментов в разных тканях. Примеры: -амилаза – маркер панкреатита. Кислая фосфатаза – маркер рака предстательной железы. Изоферменты – это разные формы одного и того же фермента, катализирующие одну и ту же химическую реакцию. Как правило, это Олигомерные белки, состоящие из разного набора субъединиц. Изоферменты отличаются друг от друга: 1) Локализацией; 2) Электрофоретической подвижностью; 3) Сродством к субстрату. Примеры: 1) 3 изофермента для Креатинкиназы: И в мышцах, и в сердце, и в головном мозге есть этот фермент, но наблюдаются разные соотношения его изоферментов. Если в крови преобладает форма фермента BM, значит у человека – инфаркт миокарда. Причем, активность КК1 (BM) достигает своего пика в крови в 1-2 сутки от начала инфаркта миокарда. 2) Лактатдегидрогеназа (ЛДГ) – тетрамер, состоящий из 2-х типов субъединиц (H и M): 5 изоферментов для ЛДГ: Пример: Увеличение в крови активности ЛДГ1 и ЛДГ2 – наиболее характерно для инфаркта миокарда. Причем, их активность повышается ко 2 суткам от начала инфаркта миокарда, достигает своего пика к 3-4 суткам и затем медленно уменьшается в течение нескольких дней. Использование ингибиторов ферментов в качестве лекарств. 1. Для лечения миастении используются лекарства – ингибиторы фермента ацетилхолинэстеразы: Пример: Прозерин – обратимый конкурентный ингибитор ацетилхолинэстеразы: 2. Для лечения бактериальной инфекции используются Сульфаниламидные препараты: Например: 3. Аспирин – противовоспалительное средство. Является необратимым специфическим ингибитором ферменты Циклооксигеназы, участвующего в синтезе простагландинов: O OH + H3C C O COOH Аспирин E Циклооксигеназа O O C CH3 + HO COOH Салициловая Ацетилированный кислота фермент 4. Панкреатит – заболевание, связанное с самоперевариванием поджелудочной железы из-за преждевременной активации трипсина в самой поджелудочной железе. В качестве лекарств используют трасилол и его аналоги (контрикал, гордокс), которые являются обратимыми конкурентными ингибиторами трипсина. 5. Аллопуринол – для лечения подагры. Заболевание связано с образованием кристаллов солей мочевой кислоты – уратов в области мелких суставов, что вызывает воспаление и острую боль. Мочевая кислота – трудно растворимое вещество. Кристаллы уратов образуются даже при незначительном увеличении ее концентрации. Реакцию образования мочевой к-ты в организме катализирует фермент ксантиноксидаза: OH O N N N N H 2O2 2H2O 2H2O2 HN ксантиноксидаза O гипоксантин NH N H N H O мочевая кислота Для лечения подагры используют аллопуринол – специфический ингибитор ксантиноксидазы: OH N N N H N аллопуринол При ингибировании ксантиноксидазы вместо мочевой к-ты начинает накапливаться гипоксантин, но т.к. он растворим в воде ~ в 10 раз лучше, чем мочевая к-та, то он легко выводится с мочой и кристаллов не образуется. 6. Использование ингибиторов фосфодиэстеразы: а) кардиотонические средства для терапии при острой сердечной недостаточности; б) для лечения бронхиальной астмы. Пример: препарат Теофеллин. Лечение основано на ингибировании фосфодиэстеразы = [цАМФ] = активность протеинкиназы A. Это увеличивает сократительную деятельность миокарда, расширяет сосуды; а также оказывает бронхорасширяюший эффект. МАТРИЧНЫЕ БИОСИНТЕЗЫ. ОСНОВЫ МОЛЕКУЛЯРНОЙ ГЕНЕТИКИ. Авторы – доцент В.А.Голенченко, доцент Д.В.Астахов СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ. ДНК – дезоксирибонуклеиновая кислота РНК – рибонуклеиновая кислота Нуклеиновые кислоты – это линейные полимеры, построенные из 4 типов мономерных звеньев – нуклеозидмонофосфатов (НМФ). Линейный полимер: ……………. полимер На рисунке: – мономерное звено нуклеозидмонофосфат (нуклеотид) Строение мономерного звена Фосфорный остаток – пентоза – азотистое основание СОСТАВ НУКЛЕОТИДОВ 1 МОНОМЕРЫ ДНК АЗОТИСТЫЕ ОСНОВАНИЯ ПЕНТОЗА А- аденин д-рибоза (дезоксирибоза) G – гуанин C – цитозин Т – тимин МОНОМЕРЫ РНК АЗОТИСТЫЕ ОСНОВАНИЯ ПЕНТОЗА А- аденин рибоза G – гуанин C – цитозин U – урацил ПЕРВИЧНАЯ СТРУКТУРА ДНК – это порядок чередования дезоксирибонуклеозидмонофосфатов в полинуклеотидной цепи. В цепи ДНК различают 5′-конец и 3′-конец. На 5′конец находится фосфорный остаток, связанный с 5′ углеродным атомом дезоксирибозы. На 3′-конце цепи ДНК – гидроксильная группа у 3′-го углеродного атома д-рибозы. Добавление к номеру углеродного атома в рибозе или дезоксирибозе значка штрих (′) было введено, чтобы отличать номера атомов в азотистых основаниях от номеров атомов в пентозах. 2 Первичная структура нуклеиновых кислот. Х=Н для ДНК, Х=ОН для РНК Связи в молекуле нуклеиновых кислот: 1 – 5′- фосфоэфирная; 2 – N-гликозидная связь; 3 – 3′5′- фосфодиэфирная. Линейная последовательность записывается с помощью однобуквенного кода, от 5′ к 3′-концу. Например, если на рисунке фрагмент цепи РНК, то 5′ -А-С-G-3′-, если ДНК – 5′-дА-C-G- 3′ВТОРИЧНАЯ СТРУКТУРА ДНК Молекула ДНК состоит из 2-х антипараллельных цепей с комплементарной последовательностью нуклеотидов. Между комплементарными азотистыми основаниями возникают водородные связи, например: 5′-А-G-C-Т-А-……….-А-Т-3′ 3′-Т-C-G-А-Т-……….-Т-А-5′ 3 По правилу ЧАРГАФФА – в молекуле ДНК количество пуринов равно количеству пиримидинов, т.е.: А + G = Т + C, а также количество А = Т и G = C; Цепи ДНК закручены относительно друг друга и вокруг общей оси. Комплементарные основания уложены в стопку в сердцевине спирали между ними возникают гидрофобные взаимодействия. Двойная спираль ДНК ТРЕТИЧНАЯ СТРУКТУРА ДНК Одна молекула ДНК называется хромосомой, в ядре диплоидных клеток 46 хромосом. В ядре половых клеток 23 хромосомы гаплоидный (одинарный) набор. Длина ДНК всех хромосом 1,74 м. В компактизации ДНК участвуют белки: ● гистоны; ● негистоновые белки. Белки гистоны содержат много остатков Арг и Лиз, поэтому имеют суммарный положительный заряд. Отрицательно заряженные (фосфатные) группы цепей ДНК образуют 4 ионные связи с положительно заряженными радикалами лизина и аргинина. Гистоны участвуют в: ● Компактизации ДНК ● Защите ДНК от действия нуклеаз ● Регуляции матричных процессов. ДНК и все связанные с ней белки и ферменты называются хроматином. ПЕРВИЧНАЯ СТРУКТУРА РНК – это порядок чередования рибонуклеозидмонофосфатов в полинуклеотидной цепи. Молекула РНК состоит из одной полинуклеотидной цепи. На 5′-конце находится свободная фосфорная группа, на 3′-конце – ОН- группа рибозы. 5 ВТОРИЧНАЯ СТРУКТУРА РНК. Отдельные антипараллельные участки цепи ДНК образуют спирализованные петли («шпильки») за счет водородных связей между азотистыми основаниями – пуринами и пиримидинами: А-U и G-C. В цитоплазме клеток присутствуют три типа рибонуклеиновых кислот – тРНК – транспортная РНК мРНК – матричная РНК рРНК – рибосомальная РНК Они различаются по: ● первичной структуре; ● молекулярной массе; ● конформации; ● продолжительности жизни; ● функциональной активности. 6 РЕПЛИКАЦИЯ Процесс удвоения хромосом называется репликацией. Этот процесс предшествует делению ядра. Репликация происходит: ● в ядре; ● в S-фазу клеточного цикла. Матрицей являются обе цепи (родительской) ДНК. Субстраты и источники энергии – дНТФ (дезоксирибонуклеозидтрифосфаты: дАТФ, дГТФ, дТТФ, дЦТФ), для активации которых необходимы ионы Мg2+; Синтез новых (дочерних) цепей идет от 5′ к 3′-концу антипараллельно матрице. ЭТАПЫ РЕПЛИКАЦИИ Инициация – образование репликативной вилки. На этом этапе участвуют: ● регуляторные белки – факторы роста; ● ферменты – ДНК-топоизомераза I и ДНК-хеликаза; ● SSB- белки. Источник энергии – молекулы АТФ 7 ● Фермент ДНК-топоизомераза I расщепляет 5′,3′-фосфодиэфирную связь в одной из цепей ДНК и присоединяется ковалентно к фосфорному остатку на 5′-конце в точке разрыва. ● Происходит локальное раскручивание двойной спирали ДНК при участии ДНК-хеликазы, которая используя в качестве источника энергии АТФ разрывает водородные связи между цепями ДНК. ● По окончании формирования репликативной вилки ДНК-топоизомераза I восстанавливает целостность цепи (фосфоэфирную связь) и отделяется от ДНК. SSВ-белки присоединяются к одноцепочечной ДНК, таким образом предотвращают их комплементарное скручивание и образование двухспиральной структуры. ЭЛОНГАЦИЯ. Репликацию (удвоение) ДНК катализируют ДНК-полимеразы α (альфа), β (бета), δ (дельта), ε (эпсилон), которые имеют в активном центре ион Zn2+. Недостаток этого микроэлемента в организме приводит снижению скорости ранозаживления, к задержке роста и развития у ребенка. ДНК-полимеразы β, δ и ε не имеют сродства к одноцепочечной ДНК, поэтому не могут инициировать процесс. Начинает элонгацию репликации ДНК-полимераза α. 8 9 1. ДНК-полимераза α присоединяется к определенному сайту (участку) одноцепочечных ДНК и синтезирует по принципу комплементарности фрагменты РНК – праймеры, состоящие из 8-10 рибонуклеозидов. Удлинение праймеров на ~ 150 дезоксирибонуклеотидов катализирует тоже ДНК-полимераза α. 2. Наличие 2-х цепочечных участков на каждой цепи позволяет ферментам ДНК- полимеразам δ и ε одновременно продолжить синтез новых (дочерних) цепей в направлении от 5′ к 3′-концу, антипараллельно матрице (3′,5′). Одна цепь растет по ходу раскручивания репликативной вилки, ее называют лидирующей цепью. 3. Вторая дочерняя цепь синтезируется (растет) против движения репликативной вилки. По мере раскручивания и доступности определенного сайта ДНК-полимераза α на второй матричной цепи синтезирует новый праймер, к которому присоединяется ДНК-полимераза ε или δ. Образованные ДНК-полимеразой α короткие фрагменты получили название – фрагменты Оказаки по имени ученого, впервые обнаружившего их образование. Цепь состоящая из фрагментов Оказаки (~ из 200 нуклеотидов) называется запаздывающей или отстающей. 4. Эндонуклеаза и РНК-гидролаза (РНКаза) удаляют праймеры, поэтому на лидирующей цепи образуется одна брешь, а на запаздывающей – несколько. 5. ДНК-полимераза β заполняет брешь, используя в качестве субстратов и источников энергии – дНТФ (дАТФ, дГТФ, дЦТФ, дТТФ). 6. ДНК-лигаза, с затратой энергии АТФ соединяет фрагменты Оказаки, состоящие после удаления РНК только из дНМФ, связанных 3′,5-фосфодиэфирной связью. Дочерние цепи комплементарны матричным и антипараллельны. Продукт (2 новые цепи) идентичен матричной ДНК. Образованные молекулы ДНК содержит одну матричную и одну дочернюю цепь. Такой механизм удвоения получил название – полуконсервативная репликация. 10 СУММАРНОЕ УРАВНЕНИЕ РЕПЛИКАЦИИ: РЕПАРАЦИЯ – это процесс исправления повреждений в ДНК. Процесс обеспечивает сохранение генетического материала на протяжении всей жизни организма. Репарация возможна только для 2-х цепочечных нуклеиновых кислот, при условии, если повреждение произошло в одной цепи (это правило для половых клеток). В клетках с диплоидным набором хромосом, даже при повреждении комплементарной пары нуклеотидов репарация возможна. Снижение скорости процесса приводит к накоплению повреждений в ДНК. Типы повреждений: ― спонтанные ● дезаминирование оснований (из цитозина образуется урацил); ● депуринизация – отщепление пуриновых оснований А и G ● ошибки репликации (замены, вставки или выпадения нуклеотидов) ― индуцируемые ● образование пиримидиновых димеров, например, тимидиловых, под действием УФО ● повреждение оснований (A, G, C или T) ДНК химическими мутагенами. Репарация: ● происходит в ядре; ● не связана с фазами клеточного цикла. 11 Матрица – одна цепь (неповрежденная) ДНК Субстраты и источники энергии – дНТФ (дезоксирибонуклеозидтрифосфаты: дАТФ, дГТФ, дТТФ, дЦТФ), для активации которых необходимы ионы Мg2+; Продукт процесса, исправленный участок цепи ДНК, комплементарен матрице Универсальная схема репарации: 12 В зависимости от типа повреждения количество этапов и ферментов, участвующих в его устранении может быть разным. Репарация с участием ДНК-N-гликозилазы: 13 Удаление неправильного азотистого основания и образование АП-сайта (участка лишенного азотистого основания) может проходить при участии ДНК-N-гликозилазы, которая расщепляет N-гликозидную связь между поврежденным (или неправильным) основанием и дезоксирибозой. В дальнейшем возможны 2 пути реализации репарации: ● без разрыва 3′,5-фосфодиэфирной связью, при участии фермента ДНК-инсертаза, который присоединяет к дезоксирибозе основание по принципу комплементарности. ● с разрывом 3′,5-фосфодиэфирной связью, при участии фермента АП-эндонуклеаза, а далее репарация идет по универсальному механизму. Репарация – устранение димеров тимина Фермент фотолиаза активируется на свету. При недостаточности фотолиазы развивается заболевание пигментная ксеродерма Если повреждения не исправляются ферментами репарации – они сохраняются в геноме. ТРАНСКРИПЦИЯ – первый этап реализации генетической информации. В ходе транскрипции образуются молекулы РНК мРНК – матрица для синтеза белка тРНК – выполняющие адапторную функцию 14 рРНК – входят в состав рибосом, на которых идет синтез белка. Транскрипция: ● происходит в ядре, ● не связана с фазами клеточного цикла. Матрица – 3′-5 цепь ДНК Субстраты и источники энергии – НТФ (рибонуклеозидтрифосфаты: АТФ, ГТФ, УТФ, ЦТФ), для активации которых нужны ионы Мg2+; Продукт процесса – молекула РНК комплементарна матричной ДНК Синтез идет от 5′ к 3′-концу, антипараллельно матрице. Основным ферментом процесса является Zn2+-зависимая РНК-полимераза, активность которой регулируется факторами инициации, элонгации и терминации. В основе процесса лежит принцип комплементарного спаривания оснований: СУММАРНОЕ УРАВНЕНИЕ ТРАНСКРИПЦИИ: 15 ЭТАПЫ ТРАНСКРИПЦИИ Инициация Присоединение ТАТА-фактора к промотору (определенный сайт на 3′-5′-цепи ДНК), включающему специфическую последовательность нуклеотидов -ТАТА- увеличивает его сродство к РНК-полимеразе и факторам инициации транскрипции. Образованный комплекс обеспечивает локальное расхождение цепей ДНК. 16 Элонгация На этом этапе фермент теряет связь с факторами инициации, но приобретает сродство к факторам элонгации. РНК-полимераза используя субстраты АТФ, ГТФ, ЦТФ и УТФ синтезирует молекулу РНК по принципу комплементарности, связывая нуклеотиды 3′5′фосфодиэфирной связью. Терминация Когда РНК-полимераза достигает сайта терминации (специфическая последовательность нуклеотидов) факторы элонгации отделяются от фермента, а присоединяются факторы терминации. Эти регуляторные белки облегчает отделение пре-РНК (первичного транскрипта) от матричной цепи ДНК. Образованные молекулы РНК функционально не активны. Их «созревание» происходит в ходе посттранскрипционных модификаций. «Созревание» (посттранскрипционные модификации) пре-мРНК : ● «Кэпирование» 5′-конца; ● Полиаденилирование 3′-конца; ● Сплайсинг (вырезание интронов и сшивание экзонов). «Зрелая» мРНК На этапе элонгации происходит построение на 5′-конце «кэпа» – 7-метилгуанозина, который связан с первым нуклеотидом пре-мРНК 5′,5′-фосфодиэфирной связью, причем между нуклеозидами не один фосфорный остаток, а три. 17 Функция кэпа: ● защита 5′-конца от действия нуклеаз (гидролаз) в цитозоле; ● обеспечение инициации трансляции. По завершении транскрипции фермент поли(А)-полимераза формирует поли-Апоследовательность (100-200 А) на 3′-конце. Функция поли-А: ● защита 3′-конца от действия нуклеаз в цитозоле; ● облегчение выхода мРНК из ядра. Сплайсинг Последовательности нуклеотидов присутствующие в матричной цепи ДНК но не входящие в «зрелую» РНК называется интронами. А участки сохраняющиеся в «зрелой» РНК называют кодирующими или экзонами. Вырезание интронов и сшивание экзонов происходит при участии малых ядерных рибонуклеопротеинов (мяРНП) - сплайсосом, которые разрывают 3′5′-фосфодиэфирную связь на границе: ↓ ↓ –экзон-интрон-экзон – Интроны удаляют, а экзоны соединяют, образуя между ними 3′5′- фосфодиэфирную связь. «Зрелая» мРНК выходит из ядра в цитоплазму. Альтернативный сплайсинг Альтернативный сплайсинг – это образование разных мРНК из одной и той же пре-мРНК, синтезированной с одного гена. В ходе этого процесса происходит соединение экзонов в разных комбинациях с формированием различных по первичной структуре «зрелых» мРНК. 18 Белки, которые синтезируются на основе этих мРНК выполняют сходные или различные функции. Таким образом, альтернативный сплайсинг позволяет увеличить разнообразие белковых продуктов генов, не увеличивая пропорционально этому размер генома, в том числе не создавая дополнительных копий генов. 19 Помимо сплайсинга разные РНК подвергаются и другим типам модификаций. Молекулы рРНК связывается со специфическими белками, образуются 40S и 60Sсубъединицы рибосом. У тРНК достраивается одинаковый для всех тРНК 3′-конец – ССА, модифицируются азотистые основания. Все пре-РНК комплементарны матричной цепи ДНК, «зрелые» РНК — нет. 20 МАТРИЧНЫЕ БИОСИНТЕЗЫ. ОСНОВЫ МОЛЕКУЛЯРНОЙ ГЕНЕТИКИ. Авторы – доцент В.А.Голенченко, доцент Д.В.Астахов Трансляция – процесс, в ходе которого «язык нуклеотидов», а именно линейная последовательность нуклеотидов зрелой мРНК «переводится на язык аминокислот». Словарь для этого перевода – генетический код, открытый в 60-годах ХХ столетия. СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА 1. Триплетность – одной аминокислоте соответствует кодон, состоящий из 3-х нуклеотидов (триплет); 2. Универсальность – смысл кодонов одинаков почти для всех живых организмов на Земле. 3. Специфичность – каждый кодон соответствует только одной аминокислоте; 4. Вырожденность – одну аминокислоту могут кодировать несколько триплетов (кодонов); 5. Однонаправленность – кодоны мРНК считываются от 5′ к 3′-концу; 6. Коллинеарность – последовательность триплетов в зрелой мРНК соответствует последовательности аминокислот в белке, синтезированном на этой матрице. 1 ТРАНСЛЯЦИЯ – БИОСИНТЕЗ БЕЛКА. ● Локализация процесса – цитозоль клеток; ● Матрица процесса – мРНК; ● Субстраты – 20 аминокислот; ● Источники энергии – макроэргические связи АТФ и ГТФ; ● тРНК – адапторы аминокислот к кодонам мРНК; ● Ферменты – аминоацил-тРНК синтетазы, катализирующие присоединение аминокислоты к тРНК. ● Регуляторные факторы – IF – фактор инициации; EF1 – фактор элонгации 1; EF2 – фактор элонгации 2; RF1, RF3 – факторы терминации. ● Кофактор – Мg2+; ● Рибосомы – рибонуклеопротеины (комплексы РНК и белков), на которых идет синтез белка; ● Синтез белка идет от N- к С-концу; ● Продукт процесса (белок) – коллинеарен матрице. АКТИВАЦИЯ АМИНОКИСЛОТ В активации аминокислот принимают участие тРНК. Эти нуклеиновые кислоты называют «адапторными молекулами», они помогают переводить «язык нуклеотидов» на «язык аминокислот». К акцепторному концу (3′-концу), который у всех тРНК имеет одинаковый триплет –ССА, ферменты аа-тРНК-синтетазы (аминоацил-тРНК-синтетазы) с затратой энергии АТФ присоединяют аминокислоту. Между остатком от α-карбоксильной группы аминокислоты и ОН-группой на 3′-конце тРНК формируется макроэргическая связь (~). 2 Другой триплет (кодон) тРНК находящийся в центре нуклеиновой кислоты, называется антикодоном, который присоединяется по принципу комплементарности и антипараллельности к кодону на мРНК. ЭТАПЫ ТРАНСЛЯЦИИ ИНИЦИАЦИЯ На этом этапе мРНК взаимодействует с: ● субъединицей 40S в области кэпа; ● Мет-тРНКМет; антикодон которой присоединяется к инициирующему кодону (AUG) мРНК по принципу комплементарности (между ними образуются водородные связи). ● 60S-субъединицей рибосомы. Этап протекает с затратой энергии ГТФ и при участии фактора инициации (IF). 3 Завершается инициация формированием 80S-рибосомы с двумя активными центрами: Р-пептидильным и А-аминоацильным. В Р-центре связана Мет-тРНКМет. ЭТАП – ЭЛОНГАЦИЯ состоит из 3-х повторяющихся стадий: 1. СТАДИЯ – связывание Про-тРНКПро в А-центре. Идет с затратой энергии ГТФ и при участии регуляторного белка EF1 – фактора элонгации 1. 2. СТАДИЯ – образование пептидной связи. Происходит пептидилтрансферазная реакция, разрывается макроэргическая связь между Мет и тРНКМет. Энергия связи используется на образование петидной связи между NH2- группой Про и СО-группой Мет. В А-центре образуется дипептидил-тРНК (Мет-Про-тРНКПро). 4 3. СТАДИЯ - транслокация С затратой энергии ГТФ и при участии фактора транслокации (IF2) происходит перемещение рибосомы на один кодон в направлении от 5′ к 3′-концу. Мет-Про-тРНКПро оказывается в Рцентре. Далее стадии элонгации продолжаются пока в А-центр не включиться один из терминирующих кодонов – UАG, UGА, UАА. ЭТАП – ТЕРМИНАЦИЯ Факторы терминации RF1, RF3 «узнают» терминирующие кодоны и обеспечивают: ● отделение белка от тРНК; ● разведение субъединиц рибосомы; ● разрыв водородных связей (RF1, RF3) между антикодоном тРНК и кодоном мРНК. Этап идет с затратой энергии АТФ. 5 ПОСТРАНСЛЯЦИОННЫЕ ПЕРЕСТРОЙКИ БЕЛКОВ Образованный полипептид функционально не активен. Для приобретения биологической активности белок подвергается посттрансляционным перестройкам (модификациям). 1. Фолдинг молекул. Формирование термодинамически наиболее выгодной пространственной структуры с участием белков шаперонов. 2.Частичный протеолиз. 3. Модификации радикалов аминокислот. Может идти как во время синтеза белка на рибосоме, так и после его завершения, например, ацилирование, гидроксилирование, гликозилирование, карбоксилирование, метилирование и др. 6 4. Присоединение простетической (небелковой) группы. Например, связывание кофермента в активном центре фермента, гема с протомерами гемоглобина и другие белки. 5. Сборка протомеров в олигомерные белки. Например, гемоглобин, лактатдегидрогеназа, синтаза жирных кислот и другие ферменты. 6.Образование дисульфидных (-S-S-) мостиков. Например, при формировании белкового гормона инсулина дисульфидные связи образуются по завершении частичного протеолиза. 7 ИСПОЛЬЗОВАНИЕ ИНГИБИТОРОВ СИНТЕЗА НУКЛЕОТИДОВ И МАТРИЧНЫХ БИОСИНТЕЗОВ В КАЧЕСТВЕ ПРОТИВООПУХОЛЕВЫХ И АНТИБАКТЕРИАЛЬНЫХ ПРЕПАРАТОВ. АНТИБАКТЕРИАЛЬНЫЕ препараты могут оказывать свое действие, подавляя синтез нуклеотидов у бактерий. Например, Сульфаниламидные препараты снижают: ● синтез нуклеотидов у микроорганизмов; ● синтез нуклеиновых кислот – РНК и ДНК; ● размножение микроорганизмов. АНТИБАКТЕРИАЛЬНЫЕ препараты могут ингибировать бактериальные ферменты: ДНКгиразу (аналог ДНК-топоизомеразы II у человека), РНК-полимеразу, поэтому снижается: ● скорость размножения бактерий (при действии ингибиторов ДНК-гиразы); ● синтез РНК у микроорганизмов (под действием ингибиторов РНК-полимеразы); ● синтез белков и размножение бактерий. АНТИБАКТЕРИАЛЬНЫЕ препараты вызывают нарушение структуры рибосом (30S и 50S) микроорганизмов, поэтому снижают: ● синтез белка у бактерий и их размножение. ПРОТИВООПУХОЛЕВЫЕ препараты могут подавлять синтез нуклеотидов в опухолевых клетках, поэтому снижается: ● скорость метаболических путей образования нуклеотидов; ● синтез ДНК и рост опухоли. ПРОТИВООПУХОЛЕВЫЕ препараты могут оказывать свое действие, нарушая структуру ДНК матрицы, так как: ● специфически связываются с ДНК; ● нарушают процесс репликации и транскрипции; ● снижают скорость деления опухолевых клеток. Эти препараты не нарушают метаболических процессов в покоящихся клетках. Но они оказывают влияние и на нормальные (здоровые) клетки быстроделящихся тканей, например: ● иммунной системы; 8 ● стволовые клетки кроветворной системы; ● эпителиальной ткани; ● фолликулы волос. Степень поражения выше у опухолевых клеток, так как: ● они более проницаемы для лекарственных препаратов; ● для них характерен активный метаболизм (рост, деление). ВИРУСНЫЕ ИНФЕКЦИИ – ИНГИБИТОРЫ МАТРИЧНЫХ БИОСИНТЕЗОВ После попадания вируса в клетку, белок-синтезирующий аппарат клеток почти полностью переключается на синтез белков вируса. ИНТЕРФЕРОНЫ – это белки, регулирующие реакцию клетки на вирусную инфекцию. Механизм действия интерферонов При заражении клетки активируется размножение вируса. Инфицированная клетка одновременно с этим начинает продукцию белка интерферона, который выходит из клетки и оказывает влияние на клетки, соседние с инфицированной (готовит клетку к вирусной «атаке»). Интерфероны являются сигналом для зараженных клеток, чтобы они не репродуцировали вирус, а для здоровых клеток, чтобы они не допускали его ядро. В ответ на воздействие интерферона клетки вырабатывают большое количество ферментов рибонуклеазы и протеинкиназы R. Рибонуклеаза (РНК-гидролаза) расщепляет клеточные РНК. Протеинкиназа R фосфорилирует и инактивирует фактор инициации трансляции IF2. В результате синтез белка в клетке снижается. 9 РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ В ЭУКАРИОТИЧЕСКИХ КЛЕТКАХ Одним из основных путей адаптации организмов к изменяющимся условиям окружающей среды является регуляция на стадии транскрипции. Индукторами или корепрессорами у эукариотов служат S (субстраты), Р (продукты) метаболических путей, гормоны. Рассмотрим на примере, действия стероидного гормона кортизола. ● гормон проходит через клеточную мембрану; ● гормон взаимодействует с цитоплазматическим рецептором; ● комплекс гормон-рецептор проходит в ядро; ● образованный комплекс взаимодействует с гормон-зависимой регуляторной зоной – энхансером или сайленсером; ● повышается или уменьшается сродство промотора к РНК-полимеразе; ● РНК-полимераза взаимодействует или не взаимодействует с промотором; ● возрастает или снижается скорость транскрипции структурных генов; ● увеличивается или уменьшается количество белков в клетке. 10 Более подробно «включение» транскрипции при прохождении индуктора (например, гормона) рассмотрено на следующем рисунке. 11 Но воздействие индукторов и корепрессоров явление временное, оно может меняться в зависимости от состояния клетки и действия на нее регуляторных факторов.. СТОЙКАЯ, СОХРАНЯЮЩАЯСЯ НА ПРОТЯЖЕНИИ ВСЕЙ ЖИЗНИ КЛЕТКИ ИНДУКЦИЯ ИЛИ РЕПРЕССИЯ. Регуляторные факторы вызывающие стойкую репрессию (препятствующие транскрипции): ● белки-гистоны; ● фермент ДНК-метилаза, присоединяющий СН3-группы к остаткам дезоксицитидина. Эта модификация меняет конформацию хроматина и препятствует транскрипции; ● белки, вызывающие конденсацию ДНК. 12 Все эти факторы препятствуют транскрипции определенных генов хромосом. В клетках разных тканей стойко репрессированы различные гены, поэтому разные клетки одного организма различаются по белковому составу и, следовательно, по обмену веществ и функциям. Образование зон стойкой репрессии происходит на стадии дифференцировки клетки. МЕХАНИЗМЫ ГЕНЕТИЧЕСКОЙ ИМЕНЧИВОСТИ. ПОЛИМОРФИЗМ БЕЛКОВ. НАСЛЕДСТВЕННЫЕ БОЛЕЗНИ. Нерепарированные изменения в первичной структуре ДНК называют мутациями. ГЕННЫЕ МУТАЦИИ – это изменения в нуклеотидной последовательности гена. Молекулярные механизмы генных мутаций. Рассмотрим на примерах. ● Замена нуклеотида а) без изменения смысла кодона (так как код вырожден) ДНК РНК аминокислота в белке АCА UGU цистеин АCG UGC цистеин Изменений в структуре белка нет. б) с изменением смысла кодона ДНК РНК аминокислота в белке АCА UGU цистеин АCC UGG триптофан В первичной структуре белка вместо аминокислоты цис появляется триптофан, происходят изменения в первичной структуре белка, а значит, возможно и снижение его активности. в) с изменением смысла и образованием терминирующего кодона ДНК РНК аминокислота в белке АСА UGU цистеин АСТ UGА терминирующий (стоп) кодон Образование стоп кодона приводит к нарушению синтеза и образованию неполного (незавершенного) белка. 13 Пример: В яйцеклетке, в гене кодирующем строение фермента диоксигеназы гомогентезиновой кислоты произошла замена 7-го нуклеотида с образованием терминирующего кодона. В ходе трансляции этого гена образуется дипептид, который не проявляет ферментативной активности. Отсутствие фермента диоксигеназы гомогентезиновой кислоты приводит к заболеванию – алкаптонурия (нарушение катаболизма тирозина). ● Вставка нуклеотидов а) кратная трем -ААА-GАG-CТА- ДНК -ААА-GАG-CCТ-CТА- ДНК -UUU-CUC-GАU- РНК -UUU-CUC-GGА-GАU- РНК -Фен- Лей- Асп- белок -Фен- Лей- Глу- Асп- белок Вставка нуклеотидов, кратная трем, приводит к изменению 1-ой структуры белка и его удлинению на 1, 2, или несколько аминокислот. Активность белка, как правило, снижается. б) не кратная трем (1, 2, 4, ) -ААА-GАG-CТА- ДНК -ААА-Т-GАG-CТА- ДНК -UUU-CUC-GАU- РНК -UUU-АCU-CGА-U- РНК -Фен- Лей- Асп- белок -Фен-Тре - Арг-белок Вставка нуклеотидов, не кратная трем, приводит к сдвигу рамки считывания, и образованию после точки мутации бессмысленной последовательности аминокислот. Происходит нарушение 1-ой, 2-ой, 3-ой структур белка и потеря активности. ● Выпадение нуклеотидов (делеция) а) кратная трем Выпадение нуклеотидов, кратное трем, приводит к укорочению белка на 1, 2, или несколько аминокислот, изменению его конформации и активности. б) не кратная трем Выпадение нуклеотидов, не кратное трем, приводит к сдвигу рамки считывания, и образованию после точки мутации бессмысленной последовательности аминокислот. Происходит нарушение 1-ой, 2-ой, 3-ой структур белка и потеря, активности. 14 Возможные причины мутаций: ● Ошибки репликации ● Повреждения ДНК под действием УФ, ● Повреждение ДНК ионизирующей радиацией. ●Повреждение ДНК химическими соединениями окружающей среды. Возможные биологические последствия "вредных" мутаций: ● Предрасположенность к некоторым болезням ● Возникновение наследственных болезней ● Непереносимость некоторых лекарственных препаратов и пищевых веществ ● Трансформация клеток и развитие опухолей. Мутации в соматических клетках не наследуются, но могут вызывать различные заболевания. Мутации в половых клетках передаются по наследству и проявляются как наследственная болезнь, связанная с малой активностью или полным отсутствием активности того или иного белка (фермента). ПОЛИМОРФИЗМ БЕЛКОВ Полиморфные белки – это множественные молекулярные формы белка в пределах организмов одного биологического вида. Полиморфные белки – это результат мутаций в копиях генов и рекомбинаций в процессе мейоза. Полиморфные белки: ● могут иметь незначительное различие в аминокислотном составе; ● имеют близкую пространственную структуру; ● выполняют одну и ту же или близкие функции; ● в популяции могут быть представлены большим числом вариантов; ● каждый индивид имеет не более 2-х вариантов этого белка. ПРИМЕРЫ ПОЛИМОРФНЫХ БЕЛКОВ Гемоглобины человека представлены группой полиморфных белков, например: НbA(2α2β); HbS(2α2β*); 15 HbC(2α2β**), которые имеют различие в первичной структуре β-цепи Гены α и β-цепей НbA, HbS, HbC аллельны, т.е. на хромосоме они занимают один и тот же локус. Но в 6 положении β-цепей НbA, HbS, HbC находятся разные аминокислотные остатки. Это результат мутации – замена нуклеотида. ~ Три – Про – Глу – Глу – Лиз ~ 4 5 6 7 β-цепь HbА 8 ~ Три – Про – Вал – Глу – Лиз ~ β-цепь HbS 4 5 6 7 8 В результате замены Глу на Вал на поверхности молекул Hb: ● появляются неполярные участи; ● происходит слипание молекул Нb; ● изменяется форма эритроцита (форма серпа); ● уменьшается продолжительность его жизни; ● снижается уровень Нb в крови и доставка О2 в ткани. Гомозиготные носители HbS (серповидно-клеточная анемия) умирают в раннем возрасте. Гетерозиготы практически здоровы (в редких случаях –легкая форма анемии). Выявить гетерозиготных носителей можно, используя методы молекулярной диагностики. ДНК – ДИАГНОСТИКА (МОЛЕКУЛЯРНАЯ ДИАГНОСТИКА) С помощью этого метода можно: ● выявить мутации; ● установить включение вирусной или бактериальной ДНК в состав ДНК-человека; ● проводить пренатальную диагностику наследственных болезней; ● выявлять гетерозиготных носителей дефектных генов; ● идентифицировать ДНК для: установления личности установления родства. 16 Для проведения этого метода выделяют ДНК из: ● клеток крови (лейкоцитов) ● гистологических срезов ● биоптата ткани ● мочи ● слюны Содержание ДНК в клетках невелико, а исследуемый ген (фрагмента гена) может составлять 1/1 000 000 всей ДНК клетки. Поэтому для исследования количество изучаемого гена или фрагмента гена необходимо увеличить в несколько сот раз. ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ (ПЦР) С помощью ПЦР можно синтезировать in vitro небольшие участки ДНК длиной от нескольких десятков до нескольких сотен пар нуклеотидов. РЕАКЦИОННАЯ СМЕСЬ ДЛЯ ПЦР включает: 1. ДНК выделенную из образца; 2. Субстраты – дАТФ, дГТФ, дЦТФ, дТТФ; 3. Термостабильную Taq-полимеразу; 4. Буфер для поддержания рНопт, содержащий ионы Mg2+; 5. 2 праймера. Праймеры синтезируются в лаборатории и представляют собой фрагменты одноцепочечной ДНК из 20-30 дезоксирибонуклеотидов. Один праймер должен быть комплементарен 3′-концу одной цепи ДНК. Второй праймер должен быть комплементарен 3΄-концу другой цепи ДНК. ПЦР проходит в три этапа: 1. Денатурация при 90-95˚С. Водородные связи между цепями ДНК разрываются и цепи расходятся. 2. Гибридизация (50-60˚С) цепей ДНК с праймерами и образование между ними водородных связей. 17 3. Элонгация (полимеризация), Taq-полимераза присоединяется к 2-х цепочечному участку ДНК и по принципу комплементарности удлиняет праймер от 5΄ к 3΄ концу. Этот 3-х этапный цикл повторяется много раз (задает экспериментатор), пока не образуется достаточно материала, необходимого для дальнейших исследований, которые обязательно включают электрофорез. 18 Биологические мембраны Мембраны – это биологические структуры, которыми окружены все клетки, а также различные клеточные органеллы. Мембраны ответственны за выполнение многих важнейших функций клетки. Основные функции мембран: 1) Отделяют клетки от окружающей среды и делят ее на компартменты (отсеки); 2) регулируют транспорт веществ в клетку и органеллы или в обратном направлении; 3) обеспечивают специфику межклеточных контактов; 4) воспринимают, усиливают и передают внутрь клетки сигналы из внешней среды. Основные типы биологических мембран: 1) плазматическая 2) ядерная 3) эндоплазматический ретикулум (ЭР) 4) мембрана аппарата Гольджи 5) митохондриальная мембрана 6) мембрана лизосом Строение и состав мембран: Биологические мембраны построены из липидов и белков, связанных друг с другом с помощью нековалентных взаимодействий. Основу мембран составляет двойной липидный слой, в формировании которого участвуют фосфолипиды, гликолипиды и холестерин. Поперечный разрез плазматической мембраны Липиды мембран Липиды мембран амфифильны, т.е. в молекуле одновременно есть гидрофильные группы (полярные «головки») и гидрофобные «хвосты» (алифатические радикалы), самопроизвольно формирующие бислой. В мембранах присутствуют липиды 3 главных типов: 1) Фосфолипиды; 2) Гликолипиды; 3) Холестерол 1) Фосфолипиды глицерофосфолипиды Производные фосфатидной кислоты сфингофосфолипиды (сфингомиелины) Производные N-ацилсфингозина (церамида) 2) Гликолипиды Углеводные производные N-ацилсфингозина (церамида) Цереброзиды Углеводный компонент моно- или олигосахарид Ганглиозиды Углеводный компонент разветвленный олигосахарид 3) Холестерол Содержится в мембранах клеток всех животных, придает мембранам жесткость и снижает жидкостность (текучесть) их гидрофобного слоя. Функции липидов мембран: 1) формируют двойной липидный слой — структурную основу мембран; 2) обеспечивают необходимую для функционирования мембранных белков среду; 3) участвуют в регуляции активности ферментов; 4) служат «якорем» (местом прикрепления) для поверхностных белков; 5) участвуют в передаче гормональных сигналов. Белки мембран Белки отвечают за функциональную активность мембран Функции белков мембран 1) избирательный транспорт веществ; 2) передача гормонального сигнала; 3) участие в иммунных реакциях; 4) участие в качестве ферментов; 5) участие во взаимодействии клеток друг с другом, обеспечивая образование тканей и органов. Белки мембран различаются по своему положению в мембране. Белки мембран интегральные (трансмембранные) белки полностью или частично погружены в мембрану поверхностные (периферические) белки располагаются на поверхности мембраны Интегральные белки содержат много гидрофобных аминокислот, с помощью которых они взаимодействуют с гидрофобной областью липидного бислоя мембраны. Участки белка, которые находятся в области полярных «головок», содержат много гидрофильных остатков аминокислот. Поверхностные белки разными способами прикрепляются к мембране: 3 – связанные с интегральными белками; 4 – присоединенные к полярным «головкам» липидного слоя; 5 – «заякоренные» в мембране с помощью короткого гидрофобного концевого домена; 6 – «заякоренные» в мембране с помощью ковалентно связанного остатка жирной кислоты. Транспорт веществ через мембрану Одна из главных функций мембран — регуляция переноса веществ. Пример: плазматическая мембрана должна впустить в клетку и удержать вещества, которые нужны клетке, и освободиться от ненужных. Типы транспорта активный пассивный перенос веществ по градиенту концентрации перенос веществ против градиента концентрации первично-активный транспорт с затратой энергии АТФ при участии транспортных АТФ-аз H+-АТФ-аза вторично-активный транспорт за счет энергии градиента концентрации другого переносимого вещества Ca2+-АТФ-аза Na+,K+-АТФ-аза активный симпорт активный антипорт Пассивный транспорт Наружная поверхность мембраны Внутренняя поверхность мембраны Простая диффузия – без участия белков-переносчиков (O2, NH3, СO2 и др.) ( ). Облегченная диффузия – при участии белков-переносчиков (Глюкозы) ( ). Пассивный симпорт: перенос 2-х ионов по градиенту концентрации в одном направлении. Например, HPO42- ( ) и Н+ ( ). Пассивный антипорт: перенос 2-х ионов по градиенту концентрации в противоположных направлениях. Например, НСО3- ( ) и Cl- ( ). Активный транспорт Пример: Na+-K+-АТФ-аза транспорт): (первично-активный Концентрация ионов Na+ снаружи клетки всегда больше, чем внутри. Это обеспечивается благодаря работе Na+-K+-АТФ-азы (Na+-K+-насоса): 1) 3 иона Na+ присоединяются к Na+-K+-АТФ-азе в центрах, обращенных в цитозоль => меняется заряд и конформация фермента; 2) Повышается сродство фермента к АТФ и активируется реакция аутофосфорилирования по OH-группе Сер или Тре; 3) Присоединение фосфатной группы к ферменту, приводит к изменению его конформации и Na+-K+АТФ-аза закрывается с внутренней стороны мембраны и открывается с наружной; 4) Снижается сродство центров связывания + фермента к ионам Na и они отделяются от фермента, оказываясь снаружи клетки; 5) С наружной стороны к ферменту присоединяются меняется заряд и 2 иона K+ => снова конформация фермента и активируется реакция аутодефосфорилирования; 6) Na+-K+-АТФ-аза теряет фосфатную группу и сродство к ионам K+, которые оказываются внутри клетки; 7) Изменяется конформация фермента и Na+-K+АТФ-аза возвращается в исходное состояние. Вторично-активный транспорт: Происходит за счет затрат энергии градиента концентрации одного из переносимых веществ. 1) Активный симпорт: Перенос одновременно 2-х веществ в одном направлении. Одно из веществ (в-во Y ) перемещается против градиента концентрации за счет перемещения другого в-ва (в-во X ) по градиенту концентрации. Например: Na+ ( глюкозы ( ). ) – зависимый транспорт 2) Активный антипорт: Перенос одновременно 2-х противоположных направлениях. Например: Na+ ( Ca2+ ( ). веществ в ) – зависимый переносчик ионов Трансмембранная передача сигнала (трансдукция сигнала через мембрану) Важное свойство мембран — способность воспринимать и передавать внутрь клетки сигналы из внешней среды. Внеклеточные химические сигналы Гормоны Нейромедиаторы Эйкозаноиды «Узнавание» сигнальных молекул помощью белков-рецепторов. Клетку-мишень определяют по избирательно связывать данную молекулу с помощью рецептора. другие сигнальные молекулы происходит с способности сигнальную Рецепторы (R) могут быть: 1) встроены в мембрану клеток-мишеней; 2) находиться в клетке (в цитозоле или в ядре). Гормоны — это молекулы, которые вырабатываются специализированными клетками, секретируются в кровь в ответ на изменение какого-либо специфического параметра внутренней среды организма и оказывают влияние на метаболизм и функциональное состояние клеток-мишеней. Классификация гормонов по механизму передачи сигнала Гормоны взаимодействующие с мембранными рецепторами Гидрофильные по химической природе адреналин глюкагон передающие сигнал через внутриклеточные рецепторы Гидрофобные по химической природе стероидные гормоны тиреоидные гормоны инсулин Передача сигнала с помощью мембранных рецепторов. Примеры мембранных рецепторов: 1) рецепторы, сопряженные с G-белками; 2) каталитические рецепторы Взаимодействие гормонов с рецепторами, сопряженными с G-белками, приводит к активации аденилатциклазной или инозитолфосфатной регуляторных систем. Аденилатциклазная система Аденилатциклазная система (АЦ-система) включает 5 мембранных белков: Интегральные белки цитоплазматической мембраны: 1)Rs – рецептор активатора аденилатциклазной системы; 2)Ri – рецептор ингибитора аденилатциклазной системы; 3)Фермент аденилатциклаза. «Заякоренные» белки: 4)Gs - ГТФ-связываюший белок, состоит из αsβγсубъединиц; 5)Gi — ГТФ-связываюший белок, состоит из αiβγсубъединиц. В АЦ-систему также входит один фермент – протеинкиназа А (ПКА). цитозольный С G-белками в состоянии покоя связана молекула ГДФ. Последовательность событий, приводящая к активации АЦ-системы: 1) Гормон присоединяется к рецептору (Rs) => => меняется конформация рецептора и происходит контакт рецептора с αsсубъединицей Gs-белка. 2) Это приводит к изменению конформации αsсубъединицы, которая теряет сродство к ГДФ, αsно приобретает сродство к ГТФ. => От субъединицы отсоединяется ГДФ, а присоединяется ГТФ. 3) В ходе присоединения ГТФ, снова меняется конформация αs-субъединицы => происходит отсоединение комплекса αsсубъединицы-ГТФ от β- и γ-субъединиц. 4) αs-ГТФ перемещается в липидном слое с помощью латеральной диффузии и присоединяется к аденилатциклазе (АЦ). 5) Присоединение αs-ГТФ вызывает изменение конформации АЦ => происходит активация АЦ и она начинает катализировать реакцию превращения АТФ в цАМФ. В клетке повышается концентрация цАМФ – вторичного вестника гормонального сигнала. 6) цАМФ активирует протеинкиназу A (ПКA), отсоединяя от неё регуляторные субъединицы. 7) Активная ПКA при участии АТФ фосфорилирует специфические белки клетки по OH-группе Сер и Тре. 8) Фосфорилирование специфических белков и ферментов повышает или понижает их => изменяется скорость активность метаболических процессов, в которых участвуют эти ферменты. Инактивация АЦ-системы: 1) При присоединении αs-ГТФ к АЦ меняется конформация αs-субъединицы и происходит ее активация. 2) => Далее αs-субъединица с медленной скоростью дефосфорилирует ГТФ, то есть проявляет ГТФ-фосфатазную (ГТФазную) активность. 3) Через некоторое время, в активном центре αsсубъединицы остается молекула ГДФ (αs-ГДФ). 4) => Изменяется конформация αs-субъединицы, она теряет сродство к АЦ и отсоединяется от нее. 5) αs-ГДФ взаимодействует с субъединицами β•γ и Gs-белок возвращается к неактивной форме αsβγ-ГДФ. 6) Фосфодиэстераза (ФДЭ) превращает цАМФ в АМФ => снижается концентрация цАМФ в клетке и активность протеинкиназы А (ПКА) скорость фосфорилирования 7) Замедляется ферментов, изменяется их активность и скорость метаболических процессов в которых они участвуют. 8) Фосфорилированные ферменты (и белки) под действием фосфопротеинфосфатаз переходят в дефосфорилированную форму. => меняется их конформация, активность и скорость метаболических процессов, в которых участвуют эти ферменты. В результате, АЦ-система переходит в исходное неактивное состояние. Взаимодействие сигнальной молекулы с Ri: При присоединении гормона (или другой сигнальной молекулы) к Ri (ингибирующему рецептору) АЦсистемы происходит та же последовательность событий, что и при присоединении к Rs, но αi-ГТФ при взаимодействии с АЦ подавляет ее активность. => Снижается образование цАМФ и активность протеинкиназы А. АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА (подробная): (идеально подходит для устного ответа) Схемы для подготовки к тестированию: Инозитофосфатная система Инозитолфосфатная система (ИФ-система) включает 3 мембранных белка: R — рецептор активатора инозитолфосфатной системы — интегральный белок; Фермент фосфолипаза C (ФЛС) — поверхностный белок; Gplc — ГТФ-связывающий белок, состоит из αplcβγсубъединиц — «заякоренный» белок. В ИФ-систему также входят: белок кальмодулин, фермент протеинкиназа C (ПКС), регулируемые Ca2+каналы мембраны эндоплазматического ретикулума (ЭР), Са2+-АТФ-аза клеточной и митохондриальной мембран. Последовательность событий, приводящая к активации ИФ-системы: ………………………………………. 1) – 3) см. АЦ-систему. 4) αplc-ГТФ (αФЛС-ГТФ) перемещается в липидном слое с помощью латеральной диффузии и присоединяется к ФЛС. 5) Присоединение αplc-ГТФ вызывает изменение конформации ФЛС => происходит активация ФЛС и она начинает катализировать реакцию гидролиза фосфатидилинозитол-4,5-бифосфата до инозитол-3-фосфата (ИФ3) и (ФИФ2) диацилглицерола (ДАГ). 6) ДАГ остается в мембране и участвует в активации фермента (ПКС). ИФ3 уходит в цитозоль. Т.е. сигнал принятый рецептором – раздваивается. 7) ИФ3 связывается специфическими центрами Са2+канала мембраны ЭР => меняется конформация Са2+-каналов и они открываются. Ионы Са2+ по градиенту концентрации поступают в цитозоль. В отсутствие ИФ3 в цитозоле канал закрыт. Ионы Са2+ оказывают 2 действия: 1. Активация Са2+-кальмодулинзависимых ферментов: 1) Ионы Са2+ присоединяются к небольшому цитозольному белку кальмодулину, на котором есть 4 центра связывания с ионами Са2+ => образуется 2+ комплекс Кальмодулин 4Ca . . . 2) Комплекс Кальмодулин 4Ca2+присоединяется к Ca2+-кальмодулинзависимым протеинкиназам => повышается скорость фосфорилирования определенных белков и ферментов по OH-группе Сер и Тре, что меняет их конформацию и повышает или скорость понижает их активность => изменяется определенных метаболических процессов. 2. Активация протеинкиназы C: 1) Ионы Са2+ соединяются с цитозольным ферментом ПКC => меняется его конформация. 2) У ПКC, связанной с ионами Са2+, образуются 2 центра: один – комплементарен фосфатидилсерину (ФС), а второй – ДАГ. 3) ПКC присоединяется к мембране на её внутренней стороне 2+ ФС ДАГ . => образуется комплекс ПКC Ca Это активная форма фермента ПКC, которая фосфорилирует специфические белки по Сер и Тре. . . . Каталитические рецепторы Каталитические рецепторы – это рецепторы гормонов или других сигнальных молекул, которые одновременно являются ферментами. Примеры: 1) Рецептор инсулина – тирозиновая протеинкиназа (ТПК). ТПК – это ферменты, которые в активной форме фосфорилируют специфические белки по OH-группе Тир. Строение инсулина: и функционирование рецептора Рецептор инсулина состоит из 2-х α- и 2-х βсубъединиц. α-субъединицы расположены на внутренней поверхности мембраны, β-субъединицы пронизывают мембранный бислой. Центр связывания инсулина образован N-концевыми доменами α-субъединиц. Каталитический центр рецептора находится на внутриклеточных доменах β-субъединиц. Цитозольная часть рецептора содержит несколько остатков Тир. Присоединение образованный кооперативные субъединиц. инсулина в центр связывания, α-субъединицами, вызывает изменения конформации всех Изменение конформации β-субъединиц повышает их каталитическую (тирозинкиназную) активность. Происходит аутофосфорилирование β-субъединиц по нескольким остаткам тирозина. β-субъединиц Фосфорилирование изменению заряда, конформации специфичности фермента ТПК. приводит к и субстратной ТПК фосфорилирует определенные клеточные белки по OH-группам Тир. Эти белки получили название субстратов рецептора инсулина (IRS). IRS-белки участвуют в активации: фосфопротеинфосфатазы (ФПФ), фосфодиэстеразы (ФДЭ), ГЛЮТ-4 – переносчиков глюкозы в инсулинзависимые ткани (увеличивают поступление глюкозы в клетки мышц и жировой ткани), регуляторных белков ядра (повышают или понижают экспрессию генов определенных ферментов). 2) Гуанилатциклазная система. Мембранно-связанная Гуанилатциклаза трансмембранный гликопротеин. (ГЦ) – Одним из первичных мессенджеров протеинкиназы G является предсердный натриуретический фактор (ПНФ), регулирующий гомеостаз жидкости в организме. Энергетический обмен Взаимосвязь обмена веществ и энергии. Обмен веществ включает три этапа: 1) поступление веществ в организм; 2) метаболизм, или промежуточный обмен; 3) выделение конечных продуктов обмена. Вещества поступают в организм в результате дыхания (кислород), питания и пищеварения. Основные вещества, поступающие с пищей – это макромолекулы: – полисахариды; – жиры; – белки; В ходе пищеварения макромолекулы расщепляются на более мелкие молекулы (глюкоза, жирные кислоты, глицерол, аминокислоты). В клетках организма эти вещества подвергаются превращениям, включаясь в метаболизм (обмен веществ). Метаболизм – это взаимосвязь химических процессов, происходящих в организме. Метаболизм Катаболизм Анаболизм процесс распада органических веществ до конечных продуктов (CO2, H2O и мочевина) с выделением энергии (~ 40-50% в виде АТФ) процесс синтеза сложных органических веществ (в основном макромолекул) с использованием энергии, выделяемой в реакциях катаболизма Пищевые вещества (полисахариды, жиры, белки) ПИЩЕВАРЕНИЕ Глюкоза, Глицерол, Жирные к-ты, Аминок-ты КАТАБОЛИЗМ CO2, H2O, Мочевина АДФ + H3PO4 АТФ АНАБОЛИЗМ Гликоген, Жиры, Белки, ДНК, РНК Катаболизм Аэробный Анаэробный происходит при участии O2 происходит без участия O2 Вся энергия необходимая для человека образуется в реакциях катаболизма. В организме катаболизм. человека преобладает аэробный В ходе аэробного катаболизма образуется до 95% всей энергии необходимой человеку. Аэробный катаболизм веществ в тканях сопровождается потреблением O2 и выделением CO2. Окисление органических веществ в организме кислородом (воздуха) с образованием воды и CO2 называется тканевым дыханием. 3 этапа аэробного катаболизма (тканевого дыхания): 1) Специфические (частные) пути катаболизма. В ходе них образуется одно из 2-х веществ: – пируват; – Ацетил-KoA. Примеры: гликолиз, β-окисление жирных кислот и др. 2) Общий путь катаболизма. Включает 2 стадии: а) окислительное декарбоксилирование пирувата; б) цикл Кребса. 3) Цепь переноса электронов (ЦПЭ) и сопряженное с ней окислительное фосфорилирование. Энергия, которая выделяется при тканевом дыхании, используется для функционирования клеток. Например, окисление 1 моль глюкозы происходит с выделением большого кол-ва энергии: ЦПЭ * C6H12O + 6O 2 .6 * 6CO + 6H O 2 + 2780 кДж/моль .2 Специфические и общий пути катаболизма ~ 40-50% энергии окисляющихся веществ используется клетками для синтеза АТФ из АДФ и H3PO4: АДФ + H3PO4 → АТФ + H2O ! Реакция идет с затратой большого кол-ва энергии. Для синтеза АТФ используется 2 источника энергии: окислительное и субстратное фосфорилирование. 1) Окислительное фосфорилирование происходит за счет энергии переноса электронов от органических веществ к O2. ! Может происходить только в аэробных условиях. Этим способом образуется ~ 95% всего АТФ в организме. 2) Субстратное фосфорилирование происходит за счет энергии макроэргических связей некоторых соединений. Например: 1,3-бифосфоглицерат, фосфоенолпируват, сукцинил-KoA, креатинфосфат и др. S – O ~ PO3H2 Фермент (киназа) АДФ S – OH АТФ Может происходить как в матриксе митохондрий, так и в цитоплазме клеток независимо от присутствия O2. ! Это вспомогательный путь синтеза АТФ в организме. (образуется ~ 5 % от всего АТФ организма) Содержание АТФ в организме человека составляет всего ~ 50 г. Т.к. клетки не способны накапливать АТФ, а расход энергии происходит постоянно, в организме также постоянно идет синтез АТФ из АДФ и Н3РO4. За сутки в организме человека может синтезироваться до 60 кг АТФ (столько же и расходуется). Фосфорилирование АДФ и последующее использование АТФ в качестве источника энергии образуют циклический процесс (цикл АДФ-АТФ): Цикл АТФ-АДФ: Катаболизм веществ Энергия АДФ + H3PO4 АТФ + H2O Энергия Анаболизм Активный транспорт Сокращение мышц Продукция тепла Цикл АДФ-АТФ работает постоянно. В ходе него образуется то кол-во АТФ, которое было израсходовано клеткой. Образующийся нуклеотид АТФ имеет две макроэргические связи (~) и используется в клетках как универсальный источник энергии для синтеза веществ и других видов работы (активный транспорт веществ через мембраны, мышечное сокращение и т.д.). Цепь переноса электронов (ЦПЭ). Цепь переноса электронов (дыхательная цепь) – это сложная система переносчиков, при участии которой происходит процесс последовательного переноса электронов от NADH и FADH2 на O2. Дыхательной цепи предшествует отнятие атомов водорода от окисляемых субстратов (реакции дегидрирования). Эти реакции относятся восстановительным. к Их катализируют ферменты (подкласс: дегидрогеназы). окислительнооксидоредуктазы Бóльшая часть окислительно-восстановительных реакций в клетке происходит в матриксе митохондрий. Реакции дегидрирования происходят специфических и общего пути катаболизма. в ходе Выделяют 2 типа дегидрогеназ, катализирующие реакции дегидрирования, предшествующие ЦПЭ: 1. NAD+ – зависимые (предшественник: витамин PP); дегидрогеназы ! Субстратов NAD+ – зависимых дегидрогеназ большинство. Примеры: малат, изоцитрат, глутаминовая кислота и др. α-кетоглутарат, Кофермент NAD+ не образует прочной постоянной связи с ферментом. Он соединяется с АЦ фермента уже в ходе реакции и после её завершения уже в восстановленной форме отсоединяется. 2. FAD – зависимые дегидрогеназы (предшественник: витамин B2) Кофермент FAD прочно связан с активным центром фермента как в окисленной, так и в восстановленной форме. Включаясь в реакцию, он принимает 2H+ и 2 e- и переходит в восстановленную форму. Субстраты FAD-зависимых дегидрогеназ: сукцинат, ацил-KoA, глицерол-3-фосфат (α-глицерофосфат). Структурная организация цепи переноса электронов (ЦПЭ). В основе функционирования ЦПЭ лежит работа 5 основных компонентов, обеспечивающих перенос электронов от NADH и FADH2 на O2. В их число входит 3 сложных ферментных комплексов, а также 2 низкомолекулярных переносчика. Компоненты ЦПЭ: 1) NADH-дегидрогеназа (комплекс I); 2) сукцинатдегидрогеназа (комплекс II); 3) низкомолекулярный переносчик: убихинон (кофермент Q); 4) Убихинолдегидрогеназа (QН2-дегидрогеназа) (комплекс III); 5) низкомолекулярный переносчик (цитохром с); 6) цитохромоксидаза (комплекс IV) Все компоненты ЦПЭ располагаются во внутренней мембране митохондрий в порядке возрастания окислительно-восстановительных потенциалов (редокс-потенциалов). Самый низкий редокс-потенциал у NADH (-0,32В), а самый высокий редокс-потенциал у кислорода (+0,82В). Это обеспечивает последовательное перемещение электронов от NADH (или FADH2) на O2, при котором происходит выделение энергии на каждом этапе ЦПЭ. ЦПЭ обеспечивает взаимосвязь (сопряжение) процессов окисления и фосфорилирования АДФ. Основная роль в переносе электронов принадлежит 3 важнейшим ферментным комплексам (I, III, IV). Это сложные олигомерные белки, расположенные во внутренней мембране митохондрий. Они являются интегральными белками. Компонент ЦПЭ NADH-дегидрогеназа (I ферментный комплекс) Убихинон Окисл. Восст. форма: Q форма: QH2 QH2-дегидрогеназа (убихинолдегидрогеназа) III ферментный комплекс, включает цитохромы b и c1 Кофермент Окисл. Восст. форма форма FMN FMN·H2 Гем (Fe3+) Гем (Fe2+) Цитохром c Гем (Fe3+) Гем (Fe2+) Цитохромоксидаза IV ферментный комплекс, включает цитохромы a и a3 Гем (Fe3+), Cu2+ Гем (Fe2+) Cu+ Сукцинатдегидрогеназа (II ферментный комплекс) FAD FAD·H2 Донор ē Акцептор ē NADH + H+ Убихинон (Q) NADHQH2-дегидрогеназа дегидрогеназа (цит. b и c1) (I комплекс) (III комплекс) QH2 Цитохром c QH2Цитохромоксидаза дегидрогеназа (IV комплекс) (III комплекс) Цитохром c O2 Сукцинат Убихинон (Q) Этапы ЦПЭ: 1 этап: I ферментный комплекс: NADH-дегидрогеназа. Это крупный интегральный белок внутренней мембраны митохондрий, состоящий из большого числа протомеров. Коферментом NADH-дегидрогеназы служит Рабочая часть: рибофлавин (витамин B2). FMN. NADH-дегидрогеназа катализирует перенос 2 атомов H с кофермента (NADH + H+) на FMN, который переходит в форму FMNH2: NADH + H+ + E-FMN → NAD+ + E-FMNH2 Восстановление кофермента FMN повышает сродство NADH-дегидрогеназы к убихинону. Убихинон (кофермент Q10) – жирорастворимое витаминоподобное вещество. Убихинон (Q) способен восстанавливаться и превращаться в QH2 (убихинол), принимает от FMNH2 2 e- . Протоны (Н+) при этом поступают в межмембранное пространство. Необходимые для образования QH2 протоны при этом поступают из матрикса. (На этом этапе ЦПЭ происходит разделение e- и H+). Таким образом, в результате работы NADHдегидрогеназы происходит перенос 2 e- от NADH на убихинон. Сумм. у-е 1 этапа: NADH + Н+ + Q → NAD+ + QH2 ! При этом происходит перенос нескольких протонов из матрикса митохондрии. в межмембранное пространство 2 этап: III Ферментный комплекс QH2-дегидрогеназа (убихинолдегидрогеназа) – сложный олигомерный белок, включающий 2 протомера: цитохром b и цитохром с1. Цитохромы различаются по структуре полипептидных цепей. Рабочей частью всех цитохромов является гем, содержащий ион Fe3+, который может принимать e- и превращаться в Fe2+: Fe3+ + e → Fe2+. QH2-дегидрогеназа окисляет убихинол, электроны при этом последовательно поступают сначала на цитохром b, затем – на цитохром с1. Далее происходит последовательный перенос 2 электронов на небольшой белок-гемопротеин цитохром c. Сумм. у-е 2 этапа: QH2 + 2 цит. с (Fe3+) → Q + 2 цит. c (Fe2+) + 2Н+ ! За счет энергии реакции III комплекс, как и комплекс I переносит несколько H+ в межмембранное пространство митохондрий. 3 этап: IV Ферментный комплекс – цитохромоксидаза (цитохром-c-оксидаза). Это сложный олигомерный белок, включающий 2 протомера-гемопротеина: цитохром a и цитохром a3, каждый из которых содержит центр связывания с O2. Цитохромы a и a3 содержат гем (Fe3+), а также ионы Cu2+. Ионы Fe3+ гема, а также ионы Cu2+ участвуют в переносе электронов на O2: Fe3+ + e- → Fe2+ Cu2+ + e- → Cu+ Кислород, поступающий в митохондрии из крови, связывается с атомом Fe в геме цитохрома a3 в форме молекулы O2 (подобно тому, как он связывается с гемоглобином). Цитохромоксидаза имеет гораздо более низкую Км (т.е. более высокое сродство) к O2, чем Hb, и поэтому клетки «высасывают» O2 из крови. Электроны от цитохрома c последовательно 3+ присоединяются сначала к ионам Fe цитохромов a и a3, затем к ионам Cu2+ и, затем, попадают на O2. При этом также происходит присоединение к кислороду H+ из матрикса => образуется метаболическая вода: ½ O2 + 2 e- + 2H+ → H2O Таким способом в организме человека синтезируется ~ 400 мл метаболической воды в сутки. Сумм. у-е 3 этапа: 2 цит. с (Fe2+) + ½ O2 + 2 H+ → 2 цит. c (Fe3+) + Н2O ! Работа IV комплекса цитохромоксидазы также сопровождается переносом H+ из матрикса в межмембранное пр-во. Использование энергии ЦПЭ для синтеза АТФ. Хемиосмотическая теория П. Митчелла, которая объясняет механизмы сопряжения процессов биологического окисления и фосфорилирования АДФ: При движении от окисляемого субстрата к кислороду электроны теряют часть своей потенциальной энергии. Комплексы (I, III и IV) внутренней мембраны митохондрий используют эту энергию электронов для переноса H+ из матрикса в межмембранное пространство митохондрий против градиента концентрации. Т.е. комплексы ЦПЭ (I, III и IV) работают как протонные насосы, перекачивая H+ из матрикса в межмембранное пр-во митохондрий за счет энергии переноса электронов по ЦПЭ. В результате возникает градиент концентрации H+ и градиент «+» заряда – протонный электрохимический потенциал (ΔμH+). При накоплении в межмембранном пространстве достаточного кол-ва H+, происходит их присоединение к ферменту => изменяется заряд и конформация фермента. Это приводит к раскрытию протонного канала АТФсинтазы => H+ переносятся в матрикс по градиенту концентрации. Это приводит к активации поверхностной части фермента, кот. катализирует синтез АТФ: АДФ + H3PO4 → АТФ + H2O Было установлено, что при переносе электронов через поверхностную часть АТФ-синтазы происходит её вращение по кругу. И каждый поворот на 120ο приводит к синтезу 1 молекулы АТФ. ! Окислительное фосфорилирование – это способ синтеза АТФ из АДФ и неорганического фосфата с использованием энергии протонного электрохимического потенциала (ΔμH+), который образуется за счет переноса электронов по ЦПЭ. Количество АТФ, которое образуется в результате сопряжения работы ЦПЭ и АТФ-синтазы, оценивается с помощью коэффициента фосфорилирования. Коэффициент окислительного фосфорилирования (Р/О) показывает, какое количество неорганического фосфата затрачивается для фосфорилирования АДФ при использовании 1 атома кислорода на образование 1 молекулы воды. ! Коэффициент окислительного фосфорилирования численно равен количеству моль АТФ, синтезированных в результате окислительной реакции. Энергия электронов и протонов, перенесенных комплексами ЦПЭ (I, III и IV) от субстратов NAD+зависимых дегидрогеназ, достаточна для активации АТФ-синтазы и синтеза 3 моль АТФ в расчете на 1 моль дегидрируемого субстрата. Т.е. для всех субстратов NAD+-зависимых дегидрогеназ Р/О ≤ 3. Таким образом, дегидрирование 1 моль любого субстрата NAD+-зависимых дегидрогеназ сопряжено с синтезом 3 моль АТФ. При дегидрировании веществ с участием FADзависимых дегидрогеназ происходит передача протонов и электронов на убихинон без участия комплекса I: 1) DH2 + E-FAD → D + E-FADH2 2) E-FADH2 + Q → E-FAD + QH2 Основной FAD-зависимой дегидрогеназой явл-ся Сукцинатдегидрогеназа. Это поверхностный белок внутренней мембраны митохондрий (комплекс II). Сукцинатдегидрогеназа катализирует превращение сукцината в фумарат, являющееся одной из реакций цикла Кребса: Сукцинат + E-FAD → Фумарат + E-FADH2 В этом случае на пути электронов от субстрата на O2 только 2 участка переноса H+ в межмембранное пр-во митохондрий (комплексы III и IV) Таким образом, энергия электронов и протонов, поступивших в ЦПЭ от субстратов FAD-зависимых дегидрогеназ, достаточна для активации АТФ-синтазы и синтеза 2 моль АТФ в расчете на 1 моль дегидрируемого субстрата. Т.е. для субстратов FAD-зависимых дегидрогеназ (сукцинат, ацил-KoA, глицерол-3-фосфат): Р/О ≤ 2. Витамин C (аскорбиновая к-та) способна восстанавливать ионы Fe3+ цитохрома c. При этом Fe3+ принимает e- и переходит в Fe2+. На пути электронов от цитохрома c на O2 1 участок переноса H+ в межмембранное пр-во (IV комплекс). В этом случае Р/О ≤ 1. Коэффициент Р/О показывает только теоретический максимум синтеза АТФ. На самом деле, для синтеза АТФ используется ~ 4050% энергии ЦПЭ. Остальная часть энергии выделяется в виде тепла, а также используется для других видов работ клеток: Распределение энергии, выделяемой в ходе работы ЦПЭ Синтез АТФ ~ 40-50% Механическая работа ~ 20% Выработка тепла > 30% Итак, биологические функции ЦПЭ: 1) Использует энергию окисления субстратов для синтеза АТФ путём окислительного фосфорилирования; 2) Обеспечивает поддержание температуры тела человека. Дыхательный контроль Окисление субстратов и фосфорилирование АДФ в митохондриях прочно сопряжены. Скорость использования АТФ регулирует скорость потока электронов в ЦПЭ. Выполнение клеткой работы с затратой АТФ приводит к уменьшению концентрации АТФ. => Происходит накопление АДФ. Это активирует окисление субстратов и поглощение O2 митохондриями клетки. Таким образом, клетки реагируют на интенсивность метаболизма и поддерживают соотношение АТФ/АДФ на необходимом уровне. Дыхательный контроль – это зависимость интенсивности поглощения кислорода митохондриями от концентрации АДФ. Ингибиторы ЦПЭ. Ингибиторы ЦПЭ подавляют активность ферментных комплексов (I, II, III и IV) => происходит замедление или даже полное прекращение работы ЦПЭ => происходит замедление или полное прекращение синтеза АТФ. I комплекс: NADH-дегидрогеназа. Ингибиторы: ротенон и барбитураты аминобарбитал, нембутал, веронал и др.) (амитал, II комплекс: FAD-зависимая сукцинатдегидрогеназа. Обратимый конкурентный ингибитор: малонат. III комплекс: QH2-дегидрогеназа. Ингибитор: Антимицин A. IV комплекс: Цитохромоксидаза. Ингибиторы: цианид-ионы (CN–): KCN, HCN и др.; угарный газ (CO), сероводород (H2S). Антибиотик олигомицин не ингибирует саму ЦПЭ, но подавляет окислительное фосфорилирование, ингибируя АТФ-синтазу. ! При полном ингибировании in vitro любого из ферментных комплексов, который располагается на пути переноса электронов от дегидрируемого субстрата на O2, работа ЦПЭ прекращается и АТФ не синтезируется. Примеры: 1) К суспензии митохондрий, где в качестве окисляемого субстрата использовали малат, добавили амитал Na. Как при этом изменится синтез АТФ? Т.к. амитал Na ингибирует NADH-дегидрогеназу, которая расположена на пути переноса электронов от малата на O2, то скорость ЦПЭ замедляется => замедляется (или прекращается) синтез АТФ. 2) Смесь: малат + амитал + сукцинат. Р/О ≤ 2, т.к. в этом случае будет окисляться сукцинат, для которого «не нужна» NADH-дегидрогеназа. 3) Смесь: сукцинат + малонат (избыток). Синтез АТФ замедляется, т.к. малонат ингибирует сукцинатдегидрогеназу, которая располагается на пути электронов от сукцината на O2. 4) Смесь: сукцинат + малонат + изоцитрат. Р/О ≤ 3, т.к. в этом случае будет окисляться изоцитрат, для которого «не нужна» сукцинатдегидрогеназа. 5) Смесь: малат (или сукцинат) + антимицин A. Синтез АТФ замедляется, т.к. антимицин A ингибирует QH2-дегидрогеназу, которая располагается на пути электронов от этих субстратов на O2. 6) Если к 5) смеси добавить витамин C, то Р/О ≤ 1, так вит. C окисляет цитохром c (в переносе электронов не участвует QH2-дегидрогеназа). 7) CN– независимо от дегидрируемого субстрата необратимо ингибируют ЦПЭ у человека. Они присоединяются к Fe3+ цитохромоксидазы и полностью блокируют ЦПЭ. (CN– – необратимый специфический ингибитор цитохромоксидазы). Разобщение тканевого дыхания и окислительного фосфорилирования. Некоторые химические вещества (протонофоры) могут переносить H+ из межмембранного пространства митохондрии через внутреннюю мембрану в матрикс, минуя протонные каналы АТФ-синтазы => снижается (или даже полностью исчезает) ΔμH+ и замедляется (прекращается) синтез АТФ. Это явление называют разобщением тканевого дыхания и окислительного фосфорилирования. В результате разобщения количество АТФ снижается, а АДФ увеличивается. При этом, за счет дыхательного контроля, возрастает скорость поглощения O2 митохондриями и скорость работы ЦПЭ, но из-за нарушения работы АТФсинтазы коэффициент Р/О резко снижается => энергия рассеивается в виде теплоты, что приводит к повышению t° тела человека (пирогенное действие). Разобщители – это гидрофобные в-ва, которые легко проходят через мембрану митохондрий в обоих направлениях. РАЗОБЩИТЕЛИ Экзогенные 2,4-динитрофенол, дикумарол, стрептомицин Эндогенные жирные к-ты, билирубин, белок термогенин, тиреоидные гормоны Некоторые эндогенные разобщители играют роль в поддержании постоянной t° тела человека. При охлаждении стимулируется освобождение норадреналина из окончаний симпатических нервов. В результате происходят активация липазы в жировой ткани и мобилизация жира из жировых депо. Образующиеся свободные жирные кислоты служат не только «топливом», но и важнейшим регулятором разобщения дыхания и фосфорилирования. Общий путь катаболизма В пище человека практически не первичных доноров водорода для ЦПЭ. содержится В ходе переваривания пищевые макромолекулы (углеводы, жиры и белки) распадаются на мономеры, которые всасываются и поступают в клетки тканей. В клетках эти мономеры (моносахариды, глицерол, жирные к-ты, аминок-ты) вступают в специфические пути катаболизма и превращаются всего в 2 в-ва: пируват или ацетил-KoA. При этом происходит разнообразия в-в. значительное уменьшение В митохондриях клеток происходит распад пирувата до CO2 и H2O, который называется общий путь катаболизма. 2 стадии общего пути катаболизма (ОПК): 1) окислительное декарбоксилирование пирувата; 2) цикл Кребса. I. Окислительное декарбоксилирование пирувата. Процесс протекает в матриксе митохондрий. В ходе процесса из пирувата образуется ацетил-KoA: Суммарное у-е процесса: + CH3 C COOH + HSKoA + NAD O CH3 C O ПДК SKoA + NADH + H+ + CO2 _3 P/O < ЦПЭ Катаболизм основных пищевых веществ: Жиры Жирные кислоты Глицерол 2 1 Полисахариды Белки Моносахариды Аминокислоты 3 4 Пируват 5 6 Ацетил-KoA Цикл Кребса 7 CO2 и H2O 1-5: специфические пути катаболизма; 6, 7: общий путь катаболизма Превращение пирувата в ацетил-KoA – многостадийный процесс, который катализирует (ПДК), пируватдегидрогеназный комплекс расположенный на внутренней мембране митохондрии. Строение пируватдегидрогеназного комплекса: В комплекс входит 3 фермента и 5 коферментов: I фермент: Пируватдекарбоксилаза (E1). Кофермент: тиаминдифосфат (ТДФ). Витамин B1 (тиамин) – предшественник. II фермент: Дигидролипоил-ацетилтрансфераза (E2). Коферменты: липоевая к-та (ЛК) и кофермент A (HS-KoA). III фермент: Дигидролипоилдегидрогеназа (E3). Коферменты: FAD и NAD+. Коферменты ТДФ, ЛК и FAD явл-ся простетическими группами и прочно связаны с АЦ соответствующих ферментов. HS-KoA и NAD+ присоединяются к ферментам ПДК только во время реакции. Схема окислительного пирувата: декарбоксилирования Превращение пирувата в ацетил-KoA происходит в 5 стадий: 1) Субстраты фермента E1 – пируват и липоевая к-та (кофермент фермента E2). В ходе действия пируватдекарбоксилазы пируват присоединяется к ТДФ. (E1) От пирувата отщепляется COOH-группа в виде CO2, а оксиэтильная группа остается связанной с ТДФ. 2) Оксиэтильный окисляется. остаток, ( CH 3 CH OH) связанный с ТДФ, За счет этого восстанавливается S – S связь ЛК, а продукт окисления оксиэтила – ацетильный остаток переносится на дигидролипоат => образуется ацетиллипоат. Присоединение к липоевой к-те ацетильного остатка повышает сродство E2 к HS-KoA. 3) Фермент E2 переносит ацетильную группу на HS-KoA с образованием Ацетил-KoA восстановленной формы ЛК (дигидролипоата). и 4) При участии следующего фермента ПДК – E3 – происходит дегидрирование дигидролипоата при участии кофермента FAD (простетической группы E3). При этом происходит регенерация окисленной формы ЛК, способной участвовать в окислении следующей молекулы пирувата. 5) Полученный FADH2 дегидрируется с помощью NAD+ и образованием NADH + H+. NH2 + N CH2 N H3C N * S CH3 O O CH2 CH2 O P O P OH OH OH Тиаминдифосфат (ТДФ) Основной продукт реакции декарбоксилирования пирувата поступает в цикл Кребса. окислительного – Ацетил-KoA Образующийся в ходе реакции NADH + H+ вступает в ЦПЭ и способствует синтезу 3 моль АТФ в расчете на 1 моль пирувата путём окислительного фосфорилирования (P/O ≤ 3). II. Цикл Кребса. Цикл Кребса (цитратный цикл, цикл лимонной к-ты, цикл трикарбоновых к-т (ЦТК) ) – это совокупность 8 последовательных химических реакций, в ходе которых происходит распад Ацетил-KoA до 2-х молекул CO2 и образование доноров водорода для ЦПЭ (NADH + H+ и FADH2). Реакции цикла митохондрий. Кребса происходят в матриксе CH3 C ~ SKoA COOH Ацетил-KoA O C O HSKoA HO CH2 H 1. ЦитратCOOH H O синтаза Оксалоацетат 2 NADH + H+ _3 P/O < 8. МалатNAD + дегидрогеназа COOH COOH COOH CH2 C COOH C H COOH Цитрат H 2O 2а. CH2 C COOH CH COOH Аконитат H2 O COOH CH2 HC COOH 2б. HC OH COOH Изоцитрат Аконитаза ЦПЭ NAD + ~ HC OH Нуклеозид3. Изоцитратдегидрогеназа _3 CH2 дифосфаткиназа АДФ АТФ P/O < + COOH NADH + H _3 Малат P/O < CO2 ГТФ 7. Фумараза + ГДФ NADH + H _2 P/O < HSKoA H2 O HSKoA COOH COOH + CO H PO FADH FAD 2 3 4 COOH COOH 2 NAD CH2 CH2 CH CH2 5. Сукцинат- CH2 6. Сукцинат4. -кетоглутаратCH2 CH2 CH тиокиназа дегидрогеназа дегидрогеназный C O C O COOH COOH комплекс SKoA COOH Фумарат Сукцинат Сукцинил-KoA -кетоглутарат Энергетическая функция ОПК. В клетках ОПК является основным поставщиком первичных доноров водорода для ЦПЭ. В ходе ОПК происходит 5 реакций дегидрирования: Из них, 4 реакции дегидрирования происходит с участием NAD+-зависимых дегидрогеназ: Одна реакция – при окислительном декарбоксилировании пирувата. И ещё 3 реакции в цикле Кребса (3, 4 и 8). Каждая из этих реакций поставляет атомы водорода в ЦПЭ и сопряжена с синтезом 3 моль АТФ путем окислительного фосфорилирования в расчете на 1 моль дегидрируемого субстрата. В 6 реакции цикла Кребса участвует FAD-зависимая сукцинатдегидрогеназа. Она сопряжена с синтезом 2 моль АТФ на 1 моль сукцината. Т.е. всего в ОПК на 1 моль пирувата образуется: (4 • 3) + 2 = 14 моль АТФ путём окислительного фосфорилирования. Из них 3 моль АТФ образуется при окислительном декарбоксилировании пирувата и 11 моль АТФ в цикле Кребса. В 5 реакции цикла Кребса, за счет энергии разрыва макроэргической связи сукцинил-KoA, образуется ГТФ из ГДФ и H3PO4 (фермент: сукцинаттиокиназа). Далее энергия и фосфатный остаток ГТФ используется для получения АТФ под действием фермента нуклеозиддифосфаткиназы. Такой способ получения АТФ называется: субстратное фосфорилирование. ! Субстратное фосфорилирование – это способ синтеза АТФ из АДФ и неорганического фосфата с использованием энергии макроэргической связи субстрата. Таким образом, за счет реакции субстратного фосфорилирования (5 реакция цикла Кребса), в ОПК образуется ещё 1 моль АТФ (на 1 моль пирувата). Т.е., всего в ОПК (на 1 моль пирувата) образуется: 14 моль + 1 моль = 15 моль АТФ. Из них, 3 моль АТФ при окислительном декарбоксилировании пирувата и 12 моль АТФ в цикле Кребса. ! ОПК может функционировать только в аэробных условиях, т.к. реакции дегидрирования сопряжены с работой ЦПЭ и синтезом АТФ путём окислительного фосфорилирования. При торможении ЦПЭ также будет замедляться и скорость ОПК. Регуляция ОПК. Выделяют 4 основных типа регуляции ОПК: 1) Регуляция «энергетическим зарядом» клетки. Это соотношение АТФ/АДФ и NADH + H+/NAD+. Если в клетке концентрация АТФ и NADH + H+ – высокая (высокий «энергетический заряд» клетки), значит, клетка мало расходует энергию и => замедляются процессы, приводящие к получению АТФ (ОПК и ЦПЭ). Если в клетке концентрация АТФ и NADH + H+ – низкая (низкий «энергетический заряд» клетки), значит, клетка интенсивно расходует энергию и => ускоряются процессы, приводящие к получению АТФ (ОПК и ЦПЭ). Т.е. АТФ и NADH – ингибируют ОПК, а АДФ, АМФ и NAD+ – активируют ОПК. 2) Регуляция метаболитами. 3) Белок-белковые взаимодействия регуляторных ферментов ОПК Кальмодулин · 4Ca2+). (активация комплексом 4) Фосфорилирование / дефосфорилирование (характерно для ПДК) Регуляторные ферменты ОПК: ПДК Цитратсинтаза Активаторы (+) NAD+, АДФ, HS-KoA, Пируват, Ca2+ Оксалоацетат Изоцитратдегидрогеназа АДФ, Ca2+ Ca2+ α-кетоглутаратдегидрогеназный комплекс Ингибиторы (–) NADH + H+, АТФ, АцетилKoA NADH + H+, АТФ, Цитрат, Сукцинил-KoA, Жирные к-ты АМФ, NADH + H+, АТФ NADH + H+, АТФ, Сукцинил-KoA 1) Регуляция ПДК. Регуляторным компонентом ПДК является фермент E1. Это аллостерический фермент. Алл. активаторы: NAD+, HS-KoA Алл. ингибиторы: NADH и Ацетил-KoA В состав ПДК, кроме основных ферментов E1, E2 и E3 входит 2 регуляторные субъединицы – киназа ПДК (фосфорилирует фермент E1) и фосфатаза ПДК (дефосфорилирует фермент E1). Киназа ПДК – аллостерический фермент: – Активаторы: NADH + H+, АТФ, Ацетил-KoA – Ингибиторы: NAD+, HS-KoA, Пируват, АДФ Фосфатаза ПДК активируется ионами Ca2+. ! Фермент E1 активен в дефосфорилированной форме. Поэтому, ингибиторы киназы ПДК – это косвенные активаторы ПДК, а активаторы киназы ПДК – косвенные ингибиторы ПДК. 2) Регуляция цикла Кребса. В цикле Кребса 3 регуляторных фермента: Цитратсинтаза, Изоцитратдегидрогеназа и αкетоглутаратдегидрогеназный комплекс. Регуляторные ферменты ЦТК – аллостерические. Поэтому, основная регуляция аллостерическая регуляция. ЦТК – это Активность регуляторных ферментов ЦТК регулируется: 1) «Энергетическим зарядом» клетки. (Высокие концентрации АТФ и NADH + H+ ингибируют регуляторные ферменты цикла Кребса); 2) Регуляция промежуточными метаболитами ЦТК. Наиболее важные регуляторные ферменты ЦТК: Цитратсинтаза; Изоцитратдегидрогеназа медленную реакцию). (катализирует самую Анаболические функции цикла Кребса. Цикл Кребса может выполнять анаболические функции. в организме Это означает, что многие метаболиты цикла Кребса служат субстратами для синтеза многих в-в организма. Примеры: 1) Ацетил-KoA → используется на синтез жирных к-т и холестерина; 2) α-кетоглутарат → на синтез аминокислот (Глу, Глн, Про); 3) Сукцинил-KoA → на синтез гема; 4) Оксалоацетат → синтез Глюкозы и аминокислот (Асп, Асн). Чтобы цикл Кребса не прекращался, существуют реакции, восполняющие утечку из него метаболитов. Основную реакцию, восполняющую утечку метаболитов из цикла Кребса, катализирует фермент пируваткарбоксилаза: COOH CH3 C O + CO2 COOH Пируват Пируваткарбоксилаза (Ko: Биотин) CH2 C O АТФ H2 O H3PO4 АДФ COOH Оксалоацетат Гипоэнергетические состояния Гипоэнергетические состояния развиваются снижении синтеза АТФ в организме. при Причины гипоэнергетических состояний Голодание Гипоксия Нарушение использования O2 в клетках Недостаток O2 в воздухе Гиповитаминозы B1, B2, PP, B5, биотина; недостаточность Q Нарушение поступления O2 в ткани (заболевания легких, нарушения в стр-ре Hb, нарушения кровообращения и др.) Основные причины нарушения использования O2 в клетках Снижение уровня гемоглобина, цитохромов Присутствие ингибиторов ЦПЭ и разобщителей тканевого дыхания и окислительного фосфорилирования Железодефицитные анемии Наследственные дефекты ферментов ЦПЭ, ОПК или АТФ-синтазы ОБМЕН УГЛЕВОДОВ ОДО Лекция № 1 Функции углеводов: 1. Энергетическая – окисление глюкозы обеспечивает половину суточной потребности организма в энергии 2. Структурная – из углеводов простроены гликозамингликаны, которые входят в состав межклеточного матрикса, р а также входят в состав гликопротеинов, гликолипидов. 3. Анаболическая – из углеводов в процессе метаболизма б образуются б исходные соединения, которые необходимы для синтеза липидов, аминокислот, нуклеотидов. 4. Защитная – глюкуроновая кислота участвует в обезвреживании токсических соединений, гепарин участвует в противосвертывающей системе крови. крови Основные углеводы пищи С Суточная потребность б в углеводах 400 400--500 гр. Полисахариды: Полисахариды: ► Крахмал Моносахариды ► Гликоген ► Глюкоза ► Целлюлоза ► Фруктоза ру Дисахариды ► Сахароза ► лактоза Строение р крахмала р z Крахмал – основной углевод пищи, пищи содержится в хлебе, картофеле, мучных изделиях. Он состоит из амилозы и амилопектина. Амилоза неразветвленный полисахарид включающий 200-300 остатков глюкозы. Амилопектин имеет структуру В местах разветвленную структуру. ветвления остатки глюкозы соединены α-1,6-гликозидными связями. Линейные участки содержат примерно 20-25 остатков глюкозы. При этом формируется древовидная структура. Крахмал высокомолекулярное соединение состоящее из сотни тысяч остатков глюкозы. Его молекулярная масса составляет 105-108Д. Строение гликогена z Гликоген – это полисахарид животного происхождения, состоит из остатков глюкозы. В клетках животных выполняет резервную функцию, но в пище содержится лишь небольшое количество гликогена, он не имеет пищевого значения. Представляет собой структурный аналог крахмала, но имеет большую степень ветвления: примерно на каждые 10 остатков глюкозы приходится одна α α1,6-гликозидная связь. Целлюлоза ((клетчатка)) Ц Гомополисахарид растительного происхождения. Состоит С из остатков глюкозы связанных между собой β-1,4р человека нет ф фермента, р гликозидной связью. В организме расщепляющего такую связь, поэтому она не переваривается в желудочно-кишечном тракте. Д Дисахариды р д z Лактоза - молочный сахар сахар, состоит из остатков галактозы и глюкозы связанные между собой β14 β-1,4-гликозидной й связью. z Сахароза - содержится в овощах и фруктах. Состоит из остатков глюкозы и фруктозы соединенных α β-1 2-гликозидной связью α,β-1,2-гликозидной связью. Основные моносахариды р д Переваривание углеводов • ПЕРЕВАРИВАНИЕ КРАХМАЛА НАЧИНАЕТСЯ В РОТОВОЙ Й ПОЛОСТИ. Большие слюнные железы синтезируют фермент ф р α-амилазу у слюны,, которая р расщепляет р α 1,4 , гликозидные связи в крахмале. Так как пища в ротовой полости находится не долго, то здесь идет частичный гидролиз крахмала до декстринов. • Основным местом переваривания углеводов является тонкий кишечник. Из поджелудочной железы туда поступают: панкреатическая α-амилаза α амилаза и бикарбонаты, бикарбонаты которые создают оптимум рН для фермента. Крахмал расщепляется ферментом до мальтозы и изомальтозы. • Дисахариды, полученные при гидролизе крахмала (мальтоза и изомальтоза), сахароза и лактоза, поступившие с пищей, расщепляются гликозидазами тонкого кишечника до моносахаридов, которые и всасываются из кишечника в кровь. Переваривание р р углеводов у д Гидролиз дисахаридов специфическими гликозидазами Гликозидазы тонкого кишечника синтезируются в клетках, но не секретируются в просвет кишечника, а образуют б на поверхности клеток крупные ферментативные комплексы с различной субстратной специфичностью: 1. Сахаразо-изомальтазный – гидролизует связи в сахарозе, изомальтозе, мальтозе. 2 Гликоамилазный – катализирует гидролиз 2. олигосахаридов, а также расщепляет связи в мальтозе. 3. β-гликозидазный – расщепляет лактозу. 4. Триголаза – расщепляет триголозу, дисахарид содержащийся в грибах грибах. Всасывание моносахаров р в энтероцит р Когда концентрация глюкозы, фруктозы, галактозы в просвете кишечника выше чем в клетке кишечника,, то они всасываются путем облегченной диффузии – по градиенту концентрации, при участии белка переносчика и без затрат энергии. Когда концентрация глюкозы и галактозы в просвете кишечника ниже чем в клетке кишечника, то всасывание идет путем Na-зависимого вторичноNa зависимого вторично активного транспорта. Na+ поступает в клетку по градиенту концентрации и «тащит»глюкозу за собой Градиент Гра е концентрации о е ра собой. Na+ создается работой Na+, К+-АТФазы. Глюкоза из энтороцитов перемещается во внеклеточную жидкость и далее с помощью облегченной диффузии в кровь. Нарушение ру переваривания р р и всасывания углеводов Дефекты ферментов, участвующих в гидролизе углеводов в кишечнике; Нарушение всасывания продуктов переваривания углеводов в клетки с з с ой оболочки слизистой обо о кишечника. е а Нарушение переваривания дисахаридов В норме натощак в крови содержится 3,5- 5,5 ммоль/л (60-100 мг/дл) глюкозы. При пищеварении (абсорбтивный период) ууровень глюкозы повышается примерно до 8 ммоль/л (150 мг/дл). Такое физиологическое повышение концентрации глюкозы в крови называют алиментарной (пищевой) гиперглюкоземией. Транспорт р р глюкозы из крови р в ткани Инсулин Рецептор р Мембрана клетки ГЛЮТ-4 Глюкоза Метаболизм глюкозы в клетках z Глюкоза, поступающая в клетки Глюкоза органов и тканей, подвергается фосфорилированию с участием АТФ Плазматическая мембрана АТФ. клеток непроницаема для фосфорилированной глюкозы и, следовательно она не может из д них выйти. Отличия гексокиназы от гюкокиназы Гексокиназа Глюкокиназа 1. По 1 П Км К ((по сродству к субстрату) Высокое сродство к В глюкозе Км<0,1 ммоль/л Низкое сродство к Н глюкозеКм=10 ммоль/л 2. По локализации в тканях Все ткани Печень, поджелудочная железа 3. По ингибированию конечным продуктом Ингибируется глюкозо-6фосфатом Не ингибируется глюкозо6-фосфатом 4. По специфичности Обладает относительной субстратной специфичность т.к. специфичность, т к может фосфорилировать и другие гексозы Обладает абсолютной субстратной специфичностью т.к. тк фосфорилирует только глюкозу Пути использования глюкозо глюкозо-6-фосфата 6 фосфата в тканях Обмен гликогена Глюкоза в абсорбтивный период (1-2 часа после приема пищи содержащей углеводы) запасается в большинстве пищи, тканей в виде гомополисахарида гликогена гликогена.. Синтез стимулируется гормоном инсулином (инсулин инсулин-- гормон сытости)). Резервная роль гликогена обусловлена двумя сытости важными свойствами: свойствами: он осмотически неактивен и сильно ветвится, е с благодаря б а о ар чему е глюкоза ю оза быстро б с ро присоединяется р сое е с к полимеру при биосинтезе и отщепляется при мобилизации.. мобилизации В наибольших количествах этот гомополисахарид содержится в мышцах ц ((≈ ≈ 350 г)) и печени ((≈100 ≈100 г). ) ММ~ ММ ~107-108 . В клетках гликоген находится в виде гранул гликогена, которые плохо растворяются в воде. Синтез гликогена Мобилизация (распад) гликогена z Происходит в интервалах между приемами пищи и при физической нагрузке и стимулируется гормонами : z Глюкагоном (гормон голода) z Адреналином (гормон ф физической й нагрузки, гормон стресса). Различия мобилизации гликогена в печени и мышцах Регуляция синтеза и распада гликогена ¾ ¾ П Переключение процессов синтеза и мобилизации б гликогена в печени и мышцах происходит при переходе р д из абсорбтивного р состояния в постабсорбтивное и из состояния покоя в режим физической работы. работы. В переключении этих метаболических путей в печени участвуют инсулин, инсулин глюкагон и адреналин адреналин,, а в мышцах – инсулин и адреналин.. адреналин Влияние этих гормонов на синтез и распад гликогена осуществляется путем изменения в противоположном направлении активности двух ключевых ферментов: ферментов: гликогенсинтазы и гликогенфосфорилазы.. гликогенфосфорилазы Изменение активности гликогенсинтазы и гликогенфосфорилазы А Активность этих ферментов ф регулируется с с помощью фосфорилирования / дефосфорилирования. ¾ Гликогенфосфорилаза существует в двух формах: Фосфорилированная – активная Дефосфорилированная – неактивная ¾ Гликогенсинтаза тоже существует в двух формах: Дефосфорилированная –активная Ф ф Фосфорилированная – неактивная Влияние инсулина на активность гликогенсинтазы и гликогенфосфорилазы ф ф Регуляция синтеза и распада гликогена в печени Инозитолфосфатный механизм регуляции синтеза и распада гликогена в печени адреналином и Са2+ Регуляция метаболизма гликогена в мышцах при физической нагрузке Гликогеновые болезни ¾ ¾ Гликогенозы (болезни накопления гликогена) обусловлены наследственным дефектом ферментов, участвующие в распаде гликогена гликогена. Гликогеноз проявляется избыточным накоплением гликогена в печени,, сердечной рд и скелетных мышцах, ц , почках , легких и др. Результатом нарушения распада гликогена является гипоглюкоземия. Агликогенозы являются следствием нарушения синтеза гликогена и сопровождаются снижением его содержания в тканях. Результатом Р также является гипоглюкоземия. По клиническим признакам различают три основные ф формы заболевания: б 1 Печеночную 2 Мышечную 3 Генерализированную П Печеночная ф форма Признаки появляются на первом году жизни, жизни начиная с 8 8--9 месяца месяца, когда отмечаются редкие приступы гипогликемии. Они характеризуются временной потерей сознания, клоническими судорогами конечностей. конечностей. Припадки начинаются до еды или утром, их появление можно предотвратить приемом сладкой воды. Х Характерный й внешний й вид детей: й маленький й рост, б большой й живот, худые конечности, "кукольное лицо". Отмечается увеличение печени. Самый опасный возраст - первые 4 4--5 лет лет. Инфекционные заболевания осложняют течение патологического процесса. С возрастом состояние больного улучшается благодаря развитию компенсаторных механизмов обмена. Интеллект, как правило, не нарушен. Мышечная ф форма р Симптомы заболевания появляются с 7-10 лет. Больные становятся малоподвижными, быстро устают при физической нагрузке. Мышечная слабость прогрессирует, появляются боли в работающих мышцах, усиленное сердцебиение, одышка. Все эти симптомы развиваются до 25-35 25 35 лет. Внешний В й вид больных б не меняется. Симптомов изменения печени и почек нет. При осмотре выраженная атрофия мышц и их гипотония. При гистохимическом исследовании видно мышечные волокна с дегенерацией, вплоть до некроза. Они содержат вакуоли, вакуоли наполненные гликогеном. гликогеном Прогноз заболевания благоприятный, но известны летальные исходы в 20-30 лет. лет Генерализированная форма При этой форме заболевания практически во всех органах и тканях накапливается большое количество гликогена. Клиника ее не похожа на а клинику у печеночной е е о о и мышечной е о фор форм,, она о а многообразна, имеет прогрессирующее течение. Гликогеновые болезни и их причины Спасибо за внимание! ОБМЕН УГЛЕВОДОВ Лекция № 2 Катаболизм глюкозы Глюкоза является универсальным источником энергии для всех клеток организма. При катаболизме глюкозы АТФ может синтезироваться как в аэробных аэробных, так и в анаэробных условиях. Гликолиз – специфический путь катаболизма глюкозы. Существует три метаболических, пути в которых р может окисляться глюкоза: Аэробный гликолиз – распад глюкозы до 2 молекул пирувата который происходит в аэробных условиях и пирувата, сопровождается синтезом 8 молекул АТФ. Анаэробный гликолиз – распад глюкозы до 2 молекул лактата без участия кислорода и митохондриальной ЦПЭ. Энергетический Ц р выход д – 2 молекулы у АТФ. Аэробный распад глюкозы до СО2 и Н2О – включает аэробный гликолиз и последующее окисление 2 молекул пирувата в общем пути катаболизма. При этом синтезируется 38 молекул АТФ. Аэробный гликолиз Аэробный гликолиз – ферментативный процесс, который й включает в себя б 10 реакций. Все эти реакции протекают в цитозоле клетки. В последовательности реакций можно выделить 2 этапа. На первом этапе из молекулы глюкозы образуются 2 триозы: глицероальдегид-3фосфат (ГАФ) и дигидроксиацетонфосфат ((ДАФ). ) Этот этап идет с затратой р энергии. р 1 этап аэробного гликолиза включает 5 реакций: 1. Фосфорилирование глюкозы гексокиназой (глюкокиназой) с образованием глюкозо глюкозо-6-фосфата, 6 фосфата, которое идет с затратой АТФ. 2. Изомеризацию глюкозо-6-фосфата во фруктозу-6-фосфат при участии фосфоглюкоизомеразы. 3. Фосфорилирование фруктозо фруктозо-6-фосфата 6 фосфата фосфофруктокиназой с затратой АТФ. 4. Альдольное расщепление фруктозо-1,6-бифосфата, катализируемое р 2 триоз р ГАФ и ДАФ. Д альдолазой,, с образованием 5. Изомеризацию ДАФ в ГАФ под действием триозофосфатизомеразы. Первый этап гликолиза требует затраты 2 молекул АТФ и приводит к образованию 2 молекул ГАФ. 2 этап аэробного р гликолиза Этот этап обеспечивает синтез АТФ. В него вступают 2 молекулы ГАФ, поэтому стехиометрический коэффициент для всех последующих у р реакций гликолиза р равен 2. 6. Дегидрирование ГАФ NAD+- зависимой глицероальдегидфосфатдегидрогеназой при участии Н3РО4, которое приводит к образованию 1,3-бифосфоглицерата, содержащего макроэргическую связь. 7. Превращение р р 1,3-бифосфоглицерата ф ф р в 3-фосфоглицерат ф ф р под действием фосфоглицераткиназы, сопровождающееся субстратным фосфорилированием АДФ. 8. Изомеризация 3 8 3-фосфоглицерата фосфоглицерата в 2-фосфоглицерат 2 фосфоглицерат, катализируемая фосфоглицератмутазой. 9. Дегидротация 2-фосфоглицерата ферментом енолазой с образование ф ф фосфоенолпирувата, содержащего макроэргическую связь. 10. Образованием пирувата из фосфоенолпирувата под действием пируваткиназы, сопряженное с субстратным фосфорелированием АДФ. Регенерация NAD+ Реакция дегидрирования ГАФ под действием NAD+зависимой глицероальдегидфосфатдегидрогеназы идет с участием кофермента NAD+ который восстанавливается в NADН+Н. При накоплении NADН+Н происходит торможение гликолиза, поэтому должна происходить регенерация NAD+, которая в аэробном гликолизе происходит в ЦПЭ, но мембрана митохондрий непроницаема для NADН+Н. Поэтому транспорт водорода через митохондриальную мембрану осуществляется с помощью особых челночных механизмов. Их два: малат-аспартатный и глицерофосфатный. Малат-аспартатный челнок • В малат-аспартатном челноке NADH+H используется для восстановления оксалоацетата цитоплазматической малатдегидрогеназой в малат. Который с помощью специфической транслоказы проходит через мембрану митохондрий и окисляется митохондриальной малатдегидрогеназой, образовавшийся при этом NADH+H отдает Н+ и ē – в ЦПЭ. В митохондриях оксалоацетат вступает в реакцию трансаминирования р р и превращается р р в аспартат, р , который р выходит при р у участии транслоказы в цитоплазму. В цитоплазме эта аминокислота включается в реакцию трансаминирования с α-кетоглутаратом и снова превращается в оксалоацетат. р/о=3. Глицерофосфатный челнок • Глицерофосфатный челнок функционирует в клетках белых мышц и печени. Образующийся в окислительной реакции гликолиза NADH является донором для восстановления ДАФ в глицерол-3-фосфат цитоплазматической глицерол3-фосфатдегидрогеназой. Глицерол-3-фосфат поступает в митохондрии и окисляется FAD-зависимой FAD зависимой митохондриальной глицерол-3глицерол 3 фосфатдегидрогеназой. р/о=2. Выход АТФ в аэробном гликолизе, сопряженным с работой малат-аспартатного челнока • Первый этап аэробного гликолиза идет с затратой 2 молекул АТФ (гексокиназная и фосфофруктокиназная реакции) -2 2 АТФ • Второй этап аэробного гликолиза идет с выходом 10 молекул АТФ: • 2 NADH+H поступают в ЦПЭ, где образуется 2х3=6 АТФ за ссчет е о окислительного с е о о фосфор фосфорилирования. ро а • Также в аэробном гликолизе есть 2 реакции субстратного у р ф фосфорилирования ф р р (фосфоглицераткиназная и пируваткиназная реакции) 2х2=4 АТФ • Σ=10-2=8 АТФ Аэробный распад глюкозы • • • • • Аэробный распад глюкозы включает реакции аэробного гликолиза (специфического пути катаболизма глюкозы) глюкозы), который заканчивается образованием 2 молекул пирувата и его окислением в щ путях у катаболизма ((ОПК)) д до СО2 и Н2О и в общих митохондриальной ЦПЭ. Реакции аэробного распада глюкозы идут в цитозоле и митохондриях. Аэробный распад глюкозы – это основной путь катаболизма глюкозы в большинстве тканей. тканей Для мозга глюкоза – основной источник энергии. р распада р д глюкозы в печени Значение аэробного заключается в образовании субстратов, необходимых для синтеза липидов. Выход АТФ при аэробном распаде глюкозы 1. Аэробный гликолиз - 8 АТФ 2 Превращение 2 молекул пирувата в 2 2. ацетил-КоА - 2х3=6 АТФ 3 Окисление 3. О 2А Ацетил-КоА К А до СО2 и Н2О - 2х12=24 АТФ Всего – 38 АТФ Анаэробный гликолиз • Анаэробный гликолиз - окисление глюкозы без кислорода. р д Он заканчивается образованием 2 молекул лактата и протекает в тканях: 1. мышцах - в первые минуты мышечной работы. 2 эритроцитах, 2. эритроцитах т.к. т к в них отсутствуют митохондрии. 3 клетках опухолевой 3. й ткани. Реакции анаэробного гликолиза • 10 реакций анаэробного гликолиза идентичны реакциям аэробного гликолиза. Они протекают в цитозоле клетки клетки. Отличаются лишь 11-ой реакцией, которая специфична для анаэробного гликолиза, гликолиза в которой идет восстановление пирувата в лактат под действием лактатдегидрогеназы. лактатдегидрогеназы Эта реакция необходима для регенерации NAD+ из NADН+Н без участия митохондриальной цепи переноса электрона в ситуациях при которых клетки недостаточно снабжаются кислородом. кислородом Лактатдегидрогеназная Ла а де дро е аз а реакция реа ц • Роль акцептора водорода выполняет пируват. • Значение этой реакции заключается не в образовании образо а лактата, ла а а, а в том, о , что оэ эта а цитозольная реакция обеспечивает регенерацию NAD+ . Вход АТФ при анаэробном гликолизе • Первый этап анаэробного гликолиза идет с затратой 2 молекул АТФ (гексокиназная и фосфофруктокиназная реакции) -2 АТФ • Второй этап анаэробного гликолиза идет с выходом 4 молекул АТФ: АТФ В анаэробном гликолизе есть 2 реакции субстратного фосфорилирования (фосфоглицераткиназная (ф ф р и пируваткиназная реакции) 2х2=4 АТФ • Σ=4-2=2 АТФ Утилизация лактата • Лактат образовавшийся в эритроцитах и мышцах поступает в кровь, а из нее в печень В печени в аэробных условиях печень. лактат превращается в пируват и исполь зуется в глюконеогенезе. • Утилизация лактата может происходить и в почках, сердце, печени. В аэробных условия лактат превращается также в пируват пируват, который включается в ОПК, где и окисляется до СО2 и Н2О и АТФ. АТФ Лактоацидоз Лактат всегда присутствует в крови, но его концентрация может повышаться при: 1. Гипоксии 2. Гиповитаминозах В1,В2,РР. 3. Наследственной недостаточности пируваткарбоксилазы и ПДК. Регуляторные у р ферменты ф р гликолиза Гексокиназа (Глюкокиназа) Фосфофруктокиназа Пируваткиназа ЭТИ ФЕРМЕНТЫ КАТАЛИЗИРУЮТ НЕОБРАТИМЫЕ РЕАКЦИИ ГЛИКОЛИЗА Регуляция гликолиза в мышцах В скелетных мышцах гликолиз имеет энергетическое значение. При высоком энергетическом потенциале NADH и АТФ аллостерически ингибируют фосфофруктопируваткиназу а также киназу и пируваткиназу, регуляторные ферменты ОПК, вследствии чего гликолиз и аэробный распад глюкозы тормозится. тормозится Снижение активности фосфофруктокиназы приводит к накоплению глюкозо-6б фосфата, который ингибирует гексокиназу. При р низком з о потенциале о е ц але клетки ле избыток АМФ аллостерически активирует фосфофруктокиназу и, таким образом, образом активирует гликолиз. гликолиз Пентозофосфатный путь глюкозы Это альтернативный путь окисления глюкозы, у которого ярко выражена анаболическая функция. • - Он является поставщиком кофермента NADPH, который используется для реакций восстановления и гидроксилирования, а также, обеспечивает клетки рибозо-5-фосфатом, рибозо 5 фосфатом необходимым для синтеза нуклеотидов изз которых о ор построены ос рое нуклеиновые е о е кислоты. Наиболее активно пентозофосфатный й путь протекает в: • • • • • • Печени Жировой р ткани Лактирующей молочной железе Эритроцитах Коре надпочечников С Семенниках Реакции пентозофосфатного пути Все реакции этого метаболического у протекают р в цитозоле ц клеток и пути их можно разделить на два этапа: • Окислительный • Неокислительный Реакции окислительного этап ПФ пути Окислительный путь Суммарное уравнение Реакции окислительного ПФ пути служат основным источником NADPH в клетках. NADPH как донор водорода участвует в анаболических б процессах: • Синтез холестерола • Синтез жирных кислот • Синтез желчных кислот NADPH также необходим как донор водорода при катаболизме лекарственных р веществ и других ру чужеродных соединений. Регуляция • Скорость окислительного этапа ф ф пути у зависит от пентозофосфатного соотношения NADPH/NADP+. Восстановленный NADPH ингибирует глюкозо-6-фосфатдегидрогеназу. Превращение NADPH в окисленное состояние NADP+ приводит к ослаблению ингибирования фермента. Неокислительный этап ПФ пути • Неокислительный путь у образования р пентоз включает реакции переноса 2 и 3 углеродных фрагментов с одной молекулы фр у на другую. ру у Этот путь служит для образования пентоз. Неокислительный путь у образования р пентоз обратим, следовательно, он может служить для образования р гексоз и пентоз. С помощью этого пути избыток пентоз, превышающий потребности р клетки, может быть возвращен в фонд гексоз. Реакции неокислительного этапа ПФ пути Седогептулозо Ферменты неокислительного ПФ пути 1.Транскетолаза (ТДФ, Вит. В1) 2 Трансальдолаза 2.Трансальдолаза 3.Транскетолаза (ТДФ, Вит. В1) 4. Эпимераза 5 Пентозофосфатизомераза 5. Пентозофосфатный цикл • Окислительный этап синтеза пентоз и неокислительный й этап составляют циклический й процесс – пентозофосфатный цикл – за один оборот цикла полностью распадается одна молекула глюкозы. Пентозофосфатный цикл функционирует фу ц ру в основном только в жировой р ткани и печени. Суммарное уравнение ПФ цикла: • Если клетке необходимы рибозо рибозо-55 фосфат для синтеза нуклеотидов и NADPН для восстановительных синтезов, то протекает окислительный этап пентозофосфатного пути пути. • Если клетке необходим только рибозо-5фосфат то идет неокислительный этап фосфат, пентозофосфатного пути. • Если Е клетке необходим б NADPH для восстановительных синтезов, то протекает пентозофосфатный цикл. Спасибо за внимание! ОБМЕН УГЛЕВОДОВ Лекция № 3 Глюконеогенез z Глюконеогенез – это процесс р ц синтеза глюкозы из веществ неуглеводной природы. Он активируется у при длительном голодании (более суток, т.к. к этому моменту исчерпываются запасы гликогена в печени) и интенсивной физической нагрузке. За счет реакций глюконеогенеза в сутки синтезируется 80-100 г глюкозы, причем 90% глюкозы синтезируется в печени и 10% в корковом слое почек и слизистой оболочке кишечника. Субстраты глюконеогенеза Пируват Оксалоацетат Лактат Глицерол Аминокислоты Включение различных субстратов в глюконеогенез зависит от физиологического состояния организма: Лактат постоянно образуется в эритроцитах и работающих мышцах в анаэробном гликолизе Глицерол образуется при распаде жиров в жировой ткани при р голодании д Аминокислоты образуются при распаде тканевых белков Включение субстратов в глюконеогенез Значение глюконеогенеза Глюконеогенез является единственным источником глюкозы в крови при длительном голодании. П Постоянное поступление глюкозы в качестве источника энергии особенно необходимо для мозга, эритроцитов, хрусталика и сетчатки глаза. Реакции глюконеогенеза • Большинство реакций глюконеогенеза являются противоположно направленными гликолизу , т.е. являются обратимыми и катализируются теми же ферментами, ф что и соответствующие реакции гликолиза. Но три реакции гликолиза являются необратимыми, поэтому в глюконеогенезе образование ф ф фосфоенолпирувата из пирувата, фруктозо-6-фосфата из фруктозо-1,6-бифосфата и глюкозы из глюкозо-6-фосфата глюкозо 6 фосфата осуществляют другие ферменты. • Реакция превращения пирувата в оксалоацетат происходит в митохондриях, а все остальные реакции в цитозоле клетки. Глюконеогенез 1-я необратимая реакция глюконеогенеза • Пируват превращается в фосфоенолфосфоенол пируват в результате двух последовательных реакций. Сначала пируват поступает в митохондрии, где превращается в оксалоацетат под действием биотинзависимого фермента пируваткарбоксилазы. Реакция идет с затратой 1 молекулы АТФ АТФ. О Оксалоацетат восстанавливается митохондриальной малатдегидрогеназой в малат. Он поступает у в цитоплазмуу по механизмуу пассивного антипорта, где окисляется цитоплазматической малатдегидрогеназой с образованием оксалоацетата. 2-я необратимая реакция глюконеогенеза • В цитоплазме оксалоацетат под действием ф ф фосфоенолпируваткарбоксикиназы ру р превращается в фосфоенолпируват, при этом затрачивается 1 молекула ГТФ. 3-я необратимая реакция глюконеогенеза • Дефосфорилирование фруктозр1,6-бифосфата под действием фермента фруктозо-1,6-бифосфатазы, которая отщепляет Н3РО4 с участием воды. 4-я необратимая реакция глюконеогенеза • Дефосфорилирование глюкозо-6бфосфата под действием фермента глюкозо-6-фосфатазы, которая отщепляет Н3РО4 с участием воды. Энергетический баланс глюконеогенеза из пирувата. Три реакции глюконеогенеза идут с затратой энергии: • Превращение пирувата в оксалоацетат (тратится 1 молекула АТФ) • 3-фосфоглицерата в 1,3бифосфоглицерат (тратится 1 молекула АТФ) • Оксалоацетат в фосфоенолпируват ф ф ру ((тратится р 1 молекула ГТФ) Учитывая, что стехиометрический коэффициент этого этапа глюконеогенеза равен 2, 2 то для синтеза 1 молекулы глюкозы из 2 молекул пирувата необходимо 4 АТФ и 2 ГТФ (всего 6 АТФ). Глюкозолактатный цикл (цикл Кори) • Г Глюкозолактатный й цикл обеспечивает использование лактата для синтеза глюкозы, предотвращая его накопление в тканях. тканях Синтез глюкозы из глицерола. • Глицерол образуется в жировой ткани при гидролизе жира, поступает в кровь и транспортируется в печень, печень где используется как субстрат глюконеогенеза. Для синтеза 1 молекулы глюкозы необходимо 2 молекулы глицерола. Фермент глицеролкиназа фосфорилирует глицерол при участии АТФ с образованием глицерол-3фосфата. Последний окисляется NAD+ - зависимой глицерол глицерол-33 фосфатдегидрогеназой с образованием промежуточного метаболита е абол а глюконеогенеза лю о ео е еза – диоксиацетонфосфата. Синтез глюкозы из глицерола. Синтез глюкозы из аминокислот. • Если в процессе катаболизма а аминокислот о с о образуются образ ю с пируват, р а оксалоацетат или промежуточные метаболиты цитратного цикла (αкетоглутарат, сукцинил-КоА, ф фумарат), ) то эти вещества могут служить субстратами глюконеогенеза, а аминокислоты, из которых р они образуются, р у , называются гликогенными. При длительном голодании или диете, содержащей только белки, уровень глюкозы в крови поддерживается за счет глюконеогенеза из аминокислот. Регуляция гликолиза и глюконеогенеза в печени • В печени, в отличии от других органов, идут реакции как гликолиза, так и глюконеогенеза. Скорость каждого определяется физиологическим состоянием организма: в абсорбтивный период ускоряется гликолиз и тормозится глюконеогенез, а при голодании наоборот, скорость гликолиза снижается, а глюконеогенеза – возрастает. Регуляция направлена на необратимые стадии гликолиза и глюконеогенеза. Активность регуляторных ферментов регулируется: • Аллостерически • Фосфорелированием/ дефосфорилированием • Количеством ферментов (индукцией и репрессией синтеза ) Схема гликолиза и глюконеогенеза Регуляция I субстратного цикла • Направление Н реакции первого субстратного цикла регулируется р у ру главным образом р концентрацией глюкозы. При пищеварении, когда концентрация глюкозы в крови повышена, активность глюкокиназы максимальна. Кроме того инсулин индуцирует синтез глюкокиназы и ускоряет тем самым фосфорилирование глюкозы. Регуляция II субстратного цикла • Активность второго субстратного б цикла регулируется аллостерически. Этот цикл составляют реакция фосфорилирования ф ф р р фруктозо-6фру фосфата с образованием фруктозо1,6-бифосфата,которую катализирует фосфофруктокиназа и реакция дефосфорилирования фруктозо фруктозо-1 1,6 6фосфата под действием фруктозо1,6-бифосфатазы. Аллостерический регулятор ферментов этого цикла фруктозо 2 6 бифосфат который фруктозо-2,6активирует фосфофруктокиназу и ингибирует фруктозо-1,6бифосфотазу. • Фруктозо-2,6- бифосфат является мощным аллостерическим активатором гликолиза и ингибитором глюконеогенеза. Бифункциональный ф фермент (БИФ) • Этот фермент может катализировать реакцию фосфорилирования фруктозо-6-фосфата с образованием фруктозо-2,6-бифосфата и обратную реакцию дефосфорилирования фруктозо-2,6-бифосфата с образованием фруктозо-6-фосфата. В зависимости от конформации, которая изменяется при фосфорилировании / дефосфорилировании, БИФ работает как киназа (БИФ-ОН) или фосфатаза (БИФ-Р). Регуляция активности бифункционального фермента Регуляция III субстратного цикла • В регуляции этого цикла основная роль принадлежит пируваткиназе ф фермента гликолиза. О Она регулируется путем фосфорилирования/дефосфорилиро вания (фосфорилированная (ф ф ф форма неактивна, а дефосфорилированная – активна) и аллостерически. • Фруктозо-1,6-бифосфат является аллостерическим активатором пируваткиназы. Регуляция активности пируваткиназы инсулином и глюкагоном При голодании ускорение глюконеогенеза и замедление гликолиза обусловлены следующими механизмами регуляции: • Фермент глюконеогенеза III субстратного цикла пируваткарбоксилаза аллостерически активируется АцетилКоА, который й при голодании образуется б в митохондриях при β-окислении жирных кислот. • При голодании увеличивается синтез и секреция стероидного гормона кортизола. Он поступает в гепатоциты, взаимодействует с внутриклеточным рецептором, ускоряет транскрипцию гена фосфоенолпируваткарбоксикиназы и таким образом увеличивает количество фермента в клетке. Регуляция гликолиза и глюконеогенеза в печени энергетическим зарядом клетки Влияние этанола на обмен углеводов • До 90% поступающего в организм этанола окисляется в печени. Сначала в цитоплазме гепатоцитов этанол под действием NAD+ зависимой алкогольдегидрогеназы в превращается в ацетальдегид, который поступает в митохондрии. В митохондриях ацетальдегид окисляется NAD+ - зависимой ацетальдегиддегидрогеназой с образованием ацетата. Ацетат поступает в кровь или под действием ацетил-КоА-синтетазы пи участии HSKoA и АТФ превращается р р щ в ацетил-КоА. ц Ацетил-КоА окисляется в ЦТК, а также использоваться для синтеза жирных кислот или кетоновых тел Окисление этанола в печени • В результате окисления этанола в печени увеличивается отношение NADH/NAD+ . Следствием увеличения образования р NADH является сдвиг д реакции, катализируемой ЛДГ, в сторону образования лактата. Изменение скорости этой реакции сопровождается со ро о дае с уменьшением у е е е содержания пирувата и увеличением лактата. Это приводит к снижению скорости глюконеогенеза. В крови снижается концентрация глюкозы (гипоглюкоземия), а лактата – повышается, что приводит к лактоацидозу. •Спасибо за внимание!! Обмен липидов Липиды – это органические вещества, для которых характерны общие свойства: гидрофобность или амфифильность. Основные классы липидов: 1) Жирные кислоты (ЖК). Входят в большинство липидов организма человека (жиры, фосфолипиды); 2) Жиры – Триацилглицеролы (ТАГ). Выполняют энергетическую функцию. Это самая компактная и очень вместительная форма хранения энергии. Жиры запасаются в жировых клетках – адипоцитах, которые входят в состав жировой ткани. 3) Фосфолипиды – амфифильные молекулы. Участвуют в формировании двойного липидного слоя мембран или монослоя на поверхности липопротеинов. 4) Стероиды Холестерол (ХС) и его производные: Желчные кислоты и их соли, стероидные гормоны. 5) Жирорастворимые витамины: A, D, E, K. Состав и строение основных высших ЖК организма человека: Жирные кислоты делятся на: насыщенные и ненасыщенные. Общая формула насыщенной жирной кислоты: CnH2n+1COOH П С Пальмитиновая 16:0 C15H31COOH Стеариновая 18:0 C17H35COOH Общая формула ненасыщенной жирной кислоты: CnH(2n+1)-2mCOOH (m - число -=- связей в молекуле ЖК). ПО О Л Л А Э Пальмитоолеиновая 16:1 C15H29COOH (Δ9) Олеиновая 18:1 C17H33COOH (Δ9) Линолевая 18:2 C17H31COOH (Δ9, 12) α-Линоленовая 18:3 C17H29COOH (Δ9, 12, 15) Арахидоновая 20:4 C19H31COOH (Δ5, 8, 11, 14) Эйкозапентаеновая 20:5 C19H29COOH (Δ5, 8, 11, 14, 17) ω6 ω3 ω6 ω3 Буква Δ означает положение 1-й двойной связи от карбоксильной группы. Последний углеродный атом метильной группы (Н3С-) независимо от длины цепи, обозначают буквой ω. ω β α Н3С-(СН2)n-СН2-СН2-СООН В организме человека синтезируются в основном ЖК с четным числом C атомов – насыщенные и мононенасыщенные. По кол-ву -=- связей ненасыщенные ЖК делятся на: а) моноеновые (ПО, О) – синтезируются в организме, в животных жирах занимают 2 (β) положение. б) полиеновые (Л, Л, А, Э). ! Полиеновые Жирные кислоты (Линолевая, Линоленовая) – это незаменимые (эссенциальные) ЖК. Они не могут синтезироваться в организме и должны обязательно поступать с пищей. Арахидоновая и Эйкозапентаеновая к-ты – условнозаменимые к-ты, могут синтезироваться из эссенциальных жирных кислот. Например: Арахидоновая из линолевой к-ты. По положению -=- связи все полиненасыщенные ЖК делят на 2 группы: ω6 и ω3 жирные к-ты. ω3: ω H3C–CH2–CH=CH….. ω6: ω H3C–(CH2)4–CH=CH….. Чтобы определить группу ω для полиненасыщенной ЖК надо от общего числа C атомов вычесть положение последней двойной связи от COOHгруппы. Наиболее полезные для организма ω3-жирные к-ты (особенно: Эйкозапентаеновая). Из нее образуются очень полезные эйкозаноиды, препятствующие атеросклерозу, инфаркту миокарда, тромбофлебиту и др. заболеваниям. Строение жиров (ТАГ): В составе жира человека преобладают насыщенные ЖК, а также мононенасыщенные: ПО, О. Также может встречаться Линолевая к-та. Животные жиры твердые при комн. tο из-за преобладания в них насыщенных ЖК. В растительных жирах и рыбьем жире преобладают полиненасыщенные ЖК. Это жидкие жиры при комн. tο. Ассимиляция пищевых жиров Ассимиляция включает в себя переваривание и транспорт жиров. 2 стадии: I стадия: Переваривание жиров (ТАГ). В ротовой полости и в желудке переваривание жиров не происходит из-за отсутствия необходимых для этого ферментов. Переваривание жира начинается перстной кишки с его гидролиза. 1. Гидролиз жира. Происходит при участии панкреатическая липаза. в полости 12- фермента – ! Т.к. гидролиз жира происходит в водной среде, а в воде жир не растворяется, то молекулы пищевого жира собираются в большие липидные капли. ! И гидролиз ТАГ происходит только на границе раздела фаз: жир-вода. Поэтому для увеличения поверхности контакта фермента с молекулами ТАГ липидные капли подвергаются эмульгированию. В роли эмульгатора используется желчь особенно ее компонент соли желчных кислот. и Первичные желчные кислоты синтезируются в печени из холестерола и секретируются в желчный пузырь. В желчном пузыре образуются простые мицеллы желчи, в которых содержатся: 1) соли желчных кислот; 2) фосфолипиды; 3) холестерол. После приема жирной пищи, желчный пузырь сокращается и желчь поступает в просвет 12-перстной кишки. Соли желчных молекулы. кислот – это амфифильные Гидрофобными участками они окружают молекулу жира, а гидрофильные участки располагаются снаружи, контактируя с молекулами воды => липидные капли разбиваются на более мелкие капли и => образуется тонкодисперсная эмульсия. В итоге происходит смешивание жира с водой и ↑ контакта E (панкреатической липазы) молекулами ТАГ. с Далее происходит гидролиз эмульгированных жиров при участии фермента панкреатическая липаза: Панкреатическая липаза синтезируется в поджелудочной железе и выделяется в полость 12перстной кишки. Активаторы реакции гидролиза ТАГ при участии фермента панкреатическая липаза: 1) соли желчных кислот – эмульгируют жиры и стабилизируют тонкодисперсную эмульсию. 2) белок Колипаза активаторы самого фермента – 3) ионы HCO3 Белок колипаза синтезируется в поджелудочной железе в неактивной форме и секретируются в полость 12-перстной кишки вместе с ферментом панкреатическая липаза, где активируется под действием трипсина путем частичного протеолиза. С помощью Колипазы: меняется конформация Панкреатической липазы и увеличивается ее сродство к субстрату – жиру: Регуляция активности панкреатической липазы: Ионы железой. HCO3– секретируются поджелудочной Функция HCO3– : нейтрализуют HCl, поступающую из желудка и обеспечивают оптимум pH для фермента (pH ≈ 7.8). Продукты гидролиза жира – β-МАГ и жирные к-ты включаются в состав мицелл желчи и образуются смешанные мицеллы. Состав смешанных мицелл: β-МАГ и жирные к-ты, а также соли желчных к-т, фосфолипиды, холестерол и витамины A, D, E, K. Смешанные энтероциты. мицеллы далее всасываются в 2. Ресинтез жиров в энтероцитах: В энтероцитах смешанные мицеллы распадаются на составные компоненты. желчи Продукты гидролиза жиров подвергаются ресинтезу. Ресинтез жира включает 2 стадии: 1) Активация жирной кислоты: 2) Взаимодействие β-МАГ и 2-х молекул ацил-KoA: !! Главное значение ресинтеза: ресинтезированный жир отличается по составу от пищевого, т.к. в его состав включаются также и ЖК, которые синтезируются в энтероцитах. Далее происходит транспорт ресинтезированных жиров по кровотоку к органам и тканям. II стадия: Транспорт жиров (ТАГ). Жиры, а также эфиры холестерола в воде не растворяются, поэтому их транспорт по лимфе и крови происходит с помощью специальных переносчиков. Роль переносчиков липидов по кровотоку выполняют Липопротеины – специальные комплексы липидов и белков. Все Липопротеины имеют схожее строение: Интегральные белки включаются в состав ЛП на стадии их формирования: Примеры: В-48 включаются в состав ХМ в энтероцитах; В-100 включаются в состав ЛПОНП в гепатоцитах. Периферические белки могут переходить с одного ЛП на другой (C-II, E, A-I и др.) Выделяют 5 типов липопротеинов (ЛП): ХМ – хиломикроны В этом ряду сверху вниз: ЛПОНП – липопротеины очень ↑ Плотность ЛП; ↑ Кол-во белков в составе ЛП ↑ Электрофоретическая подвижность (V перемещ. в электрич. поле) низкой плотности ЛППП – липопротеины промежуточной плотности ЛПНП – липопротеины низкой плотности ЛПВП – липопротеины высокой плотности ↓ Кол-во жиров в составе ЛП ↓ Размер частицы Главная функция ХМ – транспорт экзогенных (пищевых) жиров из кишечника к органам и тканям. Одна из функций ЛПОНП – транспорт эндогенных жиров, синтезированных в печени из печени к органам и тканям. Все 5 типов ЛП используются для транспорта ХС и его эфиров (ЭХС) по кровотоку. Жиры, ресинтезированные в энтероцитах включаются в состав ХМ. В энтероцитах образуются незрелые ХМ (ХМнезр.). ХМнезр. содержит один белок – интегральный АпоB-48. Далее ХМнезр. путем экзоцитоза поступают в лимфу, затем в кровь, где происходит их созревание: В ходе созревания ХМ происходит их контакт в крови с ЛПВП: ХМ получают от ЛПВП апопротеины C-II и E: Апо C-II – активатор фермента ЛП-липазы. Апо E – лиганд к рецептору мембраны гепатоцитов. ЛПВП получают от ХМ белок Апо A-I. Далее зрелые ХМ транспортируют жиры и ЭХС к органам и тканям. действует фермент В крови на ХМзрелые Липопротеинлипаза (ЛП-липаза), который распложен на поверхности эндотелия сосудов и капилляров органов: ↑↑ в жировой и мышечной тканях, в других тканях ↓. Под действием ЛП-липазы происходит гидролиз ТАГ, в составе ХМ и ЛПОНП до глицерола и 3-х молекул ЖК: Активаторы ЛП-липазы: Апо-C-II (в составе ХМ) – главный активатор, инсулин (в жировой ткани) и гепарин. ЛП-липаза имеет центр, комплементарный апо C-II (центр "узнавания" апо C-II) и каталитический центр, в котором протекает гидролиз ТАГ ХМ до глицерола и ЖК. При контакте ЛП-липазы с белком апо C-II на пов-ти ХМ происходит "захват" ХМ из кровотока и гидролиз жиров в составе ХМ: Продукты гидролиза ТАГ – глицерол и ЖК далее поступают в ткани: глицерол – в печень; жирные к-ты – вступают в β-окисление (мышцы) или используются на синтез жира (жировая ткань и печень в абсорбтивный период). ХМост. взаимодействуют с рецепторами гепатоцитов, контактируя с ними белком апо E => запускается рецептор-опосредованный эндоцитоз и ХМост. поглощаются печенью. ХМост. в печени включаются в лизосомы и подвергаются гидролитическому действию их ферментов => освобождаются ХС, ЖК и аминокислоты. Болезни, связанные с нарушением переваривания и транспорта жиров: 1) Стеаторея (выведение жиров с калом) Возникает из-за нарушения переваривания всасывания жиров и продуктов их гидролиза. и Причины: ↓ секреции: а) желчи (желчных кислот) б) панкреатического сока (панкреатической липазы) => ↑кол-во жиров выводимых с калом, возникает стеаторея. При длительной стеаторее развивается недостаточность: а) полиеновых ЖК (в организме не синтезируются) б) жирорастворимых витаминов А, D, Е, К. 2) Семейная гиперхиломикронемия (гипертриацилглицеролемия I типа) После приема пищи, содержащей ТАГ, в крови: ↑ [ХМ] – гиперхиломикронемия физиологическая ↑ [ТАГ] – гипертриацилглицеролемия В норме: через 1-2 часа поле еды [ХМ] в крови ↑ до 1.65 ммоль/л. Сыворотка крови – мутная, но через 4-5 часов [ХМ] в крови постепенно достигает нормы и сыворотка крови становится прозрачной. В норме ХМ проходят такой путь: ХМнезрелые → ХМзрелые → ХМост. → в печень Скорость удаления ХМ из кровотока зависит от: а) активности ЛП-липазы б) содержания ЛПВП (апо С-II, Е) в) активности переноса апо С-II и апо Е с ЛПВП на ХМ Нарушение структуры любого из белков и ферментов, участвующих в этом процессе, приводит к семейной гиперхиломикронемии. Симптомы: сыворотка крови мутная даже в абсорбтивный период. В тяжелых случаях: образуются ксантомы (отложения жира) в коже и сухожилиях. Происходит сужение просвета сосудов => боли в животе, нарушение памяти, функций поджелудочной железы. Содержание ТАГ в сыворотке крови после приема пищи, содержащей жир: Липидограммы сыворотки крови человека методом электрофореза в ПААГ: Необязательное дополнение к регуляции активности панкреатической липазы: Панкреатическая липаза и ее активаторы – белок колипаза и ионы HCO3– синтезируются в поджелудочной железе и секретируются в полость 12-перстной кишки. Колипаза своим гидрофобным доменом связывается с поверхностью мицеллы эмульгированного жира. Другая часть молекулы способствует формированию такой конформации панкреатической липазы, при которой активный центр фермента максимально приближен к своим субстратам – молекулам жиров, поэтому скорость реакции гидролиза жира резко возрастает. ДЕПОНИРОВАНИЕ ЖИРОВ В абсорбтивный период происходит депонирование жиров в жировой ткани, которые используются в постабсорбтивный период, при голодании и активной физической работе (стрессе). ! Жиры – наиболее выгодная и основная форма депонирования энергии. Депонированный жир может обеспечить организм энергией ~ до 40 дней полного голодания. В абсорбтивный период: содержащей углеводы: после приема пищи, В крови ↑ концентрация: а) глюкозы (140-160 мг/дл) б) ХМ (хиломикронов) в) инсулина => ↑ инсулин/глюкагоновый индекс Инсулин активирует синтез ЖК и ТАГ. В ПЕЧЕНИ: Инсулин индуцирует синтез глюкокиназы, которая катализирует реакцию: Глюкоза + АТФ ⎯→ Глюкозо – 6 – фосфат + АДФ Эта реакция "запирает" глюкозу в клетках в виде Глк-6-фосфата, который используется в гликолизе и пентозофосфатном пути превращения глюкозы (ПФП). В ЖИРОВОЙ ТКАНИ (инсулин-зависимая ткань): а) ГЛЮТ-4 перемещается из цитозоля в клеточную мембрану. б) ↑ V транспорта глюкозы в адипоциты. в) В адипоцитах инсулин индуцирует синтез ЛПлипазы. => синтез жиров идет в основном с использованием ЖК, которые образуются в ходе гидролиза жиров в составе ХМ и ЛПОНП. И в ПЕЧЕНИ и в ЖИРОВОЙ ТКАНИ: Инсулин ↑ активность: а) гликолиза (ДАФ, пируват, АТФ) б) ПДКомплекса (ацетил-KoA) в) ПФПути (NADPH) г) Синтеза жирных кислот и ТАГ Образование субстратов для синтеза ЖК и ТАГ Из глюкозы образуются все субстраты, необходимые для синтеза ЖК и ТАГ: Субстраты, необходимые для синтеза RCOOH (ЖК): Субстраты, необходимые для синтеза ТАГ: Ацетил-KoA, АТФ, NADPH Ацетил-KoA, АТФ, NADPH, ДАФ → Глицерол-3-фосфат Синтез жирных кислот 1. Образование исходного субстрата для синтеза ЖК – Ацетил-KoA. В абсорбтивный период: в ходе гликолиза, протекающего в цитозоле, образуется пируват. Пируват с помощью белка переносчика транспортируется в митохондрии, где происходит его окислительное декарбоксилирование. => образуется Ацетил-KoA. Ацетил-KoA вступает в цикл Кребса – в 1 р-ции которого образуется Цитрат. Цитрат с помощью транслоказы переносится в цитозоль, где с помощью Цитралиазы распадается на Ацетил-KoA и Оксалоацетат: Реакция, катализируемая цитратлиазой: Синтез жирных кислот: Ацетил-KoA идет на синтез ЖК, а ОА за 2 р-ции превращается в пируват: 2. Основные источники NADPH для синтеза ЖК: 1) ПФП превращения ГЛК (E – Глк-6-фосфатдегидрогеназа): Глк-6-фосфат + NADP+ → Глюконолактон-6-фосфат + NADPH 2) Малик-фермент (NADP+-зависимая малат-ДГ) Синтез ЖК происходит в печени и с меньшей активностью в жировой ткани. Все реакции синтеза ЖК происходят в цитозоле. 1 реакция синтеза ЖК – это превращение Ацетил-KoA в Малонил-KoA: Ацетил-KoA-карбоксилаза – регуляторный фермент синтеза жирных кислот. Дальнейшие реакции синтеза ЖК происходят при участии полиферментного комплекса – синтазы жирных к-т (пальмитатсинтазы). Синтаза ЖК – полифункциональный фермент, состоящий из 2-х идентичных протомеров, каждый из которых имеет доменное строение и содержит 7 активных центров и ацилпереносящий белок (АПБ). Растущая цепь ЖК, связана с АПБ в ходе всего синтеза ЖК на пальмитатсинтазе. АПБ переносит растущую цепь ЖК из одного активного центра в другой. Каждый протомер имеет содержащих SH-группы. 2 центра связывания, Одна SH-группа принадлежит цистеину, другая производному пантотеновой к-ты (фосфопантотеину): Реально идет синтез 2-х молекул ЖК одновременно, но для упрощения синтазу ЖК рисуют так: Этапы синтеза ЖК на синтазе ЖК: ! Синтез ЖК – циклический процесс. 1. – 2. Перенос ацетильного, а затем малонильного остатков на активные центры синтазы жирных кислот. Ацетильный остаток переносится на SH-группу Цис, а малонильный на SH-группу АПБ. Вначале образуется: Ацетил-E, затем Ацетил-Малонил-Е. 3. Декарбоксилирование малонильного остатка и конденсация 2-х ацетильных остатков. Образуется: Ацетоацетил-E; 4. Восстановление β-карбонильной группы с участием Ko: NADPH. Образуется: βгидроксибутирил-E; 5. Дегидратация => образуется = связь между α- и βуглеродными атомами. Образуется: Кротонил-E (Еноил-E) 6. Восстановление и образование Бутирил-E с участием Ko: NADPH. 7. Перенос бутирила на SH-группу Цис => завершается 1 цикл синтеза ЖК. Далее повторяются аналогичные циклы р-ций, пока в ходе 7 циклов не образуется Пальмитоил-E, который под действием тиоэстеразы комплекса гидролитически отделяется от SH-группы АПБ. => образуется свободная пальмитиновая к-та. Остальные насыщенные и мононенасыщенные ЖК, которые могут синтезироваться в организме, образуются из пальмитиновой кислоты. В 1-ом цикле синтеза ЖК: образуется 4 углеродных атома (Бутирил-E): 2 C-атома из Ацетил-KoA и 2 Cатома из Малонил-KoA. За каждый цикл (кроме 1-го) углеродный скелет ЖК удлиняется на 2 C-атома. Донор диуглеродных фрагментов: Малонил-KoA => Количество циклов синтеза ЖК составляет: , где n – общее число С-атомов в молекуле ЖК. Кол-во Малонил-KoA = кол-ву циклов = Кол-во Ацетил-KoA = кол-ву циклов + 1 = . . АТФ используется только в реакции образования Малонил-KoA из Ацетил-KoA. => Кол-во АТФ = кол-ву Малонил-KoA = кол-ву циклов = . Регуляция синтеза ЖК 2 пути регуляции синтеза ЖК: 1. регуляция действия Ацетил-KoA-карбоксилазы 2. индукция синтеза ферментов, синтезе ЖК из углеводов. участвующих в 1. Ацетил-KoA-карбоксилаза – регуляторный фермент синтеза ЖК. Способы регуляции действия: а) Ассоциация / диссоциация протомеров: Объединение 3-х тетрамеров в единый комплекс стимулирует цитрат (активатор); подавляет Пальмитоил-KoA (ингибитор). !! Таким образом: Цитрат играет 2 роли в синтезе ЖК: 1) Цитрат – форма переноса Ацетил-KoA из митохондрии в цитозоль; 2) Цитрат – активатор Ацетил-KoA-карбоксилазы. б) Фосфорилирование / дефосфорилирование В абсорбтивный период: при участии инсулина (активация происходит дефосфорилирование фермента). В постабсорбтивный период и при голодании: (при участии глюкагона) или при стрессовой нагрузке (при участии адреналина) – происходит фосфорилирование (ингибирование фермента): в) Индукция синтеза фермента Инсулин в абсорбтивный период индуцирует синтез Ацетил-KoA-карбоксилазы. (↑ V транскрипции гена и => ↑ кол-во фермента). 2. индукция синтеза ферментов, участвующих в синтезе ЖК из углеводов: Инсулин в абсорбтивный период индуцирует синтез ряда ферментов, участвующих в превращении продуктов катаболизма ГЛК в ЖК: 1 – Глюкозо-6-фосфатдегидрогеназа 2 – Цитратлиаза 3 – Малик-фермент 4 – Ацетил-KoA-карбоксилаза 5 – Синтаза ЖК Синтез жиров в печени и жировой ткани: ТАГ синтезируются из Глицерол-3-фосфата и АцилKoA. ! Отличия в синтезе жиров в печени и жировой ткани: И в печени и в жировой ткани Глицерол-3-фосфат может образовываться из Глюкозы. Из Глицерола – только в печени. Жиры, синтезированные в жировой ткани, в ней депонируются. Жиры, синтезированные в печени, включаются в состав ЛПОНП и секретируются в кровь. В крови они созревают, аналогично ХМ. Далее ЛПОНПзрелые подвергаются действию ЛПлипазы. Образующиеся при гидролизе ТАГ ЖК поступают в ткани (↑ в адипоциты), а глицерол – в печень. ОЖИРЕНИЕ Ожирение – это увеличение отложения жира в адипоцитах по сравнению с нормой. Важной характеристикой содержания организме является индекс массы тела: жира в Норма для ИМТ: 20-24.9 кг/м2. Ожирение Первичное Вторичное Первичное ожирение – результат нарушения баланса: Пища – затраты источник энергии >> энергии Причины первичного ожирения: 1) до 80% случаев – генетические нарушения; 2) состав и количество пищи; 3) уровень физической активности; 4) психологические факторы. Генетические факторы в развитии ожирения: 1) различие в V функционирования «бесполезных» циклов (субстратных циклов). Например: Если V1 ≈ V2 : ↓ кол-во продуктов, доступное для синтеза ТАГ => склонность к ожирению ↓. Если V1 >> V2 : ↑ склонность к ожирению. 2) более эффективная работа ЦПЭ: – кол-во рассеянного тепла ↓ – P/O ↑ => склонность к ожирению ↑. 3) изменение соотношения V аэробного и анаэробного распада глюкозы: Vаэробного >>> Vанаэробного => склонность к ожирению ↑. (38 АТФ) (2 АТФ) Vаэробного > Vанаэробного => склонность к ожирению ↓. 4) ↓ активности Na+, K+-АТФазы (в норме потребляет ~ 30% энергии клетки) => потребление энергии ↓ – склонность к ожирению ↑. 5) структура белка лептина При насыщении: Адипоцит лептин (ob) R гипоталамус нейропептид Y стимулирует потребление пищи 1 ситуация (~80% случаев): дефект R лептина. В этой ситуации: конц-ия лептина ~ в 4 раза ↑ нормы. 2 ситуация (~20% случаев): дефект стр-ры самого лептина. => в обеих ситуациях: конц-ия нейропептида Y ↑ – склонность к ожирению ↑. Вторичное ожирение – результат какого-либо первичного, чаще эндокринного заболевания (гипотиреоз, синдром Иценко-Кушинга и др.). МОБИЛИЗАЦИЯ ЖИРОВ Мобилизация (распад) жиров из жировой происходит: а) при активной физической работе (стрессе) => ↑ концентрация адреналина б) в постабсорбтивный период в) при голодании ткани Конц-ия в крови: Глюкозы 60-80 мг/дл ↑ конц-ия глюкагона ↓ конц-ия инсулина => ↓ инсулин/глюкагоновый индекс Гормон глюкагон (адреналин) взаимодействует с R (рецептором) мембраны адипоцитов. В ходе активации АЦ-системы: ПКАакт. фосфорилирует фермент ТАГ-липазу (гормоночувствительную липазу) => ↑ активность ТАГ-липазы => происходит гидролиз жиров: Образующиеся в ходе гидролиза ТАГ Глицерол и ЖК – поступают в кровь. Пути использования Глицерола и ЖК в тканях: Глицерол – гидрофильная молекула транспортируется по крови в печень без участия переносчиков. В печени Глицерол превращается в Глицерол-3фосфат: Глицерол-3-фосфат: В постабсорбтивный период (и при голодании): Вступает в глюконеогенез: При интенсивной мышечной работе (стрессе): Вступает в гликолиз, где используется в качестве источника энергии: Жирные кислоты – гидрофобные молекулы, транспортируются к тканям по крови в комплексе с белком альбумином. ! Т.к. ЖК транспортируются по крови в комплексе с альбумином, они не диссоциируют в крови на ионы и => не влияют на pH крови. В ходе мобилизации ТАГ, концентрация ЖК в крови ↑ ~ в 2 раза. Отделившись от альбумина, ЖК проходят через клеточные мембраны в цитозоль клеток. Аэробный катаболизм жирных кислот Происходит во всех тканях, кроме: – Нервная ткань (ЖК не проходят ГЭБ); – Эритроциты (из-за отсутствия митохондрий) Больше всего ЖК используют ткани: - мышцы скелетные - сердечная - печень 3 этапа аэробного катаболизма ЖК: 1) β-окисление ЖК – специфический путь катаболизма; 2) Цикл Кребса – часть ОПК, в котором окисляются молекулы Ацетил-KoA, образованные в ходе βокисления; 3) ЦПЭ и сопряженное с ней окислительное фосфорилирование АДФ. Обеспечивают: окисление NADH и FADH2, образованных в ходе 1) и 2) этапов и синтез АТФ. 1 этапу предшествуют: а) активация жирной кислоты; б) транспорт митохондрий. ацильного остатка в матрикс а) Активация ЖК происходит в цитозоле: ! В этой реакции реально тратится 2 АТФ → 2-ая АТФ необходима для превращения АМФ в АДФ: б) Далее происходит транспорт Ацил-KoA в матрикс митохондрий. Внутренняя мембрана митохондрий непроницаема для Ацил-KoA, поэтому он переносится через внутреннюю мембрану митохондрии в комплексе с молекулой карнитина. Карнитин образуется в клетках всех органов и тканей, но особенно много его содержится в мышцах. Образование комплекса Ацильного карнитином катализирует Карнитинацилтрансфераза I. остатка с фермент: Ацил-KoA в матриксе митохондрий вступает в β-окисление. Транспорт Ацил-KoA в (альтернативная схема): матрикс митохондрий β-окисление жирных кислот ! β-окисление – циклический процесс. За каждый цикл (кроме последнего) углеродный скелет ЖК укорачивается на 2 C-атома. Отрываемый диуглеродный фрагмент: Ацетил-KoA. В последнем цикле: получается сразу 2 Ацетил-KoA. => Количество циклов β-окисления составляет: , где n – общее число С-атомов в молекуле ЖК. Кол-во Ацетил-KoA = кол-ву циклов + 1 = Σ . Кол-во АТФ, образующегося при β-окислении равно: 5 – кол-во АТФ, образованного в каждом цикле (FADH2 → 2 АТФ и 1 NADH + H+ → 3 АТФ) => 2 +3 = 5 АТФ 2 – энергия 2-х макроэргических связей используется на активацию ЖК в цитозоле. АТФ Σ Кол-во АТФ, образующегося при полном аэробном катаболизме ЖК до CO2 и H2O: 12 – кол-во АТФ, которое образуется при окислении Ацетил-KoA в цикле Кребса. Кол-во АТФ при окислении ненасыщенной ЖК: где m – кол-во = связей в молекуле ЖК. Регуляция β-окисления Скорость окисления жирных кислот зависит от: а) V цикла Кребса; б) отношения NADH / NAD+ (потребности клетки в энергии); в) поступления О2 в ткани (гемоглобин, эффект Бора); г) доступности коферментов: (витамин РР) – NAD+ – FAD ( В2) – НSКоА ( В5 – пантотеновая кислота) д) доступности субстрата – Ацил-KoA. Регуляторный фермент Карнитинацилтрансфераза I. β-окисления => регулируется не собственно β-окисление, транспорт ацильного остатка из цитозоля митохондрии. – а в Активаторы: АДФ, АМФ, Ацил-KoA Ингибиторы: АТФ, Малонил-KoA (в основном: в печени). Гормональная регуляция окисления ЖК в печени В клетках печени одновременно происходит и синтез и окисление ЖК. Малонил-KoA (основной субстрат синтеза ЖК) → основной аллостерический ингибитор Карнитинацилтрансферазы I. Кол-во Малонил-KoA регулируется гормонами: Адреналином, Глюкагоном и Инсулином. 1. ситуация: В постабсорбтивный период, при голодании и при стрессе: 2. ситуация: В абсорбтивный период: При участии Инсулина происходит дефосфорилирование Ацетил-KoA-карбоксилазы. => ↑ V синтеза ЖК => ↑ конц-ия Малонил-KoA => ингибируется Карнитинацилтрансфераза I => ↓ V βокисления ЖК. Кетоновые тела 1. Синтез кетоновых тел. В синтез кетоновых тел включается часть молекул Ацетил-KoA, которые образуются в ходе β-окисления. Синтез кетоновых тел происходит в митохондриях печени: ! Конц-ия в крови β-гидроксибутирата >> конц-ии Ацетоацетата, т.к. равновесие реакции: → β-гидроксибутират сдвинуто → из-за Ацетоацетат ← ↑ конц-ии NADH, образующегося в ходе β-окисления. Основные Кетоновые тела: Ацетоацетат и βгидроксибутират. 3-е кетоновое тело – ацетон тканями не используется. В норме: синтез кетоновых тел незначительно ↑ в постабсорбтивный период. Их конц-ия в крови при этом составляет ~ 1-3 мг/дл. Синтез кетоновых тел ↑↑: а) при голодании; б) длительной физической нагрузке (стрессе); в) диете, богатой ТАГ, но с низким кол-вом углеводов ("кремлевская" диета); г) сахарном диабете. Конц-ия кетоновых тел в крови: При длительном голодании ~ 50 мг/дл При сахарном диабете ~ 300 мг/дл Регуляция синтеза кетоновых тел: При голодании и физической нагрузке: Гормоны Глюкагон (или Адреналин) через АЦ-систему ↑ распад жира из жировой ткани → → ЖК поступают в кровь и транспортируются в комплексе с альбумином в печень → → В печени ↑ V β-окисления ЖК и образуется большое кол-во Ацетил –KoA и NADH (↑ Ацетил –KoA; ↑ NADH). Ацетил-KoA в печени может вступать в 2 процесса: 1. Синтез кетоновых тел (происходит только в печени); 2. Цикл Кребса Но, в условиях голодания (стресса): V цикла Кребса в печени существенно снижена по 2 причинам: 1) Ингибирование регуляторных ферментов цикла Кребса ↑ конц-иями АТФ и NADH (аллостерическое ингибирование) 2) Утечка оксалоацетата из митохондрий в цитозоль клеток печени на глюконеогенез => ↑ накопление Ацетил-KoA и ↓ конц-ия Оксалоацетата. Из 1 и 2 причин => избыток Ацетил-KoA используется на синтез кетоновых тел. Митохондрии печени Голодание, стресс: Синтез кетоновых тел β -окисление ЖК Ацетил-KoA ЦТК Пируват Оксалоацетат NADH + H+ NAD+ Малат Внутренняя мембрана митохондрии Цитозоль печени Малат NAD+ Глюконеогенез Глюкоза NADH + H+ Оксалоацетат Регуляторный фермент синтеза кетоновых тел: ГМГKoA-синтаза. Ингибитор фермента: HS-KoA. При голодании и стрессе: при участии Глюкагона (Адреналина) происходит мобилизация ТАГ из жировой ткани => ↑ поток ЖК в печень и образуется Ацил-KoA. Далее происходит перенос ацильных остатков через внутреннюю мембрану митохондрий в матрикс. В матриксе митохондрий: HS-KoA используется в процессе β-окисления, а также происходит взаимодействие Ацил-карнитина с HS-KoA => => ↓ конц-ия HS-KoA => => не происходит ингибирования ГМГ-KoA-синтазы и происходит ↑ V синтеза кетоновых тел. Окисление (аэробный катаболизм) кетоновых тел Кетоновые тела: Ацетоацетат и β-гидроксибутират транспортируются из печени в кровь. КТ – гидрофильные молекулы => транспортируются по крови без участия переносчиков. При избыточном накоплении КТ в крови развивается кетоацидоз, т.к. они – легко диссоциирующие кислоты: ↑ конц-ия кетоновых тел в крови – кетонемия ↑ конц-ия кетоновых тел в моче – кетонурия ↑↑↑ конц-ия кетоновых тел в крови – ацидоз, ↓ pH крови. ! Если pH крови ↓ от 7.2-7.4 до 6.8-7.0 => нарушаются функции белков и ферментов крови => развивается кома и возможен летальный исход. КТ активно окисляются в большинстве тканей организма. Окисляются кетоновые тела в митохондриях: В отличие от ЖК, они проходят ГЭБ и могут использоваться нервной тканью при длительном голодании. Не используют КТ: а) Печень → нет фермента ацетоацетаттрансферазы; Сукцинил-KoA- б) Эритроциты → нет митохондрий. Подсчет кол-ва АТФ при окислении кетоновых тел: 1) β-гидроксибутират: 1 р-ция – 3 моль АТФ. 1 моль АТФ тратится на активацию Сукцината. Из 2 моль Ацетил-KoA: в цикле Кребса образуется – 2*12=24 моль АТФ. Σ выход АТФ = 3 – 1 + 2*12 = 26 моль АТФ. 2) Ацетоацетат: Σ выход АТФ = – 1 + 2*12 = 23 моль АТФ. ЭЙКОЗАНОИДЫ ЭЙКОЗАНОИДЫ – это большая группа регуляторных молекул (паракринных гормонов). Они образуются и функционируют практически во всех органах и тканях организма. Субстраты для синтеза эйкозаноидов: Полиеновые ЖК с 20 С-атомами: Эйкозатриеновая 20:3 Эйкозатетраеновая 20:4 (Арахидоновая) ω6 жирные к-ты Эйкозапентаеновая 20:5 – ω3 жирная кислота. Основное кол-во эйкозаноидов образуется из арахидоновой к-ты. При употреблении рыбьего жира ↑ синтез эйкозаноидов из эйкозапентаеновой к-ты. Синтез эйкозаноидов Выделяют 2 пути синтеза эйкозаноидов: 1) Циклооксигеназный путь → образуются простагландины (PG) и тромбоксаны (TX). 2) Липоксигеназный путь → образуются лейкотриены (LT). Буквы: I D E F A характеризуют структуру заместителя в цикле PGE1 ← PGE2 ← кол-во двойных связей в боковых цепях PG зависит от структуры ЖК – предшественника. 20:3 → PGE1 20:4 → PGE2 20:5 → PGI3 Эйкозаноиды образуются в очень малых количествах, время их жизни – минуты или даже секунды. Эйкозаноиды секретируются в межклеточное пространство и взаимодействуют с рецепторами близлежащих клеток: Паракринный и аутокринный механизмы регуляции: Эйкозаноиды регулируют: 1. тонус ГМК (гладкомышечных клеток) => влияют на: а) АД (артериальное давление); б) состояние бронхов, кишечника 2. функции макрофагов, поэтому развитие воспалительной реакции стимулируют 3. активность тромбоцитов, которые участвуют в процессе свертывания крови; 4. агрегацию и хемотаксис лейкоцитов. Роль эйкозаноидов в регуляции свертывания крови: Типы тромбоксанов и простагландинов, образованных из ω3 и ω6 жирных кислот: Арахидоновая к-та (20:4) ω-6 PGH2 TXA2 Образуются в тромбоцитах, стимулируют агрегацию тромбоцитов и сужение сосудов PGI2 Простациклин PGD2 PGE2 PGF2 Образуются в клетках эпителия, сужение сосудов, тормозят агрегацию тромбоцитов Эйкозапентаеновая к-та (20:5) ω-3 PGH3 TXA3 Образуются в тромбоцитах, стимулируют агрегацию тромбоцитов и сужение сосудов PGI3 Простациклин PGD3 PGE3 PGF3 Образуются в клетках эпителия, сужение сосудов, тормозят агрегацию тромбоцитов Поэтому людям с повышенным риском тромбообразования рекомендуют принимать пищу (или препараты), содержащие ω-3 жирные кислоты. Обмен холестерола Холестерол – основной стероид организма человека. Основные функции холестерола: 1) компонент мембран, отвечает за вязкость гидрофобного слоя; 2) компонент монослоя липидов на поверхности липопротеинов; 3) предшественник желчных к-т, стероидных гормонов, витамина D3. В организме человека содержится 140 г холестерола и его эфиров. Ассимиляция пищевого холестерола ХС и эфиры ХС содержатся в продуктах животного происхождения (яичный желток, сливочное масло, мясо и др.) В растительных продуктах холестерола нет. В полости 12-перстной кишки молекулы экзогенного ХС и его эфиров входят в состав больших липидных капель. Ассимиляция поступающего с пищей холестерола и его эфиров включает те же этапы, что и ассимиляция жиров: 1) Эмульгирование при участии солей желчных кислот. 2) Гидролиз эфиров холестерола при участии фермента панкреатического сока холестеролэстеразы: 3) Далее ХС и ЖК включаются в состав смешанных мицелл и проникают в энтероциты. 4) Ресинтез ЭХС в энтероцитах: а) активация жирной кислоты: б) взаимодействие холестерола и ацил-KoA: АХАТ – ацил-холестерол-ацил-трансфераза. 5) ЭХС и небольшое кол-во включаются в состав ХМнезр.. свободного ХС 6) Через лимфу ХМнезр. поступают в кровь, где происходит их «созревание». 7) Зрелые ХМ подвергаются действию ЛП-липазы => образуются ХМост.. 8) ХМост. взаимодействуют с ЛПНП-рецепторами печени и поглощаются печенью с помощью рецептор-опосредованного эндоцитоза. (Лигандами ЛПНП-рецепторов служат белки апо B и апо E). 9) Компоненты ХМост. расщепляются лизосомальными ферментами. Освобождающийся ХС включается в общий фонд ХС в организме. Синтез эндогенного холестерола в печени и транспорт его в ткани Холестерол синтезируется из ацетил-КоА в основном в печени (80% всего ХС). Также ХС синтезируется: в клетках кишечника, коре надпочечников, коже и др. тканях. Исходный субстрат: ацетил-КоА Необходимый кофермент: NADPH. Источник энергии: АТФ. ! Все вещества, необходимые для синтеза ХС образуются в реакциях катаболизма глюкозы. Синтез ХС происходит в абсорбтивный период, в цитозоле и эндоплазматическом ретикулуме клеток печени одновременно с синтезом жирных кислот и ТАГ. Реакции синтеза ХС до образования сквалена происходят в цитозоле, остальные реакции – в мембранном слое ЭР, т.к. сквален и последующие метаболиты не растворимы в воде. Схема синтеза холестерола: Регуляция синтеза ХС в печени: Регуляторный фермент синтеза ХС в печени – ГМГ-КоA-редуктаза. ГМГ-КоA-редуктаза – гликопротеин, пронизывающий мембрану ЭР, активный центр которого выступает в цитозоль. 2 способа регуляции действия ГМГ-КоА-редуктазы: 1) регуляция активности фермента: Фосфорилирование и дефосфорилирование. ГМГ-КоA-редуктаза – активна дефосфорилированной форме и активируется абсорбтивный период при участии инсулина. в в Ингибируется – путем фосфорилирования под действием глюкагона (в постабсорбтивный период и при голодании). 2) регуляция количества фермента: а) регуляция на уровне транскрипции: Стероиды – холестерол и желчные кислоты – подавляют транскрипцию гена ГМГ-КоА-редуктазы и синтез этого фермента. Эстрогены – индукторы синтеза ГМГ-КоАредуктазы – повышают синтез этого фермента. б) регуляция протеолиза ГМГ-КоА-редуктазы: ХС, желчные кислоты, оксипроизводные ХС и кортикостероиды стимулируют протеолиз молекул ГМГ-КоА-редуктазы, т.е. деградацию функционально активного фермента. Большая часть синтезированного ХС и ЭХС удаляется из печени в составе ЛПОНП. ЛПОНП содержат 55% ТАГ, фосфолипиды и апобелки (в основном: апо В-100). В крови ЛПОНП созревают, подвергаются действию ЛП-липазы и превращаются сначала в ЛППП, а затем в ЛПНП. После этого → белки апо E и апо C-II переносятся обратно из ЛПНП в ЛПВП. ЛПНП – основная транспортная форма ХС, в которой он доставляется в ткани. ~ 70% ХС и ЭХС в крови находятся в составе ЛПНП. Из крови ЛПНП поступают в печень (~ 75%) и другие ткани и доставляют в них ХС. Захват ЛПНП из кровотока происходит всеми тканями организма с помощью ЛПНП-рецепторов: ЛПНП-рецепторы помимо ЛПНП могут захватывать из крови ЛППП, ЛПОНП и ХМост.. также ЛПНП-рецепторы взаимодействуют своим N-концевым гидрофобным доменом с белками апоB (B48 и B100) и апоE на поверхности липопротеинов (в основном: ЛПНП). После этого → ЛП поглощаются тканями с помощью рецептор-опосредованного эндоцитоза. «Обратный» транспорт холестерола В «обратном» транспорте ХС, т.е. выведении избытка ХС из тканей и крови в печень, главную роль играют ЛПВП. В печени образуются незрелые ЛПВП – ЛПВПпредшественники (ЛПВПпредш.) Они имеют дисковидную форму и состоят из бислоя фосфолипидов, с включенными в него белкамиапопротеинами (A-I, С-П, Е). ЛПВПпредш. практически не содержат ХС. Вначале, поступая в кровь, ЛПВПпредш. отдают белки апо C-II и апо E ХМ и ЛПОНП. Затем, ЛПВПпредш. принимают холестерол с мембран клеток и поверхности других липопротеинов (в основном: ЛПНП). Для этого к поверхности ЛПВП прикрепляется фермент лецитин-холестерол-ацилтрансфераза (ЛХАТ), поступающий в ЛПВП из крови. Активатор ЛХАТ – белок апо A-I, который располагается на поверхности ЛПВП. ХС перемещается в оболочку ЛПВПпредш. путем облегченной диффузии при участии белка ABC1 (АТФ-связывающий кассетный белок). ХС, поступающий в оболочку ЛПВПпредш. вступает в реакцию этерификации, которую катализирует ЛХАТ: В результате этой реакции образуются ЭХС, которые погружаются в гидрофобное ядро ЛПВП. Таким образом, в оболочке ЛПВПпредш. освобождается место для поступления следующей порции ХС. По мере наполнения гидрофобного ядра эфирами холестерола ЛПВПпредш. приобретают сферическую форму и превращаются в ЛПВП3: Образованный лизолецитин связывается с альбумином и уносится с поверхности ЛПВП3 током крови. Далее происходит обмен ЭХС на ТАГ между ЛПВП3 и ЛПОНП или ЛППП. ЭХС поступают из ЛПВП3 на ЛПОНП или ЛППП с помощью апоD-белка – «белка, переносящего ЭХС» (БПЭХ). А обратно из ЛПОНП или ЛППП – ЛПВП3 получают ТАГ и фосфолипиды. => ЛПВП увеличиваются в размере и превращаются в ЛПВП2. ЛПВП2 подвергаются действию печеночной липазы, которая гидролизует жиры в ЛПВП2, и превращается обратно в ЛПВП3, которые могут продолжать забирать ХС из тканей и ЛП. Поступление ЭХС, собранных с помощью ЛПВП, в печень происходит несколькими путями: 1) ЛПОНП и ЛППП под действием ЛП-липазы превращаются в ЛПНП, которые доставляют ХС в печень; 2) ЛПВП2 взаимодействуют со "скэвенджер"рецепторами печени и ЭХС переносятся непосредственно с ЛПВП2 в гепатоциты после действия на ЛПВП2 печеночной липазы; 3) В меньшей степени: Взаимодействие ЛПВП2 со специфическими рецепторами печени, комплементарными белкам апо E и апо A-I и поглощение их путем рецептор-опосредованного эндоцитоза. Таким образом, ЛПВП освобождают от избытка холестерола клеточные мембраны и липопротеины => снижают вероятность развития гиперхолестеролемии и атеросклероза. Весь избыток ХС, поступившего в печень, выводится из организма с фекалиями в основном в виде желчных кислот. Синтез желчных кислот Желчные кислоты синтезируются в печени из ХС. Синтез происходит в мембранах эндоплазматического ретикулума гепатоцитов, при участии гидроксилаз: В печени образуются первичные желчные кислоты – холевая и хенодезоксихолевая. Далее, они конъюгируются с глицином и таурином, образуя парные (конъюгированные) желчные кислоты – гликохолевую, таурохолевую, гликохенодезоксихолевую и таурохенодезоксихолевую: Конъюгация желчных кислот: Парные желчные кислоты – более амфифильны и у них лучше выражены эмульгирующие свойства, из-за наличия анионных групп, препятствующих слипанию липидных капель. Далее парные желчные кислоты секретируются в желчный пузырь и накапливаются в желчи. При приеме жирной пищи выделяется гормон холецистокинин, который стимулирует сокращение желчного пузыря и желчь поступает в тонкий кишечник, обеспечивая эмульгирование жиров и ЭХС. В нижних отделах тонкой кишки часть желчных кислот под действием ферментов бактерий теряет глицин, таурин, ОН-группу в положении 7 и превращается во вторичные желчные кислоты – литохолевую и дезоксихолевую. Отщепление полярных групп приводит к снижению растворимости кислот и затруднению их всасывания клетками кишечника. Поэтому, с фекалиями в основном удаляются вторичные желчные кислоты. С калом за сутки выводится до 800 мг желчных кислот, остальная часть через воротную вену возвращается в печень. Реабсорбированные вторичные желчные кислоты в печени опять превращаются в первичные и участвуют в эмульгировании ТАГ и ЭХС. Потери желчных кислот в кишечнике восполняется синтезом новых молекул в печени. Энтерогепатическая циркуляция желчных кислот ~ 95% желчных кислот, попавших в кишечник, возвращается в печень через воротную вену, затем опять секретируется в желчь и повторно используется в эмульгировании ТАГ и ЭХС. Каждая образованная молекула желчной кислоты проходит энтерогепатический круг 6—8 раз, прежде чем выведется из организма: Регуляция синтеза желчных кислот Регуляторный фермент синтеза желчных кислот в печени: 7-α-гидроксилаза. 2 способа регуляции действия 7-α-гидроксилазы: 1) регуляция активности фермента: Фосфорилирование и дефосфорилирование. 7-α-гидроксилаза – активна в фосфорилированной форме и активируется в постабсорбтивный период при участии глюкагона. Ингибируется – в абсорбтивный период дефосфорилирования при участии инсулина. путем 2) регуляция количества фермента: регуляция на уровне транскрипции: Холестерол и тиреоидные гормоны – индуцируют транскрипцию гена 7-α-гидроксилазы и повышают синтез этого фермента. Желчные кислоты и эстрогены – репрессируют транскрипцию гена 7-α-гидроксилазы – подавляют синтез этого фермента. Желчнокаменная болезнь При желчнокаменной болезни в желчном пузыре образуются камни в результате осаждения и кристаллизации компонентов желчи. Возникает: из-за нарушения пропорции количества ХС и желчных кислот, входящих в состав мицелл желчи. Основные причины: 1) ↑ активности ГМГ-КоАредуктазы в печени; 2) ↓ активности 7α-гидроксилазы в печени. Также, к болезни может привести: пища, богатая ХС, гиперкалорийное питание, застой желчи в желчном пузыре, нарушение энтерогепатической циркуляции, инфекции желчного пузыря. Встречается 2 типа камней: 1) однородные, состоящие в основном из холестерола (белого цвета); 2) неоднородные – холестерол-билирубиновые, включающие продукты распада гема (от коричневого до черного цвета). Лечение: 1) на начальной стадии – применяют хенодезоксихолевую кислоту, которая медленно растворяет осадок холестерола. Камень в 1 мм3, образованный в основном из ХС, растворяется почти год. Если камни неоднородные, то их удаляют в основном хирургическим путем. У женщин желчнокаменная болезнь встречается в 3-4 раза чаще, чем у мужчин. Причины: эстрогены индуцируют синтез ГМГ-КоАредуктазы, но репрессируют синтез 7α-гидроксилазы в печени => у женщин чаще нарушается правильное соотношение в желчи холестерол – желчные кислоты. Но, у женщин медленнее, чем у мужчин развивается атеросклероз. Это связано с тем, что эстрогены индуцируют синтез ЛПНП-рецепторов, поглощающих ЛПНП из кровотока, что ↓ риск развития атеросклероза. Гиперлипопротеинемии Гиперлипопротеинемии – это повышение содержания липопротеинов в плазме крови выше нормы. Основные гиперлипопротеинемии: Тип и название Генетический Изменения гиперлипопротеинемии дефект липидного обмена Тип I (наследственная Дефект ↑ в крови ХМ и гиперхиломикронемия) структуры ЛП- ЛПОНП, нет риска липазы. Дефект атеросклероза структуры апо C-II (см. 1 лекцию) Тип II (семейная Дефект ↑ концентрации гиперхолестеролемия) рецепторов ЛПНП, ЛПНП или гиперхолестеролемия, мутация гена ранний атеросклероз, апо В-100 ксантоматоз Тип III (семейная Дефект в ↑ концентрации ХМост., комбинированная структуре ЛПОНП, ЛППП, гиперлипидемия, апоЕ, синтез ЛПНП. нарушение удаления изоформы Гиперхолестеролемия, остаточных апоЕ2, которая гипертриацилглицеролипопротеинов из крови) не лемия, ранний взаимодейст- атеросклероз, вует с ксантоматоз рецепторами Типы IV и V Генетически ↑ концентрации (семейная гетерогенная ЛПОНП, ЛПНП, гипертриацилглицероле- группа гипертриациглицеромия) заболеваний. лемия, умеренная Избыточная гиперхолестеролемия. продукция Атеросклероз, ЛПОНП как снижение результат толерантности к гиперинcулине- глюкозе, ксантоматоз мии Наиболее частые гиперхолестеролемии. гиперлипопротеинемии – Причины гиперхолестеролемии: 1) Гиперкалорийное питание – распространенный фактор, т.к. для синтеза ХС необходимы только Ацетил-KoA, АТФ и NADPH. Все эти субстраты образуются из ГЛК => ↑ синтез ХС и ЛПОНП в печени => ↑ [ЛПНП] в крови => развивается гиперхолестеролемия. 2) ↓ с возрастом эффективности регуляции подавления синтеза ХС в печени, поступающим с пищей ХС => ↑ кол-во ХС в печени и ↑ синтез ЛПОНП => ↑ [ЛПНП] в крови => развивается гиперхолестеролемия. 3) При недостатке фосфатидилхолина может замедляться формирование в печени ЛПВП, что приведет к ↑ содержания в крови ЛПНП, перегруженных холестеролом, и гиперхолестеролемии. 4) Наследственный дефект ЛХАТ => нарушается способность ЛПВП забирать ХС из тканей и ЛП => ↑ риск развития гиперхолестеролемии. 5) Семейная гиперхолестеролемия – наследственный дефект ЛПНП-рецепторов или мутация в гене апо B-100. При этом заболевании наблюдается атеросклероз уже в детском возрасте. 6) Вторичные гиперхолестеролемии – химические модификации липидов и белков ЛПНП и ЛПНПрецепторов. Типы химических модификаций: а) гликозилирование апоВ-100 в составе ЛПНП или апоВ-100-рецепторов. Модифицированные ЛПНП* не узнаются рецепторами и остаются в крови. Очень активно протекает этот процесс при 1 типе сахарного диабета. б) перекисное окисление липидов (ПОЛ) ЛПНП. Происходит ПОЛ ЖК в составе липидов ЛП => образуются свободные радикалы => образуются гидроперекиси полиненасыщенных ЖК фосфолипидов, ТАГ и ЭХС => гидроперекиси нарушают структуру апо B-100 в составе ЛПНП и их взаимодействие с ЛПНП-рецепторами. АТЕРОСКЛЕРОЗ Гиперхолестеролемии создают повышенную опасность заболевания атеросклерозом. Атеросклероз – заболевание, при котором поражается внутренний слой артерий за счет отложения ХС в интиме сосуда. Для оценки риска развития атеросклероза рассчитывают коэффициент атерогенности: Kатер.= XCобщ. ХСЛПВП Утром натощак: Kатер. ≈ ХСЛПВП ХСЛПНП ХСЛПВП В норме: Для людей в возрасте 20-30 лет: Катер. = 2.0 – 2.8 Для людей старше 30 лет: Катер. = 3.0 – 3.5 При ишемической болезни атеросклероза): Катер. = 4.0 – 7.0 сердца (осложнение Патогенез развития атеросклероза: 1) Вследствие гиперхолестеролемий ↑ содержание ЛПНП в крови => ↑ время жизни ЛПНП; 2) ↑ содержание в крови поврежденных в результате ПОЛ и гликозилирования ЛПНП*; 3) ↓ поглощение ЛПНП* клетками тканей; 4) ↑ поглощение ЛПНП* макрофагами с помощью скэвенджер-рецепторов; 5) перегруженные холестеролом макрофаги превращаются в «пенистые» клетки: 6) «пенистые клетки» проникают через промежутки между клетками эндотелия в интиму сосудов; 7) при ↑ кол-ва «пенистых» клеток происходит повреждение эндотелия; 8) => происходит активация тромбоцитов. 9) Тромбоциты: а) секретируют ТХА2, который ↑ агрегацию тромбоцитов и вызывает сокращение стенки сосуда => может образоваться тромб в области атеросклеротической бляшки. б) начинают продуцировать пептид – тромбоцитарный фактор роста, который ↑ пролиферацию ГМК. 10) ГМК мигрируют из медиального слоя во внутренний слой артериальной стенки и способствуют росту бляшки; 11) бляшка прорастает (коллагеном, эластином); фиброзной тканью Происходит некроз клеток под фиброзной оболочкой, отложение ХС в межклеточном пространстве. В центре бляшки образуются даже кристаллы ХС. 12) Накопление в бляшке омертвевших тканей. Бляшка пропитывается солями Ca и становится очень плотной. Разрыв оболочки бляшки вызывает кровотечение, происходит быстрое образование тромба, закрывающего сосуд. Если эти события произошли в коронарной артерии, развивается инфаркт миокарда, в артериях мозга – инсульт, в сосудах десны – пародонтоз. Способы лечения атеросклероза: Подходы к снижению уровня холестерола в крови: 1) Диета с низким кол-вом ХС; 2) Гипокалорийная диета с низким содержанием углеводов и липидов => ↓ синтез ХС в печени и ↓ [ЛПНП] в крови; 3) Прием витаминов С, Е, А, которые снижают ПОЛ и замедляют образование ЛПНП* с повышенной атерогенностью; 4) Использование препаратов, например, «Омакор», содержащих ω-3 жирные кислоты – эйкозапентаеновую (С20:5) и доказапентаеновую (С22:5), так как они являются предшественниками простациклинов (РGI3) и тромбоксанов (ТХА3), которые снижают риск тромбообразования. 5) Использование препаратов, адсорбирующих желчные кислоты в кишечнике и «размыкающих» цикл энтерогепатической циркуляции, например «Холестид», «Холестирамин» («секвестранты желчных кислот»). 6) Использование ниацина (никотиновой кислоты и ее производных), который уменьшает образование ЛПОНП и липолиз в жировой ткани => ↓ [ЛПНП] в крови и замедляется ПОЛ; 7) Фибраты, которые, действуя через рецепторы, ↑ катаболизм ЛПОНП, увеличивая синтез ЛПлипазы; ↑ синтез апоА-I, апоА-П, ↓ содержание ТАГ в крови и ↑ содержание ЛПВП. 8) При хронической гиперхолестеролемии и атеросклерозе назначают статины – обратимые конкурентные ингибиторы ГМГ-КоА-редуктазы (Мевакор, Закор и др.). Статины полностью подавляют синтез ХС в печени. => в печени ↓ образование ЛПОНП и => ↓ образование из них ЛПНП в крови. Также, в печени ↑ синтез RЛПНП и ↑ эндоцитоз ЛПНП. Дополнение к лекции: «Развитие атеросклеротической бляшки в клетках эндотелия кровеносных сосудов» (схема из большого синего (зеленого) учебника): Обмен аминокислот Функции аминокислот: 1) Структурная – синтез белков организма (до 400 г/сут). (Кол-во белков взрослого человека ~ 12-15 кг). 2) Анаболическая – предшественники биологически активных соединений (гормоны, нуклеотиды, гем, креатин, карнитин, фосфолипиды, нейромедиаторы и др.) 3) Энергетическая – источники энергии в экстремальных условиях (длительное голодание, избыточное потребление белков с пищей). Источники свободных аминокислот в клетках: Белки пищи (основной источник), распад собственных белков тканей, синтез аминок-т из глюкозы и метаболитов ОПК. Биологическая роль аминокислот: Пул свободных аминокислот организма составляет 30—100 г, содержание их в крови — в среднем 35— 65 мг/дл. Азотистый баланс ~95% всего аминокислоты. азота организма содержат Поэтому, состояние белкового и аминокислотного обмена оценивается по азотистому балансу. Азотистый баланс – это разница между кол-вом азота, поступающим с пищей, и выводимым из организма. N выводится из организма в виде мочевины и аммонийных солей почками. Виды азотистого баланса Нулевой Отрицательный Положительный 1) Нулевой (азотистое равновесие) – количество азота, поступающего в сутки с пищей равно количеству азота, выводимого из организма. Характерен для взрослого здорового человека при сбалансированном питании. – количество азота, 2) Положительный поступающего в сутки с пищей больше, чем выводится из организма. Характерен для детей, а также для людей, выздоравливающих после тяжелого заболевания. Также встречается при обильном белковом питании; в период беременности. Кроме того, характерен при восстановлении после предшествующего голодания. – количество азота, 3) Отрицательный выводимого в сутки из организма превышает его поступление с пищей. Характерен при тяжелом заболевании, при старении, а также при голодании. Для поддержания азотистого равновесия: минимальное кол-во белков в пище ~ 30—50 г/сут; при большой физической нагрузке ~ 120—150 г/сут. Норма: ~ 100г/сут. Пищевая ценность белков Выделяют 4 группы, на которые делятся все аминокислоты по возможности синтеза в организме: 1) Заменимые аминокислоты – синтезируются в организме в кол-ве достаточном для нужд организма: Ала, Про, Гли, Сер, Асн, Асп, Глн, Глу 2) Незаменимые аминокислоты – не синтезируются в организме: Вал, Лей, Иле, Мет, Фен, Три, Тре, Лиз 3) Частично-заменимые аминокислоты – синтезируются в организме, но в кол-ве, недостаточном для нужд организма: Арг, Гис 4) Условно-заменимые аминокислоты – синтезируются из незаменимых аминокислот: Цис (из Мет), Тир (из Фен) Пищевая ценность белков определяется способностью перевариваться в ЖКТ и содержанием незаменимых аминокислот. Например, белки яиц, перевариваются полностью; молока, мяса Растительные белки, особенно злаков, полностью, как правило, не перевариваются. Фибриллярные белки (волос, шерсти, ногтей и т.п.) не перевариваются. Белки, способные перевариваться в ЖКТ, по пищевой ценности делятся на: 1) Полноценные – белки, в которых содержатся все незаменимые и частично заменимые аминокислоты. (Многие животные белки: белки мяса, яичный альбумин, казеин молока) 2) Неполноценные – белки, в которых нет хотя бы одной незаменимой аминокислоты. (В основном, растительные белки, в которых редко встречаются: Мет, Лиз, Три, Тре) Переваривание белков В пище очень мало свободных аминокислот, поэтому, они поступают в ЖКТ в составе белков. В ходе переваривания белков в ЖКТ образуются свободные аминок-ты. Переваривание белков начинается в желудка и продолжается в тонкой кишке. полости Реакции расщепления пептидных связей катализируют ферменты пептидгидролазы (пептидазы). Они синтезируются в клетках желудка, поджелудочной железы и тонкой кишки. Пептидазы обладают относительной субстратной специфичностью, но преимущественно гидролизуют пептидные связи, образованные определенными аминокислотами. Все пептидазы делятся на 2 группы в зависимости от места расположения в пептиде гидролизуемой связи: Группы пептидаз Эндопептидазы Экзопептидазы 1) Эндопептидазы расщепляют пептидные связи, удаленные от концов пептидной цепи (пепсин, трипсин, химотрипсин, эластаза) 2) Экзопептидазы действуют на пептидные связи, образованные концевыми остатками аминокислот. (карбоксипептидазы А и В, аминопептидаза) Пептидазы, которые действуют в желудке и полости кишечника синтезируются в секреторных клетках в неактивной форме в виде проферментов. Проферменты выделяются к месту действия в просвет желудка или кишечника, где активируются путем частичного протеолиза. Такой механизм активации протеолитических ферментов защищает секреторные клетки желудка и поджелудочной железы от самопереваривания. Частичный протеолиз: Энтеропептидаза Переваривание белков в желудке: Происходит при участии фермента пепсина: Профермент пепсиноген синтезируется желудочными железами и при поступлении пищи секретируется в полость желудка. В желудочном соке пепсиноген превращается в активный фермент пепсин путем частичного протеолиза. В пищевых белках пепсин гидролизует связи, образованные аминогруппой ароматических аминокислот Фен и Тир, а также Глу и Асп с любой другой аминокислотой. Полипептиды, которые образовались в результате действия пепсина в желудке, поступают в тонкую кишку. Переваривание белков в кишечнике: ! В тонкой кишке происходит бóльшая часть работы по перевариванию белков. Содержимое желудка (химус) поступает в полость 12-перстной кишки. В полости 12-перстной кишки на полипептиды действует набор панкреатических пептидаз. Ферменты – панкреатические пептидазы: трипсин, химотрипсин, эластаза, карбоксипептидазы А и В синтезируются в клетках поджелудочной железы в виде неактивных проферментов. Активация ферментов – панкреатических пептидаз происходит в полости 12-перстной кишки путем частичного протеолиза. Основная панкреатическая пептидаза – трипсин. 2 функции трипсина Участвует в гидролизе пищевых белков Активирует все остальные панкреатические пептидазы В результате совместного действия набора активных панкреатических пептидаз образуются: ди- и трипептиды, аминокислоты и небольшое кол-во коротких пептидов (олигопептидов). Завершающий этап переваривания – гидролиз небольших пептидов до свободных аминокислот происходит в пристеночном слое либо уже в клетках кишечника после их всасывания. ! Ферменты завершающего этапа: аминопептидаза, дипептидаза, трипептидаза синтезируются клетках тонкой кишки сразу в активной форме. в Это связано с тем, что эти ферменты действуют только на короткие пептиды, не оказывая никакого влияния на белки и не опасны для белков кишечника. Аминопептидаза отщепляет N-концевые остатки аминокислот от олигопептидов. Ди- и трипептидазы, расщепляют ди- и трипептиды до свободных аминокислот. Образовавшиеся свободные аминокислоты поступают в кровь воротной вены, затем в клетки печени и затем в другие ткани. В тканях аминокислоты используются для синтеза белков и различных биологически активных в-в или вступают в реакции катаболизма. Трансаминирование аминокислот Трансаминирование – это реакция переноса аминогруппы с аминокислоты на кетокислоту. Аминокислота выполняет функцию донора аминогруппы, кетокислота – функцию акцептора аминогруппы. R1 HC R2 NH2 + COOH Аминотрансфераза R2 R1 + C O C O COOH Ko - Пиридоксальфосфат COOH HC NH2 COOH Трансаминироваться могут все аминок-ты, кроме Лиз, Тре, Про и Гли. Название фермента строится так: сначала название донора аминогруппы (аминок-ты), затем название акцептора аминогруппы (кеток-ты) и затем слово аминотрансфераза (или трансаминаза): Пример: COOH CH3 HC NH2 COOH Аланин + COOH CH2 CH3 C O C O COOH Оксалоацетат COOH Пируват CH2 + HC NH2 COOH Аспартат Название фермента по прямой реакции: Аланин-оксалоацетатаминотрансфераза. По обратной реакции: Аспартат-пируватаминотрансфераза Основной донор аминогруппы: Глутамат; Основной акцептор аминогруппы: α-кетоглутарат. В случае, если акцептор NH2-группы – α-кетоглутарат, то его название не произносится. Например: COOH CH3 HC NH2 COOH аланин + COOH (CH2)2 CH3 C O C O COOH α- кетоглутарат COOH пируват (CH2)2 + HC NH2 COOH глутамат Фермент: Аланинаминотрансфераза (аланинтрансаминаза) (АЛТ) По обратной р-ции: Глутаматпируватаминотрансфераза (ГПТ) COOH COOH CH2 HC NH2 (CH2)2 COOH CH2 C O C O COOH COOH COOH аспартат α- кетоглутарат + COOH (CH2)2 + оксалоацетат HC NH2 COOH глутамат Фермент: Аспартатаминотрансфераза (АСТ) По обратной р-ции: Глутаматоксалоацетатаминотрансфераза (ГОТ) Активность ферментов АЛТ (ГПТ) и АСТ (ГОТ) в крови измеряют для диагностики болезней сердца и печени. В целях энзимодиагностики, определяют соотношение активности АСТ/АЛТ в сыворотке крови – коэффициент де Ритиса: Норма: ~ 1.33 ± 0.42 При гепатите: активность АЛТ > АСТ. Коэффициент де Ритиса ↓ до ~ 0.6 При циррозе печени: коэффициент де Ритиса ~ 1.0, т.к. развивается некроз тканей и в кровь поступают 2 фракции фермента ACT — и цитоплазматическая, и митохондриальная. При инфаркте миокарда коэффициент де Ритиса резко ↑. Активность (Е) аминотрансфераз сыворотки крови при остром гепатите: Активность (Е) аминотрансфераз сыворотки крови при инфаркте миокарда: Дезаминирование аминокислот Дезаминирование – это отщепление α-аминогруппы от аминокислоты в виде NH3. Дезаминирование, в отличие от трансаминирования, уменьшает общее кол-во аминокислот. Реакции дезаминирования – это начальный этап катаболизма, общий для всех аминокислот. Дезаминируются все аминокислоты, кроме Лиз, Про и Гли. Типы дезаминирования Прямое Окислительное Неокислительное Непрямое Внутримолекулярное Прямое дезаминирование – отщепление группы происходит непосредственно аминокислоты. Прямому окислительному подвергается только глутамат: NH2от дезаминированию Обе эти реакции – обратимые. Поэтому, при ↑ в клетках концентрации NH3 могут протекать в обратном направлении и использоваться для его обезвреживания. Глутаматдегидрогеназа активна во всех тканях, кроме мышц. Сер и Тре подвергаются неокислительному дезаминированию: прямому Прямому внутримолекулярному дезаминированию подвергается Гис: Гистидин дезаминируется только в печени и коже человека, в крови гистидаза отсутствует. Поэтому, определение активности гистидазы в крови – используется при диагностике вирусных и токсических поражений печени, рака печени и кожи. Большинство аминокислот подвергается в клетках непрямому дезаминированию: В большинстве тканей окислительному пути. протекает по Непрямое дезаминирование включает 2 стадии: 1) Трансаминирование аминокислоты с α-кетоглутаратом с образованием глутамата: COOH R HC (CH2)2 NH2 + COOH R C O C O COOH COOH COOH а/к-та α -КГ кеток-та (CH2)2 + HC NH2 COOH глутамат 2) Прямое окислительное глутамата: дезаминирование Непрямое дезаминирование по неокислительному пути (цикл АМФ-ИМФ) активно протекает в мышцах из-за низкой активности глутаматдегидрогеназы. Обмен аммиака ! В ходе дезаминирования аминокислот в тканях образуется большое кол-во NH3. Также NH3 образуется при распаде нуклеотидов, биогенных аминов и при гниении белков в кишечнике в результате воздействия бактерий. Дезаминирование аминокислот и образование NH3 происходит во всех тканях. В клетках образованный обезвреживанию. NH3 подвергается Выводится из организма NH3 почками в виде мочевины и солей аммония. Мочевина образуется в печени; Соли аммония – образуются в самих почках. Источники и продукты обезвреживания NH3 в организме: Механизм токсического действия аммиака NH3 при его избытке в организме оказывает токсическое действие. Особенно сильно он воздействует на мозг и ЦНС. Состояния, при которых повышена конц-ия NH3 в крови, называются гипераммониемии. Симптомы гипераммониемии: головокружение, галлюцинации, тошнота, рвота, нечленораздельная речь, судороги, потеря сознания и др. Причины токсического действия NH3: α-КГ, связывая избыток NH3, превращается в Глу => снижается концентрация α-КГ: Это вызывает: 1) ↓ скорости цикла Кребса, что вызывает гипоэнергетическое состояние. 2) Нарушается трансаминирование аминокислот и синтез ряда биогенных аминов (ацетилхолин, дофамин и др.) Высокие концентрации NH3 стимулируют синтез Глн из Глу в нервной ткани: Глу ↓ + NH3 ↑ + АТФ → Глн ↑ + АДФ + H3PO4 Это вызывает: 1) Накопление глутамина в клетках нейроглии приводит к повышению в них осмотического давления => происходит набухание астроцитов. Большие концентрации глутамина могут вызвать отек мозга. 2) Уменьшение концентрации Глу ↓ => подавляется синтез ряда нейромедиаторов и особенно γ-аминомасляной к-ты (ГАМК): Глутаматдекарбоксилаза Глу ↓ ⎯⎯⎯⎯⎯⎯⎯⎯→ ГАМК ↓ + CO2 ГАМК – основной тормозной медиатор. Его недостаток нарушает проведение нервного импульса => повышается нервно-мышечная возбудимость и возникают судороги. ↑ концентрации NH3 в крови сдвигает рН в щелочную сторону (вызывает алкалоз): H+ ↓ + NH3 ↑ → NH4+ ↑ => нарушается эффект Бора: H+ ↓ + Hb(O2)4 → H+Hb + 4O2 ↓ Происходит увеличение сродства гемоглобина к O2, что нарушает поступление O2 в ткани и приводит к гипоксии. Из-за этого, также происходит накопление CO2 в тканях. ↑ конц-ии ионов NH4+ в крови нарушает трансмембранный перенос ионов Na+ и К+ из-за конкуренции с ними за ионные каналы. Это также нарушает проведение нервных импульсов. Способы обезвреживания аммиака в разных тканях: Основная реакция связывания NH3, протекающая во всех тканях организма – это синтез глутамина из глутамата: Глутаминсинтетаза расположена в митохондриях клеток. Кофактор: ионы Mg2+. Через клеточные мембраны глутамин легко транспортируется в кровь путём облегчённой диффузии. Больше всего глутамина образуется в мышцах и мозге. С током крови глутамин транспортируется в кишечник, почки и в небольшом кол-ве в печень. Обезвреживание NH3 в почках: В почках происходит гидролиз глутамина до глутамата с образованием NH3: Образующийся NH3 используется на синтез солей аммония, которые выделяются с мочой из организма. ! Помимо обезвреживания NH3, эта реакция имеет более важное значение – компенсация ацидоза. При ацидозе избыток кислот поступает в почки. Избыток H+ в почках индуцирует синтез глутаминазы => образующийся NH3 нейтрализует кислоты, образуя соли аммония. При дефекте глутаминазы в почках, для выведения избытка кислот используются ионы Na+ и K+, что может привести к их потере. Обезвреживание NH3 в кишечнике: В клетках кишечника также происходит гидролиз глутамина до глутамата с образованием NH3 при участии глутаминазы: Глн + H2O → Глу + NH3 ~ 5% образовавшегося кишечник. NH3 Небольшая часть попадает в печень. через NH3 удаляется через воротную вену Остальные ~ 90% NH3 поступают в почки и выводятся с мочой в виде солей аммония. Глутамат трансаминируется с пируватом при участии АЛТ: COOH COOH (CH2)2 HC NH2 COOH глутамат + CH3 (CH2)2 C O C O COOH пируват CH3 + COOH кетоглутарат HC NH2 COOH аланин α-Аминогруппа включается в аланин, который поступает из кишечника в кровь воротной вены и поглощается печенью. Обезвреживание NH3 в мозге и некоторых других тканях: В мозге и некоторых других тканях NH3 может связываться α-кетоглутаратом с образованием глутамата. Фермент: глутаматдегидрогеназа. Но, в норме эта реакция протекает медленно, т.к. используется в основном для дезаминирования глутамата. Глутамат может связывать еще одну молекулу NH3, превращаясь в глутамин: При гипераммониемии, эти реакции очень важны, т.к. за их счет обезвреживается сразу 2 молекулы NH3. Обезвреживание NH3 в мышцах: В мышцах очень глутаматдегидрогеназы. низкая активность Поэтому, основной способ обезвреживания NH3 в мышцах – глюкозо-аланиновый цикл. Глюкозо-аланиновый цикл – это образование аланина в мышцах, его транспорт в печень для синтеза глюкозы, а также поступление глюкозы, синтезированной в печени, в обратном направлении. Образование аланина – это дополнительный механизм выведения азота аминокислот из клеток. Глюкозо-аланиновый цикл: Обезвреживание NH3 в печени: Самые большие кол-ва NH3 обезвреживаются в печени. В печени NH3 в основном образуется: при дезаминировании аминокислот и при участии глутаминазы. Часть NH3 поступает с кровью воротной вены из кишечника. В печени NH3 связывается с CO2 с образованием карбамоилфосфата: Продукт реакции – карбамоилфосфат вступает затем в орнитиновый цикл. Орнитиновый цикл. ~ 90% N удаляется из организма с мочой почками в составе мочевины. Синтез мочевины происходит в печени в процессе, который называется орнитиновый цикл (цикл Кребса – Гензелейта). Орнитиновый цикл – это циклический процесс. Ключевое соединение, замыкающее цикл – αаминокислота орнитин. Источники N в молекуле мочевины: Молекула мочевины содержит 2 атома азота: 1 атом: образуется из NH3 2 атом: образуется из аспартата И NH3 и аспартат образуются из атомов N 2-х разных аминокислот: (2 версия схемы) Основные функции орнитинового цикла: 1) включение N аминокислот в мочевину, которая экскретируется и предотвращает накопление токсичного NH3. 2) синтез частично-заменимой аминокислоты аргинина. Энергетический эффект орнитинового цикла: В реакциях орнитинового цикла расходуется 4 макроэргические связи 3-х молекул АТФ на каждый оборот цикла. АТФ также расходуется при трансмембранном переносе веществ, связанном с синтезом и экскрецией мочевины. Пути компенсации энергозатрат: В цикле регенерации аспартата из фумарата на стадии дегидрирования малата в ЦТК образуется NADH => образуется 3 моль АТФ путём окислительного фосфорилирования при участии ЦПЭ. При окислительном дезаминировании глутамата в печени и других органах также образуется NADH => еще 3 моль АТФ образуется путем окислительного фосфорилирования. Гипераммониемии Гипераммониемия – это повышенное содержание NH3 в крови. Причины гипераммониемии: заболевания печени (гепатит, цирроз) или наследственный дефект ферментов орнитинового цикла. 5 основных типов гипераммониемий связаны с наследственным дефектом одного из 5 ферментов орнитинового цикла: 1) гипераммониемия I типа – дефект фермента: карбамоилфосфатсинтетазы I; 2) гипераммониемия II типа – дефект фермента: орнитинкарбамоилтрансферазы; 3) цитруллинемия – дефект аргининосукцинатсинтетазы; фермента 4) аргининосукцинатурия – дефект фермента аргининосукцинатлиазы; 5) гипераргининемия аргиназы. – дефект фермента При всех этих болезнях наблюдается накопление метаболитов, предшествующих ферментному блоку, а также значительное накопление в крови NH3. Пути обмена безазотистого остатка аминокислот В ходе катаболизма от аминокислоты отщепляется аминогруппа в виде NH3 и образуется безазотистый остаток – как правило: кетокислота. Далее безазотистый остаток аминокислот превращается в один из 6 метаболитов, вступающих в ОПК: пируват, ацетил-KoA, αкетоглутарат, сукцинил-KoA, фумарат, оксалоацетат. По судьбе безазотистого остатка аминокислоты делятся на 3 группы: все Гликогенные: безазотистый остаток аминокислоты может включаться в глюконеогенез. Кетогенные: безазотистый остаток аминокислоты превращается в Ацетил-KoA или ацетоацетат и может использоваться в синтезе кетоновых тел. Или сгорает в цикле Кребса до CO2. Смешанные (глико-кетогенные): распадаются на 2 продукта. Один включается в глюконеогенез, 2 может использоваться в синтезе кетоновых тел. Чисто кетогенных аминокислот всего 2: лейцин и лизин. Смешанные аминокислоты: изолейцин, триптофан, фенилаланин, тирозин. Все остальные аминокислоты: чисто гликогенные. Выделяют 5 анаплеротических реакций (путей включения аминокислот в глюконеогенез): 1) через пируват; 2) через α-кетоглутарат; 3) через сукцинил-KoA; 4) через Фумарат; 5) через Оксалоацетат Синтез заменимых аминокислот В организме человека может синтезироваться 8 заменимых аминокислот: Ала, Про, Гли, Сер, Асн, Асп, Глн, Глу. Сначала из глюкозы образуется α-кетокислота – углеродный скелет. Затем в α-кетокислоту вводится α-аминогруппа в результате реакций трансаминирования: Глутамат: COOH R HC NH2 + COOH (CH2)2 C O Аминотрансфераза R C O + COOH COOH Аминок-та COOH α -КГ (CH2)2 HC NH2 COOH Кеток-та Глутамат Аланин: COOH (CH2)2 HC NH2 COOH Глутамат CH3 + C O COOH Пируват COOH АЛТ (CH2)2 C O COOH α -КГ CH3 + HC NH2 COOH Аланин Аспартат: COOH (CH2)2 HC NH2 COOH Глутамат COOH + CH2 COOH COOH АСТ (CH2)2 + CH2 HC NH2 C O C O COOH COOH α -КГ Аспартат под действием Оксалоацетат Глутамин: Синтезируется из глутамата глутаминсинтетазы: COOH Аспарагин: Синтезируется из аспартата и глутамина под действием аспарагинсинтетазы: Серин образуется из 3-фосфоглицерата промежуточного продукта гликолиза. – Глицин синтезируется из серина. Пролин синтезируется из глутамата: Глутамат → γ-Полуальдегид глутамата → Пролин Частично заменимые аминок-ты: Аргинин образуется в орнитиновом цикле. Гистидин синтезируется из АТФ и рибозы. Условно заменимая аминок-та Цистеин образуется из серина и метионина. Обмен отдельных аминокислот Обмен Серина и Глицина 1) Синтез серина из глюкозы: 2) Синтез глицина из серина: Глицин – заменимая аминокислота, синтезируется из серина с участием кофермента H4-фолата: 3) Катаболизм глицина также происходит с участием H4-фолата: H4-фолат – это производное фолиевой кислоты. Фолиевая к-та – витамин Bc (или B9) поступает с пищей, а также синтезируется микрофлорой кишечника. Фолиевая кислота: H4-фолат: H4-фолат выполняет роль кофермента в реакциях превращения Гли и Сер. ! Главное значение реакций синтеза Гли из Сер и катаболизма Гли – образование Метилен-H4-фолата. образуются другие Из Метилен-H4-фолата производные: Метил-H4-фолат, Метенил-H4-фолат, Формил-H4-фолат и др. ! Все производные H4-фолата – это доноры одноуглеродных фрагментов в синтезе многих соединений. Например: синтез пуриновых и пиримидиновых нуклеотидов; регенерация метионина; синтез ряда биогенных аминов и др. Синтез H4-фолата происходит в печени: Недостаточность фолиевой кислоты Гиповитаминоз фолиевой кислоты у человека возникает редко. Основные причины: 1) нарушение всасывания фолиевой кислоты в кишечнике; 2) снижение активности ферментов фолатредуктазы или дигидрофолатредуктазы, участвующих в синтезе H4-фолата (при гепатите, циррозе печени и др. заболеваниях печени); 3) Недостаточное потребление с пищей овощей, фруктов и мясных продуктов – основных источников фолиевой к-ты; 4) Беременность, при которой увеличена потребность в H4-фолате. При дефиците фолиевой кислоты нарушается обмен одноуглеродных фрагментов и развивается мегалобластная (макроцитарная) анемия. 2-ая причина этого заболевания: недостаток витамина B12, использование которого связано с обменом фолиевой к-ты. Основные симптомы заболевания: уменьшение количества эритроцитов, снижение содержания в них гемоглобина и увеличение размера эритроцитов. Причины возникновения симптомов: нарушение синтеза ДНК и РНК из-за недостатка их предшественников – тимидиловой кислоты и пуриновых нуклеотидов, для синтеза которых необходимы производные H4-фолата. Клетки кроветворных органов быстро делятся. Поэтому, они в 1 очередь реагируют на нарушение синтеза нуклеиновых кислот снижением скорости эритропоэза. Обмен метионина Метионин – незаменимая аминок-та. Используется: в синтезе белка, источник атома S для синтеза цистеина, включается в глюконеогенез. CH3-группа метионина используется в синтезе многих биогенных аминов: Биологическая роль метионина: Активация метионина Метионин играет очень важную роль в реакциях трансметилирования. Его активная форма (SAM) – универсальный донор CH3-группы. ! В SAM, в отличие от метионина, «+» на атоме S. Из-за этого: CH3-группа становится очень подвижной и легко отрывается, переходя на молекулу-акцептор в реакциях трансметилирования. Реакции трансметилирования ферменты: метилтрансферазы. катализируют SAM, отдавая CH3-группу, превращается в Sаденозилгомоцистеин (SAГ). Реакции трансметилирования 1) Синтез фосфатидилхолина: Активно происходит в печени, кишечнике, молочной железе и некоторых др. тканях. 2) Синтез карнитина: Синтез происходит путем метилирования ГАМК с участием 3 молекул SAM: H2N CH2 CH2 CH2 COOH ГАМК 3SAM 3SAГ Метилтрансфераза H3C + H3C N CH2 CH2 CH2 COOH H3C γ -бутиробетаин O2 H3C + H3C N CH2 CH CH2 COOH OH H3C КАРНИТИН 3) Синтез креатина: Креатин синтезируется в 2 стадии в почках и печени при участии 3 аминокислот: аргинина, глицина и метионина: В почках: из аргинина и глицина образуется гуанидинацетат. Гуанидинацетат транспортируется в печень. происходит метилирование В печени: гуанидинацетата при участии SAM и образуется креатин. Креатин с током крови транспортируется в мышцы и клетки мозга, где из него образуется соединение с макроэргической связью – креатинфосфат. Реакция синтеза креатинфосфата легко обратима. Креатинфосфат накапливается в мышечной и нервной тканях и служит резервной формой АТФ. ! В работающей мышце первые секунды работы происходит реакция расщепления креатинфосфата с образованием АТФ при участии креатинкиназы (субстратное фосфорилирование). Чуть позже включается анаэробный гликолиз. И только при дальнейшей работе включаются βокисление жирных кислот и аэробный распад глюкозы. В мышцах: часть креатинфосфата неферментативно превращается в креатинин. Креатинин не используется клетками и выводится с мочой. В норме: с мочой выделяется ~ 1-2 г креатинина в сутки. Определение содержания креатинина в моче используется для характеристики клубочковой фильтрации и диагностики заболеваний почек. Регенерация метионина В клетках организма происходит очень большой расход метионина, т.к. реакции трансметилирования происходят очень интенсивно. Т.к. метионин – незаменимая аминок-та, то большое значение имеет регенерация метионина при участии заменимых аминокислот серина и глицина: 1) В ходе реакций трансметилирования от SAM отщепляется CH3-группа и он превращается в SAГ: R + SAM–S+–CH3 → R–CH3 + SAГ 2) SAГ под действием гидролазы распадается на аденозин и гомоцистеин: SH CH2 Аденозин + SAГ + H2O CH2 HC NH2 COOH Гомоцистеин 3) Гомоцистеин может превращаться в метионин при участии фермента гомоцистеинметилтрансферазы. Метилен-H4-фолат Гли SH CH2 CH2 Метил-H4-фолат HC NH2 COOH Гомоцистеин B12 Сер H4-фолат CH3 S CH2 CH2 HC NH2 COOH Метионин Донор метильной группы в этой реакции – метилH4-фолат. Промежуточный переносчик метильной группы – метилкобаламин (производное витамина B12). 4) Образующийся метионин может снова активироваться и повторно использоваться в реакциях трансметилирования: Метионин + АТФ → SAM + PPi + Pi Обмен Фенилаланина и Тирозина Фенилаланин – незаменимая аминокислота. Используется в организме только в 2-х процессах: Субстрат для синтеза белков и превращается в тирозин. ! Превращение Фен в Тир необходимо в 1 очередь для удаления избытка Фен, т.к. его высокие концентрации токсичны для клеток. В Тир превращается ~ 90% Фен: Превращение Фен в Тир – это 1-ая реакция основного пути метаболизма Фен. Все дальнейшие превращения происходят уже с Тир. в организме При нарушении протекания реакции превращения Фен в Тир возникает заболевание – фенилкетонурия (фенилпировиноградная олигофрения). Из-за невозможности превращения Фен в Тир, катаболизм фенилаланина протекает по альтернативному пути: При этом в крови и моче повышается содержание фенилаланина и метаболитов альтернативного пути: фенилпирувата, фениллактата, фенилацетата и др. Симптомы: Резкое нарушение умственного и физического развития, судорожный синдром, «мышиный» запах. Также встречается: нарушение пигментации кожи. Фен и его производные, при их избытке, оказывают токсическое действие на клетки мозга, поскольку: ограничивают транспорт Тир и Три через гематоэнцефалический барьер и тормозят синтез нейромедиаторов (дофамина, норадреналина, серотонина). Без лечения больные доживают до 30 лет. фенилкетонурией Заболевание наследуется рецессивному типу. по не аутосомно- Выделяют 2 формы фенилкетонурии: 1) Классическая фенилкетонурия: Причина: наследственный дефект фермента фенилаланингидроксилазы. Частота заболевания: 1 случай на ~ 10000 новорожденных. 2) Вариантная фенилкетонурия: (коферментзависимая гиперфенилаланинемия) Причина: мутации в генах, контролирующих метаболизм H4-биоптерина. Встречается: 1-2 случая на ~ 1 млн. новорожденных. H4-биоптерин необходим для гидроксилирования не только Фен, но и Тир и Три, поэтому, при этой форме заболевания нарушен метаболизм всех 3 аминокислот, а также синтез многих нейромедиаторов. При этой форме заболевания возникают тяжелые неврологические нарушения и ранняя смерть. Лечение фенилкетонурии: диета, с почти полным исключением из пищи фенилаланина. ! Начинать: сразу после рождения ребенка. Для диагностики фенилкетонурии определяют концентрацию фенилаланина и патологических метаболитов в крови и моче больного. В генетической консультации, для выявления гетерозиготного носителя заболевания, используют тест толерантности к фенилаланину: Разработаны специальные схемы скрининга для выявления новорождённых детей с ФКУ. В настоящее время для выявления мутантного гена фенилаланингидроксилазы у гетерозиготных носителей ФКУ, используют также ПЦРдиагностику. Особенности обмена тирозина в разных тканях Кроме использования в синтезе белков, Тир в разных тканях используется для синтеза многих биологически-активных соединений. Катаболизм Тир до конечных продуктов происходит в печени. Катаболизм Тирозина в печени: В печени происходит катаболизм Тир до конечных продуктов: Ферменты: 1 – тирозинаминотрансфераза (кофермент: ПФ); 2 – п-гидроксифенилпируватдиоксигеназа (кофакторы: вит. C и Fe2+); 3 – диоксигеназа гомогентизиновой кислоты (кофакторы: вит. C и Fe2+); 4 – фумарилацетоацетатгидролаза. При наследственном дефекте гена фермента диоксигеназы гомогентизиновой кислоты возникает заболевание – Алкаптонурия («чёрная моча»). При этом заболевании с мочой выделяется большое количество гомогентизиновой кислоты. При её окислении O2 воздуха образуются алкаптоны – темные пигменты. Симптомы: 1) Моча приобретает черный цвет (у грудных детей – темные пятна на пеленках); 2) Охронозы – черные пятна в хрящах. (Очень часто: на кончике носа и мочках ушей); 3) Артриты – из-за отложения алкаптонов в суставах. Частота заболевания: 2-5 случаев на ~ 1 млн. новорожденных. Заболевание наследуется по аутосомнорецессивному типу. Диагностических методов выявления гетерозиготных носителей – не найдено. Превращение тирозина в меланоцитах. В меланоцитах (пигментных клетках) Тир превращается в темные пигменты – меланины: При наследственном дефекте гена фермента тирозиназы возникает заболевание – Альбинизм. Симптомы: отсутствие пигментации кожи и волос. У больных часто снижена острота зрения, возникает светобоязнь. Длительное пребывание под открытым солнцем приводит к раку кожи. Частота заболевания: 1 случай на ~ 20 000 человек. Превращение тирозина в надпочечниках и нервной ткани (синтез катехоламинов). В мозговом веществе надпочечников и нервной ткани из тирозина синтезируются катехоламины (дофамин, норадреналин и адреналин): Дофамин и норадреналин – нейромедиаторы разных отделов головного мозга. Участвуют в синаптической передаче нервных импульсов. Адреналин – гормон широкого спектра действия, регулирующий энергетический обмен, работу сердечно-сосудистой системы и др. С нарушением синтеза катехоламинов связано несколько заболеваний: 1. Болезнь Паркинсона (Паркинсонизм) Возникает из-за недостаточности дофамина в черной субстанции мозга. Причины: снижение активности фермента ДОФАдекарбоксилазы, реже – тирозингидроксилазы. Основные симптомы: тремор (непроизвольное дрожание), акинезия (скованность движений), ригидность (напряжение мышц). Паркинсонизм – одно из самых распространенных неврологических заболеваний у людей старше 60 лет. Лечение: Т.к. дофамин не проходит через гематоэнцефалический барьер, то для лечения Паркинсонизма используют: 1) Заместительная терапия: препаратыпредшественники дофамина (производные ДОФА) – леводопа, мадопар, наком и др. 2) Подавление инактивации дофамина: ингибиторы МАО (депренил, ниаламид, пиразидол и др.). 2. Различные депрессивные состояния часто связаны со снижением в нервных клетках содержания дофамина и норадреналина. 3. Шизофрения Одна из причин шизофрении – гиперсекреция дофамина в височной доле мозга. Превращение тирозина в щитовидной железе В щитовидной железе из Тир синтезируются и выделяются гормоны йодтиронины – тироксин (тетрайодтиронин) и трийодтиронин. По химической структуре йодтиронины – это конденсированные йодированные остатки тирозина: OH I OH I I O I O I CH2 H2N CH COOH Тироксин I I CH2 H2N CH COOH Трийодтиронин При нарушении синтеза йодтиронинов возникает гипотиреоз. Причины гипотиреоза: 1) наследственный дефект ферментов, участвующих в синтезе йодтиронинов. (болезни: микседема (слизистый отёк), кретинизм); 2) удаление щитовидной железы при базедовой болезни; 3) недостаток йода в пище и воде в регионе (болезнь: эндемический зоб). Декарбоксилирование аминокислот. Биогенные амины. Большую роль в организме человека играют непептидные производные аминокислот – биогенные амины. Многие биогенные амины образуются декарбоксилировании аминокислот и производных. при их Реакции декарбоксилирования – это реакции отщепления α-карбоксильной группы. Реакции декарбоксилирования – необратимые. Р-ции декарбоксилирования катализируют ферменты – декарбоксилазы (Ko: ПФ): Виды биогенных аминов Гормоны Надпочечников Адреналин Нейромедиаторы Щитовидной железы Медиаторы ЦНС Медиатор воспаления Тироксин, Трийодтиронин Ацетилхолин, ГАМК и др. Гистамин Синтез биогенных аминов. Синтез серотонина: Серотонин – нейромедиатор проводящих путей. Синтезируется из триптофана в надпочечниках, ЦНС и в небольших количествах в слюнных железах (См. схему синтеза серотонина из Три выше). Серотонин — биологически активное вещество широкого спектра действия. Стимулирует сокращение гладкой мускулатуры, перистальтику кишечника, оказывает сосудосуживающее действие, регулирует артериальное давление, температуру тела, дыхание, обладает антидепрессантным свойством («гормон удовольствия») и др. В эпифизе (шишковидной железе) и ряде других тканей серотонин может превращаться в гормон мелатонин. Мелатонин регулирует суточные биоритмы организма человека, обладает мощным антиоксидантным действием, защищает организм от рака, повышает иммунитет и др. Синтез ацетилхолина: Синтезируется в нервной ткани из серина: H2N CH C Этаноламин трансметилаза Сериндекарбоксилаза O OH CH2 H2N CH2 CH2 OH ПФ CO2 Этаноламин 3 SАМ 3 SАГ OH Серин H 3C + H3C N CH2 CH2 OH H 3C Холин Холинацетил трансфераза Ацетил-KoA H 3C + H3C N CH2 CH2 O C CH3 H 3C O HS-KoA Ацетилхолин Ацетилхолин – один из важнейших возбуждающих нейромедиаторов вегетативной нервной системы. Синтез γ-аминомасляной кислоты (ГАМК): ГАМК образуется в нервных клетках в ходе декарбоксилирования глутамата: H2N CH CH2 CH2 COOH Глутамат Глутаматдекарбоксилаза H2N CH2 CH2 CH2 COOH COOH ПФ CO2 ГАМК ГАМК – основной тормозной медиатор высших отделов мозга. ГАМК увеличивает проницаемость постсинаптических мембран для ионов K+, что вызывает торможение нервного импульса. Повышает дыхательную активность нервной ткани, улучшает кровоснабжение головного мозга. Синтез гистамина: Гистамин образуется при декарбоксилировании гистидина в тучных клетках соединительной ткани: Функции гистамина: 1) стимулирует секрецию желудочного сока, слюны, т.е. является пищеварительным гормоном; 2) повышает проницаемость капилляров, вызывает отеки, снижает артериальное давление (но увеличивает внутричерепное давление, вызывает головную боль); 3) сокращает гладкую мускулатуру легких, вызывает удушье; 4) участвует в формировании воспалительной реакции – вызывает расширение сосудов, покраснение кожи, отечность ткани; 5) вызывает аллергическую реакцию; 6) выполняет роль нейромедиатора; 7) является медиатором боли. Инактивация биогенных аминов. 2 основных способа инактивации аминов и некоторых гормонов: биогенных 1) Метилирование с участием SAM под действием метилтрансфераз. Таким способом могут инактивироваться различные биогенные амины, но чаще всего адреналин и гистамин: OH OH OH CH OH SAM SAГ Адреналин-Oметилтрансфераза CH2 NH CH3 Адреналин OH CH O CH3 CH2 NH CH3 Метиладреналин Метилированные производные биогенных аминов обычно теряют биологическую активность, в печени подвергаются конъюгации с глюкуроновой или серной кислотой и выводятся из организма или же окисляются МАО. 2) Окисление ферментами моноаминооксидазами при участии кофермента FAD. Таким путем чаще происходит инактивация дофамина, норадреналина, серотонина, ГАМК. При этом происходит окислительное дезаминирование биогенных аминов с образованием альдегидов, а затем соответствующих кислот, которые хорошо растворимы в воде и выводятся почками: Гормональная р р регуляция у обмена веществ и ф функций й организма Нормальное р функционирование фу р многоклеточного организма, р в основе которого лежит взаимосвязь между отдельными клетками, тканями и органами осуществляется четырьмя основными системами регуляции. • • • • Центральная и периферическая нервные системы через нервные импульсы и нейромедиаторы; Эндокринная система через эндокринные железы и гормоны, которые секретируются в кровь и влияют на метаболизм различных клеток-мишеней; Паракринная и аутокринная системы посредством различных соединений, которые секретируются в межклеточное пространство и взаимодействуют й с рецепторами либо б близлежащих клеток, либо той же клетки (простагландины, гормоны ЖКТ, гистамин и др.); Иммунная система через специфические белки (цитокины, антитела). Системы р регуляции у обмена веществ и фу функций организма р образуют р у три р иерархических уровня. Первый р уровень - Ц ур ЦНС. Нервные р клетки получают у сигналы, поступающие из внешней и внутренней среды, преобразуют их в форму нервного импульса и передают через синапсы, используя химические сигналы - медиаторы. Медиаторы вызывают изменения метаболизма в эффекторных клетках. Второй уровень - эндокринная система. Включает гипоталамус, гипофиз, периферические эндокринные железы ((а также отдельные клетки), синтезирующие гормоны и высвобождающие их в кровь при действии соответствующего стимула. Третий р уровень - внутриклеточный. ур у р Его составляют изменения метаболизма в пределах клетки или отдельного метаболического пути, происходящие в результате: •изменения активности ф ферментов р путём у активации ц или ингибирования; •изменения количества ферментов по механизму индукции или репрессии синтеза белков или изменения скорости их разрушения; •изменения скорости транспорта веществ через мембраны клеток. Координация взаимодействия нервной и эндокринной систем Внешние и внутренние сигналы ЦНС - петли обратной связи действуют в системах регуляции гормонов надпочечников, щитовидной железы, половых желёз - + Гипоталамус + Статины Либерины Гипофиз + Тропные гормоны + Эндокринные железы + Гормоны ор о 1. Изменение активности ферментов 2. Изменение количества белков и ферментов 3. Изменение скорости транспорта веществ через мембраны клеток + Клетки-мишени Изменение концентрации метаболитов Гормоны – это интегрирующие молекулы, которые связывают различные регуляторные механизмы и метаболизм в разных органах. Они функционируют как химические посредники и переносят сигналы, которые возникают в ЦНС, различных органах и тканях, к соответствующим у щ клеткам-мишеням. Ответная реакция клетки определяется химическим строением гормона и типом клетки. В норме концентрация гормонов очень низкая. Синтез и секреция гормонов стимулируются внешними и внутренними сигналами, поступающими в ЦНС. Эти сигналы по нейронам поступают в гипоталамус, где стимулируют синтез пептидных рилизинг-гормонов (либеринов ( б и статинов), ) которые, соответственно, стимулируют или ингибируют синтез и секрецию гормонов передней доли гипофиза. Гормоны р передней р д д доли гипофиза ф стимулируют у ру образование р и секрецию р ц гормонов р периферических эндокринных желёз, которые поступают в общий кровоток и взаимодействуют с клетками-мишенями. Поддержание уровня гормонов в организме обеспечивает механизм отрицательной й обратной б й связи. Изменение концентрации метаболитов в клетках-мишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные д р железы,, либо на гипоталамус. у Синтез и секреция тропных гормонов подавляется гормонами эндокринных периферических желёз. Классификация гормонов по химическому строению Пептидные гормоны Адренокортикотропный гормон (кортикотропин; АКТГ) Гормон роста (соматотропин; СТГ) Тиротропный гормон (тиротропин; ТТГ) Фолликулостимулирующий гормон (ФСГ) Меланоцитстимулирующий гормон р ((МСГ)) Антидиуретический гормон (вазопрессин; АДГ) Паратиреоидный гормон (паратгормон; ПТГ) Кальцитонин Инсулин Глюкагон Стероиды Альдостерон Кортизол Кальцитриол Тестостерон Эстрадиол Прогестерон Производные аминокислот Адреналин Норадреналин Трийодтиронин (Т3) Тироксин (Т4) Классификация гормонов по биологическим функциям Регулируемые процессы Гормоны Обмен углеводов, углеводов липидов, липидов аминокислот Инсулин, глюкагон, адреналин, кортизол, тироксин, соматотропин Водно-солевой обмен Альдостерон, антидиуретический гормон Обмен кальция и фосфатов Паратгормон, кальцитонин, кальцитриол Репродуктивная функция Эстрадиол, тестостерон, прогестерон, гонадотропные гормоны Синтез и секреция гормонов эндокринных желез Тропные гормоны гипофиза гипофиза, либерины и статины гипоталамуса Изменение метаболизма в клетках, синтезирующих гормоны Эйкозаноиды, гистамин, секретин, гастрин, соматостатин, цитокины Гормоны распознаются и связываются особыми белками клетки рецепторами обладающими высокой специфичностью. рецепторами, специфичностью Физиологический эффект фф гормона р определяется р разными ф р факторами: р •концентрацией гормона (которая определяется скоростью инактивации в результате распада гормонов, протекающего в основном в печени, и скоростью выведения гормонов и его метаболитов из организма) •его сродством к белкам-переносчикам белкам переносчикам (стероидные и тиреоидные гормоны транспортируются по кровеносному руслу в комплексе с белками) •количеством и типом р рецепторов р на поверхности р клеток-мишеней. Передача гормонального сигнала в клетку зависит от химической природы гормона и происходит при участии соответствующих рецепторов. рецепторов Рецепторы пептидных гормонов и адреналина располагаются на поверхности клеточной мембраны (мембранные рецепторы). Рецепторы стероидных и тиреоидных гормонов находятся внутри клетки (внутриклеточные рецепторы). Причем для одних гормонов, например глюкокортикоидов, они локализованы в цитозоле, для других, таких как андрогены, эстрогены, тиреоидные гормоны, расположены в ядре клетки. клетки Типы рецепторов, локализованных в мембране (пептидные гормоны и адреналин) 1 – рецепторы, сопряженные с ионными каналами рецепторы ц р с каталитической активностью ((после связывания с гормоном) р ) 2–р 3 – рецепторы, сопряженные с G-белками и аденилатциклазой 4 - рецепторы, сопряженные с G-белками и фосфолипазой С Гормоны, связываясь с мембранными рецепторами, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника (гормона) в изменение концентрации вторичных посредников - цАМФ, цАМФ цГМФ, цГМФ ИФ3, ДАТ, ДАТ Са22+, NO. NO 9 Гормоны Гормоны, взаимодействие которых с рецептором клетки-мишени клетки мишени приводит к образованию цАМФ, действуют через трёхкомпонентную систему, которая включает белок-рецептор, Gб G-белок иф фермент аденилатциклазу. Образующийся Об й под действием аденилатциклазы цАМФ активирует протеинкиназу А, фосфорилирующую ферменты и другие белки. В случае рецепторов, сопряжённых с Gs-белком, субъединица αs-ГТФ активирует аденилатциклазу. В случае рецепторов, сопряжённых с Giбелком, субъединица αiГТФ ингибирует б аденилатциклазу 9 Другая система, генерирующая цГМФ как вторичный посредник, сопряжена р с гуанилатциклазой. у ц Цитоплазматический д Ц домен такого типа рецепторов обладает активностью гуанилатциклазы, которая катализирует реакцию образования цГМФ из ГТФ (подобно аденилатциклазе). Молекулы цГМФ могут активировать ионные каналы либо активировать цГМФ-зависимую протеинкиназу G, участвующую в фосфорилировании других белков в клетке. клетке 9 Некоторые гормоны (например, (например вазопрессин или адреналин) адреналин), образуя комплекс с соответствующими рецепторами (рецептор V1 для вазопрессина и αi-рецептор для адреналина), через а активацию а ю соответствующих соо е с ю G-белков G бе о а активируют р ю фосфолипазу фосфо аз С, С в результате чего в клетке появляются вторичные посредники ИФ3, ДАГ. Молекула ИФ3 стимулирует высвобождение Са2+ из ЭР. Кальций связывается с белком кальмодулином. Этот комплекс активирует Са2+-кальмодулинзависимую протеинкиназу. Ионы кальция и ДАГ участвуют в активации протеинкиназы С . Структура рецепторов стероидных гормонов и гормонов щитовидной железы Рецепторы стероидных гормонов - полипептиды с мол. мол м. м 50-80 50 80 тыс. тыс Находятся в цитоплазме или в ядре клеток. N-конец С-конец На С-концевом участке находится домен узнавания и связывания гормона. гормона Центральная часть рецептора включает домен связывания ДНК. На N-концевом участке располагается домен, называемый вариабельной областью рецептора, отвечающий за связывание с другими белками, участвующими в регуляции транскрипции. Передача сигнала стероидных гормонов через внутриклеточные у р рецепторы Плазматическая мембрана Пептидные гормоны и адреналин - через мембранные рецепторы Стероидные гормоны и тироксин - через внутриклеточные у рецепторы Гормон Гормон Рецептор на мембране Рецептор внутри клетки G-белок Фермент р (аденилатциклаза, гуанилатциклаза фосфолипаза С) 2-й посредник (цАМФ, цГМФ, Са+2' ИФз,, Д ДАГ,, NO)) Аутофосфорилирование рецептора Каскад фосфорилирования белков Активация ферментов А ф и факторов транскрипции Комплекс гормон-рецептор р р ц р Транспорт комплекса гормон — рецептор в ядро Взаимодействие с ДНК (энхансер, сайленсер) Изменение количества белков (ферментов) Индукция или репрессия синтеза белков Протеинкиназы Фосфорилирование белков Изменение фу функциональной активности ферментов и факторов транскрипции Изменение количества белков (ферментов) Изменение скорости метаболизма Строение и биологические функции гормонов гипоталамуса Г Гормон С Строение Биологическая Б функция Тиреотропин-рилизинггормон (тнреолиберин, ( б ТРФ) Пептид, 3 а.к. Стимулирует секрецию тиреотропина и пролактина. Кортикотропин-рилизинггормон р ((кортиколиберин, р р , КРФ). Гонадотропин-рилизинггормон (гонадолибернн, ГРФ) Полипептид, 41 а.к. Стимулирует секрецию кортикотропина р р Полипептид, 10 а.к. Стимулирует секрецию ЛГ и ФСГ Соматотропин-рилизинггормон (соматолиберин, СРФ)) Соматостатин (соматотропин ингибирующий гормон) Полипептид, 40 или 44 а.к. Стимулирует секрецию соматотропина. Полипептид, 14 или 28 а.к. Ингибирует секрецию соматотропина. Пролактолиберин Еще неизвестно Стимулирует секрецию пролактина Пролактостатин Полипептид, 5б а.к. Ингибирует секрецию пролактина Строение и биологические функции гормонов передней доли гипофиза Гормон Строение Биологическая функция Гормон роста (ГР), соматотропный гормон (СТГ) Тиреотропин, тиреотропный гормон (ТТГ) Пептид,191а.к. Димер (αβ) α-полипептид, 96 а.к., β β-полипептид, 112 а.к. Пролактин (ПРЛ) Лютеинизирующий ру щ гормон р (ЛГ) Фолликулостимулирующий Ф й гормон (ФСГ) Кортикотропин, адренокортикотропный гормон (АКТГ) Стимулирует постнатальный рост скелета и мягких тканей, участвует в регуляции энергетического обмена Стимулирует синтез йодтиронинов Полипептид, 197 а.к. Стимулирует лактацию Димер (αβ) α-полипептид, 96 а.к., β-полипептид, 112 а.к. У женщин стимулирует овуляцию, у мужчин индуцирует синтез андрогенов в клетках Лейдига Димер (αβ) α-полипептид, 96 а.к., β-полипептид, β д, 112 а.к. У женщин стимулирует рост фолликулов, у мужчин стимулирует сперматогенез Полипептид, 39 а.к. β-Липотропин (β-ЛТГ) Полипептид, 93а.к. Стимулирует рост надпочечников и синтез кортикостероидов Стимулирует липолиз Действие гормона роста через ИФР Физиологические мишенях: эффекты ГР в тканях- Печень – стимуляция глюконеогенеза и П синтеза белка. Кости – стимуляция роста и синтеза белка. Жировая ткань – стимуляция липолиза и снижение утилизации глюкозы. Мышцы - снижение утилизации глюкозы, стимуляция синтеза белка. белка 1 - ГР индуцирует синтез ИФР. 2 - ИФР взаимодействуют со специфическими рецепторами клеток той же или других тканей. 3 - ИФР стимулируют фосфорилирофосфорилиро вание и активацию белков, участвующих в митозе и росте. Нарушения функций гипоталамо гипофизарной системы гипоталамо-гипофизарной Гипофункция – уменьшение или подавление продукции тропных гормонов – ведет к резкому снижению функции периферических эндокринных желез. Гипофункция гормона роста 9Гипофизарный нанизм, или карликовость – причина – дефицит ГР, часто из за мутаций в гене ГР. из-за ГР 9Нанизм Ларона – дефект рецепторов ГР гепатоцитов и снижение синтеза ИФР-1 и ИФР-2. Концентрация ГР повышена. 9Карликовость африканских пигмеев – нарушена пострецепторная передача сигнала ГР. Концентрация ГР в норме, а ИФР-1 - снижена Гиперфункция гормона роста обычно возникает в результате образования гормонпродуцирующей опухоли соматотрофных клеток гипофиза 9Гигантизм - возникает у детей и подростков с незакончившимся процессом окостенения эпифизарных хрящей, но продолжающимся ростом длинных костей 9Акромегалия - гиперсекреция гормона роста у взрослых людей при которой рост тела ускоряется, но не в длину, а в ширину с диспропорциональным увеличением размеров лица, кистей рук, стоп, черепа, увеличением размеров внутренних органов Структура и синтез гормонов щитовидной щ д железы Йодтиронины синтезируются в составе белка тиреоглобулина (Тг) в фолликулах щитовидной железы. Тиреоглобулин синтезируется на рибосомах, далее поступает в аппарат Гольджи, а затем во внеклеточный коллоид, где он хранится. Там же происходит йодирование остатков тирозина. Образование йодтиронинов осуществляется в несколько этапов: транспорт йода в клетки щитовидной железы; окисление йода; йодирование остатков тирозина; образование йодтиронинов; транспорт йодтиронинов в кровь. I- Регуляция синтеза гормонов щитовидной железы Ц р у ц в крови Циркуляция р в связанной ф форме р в комплексе с белками: с тироксинсвязывающим глобулином (ТСГ) и тироксинсвязывающим преальбумином (ТСПА). Активна свободная форма. Скорость синтеза и секреции йодтиронинов регулируются гипоталамо гипофизарной системой гипоталамо-гипофизарной по механизму обратной связи. Стимулом для повышения секреции тиреолиберина и тиреотропина служит снижение концентрации йодтиронинов в крови. крови Основные эффекты йодтиронинов - результат их взаимодействия с внутриклеточными рецепторами, которые в комплексе с гормонами постоянно находятся в ядре и взаимодействуют с определёнными последовательностями д ДНК,, у Д участвуя у вр регуляции у ц экспрессии р генов. Йодтиронины 9повышают поглощение клетками кислорода во всех органах, кроме мозга, РЭС и гонад. 9в разных клетках стимулирует работу Nа+,К+-АТФ-азы, на что затрачивается значительная часть энергии, утилизируемой клеткой. 9участвуют в формировании ответной реакции на охлаждение увеличением теплопродукции, повышая чувствительность симпатической нервной системы к норадреналину и стимулируя секрецию норадреналина Очень высокие концентрации Т3 тормозят синтез белков стимулируют стимулируют катаболические процессы (отрицательный азотистый баланс) При физиологической концентрации проявляется в стимуляции 9белкового синтеза 9процессов роста 9клеточной дифференцировки 9транскрипции гена гормона роста йодтиронинов их действие В печени уускоряют р 9Гликолиз 9синтез холестерола 9синтез жёлчных кислот В печени и жировой ткани повышают чувствительность клеток к действию адреналина и косвенно 9стимулирует липолиз в жировой ткани 9мобилизацию гликогена в печени В мышцах увеличивают 9потребление глюкозы 9стимулирует синтез белков и увеличение мышечной массы 9 повышает чувствительность мышечных клеток к действию адреналина Заболевания щитовидной железы 1. Гипотиреоз 1 Г - развивается вследствие недостаточности йодтиронинов. й 9У новорожденных приводит к развитию кретинизма (множественные врождённые нарушения и необратимая задержка умственного развития). 9У взрослых связан с недостаточностью функции ЩЖ, ЩЖ может возникать и при заболеваниях гипофиза и гипоталамуса. Наиболее тяжёлые формы гипотиреоза, сопровождающиеся слизистым отёком кожи и подкожной клетчатки обозначают термином "микседема" клетчатки, микседема . 9Хронический аутоиммунный тиреоидит, приводящий к снижению синтеза йодтиронинов, - зоб Хашимото. 9Эндемический зоб (нетоксический зоб) - результат недостаточного поступления йода в организм при его низком содержании в воде и почве и поступлении в организм менее 100 мкг/сут. При р э этом ууменьшается продукция р ду ц йодтиронинов, д р , что приводит р д к уусилению секреции ТТГ, под влиянием которого происходит компенсаторное увеличение размеров ЩЖ (гиперплазия), но продукция йодтиронинов при этом не у увеличивается. Характерные проявления: снижение частоты сердечных сокращений, вялость, сонливость, непереносимость холода, сухость кожи. Эти симптомы развиваются вследствие снижения основного обмена, скорости гликолиза, мобилизации гликогена и жиров, потребления глюкозы мышцами, уменьшения мышечной массы и снижения теплопродукции. Заболевания щитовидной железы 2. Гипертиреоз - развивается вследствие повышенной продукции йодтиронинов. Диффузный токсический зоб (базедова болезнь, болезнь болезнь Грейвса) – наиболее распространённое заболевание ЩЖ. У больных увеличены размеры ЩЖ (зоб), повышена концентрация йодтиронинов в 2 - 5 раз и развивается тиреотоксикоз. тиреотоксикоз Причины: опухоли ЩЖ, тиреоидит, избыточное поступление йода и йодсодержащих препаратов, аутоиммунные реакции. Болезнь Грейвса возникает в результате образования антител, имитирующих ру щ д действие тиреотропина, р р что приводит р д кд диффузному ффу у разрастанию ЩЖ и неконтролируемой продукции Т3 и Т4. Уровень ТТГ при этом заболевании снижен вследствие подавления функции гипофиза высокими концентрациями йодтиронинов. Характерные признаки тиреотоксикоза: увеличение основного обмена, учащение сердцебиений, мышечная слабость, снижение массы тела, повышенный аппетит, потливость, повышение температуры тела, тремор и экзофтальм (пучеглазие). отрицательный азотистый баланс. Гормоны коры надпочечников (кортикостероиды) Биологически активные кортикостероиды объединяют в 3 основные класса в зависимости от их преобладающего действия Глюкокортикоиды, С21-стероиды, играют важную роль в адаптации к стрессу. Оказывают разнообразные эффекты, наиболее важный стимуляция у ц глюконеогенеза Основной глюкокортикоид человека - кортизол. Минералокортикоиды, С21-стероиды, необходимы для поддержания уровня Na+ и К+. Самый активный гормон этого класса – альдостерон. Андрогены - С19-стероиды. В коре надпочечников образуются предшественники андрогенов, из которых наиболее активный дегидроэпиандростерон (ДЭА) и слабый - андростендион. Самый мощный андроген надпочечников тестостерон синтезируется в надпочечниках в небольшом б количестве. Эти Э стероиды превращаются в более б активные андрогены вне надпочечников. Тестостерон в незначительных количествах может превращаться в надпочечниках в эстрадиол. Но в норме продукция этих гормонов надпочечниками не играет существенной роли. Общим предшественником кортикостероидов служит холестерол. который в основном поступает из крови в составе ЛПНП или синтезируется в клетках из ацетил-КоА. В митохондриях холестерол превращается в прегненолон С21-стероида при участии гидроксилазы, гидроксилазы относящейся к группе цитохромов Р450. Дальнейшее превращение прегненолона происходит под действием различных гидроксилаз с участием молекулярного кислорода и NADPH, NADPH а также дегидрогеназ, изомераз и лиаз. Эти ферменты имеют различную внутри- и межклеточную локализацию. В коре надпочечников различают 3 типа клеток, образующих 3 слоя, или зоны: клубочковую, пучковую и сетчатую. Ферменты, необходимые для синтеза 9альдостерона, присутствуют только в клетках клубочковой зоны. 9глюкокортикоидов и андрогенов локализованы локализованы в пучковой и сетчатой зонах. Синтез основных кортикостероидов 1. Превращение холестерола в прегненолон 2. Образование прогестерона 3 Гидроксилировагние прогестерона 3. (17-21-11) и образование кортизола 4. Гидроксилирование прогестерона (21-11) и образование альдостерона 5. Путь синтеза андрогенов и эстрогенов О О О О Путь биосинтеза кортизола: Под влиянием АКТГ происходит активация специфической эстеразы, эстеразы и свободный холестерол транспортируется в митохондрии. • В митохондриях от холестерола отщепляется боковая цепь с образованием в прегненолона. • В цитозоле клеток пучковой зоны коры надпочечников прегненолон превращается в прогестерон. • В мембранах ЭР происходит гидроксилирование прогестерона по С17 с образованием 17 17-гидроксипрогестерона гидроксипрогестерона. Далее 17 17-ОН-прогестерон ОН прогестерон превращается в 11 11дезоксикортизол • Во внутренней мембране митохондрий гидроксилирование приводит к образованию кортизола. Стимулы синтеза и секреции кортизола: 9 травма 9 инфекция 9 понижение концентрации глюкозы в крови. 9 стресс Синтез кортизола регулируется по механизму отрицательной обратной связи. Транспорт кортикостероидов Кортизол в плазме крови находится в комплексе с α-глобулином транскортином и в небольшом количестве в свободной форме. Синтез транскортина протекает в печени и стимулируется эстрогенами. Биологически активным явлется свободный кортизол. Внутриклеточная локализация синтеза кортизола Ткани-мишени и биологические функции кортизола В печени (анаболические функции): 9стимулирует у ру глюконеогенез,, 9индуцирует синтез ферментов катаболизма аминокислот р ф р тканях В периферических (катаболические функции): 9стимулирует распад белков и высвобождение аминокислот; 9тормозит потребление глюкозы; 9стимулирует липолиз в конечностях и липогенез в др. органах. В высоких дозах - иммунодепрессант, обладает противовоспалительной активностью. При длительном применении Ингибирует активность остеобластов, Повышает резорбцию кости и выведение Са2+, что приводит к развитию остеопороза. Выводится в виде 17-окси и 17-кетостероидов в виде конъюгатов с глюкуроновой и серной кислотой ГИПЕРКОРТИЦИЗМ Клинические проявления: - гипергликемия, снижение толерантности к глюкозе (стероидный диабет); усиление катаболизма белков; снижение массы тела; истончение кожи; остеопороз; инволюция лимфоидной ткани; перераспределение отложения жира (лунообразное лицо, выступающий живот); гипернатриеимия, гипертензия, гипокалиемия. Причины: 9Нарушение регуляторных механизмов синтеза кортизола при опухолях гипофиза (болезнь Иценко-Кушинга) и опухолях других клеток, вырабатывающих кортикотропиноподобные вещества вещества. 9Избыточный синтез кортизола при гормонально-активных опухолях коры надпочечников (синдром Иценко-Кушинга). Диагностика: 1.Определение концентрации АКТГ в плазме крови. 2. Проба с дексаметазоном - при болезни Иценко-Кушинга концентрация кортизола снижается на 50%. Отсутствие реакции свидетельствует об опухоли у надпочечников или внегипофизарной ф р опухоли, у , секретирующей р ру АКТГ. ПРИОБРЕТЕННАЯ НЕДОСТАТОЧНОСТЬ НАДПОЧЕЧНИКОВ (БОЛЕЗНЬ АДДИСОНА) Клинические проявления: снижение массы тела, тела общая слабость, слабость снижение аппетита, тошнота, рвота, снижение АД, гипонатриемия, гиперпигментация кожи ("бронзовая болезнь"). Причина гиперпигментации - повышение продукции р у ПОМК. Причины: поражение надпочечников туберкулезным или аутоиммунным процессом; длительное применение кортикостероидных препаратов ВРОЖДЕННАЯ АДРЕНОГЕНИТАЛЬНАЯ ДИСТРОФИЯ Клинические проявления: усиление роста тела, раннее половое созревание у мальчиков и развитие мужских половых признаков у девочек. Причины: дефицит C21-гидроксилазы (обнаруживается в 95% случаев), реже C11-гидроксилазы. C11 С Снижение продукции кортизола сопровождается увеличением секреции АКТГ и накоплением промежуточных продуктов синтеза кортизола, повышением секреции 17-ОН-прогестерона, в том числе предшественников р д андрогенов. др ОСТРАЯ НЕДОСТАТОЧНОСТЬ НАДПОЧЕЧНИКОВ Клинические проявления: - декомпенсация всех видов обмена и процессов адаптации - сосудистый коллапс, коллапс резкая адинамия, адинамия потеря сознания. сознания Может быть следствием декомпенсации при длительных хронических заболеваниях, может возникать при резкой отмене гормональных препаратов после длительного лечения глюкокортикоидамми. Причины: нарушение обмена электролитов (потеря Na+, Сl-, обезвоживание, увеличение концентрации К+ в сыворотке крови, снижение уровня сахара в крови и запаса гликогена Гормоны мозгового слоя надпочечников 9 дофамин 9 адреналин 9 норадреналин (катехоламины) Непосредственным предшественником гормонов ор о о сслужит у тирозин. роз Синтез катехоламинов происходит в цитоплазме и гранулах клеток мозгового слоя надпочечников. В гранулах происходит также запасание катехоламинов. Механизм действия катехоламинов Катехоламины действуют на клетки-мишени через мембранные рецепторы 9 α-адренергические 9β-адренергические Норадреналин в физиологических концентрациях главным образом взаимодействует с α-рецепторами. Адреналин взаимодействует с α- и β-рецепторами адреналин + ββ рецептор активация аденилатциклазы ингибирование аденилатциклазы адреналин + α2-рецептор адреналин + α1-рецептор активация фосфолипазы С, стимуляция инозитолфосфатного пути передачи сигнала Клетки-мишени и биологические эффекты адреналина печень 9стимуляция распада гликогена 9ингибирование синтеза гликогена 9активация глюконеогенеза скелетные мышцы 9стимуляция распада гликогена 9ингибирование синтеза гликогена жировая ткань 9активация липолиза Структура инсулина человека Инсулин - гормон поджелудочной железы - полипептид, состоящий из двух полипептидных цепей. Цепь А содержит 21 аминокислотный остаток, цепь В - 30 аминокислотных остатков. Обе цепи соединены между собой двумя дисульфидными мостиками Молекула инсулина содержит также внутримолекулярный й дисульфидный ф й мостик, соединяющий шестой и одиннадцатый остатки в А-цепи. А-цепь В-цепь Участок связывания с рецептором Процессинг и секреция инсулина Биосинтез препроинсулина начинается с образования сигнального пептида на полирибосомах, связанных с ЭР. После окончания синтеза препроинсулина сигнальный пептид, включающий 24 аминокислотных остатка, отщепляется. Проинсулин (86 аминокислотных остатков) поступает в аппарат Гольджи, где под действием специфических протеаз расщепляется с образованием инсулина (51 а.к. остаток) и С-пептида (31 а.к остаток) Инсулин и С-пептид в эквимолярных количествах включаются в секреторные гранулы и секретируются во внеклеточную жидкость в результате экзоцитоза. Глюкоза - главный регулятор секреции инсулина Действие инсулина начинается с его связывания со специфическим гликопротеиновым рецептором на поверхности клетки-мишени. Инсулин - главный анаболический гормон. Эффекты инсулина могут проявляться в течение секунд и минут (транспорт веществ фосфорилирование и веществ, дефосфорилирование белков, активация и ингибирование ферментов, синтез РНК) или через несколько часов (синтез ДНК, белков, рост клеток Механизм действия инсулина Активация Ras-пути Активация фосфодиэстеразы адипоцитов Активация гликогенсинтазы Инсулин Инсулин Инсулин IRS-1-P, Shc-P IRS-1-P, Shc-P IRS-1-P, Shc-P Комплекс IRS-1-P, Shc-P с белком GRB 2/m SOS Комплекс IRS-1-P, Shc-P с ФИ-3-киназой Комплекс IRS-1-P, Shc-P с белком GRB 2/m SOS Активация Ras Активация ПК-В Активация Ras Активация киназы Raf-1 Активация ФДЭ Активация киназы Raf-1 Активация киназы рр90S6 Активация МАПКК Снижение концентрации цАМР Активация МАПК Пролиферация Метаболизм Активация протеинфосфатазы 1. Инактивация киназы фосфорилазы и гликогенфосфорилазы 2. Активация гликогенсинтазы 1.Торможение мобилизации гликогена 2.Стимуляция синтеза гликогена Влияние инсулина на ключевые ферменты метаболизма Печень 1. 2. 3 3. 4. Фосфодиэстераза Фосфофруктокиназа Пируваткиназа Пируватдегидрогеназный комплекс 5. Фосфатаза гликогенсинтазы и гликогенфосфорилазы 6 Ацетил-КоА6. карбоксилаза 1. 2. 3 3. 4. 5. Мышцы Активация Фосфодиэстераза Фосфофруктокиназа Пируваткиназа Пируватдегидрогеназный комплекс Фосфатаза гликогенсинтазы Жировая ткань 1. 2. 3 3. 4. ЛП-липаза Фосфофруктокиназа Пируваткиназа Ацетил-КоАкарбоксилаза Индукция 1. 2. 3. 4. 5 5. Глюкокиназа Цитратлиаза Пальмитатсинтаза Пируваткиназа Ацетил-КоАкарбоксилаза 6. Глюкозо-6-фосфатдегидрогеназа 1. Глицеральдегидфосфатдегидрогеназа 2. Пальмитатсинтаза Репрессия 1. Фосфоенолпируваткарбоксикиназа Глюкагон Глюкагон – 29 а.к., синтезируется в α-клетках поджелудочной железы и в L-клетках кишечника, в крови – гормон поджелудочной железы, Т1/2 – 5 мин, Механизм действия обусловлен у повышением концентрации цАМФ. Органы мишени и биологические эффекты: Печень 1. Активация гликогенфосфорилазы, снижение активности гликогенсинтазы мобилизация ц гликогена 2. Фосфорилирование пируваткиназы и БИФ торможение гликолиза и стимуляция глюконеогенеза 3. Индукция синтеза глюкозо-6-фосфатазы, фосфоенолпируваткарбоксикиназы, фруктозо-1,6-бисфосфатазы стимуляция глюконеогенеза Жировая ткань Активация гормончувствительной ТАГ-липазы стимуляция липолиза РЕГУЛЯЦИЯ ОБМЕНА ОСНОВНЫХ ЭНЕРГОНОСИТЕЛЕЙ ИЗМЕНЕНИЯ ГОРМОНАЛЬНОГО СТАТУСА И МЕТАБОЛИЗМА ПРИ САХАРНОМ ДИАБЕТЕ Гормональная регуляция энергетического обмена Гормоны регулируют координацию скоростей 9 гликолиза, гликолиза 9 глюконеогенеза, 9 синтеза и распада гликогена 9 синтеза и распада ТАГ 9 распада белков и аминокислот Гормональная регуляция обеспечивает 9предотвращение чрезмерного повышения концентраци глюкозы после приема пищи; 9запасание гликогена и его использование между приемами пищи; 9снабжение мышц глюкозой при мышечной работе; 9снабжение глюкозой клеток мозга, эритроцитов и некоторых других тканей при голодании Энергетический гомеостаз обеспечивается 9 инсулином у и контринсулярными гормонами: 9глюкагоном 9кортизолом 9адреналином 9соматотропином 9гормонами щитовидной железы Инсулин и глюкагон играют главную роль в регуляции метаболизма при смене абсорбтивного и постабсорбтивного периодов и при голодании. Режим запасания включается после приёма Р ё пищи и сменяется режимом мобилизации запасов после завершения пищеварения. Главные органы, связанные с изменениями потока метаболитов при смене режимов мобилизации и запасания энергоносителей, - печень, жировая ткань и мышцы. изменения концентрации глюкозы, инсулина и глюкагона в крови в абсорбтивный период Абсорбтивный период характеризуется временным повышением концентрации глюкозы, аминокислот и жиров в плазме крови. Клетки поджелудочной железы отвечают на это усилением секреции инсулина и снижением секреции глюкагона. Увеличение отношения инсулин/глюкагон вызывает ускорение использования метаболитов для запасания энергоносителей: происходит синтез гликогена, жиров и белков. Концентрация глюкозы в крови в норме 60 - 100 мг/дл (3,5 - 5,5 моль/л) Алиментарная гипергликемия - до 150 мг/дл (8 ммоль/л) через 1 час после приема углеводной пищи, возврат к норме - через 2 часа. Энергетическая ценность основных энергоносителей Источник энергии Энергетическая ценность ккал/г Углеводы 4 Жиры 9 Белки 4 Суточная потребность взрослого человека – Д ) 2000 – 3000 ккал ((8000-12000 кДж) Источники энергоносителей Абсорбтивный период Постабсорбтивный период и голодание Пищеварительный тракт Глюкоза Жирные кислоты Аминокислоты Депонированные энергоносители Гликоген Жиры Белки Абсорбтивный период Абсорбтивный период Печень 9после приёма пищи главный потребитель глюкозы, почти 60 из 100 г глюкозы, транспортируемой портальной системой, задерживается в печени - следствие ускорения синтеза гликогена и жиров Жировая ткань 9ускорение транспорта глюкозы в адипоциты 9активация ключевых ферментов гликолиза 9стимуляция пентозофосфатного пути 9торможение липолиза 9запасание э энергоносителей р в форме ф р триацилглицеролов р ц ц р Мышцы 9ускорение транспорта глюкозы в клетки мышечной ткани 9окисление глюкозы для обеспечения клетки энергией и синтеза гликогена. Абсорбтивный период Кровь и органы Плазма П крови Особенности метаболизма Г Глюкоза ↑ аминокислоты ↑, ↑, ↑ жиры ↑ Инсулин ↑, глюкагон ↓ (инсулин-глюкагоновый индекс ↑) Печень Потребление глюкозы ↑, синтез гликогена ↑, гликолиз ↑, ↑ синтез жирных кислот↑, ↑ синтез белка ↑ , синтез жиров ↑ Жировая р ткань (адипоциты) Мышцы Транспорт глюкозы и жирных кислот в адипоциты ↑, активность ЛП-липазы капилляров↑, гликолиз ↑, пентозофосфатный путь ↑, синтез ТАГ ↑, активность ТАГ-липазы↓, липолиз ↓ Транспорт глюкозы и аминокислот ↑, гликолиз ↑ синтез гликогена ↑, синтез белка ↑ Гормоны, регулирующие обмен углеводов, белков и жиров Гормон, место с-за Инсулин, β-клетки поджелудоч д уд -ной железы Сигнал к секре секреции С глк Ор а Органы-мишени е Печень 1. Ускорение синтеза гликогена 2. Ускорение синтеза белка 3. Торможение р глюконеогенеза Мышцы 1. Ускорение синтеза гликогена 2. Ускорение синтеза белка 3. Ускорение транспорта глюкозы в клетку Жировая ткань 1. Ускорение синтеза жиров из глюкозы 2. Ускорение транспорта глюкозы в клетку Печень 1. Ускорение распада гликогена 2 Ускорение глюконеогенеза 2. Жировая ткань 1. Ускорение липолиза Печень 1. Ускорение распада гликогена Мышцы 1. Ускорение распада гликогена Жировая ткань 1. Ускорение липолиза Печень 1. Ускорение глюконеогенеза 2. Индукция ферментов глюконеогенеза и катаболизма б а/к / Мышцы 1. Ускорение распада аминокислот 2. Снижение скорости поступления а/к в крови Глюкагон, α-клетки поджелудоч -ной железы Адреналин, мозговое вещество надпочечни -ков Кортизол, кора надпочечни -ков С глк Изменение метаболизма в клеткахмишенях е в крови Сигнал ЦНС С глк в крови Источники и пути расходования глюкозы крови Мышцы Кишечник Окисление Стимулирует - И Синтез гликогена Стимулирует - И Переваривание углеводов Печень Глюконеогенез Стимулирует - К, Г Мобилизация гликоген Стимулирует - А, Г Синтез гликогена Стимулирует - И Синтез жиров Стимулирует - И Окисление Глюкоза Мобилизация гликогена Стимулирует - А Жировая ткань Окисление Стимулирует - И С Синтез жиров Стимулирует - И Мозг Окисление Постабсорбтивный период период после завершения пищеварения до следующего приёма пищи Изменения метаболизма направлены на поддержание концентрации в крови глюкозы, которая служит основным энергетическим субстратом для мозга и единственным источником энергии для эритроцитов. Основные изменения метаболизма в этот период происходят в печени и жировой ткани. Печень 9ускоряется мобилизация гликогена 9главным источником глюкозы становится глюконеогенез (субстратами служат глицерол, аминокислоты и лактат) 9снижается скорость синтеза жирных кислот 9возрастает скорость р-окисления 9увеличивается снабжение печени жирными кислотами из жировых депо 9ацетил-КоА используется в печени для синтеза кетоновых тел Жировая ткань 9снижается скорость синтеза ТАГ 9стимулируется липолиз 9жирные кислоты становятся важными источниками энергии в печени, мышцах и жировой ткани 9концентрация глюкозы в крови поддерживается на уровне 80-100 мг/дл, а уровень жирных кислот и кетоновых тел возрастает Постабсорбтивный период Постабсорбтивный период Кровь и органы Особенности метаболизма Плазма крови Глюкоза ↓ Инсулин ↓, глюкагон ↑ (инсулин-глюкагоновый индекс ↓) Печень Потребление глюкозы ↓, гликолиз ↓, синтез гликогена ↓, распад гликогена ↑, глюконеогенез ↑, синтез жирных кислот ↓, поступление жирных кислот из депо↑ , ββ окисление жирных кислот ↑ синтез кетоновых тел↑ Жировая Т Транспорт глюкозы и жирных кислот в адипоциты↓, ↓ ткань синтез ТАГ ↓, липолиз ↑ (адипоциты) Мышцы Транспорт глюкозы ↓ Голодание Голодание может быть неполным (недоедание) и полным. Главные патологические проявления неполного голодания связаны с белковой недостаточностью В изменениях обмена веществ при полном голодании можно выделить три фазы фазы. Фазы голодания Длительность голодания I фаза В течение суток II фаза В течение недели фаза III ф Несколько недель д Глюкоза ↓ инсулин ↓ глюкагон ↑ (инсулин-глюкагоновый индекс ↓) Контринсулярные гормоны ↑ Контринсулярные гормоны 9 поддерживают уровень глюкозы в крови на уровне нормы для обеспечения глюкозозависимых тканей 9 мобилизуют другие источники энергии для обеспечения энергией остальных тканей Голодание Изменение гормонального статуса и метаболизма при голодании Фаза Дни Изменение гормонов I 1 Инсулин Г Глюкагон II 7 Инсулин Глюкагон Кортизол III 30 40 Инсулин Глюкагон Кортизол Изменения в обмене веществ • Мобилизация гликогена (печень) • Распад жира (жировая ткань) • Распад жира (жировая ткань) • Окисление жирных кислот • Синтез кетоновых тел (печень) • Глюконеогенез из аминокислот и глицерина - 150 г/сутки единственный источник глюкозы. • Мышцы и адипоциты не используют глюкозу. • Печень прекращает использовать глюкозу в качестве источника энергии • Синтез кетоновых тел (печень) • Глюконеогенез из аминокислот и глицерина - 80 г/сутки • Все ткани ткани, кроме эритроцитов и мозгового слоя почек, используют альтернативные источники энергии. Кетоновые тела, мг/дл Глюкоза, мг/дл 2 60-70 30 Поддерживается на нижнем уровне нормы во все фазы голодания 30 ИЗМЕНЕНИЯ ГОРМОНАЛЬНОГО СТАТУСА И МЕТАБОЛИЗМА ПРИ САХАРНОМ ДИАБЕТЕ Сахарный диабет - заболевание, возникающее вследствие абсолютного или относительного дефицита инсулина. инсулина I. Инсулинзависимый сахарный диабет (ИЗСД) – диабет б I типа Заболевание вызвано абсолютным дефицитом инсулина Причина 9 Деструкция β-клеток в результате аутоиммунных реакций Провоцирующие факторы: 9 β-цитотропные вирусы ( вирусы оспы, краснухи, кори, цитомегаловирус и др.)) 9 токсические вещества (производные нитрозомочевины и другие нитро- или аминосодержащие соединения)) II.Инсулинонезависимый сахарный диабет (ИНСД) – диабет II типа З б Заболевание вызвано относительным дефицитом ф инсулина Причины: 9нарушение секреции инсулина; 9нарушение превращения проинсулина в инсулин; 9повышение скорости распада инсулина; 9дефекты дефекты рецептора инсулина; 9дефекты пострецепторных механизмов передачи сигнала инсулина. у Особенности: позднее проявление – после 40 лет; высокая частота семейных форм Факторы риска: ожирение, неправильный режим питания, малоподвижный образ жизни, стресс. 9 9 9 9 9 9 9 9 9 9 Симптомы сахарного диабета Гипергликемия (толерантность к глюкозе) Глюкозурия (почечный порог 160 мг/мл) Кетонемия (свыше 20 мг/дл) Кетонурия Ке оа Кетоацидоз оз Гиперлипопротеинемия Азотемия Азотурия Полиурия (3-4 л/сутки) Полидипсия, полифагия, сухость во рту Изменение толерантности к глюкозе у больного ИНСД. Тест с сахарной нагрузкой. 1 – здоровый человек 2 – больной ИНСД Причины гиперглюкоземии при диабете 9 Потребление глюкозы мышцами и жировой тканью снижено (из-за отсутствия инсулина ГЛЮТ-4 отсутствует на мембране миоцитов и адипоцитов, глюкоза в эти клетки не поступает и не используется ими для запасания в форме гликогена и жиров). 9 В печени глюкоза не используется для запасания в форме гликогена, т.к. из-за низкой концентрации инсулина и высокой глюкагона гликогенсинтаза находится в фосфорилированной ф ф й неактивной й форме. 9 В печени глюкоза не используется для синтеза жиров, т. к. активность гликолиза и пируватдегидрогеназы снижены и превращение глюкозы в ацетил-КоА замедлено. 9 В печени активирован глюконеогенез из аминокислот и глицерина из-за низкой концентрации инсулина и высокой глюкагона. Причины гиперлипопротеинемии, повышения свободных жирных кислот и кетоновых тел при диабете б Пищевые жиры не депонируются в жировой ткани, ткани т.к. т к цАМФзависимая ТАГ-липаза адипоцитов находится в фосфорилированной (активной) форме. Содержание свободных жирных кислот в крови повышено изза активности ТАГ-липазы. Жирные кислоты поглощаются печенью, часть их превращается в ТАГ, которые в составе ЛПОНП секретируются в кровь. (Предрасположенность к развитию атеросклероза и гипертонии). Часть жирных кислот подвергается β-окислению в митохондриях печени и образующийся ацетил-Ко используется для синтеза кетоновых тел. тел Острые осложнения сахарного диабета Нарушения обмена углеводов, жиров и белков при сахарном диабете могут приводить к развитию острых осложнений - коматозным состояниям Формы коматозных состояний Кетоацидотическая кома o Гипергликемия р ((2030 ммоль/л) o Кетоацидоз o (высокий уровень кетоновых тел o >100 ммоль/л) o Гиперосмолярность o Полидипсия o Полиурия o Потеря жидкости и электролитов o Дегидратация Гиперосмолярная кома o Гипергликемия р ((>40 ммоль/л) o Ацидоз отсутствует o Кетоновые тела в норме Гиперосмолярность Полидипсия Полиурия Потеря жидкости и электролитов o Дегидратация o o o o Лактоацидотическая кома o Гипотония o Снижение периферического кровообращения o Гипоксия тканей o Анаэробный гликолиз в тканях o Лактоацидоз Разные варианты р диабетической комы в чистом виде практически не встречаются. Их возникновение может быть обусловлено разными факторами: o o o o o o o o o o инфекции ф тиреотоксикоз синдром Кушинга инфаркт инсульт иравма хирургические вмешательства глюкокортикоиды Эстрогены стресс ДИАБЕТИЧЕСКАЯ КОМА – резкое нарушение всех функций ф й организма с нарушением сознания. Механизм развития комы при дефиците инсулина Диабетическая кома развивается медленно, в течение нескольких дней, но иногда может возникнуть и в течение нескольких часов часов. Первыми признаками могут быть тошнота, рвота, заторможенность. АД у больных снижено. Диабетическая кома проявляется в резком нарушении всех функций организма с потерей сознания. Основные предшественники диабетической комы ацидоз дегидратация д др ц тканей. Параллельно кетоацидозу развивается нарушение водно-электролитного обмена. В его основе – гипергликемия, повышение осмотического давления в сосудистом русле. Компенсаторное перемещение жидкости из клеток и внеклеточного пространства в сосудистое русло для сохранения осмолярности ведёт к потере тканями воды и электролитов, прежде всего ионов Na+, K+, С1-, НСО3. Развиваются тяжёлая клеточная дегидратация, дефицит внутриклеточных ионов (прежде всего К+), возникает общая дегидратации, снижается периферического кровообращения, б уменьшение мозгового и почечного кровотока и гипоксия. Поздние осложнения сахарного диабета Диабетические ангиопатии • макроангиопатии проявляются в поражениях крупных и средних сосудов сердца, мозга, оз а, нижних конечностей о е ос е • микроангиопатии повреждения капилляров и мелких сосудов ((нефропатия, фр р ретинопатия, нефропатия) фр ) Главная причина поздних осложнений гипергликемия Механизмы развития поздних осложнений 9 гликозилирование белков • • неферментативное - гликопротеиновые белки ферментативное ф р - гемоглобин, кристаллины, р белки межклеточного матрикса, р белки базальных мембран) 9 повышение скорости превращения глюкозы в сорбитол и его накопление в клетках ((токсичен для клеток)) Диагностика сахарного диабета Тест толерантности к глюкозе уровень глюкозы в плазме крови выше 10 ммоль/л через 2 ч после сахарной нагрузки (N – 3,5-5,5 ммоль/л) 9 определение гликозилированного HbA1c уровень НbА1с, в норме составляет около 5% от всего содержания гемоглобина, при СД увеличивается в 2-3 раза 9определение уровня инсулина и С-пептида в крови и моче в норме инсулин и С-пептид секретируются в эквимолярных концентрациях. Поскольку печенью задерживается примерно 2/3 инсулина, соотношении соотношение инсулин/С-пептид в воротной вене и периферических сосудах в норме составляет 1/3 9 определение уровня альбумина б в моче суточное выведение альбумина 30-300 мг (N – 8 мг) Группы лекарственных средств для медикаментозной терапии СД 2 типа ингибиторы всасывания углеводов (гуарем, акарбоза, меглитол и др.); в желудочно-кишечном желудочно кишечном тракте бигуаниды - угнетают глюконеогенез, глюконеогенез уменьшают всасывание глюкозы из ЖКТ и повышают ее утилизацию в тканях; снижают концентрацию ТАГ, холестерина и ЛПНП, не изменяя концентрацию ЛПВП крови; стабилизируют массу тела. тела При отсутствии инсулина терапевтический эффект не проявляется. (Метформин, глиформин, сиофор и др.) производные ро з од е сульфонилмочевины су фо о е - сстимуляторы у ор се секреции рец инсулина, су а, связываются с рецепторами мембраны β-клеток, блокируют АТФчувствительные К+ каналы, мембрана деполяризуется, открываются потенциал-зависимые К+ каналы, возрастает р концентрация р К+ в клетках, что стимулирует секрецию инсулина. (Глибенкламид, глипизид, гликлазид, гликвидон и глимепирид) тиазолидиндионы, или глитазоны: избирательно стимулируют рецептор PPARγ и PPAR-α, изменяют транскрипцию инсулин чувствительных генов, контролирующих метаболизм глюкозы и жиров в жировых тканях, мышцах и печени, повышают чувствительность к инсулину в печени и периферических тканях, снижают уровень глюкозы, инсулина и гликозилированного Hb (Росиглитазон - авандия, и пиоглитазон - актос) ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ВОДНО-СОЛЕВОГО Д ОБМЕНА РЕГУЛЯЦИЯ ОБМЕНА ИОНОВ КАЛЬЦИЯ И ФОСФАТОВ ЭЛЕКТРОЛИТНЫЙ СОСТАВ ЖИДКОСТЕЙ ОРГАНИЗМА Важнейшие параметры вводно вводно-солевого солевого гомеостаза - осмотическое давление, давление рН и объём внутриклеточной и внеклеточной жидкости. Изменение этих параметров может привести к изменению АД, ацидозу или алкалозу, дегидратации и отёкам тканей. а е Основные гормоны, участвующие в тонкой регуляции водно-солевого баланса и действующие на дистальные извитые канальцы и собирательные трубочки почек: антидиуретический ур гормон р ((АДГ), Д ) альдостерон р и предсердный р р натриуретический фактор (ПНФ) Вещество Плазма крови, ммоль/л / Межклеточная жидкость, ммоль/л Внутиклеточная жидкость, ммоль/л Na+ 140 140 10 K+ 5 16 160 Mg2+ 1 0,8 7 Ca2+ C 25 2,5 13 1,3 - Cl- 100 110 2 HCO3- 30 25 8 H3PO4 1,2 1,2 - Белки (%) 7 0,5 20 Схема функционирования почек Почечное тельце Проксимальный извитой каналец Собирательная р трубочка Нисходящая часть петли Генле Восходящая часть петли Генле (4) и дистальный извитой каналец (5) Антидиуретический гормон (AДГ, вазопрессин) П Пептид из 9 а.к., молек. масса -1000 1000 Д Да, 1 S S-SS мостик Место синтеза — нейроны гипоталамуса. Транспорт в нервные окончания в задней доле гипофиза. Т Транспортный йб белок — нейрофизин. й ф Стимулы секреции - [Na+] и осмотического давления внеклеточной жидкости. Улавливают сигнал осморецепторы гипоталамуса. Р Рецепторы — V1 (на ( мембране б ГМК сосудов)) и V2 (на ( базолатеральной б й мембране клеток собирательных трубочек и дистальных канальцев). V2 АЦ V1 ФЛС ПКА ИФЗ+ДАГ экспрессия гена белка аквапорин-2 Ca+2 задержка воды сосудосуживающее действие Биологическое действие АДГ в клетках почечных канальцев Нейрофизин АДГ АДГ синтезируется ру в нейронах р гипоталамуса у в виде предшественника препрогормона, который поступает в аппарат Гольджи и превращается в прогормон. В составе нейросекреторных гранул прогормон переносится в нервные окончания задней доли гипофиза (нейрогипофиз). Во время транспорта гранул происходит процессинг прогормона, в результате чего он расщепляется на зрелый гормон и транспортный белок б - нейрофизин. й Гранулы, содержащие зрелый антидиуретический гормон и нейрофизин, хранятся в терминальных расширениях аксонов в задней й доле гипофиза, ф из которых секретируются в кровоток при соответствующей стимуляции. Стимул секреции АДГ oповышение концентрации ионов натрия oувеличение осмотического давления внеклеточной жидкости Несахарный диабет (нерегулируемая экскреция воды) Центральный несахарный диабет Причины 9 генетические дефекты ф синтеза препро-АДГ АДГ 9 генетические дефекты процессинга и транспорта про-АДГ 9 повреждения гипоталамуса или нейрогипофиза Основное проявление o гипотоническая полиурия o усиленное потребление воды воды. Диагностические критерии oвыраженная полиурия (до 20 л/сутки); oплотность мочи <1,001 (N — 1,002). Нефрогенный несахарный диабет Причина р oмутации гена V2 рецептора АДГ - почки не реагируют на гормон. Альдостерон (минералкортикостероид) Место синтеза — кора надпочечников Транспортых белков нет, может образовывать связи с альбумином. Стимулы у секреции р ц – [[Na,, [К [ +] в плазме крови, р , АКТГ,, анигиотензин II. Рецепторы — локализованы в ядре и в цитоплазме. Комплекс гормон-Ре взаимодействует с ДНК и изменяет скорость транскрипции специфических генов. : Альдостерон в клетках почечных канальцев вызывает oиндукцию синтеза белков белков-транспортеров транспортеров Na+ из канальца в клетку; oиндукцию синтеза Na+, К+-АТФазы (перенос Na+ из клетки в МКП, а К+ - из МКП в клетку; oиндукцию белков-транспортеров К+ из клеток канальца в мочу; oиндукцию синтеза митохондриальных ферментов ЦТК, стимулирующих oобразование АТФ для активного транспорта ионов Суммарный эффект действия альдостерона увеличение реабсорбции ионов Na+ в канальцах нефронов задержка Na+ в организме экскреции К+ Гормон стероидной природы Механизм действия альдостерона Юкстагломерулярный аппарат 1 – приносящая артериола 2 – выносящая артериола 3 – дистальный й извитой й каналец 4 – плотное пятно 5 – юкстагломерулярные клетки с гранулами ренина 6 – юкставаскулярные клетки Снижение давления в приносящих артериолах – стимул для секреции ренина. АНГИОТЕНЗИН II Ангиотензин-превращающий Ангиотензин превращающий фермент Ренин (карбоксидипептидилпептидаза) Ангиотензиноген Асп-Арг-Вал-Тир-Иле-Гис-Про-Фен-Гис-Лей-Лей Асп-Арг-Вал-Тир-Иле-Гис-Про-Фен-Гис-Лей-Лей………………~400а.к ~400а к 1 2 3 4 5 6 7 8 9 10 11 Ангиотензин I Ангиотензин II Р Рецепторы ангиотензина II клетки клубочковой зоны коры надпочечников клетки гладких мышц Ангиотензин II Рецептор ангиотензина II ФЛС ДАГ+ИФ3 ПКС Система ренин-ангиотензин-альдостерон Н2О АДГ Альдостерон Главный механизм регуляции синтеза и секреции альдостерона – Система ренин-ангиотензина-Альдостерон Ренин, протеолитический фермент, катализирует превращение ангиотензиногена ((гликопротеина) р ) в ангиотензин I ((декапептид). ) Ангиотензин I превращается в ангиотензин II под действием АПФ, отщепляющего два аминокислотных остатка от декапептида Ангиотензин II стимулирует синтез и секрецию альдостерона Ангиотензин II вызывает сужение сосудов периферических артерий Альдостерон А стимулирует реабсорбцию б б N Na+ и экскрецию К+ Секреции ренина и альдостерона регулируется по механизму отрицательной обратной связи (пунктирные линии) ГИПЕРАЛЬДОСТЕРОНИЗМ Первичный гиперальдостеронизм – синдром Конна Причины 9аденома надпочечников – 80% случаев 9диффузная д ффу гипертрофия р р ф клеток клубочковой у зоны,, вырабатывающих альдостерон – 20% случаев. Следствие: усиление реабсорбции Na в почечных канальцах → увеличение [Na] в плазме → увеличение секреции АДГ → задержка воды почками, гипернатриемия, гипертония, гиперволемия, отеки, гипокалиемия → мышечная слабость, дефицит магния, легкий метаболический алкалоз. Вторичный гиперальдостеронизм Причины П 9сердечная недостаточность 9хронические заболевания почек 9опухоли секретирующие ренин 9опухоли, Следствие: Повышен уровень ренина и ангиотензина II → стимуляция секреции альдостерона. Клинические симптомы менее выражены. Дифференциальный диагноз первичного и вторичного гиперальдостеронизма определение ренина и альдостерона в плазме крови Активность ренина Концентраци я альдостерона Первичный гиперальдостеронизм Вторичный гиперальдостеронизм Снижена Повышена Повышена Повышена МЕХАНИЗМ ВОЗНИКНОВЕНИЯ ПОЧЕЧНОЙ ГИПЕРТЕНЗИИ Стеноз почечной артерии Снижение перфузионного рфу давления д в почках Секреция ренина Образование ангиотензина II Сужение сосудов Секреция альдостерона Жажда Секреция вазопрессина Потребление П б воды АД Задержка З NaCl Задержка воды ИНГИБИТОРЫ АПФ В ЛЕЧЕНИИ АРТЕРИАЛЬНОЙ ГИПЕРТЕНЗИИ Ингибиторы АПФ приводят также к снижению притока крови к сердцу, что уменьшает нагрузку на сердце сердце, и защищают почки от воздействия гипертензии. гипертензии Ингибиторы АПФ используются и для лечения сердечной недостаточности, инфаркта миокарда и для предупреждения патологии почек вследствие гипертензии гипертензии. o o o o o o o o o o Капотен (каптоприл) Энам (энап, эналоприл, вазотек) Привинил (лизиноприл) Лотензил (беназеприл) Моноприл (фозиноприл) Алтейс (рамиприл) Аккуприл (квиноприл) Асеон (периндоприл) Мавик (трандолаприл) У Униваск (моэксиприл) ( ) ПРЕДСЕРДНЫЙ НАТРИЙУРЕТИФЕСКИЙ ФАКТОР (ПНФ) МЕСТО СИНТЕЗА — КАРДИОМИОЦИТЫ ПРЕДСЕРДИЙ СТИМУЛЫ CEKPЕЦИИ: ПОВЫШЕНИЕ АД ПОВЫШЕНИЕ ОСМОЛЯРНОСТИ ПЛАЗМЫ ПОВЫШЕНИЕ ЧАСТОТЫ СЕРДЕЧНЫХ СОКРАЩЕНИЙ Й ПОВЫШЕНИЕ КОНЦ-ИИ КАТЕХОЛАМИНОВ ПОВЫШЕНИЕ КОНЦ-ИИ ГЛЮКОКОРТИКОИДОВ МИШЕНИ ПНФ: ПНФ ПОЧКИ, ПОЧКИ ПЕРИФЕРИЧЕСКИЕ АРТЕРИИ ПОЧКИ — РАСШИРЕНИЕ ПРИНОСЯЩИХ АРТЕРИОЛ УСИЛЕНИЕ ПОЧЕЧНОГО КРОВОТОКА УВЕЛИЧЕНИЕ СКОРОСТИ ФИЛЬТРАЦИИ УВЕЛИЧЕНИЕ ЭКСКРЕЦИИ ИОНОВ Na+ ПЕРИФЕРИЧЕСКИЕ АРТЕРИИ — СНИЖЕНИЕ ТОНУСА ГЛАДКИХ МЫШЦ, РАСШИРЕНИЕ АРТЕРИОЛ РЕЗУЛЬТАТ ДЕЙСТВИЯ ПНФ — ПОВЫШЕНИЕ ЭКСКРЕЦИИ ИОНОВ Na+, ПОНИЖЕНИЕ АД РЕЦЕПТОР ПНФ — ИМЕЕТ ДОМЕН, ОБЛАДАЮЩИЙ Й ГУАНИЛАТЦИКЛАЗНОЙ Й АКТИВНОСТЬЮ: ГТФ цГМФ ПKG Регуляция активности мембранной (1) и цитоплазматической (2) гуанилатциклазы Предсердный натрийуретический пептид состоит из 28 аминокислот. В центре молекулы 17 аминокислот образуют кольцевую структуру с помощью дисульфидной связи между двумя остатками цистеина в 7 и 23 положениях. Биологические последствия действия ПНФ ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ИОНОВ КАЛЬЦИЯ И ФОСФАТОВ Ионы кальция 9 увеличивают проницаемость мембраны клеток для ионов К+, 9 влияют на натриевую проводимость клеток, 9 влияют на р работу у ионных насосов,, 9 способствуют секреции гормонов, 9 участвуют в каскадном механизме свёртывания крови, 9 уучаствуют у вр регуляции у активности ферментов, ф р 9 участвуют во внутриклеточной передаче сигналов. И Изменение уровня кальция может приводить: o к нарушению функционирования кальциевого насоса o к снижению активности ферментов o к нарушению гормональной регуляции метаболизма o к изменению порога возбудимости нервных и мышечных клеток Роль транспортных белков клеточной мембраны и эр в поддержании внутриклеточной концентрации Са2+ Na+, Са 2+-переносчик Белки млекопитающих, связывающие кальций Белок Са2+-АТФаза Na+/Са2+- обменник Рецептор рианодина Рецептор ИФ3 Са2+-активируемый калиевый канал Кальмодулин Протеинкиназа С Кальпаин К Кальцинейрин В Тропонин С Фосфолипаза А2 Фосфолипаза С Парвальбумин Кальсеквестрин Функция в клетке Осуществляет активный транспорт Са2+ из клетки или в цистерны ЭПР Осуществляет ущ выход д Са2+ из клетки в обмен на вход ионов Na (активный антипорт) Вызывает выход Са2+ из ЭПР Вызывает выход Са2+из ЭПР Вызывает гиперполяризацию мембраны Активатор многих кальмодулин-зависимых ферментов Фосфорилирование белков по остаткам серина и треонина П Протеаза Фосфатаза фосфобелков Регулятор сокращения скелетных и сердечной мышц Образует арахидоновую кислоту Образует ИФ3 и ДАГ Буфер кальция Буфер кальция КАЛЬЦИЙ Основное депо - кости. o99% 99% кальция - в виде гидроксиапатита [Са10(РО4)6(ОН)2Н2О]. О] o1% кальция - виде фосфатных солей. Он играет роль буфера при изменениях концентрации Са в плазме крови. Другой фонд Са (1% от общей массы Са) – oкальций плазмы крови. Концентрация Са2+ в плазме крови - 2,12-2,6 ммоль/л, или 9-11 мг/дл Кишечник Костная ткань Плазма крови Кальций присутствует в плазме крови : o в виде несвязанного ионизованного кальция – 50%; o в соединении с белками б – 45%; 45% o в виде недиссоциирующих комплексов с цитратом, сульфатом, фосфатом и карбонатом (5%). Ионы кальция 9 увеличивают проницаемость мембраны клеток для ионов К+, 9 влияют на натриевую проводимость клеток, 9 влияют на р работу у ионных насосов, 9 способствуют секреции гормонов, 9 участвуют в каскадном механизме свёртывания крови, 9 участвуют в регуляции активности ферментов, 9 участвуют во внутриклеточной передаче сигналов. Изменение уровня кальция может приводить: o к нарушению функционирования кальциевого насоса o к снижению активности ферментов o к нарушению гормональной регуляции метаболизма o к изменению порога возбудимости нервных и мышечных клеток Паратгормон (ПТГ) Строение ПТГ - полипептид, 84 а.к.(9,5 кД), препрогормон содержит 115 а.к. Стимул секреции - снижении концентрации Са22+ Механизм действия – через мембранные рецепторы и активацию аденилатциклазы Органы-мишени – кости и почки Эффекты В костной ткани рецепторы ПТГ обнаружены в остеобластах и остеоцитах. 9стимулирует у ру мобилизацию ц ионов Са и фосфатов ф ф из кости во внеклеточную у жидкость. В почках 9стимулирует реабсорбцию Са2+ в дистальных извитых канальцах, 9 снижает экскрецию Са с мочей, 9 уменьшает реабсорбцию фосфатов, 9 стимулирует синтез кальцитриола. Биологическое действие паратгормона Избыток паратгормона Гиперпаратиреоидизм – при опухолях из клеток паращитовидных желез Увеличение мобилизации Са из костей Увеличение реабсорбции Са в почках Увеличение концентрации Са в крови (гиперкальциемия) Кальцификация ц ф ц мягких тканей Образование камней в почках ДЕФИЦИТ ПАРАТГОРМОНА ГИПОПАРАТИРЕОИДИЗМ – при у удалении или повреждении паращитовидных желез Уменьшение мобилизации У б С Са из костей Увеличение выведения Са У С с мочей Снижение концентрации Са в крови ( (гипокальциемия) ) Повышение нервно-мышечной возбудимости (снижение порога возбуждения клеток) Судороги, гиперрефлексы, спазмы гортани Витамины группы D УФО Витамин D3 Холекальциферол 7-дегидрохолестерол УФО Эргостерин Витамин D2 Эргокальциферол Схема синтеза кальцитриола Холестерол Кожа 7-Дегидрохолестерол УФ Холекальциферол Витамин D3 Печень Кальцидиол Почки Кальцитриол Транскальциферин р ф р - транспортный р р белок для холекальциферола, кальцидиола и кальцитриола. Мишени кальцитриола – тонкий кишечник, почки и кости. Недостаточность витамина Д (Д2 и Д3) Суточная потребность у детей - 12 – 50 мкг (500 – 1000 МЕ) Рахит Остеомаляция Остеопороз Роль витамина Д вне метаболизма кальция Контроль состояния мышц, ССС, участвует в регуляции роста, важен в предупреждении рака, диабета, ревматоидного артрита, расеянного склероза Причины и механизмы развития рахита Недостаток УФ Нарушение всасывания жирорастворимых витаминов Снижение активности гидроксилаз Дефицит D3 Нарушение образования кальцитриола Нарушение всасывания Са в кишечнике Снижение концентрации Са в крови Стимуляция секреции паратгормона Вымывание Са из костей Нарушение образования костной ткани ткани, деформация скелета Гипервитаминоз D Гипервитаминоз D (интоксикация витамином D) - состояние, обусловленное как прямым токсическим действием препарата на клеточные мембраны, так и повышенной концентрацией ц р ц в крови р солей кальция, ц , откладывающихся д щ в стенках сосудов внутренних органов, в первую очередь почек и сердца. Острая интоксикация витамином D - нейротоксикоз О й или кишечный й токсикоз: резко снижен аппетит, жажда, рвота, быстро уменьшается масса тела, развивается обезвоживание, появляются запоры (возможен неустойчивый и жидкий й стул). ) У некоторых детей й регистрируют кратковременную потерю сознания, возможны тонико-клонические судороги. Хроническая интоксикация витамином D возникает на фоне длительного (6-8 мес и более) применения препарата в превышающих физиологическую потребность дозах. дозах Клиническая картина выражена менее отчётливо и включает повышенную раздражительность, плохой сон, слабость, боли в суставах, постепенное нарастание дистрофии, преждевременное закрытие большого родничка, изменения со стороны сердечно-сосудистой сердечно сосудистой и мочевой систем. Кальцитонин Природа гормона Пептидный гормон гормон, содержит 32 аминокислоты аминокислоты, секретируется парафолликулярными клетками щитовидной железы (С-клетками) при повышении концентрации Са2+ в крови. Механизм действия Рецептор кальцитонина - трансмембранный белок (7 трансмембранных сегментов). Связывание кальцитонина сопровождается активацией аденилатциклазной системы, системы фосфоинозитольного и Са 2+-зависимого внутриклеточных сигнальных путей. Эффекты В Вызывает угнетение активности остеокластов и остеоцитов и ингибирует б резорбцию костной ткани. Стимулирует остеобласты. В почках кальцитонин усиливает экскрецию фосфатов, ионов кальция и натрия, натрия угнетает активность 1α-гидроксилазы, 1α гидроксилазы что уменьшает синтез кальцитриола. В тонкой кишке кальцитонин тормозит всасывание кальция. Кальцитонин - антагонист ПТГ ПТГ. Уровень секреции кальцитонина зависит от концентрации эстрогенов. При дефиците эстрогенов (менопауза или заболевания яичников) секреция кальцитонина снижается, что способствует б ускоренной й резорбции б костной й ткани и приводит к остеопорозу и переломам костей. Лекция « МЕТАБОЛИЗМ ГЕМА И ОБМЕН ЖЕЛЕЗА» Авторы – доцент Т.А.Титова и доцент Е.С.Зыкова СИНТЕЗ ГЕМА Гем является простетической группой гемоглобина, миоглобина, цитохромов, а также коферментом каталазы, пероксидазы, цитохромоксидазы. Гем синтезируется в большинстве клеток, за исключением эритроцитов. Железо, необходимое для синтеза гема, поступает с пищей или освобождается при распаде гема. Гем состоит из иона Fe+2 , входящего в структуру порфирина, который состоит из 4-х пиррольных колец, связанных между собой метеновыми «мостиками». Порфирины могут отличаться заместителями в пиррольных кольцах. Гем гемоглобина содержит протопорфирин IХ, имеющий два винильных, четыре метильных радикала и два остатка пропионовой кислоты (рис.1). Рис. 1. Субстратами для синтеза гема являются глицин, сукцинил-КоА и Fe+2 Первая и три последние реакции синтеза идут в матриксе митохондрий, а остальные – в цитоплазме. В матриксе митохондрий из глицина и сукцинил-КоА под действием пиридоксальфосфат зависимого фермента 5-аминолевулинатсинтазы образуется 5-аминолевулиновая кислота, которая поступает в цитоплазму. В цитоплазме фермент 5-аминолевулинатдегидратаза катализирует конденсацию двух молекул 5-аминолевулината с образованием порфобилиногена. Далее из двух молекул порфобилиногена последовательно образуются промежуточные метаболиты порфириногены. Последний из них поступает в митохондрии и превращается в протопорфирин IХ. Фермент феррохелатаза присоединяет Fe+2 к протопорфирину IХ и таким образом образуется гем (рис.2). Рис. 2. Две первые реакции синтеза гема аллостерически ингибирует гем по принципу отрицательной обратной связи. Синтез аминолевулинатсинтазы индуцирует Fe+2 . В свою очередь сам гем индуцирует синтез α- и βполипептидных цепей гемоглобина (рис.3). Рис. 3. Нарушения синтеза гема могут быть наследственными и приобретенными. Они сопровождаются повышением содержания промежуточных метаболитов синтеза гема, а также продуктов их окисления в клетках и называются порфириями. Эти вещества поступают в мочу, окрашивая ее в красный цвет. При генетических дефектах ферментов, катализирующих превращения промежуточных метаболитов синтеза гема порфириногенов, наблюдаются наследственные порфирии. Порфириногены являются нейротоксинами и ускоряют образование активных форм кислорода, которые стимулируют перекисное окисление липидов (ПОЛ) и разрушение мембран клеток. Легкие формы порфирий могут протекать бессимптомно, но прием лекарств, являющихся индукторами синтеза 5аминолевулинатсинтазы (сульфаниламиды, барбитураты, стероиды) могут вызвать обострение болезни. Порфирии сопровождаются нейропсихическими расстройствами, нарушениями ретикулоэндотелиальной системы, фотосенсибилизацией. ОБМЕН ЖЕЛЕЗА Железо входит в состав гемсодержащих белков и ферментов (миоглобина, гемоглобина, цитохромов, цитохромоксидазы, пероксидазы, каталазы), железофлавопротеинов ( NADH-дегидрогеназы, сукцинатдегидрогеназы, ксантиноксидазы, ацил-КоАдегидрогеназы) и железосодержащих белков трансферрина и ферритина. В организме здорового человека содержится 3-4г железа. Оно поступает с пищей или освобождается при распаде гема и железосодержащих белков. В пище содержится Fe+2 и Fe+3 . Наиболее богаты железом печень и почки животных, яйца, мясо. В растительных продуктах железа содержится меньше, вместе с тем им относительно богаты овсянка, персики, грибы, гранаты. Однако в растениях железо связано с фитиновой кислотой и поэтому плохо усваивается. Ионы железа легко вступают в окислительно-восстановительные реакции и образуют хелатные соединения со многими веществами. Это может привести к изменению их свойств и функций. Поэтому железо транспортируют и депонируют особые белки. По крови железо транспортирует трансферрин, а в клетках депонирует ферритин. При связывании железа с белками оно окисляется, а при освобождении из белков – восстанавливается. В суточном количестве пищи содержится около 20 мг железа, а всасывается не более 2 мг. В полости кишечника железо освобождается из белков и органических солей пищи. В клетки слизистой кишечника всасывается закисное железо - Fe+2 . Витамин С восстанавливает железо и улучшает его усвоение. В клетках слизистой кишечника избыток железа соединяется с белком апоферритином, который окисляет железо и превращается в ферритин. Поступающее из клеток кишечника в кровь железо окисляется ферментом плазмы крови ферроксидазой. В крови Fe+3 транспортирует белок трансферрин. В клетках железо используется для синтеза железосодержащих белков или депонируется в ферритине (рис.4). Рис. 4. Ферритин это олигомерный белок, состоящий из 24 протомеров. Они образуют полую сферу, внутри которой может содержаться около 3000 ионов железа в составе гидроксифосфата. Ферритин окисляет Fe+2 . В белковой части этого белка есть каналы, через которые железо может поступать и выходить из полости сферы. Из организма в сутки со слущенным эпителием выводится около 1 мг железа. При нарушениях метаболизма железа возникают железодефицитная анемия или гемохроматоз. Железодефицитная анемия развивается при недостатке железа в организме, который может возникнуть при частых кровотечениях, анацидном гастрите, беременности, родах, язвах и опухолях желудочно-кишечного тракта, снижении всасывания железа в кишечнике. При этой анемии уменьшается размер эритроцитов, снижается интенсивность их окраски, содержание гемоглобина в крови, насыщение железом трансферрина, содержание ферритина в клетках. Причина этих изменений – снижение синтеза гема, гемоглобина и апоферритина из-за дефицита железа. Больных лечат лекарствами, содержащими железо. Гемохроматоз - это наследственное заболевание, которое сопровождается повышением всасывания железа в кишечнике. При этом железо откладывается в белковой части ферритина, который в результате этого превращается в гомосидерин. Накопление гранул гомосидерина в клетках печени, селезенки, поджелудочной железы приводит к гемохроматозу. Содержание железа в организме больных может достигать 40 г. Накопление железа в клетках поджелудочной железы приводит к сахарному диабету, в гепатоцитах – к циррозу печени, в кардиоцитах – к сердечной недостаточности. Больных лечат кровопусканиями и препаратами, связывающими железо. КАТАБОЛИЗМ ГЕМА Наибольшее количество гема содержит гемоглобин, который находится в эритроцитах. Они циркулируют в крови около 120 дней, а затем распадаются в клетках ретикулоэндотелиальной системы селезенки, костного мозга и печени. В процессе распада гема можно выделить несколько последовательных этапов (рис.5). Гемоглобин Гем Гемоксигеназная система Биливердин Биливердинредуктаза Билирубин Непрямой билирубин Билирубин 2УДФ-глюкуронат Глюкуронилтрансфераза I и II Рис. 5. • В клетках ретикулоэндотелиальной системы гемоглобин под действием фермента гемоксигеназы превращается в вердоглобин. Вердоглобин состоит из глобина, Fe+2 и линейного тетрапиррола биливердина. Теряя глобин и железо, он превращается в желчный пигмент желто-зеленого цвета биливердин. • Биливердинредуктаза восстанавливает биливердин в желчный пигмент красного цвета билирубин, который поступает в кровь. • В крови гидрофобный билирубин соединяется с белком альбумином. Билирубин, связанный с альбумином, называют непрямым (неконъюгированным), так как он не дает положительную качественную реакцию (розовое окрашивание) с диазореактивом. Непрямой билирубин плохо растворим в воде, токсичен и не обнаруживается в моче, так как связан с альбумином. • Неконъюгированный билирубин поступает в клетки печени. В эдоплазматическом ретикулуме гепатоцитов билирубин под действием фермента глюкуронилтрансферазы соединяется с глюкуроновой кислотой с образованием диглюкуронидбилирубина. Диглюкуронидбилирубин называют прямым билирубином. Он растворим в воде, не токсичен и взаимодействует с диазореактивом. Около 75% прямого билирубина из клеток печени поступает в желчные капилляры и тонкий кишечник, а остальное - в кровь. • Под действием микрофлоры кишечника прямой билирубин освобождается от глюкуроновой кислоты и превращается в уробилиноген. До 95% уробилиногена поступает в толстый кишечник и называется стеркобилиногеном. На свету стеркобилиноген, окисляясь превращается в пигмент кала стеркобилин (выводится 200-300мг в сутки). Примерно 5% уробилиногена из кишечника идет в кровь, откуда частично улавливается печенью, где распадается до пирролов, поступающих в желчь. Другая часть из крови поступает в почки и мочу. На свету уробилиноген превращается в пигмент мочи уробилин (выводится 3-4мг в сутки). В норме в крови содержится 2,0-20,0 мкмоль/л (0,1-1 мг/дл) билирубина. ДИАГНОСТИЧЕСКОЕ ЗНАЧЕНИЕ СОДЕРЖАНИЯ БИЛИРУБИНА, СТЕРКОБИЛИНА В КРОВИ И МОЧЕ ОПРЕДЕЛЕНИЯ УРОБИЛИНА И Повышение концентрации билирубина в крови называется билирубинемия, а присутствие прямого билирубина в моче – билирубинурия. Моча при этом приобретает коричневый цвет. При повышении концентрации билирубина в крови более 2-3 мг/дл кожа, склеры глаз, слизистые оболочки желтеют и такие симптомы называются желтухой. Для дифференциальной диагностики желтух в крови определяют прямой и непрямой билирубин, а к моче и кале уробилин и стеркобилин соответственно. • Гемолитическая желтуха возникает при усиленном гемолизе эритроцитов. В этом случае повышается поступление билирубина в кровь по сравнению с нормой, и концентрация прямого билирубина увеличивается до верхней границы нормы. Так как печень не справляется с обезвреживанием повышенного количества билирубина, то в крови резко возрастает концентрация непрямого билирубина. Непрямой билирубин в мочу не поступает, так как связан с альбумином. В моче больных билирубина нет, но повышено содержание уробилина, а в кале – стеркобилина. • Механическая (обтурационная) желтуха наблюдается при закупорке желчных протоков камнями или опухолью. В этом случае прямой билирубин, который образуется в печени, не может поступать в кишечник, а идет в кровь. Концентрация прямого билирубина при механической желтухе превышает почечный порог, поэтому он поступает в мочу, окрашивая ее в коричневый цвет. В моче и кале отсутствуют желчные пигменты, поэтому кал – обесцвеченный (ахолический). • Паренхиматозная (печеночно-клеточная) желтуха развивается при вирусном гепатите. При этом заболевании снижается способность гепатоцитов улавливать непрямой билирубин из крови и экскретировать прямой билирубин в кишечник. Поэтому в крови повышается прямой и непрямой билирубин, а в моче и кале снижается содержание уробилина и стеркобилина соответственно. У больных моча коричневая, а кал светлый. • Желтуха новорожденных обусловлена запаздыванием включения гена глюкуронилтрансферазы и недостаточной способностью рецепторов гепатоцитов улавливать непрямой билирубин из крови. • Наследственные желтухи связаны с генетическими дефектами ферментов и белков, участвующих в метаболизме билирубина. Например, при генетическом дефекте белков, захватывающих билирубин из крови, развивается синдром Жильбера, при дефекте белков экскретирующих билирубин в кишечник – синдром Дубина-Джонсона, а при нарушении структуры глюкуронилтрансферазы – синдром Криглера-Найяра. Лекция «БИОХИМИЯ КРОВИ» Авторы – доцент Т.А.Титова и доцент Е.С.Зыкова Кровь циркулирует по системе кровеносных сосудов и, транспортируя различные вещества, выполняет следующие функции: • дыхательную (гемоглобин, находящийся в эритроцитах, переносит О2 из легких в ткани и СО2 из тканей в легкие); • трофическую (транспорт продуктов переваривания пищи из клеток кишечника в ткани); • выделительную (уносит продукты катаболизма в выделительные органы); • регуляторную (транспортирует сигнальные молекулы к тканяммишеням); • кровь сохраняет кислотно-щелочной и водный баланс и поддерживает температуру тела; • защитную (кровь содержит клеточные и гуморальные компоненты иммунного ответа). К этой функции относится также способность крови свертываться. БЕЛКИ ПЛАЗМЫ КРОВИ Плазма крови содержит 7% белков всего организма, причем концентрация альбумина составляет 40-50г/л, глобулинов 20-30г/л, общий белок плазмы крови – 60-70г/л. Белки плазмы крови методом электрофореза можно разделить на фракции, количество которых в зависимости от типа электрофореза может составлять от 5 до 17. Электрофорезом на бумаге можно обнаружить 5 фракций: альбумин, α1, α2, β и γ- глобулины. Большинство белков плазмы синтезируются в печени, но некоторые образуются и в других органах. Например, γ- глобулины синтезируются В-лимфоцитами, большинство пептидных гормонов секретируют эндокринные железы, а гормон эритропоэтин образуется в почках. Белок альбумин синтезируется в печени и имеет небольшую молекулярную массу - 69 КДА. Молекула альбумина содержит много дикарбоновых аминокислот, поэтому удерживает катионы Na+2 , Ca2+ , Zn2+. Небольшая молекулярная масса альбумина и высокая концентрация в крови обеспечивает до 80% осмотического давления плазмы. Альбумин является важнейшим транспортным белком. Он транспортирует, например, свободные жирные кислоты, неконъюгированный билирубин, триптофан, тироксин, трииодтиронин. Глобулины делят на 4 фракции α1, α2, β и γ. В эти фракции входят белки, которые выполняют специфические и защитные функции. К глобулинам относятся, например, тироксин- и кортизолсвязывающие белки, трансферрин, гаптоглобин, церулоплазмин, интерфероны, С реактивный белок , появляющийся в крови при некоторых воспалительных заболеваниях. При ряде заболеваниях содержание белков в плазме крови изменяется. Такие изменения называют диспротеинемиями. Гиперпротеинемия – это повышение концентрации белков в плазме крови. Она может быть первичной и вторичной. Первичная гиперпротеинемия обусловлена повышением концентрации γ-глобулинов и некоторых специфичесеских белков при острых воспалительных заболеваниях. Такие белки называют белками острой фазы. К ним относят С-реактивный белок, α1-антитрипсин, гаптоглобин, фибриноген. Синтез этих белков в печени индуцирует белок мононуклеарных фагоцитов интерлейкин-1. Вторичная гиперпротеинемия обусловлена потерей воды при диарее, рвоте, полиурии. Гипопротеинемия наблюдается главным образом при потере организмом альбумина при нефрите и снижении его синтеза, например, при гепатитах, циррозе печени, длительном голодании. Уменьшение содержания альбумина в крови приводит к снижению осмотического давления и нарушению распределения жидкости между сосудистым руслом и межклеточным пространством. Клинически это проявляется в виде отеков. МЕТАБОЛИЗМ ЭРИТРОЦИТОВ Эритроциты – это высокоспециализированные клетки, которые не имеют клеточных органелл. Гемоглобин составляет 95% их сухого остатка. Эритроциты циркулируют в крови около 120 дней и разрушаются в печени, селезенке и костном мозге. Эритропоэз регулирует гормон эритопоэтин, синтезирующийся в почках. Двояковогнутая форма эритроцитов и особенности строения их плазматической мембраны обеспечивают пластичность этих клеток при прохождении мелких капилляров. В эритроцитах нет митохондрий, поэтому единственным источником энергии является анаэробный гликолиз. Глюкоза поступает в эритроциты с помощью белков-переносчиков ГЛЮТ-2. В анаэробном гликолизе используется примерно 90% глюкозы, содержащейся в эритроцитах, остальное - в пентозофосфатном пути. Анаэробный гликолиз: а) обеспечивает синтез АТФ для работы транспортных АТФ-аз и гексокиназной реакции гликолиза; б) необходим для образования NADH+H + , участвующего в восстановлении метгемоглобина в гемоглобин; в) является источником 1,3-бисфосфоглицерата, который ферментом бисфосфоглицератмутазой превращается в 2,3 бисфосфоглицерат. Последний является аллостерическим регулятором гемоглобина (снижает его сродство к кислороду). В пентозофосфатном пути превращения глюкозы образуется NADРH+H+, являющийся донором водорода в реакции восстановления глутатиона. Кислород, содержащийся в эритроцитах, потенциально токсичен, так как, окисляя Fe+2 и присоединяя электрон, он становится супероксидным анионом, а гемоглобин превращается в метгемоглобин, который не транспортирует кислород. Восстановление метгемоглобина в гемоглобин катализирует метгемоглобинредуктазная система. Коферментом + метгемоглобинредуктазы является NADH+H . Супероксидный анион (О2-) и гидроксил радикал (ОН.) вызывают образование гидроперекисей остатков ненасыщенных жирных кислот в фосфолипидах клеточных мембран, а это приводит к их разрушению и гемолизу эритроцитов. Эритроциты содержат систему ферментов, обезвреживающих активные формы кислорода (рис.1): • супероксиддисмутаза превращает супероксидный анион в пероксид водорода; • глутатионпероксидаза превращает пероксид водорода в воду, а глутатион в глутатион S-Sглутатион; • пероксид водорода может расщепляться каталазой; • окисленный глутатион (G-S-S-G) восстанавливается NADРHзависимой глутатионредуктазой. Рис.1. Любое нарушение системы обезвреживания О2 в эритроцитах приводит к увеличению содержания активных форм кислорода и гемолизу эритроцитов. Гемолиз эритроцитов наблюдается при: • генетических дефектах фермента пентозо-фосфатного пути глюкозо-6-фосфатдегдрогеназы. Лечение пациентов с таким генетическим дефектом лекарствами, обладающими сильными окислительными свойствами (аспирин, примахин, сульфаниламиды, парацетамол) может вызвать гемолиз эритроцитов. • генетических дефектах любого фермента гликолиза, так как при этом снижается синтез АТФ и NADH+H, являющегося коферментом метгемоглобинредуктазы. Падение активности фермента приводит к увеличению содержания метгемоглобина, молекулы которого соединяются дисульфидными связями, образуя тельца Хайнца. Это вызывает разрушение эритроцитов, попадающих в мелкие капилляры. СВЕРТЫВАЮЩАЯ И ПРОТИВОСВЕРТЫВЮЩАЯ СИСТЕМЫ КРОВИ Свертывание крови – это защитная реакция организма. Основную роль в этом процессе играют тромбоциты и белки плазмы крови, которые называют факторами свертывания и обозначают римскими цифрами. Прекращение кровотечения после травмы сосуда называют гемостазом. Гемостаз имеет 4 этапа: I – сокращение поврежденного сосуда. II – образование в месте повреждения в течение 3-5 минут рыхлой тромбоцитарной пробки (белого тромба). Он образуется при взаимодействии поврежденного эндотелия с тромбоцитами, в результате чего изменяются свойства тромбоцитов. Они прилипают к месту повреждения (адгезируют) и друг к другу (агрегируют). III этап заключается в превращении растворимого белка плазмы крови фибриногена в нерастворимый фибрин. Фибрин вместе с эритроцитами откладывается между тромбоцитами белого тромба и таким образом образуется красный тромб (рис.2). Рис.2. IV – растворение тромба. Снижение свертываемости крови ведет к повышению кровоточивости. При повышении свертываемости образуются внутрисосудистые тромбы, которые могут закупорить неповрежденный сосуд и вызвать тромбоз. Нарушения системы свертывания могут быть наследственными и приобретенными. Образование фибринового тромба включает несколько этапов: • Образование мономера фибрина. Растворимый белок плазмы крови фибриноген (I) превращается в нерастворимый мономер фибрин под действием протеолитического фермента крови тромбина (IIа). Фибриноген состоит из шести полипептидных цепей трех типов 2Аα, 2Вβ и 2γ. Цепи связаны между собой дисульфидными связями и образуют три домена. На N-концах Аα, и Вβ цепей находятся А и В участки, выступающие из центрального домена. Эти А и В пептиды содержат много остатков асп и глу, следовательно, заряжены отрицательно, поэтому молекулы фибриногена отталкиваются друг от друга и не агрегируют (рис.3). Рис. 3. • Сериновая протеаза тромбин частичным протеолизом удаляет А и В пептиды и превращает фибриноген в фибрин -мономер. • Образование геля фибрина. В результате изменения конформации в фибрине-мономере образуются комплементарные участки – центры связывания. Это приводит к полимеризации фибрина и образованию нерастворимого геля фибрина. Он непрочен, так как образован не ковалентными связями между комплементарными доменами. • Стабилизация геля. Гель стабилизирует фермент трансглутамидаза (XIIIа), который катализирует образование изопептидных связей между радикалами глутамина и лизина мономеров фибрина. • Сжатие геля осуществляет белок тромбостенин при участии АТФ. Для образования фибринового геля необходим фермент тромбин. Активный тромбин образуется в крови из неактивного протромбина, путем частичного протеолиза. Активацию протротромбина обеспечивает ряд последовательных реакций. Эти реакции инициируются (запускаются) на поврежденной или измененной тромбогенным сигналом клеточной мембране. Каскад реакций свертывания крови ускоряется на каждом этапе и заканчивается образованием тромба (коагуляцией). Реакции, предшествующие превращению фибриногена в фибрин, составляют прокоагулянтный путь свертывания крови. В прокоагулянтном пути последовательно образуются три ферментных комплекса. Каждый комплекс включает: • отрицательно заряженный участок мембран клеток эндотелия или тромбоцитов, • белок-активатор ТФ (тканевой фактор), VIIIa или Va; • один из протеолитическиx ферментов (сериновую протеазу) VIIa, IXa, Xa; эти ферменты в своем составе содержат остатки γ- карбоксиглутаминовой кислоты. Они образуются при посттрансляционном карбоксилировании глутамата при участии витамина К; • Са+2 Таким образом, на отрицательно заряженном участке мембраны последовательно формируются три мембранных комплекса: VIIa-ТФ- Са+2 IXa- VIIIa- Са+2 Xa- Va- Са+2 Фактор VII в составе инициирующего комплекса частичным протеолизом активирует факторы IX и X других комплексов. Тканевой фактор (ТФ)- это белок, который экспонирован на поверхности многих клеток, например, моноцитов, фибробластов, макрофагов, тромбоцитов. Он не требует протеолитической активации, а фактор VII обладает небольшой эндогенной активностью, но может активироваться частичным протеолизом тромбином. Комплекс Xa- Va- Са+2-мембрана протеолитически активирует протромбин, поэтому называется протромбиназным (рис.4). Рис. 4. Фактор Xa в составе этого комплекса гидролизует две пептидные связи в протромбине и превращает его в тромбин. Тромбин частичным протеолизом превращает фибриноген в фибрин, активирует фактор XIII и по принципу положительной обратной связи факторы V, VIII, VII (рис. 5). Рис.5 Известно, образование тромба могут вызвать повреждение эндотелия сосудов при травме или некоторые сигнальные молекулы, например, тромбоксан, катехоламины, действующие через инозитолфосфатную систему. В наружном слое плазматической мембраны обычно преобладают нейтральные фосфолипиды, а во внутреннем - отрицательно заряженные. Такое распределение липидов поддерживает специфическая транслоказа. Активация ИФ-системы вызывает повышение концентрации Са+2, который ингибирует транслоказу. Вследствие этого во внешнем липидном слое мембраны появляются отрицательно заряженные липиды. При травме внутренняя отрицательно заряженная поверхность мембраны становится доступной для взаимодействия с факторами свертывания крови. Контактная фаза свертывания (внутренний путь) включает проферменты факторы XI и XII, прекалликреин. Они путем частичного протеолиза, превращаются в активные ферменты, которые образуют мембранные комплексы с белком-активатором высокомолекулярным кининогеном (ВКМ) (рис.6). . Рис. 6. Проферменты внутреннего пути не содержат карбоксиглутамат. Вместе с тем фактор XIIа и калликреин являются важными компонентами ряда защитных систем организма: свертывающей, калликреин-кининовой и фибринолитической. Поэтому считают, что контактная активация служит для сопряжения системы гемкоагуляции с другими регуляторными системами организма. В крови и в норме есть клетки с тромбогенными участками, однако кровь остается жидкой. Это обеспечивает антисвертывающая система крови. Она включает антитромбин III и гепарин, плазменные ингибиторы протеаз и антикоагулянтную систему (систему протеинаС). Антитромбин III ингибирует все сериновые протеазы, причем его активатором является гетерополисихарид гепарин (рис.7). Рис.7. Белок α2 - макроглобулин образует комплекс с сериновыми протеазами, в котором они теряют протеолитическую активность. Антикоагулянтная система включает два ферментных комплекса. Первый из них формируется на мембране и содержит фермент тромбин, белок-активатор тромбомодулин, Са+2 . Интегральный белок плазматической мембраны эндотелиальных клеток тромбомодулин повышает сродство тромбина к протеину С. В составе мембранного комплекса тромбин-тромбомодулин-Са+2 тромбин частичным протеолизом активирует протеин С. Активный протеин С (Са) в составе мембранного комплекса Са-S- Са+2 гидролизует белки-активаторы Vа и VIIIа (рис. 8). Это приводит к торможению каскада реакций свертывания крови. Рис.8. Растворение фибринового тромба – фибринолиз- катализирует белок плазмы крови плазмин. Плазмин образуется в результате частичного протеолиза плазминогена под действием калликреина, фактора XIIа или тканевого активатора плазминогена (ТАП), который содержится в клетках эндотелия всех тканей кроме печени. Поступая в кровоток ТАП ингибируется специфическими ингибиторами первого (и-ТАП-1) и второго (и-ТАП-2) типов. Плазмин в кровотоке ингибируют неспецифические ингибиторы сериновых протеаз (рис. 9). Рис. 9. Биохимия печени Печень – самая крупная железа организма. Это своеобразная «биохимическая лаборатория», которая играет важную роль в белковом, углеводном и липидном обменах. В печени синтезируются важнейшие белки плазмы крови: альбумин, фибриноген, протромбин, церулоплазмин, трансферрин, ангиотензиноген и др. Эти белки участвуют в регуляции АД и объёма циркулирующей крови, необходимы для свёртывания крови, метаболизма железа и др. Основные функции печени 1) Обмен углеводов: – Глюконеогенез – Синтез и распад гликогена – Пентозофосфатный путь 2) Обмен липидов и их производных: – – – – – – Синтез ЖК и жиров из углеводов Синтез и выведение холестерола Формирование липопротеинов -окисление жирных к-т Синтез кетоновых тел Синтез желчных кислот 3) Обмен белков – – Синтез белков плазмы крови (включая некоторые факторы свертывания крови) Орнитиновый цикл (обезвреживание NH3) 4) Обмен гормонов: – Метаболизм и выделение стероидных гормонов – Метаболизм полипептидных гормонов 5) Депонирование – – – – гликогена витамина A витамина B12 железа 6) Метаболизм и экскреция билирубина 7) Лекарства и чужеродные вещества – Метаболизм и экскреция Обезвреживающая функция печени. ! Самая важная функция печени: обезвреживание веществ. токсических Ксенобиотики – это чужеродные вещества, попадающие в организм из окружающей среды и не использующиеся для пластических и энергетических целей организма. Они попадают в организм с пищей, через кожу и легкие. Примеры: нефтепродукты, пластмассы, моющие средства, парфюмерия, красители, пестициды и др. Гидрофильные ксенобиотики выводятся из организма в неизменённом виде в основном с мочой. Гидрофобные ксенобиотики могут накапливаться и, взаимодействуя с белками и липидами клеток, нарушать их структуру и функции. Обезвреживание ксенобиотиков происходит во многих тканях, но наиболее активно в печени. Обезвреживание веществ в печени может состоять из одного или двух этапов. Этапы обезвреживания веществ в печени: 1) повышение гидрофильности чужеродных веществ. Включает реакции их гидролиза, окисления, гидроксилирования, восстановления и др. ! Наиболее частая гидрофобного в-ва на гидроксилирование. модификация 1 этапе – 2) конъюгация неизмененных или химически модифицированных на 1 этапе веществ с рядом метаболитов. Если вещество гидрофобно, то его обезвреживание проходит в 2 этапа, если – гидрофильно, то 1 этап может отсутствовать. Некоторые полярные ксенобиотики выводятся из организма, не подвергаясь никаким превращениям. Метаболизм организма: и выведение ксенобиотиков из RH – ксенобиотик; K – радикал, используемый при конъюгации. Примеры стадии: – – – – – модификации ксенобиотиков гидроксилирование окисление по атому S (сульфоокисление) Окислительное дезаминирование Дезалкилирование по N, O, S. Эпоксидирование в 1 1) Система микросомального окисления За 1 стадию обезвреживания отвечает система микросомального окисления (СМО), локализованная в мембранах ЭР. СМО работает практически во всех тканях организма, но наиболее активно в печени. В печени существуют 2 электронтранспортные цепи СМО, которые катализируют гидроксилирование субстратов и являются монооксигеназами. 1 цепь включает: 1) цитохром P450 (гемопротеин), который имеет центры связывания для O2 и гидрофобного субстрата и обладает широкой субстратной специфичностью; 2) фермент NADPH-цитохром P450-редуктазу, содержащий коферменты FAD и FMN; 3) NADPH+H+ – донор ē и Н+ в этой электронтранспортной цепи; 4) O2. 2 цепь включает: 1) цитохром P450; 2) фермент NADH-цитохром b5-редуктазу, коферментом которой является FAD; 3) цитохром b5 – гемопротеин, переносящий ē от NADH-цитохром b5-редуктазы на цитохром P450; 4) NADH + Н+ – донор ē и Н+; 5) O2. Цитохром P450 один атом O2 включает в молекулу субстрата, а 2-й восстанавливает с образованием H2O за счет переноса ē и Н+ от NADPH+H+ при участии цитохром P450-редуктазы (или от NADH+H+ с помощью цитохром b5редуктазы и цитохрома b5). Примеры модификации обезвреживания: ксенобиотиков в 1 стадии Появление в молекулах субстратов полярных групп в результате микросомального окисления повышает гидрофильность веществ и обеспечивает возможность их вступления в реакции конъюгации. Второй этап инактивации – это реакции конъюгации модифицированных на 1 этапе или содержащих полярные группы веществ. Реакции конъюгации – присоединение к гидрофильным группам какого-либо вещества (глюкуроновая кислота, серная кислота, глицин, глутамин, ацетильный остаток и др.). Эти реакции катализируют трансферазы, имеющие широкую субстратную специфичность. Конъюгация снижает реакционную способность веществ и, следовательно, уменьшает их токсичность, повышает гидрофильность и способствует выведению их из организма. ! Донором CH3– группы является SAM, группы –SO3H – активная форма серной кислоты (ФАФС), ацетильной группы – ацетил-КоА, остатка глюкуроновой кислоты – УДФглюкуронат. Примеры реакций конъюгации: 1) Присоединение остатка глюкуроновой к-ты. Ферменты: УДФ-глюкуронилтрансферазы Активная форма: УДФ-глюкуронат: Реакция в общем виде: ROH + УДФ–C6H9O6 RO–C6H9O6 + УДФ 2) Присоединение остатка серной к-ты. Ферменты: сульфотрансферазы Активная форма: ФАФС фосфоаденозинфосфосульфат: Реакция в общем виде: ROH + ФАФ–SO3H RO–SO3H + ФАФ – 3) Глутатионтрансферазы (ГТазы). Для работы ферментов требуется глутатион. Глутатион – трипептид глу-цис-гли. Остаток Глу присоединен к Цис COOH-группой радикала. Глутатион: ГТазы имеются во всех тканях (больше всего: в печени). ! Играют важную роль в обезвреживании собственных метаболитов: некоторых стероидных гормонов, простагландинов, билирубина, желчных кислот, продуктов ПОЛ и др. 3 пути обезвреживания участием ГТазы: ксенобиотиков с 1) конъюгация субстрата R с глутатионом (ГSH) под действием глутатионтрансферазы: R + ГSH ГSRH 2) нуклеофильные замещения: RX + ГSH ГSR + НХ Например, 1-хлор-2,4-динитробензол обезвреживается следующим образом: 3) восстановление органических пероксидов до спиртов под действием глутатионпероксидазы: R–HC–O–OH + 2 ГSH R–HC–OH + ГSSГ + H2O. –ООН – гидроперекисная группа, ГSSГ – окисленный глутатион. Участие P-гликопротеина обезвреживании ксенобиотиков. в P-гликопротеин необходим для выведения из клетки гидрофобных ксенобиотиков. Также он участвует в выведении из клетки лекарственных в-в, использующихся в химиотерапии. P-гликопротеин фосфогликопротеин, который присутствует в мембране многих клеток, особенно почек и кишечника. Содержит 12 трансмембранных доменов и 2 АТФ-связывающих центра. Гидрофобное в-во, проникая выводится из нее с участием АТФ: в клетку С этим механизмом связана 1 из причин множественной лекарственной устойчивости. Примеры обезвреживания токсических в-в в печени: 1) Обезвреживание жизнедеятельности кишечника: продуктов микрофлоры В толстой кишке происходит гниение белков – это образование токсичных продуктов из аминокислот под действием ферментов микроорганизмов кишечника. В основе гниения лежат реакции декарбоксилирования и дезаминирования аминокислот. В ходе гниения, из тирозина образуются фенол и крезол, из триптофана – скатол и индол. Эти вещества всасываются клетками кишечника, транспортируются кровью по воротной вене в печень, где они конъюгируют с серной или глюкуроновой кислотой. Если в организм попадает бензол, то его обезвреживание происходит в 2 этапа. Сначала он гидроксилируется с помощью СМО и превращается в фенол, а потом вступает в реакции конъюгации с ФАФС или УДФглюкуронатом. Реакции обезвреживания фенола, крезола и бензола: Реакции обезвреживания индола и скатола: Биотрансформация лекарств в печени Лекарства либо выводятся из организма в неизмененном виде, либо подвергаются химической модификации. В результате биотрансформации лекарственных веществ может произойти: 1) инактивация лекарственных в-в, т.е. снижение их фармакологической активности (фенобарбитал, нитриты, эфедрин и др.) 2) повышение активности лекарственных в-в (ловастатин, метилдофа, норморфин и др.) 3) образование токсических метаболитов (фенацетин, сульфаниламиды) Инактивация лекарственных в-в (как ксенобиотиков) происходит в 1 или в 2 фазы. и Примеры инактивации лекарственных в-в: Аспирин (ацетилсалициловая кислота) сначала гидролизуется и превращается в салициловую кислоту, которая конъюгирует с УДФ-глюкуронатом: Сульфаниламиды (альбуцид, сульгин, сульфаметоксазол) инактивируются в результате реакций ацетилирования: Реакции инактивации аспирина: Инактивация сульфаниламидов: Пример повышения активности лекарственных в-в: Превращение имипрамина в более активное в-во – дезметилимипрамин: Дозы некоторых лекарств при частом приеме необходимо увеличивать, так как их действие на организм ослабляется. Это происходит потому, что эти лекарства, как и другие чужеродные соединения, индуцируют синтез ферментов СМО и реакций конъюгации. На этой особенности основано привыкание к лекарствам и некоторым ядам. ("Эффект Митридата") Аналогичное действие вызывает и этанол. Поэтому, в состоянии алкогольного опьянения существенно снижается эффективность многих лекарственных в-в. Химический канцерогенез В основе химического канцерогенеза – повреждения ДНК под действием химических канцерогенов. ! Химические канцерогены способствуют превращению протоонкогена в онкоген. Протоонкоген – ген, содержащий информацию о белке, регулирующем нормальную пролиферацию клеток, и способный в результате изменения структуры превращаться в онкоген. Известно несколько сотен протоонкогенов, мутации в которых могут привести к превращению нормальной клетки в опухолевую. Онкоген – ген, экспрессия которого приводит к неконтролируемой пролиферации клеток. Для превращения протоонгкогена в онкоген требуются определенные изменения в его регуляторной или структурной части. Многие в-ва, попадающие в организм, являются проканцерогенами. Они превращаются в канцерогены в ходе «обезвреживания» в печени. Некоторые химические проканцерогены: Класс соединений Представители Полициклические Бензантрацен, углеводороды метилхолантрен, диметилбензантрацен Источники Выхлопные газы, продукты горения, сигаретный дым, коксохимическое производство Производство красителей Ароматические амины Метиламинобензол, нафтиламин Диоксины Тетрахлорбензодиоксин Производство дефолиантов и ростовых веществ, целлюлозно-бумажная промышленность, хлорирование воды, горящие свалки Афлатоксин В1 Плесневые грибы Микотоксины Нитрозамины Диметилнитрозоамин, диэтилнитрозоамин Образуются при употреблении нитратсодержащих продуктов Примеры канцерогенных в-в: 1) Ароматические амины. Используются в производстве анилиновых красителей и резиновой промышленности. Активный канцероген вызывает рак мочевого пузыря. 2-амино-1-нафтол 2) Полициклические углеводороды (ПУ). Яркий представитель - бензантрацен. Содержится в выхлопных газах и сигаретном дыме. Активный канцероген: эпоксид бензантрацена. Может вызвать рак легких, печени и желудка. Эпоксиды обладают высокой химической активностью. Они могут участвовать в реакциях неферментативного алкилирования ДНК, РНК и белков, что может способствовать перерождению нормальной клетки в опухолевую. 3) Микотоксины. Типичные представители: Афлатоксины – метаболиты некоторых видов плесени. Образуются при неправильном хранении зерна, круп, орехов. Яркий представитель – Афлатоксин B1: Активный канцероген Эпоксид Афлатоксина B1 вызывает рак печени. 4) Канцерогены неорганической природы: Нитраты в высокой концентрации. Нитраты и окислители: особенно нитриты – сильные Они могут повреждать ДНК, окислять многие гемопротеины (Hb, цитохромы и т.п.). В организме из HNO2 и вторичных аминов образуются нитрозамины R2N–N=O, которые являются сильными мутагенами, вызывающими алкилирование азотистых оснований ДНК.