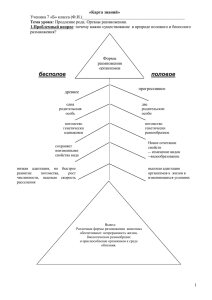
биология размножения и развития
advertisement

Министерство образования и науки Российской Федерации Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования «Иркутский государственный университет» А. В. Болотов БИОЛОГИЯ РАЗМНОЖЕНИЯ И РАЗВИТИЯ Раздел. БИОЛОГИЯ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ Учебное пособие УДК 591.3(075.8) ББК 28.63я73 Б79 Печатается по решению ученого совета биолого-почвенного факультета ИГУ Рецензенты: канд. мед. наук А. А. Бочкарёв (Иркут. филиал ФГОУ ВПО «РГУФКСМиТ») канд. биол. наук Н. Ю. Копылова (ФГОУ ВПО «ИГУ») Б79 Болотов А. В. Биология размножения и развития. Раздел. Биология индивидуального развития : учеб. пособие / А. В. Болотов. – Иркутск : Изд-во ИГУ, 2011. – 217 с. Рассмотрены с учетом новейших данных основные разделы биологии развития: развитие половых клеток, оплодотворение, дробление, процессы гаструляции и нейруляции, основные закономерности органогенезов, дифференцировка клеток и процессы роста. Рассмотрены процессы размножения разных организмов как растений, так и животных. Описаны процессы клеточного и молекулярного механизмов развития. Отдельные главы посвящены краткому изложению: половому созреванию, роли генов в процессах дифференцировки и развития и онтогенезу человека. Для студентов биологических специальностей вузов бакалавров по направлению 020400.62 «Биология» (профиль «ботаника», профиль «зоология беспозвоночных», профиль «зоология позвоночных», профиль «физиология», профиль «микробиология», профиль «биоинженерия и биоинформатика», профиль «биохимия», профиль «физико-химическая биология», профиль «микробиологическая биотехнология»), специалистов 020201.65 «Биология», специалистов 020205.65 «Физиология», специалистов 020209.65 «Микробиология», магистров по направлению 020400.68 «Биология» профиль «психофизиология», магистров по направлению 022000.68 «Экология» профиль «медицинская экология». Библиогр. 24 назв. Табл. Ил. 61. УДК 591.3(075.8) ББК 28.63я73 © Болотов А. В., 2011 © ФГБОУ ВПО «ИГУ», 2011 3 Оглавление Введение .............................................................................................. 5 МЕТОДЫ ЭМБРИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ .............. 9 ИСТОРИЯ РАЗВИТИЯ ЭМБРИОЛОГИИ .................................... 10 Развитие эмбриологии в Античную эпоху ........................... 10 Развитие эмбриологии после Античной эпохи .................... 12 Развитие эмбриологии в XVIII–XIX веках ........................... 16 Эмбриология в XX веке и до наших дней............................. 22 I. РАЗМНОЖЕНИЕ ОРГАНИЗМОВ.............................................. 29 1. Бесполое размножение ....................................................... 30 2. Половое размножение у покрытосеменных растений ..... 32 3. Половое размножение животных ...................................... 39 II. ПОЛОВОЕ СОЗРЕВАНИЕ ......................................................... 43 III. ПОЛОВАЯ СИСТЕМА МЛЕКОПИТАЮЩИХ ..................... 46 1. Мужская половая система .................................................. 46 2. Женская половая система ................................................... 56 IV. ОСНОВНЫЕ ЭТАПЫ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ ... 67 V. ГАМЕТОГЕНЕЗ .......................................................................... 69 1. Происхождение первичных половых клеток .................... 70 2. Состав, развитие и особенности половых клеток ............ 72 3. Строение половых клеток................................................... 74 4. Развитие половых клеток ................................................... 87 5. Профаза мейоза ................................................................... 88 6. Оогенез ................................................................................. 90 7. Гормональная регуляция половой функции у женщин. Овуляция ................................................................................ 100 8. Сперматогенез ................................................................... 107 9. Нарушения сперматогенеза .............................................. 112 10. Отличия сперматогенеза и овогенеза ............................ 118 11. Эволюция гаметогенеза .................................................. 119 VI. ОПЛОДОТВОРЕНИЕ.............................................................. 120 1. Дистантное взаимодействие и сближение гамет ............ 122 2. Контактное взаимодействие и активизация яйцеклетки .. 123 3. Сингамия ............................................................................ 129 4 Болотов А. В. Биология размножения и развития 4. Особенности репродукции человека ............................... 130 5. Детерминация пола. Влияние факторов среды на развитие признаков пола .................................................. 130 VII. ДРОБЛЕНИЕ ........................................................................... 133 1. Радиальное голобластическое дробление ....................... 137 2. Спиральное голобластическое дробление ....................... 140 3. Билатеральное голобластическое дробление .................. 141 4. Асинхронное, ротационное голобластическое дробление ............................................................................... 142 5. Дискоидальное меробластическое дробление ................ 145 6. Поверхностное меробластическое дробление ................ 147 VIII. ГАСТРУЛЯЦИЯ .................................................................... 149 Образование мезодермы ....................................................... 153 IX. ОСОБЕННОСТИ РАЗВИТИЯ МЛЕКОПИТАЮЩИХ ........ 156 X. ПРОВИЗОРНЫЕ ОРГАНЫ (ВНЕЗАРОДЫШЕВЫЕ) ........... 159 XI. ОРГАНОГЕНЕЗ ........................................................................ 164 1. Развитие производных эктодермы ................................... 164 2. Развитие производных энтодермы ................................... 175 3. Развитие производных мезодермы................................... 178 XII. РОСТ, РАЗВИТИЕ, ДИФФЕРЕНЦИРОВКА, МОРФОГЕНЕЗ, РОЛЬ ГЕНОВ ..................................................... 188 1. Гомеозисные гены ............................................................. 191 2. Реализация программ морфогенеза.................................. 195 XIII. ОНТОГЕНЕЗ .......................................................................... 199 1. Особенности, свойства и виды онтогенеза...................... 199 2. Онтогенез человека ........................................................... 204 3. Основные закономерности роста и развития .................. 211 Вопросы для аттестации по предмету ....................................... 212 Список использованной литературы ........................................... 214 Рекомендуемая литература .......................................................... 216 ВВЕДЕНИЕ Биология развития – это, вероятно, наиболее широкая из всех биологических дисциплин. Биология индивидуального развития – это область знаний, формирующаяся путём синтеза ряда биологических дисциплин – эмбриологии, генетики, цитологии, биохимии, молекулярной биологии, биофизики, физиологии. Исследователь, работающий в этой области, не ограничен какимлибо иерархическим уровнем организации. В своих исследованиях он не ограничен также какой-либо конкретной группой организмов или системой органов в организме. Можно сказать, что биология развития включает и интегрирует их все. Единственный путь возникновения всего лежит через развитие, и биология развития – ни что иное, как изучение каждой молекулы в клетке, клетки, ткани, органа и организма как функций времени. Теперь даже эволюцию рассматривают как функцию развития. Эволюция осуществляется путем наследственных изменений, происходящих в зародыше, и изменения, которые создали современную лошадь из ее пятипалого предка – это изменения, происходившие в зародыше. Таким образом, биология развития является наукой, охватывающей наибольший спектр биологических проблем. Это также одна из самых своеобразных наук. Биология развития – наука о становлении, а не о существовании. Она свергает гегемонию взрослого организма. Взрослая особь – только конечная стадия длинного ряда взаимодействий, создавших ее. Для исследователя в области биологии развития процесс возникновения намного важнее и интереснее, чем процесс поддержания созданной формы. Биология развития является также областью (если можно назвать областью то, что объясняет всё в живой природе), в которой постановка проблем имеет такое же большое значение, как и их решение. Нами исследована только незначительная часть процессов развития, и объяснять механизмы развития на основе этих знаний примерно то же, что описывать жизнь океана, изучая оставшиеся после отлива лужи. Большую часть процесса развития еще предстоит познать. На протяжении последних сорока лет биология развития накапливала новые, всё более утонченные методы исследования. В 6 Болотов А. В. Биология размножения и развития частности, объединение эмбриологии с молекулярной биологией и иммунологией дало в руки ученых могущественные орудия. Моноклональные антитела, меченные флуоресцентными соединениями, являются красителями, которые способны различать в пептиде индивидуальные аминокислоты. Радиоактивная рекомбинантная ДНК может обнаружить в популяции клеток одну единственную молекулу мРНК. Эти новые методы используются теперь для того, чтобы ответить на вопросы о процессах индукции и дифференцировки, которые эмбриологи-экспериментаторы задавали шестьдесят лет тому назад. Но это только начало революции, включающей соединение генетики развития с эволюционной биологией для создания новой теории эволюции, которая могла бы объяснить макроэволюционные изменения. Биология развития, или онтогенетика – раздел биологии, всесторонне изучающий процессы и движущие силы индивидуального, или онтогенетического развития организма, включающий период развития половых клеток, от оплодотворения и эмбрионального развития до рождения и смерти. Науку о закономерностях индивидуального развития стали называть «биологией развития». Биология индивидуального развития – область науки, изучающая закономерности онтогенетического развития организмов. Предметом изучения биологии индивидуального развития (животных организмов) являются: 1) предзародышевый период развития – формирование половых клеток; 2) зародышевый период развития – от оплодотворения яйцеклетки до выхода организма из яйцевых оболочек; 3) постэмбриональный период – развитие после рождения, метаморфоз личинок и т. д. В целом можно сказать, что предметом биологии индивидуального развития могут быть различные аспекты изучения морфогенетических процессов (морфологические, физиологические, биохимические, биофизические, генетические, экологические, филогенетические и др.). Так же в области рассмотрения современной биологии индивидуального развития находятся такие проблемы, как: регенерация, бесполое и вегетативное размножение, уродливые патологические формообразования. Введение 7 Биология индивидуального развития исследует как общие закономерности индивидуального развития, так и частные особенности процесса онтогенеза. Онтогенез – процесс индивидуального развития организма, проходящий весь жизненный цикл (от зиготы до смерти). В ходе онтогенеза выделяют определенные фазы: 1) эмбриогенез; 2) созревание; 3) взрослое состояние; 4) старение. Содержание же понятия «биология развития» в значительной степени определяется молекулярно-генетическими аспектами изучения индивидуального развития, что недостаточно для причинного анализа любого из перечисленных ранее морфогенетических процессов. Широкое использование термина «биология развития» – это дань уважения успехам в области молекулярной генетики, биохимии и цитологии, но вряд ли из-за этого надо отказываться от устоявшегося названия «эмбриология». Название «эмбриология» происходит от греческих слов (embrion – зародыш и logos – учение) – учение о зародыше, буквально. Но эмбриология изучает не только зародышевое развитие (пренатальный период), но и предзародышевый период (гаметогенез) и период после рождения или постэмбриональный период. Выделяют несколько направлений в изучении биологии развития: 1) общее вопросы развития; 2) частные вопросы развития; 3) развитие в сравнении; 4) развитие с эволюционной точки зрения; 5) использование экспериментальных методов для исследования процессов развития. При изучении регуляторных механизмов ведущее звено принадлежит генетическому подходу, который приобрел повышение статуса, и появилось направление исследования – генетика развития, которая изучает реализацию наследственной информации в ходе индивидуального развития. 8 Болотов А. В. Биология размножения и развития Таким образом, биология развития на современном этапе наметила три направления дальнейших исследований: Во-первых, изучение механизмов регуляции функций генов. Во-вторых, каким образом вновь синтезированные белки определяют форму, функцию и поведение клеток. Это изучение микротрубочек, клеточных мембран. В-третьих, и наиболее сложное направления – это становление формы органов и организма. Таким образом, биология развития – одно из магистральных направлений современной науки. Вобрав в себя концептуальные и методические достижения многих биологических наук. Сейчас она все более активно участвует в исследовании функции генома на ранних стадиях онтогенеза, анализе и контролирующих механизмов нормального и патологического развития и многих других. Значительны ее практические успехи. Так, уже решена проблема глубокого замораживания и длительной криоконсервации гамет и ранних зародышей, разработаны научные основы получения животных и растений с заданными наследственными свойствами, быстро совершенствуются методы ранней дородовой диагностики наследственных болезней, значительно повысилась эффективность лечения различных форм бесплодия. Всё это – итог стремительного прогресса возможностей. Именно эта отрасль биологии сегодня может служить примером положения, что прогресс науки связан с прогрессом или развитием ее методов. МЕТОДЫ ЭМБРИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ Методы эмбриологических исследований очень разнообразны и включают в себя различные методы физики и химии, биологии, гистологии, цитологии, эмбриологии, биохимии, физиологии, генетики, молекулярной биологии. При морфологических исследованиях пользуются всевозможными видами световой микроскопии и электронной микроскопией. Визуальное наблюдение развития зародышей фиксируемое микрофотосъемкой или видеосъемкой. Особенно важны методы прижизненного наблюдения, в частности – прослеживание перемещений эмбрионального материала (морфогенетических движений) при помощи меток, наносимых на зародыш прижизненными красителями, а также методы гистохимии, применение радиоактивных изотопов, антител к определенным белкам развивающегося зародыша, меченные флуоресцином. В основе экспериментальных методов лежит удаление и трансплантация различных частей зародыша. Начиная с 50-х гг. XX в., преимущественное значение приобрели биохимические методы. Методы трансплантологии, выращивание культуры тканей, использование стволовых клеток, методы ЭКО (экстракорпоральное оплодотворение). ИСТОРИЯ РАЗВИТИЯ ЭМБРИОЛОГИИ Развитие эмбриологии в Античную эпоху Биология индивидуального развития – это область, формирующаяся в последние десятилетия путем синтеза ряда биологических дисциплин – эмбриологии, генетики, цитологии, биохимии и физико-химических направлений биологии (молекулярной биологии, биоорганической химии, биофизики). А основу биологии индивидуального развития составляет – эмбриология – одна из наиболее древних наук. И на протяжении длительной истории эта наука, олицетворяя глубокий интерес человека к тайнам зарождения живых существ, впитывала научно-технические и идейно-философские достижения, способствовала формированию научной идеологии. Вопросы зарождения и развития новых особей были предметом многочисленных легенд и мифов. Не будем останавливаться на моментах предыстории и обратимся к Древней Греции – стране, где впервые в истории человечества началась осознанная работа над теоретико-философскими основами наук. Первые регулярные знания в области эмбриологии связывают с именем Гиппократа (460–377 гг. до н. э.). Эмбриологические знания ученого связаны главным образом с акушерством и гинекологией. Самые важные для истории эмбриологии книги Гиппократа – «О диете», «О семени», «О природе ребенка». Гиппократ выдвинул двухсеменную теорию развития через смешивание мужского и женского семени. В его сборниках содержатся первые научные высказывания о развитии эмбрионов и факторах формообразования. В них он высказывает идеи, согласно которым каждому телу присущи три первичных свойства – тепло, сухость и влажность. Данные свойства никогда не встречаются раздельно. При этом каждое тело способно привлекать себе подобное. Жизнь заключается в одновременном высушивании влаги огнем, в увлажнении огня влагой, в поглощении огнем пищи, поступающей в тело, и поэтому требуется новая пища. Процессы, происходящие в организме, Гиппократ сравнивает с процессами в неорганических телах – небесных и земных, а также с трудовой деятельностью. Гиппократ предвосхитил идею преформации: «Все части зародыша 11 История развития эмбриологии образуются в одно и то же время. Все члены отделяются друг от друга одновременно и таким же образом растут. Ни один не возникает раньше или позже другого, но те, которые по природе своей толще, появляются прежде тонких, не будучи сформированы раньше». Гиппократ Аристотель Он заложил принципы, которые позже стали характерны для концепции развития – преформизма (от греч. prae – впереди и forme – формы). Основная черта преформационных концепций – признание существования изначальных различий между частями зародыша и мнение, что «отделение» частей (дифференцировка) происходит лишь в некоторый начальный момент развития, а в дальнейшем разделившиеся части только растут. Во многом своеобразными были взгляды одного из величайших мыслителей древности, основоположника естественных наук – Аристотеля (384–322 до н. э.). Эмбриологические факты, которыми располагал Аристотель, и его воззрения на развитие жизни подробно изложены в сочинении «О возникновении животных». Он имел немалые сведения по анатомии и физиологии организмов. Однако огромное влияние Аристотеля на последующую науку связано не столько с его фактическими наблюдениями, сколько с теоретико-философскими представлениями. Аристотель счел недостаточными для объяснения развития те, 12 Болотов А. В. Биология размножения и развития по современной терминологии, «механистические» причины, к которым сводил развитие Гиппократ. Для объяснения и познания сущего Аристотель выдвинул четыре принципа – четыре причины: материальную («неоформленный» материал – causa materialis) причину; действующую причину (своего рода триггер предстоящих событий – causa efficiens); активное начало, обладающее энергией, способное формировать пассивную материю (формальная причина – causa formalis); финальную причину (causa finalis) – причину всех причин. Последняя есть «цель» данного процесса, та форма, к которой этот процесс стремится. В противоположность Гиппократу, Аристотель считал и аргументировал конкретными примерами то, что органы возникают не все сразу, а постепенно, один вслед за другим из бесструктурной вначале массы. Такое представление сделало Аристотеля основателем эпигенеза (от греч. epi – после и genesis – происхождение) – противоположного преформизму учения о постепенном развитии, связанном с усложнением организации. Развитие эмбриологии после Античной эпохи Первые после античной эпохи систематические наблюдения за развитием зародыша цыпленка относятся, по-видимому, к концу XVI в. и принадлежат итальянскому натуралисту Улиссе Альдрованди (1522–1605). Вслед за ним еще более подробные описания дали его ученик Волхер Койтер (1534–1576) и Джироламо Фабриций (1533– 1619). Фабриций описал и изобразил зародышей многих позвоночных, но его работы имели анатомический характер и не давали представлений о последовательном ходе развития. Койтер в 1572 г. первый дал научное описание развития куриного зародыша, что позволяет говорить о нем как об основоположнике эмбриологии. Огромное значение для развития эмбриологии, как и всей биологии, сыграло появление первых простых микроскопов. И первым из них был голландский очковый мастер Захариус Йансен (1580–1638) в 1595 г. Его микроскоп увеличивал всего в 5 раз. Но это была лишь начальная ступенька на этом пути. 13 История развития эмбриологии Улиссе Альдрованди Волхер Койтер Джироламо Фабриций Первые сложные микроскопы были созданы в начале XVII в. голландским изобретателем Корнелием Дребелем (1572–1633) в 1621 г. Этот микроскоп стал прототипом всех последующих микроскопов. Позднее, в 1663 году микроскоп Дребеля усовершенствовал английский физик Роберт Гук (1635–1703) (появился тубус, определенного размера, конденсор). Захариус Йансен Корнелий Дребель Роберт Гук С необозримым миром микроскопических объектов, открывшимся исследователям, знакомились бессистемно. Из числа первых исследователей, применивших микроскоп, были англичанин Р. Гук (срез пробки – клетка, 1665 г.), голландцы Антонио ван Левенгук (1632–1723) и Ян Сваммердам (1637–1680) и итальянец Марчелло Мальпиги (1628–1694). Одно из важнейших открытий эмбриологии – обнаружение Левенгуком сперматозоидов в 1677 г.; он также изучал партеногенез у тлей. 14 Болотов А. В. Биология размножения и развития Антонио Левенгук Ян Сваммердам Марчелло Мальпиги Сваммердаму принадлежат первые работы по метаморфозу насекомых, а М. Мальпиги – по многим вопросам микроскопической анатомии, а также органогенезу зародыша курицы. Быстрое накопление фактического материала оживило и теоретические аспекты эмбриологии; возникло стремление осмыслить полученные факты. И в 1651 г. Вильям Гарвей (1578–1657) в книге «О произрождении животных» впервые после Аристотеля дополнил знания по эмбриологии систематическими данными о развитии куриного яйца, т. е. фактически по оогенезу, а также о развитии яйцевых оболочек. При этом ученого ставили в тупик такие наблюдения: почему теплота способствует развитию цыпленка из хорошего яйца, а из плохого яйца под действием теплоты развитие становится хуже? Будучи придворным медиком, Гарвей получил разрешение на вскрытие животных (косули, лани), добытых на охоте. Таким образом, он имел возможность изучать внутриутробное развитие млекопитающих и анатомию животных. В. Гарвей стал автором термина «эпигенез». Он открыл замкнутые круги кровообращения и выдвинул в своей книге знаменитый принцип «Все (живое) из яйца». С этого времени и на протяжении более двухсот лет теоретическая работа в эмбриологии концентрировалась вокруг борь- История развития эмбриологии 15 бы двух зародившихся еще в античные времена основных идейных течений – преформизма и эпигенеза. Но, несмотря на огромный по тому времени технический прогресс, предпочтение в объяснении развития отдавалось не эпигенезу, а преформизму. Действительно, основной тезис древних эпигенетиков о том, что наиболее ранние зачатки зародыша бесструктурны, легко опровергался данными микроскопии, которые свидетельствовали о существовании микроструктуры в ранних зачатках. В особенно яркой форме детальные микроструктуры, предвосхищающие организацию взрослых особей, обнаруживались в личинках и куколках насекомых, в бутонах растений. Сторонником мнения, что все детали строения предшествуют и распределены в сперматозоиде, стали называть преформистами – анималькулистами. Были и преформисты-овисты, считавшие, что организация будущей особи полностью представлена в яйце. Поводом к этому послужило открытие партеногенеза. Преформизм XVII в. – это учение, возникшее в основном благодаря необоснованной экстраполяции только что полученных микроскопических данных и утверждающее, что все детали строения будущего организма предшествуют и предраспределены с самого начала развития в том же пространственном порядке, в каком они расположены у взрослого животного. Преформисты не допускали новообразование частей, а лишь их рост. Если быть последовательным преформистом, необходимо допустить, что в яйце или сперматозоиде заготовлена структура организма не только ближайшего, но и всех последующих (потенциально бесконечного числа) поколений. Но такое допущение не казалось преформистам абсурдным. Оно было высказано в виде широко распространенной «гипотезы вложения», согласно которой тела потомков действительно вложены друг в друга, как матрешки. Некоторые только что открытые явления, например личиночное размножение у тлей (в теле личинки обнаруживали зародыша будущего поколения), рассматривались как прямое подтверждение этой гипотезы. Сторонником гипотезы вложения был, в частности, немецкий философ и математик Готфрид Вильгельм фон Лейбниц (1646–1716). 16 Болотов А. В. Биология размножения и развития Развитие эмбриологии в XVIII–XIX веках Решительный поворот в эмбриологии был осуществлен в 1759 г. петербургским академиком Каспаром Фридрихом Вольфом (1734–1794). В этом году Каспар Вольф Альбрехт Галлер Xристиан Пандер Вольф представил свою диссертацию под названием «Теория зарождения». В те времена господствовало мнение швейцарского учёного физиолога и анатома Альбрехт фон Галлер (1708– 1777) о том, что трубчатые и мешкоподобные структуры зародыша (кишечник, например) с самого начала имеют такую же форму, но это трудно заметить из-за тонкости стенок и их плотного слипания. Позже происходит их простое раздувание. Такое толкование строго соответствовало преформационной теории. Вольф установил совершенно иное. Кишечник, а также зачаток нервной системы сначала представляет собой пласты, которые лишь позже скручиваются в трубки. В ходе развития образуются новые формы. По сути дела, Вольф открыл формообразование и тем самым дал первый позитивный и неопровержимый аргумент в пользу эпигенеза. Судьба этих открытий была трудной. Под давлением господствующих авторитетов выводы Вольфа отвергались, и его работы были на некоторое время забыты. Еще при жизни Вольфа с весьма остроумными доводами в пользу эпигенеза выступил немецкий профессор Иоганн Фридрих Блюменбах (1752–1840). Он впервые указал на несовместимость с преформизмом всевозможных случайных новообразова- 17 История развития эмбриологии ний (например, галлы у растений) или регенерации гидры из любого, произвольно выбранного участка тела, т. е. он обнаружил регуляции формы организма, не связанные с его ростом. Так, целая гидра восстанавливается из своей продольной половинки простым схождением краев разреза, тогда как, по убеждению преформистов, такой процесс должен был быть обязательно связан с ростом. Таким образом, для Блюменбаха, как и для Вольфа, одним из основных аргументов против преформизма было обнаружение «чистого», не связанного с ростом формообразования. Но, несмотря на эти единичные догадки, уровень естественных наук не позволял еще обрести им прочную основу. Иоганн Блюменбах Мартин Ратке Карл Бэр Дальнейший прогресс в эмбриологии позвоночных связан с именами Мартина Генриха Ратке (1814–1860), Xристиана Пандера (1794–1865), и Карла Эрнста фон Бэра (1792–1876). Пандер в 1817 г. впервые описал зародышевые листки. Он нашел, что зародыш цыпленка на определенной стадии состоит из трех пластов: наружного – серозного, самого глубокого – слизистого, и промежуточного – кровяного. К. Бэр распространил этот структурный принцип на всех позвоночных, обнаружив такие же листки в развитии рыб, лягушки и черепахи. Однако у зародышей птиц он насчитывал четыре листка, считая за отдельный листок каждый из двух слоев, на которые в ходе развития расслаивается мезодерма. Таким образом, Бэр установил единство плана строения зародышей разных классов позвоночных. Это привело его к важнейшему обобщению – «закону зародышевого сходст- 18 Болотов А. В. Биология размножения и развития ва». Бэр утверждал, что зародыши разных видов, относящихся к одному типу, более сходны между собой, нежели взрослые формы, и что их видовые различия в ходе развития постепенно нарастают. Иными словами, сначала в развитии проявляются черты типа, потом класса и т. д. Бэр – автор многих важнейших открытий. Он впервые правильно описал яйцо млекопитающих и человека (1827 г.) и хорду зародышей позвоночных. В споре преформистов с эпигенетиками Бэр занимал осторожную промежуточную позицию. Всецело соглашаясь с фактическими выводами Вольфа, он выступал против утверждений о полной «бесструктурности» ранних закладок. Бэр подчеркивал преемственность каждого этапа развития – от более простого к более сложному. По его словам, развитие есть не предобразование, не новообразование, а преобразование. Такая точка зрения полностью подтверждена последующим ходом развития науки. Следующий важнейший идейный перелом в эмбриологии, как и вообще в биологии, связан с выходом в свет в 1859 г. «Происхождения видов» Ч. Дарвина. Дарвинизм прежде всего подрывал главную опору телеологического мировоззрения, указывая на относительность органической целесообразности и на возможность достижения ее методом «проб и ошибок» (теория естественного отбора). Именно это произвело наибольшее впечатление на современников. Но не только этим своим аспектом дарвинизм повлиял на развитие эмбриологии. Наряду с палеонтологией и сравнительной анатомией Дарвин обращался к эмбриологии в поисках подтверждения своей эволюционной теории. Дарвин предлагал эволюционное истолкование закона Бэра. В более категорической форме это же положение было выражено в биогенетическом законе Эрнста Геккеля (1834–1919): «онтогенез есть краткое повторение филогенеза». 19 История развития эмбриологии Эрнст Геккель А. О. Ковалевский И. И. Мечников Гипотеза Дарвина оказалась мощным стимулом к эмбриологическим исследованиям. На основе эволюционной теории ученые разных стран за считанные годы выяснили развитие обширных, раннее совершенно не изученных групп организмов. Среди них первыми были русские эмбриологи Александр Онуфриевич Ковалевский (1840–1901) и Иван Ильич Мечников (1845–1916). Особое значение имели работы Ковалевского по развитию ланцетника и асцидий, в которых продемонстрированы сходные черты в развитии позвоночных и беспозвоночных животных. Эти исследования способствовали укреплению взглядов на эволюцию как на сквозной монофилетический процесс. А. О. Ковалевский – один из основоположников теории зародышевых листков (эктодерма, мезодерма, энтодерма). Он обнаружил эти зародышевые листки у всех типов животных. Термины предложил Геккель. Таким образом, Ковалевского можно считать основателем современной эмбриологии. В 70–80 гг. XIX в. зародилось новое направление эмбриологии, явившееся до некоторой степени реакцией на господство филогенетических принципов. В противоположность Геккелю его создатели подчеркивали необходимость изучения непосредственных причин развития с применением специальных экспериментов. Началось формирование аналитической и экспериментальной эмбриологии, внесшей наибольший вклад в изучение факторов и механизмов индивидуального развития. Основоположниками этого направления были немецкие ученые Вильгельм Гис (1831–1904) и Вильгельм Ру (1850–1924). 20 Болотов А. В. Биология размножения и развития Вильгельм Гис Вильгельм Ру Зарождение аналитического направления в эмбриологии связано с деятельностью В. Гиса, анатома и эмбриолога, который первым стал внедрять в эмбриологию методы химии и физики. Неоценимое значение этих фундаментальных для биологии наук полностью раскрылось в наше время. Так, Гис указывал на важность выяснения механических сил, вызывающих изменения формы развивающегося зародыша, т. е. процессов морфогенеза. Он стремился исследовать самые ранние стадии зародышевого развития (морфогенез которых, как известно сейчас, происходит на уровне макромолекул) и выявить их роль в возникновении зачатков и органов на последующих стадиях. Тем самым В. Гис создал основы аналитической эмбриологии, одна из задач которой – целенаправленный анализ ранних стадий развития органов и тканей, выявление их еще незримых, неоформившихся зачатков. В. Гис считал, что невидимые предшественники будущих органов локализованы в еще недифференцированном зародыше и даже в яйце не беспорядочно, а пространственно упорядочено, так, что их можно картировать и представляет собой как бы мозаику из различно детерминированных бластомеров, неспособных изменять направление своего развития (мозаичная теория). Следующим этапом в развитии новых тенденций в эмбриологии был казуально-аналитический подход. Его создатель В. Ру – крупнейший экспериментатор, оказавший большое влияние на развитие цитологии, генетики и особенно эмбриологии. В. Ру считал, что для раскрытия механизмов развития недостаточно История развития эмбриологии 21 чисто аналитического описания нормального развития, а необходимы эксперименты, для того чтобы установить причинноследственные взаимоотношения между частями, выявить факторы, которые определяют, детерминируют пути развития частей зародыша и их дифференцировку. Таким образом, в основу своей методологии Ру положил эксперимент, а главной задачей провозгласил поиск и анализ причинных факторов, определяющих развитие. Он считал, что факторы, детерминирующие зачаток, могут быть как внутри, так и вне его, и в зависимости от этого механизм их действия неодинаков и стал основателем «механики развития организмов». Укреплению эпигенетического принципа развития в большой мере способствовали эксперименты Ганса Дриша (1867– 1941), который идеологически занимал прямо противоположную позицию. В одном из экспериментов Дриш воспроизвел опыт Ру, но использовал иной технический прием: он отделил друг от друга два бластомера морского ежа, показав, что каждый из них способен развиваться в полноценный организм. В последующем аналогичные результаты были получены на множестве других объектов. Способность части зародыша развиваться в целостную особь, т. е. компенсировать отсутствующую часть, Дриш назвал эмбриональной регуляцией. Открытие эмбриональной регуляции было событием величайшего значения, но сильно затруднило дальнейшее применение казуально аналитического подхода. С позиций сегодняшнего дня можно сказать, что оба подхода имели огромное значение и в принципе не противоречили друг другу. Историческая заслуга Дриша состоит в том, что он проанализировал открытую им эмбриональную регуляцию развивающегося зародыша и сформулировал известный, носящий его имя закон о том, что путь развития части зародыша есть функция положения этой части относительно целого. 22 Болотов А. В. Биология размножения и развития Эмбриология в XX веке и до наших дней Направление, намеченное Ру в экспериментальной эмбриологии, наибольшее воплощение нашло в работах школы Ганса Шпемана (1869–1941). Уже в начале века, в 1901 г., Шпеман в эксперименте по разделению раннего зародыша тритона получил результат, подтверждающий одновременно и «мозаичную» теорию Гиса) и концепцию эмбриональных регуляций Дриша. Оказалось, что дальнейшее развитие разделенных частей зависит от того, в какой плоскости зародыш расчленяется. Шпеман экспериментально продемонстрировал, что именно в процессе взаимодействия частей зародыша в 1924 г. было обнаружено явление эмбриональной индукции. Это было одним из крупнейших открытий в биологии первой половины XX в. Как и в первых экспериментах с разделением зародыша, явление индукции показывало, что детерминация и регуляция два взаимодополняющих фундаментальных принципа индивидуального развития целостной живой системы. Выяснилось, что процесс индивидуального развития зависит от множества переплетающихся внутренних и внешних факторов. В разработке такого понимания процессов развития, при котором одна и та же система факторов развития зародыша определяет и ее целостность, и ее дифференциацию на части, важная заслуга принадлежит советскому биологу Александру Гавриловичу Гурвичу (1874–1954). Он искал те факторы, действием которых можно было бы объяснить интегрированность процессов развития, их пространственную организацию. Гурвич начинал свою научную деятельность под сильным влиянием идей Дарвина, но не разделял его взглядов на непознаваемость целостных факторов развития. Он описывал эти факторы и их отношения с частями организма в точных математических выражениях, что для того времени было новым и непривычным. Гурвич первый ввел статистические методы в эмбриологию и обнару- 23 История развития эмбриологии жил явление «нормировки» клеточных делений в целом организме, а также участие случайных событий в развитии. Ему принадлежат первые математические модели развивающихся систем, в ряде отношений предвосхитившие современные. Своей основной целью Гурвич считал построение теории биологического поля, которую он постоянно видоизменял и совершенствовал, стремясь найти все более точные и близкие к физике формулировки взаимодействия частей в развивающемся организме. А. Г. Гурвич М. М. Завадовский Чарлз Чайлд Новый этап в изучении механизмов развития связан с исследованиями школы Михаила Михайловича Завадовского (1891–1957), создавшего направление, названное им динамикой развития. Завадовский придавал большое значение физиологическим аспектам развития, гуморальной среде, гормонам и т. д. Эти исследования привели к формулированию представлений о полях (А. Г. Гурвич, Н. К. Кольцов и др.), физиологических градиентах Чайлд Чарлз Мэннинг (1869–1954) или градиент-полях. Постепенно формировалось направление «химической эмбриологии» Джозефа Нидхем (1903–1995), интенсивно развивалась цитология и генетика индивидуального развития. Все это открывало новые возможности для более детального изучения различных сторон индивидуального развития, для углубления в детали процессов. Вместе с тем основные принципы и категории науки об индивидуальном развитии, ее методология, сформированные в аналитической и экспериментальной эмбриологии, не испытывали сильного влияния со стороны других дисциплин. 24 Болотов А. В. Биология размножения и развития Джозеф Нидхем Д. П. Филатов П. П. Иванов Одно из важных направлений эмбриологии, возникшее в бывшем СССР, связано с именем Дмитрия Петровича Филатова (1876–1943). Филатов обосновал сравнительно морфологический подход в экспериментальной эмбриологии, который был направлен на устранение накопившихся к тому времени противоречий между сравнительно-эволюционной и экспериментальной эмбриологией. Он ввел представление о «формообразовательном аппарате» как системе двухсторонних взаимодействий между индуктором и реагирующей тканью, первым отметил неспецифичность ранних этапов детерминационного процесса, обосновал принцип комплексности развития. Д. П. Филатов создал крупную школу советских эмбриологов-экспериментаторов, из которых многие (В. В. Попов, А. Т. Детлаф) внесли существенный вклад в науку. Из других выдающихся эмбриологов следует отметить Петра Павловича Иванова (1878–1942) – автора теории о лавральном и постлавральном отделах тела первичноротых, которая в наше время успешно применена к позвоночным животным. Большой вклад в развитие эмбриологии внес ученик и последователь П. П. Иванова Павел Григорьевич Светлов (1892– 1974), который уделил внимание изучению роли ряда экологических факторов (температура, голодание, ионизирующая радиация и др.) в ходе эмбриогенеза. Им установлены критические периоды развития у всех животных, во время которых зародыши оказываются легкоранимыми. Теория критических периодов, разработанная П. Г. Светловым, имеет большое значение для биологии и медицины, так как позволяет прогнозировать возможность возникновения патологии развития и уродств. 25 История развития эмбриологии П. Г. Светлов А. А. Заварзин Г. А. Шмидт Развитие современной эмбриологии в значительной мере связано с деятельностью советских ученых. Георгий Александрович Шмидт (1896–1975) исследовал ряд проблем сравнительной эмбриологии позвоночных и беспозвоночных животных, а также связь развивающегося зародыша птицы с окружающей средой, которую он видел в развитии эмбриональных приспособлений к обмену веществ, а они обусловливают формообразовательные процессы самого зародыша. В последствии Г. А. Шмидт установил 6 стадий развития зародыша курицы. Борис Петрович Топкин (1900–1984) развивал учение о соматических эмбриогенезах, т. е. развитие организмов из соматических клеток. Алексей Алексеевич Заварзин (1886–1845) сформировал понятие «эволюционной динамики тканей» как исторически обусловленные изменения свойств и потенций к совершенствованию в определенном направлении функционально аналогичных тканей. Он рассматривал проблему эволюционной динамики тканей как основную в эволюционной гистологии. Николай Григорьевич Хлопин (1897–1961) сделал крупное теоретическое обобщение в области изучения эволюционного развития тканей («теория дивергентного развития»). Отечественный эмбриолог Алексей Георгиевич Кнорре (1914–1981) внес ценный вклад в учение о эмбриональных гистогенезах. 26 Болотов А. В. Биология размножения и развития А. Г. Кнорре Н. Г. Хлопин Б. П. Токин Леонид Яковлевич Бляхер (1900–1987) изучал проблемы морфологии животных: соотношению формы и функции, аналогии и гомологии образования клеток в онтогенезе. Георгий Викторович Лопашов (1913–2010) стоял у истоков клонирования. Разработал метод пересадки ядер в яйцеклетку лягушки. Сформулировал положения концепции «клональной теории иммунитета», а также изменил представления о ходе и принципах эмбриогенеза. В 2003 г. российские биологи под руководством профессора Георгия Викторовича Лопашова первыми в мире вырастили новый хрусталик в глазу крысы. При разработке методик культивирования тканей и зачатков органов вне организма внесли большой вклад в науку о культуре клеток, тканей и органов вне организма (Ж. Лёб, А. Каррель, Н. Г. Хлопин, Э. Вольф, У. Фелл и др.). А открытие метода тканевых культур принадлежит Россу Гаррисону Гренвиллу (1870–1959). В 30–40-е гг. XX в. успешно разрабатывались вопросы э в о л ю ц и о н н о й э м б р и о л о г и и большим отрядом отечественных эмбриологов во главе с академиком Алексеем Николаевичем Северцовым. В настоящее время это направление продолжает разрабатываться эмбриологами. А. Н. Северцов (1866–1936) и его сотрудники на основе сравнительно-эмбриологического материала о развитии органов, решили вопрос об эволюции всех низших позвоночных – от древнейших предков хордовых до появления первых наземных четвероногих. Установлено два типа закладки изменяющихся во История развития эмбриологии Г. В. Лопашов 27 Росс Гарисон Гренвилл А. Н. Северцов время эволюции органов. Один тип характеризуется поздними «прибавлениями» к зародышевым признакам предков. При другом – новые изменения органа возникают на ранних стадиях развития, перестраивая дальнейший ход последнего. Это послужило основанием для уточнения биогенетического закона Мюллера – Геккеля о том, что онтогенез повторяет филогенез. Оказалось, что этот закон справедлив лишь по отношению к животным первого типа развития. Такое представление о взаимоотношениях между онтогенией и филогенией вошло в науку под названием филэмбриогенеза. Филэмбриогенезы – изменения, приобретенные в процессе эмбрионального развития, сохраняющиеся у взрослых животных и передающиеся потомству, т. е. эмбриональные изменения, связанные с филогенетическим развитием взрослого организма, – эволюционно-значимые изменения в ходе индивидуального развития. Филэмбриогенезы тканей – это эволюционный механизм тканевой дивергенции. А. Н. Северцов выделил три основных механизма филэмбриогенеза: 1) анаболию – надставку конечной стадии развития ткани или органа, при которой онтогенез продолжается после достижения той стадии, на которой у предков он закончился (например, ороговение покровного эпителия); 2) девиацию – отклонение в развитии на промежуточной стадии (например, развитие перьев у птиц и волос у млекопитающих); 28 Болотов А. В. Биология размножения и развития 3) архаллаксис – изменение первичных зачатков органа, при котором с самого начала онтогенез идет иначе, чем у предков (например, образование из эктодермы нервной трубки у хордовых животных или появление многослойного эпидермиса у позвоночных). Современная биология развития ведет интенсивные исследования молекулярных, биохимических и генетических механизмов различных формообразовательных процессов, активно изучает особенности субклеточной и клеточной организации зародышей. Естественно, что не утратили своего значения гистологические и анатомические исследования. Безусловную актуальность сохраняют исследования эволюции онтогенеза, и, прежде всего, исследования эволюции механизмов онтогенеза. Это новое направление – эволюционная биология развития в англоязычной литературе получило сокращенное наименование «Evo-Devo». В настоящее время прикладная биология стремится не только восстановить нарушенное какими-либо воздействиями нормальное течение жизненных процессов (как в отдельном организме, так и в экологическом сообществе), но и создавать новые, не существовавшие в природе биологические конструкции. Это направление получило название биотехнологии, или биоинженерии. Прикладная эмбриология является неотъемлемой и важной ее частью. Активно развиваются такие ее направления, как криоконсервация (хранение при низких температурах) эмбрионального материала, различные способы искусственного (в том числе – экстракорпорального, или «пробирочного») оплодотворения. На очереди – захватывающие задачи создания «банков» тканей и органов из стволовых клеток для трансплантаций взамен органов искусственных или взятых от доноров. I. РАЗМНОЖЕНИЕ ОРГАНИЗМОВ Размножение – свойство живых организмов воспроизводить себе подобных, это процесс смены поколений. Размножение происходит на разных уровнях организации: молекулярно-генетическом (репликация ДНК), клеточном (амитоз, митоз) и организменном. Биологическая роль размножения заключается в том, что происходит смена родительских поколений на дочерние которые несут в себе индивидуальные изменения, что способствует эволюционным процессам. Задачи размножения – увеличения числа особей, сохранение складывающихся в процессе эволюции типов структурнофизиологической организации (сохранение биологической информации), что делает возможным естественный отбор и, следовательно, эволюцию жизни. Последнее связанно с тем, что при размножении возникает изменчивость дочерних организмов. Готовность к размножению и приуроченность времени спаривания к определенному периоду обеспечивается сложным регуляторным механизмом, действующим по сигналам внешней среды. В умеренных и высоких широтах таким сигналом служит изменение длины светового дня. Для видов с весенней течкой сигнальное значение имеет увеличение длины дня (многие звери), для спаривающихся осенью копытных – ее сокращение, а для волков – короткий зимний день. Изменение светового режима воздействует на нейрогипофиз, стимулирующий выделение гонадотропных гормонов передней долей гипофиза, влияющих на гонады. В низких широтах значение сигнального фактора могут принимать другие периодические явления природы, предшествующие приближению благоприятного для размножения времени (в пустынях и саваннах – осадки, в тропических лесах – созревание основных кормовых растений или их плодов и т. п.). Даже в тропических лесах с их устойчиво теплым и влажным климатом размножение млекопитающих обычно сезонно, хотя у разных видов может приходиться на разные периоды года. Размножение может быть бесполым и половым. 30 Болотов А. В. Биология размножения и развития 1. Бесполое размножение Признаки бесполого размножения: 1) в воспроизведении участвует одна родительская особь; 2) источник генетической информации – соматические клетки; 3) генотипы дочерних особей идентичны родительскому; 4) быстрое увеличение числа особей; 5) обеспечивает существование вида в неизменяющихся условиях среды; 6) дочернее поколение у одних видов возникает из одной или группы клеток материнского организма, у других видов – в специализированных органах. Способы бесполого размножения: А. Бесполое размножение у одноклеточных организмов чередуется с половым процессом (см. рис. 1): 1) деление клеток надвое (бактерии, простейшие, водоросли) (рис. 1, 1). 2) шизогония (рис. 1, 2) – множественное деление (споровики). Шизогония свойственна малярийным плазмодиям, относящимся к простейшим. При этом содержимое материнской клетки многократно делится. 3) почкование – на материнской клетке появляется небольшой бугорок, который увеличивается, достигает размеров материнского организма и отделяется от него (дрожжи). 4) спорогония (рис. 1, 8) (споровики). Б. У многоклеточных организмов бесполое размножение идет в целом в двух вариантах (вегетативный и спорообразование). Вегетативное – размножение частью материнского организма. Спорообразование связано с образованием специализированных клеток – спор, которые являются зачатком нового организма. Формы бесполого размножения у многоклеточных организмов: 1. При помощи вегетативных (рис. 1, 5): органов растений, например: стебель (смородина), лист (фиалка), корень (осот), клубень (картофель), луковица (лук). 2. Спорообразование (рис. 1, 6). У грибов споры образуются из любой клетки митозом, а у высших споровых растений – в спорангиях мейозом. I. Размножение организмов 31 3. Почкование (гидра) (рис. 1, 3). 4. Полиэмбриония – разделение клеточного материала зародыша на стадии ранней бластулы на несколько частей, из которых развиваются отдельные особи (броненосцы, наездники). К полиэмбрионии относится образование однояйцевых близнецов у человека. В этом случае зигота, возникшая в результате оплодотворения, дробясь, образует зародыш, который разделяется на несколько частей, в результате чего рождаются младенцы обязательно одного пола (близнецы). 5. Фрагментация (рис. 1, 4) – (неупорядоченное деление) материнская особь делится на несколько неравных частей и каждая из них развивается в самостоятельный организм (ресничные и кольчатые черви). В основе фрагментации лежит способность организмов к регенерации. 6. Стробиляция (рис. 1, 7) – упорядоченное деление материнской особи на несколько частей (медузы). Рис 1. Виды бесполого размножения. 1 – деление; 2 – шизогония; 3 – почкование; 4 – фрагментация; 5 – вегетативное размножение; 6 – спорообразование В. Клонирование – комплекс методов, используемых человеком для получения генетически идентичных копий клеток или особей. Клон – совокупность клеток или особей, произошедших от общего предка путем бесполого размножения. В основе получения клона лежит митоз (у бактерий – простое деление). 32 Болотов А. В. Биология размножения и развития 2. Половое размножение у покрытосеменных растений Органом полового размножения покрытосеменных растений является цветок. Цветок – видоизмененный, укороченный, неразветвленный побег, предназначенный для образования спор и гамет и полового процесса, завершающегося образованием семян и плода. Строение цветка У цветка различают цветоножку, цветоложе, околоцветник, тычинки и пестики (рис. 2). У некоторых цветков отдельные части могут отсутствовать. Цветки большинства видов растений имеют и тычинки, и пестики. Такие цветки называют обоеполыми (вишня, горох и др.). Цветки, которые имеют только пестики, называют пестичными (женскими). Цветки, которые имеют только тычинки, называют тычиночными (мужскими). В зависимости от распределения однополых цветков на растениях различают: однодомные растения – растения, у которых на одних и тех же экземплярах располагаются и женские, и мужские цветки (огурец, кукуруза, дуб и др.); двудомные растения – растения, у которых на одних экземплярах располагаются женские, а на других – мужские цветки (крапива двудомная, конопля, облепиха); многодомные растения – растения, у которых на одних и тех же экземплярах встречаются как обоеполые, так и однополые цветки в различных количественных соотношениях (гречиха, некоторые виды ясеня, клена и др.). Цветоножка (рис. 2, 1) – междоузлие под цветком. Цветки, лишенные цветоножки, называются сидячими (цветки в соцветии корзинка у подсолнечника, астры, одуванчика). Цветоложе (рис. 2, 2) – укороченная стеблевая часть цветка. На ней располагаются все остальные части цветка. Околоцветник – стерильная часть цветка, его покров. Околоцветник может быть простым (не дифференцированным на чашечку и венчик, образованным совокупностью однородных листочков (рис. 2, 4), имеющих одинаковые размеры и окраску) и двойным (дифференцированным на чашечку и венчик, отличающиеся друг от друга размерами и окраской. Простой около- I. Размножение организмов 33 цветник может быть венчиковидным (образованным ярко окрашенными листочками) или чашечковидным (образованным зелеными листочками). Цветки, лишенные околоцветника (ива, тополь), называются голыми. Рис. 2. Строение цветка. 1 – цветоножка; 2 – цветоложе; 3 – чашелистики; 4 – лепесток; 5 – завязь; 6 – тычинка; 7 – пыльник; 8 – тычиночная нить; 9 – столбик; 10 – рыльце; 11 – околоцветник; 12 – семязачаток Чашечка – наружная часть двойного околоцветника, представляет собой совокупность чашелистиков (рис. 2, 3) – видоизмененных прицветных листьев. Обычно чашелистики имеют небольшие размеры и зеленую окраску. Они сходны с обычными листьями, но устроены проще. Различают: раздельнолистную чашечку – чашечку, образованную свободными (несросшимися) чашелистиками (капуста, лютик и др.); сростнолистную чашечку – чашечку, образованную частично или полностью сросшимися чашелистиками (картофель, табак, горох и др.). Венчик – внутренняя, обычно окрашенная часть двойного околоцветника. Представляет собой совокупность лепестков (рис. 2, 4), часто имеющих яркую окраску. Количество лепестков венчика может быть различным. Лепестки могут быть более или менее одинаковыми (лютик, яблоня и др.) либо отличаться размерами и формой (фиалка, горох и др.). В результате венчик мо- 34 Болотов А. В. Биология размножения и развития жет быть правильным, неправильным или асимметричным. Венчик, как и чашечка, может быть раздельнолепестным и сростнолепестным. Раздельнолепестной венчик состоит из свободных, несросшихся лепестков. Сростнолепестной венчик состоит из сросшихся в той или иной степени лепестков. Главная функция венчика – привлечение опылителей. Андроцей – совокупность тычинок (рис. 2, 6) одного цветка. Количество тычинок в цветке – от одной (орхидные) до нескольких сотен (некоторые кактусы). У большинства растений тычинка состоит из тычиночной нити (рис. 2, 8) и пыльника (рис. 2, 7). Рис. 3. Строение тычинки (А) и пыльника (поперечный разрез – Б). 1 – тычиночная нить (филамент); 2 – пыльцевой мешок (пыльник); 3 – связник; 4 – надсвязник; 5 – сосудистый пучок; 6 – гнездо пыльника (соответствующее одному микроспорангию); 7 – раскрывающаяся половинка пыльника с высыпающимися пыльцевыми зернами; 8 – стенка пыльника Тычиночная нить (рис. 2, 8; 3, 1) – нижняя, как правило, суженная стерильная часть тычинки. Нижний конец тычиночной нити отходит от цветоложа, а верхний конец несет пыльник. Обычно тычиночные нити тонкие, длинные, округлые в сечении. Пыльник (рис. 2, 7; 3, 2) – верхняя расширенная фертильная часть тычинки. Пыльник состоит из двух половинок, соединенных связником. Каждая половинка имеет два пыльцевых гнезда I. Размножение организмов 35 (микроспорангия (рис. 3, 4)), в которых происходит образование микроспор, а впоследствии пылинок. Связник (рис. 3, 3) является продолжением тычиночной нити, через него в пыльник поступают питательные вещества. Микроспорогенез Микроспорогенез – процесс образования микроспор в микроспорангиях (гнездах пыльника). Микроспоры формируются из материнских клеток – микроспороцитов, имеющих диплоидный набор хромосом. В результате мейоза каждая материнская клетка образует четыре гаплоидных микроспоры. Микроспоры быстро обособляются друг от друга. Микрогаметогенез Микрогаметогенез – процесс образования мужских половых клеток (спермиев), происходит в пыльцевом зерне, которое является мужским гаметофитом покрытосеменных растений. Развитие мужского гаметофита происходит также в гнездах пыльников тычинок и сводится к одному митотическому делению микроспоры и формированию оболочек пыльцевого зерна. Оболочка пыльцевого зерна состоит из двух слоев: интины (внутренней, тонкой) и экзины (наружной, толстой). Каждое пыльцевое зерно содержит две гаплоидные клетки: вегетативную и генеративную. Из генеративной (спермагенной) далее образуются два спермия. Из вегетативной (сифоногенной) впоследствии образуется пыльцевая трубка. Гинецей Гинецей – совокупность пестиков одного цветка. Обычно в пестике выделяют три части: завязь, столбик и рыльце (рис. 2, 5; 2, 9; 2,10; 4, 1–3). Завязь (рис. 2, 5; 4, 3) – замкнутая, нижняя, полая часть пестика, несущая и защищающая семязачатки. Завязь бывает: верхняя, нижняя, полунижняя. В завязи может располагаться от одного (пшеница, вишня) до нескольких тысяч (мак) семязачатков. Стенки завязи выполняют функцию защиты семязачатков от неблагоприятных факторов среды (высыхание, колебание температур, поедание насекомыми и т. д.). Внутри завязи (в семязачатках) происходит мегаспорогенез и мегагаметогенез, они принимают участие в образовании околоплодника. 36 Болотов А. В. Биология размножения и развития Столбик (рис. 2, 9; 4, 2) – средняя более или менее удлиненная стерильная часть пестика, отходящая обычно от верхушки завязи, соединяет завязь и рыльце. Рыльце (рис. 2, 10; 4, 1) – верхняя расширенная часть пестика, предназначено для восприятия пыльцы. Рыльце может быть разнообразной формы (двухлопастное, звездчатое, перистое и т. д.) и размера в зависимости от особенностей опыления. При отсутствии столбика рыльце называют сидячим. Рис. 4. Строение пестика. 1 – рыльце; 2 – столбик; 3 – завязь; 4 – семязачаток; 5 – зародышевый мешок; 6 – плацента; 7 – пыльцевое зерно; 8 – пыльцевая трубка Рис. 5. Строение завязи (семязачатка) и зародышевого мешка (женского гаметофита) покрытосеменных растений. 1 – покровы семязачатка, или интегументы (а – наружный, б – внутренний); 2 – микропиле; 3 – халаза; 4 – семяножка (фуникулус); 5 – нуцеллус; 6 – зародышевый мешок; 7 – яйцеклетка; 8 – синергиды; 9 – антиподы; 10 – вторичное ядро; 11 – плацента; 12 – проводящий пучок Семязачаток (рис. 2, 12; 5) состоит из нуцеллуса (ядра) (рис. 5, 5) – центральной части, являющейся мегаспорангием, двух покровов – интегументов (рис. 5, 1), которые при смыкании образуют узкий канал – микропиле (рис. 5, 2), или пыльцевход, через I. Размножение организмов 37 который пыльцевая трубка проникает к зародышевому мешку. С помощью семяножки (рис. 5, 4) семязачаток прикрепляется к плаценте (рис. 5, 3). Место прикрепления семязачатка к семяножке называют рубчиком. Противоположную микропиле часть семязачатка, где сливаются нуцеллус и интегументы, называют халазой (рис. 5, 10). В семязачатке происходят мегаспорогенез, мегагаметогенез и процесс оплодотворения. После оплодотворения (реже без него) из семязачатка формируется семя. Мегаспорогенез Процесс формирования мегаспор называется мегаспорогенезом. Он происходит в нуцеллусе семязачатка. После заложения семязачатка и формирования нуцеллуса в области микропиле начинает разрастаться одна археспориальная (спорогенная) клетка – мегаспороцит, или материнская клетка мегаспор. Материнская клетка мегаспор имеет диплоидный набор хромосом. У большинства покрытосеменных из нее путем мейоза образуются четыре гаплоидные мегаспоры. Из них лишь одна (обычно нижняя, обращенная к халазе, реже верхняя, обращенная к микропиле) дает начало женскому гаметофиту – зародышевому мешку. Остальные мегаспоры отмирают. Мегагаметогенез Процесс формирования женских половых клеток происходит в зародышевом мешке. Формирование женского гаметофита начинается с разрастания мегаспоры, которая далее три раза делится митозом. В результате этого образуются восемь клеток, которые располагаются следующим образом: три – на одном полюсе зародышевого мешка (микропилярном), три – на другом (халазальном), две – в центре. Две оставшиеся сливаются в центре клетки, образуя диплоидную центральную клетку зародышевого мешка. Одна из трех клеток, расположенных на микропилярном полюсе, отличается большими размерами и является яйцеклеткой (рис. 5, 7). Две рядом расположенные клетки являются вспомогательными и называются синергидами (рис. 5, 6). Группа из трех клеток, находящихся на противоположном, халазальном полюсе, называется антиподом (рис. 5, 9). Таким образом, сформированный женский гаметофит включает шесть гап- 38 Болотов А. В. Биология размножения и развития лоидных клеток (яйцеклетка, две клетки-синергиды, три клеткиантипода) и одну диплоидную клетку. Оплодотворение. Образование семян и плодов Процессу оплодотворения предшествует опыление – перенос пыльцы от пыльцевых мешков тычинок к рыльцам пестиков. Попав на рыльце пестика, под воздействием веществ, выделяемых пестиком, пыльца начинает прорастать: образуется пыльцевая трубка, внедряющаяся в ткань рыльца. Кончик пыльцевой трубки выделяет вещества, размягчающие ткань рыльца и столбика. В процессе формирования пыльцевой трубки принимает участие сифоногенная клетка. По мере роста пыльцевой трубки в нее переходит спермагенная клетка, которая делится митозом с образованием двух спермиев (у некоторых растений спермагенная клетка дает начало двум спермиям еще до прорастания пыльцы). Пыльцевая трубка продвигается по столбику пестика и врастает в зародышевый мешок, как правило, через микропиле. После проникновения в зародышевый мешок кончик пыльцевой трубки разрывается, и спермии попадают внутрь. Один из спермиев сливается с яйцеклеткой, образуя диплоидную зиготу. Второй спермий сливается с центральной клеткой зародышевого мешка, образуя триплоидную клетку, из которой далее формируется эндосперм (питательная ткань) семени, обеспечивающий питание зародыша. Синергиды и антиподы дегенерируют. Вышеописанный процесс получил название двойного оплодотворения. Двойное оплодотворение у цветковых растений было открыто в 1898 г. русским ботаником Сергеем Гавриловичем Навашиным (1857–1930). После двойного оплодотворения из яйцеклетки формируется зародыш семени, из центрального ядра зародышевого мешка – эндосперм, из интегументов – семенная кожура, из всего семязачатка – семя, а из стенок завязи – околоплодник. В целом из завязи пестика формируется плод с семенами. I. Размножение организмов 39 Среди российских ученых, внесших весомый вклад в формирование эмбриологии и биологии развития растений, следует отметить: Йозефа Готлиба Кельрейтера (1733–1806), который является основоположником учения о биологии цветения и гибридизации; Андрея Тимофеевича Болотова (1738–1833) с работами по биологии цветения и плодообразованию у некоторых культурных растений; Ивана Николаевича Горожанкина (1848–1904), выполнившего работы по развитию гаметофитов и оплодотворению у голосеменных; Владимира Ивановича Беляева (1855–1911), исследовавшего спермиогенез у высших растений (хвощи, папоротники), описавшего редукционное деление у голосеменных и покрытосеменных. 3. Половое размножение животных Половое размножение является более прогрессивным, поскольку обеспечивает наследственное разнообразие. Половое размножение – совокупность процессов гаметогенеза, осеменения и оплодотворения, приводящих к воспроизведению. При половом размножении происходит образование половых клеток или гамет и последующее их слияние (гаметическая копуляция). Таким образом, половой процесс и размножение объединяются. Название гаметы произошло от греческих слов gamete – «жена», gametes – «муж». Половое размножение может быть с оплодотворением и без оплодотворения – партеногенез. Партеногенез Особая разновидность полового размножения – партеногенез – развитие из неоплодотворенной яйцеклетки (девственное размножение). Партеногенез обеспечивает рост численности особей в условиях, затрудняющих встречу партнеров противоположного пола. Партеногенез – это особый вид полового размножения, при котором новый организм развивается из неоплодотворенной яйцеклетки, таким образом, обмена генетической 40 Болотов А. В. Биология размножения и развития информацией не происходит, как и при бесполом размножении. Аналогичный процесс у растений называется апомиксис. Существует два типа партеногенеза: облигатный (т. е. обязательный) и факультативный. Первый свойствен видам, у которых самцов либо нет совсем, либо они редки и не способны функционировать. К таким видам относятся некоторые тли, палочники, сверчки, бабочки; популяции без самцов изредка встречаются у рыб, например у серебряного карася. При факультативном партеногенезе яйца могут развиваться как партеногенетически, так и в результате оплодотворения, причем партеногенетическое размножение может преобладать в условиях, когда слишком редки контакты разнополых особей, например, на границе ареала распространения вида. Известен также циклический партеногенез, при котором размножение с участием обоих полов чередуется с партеногенетическим. Например, многие виды тлей дают несколько партеногенетических поколений в течение короткого теплого периода лета, а на зиму откладывают оплодотворенные яйца, которые покрыты плотной оболочкой и способны перезимовывать; весной из них выходят только самки, но осенью появляется поколение с некоторым количеством самцов – и цикл возобновляется. Аналогичным образом размножаются и некоторые другие виды с высокой сезонной смертностью, например коловратки. Циклический партеногенез наблюдается также у видов с личиночным размножением; при этом оплодотворенные яйца откладывают обычно только зрелые особи, а у личинок они развиваются партеногенетически. Гиногенез – развитие организма на основе ядра яйцеклетки, диплоидный набор хромосом восстанавливается путем слияния ядра ооцита и редукционного тельца. Андрогенез – развитие из клетки с цитоплазмой ооцита и хромосомами двух сперматозоидов. Ядро женской гаметы в случае андрогенеза разрушается. В основе полового размножения с оплодотворением лежит половой процесс – обмен генетической информацией между особями вида (конъюгация), или ее объединение (копуляция). Формами полового процесса являются конъюгация и копуляция (рис. 6, 1–2). I. Размножение организмов 41 Рис. 6. Формы полового процесса (1, 2) и формы копуляции (а, б, в) Конъюгация характерна для бактерий и инфузорий. Две инфузории сближаются, между ними возникает цитоплазматический мостик, и после преобразования микронуклеусов происходит обмен мигрирующими ядрами (рис. 6, 2). Копуляция – половой процесс, при котором две особи соединяются друг с другом при половом акте (одноклеточные) или происходит слияние женских и мужских половых клеток и образуют зиготу. Формы копуляции: • изогамия – гаметы не имеют морфологических различий (жгутиковые Polytoma (рис. 6, 1а); • анизогамия – гаметы дифференцированы на крупные и мелкие подвижные клетки (колониальные жгутиковые Pandorina morum) (рис. 6, 1б); • оогамия – крайняя степень дифференцировки: большая гамета – неподвижна, а мелкая – подвижна (Volvox globator) (рис. 6, 1в). Гаметогенез – процесс образования половых клеток В гаметогенезе различают три стадии: размножения, роста, созревания. В стадии размножения происходит деление гаметогоний митозом. В стадии роста гаметоциты 1-го порядка достигают размеров, характерных для половых клеток. В стадии созревания после редукционного деления образуются гаметоциты 2-го порядка, а после эквационного – яйцеклетки и сперматиды, которые, проходят еще одну стадию – формирование сперматозоидов. 42 Болотов А. В. Биология размножения и развития Характеристика полового размножения с копуляцией: • в воспроизведении участвует две родительские особи; • источник генетической информации – половые клетки родителей; • генотипы дочерних особей отличаются от родительских, вследствие комбинативной изменчивости; • способствует приспособлению организмов в изменяющихся условиях среды. Половое размножение объединяет обширный круг морфологических и физиологических явлений: половые различия; строение и развитие органов, в которых развиваются половые клетки; физиологические особенности самцов и самок и морфологические особенности органов размножения. Форма и размеры органов размножения должны соответствовать друг другу, т. е. подходить друг к другу как «ключик к замочку», в противном случае, оплодотворение будет затруднено или станет невозможным. Эволюция размножения Эволюция размножения шла, как правило, в направлении от бесполых форм к половым, от изогамии к анизогамии, от участия всех клеток в размножении к разделению клеток на соматические и половые, от наружного оплодотворения к внутреннему с внутриутробным развитием и заботой о потомстве. Темп размножения, численность потомства, частота смены поколений наряду с другими факторами определяют скорость приспособления вида к условиям среды. Например, высокие темпы размножения и частая смена поколений позволяют насекомым в короткий срок вырабатывать устойчивость к ядохимикатам. В эволюции позвоночных – от рыб до теплокровных – наблюдается тенденция к уменьшению численности потомства и увеличению его выживаемости. II. ПОЛОВОЕ СОЗРЕВАНИЕ Поскольку потомство животных рождается с недоразвитыми половыми органами, способность к размножению проявляется лишь в определенном возрасте, с наступлением полового созревания животных. Это состояние характеризуется следующими основными признаками: 1) половые органы достигают полного развития; в яичниках и семенниках созревают половые клетки (соответственно яйцеклетки и спермии); 2) формируется половое поведение животных (возбуждение, влечение, половые рефлексы); 3) в эндокринной ткани гонад начинают усиленно вырабатываться женские и мужские половые гормоны; 4) у самок проявляются циклические изменения в половых путях. Следовательно, половое созревание – это морфологическое и функциональное оформление полового аппарата, когда самец становится способным оплодотворить самку, а самка – забеременеть. Половое созревание – период жизни, в течение которого организм достигает биологической половой зрелости и характеризуется появлением вторичных половых признаков и окончательным формированием половых органов и желез. Сроки полового созревания различны, они зависят от наследственных факторов, условий жизни, питания, состояния здоровья и климатических факторов. У самок и самцов они происходят, примерно, в одни сроки: Кролик Кошка Свинья, хряк Женщина Животные, месяц 5 Собака 5 Корова, бык 6 Кобыла, жеребец Человек, год 12–15 Мужчина 7 7,5 16 13–16 Одним из главных стимулов, запускающих развитие и функционирование половой системы, является начало функцио- 44 Болотов А. В. Биология размножения и развития нирования цирхоральной системы. В нейронах медиобазального гипоталамуса происходит пульсирующая секреция РГЛГ(рилизинг-гормон ЛГ (РГ-ЛГ люлиберин) в часовом ритме. По аксонам нервных клеток нейросекрет (РГ-ЛГ) поступает в портальную систему и с кровью переносится в переднюю долю гипофиза. Образование двух гонадотропинов: фолликулостимулирующего гормона (ФСГ) и лютиенизирующего гормона (ЛГ), ведет к увеличению размеров гонад, повышению их генеративной и гормональной активности. В онтогенезе эндокринная функция яичка устанавливается раньше, чем генеративная. Мужской половой гормон – тестостерон начинает вырабатывать у зародыша человека с 8–10-й недели внутриутробного периода. В зародышевом яичке еще до начала биосинтеза тестостерона образуются поочередно несколько белковых (пептидных) андрогенов, принадлежащих к ингибинам. Первым на стадии индифферентной гонады появляется гормон, под влиянием которого происходит редукция парамезонефрального протока, и с этого момента индифферентный зачаток половой системы дифференцируется по мужскому типу. В женском организме эндокринная функция гонад возникает впервые. Вырабатываемые половые гормоны, помимо их влияния на развитие экстрагенитальных половых признаков, необходимы для процессов спермиогенеза и овогенеза. В половом созревании участвует центральная нервная система – преоптическая область гипоталамуса, миндалевидные ядра, лимбическая система мозга, однако клеточные и молекулярные механизмы этих процессов неясны. Четко очерченного полового центра в ЦНС нет. При половом созревании в разных отделах мозга (прежде всего в гипоталамусе) возникает половая доминанта эндогенного (гормонального) происхождения, лежащая в основе мотивационного возбуждения и формирования соответствующего поведения животных. Степень половой активности зависит от изменений гормонального статуса организма. Способность к воспроизводству у животных возникает раньше, чем заканчивается их рост и развитие, т. е. раньше, чем происходит полное «физиологическое созревание». Этим термином обозначают состояние, когда организм приобретает формы, свойственные взрослому животному данного пола и достигает 45 II. Половое созревание 70–75 % его живой массы. У самок при этом стабилизируются циклические проявления половой функции. Период физиологического созревания является оптимальным временем включения животных в репродуктивный процесс. Он наступает в следующем возрасте: Кролик Кошка Свинья, хряк Женщина Животные, месяц 6 Собака 10 Корова, бык 10 Кобыла, жеребец Человек, год 20 Мужчина 12 17 36 21 Использовать животных в качестве производителей можно в течение всего периода, пока они сохраняют половую потенцию. Однако в условиях интенсивной эксплуатации эти сроки, особенно для самок, значительно сокращаются. Становление человека как индивида и личности предполагает диалектическое взаимодействие двух относительно автономных, но неразрывно связанных элементов развития – биологического и социального. Социальное созревание – сложный процесс, темпы и этапы которого определяются разными критериями. Социальное созревание по времени чаще всего не совпадает с физическим и обычно наступает после физического. III. ПОЛОВАЯ СИСТЕМА МЛЕКОПИТАЮЩИХ Половые органы у высших животных и человека имеют сходное строение. Поэтому строение половых органов рассматривается на примере строения их у человека. Половые органы подразделяются на мужские и женские. Наиболее существенной частью их являются половые железы: яичники у женщин и яички (семенники) у мужчин. По расположению половые органы делят на наружные и внутренние. 1. Мужская половая система К мужским половым органам относятся: яички с их оболочками, семявыносящие протоки и придаточные половые железы: семенными пузырьками, предстательная железа, бульбоуретральные (куперовы) железы – внутренние половые органы; половой член и мошонка – наружные половые органы. Семенники Яички (testes), или семенники представляют собой овальные тела, несколько сплющенные с боков, которые расположены в мошонке. Это обеспечивает поддержание оптимальной температуры для сперматогенеза не выше 35 ºС. Функции семенников – в них образуются мужские половые клетки – сперматозоиды и мужской половой гормон – тестостерон. В среднем длина яичка равна 4 см, ширина – около 3 см. Вес 20–30 г. В яичке различают латеральную и медиальную поверхности, передний и задний края, верхний и нижний концы. Левое яичко обычно опущено несколько ниже, чем правое. У заднего края яичка располагаются придаток яичка (epididymis) и семенной канатик (рис. 7). Придаток представляет собой узкое тело, в котором различают верхнюю часть – головку, среднюю – тело и нижнюю – «хвост» (рис. 7). При коитусе спермии освобождаются не прямо из семенника, а из каудальной части («хвоста») придатка семенника. В канале придатка спер- III. Половая система млекопитающих 47 мии накапливаются в больших количествах (20–40 млрд у быка). Здесь они претерпевают дальнейшие морфофункциональные изменения («дозревают») в течение 8–20 дней. В кислой бескислородной среде канала придатка спермии впадают в состояние, подобное анабиозу, приобретают уплотненную липопротеидную оболочку и отрицательный заряд, что предохраняет их от действия кислых продуктов и от агглютинации в половых путях самки. В придатке изменяются также антигенные свойства поверхности спермиев. Оплодотворяющую способность спермии сохраняют в придатках семенника до 2–3 мес. Достигшие каудального отдела придатка спермии обладают высокой оплодотворяющей способностью и могут высвобождаться при эякуляции. Рис. 7. Семенник с придатком и семявыносящим протоком Яичко окружено плотной фиброзной оболочкой беловатого цвета – белочной оболочкой (рис. 7; 22, А), лежащей непосредственно на его паренхиме. Паренхима семенника состоит из множества извитых канальцев и расположенных между ними клеток Лейдига (рис. 22, А, Б) и прямых семенных канальцев. В извитых канальцах происходит образование мужских половых клеток – сперматозоидов. Каждый семенной каналец имеет диа- 48 Болотов А. В. Биология размножения и развития метр от 150 до 250 мкм и длину от 30 до 70 см, а общая длина извитых канальцев у мужчины составляет 300–400 м. Для сравнения в одном семеннике взрослого кабана она составляет более 3000 м. У заднего края она образует утолщение – средостение яичка, от которого вперед лучеобразно расходятся фиброзные тяжи перегородки яичка, подходящие к внутренней поверхности белочной оболочки и делящие яичко на дольки (рис. 7). Всего долек от 100 до 300. Прямые канальцы открываются в сеть ходов в средостении яичка (рис. 7), откуда начинается 12–15 выносящих канальцев яичка, направляющихся в головку придатка. По выходе из яичка они прямые, а затем извиваются и образуют дольки придатка. Выносящие канальцы открываются в проток придатка яичка, который тянется, изгибаясь от головки до хвоста, где продолжается в семявыносящий проток. Выносящие канальцы семенника, выстланные однослойным плоским эпителием. Прямые семенные канальцы выстланы призматическим эпителием, канальцы сети – кубическим, а выносящие канальцы – реснитчатым эпителием. Выстилка извитых канальцев образована сперматогенным эпителием, между клетками которых находятся поддерживающие клетки (клетки Сертоли) (рис. 22, А, Б). Клетки сперматогенного эпителия лежат на базальной мембране. Базальная мембрана окружена рыхлой (снаружи) соединительной тканью (рис. Б-1), содержащей непрерывный миоидный слой (рис. 22, Б) (сократительные клетки, содержащие актин). Мошонка Мошонка (scrotum) представляет собой кожный мешок, в котором располагаются яички, их придатки и нижний отдел семенного канатика. По средней линии мошонки проходит шов мошонки – от нижней поверхности полового члена до анального отверстия. Остальная поверхность мошонки покрыта более или менее значительным количеством морщин. Оболочки яичка образованы из различных слоев брюшной стенки, которые яичко в процессе опускания увлекает за собой. Непосредственно под кожей мошонки располагается видоизмененная соединительная ткань, лишенная жира и содержащая значительное количество гладких мышечных клеток. Она имеет красноватый цвет и называется мясистой оболочкой. Под ней III. Половая система млекопитающих 49 имеется фасциальная пластинка – наружная семенная фасция. Следующий слой, покрывающий яичко и семенной канатик, представлен двумя образованиями: фасцией и мышечными волокнами, расходящимися по поверхности яичка. Это мышца, поднимающая яичко, и ее фасция. Мышца представляет собой производное поперечной и внутренней косой мышц живота, при ее сокращении яичко подтягивается кверху. Под мышцей, поднимающей яичко, располагается внутренняя семенная фасция, являющаяся продолжением поперечной фасции живота. Влагалищная оболочка яичка представлена двумя пластинками. Одна сращена с его белочной оболочкой (висцеральная пластинка), а другая образует вокруг него замкнутый мешок (париетальная пластинка), соприкасающийся с внутренней семенной фасцией. Эта оболочка является производной влагалищного отростка брюшины, образующегося в момент опускания яичка в мошонку. В результате зарастания верхнего участка влагалищного отростка связь между брюшинной полостью и полостью серозной оболочки яичка прерывается. В случаях незарастания влагалищного отростка остается канал, через который могут выходить врожденные мошоночные грыжи. Семявыносящий проток Семявыносящий проток (liictus deferens) (рис. 8, 11) парный орган длиной около 40–45 см, в поперечнике 3 мм и диаметр просвета около 0,5 мм в своем начальном отделе также делает изгибы и позади яичка поднимается вместе с сосудами и нервами к наружному паховому кольцу в составе семенного канатика, проходит через паховый канал, у внутреннего его кольца отделяется от сосудов яичка, идет вниз и назад по боковой стенке таза. У боковой поверхности мочевого пузыря он загибается к дну его и у средней линии подходит к предстательной железе. В нижнем отделе семявыносящий проток расширен и образует ампулу) семявыносящего протока. Стенка его состоит из наружной фиброзной оболочки, средние – мышечной и внутренней слизистой, образующей продольные складки. Сперма Сперма (sperma) – мужское семя, секретируется в извитых канальцах яичка, выстланных сперматогенным эпителием. Все 50 Болотов А. В. Биология размножения и развития остальные канальцы яичка, придатка, семявыносящий и семявыбрасывающий протоки являются, путями выведения спермы. По достижении половой зрелости в извитых канальцах начинаются процессы сперматогенеза, приводящие к образованию мужских половых клеток. Яичко вырабатывает спермии непрерывно на всем протяжении половой активности организма. Спермии образуются в огромном количестве, в половых путях женщины они сохраняют оплодотворяющую способность в течение 2–3 и более суток. Жидкая часть спермы в очень небольшом количестве продуцируется яичками. В основном она является продуктом выделения придаточных желез полового аппарата – семенных пузырьков и предстательной железы, открывающихся в пути, выводящие сперму. Состав спермы Спермой называют жидкость, выделяемую самцами при половых актах. Сперма (греч. sperma – семя; синонимы – семенная жидкость, эякулят) – смесь выделяемых во время эякуляции продуктов секреции мужских половых органов: яичек и их придатков, предстательной железы, семенных пузырьков, куперовых желез, уретры. рН спермы ≈ 7,0–7,6 (слабощелочная). Состоит из сперматозоидов и семенной жидкости. Мужская сперма – это сложное вещество, состоящее на 90 % из воды и из более чем 30 различных компонентов, включающих натрия хлорид (2 г/л), глюкозу (3 г/л), фруктозу (1–6 г/л), молочную кислоту (0,9–1 г/л), белок (1,8 г/100 мл), лимонную кислоту, высококонцентрированный калий и такой важнейший элемент, как цинк, содержит микроэлементы кальция, железа, фосфора и другие минеральные вещества. В состав спермы также входят сера, медь, магний, витамин С и В12, т. е. все самые важные химические элементы для здоровья человека. Кроме того, семенные пузырьки содержат 15 различных выделений предстательной железы, которые стимулируют мышечные сокращения и расширение кровеносных сосудов. Несмотря на присутствие лимонной кислоты, сперма имеет легкую щелочную среду. Порция спермы, выделенная самцом за одну садку, называется эякулятом. Объем эякулята и концентрация в нем спермиев у нормальных самцов разных видов различны. В пределах вида III. Половая система млекопитающих 51 они зависят от интенсивности использования производителя (частоты садок), условий кормления и содержания. Часть спермы, не содержащая спермиев, называется плазмой спермы (семенной плазмой). Ее относительный объем в эякуляте зависит от вида животных (70 % у барана, 30 % у быка; 90–92 % у хряка и жеребца; у человека 95 %). В эякуляте человека сперматозоиды составляют в среднем только 5 % эякулята. Остальные 95 % – это секрет предстательной железы и жидкость семенных пузырьков: секреции семенных пузырьков (75 %), секреции простаты (20 %), секреции куперовых желез и уретры (до 5 %). В первой порции эякулята содержание сперматозоидов выше, чем в последующих. Семенная плазма Она является стабилизирующей, разбавляющей, активирующей и питательной средой для спермиев. Она стабилизирует плазменную мембрану спермиев, содержит энзимы, растворяющие акросому, простагландины, инозит, андрогены, антиагглютинины (в секрете простаты); аскорбиновую кислоту, фруктозу, лимонную кислоту. При кастрации содержание фруктозы, лимонной кислоты и тестостерона падает. Секреты придаточных желез выделяются в просвет канала в момент эякуляции, что способствует смешиванию спермы с семенной плазмой в уретре и сообщает первичный стимул подвижности спермиям. У свиней желатиновая фракция семенной жидкости (секрет луковичных желез) может образовывать пробку, препятствующую вытеканию спермы из половых путей самки. У грызунов пробку образует секрет пузырьковидных желез. Небольшое количество секрета придаточных половых желез выделяется с мочой и в период полового покоя. Факторы, отрицательно влияющие на сперму • Возраст (чем старше, тем менее активными становятся сперматозоиды, но их оплодотворяющая способность сохраняется). • Травмы яичек. • Никотин и алкоголь (мешают вызреванию сперматозоидов). Активность сперматозоидов резко снижается у мужчин, которые много курят и злоупотребляют спиртным. • Стресс (кортизол, выделяемый во время стресса, снижает уровень тестостерона). 52 Болотов А. В. Биология размножения и развития • Перегревание (горячая ванна, сауна, баня, слишком теп- лое или тесное белье). • Гиподинамия, длительная сидячая работа. • Избыточный вес тела оказывают негативное влияние на созревание сперматозоидов. • Неправильно подобранное сиденье (стул, велосипедное сиденье) (нарушение кровообращения в половых органах). • Перегрузки во время физических тренировок. При усиленных тренировках в крови снижается уровень мужского полового гормона тестостерона. • Жирное мясо (снижает эффективность тестостерона). • Кофеин (задерживает образование сперматозоидов и нарушает их метаболизм). • Недостаток витамина С и минерала цинка отрицательно влияет на подвижность сперматозоидов. Чтобы возместить нехватку столь полезных веществ необходимо съедать в день пару апельсинов, блюдо из свежей или квашеной капусты, овсяную кашу, морепродукты, кусочек мяса, горсть грецких орехов. • Анаболические стероиды, а также антибиотики и противобактериальные препараты. • Болезни мочеполовой сферы (простатит, хламидиоз, венерические заболевания), детские инфекции (краснуха, свинка), протекавшие с осложнениями, сахарный диабет, вирусный гепатит. Из-за них «качество» сперматозоидов значительно страдает. • Нарушение проходимости семявыносящего протока. Препятствиями на пути сперматозоидов могут быть врожденное сужение участка семявыносящего тракта, спайка, оставшаяся после воспалительного или инфекционного заболевания, рубец после операции, киста или опухоль. • Некоторые заболевания иммунной системы. При них организм начинает вырабатывать антитела к собственным клеткам яичек, воспринимая их как «чужаков». Заболевание встречается редко. Семенной канатик Семенной канатик (funiculus spermaticus) состоит из семявыносящего протока (рис. 8, 11), артерии и вен яичка, лимфатических сосудов и нервов, покрытых оболочками. Он идет от заднего края яичка до внутреннего кольца пахового канала, где сосуды и семявыносящий проток расходятся. Семенной канатик III. Половая система млекопитающих 53 образуется после опускания яичка из брюшной полости, где оно первоначально развивается, в мошонку. К моменту рождения оба яичка находятся в мошонке. Отсутствие яичек в мошонке называется крипторхизмом. Половой член Половой член (penis) состоит из двух пещеристых тел и губчатого тела. Губчатое тело лежит под пещеристыми и пронизано мочеиспускательным каналом. Задняя часть полового члена называется корнем и прикреплена связками к лобковым костям. Передняя часть – головка полового члена утолщена и закруглена. Промежуточная часть – тело полового члена, верхняя поверхность его называется спинкой. На головке полового члена находится наружное отверстие мочеиспускательного канала. Кожа полового члена тонкая, у основания головки образует складку – крайнюю плоть. На внутренней поверхности крайней плоти расположены сальные железы, секрет которых входит в состав препуциальной смазки. Кнутри от рыхлой подкожной клетчатки фасция охватывает все три тела, соединяя их в единое целое. Наружной оболочкой пещеристых тел является фиброзная белочная оболочка, от которой внутрь отходят многочисленные перекладины. Промежутки между ними заполнены кровью. Величина полового члена изменяется в зависимости от количества крови в пещеристой ткани. Губчатое тело образует два утолщения: головку полового члена и его луковицу (сзади) (рис. 7, 6). Мужской мочеиспускательный канал Мужской мочеиспускательный канал (urethra masculina) (рис. 8, 10) представляет собой трубку длиной около 18 см, идущую от мочевого пузыря до наружного отверстия мочеиспускательного канала на головке полового члена. Он служит не только для выведения мочи, но и для прохождения семени, которое поступает в него из семявыбрасывающих протоков. В мочеиспускательном канале выделяют три части: предстательную, перепончатую и губчатую. Предстательная часть длиной около 3 см, наиболее широкая, является начальным отделом мочеиспускательного канала и проходит через предстательную железу. На задней стенке имеется небольшое возвышение – семенной холмик. Здесь открываются семявыбрасывающие протоки (рис. 8, 3), а по 54 Болотов А. В. Биология размножения и развития бокам холмика – протоки предстательной железы. По окружности начального отдела мочеиспускательного канала располагаются гладкие мышечные клетки, образующие его внутренний (пузырный) непроизвольный сфинктер. Перепончатая часть начинается от предстательной железы и тянется до луковицы полового члена. Это узкая и короткая (около 1 см) часть мочеиспускательного канала, проходящая через мочеполовую диафрагму. Здесь располагается сфинктер мочеиспускательного канала – произвольный. По сторонам от канала и сзади от него лежат бульбоуретральные железы (рис. 8, 7). Рис. 8. Придаточные половые железы мужской половой системы (по Борзяк, Бочарову, Волковой, 1986). 1 – мочевой пузырь; 2 – семенные пузырьки; 3 – семявыбрасывающий проток; 4 – перепончатая часть мочеиспускательного канала, 5 – ножки полового члена (пещеристая ткань); 6 – луковица полового члена; 7 – бульбоуретральные (куперовы) железы; 8 – простата; 9 – ампула семявыносящего протока; 10 – урета; 11 – семявыносящий проток Губчатая часть мочеиспускательного канала наиболее длинная (около 15 см), окружена тканью губчатого тела полового члена. В головке полового члена канал образует расширение – III. Половая система млекопитающих 55 ладьевидную ямку и заканчивается наружным отверстием. Эпителий, выстилающий слизистую оболочку мочеиспускательного канала, в различных участках разный (переходный, многорядный цилиндрический, однослойный цилиндрический и многослойный плоский). В канал открывается множество желез (железы Литтре). Кнаружи лежит мышечный слой, состоящий из неисчерченных мышечных клеток, расположенных продольно и циркулярно. Мочеиспускательный канал S-образно изогнут. При поднимании губчатой части передняя кривизна выпрямляется и заметна лишь задняя, более фиксированная. Семенные пузырьки Семенные пузырьки (vesicuiae seminales) – парные органы, расположенные латерально от семявыносящих протоков (рис. 8, 2), между мочевым пузырем и прямой кишкой. Длина каждого пузырька около 5 см. Книзу он суживается и переходит в узкий выделительный проток, который, соединяясь под острым углом с семявыносящим протоком, образует семявыбрасывающий проток. Он проходит сквозь толщу предстательной железы и открывается в предстательную часть мочеиспускательного канала на семенном холмике. Бульбоуретральные (куперовы) железы Бульбоуретральные (куперовы) железы (glandulae bulbourethraies) парные, величиной с горошину, плотные, располагаются в толще мочеполовой диафрагмы (рис. 8, 7). Железа имеет альвеолярно-трубчатое строение, плотную консистенцию, овальную форму, диаметр 0,3–0,8 см. Выводные протоки длиной 3–4 см открываются в губчатой части мочеиспускательного канала. Железа вырабатывает вязкую тягучую жидкость, которая защищает слизистую оболочку стенки мочеиспускательного канала от раздражения ее мочой. Предстательная железа Предстательная железа (prostata) – железисто-мышечный орган, охватывающий начальный отдел мочеиспускательного канала (рис. 8, 8). По форме и величине напоминает каштан. В ней различают основание, обращенное к мочевому пузырю, и верхушку, примыкающую к мочеполовой диафрагме. Передняя поверхность более выпуклая, обращена к лобковому симфизу. 56 Болотов А. В. Биология размножения и развития Задняя поверхность прилежит к прямой кишке и разделена бороздой на две доли – правую и левую. Участок железы, расположенный между мочеиспускательным каналом и правым и левым семявыбрасывающими протоками, называют перешейком (средняя доля). Средняя доля представляет значительный практический интерес, так как увеличение ее может нарушать мочеиспускание. Предстательная железа окружена фациальными листками, образующими капсулу. В этом вместилище находятся также клетчатка и венозное сплетение. Связками предстательная железа фиксирована к тазовому дну. Ткань ее состоит главным образом из желез и гладких мышечных клеток. Протоки желез (20–30) открываются на задней стенке мочеиспускательного канала по сторонам от семенного холмика. Секрет предстательной железы является составной частью спермы. Закладка яичек у человека появляется на 28–30-е сутки эмбриогенеза в виде парных утолщений целомического эпителия на медиовентральной поверхности первичных почек (мезонефросов). Мужская гонада начинает дифференцироваться раньше женской. 2. Женская половая система Женские половые органы делятся на наружные и внутренние. Наружные половые органы состоят из больших и малых половых губ, входа во влагалище, клитора, передней и задней спаек, лобка (бугорок Венеры), промежности и молочных желез. Внутренние половые органы состоят из половых желез или яичников (ovarii), яйцеводов или фаллопиевых труб (salpinx), матки (uterus) и влагалища (vagina). Яичник Наибольшая сложность строения у яичника. Это динамичный орган, в котором происходят постоянные изменения, связанные с гормональным статусом. Имеет среднем длину 2,5 см, ширину 1,5 см и толщину 1–1,5 см. Вес 5–8 г. Правый яичник обычно бывает больше и тяжелее левого. Типичное нормальное положение яичника – лежит у боковой стенки малого таза, где свободно и подвижно соединен собственной связкой с маткой. На другом конце он соединен с боковой стенкой таза (воронкотазовой связкой). На топографию яичника влияет его величина, III. Половая система млекопитающих 57 длина и растяжимость собственной связки, беременность матки, степень наполнения кишечника. Развивается из материала полового валика, который закладывается на 4 неделе эмбриогенеза на медиальной поверхности почек. Он образован целомическим эпителием (из висцерального листка спланхнотома) и мезенхимы. Это индифферентная стадия развития (без половых различий). Специфические различия наступают на 7–8 неделе. Этому предшествует появление в области полового валика первичных половых клеток – гоноцитов. Они содержат много гликогена в цитоплазме – высокая активность щелочной фосфатазы. Из стенки желточного мешка гоноциты по мезенхиме или с током крови попадают в половые валики, встраиваются в эпителиальную пластинку. С этого момента развитие женских и мужских половых желез различается. Образуются яйценосные шары – образования, состоящие из нескольких оогоний, окруженных одним слоем плоских эпителиальных клеток. Затем тяжи мезенхимы разделяют эти шары на более мелкие. Формируются примордиальные фолликулы, состоящие из одной половой клетки, окруженной одним слоем плоских фолликулярных эпителиоцитов. Несколько позже формируется корковое и мозговое вещество. В эмбриональном периоде в яичнике заканчивается период размножения овогенеза и начинается стадия роста, являющаяся самой продолжительной (несколько лет). Оогоний превращается в ооцит первого порядка. Белочная оболочка яичника, соединительнотканная строма, интерстициальные клетки дифференцируются из окружающей мезенхимы. Строение яичника взрослого организма в репродуктивный период Функции: эндокринная и репродуктивная. С поверхности покрыт мезотелием, под которым располагается оболочка, образованная плотной соединительной тканью – белочной оболочкой. Под ней лежит корковое вещество, а в центре – мозговое. Мозговое вещество образовано рыхлой соединительной тканью, в которой имеются химосные клетки, продуцирующие гормоны – андрогены. В мозговом веществе большое количество кровеносных, лимфатических сосудов и нервных элементов. Основу (строму) коркового вещества образует рыхлая соединительная 58 Болотов А. В. Биология размножения и развития ткань. В строме в большом количестве располагаются различные фолликулы, желтые и белые тела на разных стадиях развития. В течение репродуктивного периода в яичнике происходит рост овоцита первого порядка в фолликуле. Фолликулы созревают. Последовательные стадии развития фолликула: самый молодой (их очень много 30 000–400 000) – примордиальный фолликул, образованный ооцитом первого порядка, вокруг которого расположен один слой плоских фолликулярных эпителиоцитов, которые выполняют защитную и трофическую функции. Фолликулы располагаются на периферии. На разных стадиях онтогенеза происходит гибель женских половых клеток – атрезия. Первичные фолликулы (рис. 9, 2). Половые клетки становятся несколько больше. На периферии ооцитов первого порядка особая оболочка – блестящая. Вокруг него располагается один слой кубических или призматических фолликулярных эпителиоцитов. Прозрачная (блестящая) оболочка (рис. 10, 3) образована гликопротеидами. В образовании ее принимают участие ооцит первого порядка. В блестящей оболочке находятся радиально расположенные поры, в которые проникают микроворсинки ооцита и цитоплазматические отростки фолликулярных эпителиоцитов. Рис. 9. Схематическое изображение микроскопического строения яичника: 1 – примордиальные фолликулы; 2 – преантральные фолликулы; 3 – строма яичника; 4 – антральный фолликул; 5 – атретический фолликул; III. Половая система млекопитающих 59 6 – зрелый фолликул (Граафов пузырёк); 7 – овуляция; 8 – формирующееся желтое тело; 9 – зрелое желтое тело; 10 – яичковый эпителий; 11 – беловатое тело; 12 – кровеносные сосуды в воротах яичника Вторичные фолликулы (рис. 9, 4). Их образование уже связано с гормональным фоном (воздействие ФСГ). Под его влиянием фолликулярные эпителиоциты начинают усиленно делиться. Вокруг ооцита первого порядка формируется многослойный фолликулярный эпителий. Образование вторичных фолликулов наступает в период полового созревания. Фолликулярный эпителий синтезирует фолликулярную жидкость, которая содержит эстрогены. Образуется полость – пузырчатый фолликул, который постепенно преобразовывается в третичный фолликул. Третичный фолликул. Имеет сложноустроенную стенку, содержит ооцит первого порядка. Стенка состоит из двух частей: Рис. 10. Яйцеклетка человека (по Алмазову И. В, Сутулову Л. С., 1978): 1 – яйценосный бугорок с яйцевой клеткой (ооцитом); 2 – клетки фолликулярного эпителия, окружающие яйцеклетку (гранулеза); 3 – блестящая оболочка яйцевой клетки; 4 – лучистый венец; 5 – цито- 60 Болотов А. В. Биология размножения и развития плазма с желточными зернами; 6 – ядро с ядрышком; 7 – соединительная ткань (тека) 1. Многослойный фолликулярный эпителий – зернистый слой (гранулеза) (рис. 10, 2). Располагается на хорошо выраженной базальной мембране (стекловидная мембрана Славянского). 2. Соединительнотканная часть – тека (покрышка) (рис. 10, 7). В зрелом фолликуле (рис. 10) выделяют 2 слоя: • внутренний рыхлый (большое количество кровеносных сосудов, особые гормонально-активные клетки – текоциты (разновидность интерстициальные клетки), продуцирующие эстрогены. Являются источником образования опухолей). • фиброзный слой (плотный). Состоит из волокон. Полость фолликула заполнена фолликулярной жидкостью, которая содержит эстрогены, гонадокринин (гормон белковой природы, синтезируется фолликулярными клетками. Ответственен за атрезию фолликула). На одном из полюсов есть яйценосный бугорок или холмик (рис. 10, 1), на котором лежит ооцит первого порядка, окруженный лучистым венцом (рис. 10, 4). При образовании ЛГ происходит разрыв фолликула и выход из яичника половой клетки, т. е. происходит овуляция. Половая клетка устремляется в яйцевод, где происходит деление и созревание ее. На месте лопнувшего фолликула происходит образование желтого тела (рис. 9, 8–9). Его клетки продуцируют гормон прогестерон. Различают два вида желтых тел – менструальное и желтое тело беременности. Менструальное тело меньших размеров (1–2 см в диаметре в то время как желтое тело беременности 5–6 см), продолжительность жизни его меньше (5–6 дней против нескольких месяцев). Все преобразования в фолликуле называются овариальным циклом. Он происходит под влиянием гормонов в две фазы: • фолликулиновая фаза (под влиянием ФСГ); • лютеиновая (под влиянием ЛГ, ЛТГ). Изменения в яичниках вызывают изменения в других органах женской половой системы – яйцеводах, матке, влагалище, молочных железах. Матка III. Половая система млекопитающих 61 Матка (uterus) – полый непарный орган (рис. 11, А), в котором происходит развитие зародыша и вынашивание плода. В ней различают дно (рис. 11, А) – верхняя часть, тело – средний отдел и шейку – нижняя суженная часть. Матка имеет переднюю и заднюю поверхности. Передняя поверхность матки обращена к мочевому пузырю и называется пузырной, задняя, обращенная к прямой кишке – кишечной. Размеры матки и ее масса варьируют. Длина матки у взрослой женщины в среднем составляет 7–8 см, ширина 4 см, а толщина 2–3 см. Объем полости матки находится в пределах 4–6 см3. Расположена в полости малого таза между прямой кишкой и мочевым пузырем. Матка это мышечный орган. Имеет три оболочки – слизистая (эндометрий), мышечная (миометрий), серозная (периметрии). Эпителий слизистой дифференцируется из мезонефрального протока. Соединительная ткань, гладкомышечная ткань – из мезенхимы. Мезотелий из висцерального листка спланхнотома. Эндометрий образован однослойным призматическим эпителием и собственной пластинкой. В эпителии два вида клеток: реснитчатые эпителиоциты и секреторные эпителиоциты. Собственная пластинка образована рыхлой волокнистой соединительной тканью, в ней находятся многочисленные маточные железы (многочисленные трубчатой формы, выпячивания собственной пластинки – крипты) (рис. 11, Б, а). Их количество, размеры, глубина, активность секреции зависят от фазы овариальноменструального цикла. В эндометрии (рис. 11, А, Б) различают два слоя: глубокий базальный (образован глубокими участками эндометрия) и функциональный. Миометрий (рис. 11, А; Б, 2) образован гладкой мышечной тканью и состоит из 3 слоев: • подслизистый слой миометрия (косое расположение) (рис. 11, Б, б); • сосудистый слой (в нем располагаются крупные кровеносные сосуды) – косопродольное направление (рис. 11, Б, в); • надсосудистый слой (косопродольное направление, противоположное направлению миоцитов сосудистого слоя) (рис. 11, Б, г). 62 Болотов А. В. Биология размножения и развития Рис. 11. Внутренние женские половые органы, оплодотворение и развитие зародыша (А). Слои матки (Б). х 70. Окраска гематоксилинэозином (по Алмазову И. В, Сутулову Л. С., 1978): 1 – слизистая оболочка (эндометрий); а – маточные железы (крипты); 2 – мышечная оболочка (миометрий); б – подслизистый слой мышечной оболочки; в – сосудистый слой мышечной оболочки; г – надсосудистый слой мышечной оболочки; 3 – серозная оболочка (периметрий) III. Половая система млекопитающих 63 Состав миометрия зависит от гормона эстрогена (при его недостатке развивается атрофия). Прогестерон вызывает гипертрофические изменения. Периметрий (рис. 11, А, Б, 3). Образован двумя тканями: пластинкой гладкомышечной ткани и однослойным плоским эпителием целомического типа – мезотелием. Женская половая система характеризуется цикличностью строения и функций, которая определяется гормонами. Изменения в яичниках и матке – овариально-менструальный цикл. Продолжительность в среднем 28 дней. Весь период подразделяется н а т р и фазы: • менструальная (с первого дня менструации); • постменструальный (пролиферации); • предменструальный (секреции). Менструальная фаза примерно четыре дня. В течение этого времени происходит десквамация (гибель) тканей слизистой оболочки матки, отторжение их, а затем регенерация эпителия. Отторжение всего функционального слоя до самых глубоких участков до крипт. Пролиферация – изменение эпителия, восстановление функционального слоя эндометрия, структурное оформление маточной железы. Происходит восстановление спиральных артерий примерно 5–14 суток. На 14-е сутки происходит овуляция. Под влиянием прогестерона происходит увеличение эндометрия до 7 мм (вместо 1 мм), он становится отечным, маточная железа приобретает штопорообразный вид. Просвет переполнен секреторными продуктами, спиралевидные артерии удлиняются, закручиваются. После 23–24 суток сосуды спазмируются. Развивается ишемия и гипоксия тканей. Они некротизируются и все начинается сначала. Шейка матки Шейка матки представляет собой нижнюю суженную часть и имеет вид мышечного цилиндра (рис. 11). В центре шейки проходит шеечный, или цервикальный канал, который начинается в полости тела матки внутренним зевом. Дистальная часть шейки матки выступает во влагалище и заканчивается наружным зевом. Шейка матки состоит из тех же оболочек, что и тело. 64 Болотов А. В. Биология размножения и развития Канал шейки матки выстлан однослойным призматическим эпителием, который в области дистальной (влагалищной) части шейки соединяется с многослойным плоским неороговевающим эпителием. Последний продолжается в эпителий слизистой оболочки влагалища. Разница между многослойным и однослойным призматическим эпителием слизистой оболочки всегда четкая и ориентировочно располагается на уровне дистальной части шейки. Маточная труба (яйцевод) Яйцевод (фаллопиева труба) это парный трубчатый орган (рис. 11), дистальный конец которого имеет вид воронки, открыт и контактирует с поверхностью яичника, а проксимальный прободает стенку матки в области боковых поверхностей ее дна и сообщает трубы с маточной полостью (рис. 11). У человека длина маточных труб составляет около 10–12 см, диаметром 2-4 мм. Маточные трубы захватывают ооцит при овуляции, осуществляют eгo тpaнспорт по направлению к полости матки, создают условия для беспрепятственного продвижения спермиев навстречу ооциту, обеспечивают среду, необходимую для оплодотворения и дробления зародыша, транспортируют зародыш в полость матки. Маточные трубы развиваются из верхней части парамезонефральных (мюллеровых) протоков. Маточная труба подразделяется на четыре отдела: воронку – дистальный отдел трубы, заканчивающийся бахромками (фимбриями) (рис. 11) и открывающийся в яичниковую сумку; ампулу – следующую за воронкой наиболее широкую и протяженную часть (составляет около 2/3 длины трубы); перешеек, или истмус; и интерстициальный (интрамуральный) отдел, прободающий стенку матки. Влагалище Влагалище (vagina) – непарный полый орган в виде трубки длиной 8–10 см, толщина стенок равна 3 мм. Верхним концом оно охватывает шейку матки, а нижним через мочеполовую диафрагму таза открывается в преддверие отверстием влагалища (рис. 11). Это отверстие у девственницы закрыто девственной плевой, представляющей собой полулунную или продырявленную пластинку, которая во время полового акта разрывается, а ее лоскуты потом атрофируются. Впереди влагалища находятся III. Половая система млекопитающих 65 мочевой пузырь и мочеиспускательный канал, сзади – прямая кишка, с которой оно срастается рыхлой и плотной соединительной тканью. Во влагалище выделяют переднюю и заднюю стенки, которые соединяются одна с другой. Охватывая влагалищную часть шейки матки, они образуют вокруг нее куполообразное углубление – свод влагалища. Молочные железы Представляют собой измененные потовые железы с апокриновым типом секреции. Железистая ткань имеет эктодермальное происхождение. Дифференцировка начинается на четвертой неделе. Вдоль передней части туловища образуются продольные утолщенные линии, из которых и образуются железы. Строение до и после полового созревания резко различается. Молочные железы взрослой женщины состоят из 15–20 отдельных желез, которые имеют альвеолярно-трубчатое строение. Каждая железа образует долю, между которыми находится прослойка соединительной ткани (рис. 12). Каждая доля состоит из отдельных долек, между которыми располагаются прослойки соединительной ткани, богатые жировыми клетками (рис. 12). Молочная железа состоит из секреторных отделов (альвеолы или ацинусы) и системы выводных протоков (рис. 12). Рис. 12. Строение молочной железы 66 Болотов А. В. Биология размножения и развития В нелактирующей железе имеется большое количество протоков и очень мало секреторных отделов. До момента полового созревания в молочной железе нет концевых отделов. В лактирующей молочной железе альвеолы многочисленны. Каждая из них образована железистыми клетками (кубические лактоциты) и миоэпителиоцитами. Лактоциты продуцируют секрет – молоко. Это водная эмульсия триглицеридов, глицерина, лактоальбуминов, глобулинов, солей, лактозы, макрофагов, Т и Влимфоцитов, иммуноглобулинов А (которые предохраняют ребенка от кишечных инфекций). Белки выделяются из железистых клеток по мерокриновому типу, а жиры по апокриновому. В конечный период беременности происходит образование и накопление секрета – молозива. Оно отличается более высоким содержанием белка, чем жира. А у молока наоборот. Последовательность протоков: альвеолы – альвеолярные млечные ходы (внутри долек) – внутридольковые протоки (выстланные более высоким эпителием и миоэпителиоцитами) – междольковый проток (в прослойке соединительной ткани). Вблизи соска они расширяются и называются молочными синусами. Деятельность лактоцитов определяется пролактином. Выделению молока способствуют миоэпителиоциты. Их деятельность регулирует гормон окситоцин. Недавние исследования (2011) показали, что в грудном молоке содержатся стволовые клетки, число которых составляет около 2 %. Их доля в нем варьирует в зависимости от того, насколько полна молоком грудь женщины и насколько долго оно производится. IV. ОСНОВНЫЕ ЭТАПЫ ЭМБРИОНАЛЬНОГО РАЗВИТИЯ Деление, рост и дифференцировка оплодотворенного яйца, в результате которых и образуется такая сложная и совершенная система, как взрослое животное или растение. В каждом новом организме с необычайной точностью воспроизводятся все детали строения взрослого организма, а многие органы начинают функционировать еще в период своего формирования. Например, сердце человеческого эмбриона (в конце первого месяца). Процессы дробления яйца, образование бластулы и гаструляция происходят в самых общих чертах одинаково почти у всех многоклеточных животных. А вот дальнейшее развитие может протекать по-разному у животных, принадлежащих к разным типам, но сходно у родственных между собой форм. Все общие процессы, которые идут сходно у разных организмов в процессе развития организма можно разбить на шесть этапов (стадий): 1. Гаметогенез. Развитие новой особи начинается с образования яйцеклеток и сперматозоидов у представителей родительского поколения. 2. Оплодотворение – слияние женской и мужской половых клеток. В результате образуется новый одноклеточный организм – зигота. 3. Дробление. Серия быстро следующих друг за другом делений зиготы, при этом цитоплазма зиготы разделяется (дробится) на множество мелких клеток. Эта стадия заканчивается образованием многоклеточного зародыша и образование бластулы (рис. 13). 4. Гаструляция. В результате деления, дифференцировки, взаимодействия и перемещения клеток зародыш становится многослойным. Появляются зародышевые листки эктодерма, энтодерма и мезодерма, несущие в себе закладки различных тканей и органов (рис. 13). Из эктодермы формируются: наружный эпителий (эпидермис), кожные железы, поверхностный слой зубов, рецепторные 68 Болотов А. В. Биология размножения и развития элементы, впячивание передней и задней кишки, первичные половые клетки (млекопитающие), нервная система. Из мезодермы формируются: поперечно-полосатая скелетная ткань, сердечная ткань, гладкая мышечная ткань, выстилка целомической полости, органы размножения и выделительная система, костная и хрящевые ткани, кровь и лимфа. Из энтодермы формируются: выстилка желудочно-кишечного тракта, пищеварительные железы, железы внутренней секреции, легкие. 5. Органогенез. В ходе дифференцировки трех зародышевых листков клеточные пласты делятся на группы клеток меньшего объема, предназначенных для построения различных органов и частей взрослого организма. Каждый орган закладывается в виде эмбрионального зачатка почки – группы клеток, обособленных от остальных клеток зародыша. Из них начинают формироваться органы и системы организма (рис. 13). Рис. 13. Этапы развития животного 6. Рост и дифференцировка клеток в зачатках отдельных органов. В результате этого клетки приобретают те структурные, химические и физиологические особенности, благодаря которым они могут выполнять те или иные функции так, что развивающийся организм становится способным к независимому существованию. V. ГАМЕТОГЕНЕЗ Первым шагом развития нового организма, т. е. началом собственного эмбриогенеза, принято считать момент оплодотворения яйцеклетки сперматозоидом или же ее активацию какимлибо другим агентом. Но перед тем как стать способной к оплодотворению, яйцеклетка проходит длинный путь развития, начинающийся, как правило, еще в раннем развитии будущей половозрелой самки. Аналогичный путь проходит и сперматозоид. Процессы развития половых клеток принято называть гаметогенезом, или предзародышевым развитием; применительно к яйцеклетке говорят об оогенезе (овогенезе), а развитие сперматозоида называют сперматогенезом. Сперматозоиды и яйцеклетки обычно формируются соответственно особями мужского и женского пола. Биологические виды, у которых все организмы делятся в зависимости от производимых ими половых клеток на самцов и самок, называются раздельнополыми. Встречаются виды, у которых один и тот же организм может образовывать как мужские, так и женские половые клетки. Такие организмы называются гермафродитами (в греческой мифологии гермафродит – дитя Гермеса и Афродиты – обоеполое существо, несущее в себе и женское, и мужское начало). Гермафродитизм наблюдается у многих беспозвоночных животных (моллюсков, плоских и кольчатых червей), а также у круглоротых (миксины) и рыб (морской окунь). Классическим примером такого гермафродитизма являются гольяны (род Poeciliopsis) и европейский серебряный карась Carassius gibelio. В этом случае организмы, как правило, имеют ряд приспособлений, препятствующих самооплодотворению. У некоторых моллюсков половая железа продуцирует попеременно мужские и женские половые клетки. Это зависит от условий существования особи и ее возраста. У большинства низших животных гаметы вырабатываются в течение всей жизни, у высших – только в период половой активности, с момента полового созревания до затухания деятельности желез в старости. 70 Болотов А. В. Биология размножения и развития 1. Происхождение первичных половых клеток Все современные многоклеточные в ходе своего развития рано или поздно разделяются на генеративную часть (половые клетки) и соматическую часть, из которой развиваются все остальные органы. Такое разделение было важнейшим эволюционным событием, которое и обусловило переход от одно- к многоклеточности и сделало возможным сам процесс онтогенеза, сводящийся главным образом к прогрессивному усложнению и специализации соматической части организма. У многих животных половые клетки обособляются от соматических очень рано. Особенно характерно это для членистоногих, круглых червей, щетинкочелюстных и некоторых других беспозвоночных. Так, у двукрылых насекомых еще до начала дробления яйцеклетки в цитоплазме ее заднего полюса обнаруживаются особые базофильные гранулы, названные половыми детерминантами. Половые клетки обособляются впоследствии именно из этого участка цитоплазмы, независимо от того, какое именно клеточное ядро попадет в этот участок при дроблении. Если этот участок цитоплазмы разрушить ультрафиолетовыми лучами, возникают стерильные особи без половых клеток. У дрозофилы окончательное обособление половых клеток от соматических происходит на 13-м делении дробления. В яйцеклетке веслоногого рака циклопа также присутствуют базофильные гранулы (эктосомы), которые по ходу делений дробления всегда попадают лишь в одну из двух разделившихся клеток. В конце концов, эктосомы распределяются между двумя клетками, которые и дают начало половым. В данном случае окончательное обособление гоноцитов от соматических клеток происходит уже на 5-м делении дробления. Еще раньше (на 4-м делении дробления) это обособление происходит у ветвистоусых раков, а также у лошадиной аскариды. У позвоночных первичные гоноциты обособляются позже, но почти всегда задолго до окончания эмбрионального развития будущего организма. Так, у бесхвостых амфибий еще до начала делений дробления яйцеклетки на ее вегетативном (богатом желтком) полюсе обнаруживаются, как и у членистоногих, зерна половой цитоплазмы, которые затем перемещаются и концен- V. Гаметогенез 71 трируются в клетках внутренних областей зародыша, поблизости от так называемого бластоцеля. Окончательное обособление первичных гоноцитов происходит в этой группе животных на стадии поздней бластулы, когда зародыш насчитывает несколько сотен клеток, но основные зачатки органов еще не обособились. Значительно позже происходит обособление первичных гоноцитов у хвостатых амфибий. В противоположность бесхвостым амфибиям этот процесс идет не автономно, а под влиянием других, соседних эмбриональных тканей. Гоноциты хвостатых амфибий возникают из среднего зародышевого листка (мезодермы) под воздействием внутреннего листка (энтодермы). Это воздействие осуществляется на стадии бластулы, само же обособление гоноцитов на стадии гаструлы или даже нейрулы, т. е. в период закладки других зачатков органов. У птиц, по современным данным, первичные половые клетки возникают на ранних стадиях развития зародыша из наружного листка (эпибласта), а затем мигрируют во внутренний листок (гипобласт), в составе которого располагаются у переднего конца зародыша, в области так называемого головного серпа. Возможно, что и у млекопитающих первичные половые клетки возникают из наружного листка зародыша. Позже они перемещаются к так называемому устью желточного мешка. У зародыша человека окончательное обособление гоноцитов от соматических клеток происходит к 56-му дню развития; к этому времени сформированы зачатки основных органов. Половые клетки у круглых червей, членистоногих, позвоночных – первичные гоноциты, обособляющиеся от клеток соматических на ранних стадиях развития. Иными словами, дифференцировка клеток зародыша на половые и соматические сопровождается у данных групп животных также и их необратимой детерминацией. Однако далеко не у всех групп животных гоноциты обособляются исключительно на ранних стадиях эмбрионального развития и не могут позже пополняться за счет соматических клеток. У низших многоклеточных – губок, стрекающих кишечнополостных, мшанок, а также у некоторых кольчатых червей и полухордовых имеются особые тотипотентные стволовые клетки (способные трансформироваться в любой тип клеток данного 72 Болотов А. В. Биология размножения и развития вида), которые в течение всей жизни организма пополняют запас его половых клеток. Таким образом, у этих организмов линии половых и соматических клеток практически неотделимы. У плоских и кольчатых червей стволовые клетки, называемые необластами, присутствуют во многих тканях. Поэтому половые клетки могут у них возникать и в случае регенерации из небольших участков тела взрослых животных при полном удалении половых желез. У продолжительно голодающих планарий (плоские черви) половые клетки могут дедифференцироваться и превращаться в стволовые клетки, используемые для регенерации соматических тканей. Впоследствии из них могут вторично возникать половые клетки. Вместе с тем у многих кольчатых червей имеется и рано обособляющаяся линия эмбриональных гоноцитов. Таким образом, у них существуют два источника половых клеток; раннеэмбриональный и соматический. Важно напомнить также, что почти у всех растений (за исключением зеленых водорослей) линии половых и соматических клеток фактически неотделимы на протяжении всей жизни: половые клетки выделяются из соматических только при формировании мужских и женских гаметофитов. Каков бы ни был источник половых клеток – раннеэмбриональные первичные гоноциты или постоянно существующие резервные клетки, они должны пройти длинные пути превращений, прежде чем стать зрелыми, способными к оплодотворению половыми клетками. 2. Состав, развитие и особенности половых клеток Половые, или зародышевые, клетки (гаметы) представляют собой особые клеточные протоплазматические образования, из которых после их слияния во время оплодотворения начинает развиваться новый индивидуум. Половые клетки делятся на мужские (спермии, сперматозоиды) и на женские (яйцеклетки, овули). При оплодотворении обе половые клетки сливаются в одно общее клеточное целое (оплодотворенное яйцо, spermovium, или зигота). Процесс оплодотворения одновременно служит для зиготы импульсом к интенсивному клеточному делению V. Гаметогенез 73 (сегментации, или дробящему делению), которое является первым шагом в развитии нового организма. В соответствии с основным биогенетическим законом в живом веществе половых клеток сконцентрирована совокупность свойств, являющаяся результатом длительного процесса филогенетического развития соответствующего вида животного и всего животного мира в целом. Таким образом, эта совокупность возникает как результат влияний внутренней и внешней среды на организмы во время их филогенеза и представляет собой основу наследственности будущей особи, зафиксированную, прежде всего в ядерном веществе половых клеток. Половые клетки, из которых развивается новый организм, являются биологически наиболее сложными клетками; они характеризуются максимальными возможностями развития, в то время как все остальные клетки, возникающие из оплодотворенного яйца в процессе развития организма или же при дифференциации отдельных тканей, являются биологически менее сложными и имеют меньшие возможности развития (меньший проспективный потенциал). В связи с этим половые клетки можно считать завершением цикла развития организма, его концентрированием, а в филогенетическом отношении одновременно началом развития новых организмов. В половых клетках как бы кумулируется весь путь развития, который прошли организмы предыдущих поколений. Особенности половых клеток 1. Измененное ядерно-плазменное отношение по сравнению с соматическими клетками; 2. Гаплоидный наследственный материал (nc). 3. Измененный обмен веществ (половые клетки находятся как бы в состоянии депрессии). 4. Наличие специфических структур: акросомальный аппарат, хвостик для движения спермия, оболочки яйца, наличие кортикальных и желточных гранул. 5. Отсутствие ряда органелл в клетках: в сперматозоидах (аппарат Гольджи, эндоплазматический ретикулум, рибосомы) и яйцеклетках (центриоль). 6. Незаконченный жизненный цикл. 74 Болотов А. В. Биология размножения и развития 3. Строение половых клеток Сперматозоид Спермий, сперматозоид (мужская половая клетка) – зрелая мужская половая клетка – по своим морфологическим и физиологическим свойствам значительно отличается от женской половой клетки – яйца. Спермий был открыт в 1677 г. учеником Левенгука Л. Гаммом. Сам Левенгук считал спермий (бойко движущиеся образования с характерным хвостиком) примитивными зародышами, паразитирующими в семени и называл их «семенными зверьками». От него происходит и более старое, до сих пор иногда применяемое название мужской половой клетки – сперматозоон (от лат. animalculum – семенное животное). В соответствии с господствовавшими в то время представлениями, головка этого семенного животного содержала в миниатюре всю будущую особь. Термин «сперматозоид» ввёл в начале XIX в. Карл Эрнст фон Бэр. По своему виду спермий значительно отличается не только от яйцеклетки, но и от всех остальных клеток организма. В отличие от крупной и инертной яйцеклетки сперматозоиды значительно более мелкие, размер которых у разных видов животных, колеблется от 20 мкм (у крокодила), до 2 мм в длину (у амфибии); они активны и подвижны. Ядерно-плазменное отношение сильно смещено в сторону ядра, т. е. у них почти нет цитоплазмы. За счет уникального механизма конденсации хроматина (из ядра удаляются гистоны, и ДНК связывается с белками-протаминами), большая часть цитоплазмы выбрасывается из сперматозоида в виде так называемой «цитоплазматической капли», остаются только самые необходимые органеллы. Поэтому они утрачивают и некоторые органеллы: рибосомы, эндоплазматический ретикулум, аппарат Гольджи. В ядре сперматозоида человека содержится 23 хромосомы, одна из которых является половой (X или Y), остальные – аутосомами. Среди спермиев 50 % содержат Х-хромосому и 50 % – Y-хромосому. Показано, что масса Х-хромосомы больше массы Y-хромосомы, поэтому спермии, содержащие Х-хромосому, менее подвижны, чем содержащие Y-хромосому. Сперматозоиды, V. Гаметогенез 75 содержащие Y-хромосому, называются андроспермиями, Х-хромосому – гиноспермиями. В спермограмме здорового мужчины наряду с нормальными встречаются и патологические формы спермиев, но обычно не более 20–25 % (рис. 14, Б). Рис. 14. Разные виды сперматозоидов (А): 1 – человек; 2 – тритон; 3 – речной рак; 4 – морская свинка; 5 – хряк; 6 – бык; 7 – петух; 8 – ветвистоусый рак; 9 – десятиногий рак; 10 – лошадиная аскарида; 11 – острица. Схема строения зрелого сперматозоида человека (Б). Сперматозоиды человека нормальные (1) и аномальные (2) (В) Форма сперматозоидов очень разнообразна, но среди них можно выделить 2 главных типа: жгутиковые и безжгутиковые. Безжгутиковые формы сравнительно редки (круглые черви, высшие раки, моллюски), которые имеют причудливую форму (рис. 14, А). В жгутиковых сперматозоидах, как правило, присутствует головка, тело, шейка и хвостик (рис. 14, А). У сперматозоида человека, например, различают четыре основные части (рис. 14, Б): 76 Болотов А. В. Биология размножения и развития 1. Головка сперматозоида человека имеет форму эллипсоида, сжатого с боков, с одной из сторон имеется небольшая ямка, поэтому иногда говорят о «ложковидной» форме головки сперматозоида у человека (рис. 14, Б). Передняя часть головки покрыта тонким, прозрачным слоем плазмы, которая является довольно устойчивой и умеренно заостренной. У некоторых животных это приспособление, возможно, играет роль образования, облегчающего проникновение головки спермия в яйцеклетку во время, оплодотворения. В передней части головки расположен акросомальный аппарат (рис. 14, Б), состоящей из секреторного пузырька – акросомы (содержащий гидролитические ферменты, и позволяющий спермию проникнуть через наружные яйцевые оболочки). Акросома функционально соответствует лизосоме. Она развивается из аппарата Гольджи и содержит около 15 гидролитических ферментов – гиалуронидазу и несколько протеиназ, основным из который является трипсиноподобный акрозин – они осуществляют «переваривание» гранулярного слоя и плазматической мембраны ооцита непосредственно перед оплодотворением. Акросома лежит спереди от ядра и покрывает собой половину ядра (поэтому часто акросому сравнивают с шапочкой). При контакте с яйцеклеткой акросома выбрасывает свои ферменты наружу и растворяет небольшой участок оболочки яйцеклетки, благодаря чему образуется небольшой «проход» для проникновения сперматозоида. Почти все вещество головки – за исключением очень тонкого и почти невидимого цитоплазматического слоя на поверхности – состоит из ядерного вещества, очень сгущенного в этой части и интенсивно красящегося ядерными красками. В связи с сильной конденсацией хроматин неактивен – в ядре сперматозоида не синтезируется РНК. Иногда в ядерном веществе головки обнаруживается шаровидная вакуоль. Длина головки составляет приблизительно 4–5 мкм. 2. Шейка спермия (рис. 14, Б) представляет собой короткую, более узкую часть, образованную мягким, гомогенным плазматическим веществом. Сразу же за головкой в ней располагается проксимальная (передняя) центриоль, по мнению одних авторов, в виде двух или нескольких зернышек, по мнению других – в V. Гаметогенез 77 виде пластинки. Мягкое вещество шейки обусловливает сравнительную подвижность головки по отношению к хвосту и ее наклон под определенным, почти прямым углом. Длина шейки составляет приблизительно 0,5 мкм. 3. Соединяющая (средняя) (рис. 14, Б) часть спермия или тело отграничивается от шейки проксимальной частью дистальной (задней) центриоли в виде пластинки, которая, по мнению некоторых авторов, состоит из целого ряда мелких зернышек. Соединяющая часть оканчивается кольцеподобным образованием, исходящим из дистальной части дистальной центриоли. Посередине соединяющей части проходит осевая нить хвоста (рис. 14, Б), образованная тонкими фибриллами и исходящая, по мнению одних авторов, из передней центриоли, по мнению других – из задних зернышек. Осевая нить выходит из соединяющей части через кольцеподобное образование и проходит далее по хвосту спермия. В области соединяющей части она непосредственно покрыта цитоплазматическим слоем, вокруг которого несколько раз обвивается спиралевидное волоконце, образованное митохондриями цитоплазматического слоя (спираль) (рис. 14, Б). Длина соединяющей части составляет приблизительно 5–6 мкм; она считается двигательным центром хвоста. 4. Хвост, или жгутик (рис. 14, Б), представляет собой собственно продолжение осевой нити соединяющей, средней части. Осевая нить на отрезке длиной в среднем в 20–30 мкм покрыта тонким слоем цитоплазмы, а ее конец, длиной приблизительно в 5 мкм, обнажен и, постепенно утончаясь, заканчивается заострением. Таким образом, общая длина спермия колеблется от 50 до 60 мкм. Яйцеклетка (ovulum, или женская половая клетка) Зрелая яйцеклетка относится к наиболее крупным клеткам организма. Следовательно, яйцеклетка поставляет большую часть строительного материала, необходимого для развития нового организма. Яйцевые клетки разных видов животных и птиц сильно варьируют по размерам, при этом форма у них может быть либо шаровидная, либо овальная. Самое крупное яйцо обнаружено у китовой акулы, длина 60 см, а диаметр – 40 см. Достаточно крупные яйца у тигрового питона – 12 см в длину и с диаметром 78 Болотов А. В. Биология размножения и развития 6 см. Среди птиц самое крупное яйцо у страуса в длину 15–20 см, диаметром 15–18 см. Среди млекопитающих самые крупные яйца свойственны яйцекладущим – утконосу и ехидне. Диаметр яйца утконоса – 4,4 мм, ехидны – 3 мм. Зрелая яйцеклетка человека имеет примерно 150 мкм (0,15 мм) в диаметре, макака – резуса – 118 мкм, морской свинки – 76 мкм, кролика – 160 мкм, а мыши – 80 мкм. А самая мелкая яйцеклетка обнаружена у примитивного млекопитающего – полевки пашенной, размер которой составляет всего 40 мкм. Яйцевая клетка человека имеет правильную шаровидную форму. Величина живой яйцеклетки человека колеблется от 130 до 160 мкм, но несмотря на свои сравнительно большие размеры, она была открыта довольно поздно. Хотя о «яичке» упоминалось уже в литературе XVII в., однако в то время яйцеклеткой считали шаровидные, пузырчатые образования, хорошо видимые простым глазом в яичниках животных и человека и известные в настоящее время как овариальные фолликулы (Граафовы фолликулы – Регнерус де Грааф, 1677). Сама яйцевая клетка была открыта лишь в 1827 г. Карлом Бэром. Однако уже и другие авторы – современники Бэра – в первой половине XIX в. под различными названиями описывали некоторые самостоятельные составные части яйцеклетки. Так, Ян Пуркинье обнаружил ядро птичьей яйцеклетки и назвал его vesicula germinativa (1825), Рудольф Вагнер (1836) описал в нем ядрышко. Большую часть шаровидного клеточного тела яйцеклетки составляет цитоплазма, которая в данном случае называется ооплазмой. В живой яйцеклетке ооплазма представляет собой мягкое, прозрачное, зернистое вещество; более отчетливо зернистость видна в центральных отделах клетки, в области, расположенной вокруг ядра, в то время как более поверхностный слой ооплазмы или вообще не содержит никаких гранул, или же содержит их в малом количестве. Некоторые зернышки в ооплазме соответствуют зернистым митохондриям, которые обнаруживаются в яйцевой клетке при помощи соответствующих гистологических методов. Однако большую часть гранул составляет желтковое вещество. Объем яйца, как правило, превосходит объем спермия во много раз (у морского ежа их отношение составляет 10 000:1). Ядерные гаплоидные компоненты V. Гаметогенез 79 у них примерно равны, а вот по содержанию цитоплазматических компонентов различия велики. Цитоплазма яйца представляет собой огромную кладовую запасов, накопленных в период вителлогенеза. Эти запасы включают белки, рибосомы, транспортную и матричную РНК, морфогенетические факторы. Это молекулы, которые направляют процессы дифференцировки клеток, необходимые для образования различных тканей и органов. Они рассеяны по всему яйцу и распределяются между разными клетками при дроблении. Важной отличительной способностью яйцеклеток является то, что ядерно-плазменное отношение смещено в сторону цитоплазмы (рис. 15). Ядро яйцевой клетки – шаровидное, располагающееся чаще всего несколько эксцентрично (рис. 16, 1). В ядре яйцеклетки Рис. 15. Соотношение человека содержится 23 хромоядерного вещества к цито- сомы; одна из них является полоплазму у соматический (А) вой Х-хромосомой. клетки, яйцеклетки (Б) и На фиксированных и окрасперматозоида (В) шенных препаратах видно, что в ядре содержится мало хроматина, в связи с чем оно слабо окрашивается и имеет везикулярный (пузырчатый) вид. Хроматин носит волокнистый характер и в периферических отделах ядра образует более компактный слой, представляющий собой нечто наподобие ядерной оболочки. В ядре, как правило, находится крупное шаровидное псевдоядрышко. В зрелой яйцеклетке центриоль уже не содержится, поскольку она исчезает в процессе развития и созревания яйца. Все компоненты яйца располагаются в цитоплазме яйца ассиметрично, обеспечивая полярность яйца и эмбриона в будущем (рис. 16). То есть яйцо имеет два полюса: анимальный и вегетативный. Та часть яйца, ближе к которой располагается ядро или на котором выделяются редукционные тельца называется анимальным, а противоположный – вегетативным. А ось, которая проходит через эти два полюса называется анимально- 80 Болотов А. В. Биология размножения и развития вегетативной осью клетки, играющей большое значение в процессах дробления и дальнейшего развития организма. Эта анимально-вегетативная поляризация яйцеклетки решающим образом ориентирует последующие морфогенетические процессы: за редкими исключениями первые две борозды делений дробления оплодотворенной яйцеклетки проходят по взаимно перпендикулярным анимально-вегетативным меридианам, пересекаясь на аномальном и вегетативном полюсах, а у взрослых животных передне-задняя ось тела либо совпадает с анимальновегетативной осью яйцеклетки (так называемые протаксонные животные – многощетинковые черви, позвоночные и др.), либо перпендикулярна ей (плагиаксонные животные – малощетинковые черви, некоторые членистоногие). Рис. 16. Схема строения яйцеклетки: 1 – ядро; 2 – цитоплазма с желточными гранулами; 3 – кортикальные гранулы; 4 – микроворсинки на поверхности плазмолеммы; 5 – блестящая оболочка; 6 – зернистая оболочка. Первые морфологические проявления поляризации яйцеклетки приурочены к периоду вителлогенеза: у большинства яйцеклеток желток откладывается преимущественно в вегетативном полушарии, а ядро ооцита оттесняется в анимальное полушарие, где больше свободной цитоплазмы. Но только в период делений созревания, а точнее, в период выделения 2-го ре- V. Гаметогенез 81 дукционного тельца поляризация становится устойчивой и необратимой. Это доказывается следующим опытом, поставленным французским ученым Геррье на яйцеклетках моллюсков (виноградный слизень). Посредством центрифугирования яйцеклетки после выделения 1-го редукционного тельца веретено 2-го деления созревания смещалось со своего обычного положения и 2-го редукционное тельце выделялось не под 1-м тельцем, как обычно, а в другой точке поверхности яйцеклетки. Оказалось, что в последующем развитии таких яйцеклеток ось делений дробления, а отсюда и переднезадняя ось организма соответствовали точке выделенного не 1-го, а 2-го редукционного тельца. Материальные носители полярности яйцеклетки до сих пор полностью не выявлены. Судя по всему, они локализованы в плазматической мембране яйцеклетки, а не в цитоплазматических включениях (эти включения и особенно желток можно сместить путем центрифугирования в различные положения, а полярная ось сохранит после этого свою исходную ориентацию). Что касается мембранных структур, ответственных за полярность яйцеклетки, интересные данные получены в последние годы с применением электрофизиологических измерений: при помощи так называемых вибрирующих электродов вокруг ооцитов и яйцеклеток ряда животных и растений обнаружены электрические поля. Известно также о наличии такого поля вокруг системы «ооцит – трофоциты». Особенно подробно были исследованы электрические поля вокруг яйцеклеток бурых водорослей – фукуса и др. (работы Л. Джаффе и других исследователей). Установлено, что через яйцеклетку фукуса протекает электрический ток плотностью порядка 5 мкА/см2, причем в один из полюсов яйцеклетки (из которого впоследствии возникнет ризоид) катионы входят, а из противоположного – выходят наружу. Разность потенциалов между полюсами составляет от 2 до 20 мВ. Наличие стационарных электрических полей и электрических (ионных) токов объясняется, по-видимому, разной концентрацией ионных каналов и насосов на противоположных полюсах яйцеклеток. На ризоидном полюсе яйцеклетки фукуса сосредоточены преимущественно каналы для ионов Na+ и Са2+; на противоположном полюсе – насосы, активно откачивающие эти ионы с использованием энергии, аккумулированной в молекулах 82 Болотов А. В. Биология размножения и развития АТФ. Приложением внешнего электрического поля можно сместить каналы и насосы (которые представляют собой ассоциации белковых молекул, обладающих некоторым суммарным зарядом) с их естественных положений. В результате и яйцеклетка в целом изменит свою полярность. Из этих опытов следует, что расположение ионных каналов и насосов однозначно определяет полярность яйцеклетки. Подобные же электрические поля обнаружены вокруг яйцеклетки шпорцевой лягушки: здесь концентрация ионных каналов выше в анимальном, а насосов – в вегетативном полушарии. Электрические поля в системе «ооцит – трофоциты» связаны также с тем, что каналы локализованы преимущественно в мембране трофоцитов, а насосы – в мембране ооцита. В цитоплазме яйцеклетки содержатся специфические структуры: 1. Желточные гранулы, количество которых в разных яйцеклетках различно и зависит от экологических условий и особенностей развития организмов. 2. Слой цитоплазмы, располагающийся непосредственно под плазматической мембраной, называется кортикальным слоем или кортексом (рис. 16, 3). Кортикальная цитоплазма гораздо более вязкая, чем расположенная глубже основная масса цитоплазмы. В кортексе заключены кортикальные гранулы. Эти мембранные структуры гомологичны акросомному пузырьку спермия, поскольку они содержат протеолитические ферменты и формируются аппаратом Гольджи. Однако если каждый спермий имеет только один акросомный пузырек, то каждое яйцо содержит приблизительно 15 000 кортикальных гранул. Кроме того, в кортикальных гранулах помимо протеолитических ферментов содержатся мукополисахариды и белок гиалин. Ферменты и мукополисахариды участвуют в защите яйца от проникновения многочисленных спермиев (предотвращают полиспермию). 3. Яйцевые оболочки (рис. 16, 5, 6). Поверх плазматической мембраны яйцо может быть одето еще несколькими оболочками. Оболочек может быть одна, две или три в разных сочетаниях, в зависимости от условий развития и экологии. Функции оболочек различны и многообразны: 1) через оболочки идет процесс обмена веществ; V. Гаметогенез 83 2) участие в оплодотворении; 3) участие или ограничение полиспермии; 4) на ранних этапах эмбриогенеза обеспечивают сохранение зародышем формы, важной для дальнейших формообразовательных процессов; 5) защитная (от температуры, влажности, химических веществ, механических воздействий, инфекции и т. д.) Различают первичные (желточные), вторичные и третичные оболочки. Первичные оболочки представляют собой производные плазматической мембраны яйцеклетки. Они присущи яйцеклеткам почти всех животных (кроме губок и большинства стрекающих), но особенно хорошо развиты у позвоночных, где они пронизаны уже упоминавшимися выше выростами яйцеклетки и фолликулярных клеток. При малых увеличениях микроскопа такая оболочка выглядит радиально исчерченной и называется zona radiata. Первичная оболочка яиц млекопитающих похожа на блестящий ободок и называется zona pellucida (рис. 16, 5). Возможно, что ее внешняя часть формируется выделениями фолликулярных клеток. Первичные оболочки видоизменяются и достраиваются после оплодотворения яйцеклетки. Вторичные оболочки (хорион) образуются в яичниках и представляют собой исключительно продукт выделения фолликулярных клеток. Они есть далеко не у всех яиц и лучше выражены, например, у насекомых. В хорионе имеется одно или несколько узких отверстий (микропиле), через которые сперматозоид проникает в яйцо. У млекопитающих вторичная оболочка представлена зернистой оболочкой (рис. 16, 6). Третичные оболочки выделяются железами яйцевода. В ряде групп позвоночных (химеровые рыбы, амфибии, рептилии, птицы) они развиты очень сильно. Поразительной сложности достигают третичные оболочки у акуловых и химеровых рыб. Часто эти оболочки имеют вытянутую форму, и яйцо в начале развития заполняет только часть пространства внутри плотной роговой третичной оболочки. Последняя представляет собой как бы «люльку», размеры которой точно подогнаны к размеру и форме зародыша перед вылуплением. У птиц третичные оболочки представлены белком, двумя слоями подскорлуповой «перга- 84 Болотов А. В. Биология размножения и развития ментной оболочки» и скорлупой. Названные компоненты выделяются последовательно расположенными железами яйцевода, по мере движения яйца по яйцеводу вниз. Двигаясь по яйцеводу, яйцо вращается благодаря сокращениям гладкой мускулатуры стенок яйцевода, о чем можно судить по винтовой закрученных халаз – плотных тяжей белкового вещества, которые поддерживают желток в менее плотной массе белка. Интересно, что передне-задняя ось зародыша расположена всегда перпендикулярно направлению движения яйца по яйцеводу, а направление от хвоста зародыша к голове совпадает с направлением вращения яйца. Эта закономерность была впервые отмечена Карлом Бэром и называется правилом Бэра. До сих пор правило Бэра не получило полного объяснения. Примеры третичной оболочки: галлерта у моллюсков; студенистая оболочка у амфибий; белковая, подскорлуповая и скорлуповая оболочки у птиц, коконы многих червей и моллюсков. К третичным оболочкам относится также и белковая жидкость внутри кокона, в которой находится не одно, а несколько яиц. Присутствие всех трех типов оболочек встречается не у всех яиц; яйцо может иметь лишь одну или две из названных оболочек (первичную и вторичную или вторичную и третичную). Желток – это вещество, которое для развивающегося зародыша служит питательным и строительным материалом (у человека – только во время примитивных фаз развития, у птиц – в течение почти всего периода эмбриогенеза). В центральной части ооплазмы более или менее диффузно рассеяны желточные гранулы, в живой яйцеклетке они наблюдаются в виде маленьких образований, сильно отражающих свет. Их вещество состоит из липоидов, а в меньшей мере – из составных частей белков (аминокислот), жиров, углеводов и солей. Наличие красящего вещества липохрома, растворимого в липоидах, придает им желтый цвет. Яйцеклетка различных животных содержит различное количество желтка. Это тесно связано с филогенетическим развитием вида животного. Животные, стоящие филогенетически на самых низких ступенях развития разряда позвоночных, как, например, ланцетник (Amphioxus lanceolatus) не нуждаются для своего сравнительно несложного и недлительного развития в V. Гаметогенез 85 большом количестве питательного вещества в яйце. Однако чем сложнее филогенетическое развитие организма и чем длительнее его онтогенетическое развитие, тем больше желтка содержится в яйцах соответствующего вида животного. В яйцеклетках первого предшественника разряда позвоночных – (ланцетника) – содержится еще очень мало желточных гранул (олиголецитальные яйцеклетки). У земноводных (Amphibia), стоящих в филогенетическом отношении выше, и у некоторых низших рыб содержание желтка уже повышено, его количество можно считать средним (мезолецитальные яйцеклетки). В яйцеклетках позвоночных, стоящих на последующих ступенях филогенетического развития, желтка содержится все больше и больше, например, у высших рыб, пресмыкающихся и птиц яйцеклетки имеют сравнительно крупные размеры с большим количеством желточного вещества (полилецитальные яйцеклетки). У птиц количество желтка в яйцах филогенетически кульминирует. При последующем филогенезе процессы развития у высших животных (млекопитающих) под влиянием новых жизненных факторов становятся все более сложными, их развитие сравнительно удлиняется, а их организмы приобретают более сложное строение. Питание лишь одним желточным веществом является уже недостаточным, поэтому для эмбрионального организма этих животных возникают новые условия и новые источники питания. Самые низшие млекопитающие (яйцеродные), хотя еще и развиваются из полилецитальных яиц и вне тела матери, тем не менее, в соответствии с филогенетическим развитием млекопитающих их яйцеклетки содержат все меньше желтка. Дело в том, что развивающийся организм получает все больше питания и в более сложном виде из другого источника, а именно непосредственно из организма матери при помощи особых устройств, находящихся вне собственного тела зародыша, которые эмбрион создает для этих целей. У человека филогенетическое развитие данных устройств кульминирует; по этой причине яйцеклетка человека, как и яйца высших млекопитающих вообще, снова содержат мало желточного вещества, становясь в процессе филогенетического развития вторично олиголецитальными. С содержанием желтка в яйцеклетке тесно связан и характер процессов развития в начальной стадии эмбриогенеза, то есть 86 Болотов А. В. Биология размножения и развития первичное, иными словами, примитивное развитие эмбриона (бластогенез). Большие массы желтка в ооплазме яйцеклетки затрудняют деление оплодотворенного яйца в период дробления, а полилецитальные клетки даже при дроблении не могут разделиться полностью, в связи с чем новые клетки возникают только на одном полюсе яйца. Распределение желточного вещества в ооплазме и его локализация также играют большую роль в процессе дробления и бластуляции. В зависимости от количества и распределения желтка в ооплазме различаются следующие яйцеклетки: 1. Изолецитальные яйцеклетки – желток располагается в ооплазме более или менее диффузно и равномерно. Обычно это яйца с малым содержанием желтка (олиголецитальные яйцеклетки). Такие яйца у губок, кишечнополостных, бесчерепных (ланцетник), иглокожих и большинства млекопитающих. 2. Телолецитальные яйцеклетки – желток располагается неравномерно, причем большая его часть концентрируется в области одного полюса яйцеклетки (вегетативное полушарие), в то время как на противоположном полюсе желтка гораздо меньше (анимальное полушарие). Такие яйца у кольчатых червей, головоногих моллюсков, рыб, амфибий, пресмыкающихся, птиц и однопроходных млекопитающих. 3. Центролецитальные яйцеклетки – желток располагается в центре и его много. Такие яйца у членистоногих и некоторых кишечнополостных. 4. Алецитальные яйцеклетки – желтка очень мало, располагается в ооплазме более или менее равномерно. Такие яйца у млекопитающих. По количеству желтка в цитоплазме яйца различают: 1. Полилецитальные яйцеклетки – желтка много. Такие яйца у рыб, пресмыкающихся, птиц. 2. Мезолецитальные яйцеклетки – желтка среднее количество. Такие яйца у рыб, амфибий, пресмыкающихся, птиц. 3. Олиголецитальные яйцеклетки – желтка мало. Такие яйца у губок, кишечнополостных, иглокожих, нематод, бесчерепных (ланцетник), оболочников, и большинства млекопитающих. 87 V. Гаметогенез 4. Развитие половых клеток Миграция первичных гоноцитов. Прежде всего гоноциты должны добраться до места своего назначения, т. е. до закладки половой железы гонады. Как первичные гоноциты, так и резервные клетки типа интерстициальных способны двигаться самостоятельно, иногда (шпорцевая лягушка) по ориентированным волокнам внеклеточного матрикса. Но значительную часть пути они, повидимому, проходят пассивно: интерстициальные клетки – с током воды в гастральной полости, а гоноциты куриного зародыша, как, вероятно, и других амниот, – с током крови по эмбриональным кровеносным сосудам. Но поблизости от места своего назначения они движутся, несомненно, активно. Так, гоноциты куриного зародыша активно проползают через стенку половой железы. На этом этапе своего движения они привлекаются химическими веществами типа мукополисахаридов, выделяемых зачатком гонады. Перед нами в данном случае один из немногих достоверных примеров хемотаксиса – движения клеток вверх по градиенту концентрации химических веществ. С другими примерами хемотаксиса мы познакомимся при изучении движения сперматозоидов. Пол зародыша определяется хромосомным набором, полученным им в момент оплодотворения. Но на стадии первичных гоноцитов мужские и женские половые клетки, как правило, неотличимы. Различия проявляются лишь после их проникновения в половые железы. Надо сказать, что генетическая детерминация пола не всегда стабильна и окончательна – у некоторых животных пол может быть переопределен воздействием гормонов. Размножение и гибель половых теток. Попав в зачатки половых желез, гоноциты обоих полов усиленно размножаются путем обычных митотических делений. В течение этого периода размножения женские половые клетки называются оогониями, а мужские – сператогониями. Однако оогонии прекращают размножаться еще в эмбриональном периоде жизни самки, задолго до наступления половой зрелости. Подсчитано, что у 4месячного плода человека имеется приблизительно 2–3 млн женских половых клеток, потом наступает массовая дегенерация 88 Болотов А. В. Биология размножения и развития их, так что к моменту рождения остается около 1 млн, а к 7летнему возрасту – примерно 300 тыс. половых клеток. Размножение сперматогониев, напротив, происходит в течение всего периода половой зрелости самца, непрерывно (теплокровные животные) или с сезонной ритмикой (холоднокровные). 5. Профаза мейоза Перед вступлением клетки в профазу мейоза в предмейотической интерфазе происходит синтез белков, РНК и удвоение ДНК (4с). Профаза мейоза (рис. 17) в отличие от таковой митоза сопровождается более значительными преобразованиями ядерных структур, большей длительностью во времени, достигающей иногда многих лет (например, у птиц и млекопитающих). Рис. 17. Профаза 1-го деления мейоза Условно профазу мейоза делят на несколько стадий (рис. 17). Лептотенная (с лат. leptos – тонкий) стадия – хромосомные нити тонкие (период роста – ооцит и сперматоцит 1-го порядка) и лишь к концу стадии по длине хромосом выявляются небольшие утолщения-хромомеры, количество и расположение которых специфичны для пары гомологичных хромосом. V. Гаметогенез 89 Зиготенная стадия (с лат. zeugos – пара) – состоит в конъюгации гомологичных хромосом. Сближение гомологичных хромосом (за счет комплиметарных участков ДНК) и пары конъюгирующих хромосом называются бивалентами. Сдваивание подготавливает редукционное деление мейоза. Пахитенная стадия (с лат. pachis – толстый) – завершение конъюгации гомологичных хромосом. При усилении спирализации хромосом происходит тесное взаимное закручивание хромосом в составе каждого бивалента. В конце этой стадии в составе каждой хромосомы, образующей бивалент, делается различимой ее двухроматидная структура. С этого момента бивалент может быть назван тетрадой, т. е., две гомологичные хромосомы, конъюгируя, образуют бивалент, каждая из этих хромосом расщепляется на две хроматиды. Четырехнитчатое строение пахитенных хромосом отражает подготовленность хромосомного аппарата клетки к двум последующим мейотическим делениям, в результате которых каждая из образующих клеток получит по одному элементу из каждой тетрады. Диплотенная стадия (лат. diplos – двойной) – начинается процесс расхождения и раскручивания гомологичных хромосом в составе каждого бивалента. При этом может быть, что несестринские хроматиды в биваленте могут быть соединены между собой в одной или нескольких точках, образуя перекресты – хиазмы. Хиазмы являются следствием происходящего в предшествующий период профазы мейоза кроссинговера – обмена участками между несестринскими хроматидами, происходящего в результате разрывов и воссоединений в соответствующих локусах хроматид. Кроссинговер, приводящий к обмену генетическим материалом между хроматидами, служит одной из причин увеличения резерва наследственной изменчивости в потомстве при половом размножении. У млекопитающих хромосомы сильно спирализованные и образуют клубок той или иной плотности – кариосферу. Снабжение ооцита запасными веществами выполняют питающие клетки или клетки фолликулярного эпителия яичника. У человека еще в эмбриогенезе хромосомы, приняв морфологическую форму «ламповых щеток», прекращают дальнейшие структурные изменения на многие годы. В женской профазе 1-го 90 Болотов А. В. Биология размножения и развития деления мейоза имеется стадия диктотены (до диакинеза). При достижении женским организмом половой зрелости под влиянием лютеинизирующего гормона передней доли гипофиза один ооцит ежемесячно возобновляет мейоз, т. е. он проходит завершающую стадию профазы 1-го деления – диакинез. Диакинез отличается все большей степенью спирализации хромосом, все большим смещением хиазм к концевым участкам хромосом (терминализация хиазм). Диакинез завершает профазу мейоза, и клетка вступает в первое деление мейоза. Затем после непродолжительной интерфазы, в ходе которой не происходит удвоения ДНК, наступает профаза второго мейотического деления и затем второе деление мейоза, которое по форме напоминает митоз и не сопровождается изменением числового состава хромосом, в связи с чем его называют эквационным или уравнительным делением. Но оно принципиально отличается от митоза тем, что отходящие к двум клеткам из состава каждой хромосомы хроматиды могут быть не идентичны по генетическим свойствам вследствие происшедшего кроссинговера. Итак, в ходе делений мейоза перекомбинация генетического материала осуществляется дважды: при первом делении в связи с расхождением к каждому из полюсов веретена части материнских и части отцовских хромосом, при втором – вследствие расхождения разнокачественных по генетическому составу хроматид. Все сложные преобразования хромосомного аппарата клетки, сопровождающие мейоз, имеют два важнейших следствия: 1. Редукцию числа хромосом и соответственно количества ДНК в ядре половых клеток. 2. Перекомбинацию генетического материала между формирующимися клетками, увеличивающую резерв наследственной изменчивости будущего потомства. 6. Оогенез Яйцевые клетки образуются в женской половой железе – яичнике. Они проходят долгий путь развития, который начинается в эмбриональном и продолжается в репродуктивном периоде онтогенеза особей женского пола. Развитие яйцеклеток происходит в три стадии: 1) размножения; 2) роста; 3) созревания. V. Гаметогенез 91 1. Стадия размножения происходит у самок-эмбрионов в яичниках. При этом зачатковый эпителий, расположенный на поверхности яичника в виде тяжей, которые позже превращаются в стенки флюгеровских мешков. В эти мешки поступают первичные половые клетки – овогонии, которые многократно делятся путём митоза. Мешки постепенно распадаются на первичные или примордиальные фолликулы. Они состоят из половой клетки овогонии, окружённой одним слоем рыхлорасположенных плоских фолликулярных клеток (это бывшие клетки зачаткового эпителия). Стадия размножения заканчивается к моменту рождения самки. При этом около 200 тысяч первичных или спящих фолликулов располагаются возле наружной оболочки яичника. В течение жизни спящие фолликулы поочерёдно переходят в стадию роста. 2. Стадия роста происходит у родившихся самок в яичниках. У человека протекает в функционирующем зрелом яичнике (после полового созревания девочки) и состоит в превращении ооцита 1-го порядка первичного фолликула в ооцит 1-го порядка в зрелом фолликуле. При этом под влиянием гормонов пробуждается половая клетка, которая теперь называется ооцит 1-го порядка. В стадию роста ооцит 1-го порядка растёт и готовится к будущему делению путём мейоза, т. е. стадия роста является интерфазой перед мейозом. И начинается весьма своеобразный, только этой клетке свойственный период роста. Он связан с поступлением в яйцеклетку питательных веществ извне и с рядом синтетических процессов в самой яйцеклетке. Увеличение массы и объема яйцеклетки может быть колоссальным. Так, ооциты дрозофилы за 3 дня увеличиваются в 90 тыс. раз. У лягушек диаметр молодого ооцита примерно 50 мкм, а зрелого – 2 000 мкм, что соответствует увеличению объема в 64 тыс. раз. Рост ооцита идет сравнительно медленно: только 3-летние лягушки достигают половозрелости. Несравненно быстрее растет яйцо у птиц. Например, у курицы последние 6 дней перед овуляцией объем яйца возрастает в 200 раз. Яйца млекопитающих меньше по размеру; диаметр яйца мыши возрастает от 20 до 70 мкм, что все же соответствует росту объема более чем в 40 раз. Рост яйцеклетки млекопитающих может длиться десятки лет, например, у человека – около 30 лет. 92 Болотов А. В. Биология размножения и развития При подготовке к 1-му делению созревания ооцит длительное время находится в фазе диакинеза, когда и происходит его рост и вителлогенез. Рост ооцитов всех животных принято разделять на два периода. Первый из них называют периодом малого роста (стадия Винвартера), превителлогенеза, или цитоплазматического роста; второй – периодом большого роста, вителлогенеза, или трофоплазматического роста. Для первого периода характерно относительно малое и притом пропорциональное увеличение массы ядра и цитоплазмы; таким образам, ядерноцитоплазматическое отношение при этом не изменяется. Второй период характеризуется, напротив, резкой интенсификацией роста цитоплазматических компонентов: ядерно-цитоплазматическое отношение при этом уменьшается. В течение второго периода в ооците откладывается желток. В период вителлогенеза в ооците 1-го порядка образуется желток, а также другие питательные вещества – жиры и гликоген. Желток представляет собой высокофосфорилированный кристаллический белок. Он откладывается в виде желточных гранул, одетых пограничной мембраной. По количеству откладываемого в период вителлогенеза желтка яйцеклетки принято делить на полилецитальные, или многожелтковые (например, большинство членистоногих, некоторые рыбы, птицы), мезолецитальные – со средним количеством желтка (амфибии, осетровые рыбы), олиголецитальные – маложелтковые (большинство червей, моллюсков, иглокожих) и алецитальные, т. е. практически безжелтковые (плацентарные формы – млекопитающие, а также и некоторые беспозвоночные, например первичнотрахейные). Количество желтка строго детерминировано генетически и не зависит от условий питания самки. По способу образования желток принято разделять на экзогенный и эндогенный. Присущий большинству видов животных экзогенный желток строится на основе белка – предшественника вителлогенина, который поступает в ооцит извне. У позвоночных вителлогенин синтезируется в печени матери и транспортируется к содержащему ооцит фолликулу по кровеносным сосудам. Попадая затем в пространство, непосредственно окружающее ооцит (периооцитное пространство), вителлогенин поглощается ооцитом путем пиноцитоза, в составе так называемых V. Гаметогенез 93 окаймленных мембранных пузырьков. Каждая гранула желтка образуется в результате слияния большого количества (до тысячи) таких пиноцитозных пузырьков. При формировании гранул желтка вителлогенин расщепляется на сильно фосфорилированный белок фосвитин и другой белок – липовителлин, содержащий до 20 % липидов. Эндогенный желток синтезируется из низкомолекулярных предшественников внутри самого ооцита, в его сильно развитом эндоплазматическом ретикулуме. Желточные гранулы возникают из концевых цистерн аппарата Гольджи. Лишь немногие типы яйцеклеток развиваются исключительно за счет эндогенного желтка. Подготовка к делениям созревания и синтетические процессы в период превителлогенеза. Весь период превителлогенеза проходит на фоне подготовки ооцита 1-го порядка к последующим делениям созревания, или мейоза. Подготовка к 1му делению созревания начинается вслед за прекращением оогониальных делений с того, что ооцит вступает в S-фазу редукционного деления, т. е. в фазу удвоения ДНК (предшествующую любому клеточному делению). Удвоенное количество ДНК в геноме, достигнутое по завершении этой фазы (4с). Затем наступает профаза 1-го деления мейоза. Профаза мейоза продолжается у ооцитов млекопитающих от несколько дней (у кроликов – до 20 дней) до нескольких лет (у птиц и млекопитающих), однако по достижении фазы диплотены, когда гомологичные хромосомы уже прошли конъюгацию и начали расходиться, наступает стационарная фаза диакинеза. На этой стадии дальнейшее течение мейоза сильно замедляется или же полностью прекращается. Этот блок мейоза продолжается до достижения особью половозрелости, т. е. у ряда млекопитающих и человека много лет. Однако в период диакинеза ядерный материал ооцита не является инертным: у большинства яйцеклеток, обладающих так называемым солитарным, или фолликулярным способами питания, он выполняет роль матрицы для синтеза всех видов РНК – рибосомных, транспортных и информационных, или матричных (соответственно рРНК, тРНК и мРНК). Все эти типы РНК синтезируются преимущественно впрок, т. е. для использования уже оплодотворенной яйцеклеткой. 94 Болотов А. В. Биология размножения и развития События, происходящие в половой клетке в это время: 1) происходит накопление питательных веществ, как источника энергии; 2) удваивается молекула ДНК; 3) удваивается клеточный центр; 4) синтезируется специальный белок типа тубулина для построения нитей веретена деления; 5) происходит конъюгация и кроссинговер гомологичных хромосом. Кроме ооцита 1-го порядка в стадию роста начинают расти и вспомогательные фолликулярные клетки. С этого момента фолликулы получают название вторичных или растущих фолликулов. Постепенно плоские фолликулярные клетки становятся кубическими, а затем и высокими – призматическими. Они формируют вторичную оболочку – лучистый венец. Затем, за счёт веществ, вырабатываемых ооцитом 1-го порядка и клетками лучистого венца, между ними появляется блестящая или прозрачная оболочка, состоящая из гликозаминогликанов (гиалуроновая кислота) и белков. Затем фолликулярные клетки лучистого венца начинают размножаться и формируют снаружи зернистый слой – это будет третья вторичная оболочка, состоящая из множества плотно прилегающих друг у другу фолликулярных клеток (отсутствуют щели между клетками). Снаружи вторичный фолликул покрывается соединительно-тканной оболочкой – текой. Итак: вторичный или растущий фолликул состоит из овоцита 1-го порядка, покрытого плазмалеммой, блестящей оболочкой, лучистым венцом (это первый слой кубических или призматических фолликулярных клеток). В некоторых фолликулах снаружи лучистого венца расположен зернистый слой из плотно прилегающих фолликулярных клеток. В период полового созревания организма фолликулярные клетки зернистого слоя вырабатывают половые гормоны (фолликулины или эстрогены), которые скапливаются в небольших щелях между клетками зернистого слоя. Такой фолликул, в котором имеются полости с гормонами, называется третичным или пузырчатым. Самые зрелые третичные фолликулы называются Граафовым пузырём. V. Гаметогенез 95 Наружная оболочка Граафова пузыря – тека, состоящая из двух слоев: наружного – фиброзного и внутреннего – сосудистого, состоящего из ретикулярной ткани, пронизанной множеством капилляров. Здесь содержится много кровеносных сосудов и интерстициальных клеток, дополнительно вырабатывающих половые гормоны. Тека отделяется от эпителиальных фолликулярных клеток зернистого слоя базальной мембраной (это студневидная прослойка). Внутренний слой зернистой оболочки представлен клетками цилиндрической формы с радиальным расположением. Этот слой называется лучистым венцом (рис. 10, 4). Своими отростками фолликулярные клетки передают питательные вещества яйцеклеткам, которые находятся в стадии роста. Среди клеток зернистого слоя появляются щели, или полости, заполненные жидкостью. Таких щелей становится все больше, они объединяются и образуют общую полость, с этого момента фолликул называется пузырчатым или Граафовым пузырьком. Граафов пузырек сильно увеличивается и выпячивает стенку яичника. Внутри фолликула появляется бугорок (яйценосный) (рис. 10, 1) – это скопление фолликулярных клеток, и на его вершине среди зернистых фолликулярных клеток располагается созревающее яйцо, окруженное высокими фолликулярными трофическими клетками, образующими лучистый венец. Стенки Граафова пузырька секретируют женский половой гормон – эстроген, которым насыщается жидкость пузырька. Попадая в кровь, он стимулирует образование лютеотропного гормона в гипофизе. Скопление половых гормонов в Граафовом пузыре приводит к тому, что пузырь лопается, а ооцит 1-го порядка в прозрачной оболочке и лучистом венце овулирует в брюшную полость (рис. 9, 7), где его подхватывает воронка яйцевода. Дальнейшее развитие половой клетки будет происходить в трубке яйцеводе. У свиньи в одном Граафовом пузырьке может созревать несколько яйцеклеток. У овец могут овулировать несколько Граафовых пузырьков. 3. Стадия созревания, или мейоз, состоит из двух быстро следующих друг за другом делений – редукционного и эквационного (уравнительного). При первом этапе (начало) мейоза ооцит 1-го порядка даёт начало двум дочерним клеткам с половинным набором хромосом. Это будет ооцит 2-го порядка и ма- 96 Болотов А. В. Биология размножения и развития ленькое направительное тельце. И завершается выходом его из яичника в результате овуляции. Рис. 18. Схема делений созревания в ооците (по Э. Вильсону, 1940): А – метафаза 1-го деления созревания; Б, В – выделение I полярного тельца (1); Г, Д – метафаза и анафаза 2-го деления созревания (одновременно делится I полярное тельце); Е – выделение II полярного тельца (2) Перед 1-м делением созревания ядро ооцита мигрирует к его поверхности. Та точка поверхности ооцита, к которой ближе всего располагается ядро, названа анимальным, противоположная – вегетативным полюсом яйцеклетки. Затем в результате 1-го деления созревания половина хромосомного набора выталкивается в очень маленькую клетку, которая называется первым редукционным, или полярным тельцем (рис. 18, В). Впоследствии эта клетка делится на две столь же малые, и никакого участия в дальнейшем развитии они не принимают (за исключением некоторых случаев партеногенеза). Яйцевая клетка после выделения 1-го редукционного тельца называется ооцитом 2-го порядка. Второе деление созревания осуществляется путем V. Гаметогенез 97 выделения 2-го редукционного тельца таких же размеров, как и 1-го. После его выделения ооцит 2-го порядка превращается в зрелое яйцо (рис. 18, Г). Биологический смысл резкой неравномерности делений созревания очевиден: невыгодно дробить на части накопленный в процессе роста яйцеклетки запас питательных веществ. Итак, блок мейоза продолжается до достижения особью половозрелости, Выход из фазы диакинеза и начало собственно делений созревания определяются половыми гормонами: гонадотропные гормоны гипофиза воздействуют на фолликулярный эпителий, продуцирующий в ответ прогестерон или его аналоги. Гормоны фолликулярного эпителия поступают в ооцит и стимулируют его к созреванию. Современные представления о механизмах этого процесса таковы. Как уже говорилось, в период роста ооцита он связан с фолликулярными клетками щелевыми контактами, через которые из последних в ооцит поступает цАМФ. Высокий уровень цАМФ блокирует (путем фосфорилирования) белковый фактор созревания, который присутствует в цитоплазме ооцита. Под действием прогестерона или его аналогов щелевые контакты размыкаются, концентрация цАМФ в цитоплазме ооцита падает, и фактор созревания деблокируется (дефосфорилируется). Далее он способен еще не вполне изученным образом размножаться (амплифицироваться) в цитоплазме: достаточно инъецировать немного этого фактора в незрелый ооцит, чтобы затем его количество возросло. Фактор созревания вызывает дезинтеграцию (разрушение) оболочки ядра ооцита, разрушение ядрышек и миграцию хромосом к будущему анимальному полюсу, где и произойдут деления созревания. Из двух делений созревания первое у большинства видов является редукционным, так как именно в ходе этого деления гомологичные хромосомы расходятся по разным клеткам. Таким образом, каждая из разделившихся клеток приобретает половинный (гаплоидный) набор хромосом, где каждый ген представлен лишь одной аллелью. Поскольку 1-му делению созревания предшествовала, как и обычному митотическому делению, Sфаза, каждая из разошедшихся хромосом содержит двойное количество ДНК (две хроматиды). Эти генетически идентичные 98 Болотов А. В. Биология размножения и развития хроматиды и расходятся по сестринским клеткам во 2-м делении созревания, которое является эквационным (как и обычное деление соматических клеток). Наследственный материал представляет из себя одну хромосому, содержащую двойное количество ДНК (1n2c) (рис. 19). Рис. 19. Схема оогенеза и сперматогенеза Эти клетки без интерфазы моментально делятся снова, проходя состояние профазы, метафазы, анафазы и телофазы. При этом из ооцита 2-го порядка образуется крупная зрелая яйцеклетка и крошечное направительное тельце. А из первого направительного тельца также образуются две дочерние клетки – два направительных тельца (рис. 18, 2; 19). V. Гаметогенез 99 Итог мейоза: четыре дочерние клетки с половинным набором хромосом (это яйцеклетка и три направительных тельца, которые почти сразу погибают и рассасываются). На месте лопнувшего Граафова пузыря в яичнике отмечается небольшое кровотечение. Здесь размножаются фолликулярные клетки, откладывается желтый пигмент. Так образуется желтое тело – железа внутренней секреции. Она вырабатывает гормон – прогестерон, тормозящий овогенез. Различают: 1) желтое тело полового цикла, оно функционирует один половой цикл и рассасывается, превращаясь сначала в белое тело (рис. 9, 11); 2) желтое тело беременности (функционирует в период беременности). Некоторые третичные фолликулы в яичнике, наработав гормоны, подвергаются разрушению. После двух делений созревания число хромосом в каждой из клеток оказывается гаплоидным (1n), количество ДНК в каждом клеточном ядре тоже будет гаплоидным (1с). Основная особенность делений созревания в ооцитах состоит в том, что эти деления резко неравномерны. Лишь у немногих видов (некоторые кишечнополостные; из иглокожих – морские ежи) мейоз доходит до конца без участия сперматозоида, внедряющегося в яйцеклетку. У большинства животных течение мейоза останавливается на некотором этапе созревания (возникает, как говорят, блок мейоза), и для дальнейшего его протекания требуется оплодотворение яйцеклетки сперматозоидом (или действие другого активирующего агента). Различают следующие три типа блока мейоза: 1. Мейоз останавливается на стадии диакинеза, т. е. участие сперматозоида необходимо для протекания обоих мейотических делений. Этот тип блока мейоза наблюдается у губок, отдельных представителей плоских, круглых и кольчатых червей, моллюсков и щетинкочелюстных. Удивительным образом сюда же относятся три представителя млекопитающих: собака, лиса и лошадь. 2. Мейоз останавливается на метафазе 1-го деления созревания. Такой блок мейоза отмечен у некоторых губок, немертин, кольчатых червей, моллюсков и почти у всех насекомых. 3. Мейоз останавливается на метафазе 2-го деления созревания. Сюда относятся почти все хордовые. У летучих мышей 100 Болотов А. В. Биология размножения и развития блок мейоза происходит на анафазе 2-го деления созревания. Именно на этих стадиях созревания происходит овуляция яйцеклетки у млекопитающих. Лишь оплодотворение позволяет завершиться ее созреванию и перейти к дальнейшему развитию. 4. Мейоз не блокируется, яйцо останавливается в развитии после завершения мейоза, когда сформирован женский пронуклеус (некоторые кишечнополостные и иглокожие – морские ежи). В большинстве случаев блок мейоза снимается после активации яйца при оплодотворении или путем искусственной активации. Механизм блока мейоза пока не выяснен. Яйцеклетки, или ооциты созревают в неизмеримо меньшем количестве, чем сперматозоиды. У женщины в течение полового цикла 24–28 дней) созревает, как правило, одна яйцеклетка. Таким образом, за детородный период образуются около 400 зрелых яйцеклеток. 7. Гормональная регуляция половой функции у женщин. Овуляция Следует отметить, что большинство ооцитов в растущих и созревающих фолликулах в разные периоды своего роста претерпевают атрезию. Завершающие этапы периода большого роста ооцита и фолликула, созревания и овуляции происходят циклически и зависят от циклической деятельности системы гипофиз – гипоталамус – яичники. Женские половые клетки, так же как и мужские, в определенной степени отделены от микроокружения гематофолликулярным барьером, который создает оптимальные условия для метаболизма ооцитов. Он состоит из соединительной ткани (теки), сосудов микроциркуляторного русла, базальной мембраны, фолликулярного эпителия и блестящей зоны. Вся система регуляции менструального цикла построена по иерархическому принципу (нижележащие структуры регулируются вышележащими, которые, в свою очередь, реагируют на изменения в нижележащих уровнях). При этом сигналы, поступающие от нижележащих структур, корректируют деятельность вышележащих. V. Гаметогенез 101 Нормальная продолжительность менструального цикла от 21 до 35 дней, продолжительность менструации в среднем 3–4 дня, до 7 дней, величина кровопотери 50–100 мл. Нормальный менструальный цикл всегда овуляторный, отсутствие выброса в брюшную полость ооцита 2-го порядка говорит об ановуляторном цикле. Однако под влиянием эндогенных и экзогенных факторов может наблюдаться ускорение или замедление овуляции, вплоть до исчезновения её. Обычно у большинства женщин наблюдается от 11 до 13 менструаций в год. На ускорение овуляции влияет повышенная масса тела, употребление пищи с фитогормонами. На замедление овуляции влияют: болезни, стресс, физические нагрузки, недостаточное питание, употребление анаболических стероидов, повышенного уровня андрогенов экзогенного или эндогенного происхождения. Ановуляция может возникнуть по следующим причинам: избыточное содержание в женском организме мужских половых гормонов; заболевания щитовидной железы (гипотиреоз, гипертиреоз); избыточная масса тела; значительное снижение массы тела; чрезмерные физические нагрузки; употребление анаболических стероидов; курение; лекарства: антибиотики, гормоны; алкоголь (регулярное употребление); употребление избытка сладкого; недостаток веса женщины – 45 кг и ниже; опухоли гипофиза и гипоталамуса. Природой задумано так, что длина первой фазы цикла может очень сильно колебаться у одной и той же женщины, в то же время длина второй фазы остается стабильной, т. е. если овуляция происходит всегда за 15 дней до начала нового цикла (рис. 20; 21). Согласно современным представлениям менструальРис. 20. Регуляция половой ная функция регулируется нейрогуморальным путем функции у женщин 102 Болотов А. В. Биология размножения и развития при участии: 1) коры головного мозга; 2) подкорковых вегетативных центров, расположенных главным образом в гипоталамусе; 3) гипофиза; 4) яичников; 5) матки, маточных труб и влагалища, содержащих рецепторы, реагирующие на действие половых гормонов яичников. Циклические функциональные изменения в системе гипоталамус – гипофиз – яичники условно объединяются в яичниковый цикл, а циклические изменения в слизистой оболочке матки – в маточный. Одновременно происходят циклические сдвиги во всем организме женщины (менструальная волна), представляющие собой периодические изменения деятельности ЦНС, обменных процессов, функции сердечнососудистой системы и терморегуляции (рис. 21). Кора головного мозга регулирует процессы, связанные с развитием менструальной функции. Через нее осуществляется влияние внешней среды на нижележащие отделы нервной системы, участвующие в регуляции менструального цикла. В гипоталамусе концентрируется влияние импульсов нервной системы и гормонов периферических желез внутренней секреции. В его клетках содержатся рецепторы для всех периферических гормонов, в т. ч. эстрогенов и прогестерона. Нейрогормоны гипоталамуса, стимулирующие освобождение тропных гормонов в передней доле гипофиза, называют рилизингфакторами или либеринами. Существуют также статины – нейрогормоны гипоталамуса, ингибирующие освобождение тропных гормонов в аденогипофизе. Секреция Рилизинг-гормон ЛГ (РГ-ЛГ люлиберин) генетически запрограммирована и происходит в определенном пульсирующем ритме с частотой примерно один раз в час (цирхоральный ритм). В передней доле гипофиза (аденогипофиз) синтезируется гонадотропные гормоны гипофиза: фолликулостимулирующий (ФСГ), лютеинизирующий (ЛГ), пролактин (лактотропный, ПРЛ). ФСГ вызывает развитие и созревание первичного фолликула. ФСГ стимулирует рост фолликула, пролиферацию гранулезных клеток, индуцирует образование рецепторов ЛГ на поверхности клеток гранулезы. ЛГ стимулирует образование андрогенов (предшественников эстрогенов) в тека-клетках, совместно с ФСГ способствует овуляции и стимулирует синтез прогестерона в лютеинизированных клетках гранулезы овулировавшего фолликула. V. Гаметогенез 103 Рис. 21. Гормональная регуляция половой функции у женщин, температурная кривая. ОВЦ – овуляторный цикл; АВЦ – ановуляторный цикл Пролактин стимулирует синтез и секрецию прогестерона, превращает не функционирующее желтое тело в функционирующее. При отсутствии пролактина наступает обратное развитие этой железы. Пролактин оказывает многообразное действие на организм женщины. Его основная биологическая роль – рост молочных желез и регуляция лактации. Он обладает также жиромобилизующим эффектом и оказывает гипотензивное действие. Увеличение секреции пролактина является одной из частых причин бесплодия, так как повышение его уровня в крови тормозит стероидогенез в яичниках и развитие фолликулов. 104 Болотов А. В. Биология размножения и развития В период полового созревания организма фолликулярные клетки зернистого слоя вырабатывают половые гормоны (фолликулины или эстрогены), которые скапливаются в небольших щелях между клетками зернистого слоя. Такой фолликул, в котором имеются полости с гормонами, называется третичным или пузырчатым и наиболее крупные третичные фолликулы с большой полостью внутри и значительным количеством фолликулярных клеток называются Граафовым пузырём. Разрыв зрелого фолликула (овуляция) происходит под влиянием ФСГ, который, влияя на стенку зрелого фолликула, побуждает к выработке в нем гармона эстрина и ЛГ. Происходит разрыв фолликула и выброс ооцита 2-го порядка в брюшную полость. Ооцит 2-го порядка, окруженный фолликулярным эпителием, из брюшной полости попадает в воронку и далее в просвет маточной трубы. Здесь (при наличии мужских половых клеток) быстро происходит второе деление созревания и образуется зрелая яйцеклетка, готовая к оплодотворению. Значительное число фолликулов не достигает стадии зрелости, а претерпевает атрезию – своеобразную перестройку деструктивного характера. Атрезия ооцитов и возникает атретическое тело (corpus atreticum), внешне несколько напоминающее желтое тело, но отличающееся от последнего наличием в центре блестящей зоны ооцита. В ходе дальнейшей инволюции атретических тел на их месте остаются скопления интерстициальных клеток. Так как интерстициальные клетки участвуют в выработке овариальных гормонов, следует допустить, что атрезия, результатом которой является увеличение количества этих клеток, необходима для гормонообразования в яичнике женщины. Таким образом, в регуляции функции репродуктивной системы основными являются цирхоральная секреция РГ-ЛГ в нейронах гипоталамуса и регуляция выделения ЛГ и ФСГ эстрадиолом по механизму отрицательной и положительной обратной связи. Признаки овуляции делятся на объективные и субъективные Объективные признаки овуляции (эти признаки основываются на лабораторных исследованиях. При помощи объективных признаков можно точно определить наступление овуляции) и субъективные признаки овуляции: V. Гаметогенез 105 1. Общая реакция организма на разрыв фолликула. 2. Гормональный изменения, сопровождающиеся межменструальными болями. 3. Изменение слизи, выделяемой шейкой матки. 4. Изменение шейки матки. 5. Уменьшение времени редукции оксигемоглобина. 6. Увеличение α и β-глобулинов. 7. Уменьшение альбуминов. 8. Повышение секреции гонадотропных гормонов. 9. Межменструальные кровотечения. 10. Падение экскрекции витамина С. 11. Увеличение либидо (половое влечение). 12. Болезненность груди. 13. Боль внизу живота. 14. Изменение шейки матки и ее выделения. 15. Изменение настроения (депрессия). 16. Изменение чувствительности рецепторных элементов. Как правило, чувствительность повышается. 17. Повышение ректальной Т ºС в фазе желтого тела и ↓Т ºС перед овуляцией: Обычно ректальная Т ºС < 37 ºС, за 4–6 дней до овуляции начинает расти, затем за 1–3 дня до овуляции ↓Т ºС. После овуляции (фаза желтого тела) Т ºС q на 0,5–0,8 ºС и держится в течение всей второй фазы, снижаясь лишь за 1–2 дня до менструации (рис. 21). Двухфазная температурная кривая указывает на чередование фолликулиновой и прогестероновой фаз и говорит об овуляторном цикле. В случае однофазной температурной кривой можно говорить об ановуляторном цикле. Желтое тело (corpus luteum) Под воздействием ЛГ образуется желтое тело (рис. 9, 8–9, 11) и синтезируется прогестерон в лютеинизированных клетках гранулезы овулировавшего фолликула. Под влиянием избытка лютеинизирующего гормона, вызвавшего овуляцию, элементы стенки лопнувшего зрелого пузырька претерпевают изменения, приводящие к формированию желтого тела – временной добавочной эндокринной железы в составе яичника. При этом в полость запустевшего пузырька изливается кровь из сосудов внутренней оболочки, целость кото- 106 Болотов А. В. Биология размножения и развития рых нарушается в момент овуляции. Сгусток крови быстро замещается соединительной тканью в центре развивающегося желтого тела. В развитии желтого тела различают четыре стадии: 1) пролиферации; 2) железистого метаморфоза; 3) расцвета; 4) инволюции. В первой стадии – пролиферации и васкуляризации – происходит размножение эпителиоцитов бывшего зернистого слоя и между ними интенсивно врастают капилляры из внутренней оболочки. Затем наступает вторая стадия – железистого метаморфоза, когда клетки фолликулярного эпителия сильно гипертрофируются и в них накапливается желтый пигмент (лютеин), принадлежащий к группе липохромов. Такие клетки называются лютеиновыми или лютеоцитами (luteocyti). Объем новообразующегося тела быстро увеличивается, и оно приобретает желтый цвет. С этого момента желтое тело начинает продуцировать свой гормон – прогестерон, переходя, таким образом, в третью стадию – расцвета. Продолжительность этой стадии различна. Если оплодотворения не произошло, период расцвета желтого тела ограничивается 12–14 днями. В этом случае оно называется менструальным желтым телом (corpus luteum menstruationis). Более длительно желтое тело сохраняется, если наступила беременность – это желтое тело беременности (corpus luteum graviditationis). Разница между желтым телом беременности и менструальным ограничивается только длительностью периода расцвета и размерами (1,5–2 см в диаметре у менструального и более 5 см в диаметре у желтого тела беременности). После прекращения функционирования как желтое тело беременности, так и менструальное претерпевают инволюцию (стадию обратного развития). Железистые клетки атрофируются, а соединительная ткань центрального рубца разрастается. В результате на месте бывшего желтого тела формируется белое тело (corpus albicans) – соединительнотканный рубец. Оно сохраняется в яичнике на протяжении нескольких лет, но затем рассасывается. 107 V. Гаметогенез Под влиянием прогестерона происходит увеличение эндометрия (слизистой матки) с 1 мм до 7 мм. Он становится отечным, маточная железа приобретает штопорообразный вид. Просвет переполнен секреторными продуктами. Такие изменения необходимы для имплантации зародыша в стенку матки. В маточных трубах и влагалище имеются также рецепторы к половым стероидным гормонам, но циклические изменения в них выражены менее отчетливо. В других органах (молочные железы, кожные покровы, кости, жировая ткань) клетки названных тканей и органов содержат рецепторы к половым гормонам. В предменструальном периоде количество железистых ходов в молочной железе увеличивается, они расширяются, доли становятся отечными, эпителиальный слой набухает, вакуолизируется и может ощущаться болезненность. В послеменструальном периоде отечность долей, инфильтрация вокруг крупных ходов исчезают и неприятные ощущения тоже исчезают. В головном мозге также обнаружены рецепторы к половым гормонам, что, по-видимому, может объяснять циклические колебания психики женщины в течение менструального цикла. 8. Сперматогенез Мужские половые клетки, как и женские, возникают из первичных гоноцитов. При сперматогенезе непосредственными потомками гоноцитов являются стволовые сперматогенные клетки (у млекопитающих называемые также сперматогониями типа А) (рис. 22, Б). Они присутствуют не только у зародышей, но и у половозрелых самцов; в семенниках млекопитающих они располагаются в пристеночном слое семенных канальцев. Стволовые клетки время от времени (нерегулярно) делятся, оставаясь в недифференцированном состоянии. Некоторые из них при этом перемещаются ближе к центру семенного канальца, их деления становятся более регулярными, а после каждого деления эти клетки изменяют свою величину и форму. Такие клетки называют либо просто сперматогониями, либо сперматогониями типа В, а их деления – сперматогониальными (рис. 22, Б). Сперматогониальные деления постоянно происходят у половозрелых самцов. Однако число делений отдельного сперма- 108 Болотов А. В. Биология размножения и развития тогония невелико (от 1 до 14) и строго определенно для каждого вида животного. Например, у паразитического червя Dicyema каждый сперматогоний делится всего один раз, у человека, повидимому, 4 раза, а у рыбки гуппи – 14 раз. Откуда же в таком случае берется столь значительное количество продуцируемых сперматозоидов (до 108 в сутки у человека и кролика, 1010 у хряка и лошади и т. д.)? Очевидно, что в основном за счет делений стволовых клеток. И действительно, подсчитано, что у дрозофилы каждая стволовая сперматогониальная клетка (а всего этих клеток порядка 28) делится 107 раз, т. е. продуцирует 2107 сперматогониев, каждый из которых затем делится еще всего 4 раза. После определенного числа делений сперматогонии передвигается еще ближе к просвету канальца, вступает в профазу 1го деления созревания и начинает называться сперматоцитом 1го порядка (рис. 22, Б). В результате 1-го деления созревания Рис. 22. Семенник, поперечный срез извитых канальцев (А). Часть поперечного среза извитого семенного канальца (Б). Окраска гематоксилин-эозином. х900 (по Алмазову, 1978). А: 1 – белочная оболочка; 2 – сосудистая оболочка; 3 – извитые семенные канальцы; 4 – интерстиций яичка; 5 – просвет извитого канальца; 6 – сперматозоиды на разной стадии развития. Б: 1 – волокнистый слой; 2 – сперматогонии А; 3 – сперматогонии В; 4 – сперматоциты 1-го порядка; 5 – сперматоциты 2-го порядка; 6 – сперматозоиды; 7 – фолликулярная клетка (клетка Сертоли); 8 – интерстициальные клетки яичка (клетки Лейдига); 9 – миоидные клетки в составе собственной оболочки канальца V. Гаметогенез 109 сперматоцит 1-го порядка делится на два одинаковых сперматоцита 2-го порядка (рис. 22, Б), а последние в ходе 2-го деления созревания – на две сперматиды, обладающие, как и зрелая яйцеклетка, гаплоидным числом хромосом и количеством ДНК, соответствующим 1с. Затем каждая сперматида в результате сложных цитологических преобразований, не сопровождающихся клеточными делениями, преобразуется в сперматозоид (рис. 22, Б). Этот процесс спермиогенеза продолжается у разных видов животных несколько дней. Как сперматогонии, так и сперматоциты и сперматиды всех исследованных видов животных почти непрерывно связаны между собой цитоплазматическими мостиками, образуя синцитий. Этим, по-видимому, объясняется высокая степень синхронности делений сперматогониев и сперматоцитов в данном отделе семенного канальца. Деления эти идут последовательными циклами с периодом в несколько дней, а вдоль семенного канальца наблюдается волна сперматогониальных делений и вхождения в спермиогенез. Большое значение для сперматогенеза имеют соматические клетки, расположенные в стенках семенных канальцев, – так называемые клетки Сертоли (рис. 22, Б). К ним прикрепляются сперматиды и уже сформированные молодые сперматозоиды. По-видимому, клетки Сертоли снабжают сперматогониальные клетки питательными веществами и гормонами, способствуют высвобождению уже сформированных сперматозоидов в просвет канальцев, а также фагоцитируют неполноценные сперматозоиды. Основные процессы, протекающие в ходе спермиогенеза (преобразования сперматиды в сперматозоид), таковы (рис. 23): 1) ядро сперматиды сильно уплотняется, хроматин конденсируется и становится синтетически совершенно инертным (рис. 23, А); 2) происходят перемещения органелл клетки; аппарат Гольджи также уплотняется, прижимается к ядру и формирует так называемую акросому, смещается на апикальный конец клетки: центриоли, напротив, смещаются на противоположный полюс ядра и располагаются одна ближе к ядру (проксимальная центриоль), другая дальше (дистальная центриоль) (рис. 23, А, Б, В); 110 Болотов А. В. Биология размножения и развития Рис. 23. А – Ж – последовательные фазы спермиогенеза. 1 – ядро сперматиды; 2 – аппарат Гольджи; 3 – центриоли; 4 – зачаток акросомы; 5 – митохондрии; 6 – жгутик; 7 – акросома; 8 – проксимальная центриоль; 9 – дистальная центриоль; 10 – ядерное вещество, сосредоточенное в головке сперматозоида; 11 – митохондриальная спираль; 12 – остатки цитоплазмы; 13 – головка; 14 – шейка; 15 – средняя часть; 16 – хвост; 17 – концевой участок хвоста (по Карлсону, 1983) 3) из дистальной центриоли начинает расти жгутик, представляющий собой орган движения сперматозоида. У некоторых видов сперматозоиды лишены жгутика (круглые черви, ракообразные). Вокруг основания жгутика, если он есть, располагаются митохондрии в виде спирального чехлика; по-видимому, они поставляют энергию для движения жгутика (рис. 23, Б, В, Г, Д); 4) вся или почти вся цитоплазма отторгается; зрелый сперматозоид практически ее лишен (рис. 23, Е, Ж). Для каждого вида животных время, необходимое для превращения сперматогония в зрелый спермий (включая время пребывания в придатке), постоянно, хотя различия между видами существенны. Продолжительность спермиогенеза составляет, в днях: V. Гаметогенез 111 Сперматозоиды, законЖивотные дни чившие форми- Морской еж рование, попа4 дают в систему Петух семявыводящих 25 путей. Хряк 34 Гаметогенез отличается Кролик 41 высокой произ- Жеребец водительно42 стью. За всю ре- Баран продуктивную 49 жизнь человека Бык (в среднем 55– 54 60 лет) в яичках Кобель формируется 56 11 12 Человек сперматозоидов 8·10 – 4,5·10 23 (т. е. от 800 Мужчина млрд до 4,5 трлн), и лишь единичные из них участвуют в оплодотворении. Длительность сперматогенеза у человека составляет 72–75 дней, но не во всех семенных канальцах созревание сперматозоидов занимает одинаковый промежуток времени и происходит на протяжении извитого семенного канальца волнообразно, каждый час их в семенниках образуется от 100 до 200 млн (Гилберт С., 1995). Сложный процесс сперматогенеза регулируется гонадотропными гормонами гипофиза и стероидными гормонами яичка. После полового созревания гипоталамус начинает выделять гонадотропный рилизинг-гормон, под влиянием которого гипофиз секретирует фолликулостимулирующий гормон (ФСГ), стимулирующий развитие и функционирование клетки Сертоли и лютеинизирующий гормон (ЛГ), стимулирующий клетки Лейдига к выработке тестостерона. Клетки Лейдига (рис. 22, Б) – гормонопродуцирующие клетки млекопитающих, расположенные между семенными канальцами в семенниках, в них производится тестостерон и другие соединения андрогенного ряда, также в них образуется небольшое количество женских половых гормонов эстрогенов и прогестинов. Кроме андрогенов клетки Лейдига вырабатывают активин, стимулирующий выработку ФСГ. Таким образом, процесс сперматогенеза регулируется множеством гормонов. Соматотропный гормон гипофиза стимулирует рост и размножение сперматогоний. ГТРГ, воздействуя на гипофиз, вызывает синтез и секрецию ЛГ и ФСГ. ФСГ обеспечивает превращение сперматогоний в спермациты. ЛГ, пролактин и тестостерон (его синтез и секрецию стимулирует ЛГ) спо- 112 Болотов А. В. Биология размножения и развития собствуют превращению сперматоцитов в сперматиды. Сперматогенез протекает с влиянием ФСГ и тестостерона. У мужчин в возрасте от 25 до 45 лет содержание тестостерона в крови колеблется в пределах от 12 до 40 нмоль/л (колебания в течение суток достигают 30 %), максимальное его количество вырабатывается утром, а в годовом цикле – весной. 9. Нарушения сперматогенеза. Сперматогенез у мужчин Сперматогенез является одним из наиболее динамичных процессов в организме человека, связанных с клеточной регенерацией и дифференцировкой. Он протекает под контролем специфических генов развивающихся гамет и регулируется совокупностью гормонов, цитокинов и факторов роста. В сутки у взрослого мужчины в яичке образуется около 100–200 млн спермиев, что соответствует продукции примерно 70–150 тыс. клеток в минуту или 15 тыс. спермиев с каждым биением сердца. Вместе с тем в эякулят поступает значительно меньшее количество спермиев, чем то, что изначально образуется в семенных канальцах яичка, вследствие их частичной гибели в самом яичке и в семявыносящих путях. В последних, в особенности в придатке яичка, происходит дозревание и накопление этих клеток. При патологии в эякуляте уменьшается количество сперматозоидов (олигозооспермия), может снижаться число подвижных форм (астенозооспермия). Иногда отсутствуют зрелые сперматозоиды, а встречаются лишь клетки сперматогенеза. Все сперматозоиды могут быть неподвижными или в сперме могут отсутствовать как сперматозоиды, так и клетки сперматогенеза (аспермия). Патологические изменения морфологического строения сперматозоида могут проявляться в размерах головки (большая и маленькая), ее формы (конусовидная), структуре (аморфная или сдвоенная). Хвост сперматозоида бывает двойным или спиралевидным. Для оценки способности мужчины к зачатию необходимо сделать анализ его спермы, который называется спермограмма. • Нормозооспермия – норма; • Олигозооспермия – снижена концентрация сперматозоидов; V. Гаметогенез 113 • Тератозооспермия – изменена морфология сперматозоидов; • Астенозооспермия – снижена подвижность сперматозоидов; • Олигоастенозооспермия – сочетание 3-х вариантов патозооспермии; • Азооспермия – нет сперматозоидов в сперме; • Аспермия – нет спермы как таковой (объем спермы 0 мл). Количественные характеристики активности сперматогенеза Несмотря на длительную историю микроскопического изучения спермиев, насчитывающую более трех веков, впервые методы количественного анализа для оценки их концентрации в эякуляте были использованы лишь с 1929 г. в связи с проблемами бесплодия. В этих исследованиях было впервые сформулировано положение о связи концентрации спермиев в эякуляте с фертильностью мужчины, определены средние и пограничные значения этого показателя у здоровых лиц, которые составляли соответственно 100 млн и 60 млн спермиев в 1 мл. Хотя вскоре было показано, что у 15–25 % здоровых мужчин, имеющих детей, отмечается более низкая концентрация спермиев в эякуляте, однако величина, принятая за норму, в течение длительного времени не пересматривалась. В 70-х годах нижняя граница концентрации спермиев в эякуляте, условно разделяющая нормоспермию и олигозооспермию, была снижена до 40 млн/мл, в конце 20 в. (1999 г.) – до 20 млн/мл, в 2010 – 15 млн/мл. В последующем в оценку состояния спермиев, наряду с их концентрацией и общим содержанием в эякуляте (которое рассчитывают с учетом объема последнего), были введены два других основных показателя, отражаемые в спермограмме – их подвижность и морфологические характеристики, характерные для нормоспермии, которые тоже менялись соответственно с 70 % и 80 % в 70-х годах, 50 % и 30 % в 1999 г., до 40 % и 4 % в 2010 г. Современные тенденции изменения активности сперматогенеза Приведенные выше сведения об изменении величин количественных показателей состояния сперматогенной функции человека, принятых за норму, могут свидетельствовать либо о за- 114 Болотов А. В. Биология размножения и развития вышении первоначально принятых критериев, либо о действительно происходящем снижении активности сперматогенеза. Последнее было подтверждено исследованиями, проведенными с 1930 по 2010 гг., посвященным количественной оценке сперматогенной функции у нормальных мужчин в разных странах мира. В анализ были включены показатели эякулята 14 947 здоровых мужчин в возрасте 17–64 лет, обследованных в различных странах мира. При использовании линейного регрессионного анализа было установлено, что концентрация спермиев за последние 70 лет снизилась – с 113 млн/мл в 1940 г. до ≈66 млн/мл в 1999 г. (в 1,7 раза). За этот же период несколько уменьшился и средний объем эякулята – с 3,4 до 2,75 мл (в 1,2 раза). Возможные механизмы снижения активности сперматогенеза Описанные изменения сперматогенной функции у мужчин (количественные или качественные дефекты выработки спермиев) могут возникать в результате воздействия повреждающих факторов либо на внутриутробно развивающийся организм, либо на организм взрослого. Действие повреждающих факторов на взрослый организм Влиянию различных факторов на активность сперматогенеза у взрослого мужчины посвящена обширная литература. Основные неблагоприятные факторы можно условно разделить на три группы: химические, физические и бытовые. Химические факторы. Как и в случае антенатального воздействия, из химических факторов особое внимание уделяется разнообразным соединениям, которые имитируют эффекты эстрогенов или являются лигандами рецепторов андрогенов. Такие вещества способны вмешиваться в естественные пути эндокринной регуляции процессов гаметогенеза и стероидогенеза в мужском организме. К веществам с указанным действием относят естественные эстрогены растительного происхождения (фитоэстрогены), попадающие в организм с продуктами питания. V. Гаметогенез 115 Фитоэстрогенами богаты все бобовые растения, особенно соя (изофлавоны генистеин, дайдзеин, глицитеин), финики, имбирь, семена подсолнечника, семена льна содержат фитоэстрогены лигнаны, которые в кишечнике человека превращаются в энтеролактон и энтеродиол, пшено, проросший овес, проросшая пшеница и хмель (фитоэстроген 8-пренилнарингенин), который находится в пиве. Выраженным действием, снижающим количество и качество спермы, обладают и некоторые напитки, такие как кока-кола и пепси-кола (рис. 14, В). Литр колы в день снижает качество и количество эякулята примерно на 30 % и вырабатывалось в среднем вдвое меньше семенной жидкости. Однако несравненно большую опасность представляют разнообразные группы искусственно созданных (антропогенных) химических соединений, обладающих эстрогенной или антиандрогенной активностью, которые практически повсеместно все в большей степени загрязняют окружающую среду. Эта группа соединений включает широко применяемые в промышленности химические вещества, компоненты топлива и соединения, образующиеся при сгорании нефтепродуктов – полициклические ароматические углеводороды, полихлорированные бифенилы, диоксины, эфиры фталата, алкилфенольные соединения, а также используемые в сельском хозяйстве и животноводстве хлорорганические пестициды, инсектициды и фунгициды (например, ДДТ). Указанные вещества проникают в организм на производстве и в быту – с водой, вдыхаемым воздухом и всеми видами пищи (как растительной, так и животной). Некоторые из них оказывают токсическое влияние непосредственно на дифференцирующиеся половые клетки, другие действуют на гормональные механизмы на уровне гипоталамуса и гипофиза. Часть их оказывает дополнительное неблагоприятное действие на репродуктивную функцию мужчин путем изменения характера секреции предстательной железы и семенных пузырьков. Помимо веществ с эстрогенным и антиандрогенным эффектом, угнетению сперматогенной функции могут способствовать соединения, оказывающие токсическое воздействие на сперматогенные клетки, клетки Лейдига, гипоталамус, гипофиз и нарушающие механизмы обратной связи в системе гипоталамус – гипофиз – яички. К таким веществам относят тяжелые металлы, 116 Болотов А. В. Биология размножения и развития нейротропные яды (фенол, толуол, бензин, хлорид аммиака), которые встречаются на производстве в различных отраслях промышленности и постоянно попадают в окружающую среду. В данный список необходимо включить и широкий спектр фармакологических препаратов – седативные средства и антидепрессанты, некоторые антибиотики и все сульфаниламиды, кетоконазол, диуретики, гиполипидемические средства, гормоны (эстрогены, андрогены), блокаторы гистаминовых рецепторов (противоязвенные препараты), антигипертензивные, средства химиотерапии опухолей. Физические факторы. Из физических факторов, влияющих на сперматогенез, наибольшее действие оказывают температура, облучение и вибрация. Температура, при которой протекает сперматогенез, оказывает влияние на его количественные и качественные показатели: повышение температуры тела (в связи с действием производственных факторов, при лихорадочных состояниях, частом приеме горячих ванн, посещении сауны) вызывает подавление сперматогенеза. Сперматогенез, как известно, страдает также при таких заболеваниях, как крипторхизм (одно или оба яичка не находятся в мошонке) и варикоцеле (расширение вен в семенном канатике), при которых одним из повреждающих факторов служит повышенная интратестикулярная температура. Как показали недавние исследования, даже умеренное локальное перегревание яичек вследствие ношения плотно прилегающего белья способно оказывать повреждающее влияние на сперматогенез. Хорошо известным фактором, нарушающим сперматогенную функцию, является облучение, эффект которого зависит от общей дозы, кратности и длительности воздействия, возраста, в котором оно происходит, и пубертатного статуса. В последние годы особое внимание уделяется долгосрочному влиянию низких доз облучения, с которым некоторые авторы связывают снижение активности сперматогенеза. Физическим фактором, оказывающим повреждающее влияние на сперматогенез является вибрация, которая служит фактором профессиональной вредности у рабочих ряда специальностей (водители, механизаторы, горняки и др.). У таких лиц повышена частота олигозооспермии и азооспермии, снижены объ- V. Гаметогенез 117 ем эякулята и доля подвижных спермиев, повышена частота морфологически аномальных спермиев (рис. 14, В). Вибрационную болезнь рассматривают как дисгормональное состояние с гиперпродукцией кортикостероидов. Бытовые факторы. Влияние бытовых факторов связано с индивидуальными особенностями образа жизни, которые систематически, а в последние годы – охватывая все более широкие массы – оказывают угнетающее влияние на сперматогенез. Хорошо известно, что алкоголь способен вызвать тяжелые нарушения сперматогенеза, повреждая сперматогенные клетки и клетки Лейдига, нарушая метаболизм половых стероидов, поражая гипоталамус и гипофиз. В яичке алкоголиков гистологически выявляется атрофия клеток Лейдига и извитых семенных канальцев с потерей сперматогенных клеток (вплоть до полной – синдрома «только клетки Сертоли»), снижается содержание зрелых спермиев и доли подвижных и морфологически нормальных форм, развивается фиброз яичка (рис. 14, В). Более 80 % хронических алкоголиков стерильны. Степень нарушения сперматогенеза отчетливо связана с количеством потребляемого алкоголя. При ежедневной дозе 80– 160 г и выше нормальный сперматогенез сохраняется только у 21–37 % мужчин, у 54–74 % отмечается частичное или полное нарушение сперматогенеза, у 4–9 % – синдром «только клетки Сертоли». Тот факт, что у пятой части алкоголиков сперматогенез сохраняется на достаточно высоком уровне, указывает на существенные индивидуальные различия в чувствительности к алкоголю. Важным фактором, оказывающим негативное влияние на активность сперматогенеза, является курение. У курящих снижены секреция тестостерона яичком, концентрация спермиев в эякуляте, их подвижность, оплодотворяющая способность, доля морфологически, генетически и функционально нормальных клеток. Этот эффект связывают с нарушением деятельности клеток Лейдига и Сертоли, а также с прямым цитотоксическим влиянием на сперматогенные клетки. Тяжелые расстройства сперматогенеза при систематическом употреблении наркотиков (в особенности марихуаны, каннабиса и героина) часто проявляются олигоастенозооспермией и некроспермией. На ультра- 118 Болотов А. В. Биология размножения и развития структурном уровне выраженные дегенеративные изменения отмечаются практически во всех отделах зрелых спермиев (рис. 14, В). К факторам, угнетающим сперматогенез, относят действие выраженного стресса (в том числе психологического), которое опосредуется рядом гормональных сдвигов – снижением уровня тестостерона и дигидроандостерона при повышении концентраций кортикостероидов. 10. Отличия сперматогенеза и оогенеза 1. При сперматогенезе более интенсивно и продолжительно идет размножение. За время половой жизни мужчины образуется от 800 млрд до 4,5 трлн сперматозоидов. На 5-м месяце эмбриогенеза в зачатке женской половой железы насчитывается 6 млн оогоний. От момента полового созревания до прекращения гаметогенеза созревает 300–400 ооцитов. 2. Стадия роста более выражена при овогенезе (состоит из 2 периодов: малого роста и большого роста), что связано с накоплением желтка. 3. Неравномерное распределение цитоплазмы при мейозе (стадия созревания) характерно для овогенеза. В сперматогенезе из одного гаметотоцита 1-го порядка образуются четыре сперматиды, при овогенезе – одна яйцеклетка и три направительных (редукционных) тельца. 4. Стадия формирования имеется только при сперматогенезе: сперматиды приобретают форму, характерную для сперматозоидов данного вида. 5. В оогенезе процесс созревания блокируется на какой-то стадии мейоза. В сперматогенезе нет блока мейоза. 119 V. Гаметогенез 11. Эволюция гаметогенеза Эволюция гаметогенеза осуществлялась по пути большей специализации и расширения функций вспомогательных структур. Фолликулярный эпителий имеет несколько функций: 1) изолирующая; 2) барьерная; 3) транспортная; 4) синтетическая (синтез трофических веществ); 5) гормонально-регуляторная (синтез гормонов и гормоноподобных веществ, регулирующих гаметогенез); 6) фагоцитарная (способен к фагоцитозу погибших половых клеток); 7) опорно-механическая; 8) морфогенетическая; 9) структурная (в оогенезе вырабатываются вещества для формирования вторичной оболочки яйца и др.). Таким образом, характеризуя взаимоотношения половых клеток с организмом в филогенезе, следует отметить две тенденции: • всё большую изоляцию половых клеток от внутренней среды организма; • всё большую зависимость развития половых клеток от вспомогательных элементов гаметогенеза. VI. ОПЛОДОТВОРЕНИЕ Оплодотворение (fertilisatio) – слияние мужской и женской половых клеток, в результате чего восстанавливается диплоидный набор хромосом, характерный для данного вида животных, и возникает качественно новая клетка – зигота (оплодотворенная яйцеклетка или одноклеточный зародыш). У животных оплодотворению предшествует осеменение, т. е. процесс, обеспечивающий встречу гамет. У животных различают два типа оплодотворения: моноспермию и полиспермию. Когда для оплодотворения требуется один сперматозоид, то это моноспермия. Она характерна для иглокожих, млекопитающих и др. Если в яйцо проникает более одного спермия, то это вызывает нарушение процесса развития, уродства. Если для оплодотворения требуется больше одного сперматозоида, то такой процесс называют полиспермией. Это наблюдается у некоторых членистоногих (насекомые, паукообразные, брюхногие), моллюсков, хордовых (акулообразные и химеровые рыбы), хвостатых амфибий, рептилий и птиц. Для оплодотворения яйца у голубя требуется 15–20 сперматозоидов. У животных встречается три вида осеменения: 1) наружное; 2) смешанное; 3) внутреннее. 1. Наружное осеменение. При наружном осеменении и сперматозоиды, и яйцеклетки попадают во внешнюю среду, где и происходит их взаимодействие и оплодотворение. Лучше всего для этих целей подходит водная среда. Так осуществляется, например, оплодотворение у рыб и бесхвостых амфибий. Рыбы во время нереста чаще всего выметывают икру большими порциями, а самцы тут же поливают ее семенной жидкостью. Такой способ экономически невыгоден. Во время нереста множество икринок могут остаться неоплодотворенными, так как на эту процедуру отпущено очень короткое время. В воде их оболочки быстро затвердевают, становятся непроницаемыми, слияние мужских и женских половых клеток – оплодотворение становится невозможным. Кроме того, половые клетки могут просто не встретиться. VI. Оплодотворение 121 2. Смешанный тип осеменения. У хвостатых амфибий наружное оплодотворение более экономично, так как икринки и семенная жидкость упакованы в специальные мешочки, которые попадают в воду, а затем самки сами захватывают их клоакой, после чего сперматофоры оказываются внутри тела самки. В этих условиях половым клеткам встретиться гораздо проще, и временной фактор не играет существенной роли. У них сперматозоиды из сперматофора проникают в слизистый икряной мешочек и там оплодотворяют яйца. Так происходит осеменение у хвостатых амфибий (тритоны, амбистомы) и др. 3. Внутреннее осеменение. При внутреннем осеменении выделяемая самцом семенная жидкость, содержащая сперматозоиды, вводится в половые пути самки. Этот тип осеменения характерен для всех наземных позвоночных (рептилий, птиц и млекопитающих). У человека сперматозоиды достигают оплодотворяющей способности в нижних отделах половых путей мужчины. После эякуляции они быстро поднимаются по половым путям женщины за счет сокращений матки и маточных труб, а собственная их подвижность становится важной на более поздних этапах. Только небольшое число из них достигает места оплодотворения в ампуле трубы, и только один из нескольких миллионов спермиев эякулята оплодотворит яйцеклетку. У наземных животных -внутреннее осеменение, при котором самцы с помощью копулятивных органов вводят сперматозоиды в половые пути самки. У животных сперма может попадать во влагалище или в матку. Влагалищный тип осеменения имеют кролики, крупный рогатый скот, овцы, козы, обезьяны и человек. Маточный тип осеменения имеют свиньи, лошади, собаки, грызуны. Оплодотворение – слияние гамет с образованием зиготы с диплоидным набором хромосом. Оплодотворению предшествует осеменение – излияние семенной жидкости в половые пути при внутреннем оплодотворении или в среду, где находятся яйцеклетки, при наружном оплодотворении. В процессе оплодотворения различают три фазы: 1) дистантное взаимодействие и сближение гамет; 2) контактное взаимодействие и активизация яйцеклетки; 122 Болотов А. В. Биология размножения и развития 3) вхождение сперматозоида в яйцо и последующее слияние – сингамия. 1. Дистантное взаимодействие и сближение гамет Обеспечивается совокупностью ряда неспецифических факторов, повышающих вероятность столкновения половых клеток. Сближению гамет способствуют: I. Координирование процессов оогенеза и сперматогенеза. II. Приспособления, обеспечивающие попадание созревших гамет в места, где происходит оплодотворение. III. Факторы, способствующие оплодотворению: 1. Концентрация спермы. Оптимальная – 40–60 млн в 1 мл. 2. Концентрация водородных ионов или рН-среды. В более щелочной среде спермии становятся более подвижными, но менее жизнеспособными. 3. Концентрация СО2. Выделяется самими спермиями. 4. Тип сперматозоида. Y-спермии более активные, но менее жизнеспособные, чем X-спермии. 5. Избыточная продукция сперматозоидов. В норме должно быть 150–240 млн спермиев. 6. Крупные размеры яйцеклеток. Диаметр яйцеклеток у человека от 100 до 200 мкм, что в среднем составляет 150 мкм. IV. Влияние химических веществ, способствующих сближению и взаимодействию половых клеток. Важную роль в этом играют химические вещества, вырабатываемые половыми клетками, – гамоны: гиногамоны (I, II), вырабатываемые яйцеклетками и андрогамоны (I, II), продуцируемые спермиями. Гиногамоны I (низкомолекулярные вещества небелковой природы, выделяемые яйцеклетками, активизируют движение спермиев. Гиногамоны II (фертилизины) – видоспецифические белки, вызывающие склеивание спермиев при их реакции с комплементарным андрогамоном II, встроенным в цитолему спермия. Склеивание спермиев предохраняет яйцеклетку от проникновения многих спермиев. Андрогамоны I – антагонисты гиногамонов I – вещества небелковой природы, подавляют подвижность спермиев. Андрогомоны II (белковой природы) участвуют в растворении оболочки яйца. VI. Оплодотворение 123 Попав в женские половые пути во время своего движения сперматозоид обычно вращается вокруг своей оси. Скорость движения сперматозоида человека может достигать 0,1 мм в 1 с или более 30 см в 1 ч. У человека приблизительно через 1–2 ч после коитуса с эякуляцией первые сперматозоиды достигают ампулярной части фаллопиевой трубы (той части, где происходит оплодотворение). Только каждый 5-й сперматозоид движется в правильном направлении, после эякуляции. Движение сперматозоидов по половым путям женщины является самостоятельным и осуществляется против движения жидкости, т. е. имеют отрицательный реотаксис. Для осуществления оплодотворения сперматозоидам необходимо преодолеть путь длинной около 20 см (цервикальный канал – около 2 см, полость матки – около 5 см, фаллопиева труба – около 12 см). Среда влагалища является губительной для сперматозоидов, семенная жидкость нейтрализует влагалищные кислоты и частично подавляет действие иммунной системы женщины против сперматозоидов. Из влагалища сперматозоиды движутся по направлению к шейке матки. Направление движения сперматозоид определяет, воспринимая pH окружающей среды. Он движется по направлению уменьшения кислотности от влагалища около pH = 6,0 к шейке матки pH = 7,2. Для успешного оплодотворения в матку должно проникнуть не менее 10 млн сперматозоидов. Из матки сперматозоиды направляются в фаллопиевы трубы, направление к которым и внутри которых сперматозоиды определяют по току жидкости. 2. Контактное взаимодействие и активизация яйцеклетки Она включает: 1) контактное взаимодействие гамет; 2) проникновение спермия в яйцеклетку, которое осуществляется с помощью акросомы и ее ферментов спермолизинов. Плазматические мембраны в месте контакта половых клеток сливаются и происходит плазмогамия – объединение цитоплазм обеих гамет. 124 Болотов А. В. Биология размножения и развития Сближение половых клеток происходит за счет пассивного движения яйцеклетки (ооцита с оболочками) вместе с током жидкости по яйцеводу к матке и вначале (во влагалище и матке) тоже пассивного, а затем (в яйцеводе) активного движения сперматозоидов. а) В яйцеводе на относительно больших расстояниях от яйцеклетки направление активного движения сперматозоидов обеспечивается отрицательным реотаксисом – способностью регистрировать направление тока жидкости и двигаться противоположно ему. б) На малых расстояниях могут иметь значение два других вида таксиса: 1) хемотаксис – движение сперматозоидов по градиенту концентрации гиногамонов – специальных химических веществ, выделяемых яйцеклеткой; 2) электротаксис – электрическое взаимодействие между разноименно заряженными белками сперматозоидов и яйцеклетки. Капацитация Во время перемещения по женским половым путям в сперматозоидах происходит капацитация – процесс активизации. В это время: а) метаболизм и подвижность сперматозоидов усиливаются; б) плазмолемма головки теряет некоторые поверхностные гликопротеины, что придает ей способность связываться с внешней оболочкой яйцеклетки, а также лабильность, необходимую для последующего разрыва в ходе высвобождения ферментов акросомы. Возможно, что капацитацию инициируют гиногамоны II, выделяемые ооцитом, и, вероятно, адреналин (содержащийся в семенной жидкости). Достигая яйцеклетки (ооцита II), многочисленные сперматозоиды (у млекопитающих) проникают между достаточно рыхло расположенными фолликулярными клетками зернистой оболочки и связываются с блестящей оболочкой. В связывании участвуют специфические рецепторы – как со стороны плазмолеммы головки сперматозоида, так и со стороны блестящей оболочки ооцита. Именно эта специфичность во многом обусловливает VI. Оплодотворение 125 трудность или полную невозможность скрещивания разных животных. Благодаря биениям жгутиков сперматозоидов образовавшийся комплекс обычно начинает вращаться с частотой четыре оборота в минуту. Одновременно продолжается пассивное поступательное движение в яйцеводе по направлению к матке. Рис. 24. Акросомная реакция у млекопитающих После капацитации следует акросомальная реакция, которая индуцируется только при контакте сперматозоида с клетками яйценосного бугорка. Чтобы оплодотворение произошло необходимы, помимо прочего, прогестерон, кальций и, как выяснилось, их белковый посредник. Прогестерон воздействует на порообразующий белок (кальциевый ионный канал CatSper), расположенный на хвостике сперматозоида; это приводит к резкому увеличению его пропускной способности и, соответственно, к быстрому увеличению концентрации Са2+ в хвостике сперматозоида. После этого запускается кальций-зависимый ответ сперматозоида, приводящий к созреванию и внедрению сперматозоида в яйцеклетку. Прокачивая ионы через мембрану жгутика, CatSper провоцирует и увеличение концентрации кальция в головной части сперматозоида. Дополнительные ионы кальция «заставляют» сперматозоид высвободить содержимое пузырька передней части – акросо- 126 Болотов А. В. Биология размножения и развития мы. Содержимое акросомы и «переваривает» защитную оболочку (zona pellucida) – мужская гамета проникает в яйцеклетку. У всех животных кальций – ключевой элемент во многих процессах, связанных с созреванием половых клеток и оплодотворением. Обнаружение особого мембранного рецептора, напрямую связанного с регуляцией кальция, – важная веха в понимании динамики этих биологических процессов. За счет изменения содержания кальция мужские и женские клетки передают сигналы друг другу. У растений процесс роста пыльцевых трубок к яйцеклетке (по аналогии с движением сперматозоида к яйцу) и последующее оплодотворение также опосредованы изменениями концентрации кальция. Сперматозоиды млекопитающих просто напичканы белками CRISP (всего четыре разновидности). CRISP всех форм и размеров – это белки, управляющие ионными каналами сперматозоидов и клеток, входящих в структуру мужского полового тракта. Примечательно то, что эти же белки входят в состав яда рептилий. Ферменты, выделяемые из акросом, – пенетраза и коллагеназа осуществляют диссоциацию и удаление оставшихся фолликулярных клеток, спермолизины (акрозин, трипсин, гиалуронидаза), разрушают лучистый венец, расщепляют гликозаминогликаны блестящей оболочки яйцеклетки. Отделяющиеся фолликулярные клетки склеиваются в конгломерат, который вслед за яйцеклеткой перемещается по трубе благодаря мерцанию ресничек эпителиальных клеток слизистой оболочки. И сперматозоид как бы «пробуравливает» для себя собственный узкий канал (рис. 24). Один из сперматозоидов, а именно тот, который первый преодолел эти оболочки, прикрепляется к плазмолемме яйцеклетки (ооцита II). В данном месте образуется выпячивание цитоплазмы яйцеклетки – воспринимающий бугорок (рис. 25, 8). В ооплазму проникает головка и промежуточная часть хвостового отдела (рис. 24, 4; 25, 9). После вхождения сперматозоида на периферии ооплазмы происходит уплотнение ее (кортикальная реакция) и образуется оболочка оплодотворения (рис. 25, 10). VI. Оплодотворение 127 Рис. 25. Стадии оплодотворения и начала дробления (схема). 1 – ооплазма; 1а – кортикальные гранулы; 2 – ядро; 3 – блестящая оболочка; 4 – фолликулярный эпителий; 5 – спермий; 6 – направительные тельца; 7 – деление созревания ооцита; 8 – бугорок оплодотворения; 9 – головка спермия с ядром; 10 – оболочка оплодотворения; 11 – мужской пронуклеус; 12 – женский пронуклеус; 13 – синкарион; 14 – первое митотическое деление зиготы; 15 – бластомеры Как показано на беспозвоночных, механизм кортикальной реакции включает: приток ионов натрия через участок мембраны сперматозоида, встроенный в поверхность яйцеклетки после завершения акросомальной реакции. В результате отрицательный мембранный потенциал клетки становится слабоположительным. Приток ионов натрия обусловливает высвобождение ионов кальция из внутриклеточных депо и увеличение его содержания в цитоплазме яйцеклетки. Вслед за этим начинается экзоцитоз кортикальных гранул. Освобождающиеся из них протеолитические ферменты разрывают связи между блестящей оболочкой (или желточной оболочки у амфибий и птиц) и плазмолеммой яйцеклетки, а также между спермиями и прозрачной оболочкой. Кроме того, выделяется гликопротеид, связывающий воду и привлекающий ее в пространство между плазмолеммой и блестящей оболочкой. Вследствие этого формируется перивителлиновое пространство. Наконец, выделяется фактор, способствующий затвердению прозрачной оболочки и образованию из нее оболочки оплодотворения (fmembrana fertilisationis). 128 Болотов А. В. Биология размножения и развития Кортикальная реакция – один из механизмов, препятствующий проникновению в яйцеклетку других сперматозоидов. Проникновение сперматозоида через несколько минут значительно усиливает процессы внутриклеточного обмена, что связано с активизацией ферментативных систем яйцеклетки, в частности окислительно-восстановительных, а позднее – белковых синтезов. Активация яйцеклетки происходит в результате контакта сперматозоида с яйцеклеткой и заключается в сложных структурных и физико-химических изменениях. Благодаря тому что участок мембраны сперматозоида проницаем для ионов натрия, последние начинают поступать внутрь яйца, изменяя мембранный потенциал клетки. Затем в виде волны, распространяющейся из точки соприкосновения гамет, происходит увеличение содержания ионов кальция, вслед за чем также волной растворяются кортикальные гранулы. Выделяемые при этом специфические ферменты способствуют отслойке желточной оболочки. У таких животных, как морской еж, костистые рыбы и земноводные, все изменения цитоплазмы сопровождаются морфологическими перестройками. Эти явления получили название расслоения или сегрегации плазмы. Активация яйцеклетки завершается началом синтеза белка на трансляционном уровне, поскольку мРНК, тРНК, рибосомы и энергия были запасены еще в овогенезе. Активация яйцеклетки может начаться и протекать до конца без ядра сперматозоида и без ядра яйцеклетки, что доказано опытами по энуклеации зиготы. Яйцеклетка в момент встречи со сперматозоидом обычно находится на одной из стадий мейоза с помощью специфического фактора. У большинства позвоночных этот блок осуществляется на стадии метафазы 2-го деления; у многих беспозвоночных, а также у трех видов млекопитающих (лошади, собаки и лисицы) блок происходит на стадии диакинеза. В большинстве случаев блок мейоза снимается после активации яйцеклетки вследствие оплодотворения. Вслед за проникновением спермия в яйцеклетку и усилением окислительно-восстановительных реакций начинается интенсивное перемещение составных частей цитоплазмы (ооплазмы) с образованием зон повышенной концентрации желточных и пигментных гранул, органелл, что носит название ооплазматической сегрегации. Методом маркировки установлено, что в ходе 129 VI. Оплодотворение дальнейшего развития каждый участок оплодотворенной яйцеклетки даст начало определенной структуре зародыша. Такие участки называются презумптивными (от лат. praesumptio – предположение, основанное на вероятности). 3. Сингамия Попавшая в яйцеклетку головка спермия поворачивается на 180°, ядро постепенно набухает, округляется, хроматин разрыхляется, и оно превращается в мужской пронуклеус (рис. 25, 11). Центриоли, внесенные мужской половой клеткой, становятся при этом центром движения внутри оплодотворенной яйцеклетки (зиготы). В то время как в яйцеклетке завершается мейоз, ядро сперматозоида, проникшее в нее, видоизменяется. Оно принимает вид интерфазного, а затем профазного ядра. В это время удваивается ДНК и мужской пронуклеус получает количество наследственного материала, соответствующего 2с, т. е. содержит гаплоидный набор редуплицированных хромосом. Ядро яйцеклетки, закончившее мейоз, превращается в женский пронуклеус (рис. 25, 12), также приобретая наследственноый материал 2с. Оба пронуклеуса проделывают сложные перемещения, которые получили название «танца ядер», затем сближаются и сливаются (синкарион), образуя общую метафазную пластинку. Это, собственно, и есть момент окончательного слияния гамет – сингамии. В результате из двух гаплоидных гамет образуется одна диплоидная клетка, называемая зиготой. Объединение двух пронуклеусов – синкарион (рис. 25, 13) (от греч. sin – связь, karyon – ядро) – приводит к восстановлению характерного для данной особи животного или человека диплоидного набора хромосом. Таким образом, зигота приобретает гены, унаследованные от обоих родителей. В реализации наследственной информации, кроме ядер, половых клеток, важная роль принадлежит цитоплазме клетки. Об этом свидетельствуют эксперименты с пересадкой ядер соматических клеток в яйцеклетку. При этом пол развивающегося организма зависит от половых хромосом. При слиянии яйцеклетки со сперматозоидом, несущим хромосому X, образуется женская особь, а при слиянии со сперматозоидом, имеющим хромосому Y, – мужская особь. 130 Болотов А. В. Биология размножения и развития В оплодотворении выделяют две фазы: – внешняя фаза начинается с дистантного взаимодействия гамет и заканчивается образованием оболочки оплодотворения (рис. 25, 9); – во внутренней фазе выделяют стадии женского и мужского пронуклеусов, синкариона и сингамии. 4. Особенности репродукции человека 1. Человек – биосоциальное существо, может сознательно регулировать свою сексуальную жизнь. 2. Способность к репродукции у женщин возможна с 14–16 лет и до 45–50 лет; у мужчин с 16–18 лет и до старости. 3. Образование гамет не имеет сезонного характера. 4. Особенности оплодотворения у человека: • эякулят спермы должен содержать не менее 150 млн. сперматозоидов, • яйцеклетка жизнеспособна 24 часа, сперматозоид способен к оплодотворению в течение 24–48 часов, • сперматозоид проникает в ооцит 2-го порядка, • через 24 часа после оплодотворения происходит 2-е деление мейоза с образованием яйцеклетки и 2-го редукционного тельца, а спустя 10 часов наступает сингамия. 5. Чем старше женщина, тем большая вероятность рождения ребенка с генетическими дефектами. Возраст отцов способствует увеличению у потомства патологии, обусловленной генными мутациями. 5. Детерминация пола. Влияние факторов среды на развитие признаков пола У высших организмов значение среды в определении пола, как правило, невелико. Вместе с тем известны случаи, когда при определенных условиях происходит переопределение пола, так как эмбрионы всех животных изначально бисексуальны. Известно, что у млекопитающих дифференцировка пола происходит рано, поэтому полное переопределение пола невозможно. Известны случаи у крупнорогатого скота, когда в разнополых двойнях рождались бесплодная телочка – интерсекс (фри- VI. Оплодотворение 131 мартин) – и бычок. Это происходило из-за того, что гормоны мужского эмбриона начинали синтезироваться раньше, чем гормоны женского эмбриона. У некоторых видов происходит переопределение пола. Так, в Тихом океане обитают рыбки Labroides dimidiatus, живущие стайками из самок и одного самца. Каждый член кроме самца находится в состоянии стресса, источником которого является сам самец. При этом можно выделить разные уровни самок. Гибель самца сбрасывает напряжение с главной самки, и она становится самцом. Описанное явление зависит от гормонов выделяемых клетками надпочечников. У морского червя Bonella viridis пол зависит от того, будет зигота развиваться в непосредственном контакте с материнским организмом или самостоятельно. Искусственное изменение условий среды Проводилось с рыбами, амфибиями, птицами. Если кормить мальков самцов аквариумных рыб до 8 месяцев кормом с добавлением женских половых гормонов (эстрогенов), приводило к превращению их в самок. Новые данные о детерминации пола у птиц Недавние исследования на гинандроморфных (у которых одна половина мужская, а другая – женская; это крайне редкое природное отклонение) цыплятах дали неожиданный результат (рис. 26). Оказалось, что эти животные – истинные химеры, то есть представляют собой «коктейль» из клеток, несущих хромосомы ZZ (определяющие мужской фенотип у птиц), и ZW (определяющие женский фенотип). Эти данные посеяли сомнения в том, что для птиц верна классическая модель детерминации пола, когда фенотип большей частью определяется гормональной секрецией половых желез, а те, в свою очередь, начинают развиваться в ответ на активацию некоего «полового» гена (например, у млекопитающих такой ген называется Sry и определяет развитие яичек). Если же организм цыпленка, несмотря на общий гормональный фон, разделяется на две разнополые половины, это говорит о том, что соматические клетки птиц определяют свой пол автономно, а не генерализованно, как считалось раньше. 132 Болотов А. В. Биология размножения и развития Чтобы проверить это предположение, были проведены эксперименты на искусственных химерах. Для этого мезодермой двухдневных GFP-эмбрионов, находящихся на 12-й стадии развития, заменяли аналогичный участок в левой части не GFP-эмбрионов такого же возраста (поскольку у птиц полностью развивается только левый яичник). На 35-й стадии развития (9 дней инкубации) эмбрионы исследовали и обнаружили (с помощью иммуногистохимии), что в разнополых химерах донорские клетки хранят верность своему первоначальному полу: то есть мужские клетки, будучи помещенными в женское общество, не начинают вырабатывать ароматазу, а женские, оказавшиеся в мужской гонаде, – наоборот, не вырабатывают анти-мюллеров гормон. Более того, одна из полученных химер образовала яичнико-яичко – то есть передняя его часть состояла из женских клеток и экспрессировала ароматазу, а задняя – из мужских клеток и вырабатывала анти-мюллеров гормон, что стало дополнительным свидетельством автономной половой детерминации клетками птиц. VII. ДРОБЛЕНИЕ Оплодотворение играет чрезвычайно важную роль в развитии, но это лишь его первая ступень. Зигота ее генетическим материалом и определенным распределением цитоплазмы приступает к созданию многоклеточного организма. У всех известных животных это наблюдается в процессе дробления – серии митотических делений, в результате которых объем цитоплазмы яйца разделяется на клетки меньшего размера, которые называются бластомерами. У большинства видов в период дробления объем зародыша не увеличивается и огромная масса цитоплазмы зиготы разделяется на всё более мелкие клетки. Это деление без роста объема сопровождается выпадением интерфазного периода роста между делениями, тогда как деление ядер следует друг за другом с большой скоростью, которая никогда не наблюдается в дальнейшем. Одним из факторов, влияющих на скорость делений является соотношение ядра и цитоплазмы, существенно отличающееся от такового у соматических клеток. В процессе делений дробления скорость их постепенно уменьшается. Таким образом дробление начинается сразу после оплодотворения и заканчивается, когда у зародыша достигается новое равновесие между ядром и цитоплазмой, соответствующее соматическим клеткам. На ранних стадиях деления дробления фаза S занимает очень маленькое время (≈10 мин), а в дальнейшем оно растет как в процессе дробления, так и в общем эмбриогенезе. Например, в период гаструляции фаза S ≈ 1 ч, а во время нейруляции – 5 ч. У некоторые животные после завершения дробления наступает пауза, которая заканчивается в определенные сроки. У летучих мышей осеменение происходит поздней осенью, тогда как развитие яйца начинается весной. Пауза типична и для ракообразных. Скорость дробления изменяется под влиянием температуры. Существует следующая закономерность: при изменении температуры на 10 ºС деление дробления ускоряется (замедляется) в 2–3 раза. Ранний период развития характеризуется синхронностью деления. Возможно, что деление дробления является еди- 134 Болотов А. В. Биология размножения и развития ницей для регуляции продолжительности различных периодов развития в дальнейшем. Поздний период дробления называется бластуляцией. Он характеризуется: 1) удлинением фазы S; 2) ускорением синтетических процессов. Геном зародыша начинает функционировать не сразу, а после прохождении определенного числа делений дробления и нормализации ядерно-плазменных отношений. По мере дробления включаются факторы взаимодействия бластомеров, факторы интеграции зародыша, а это ведет к тому, что темпы деления клеток в разных частях зародыша становятся неодинаковыми. Угнетение одних частей зародыша на ранних стадиях онтогенеза. Если темпы дробления сохранились бы прежними и дробление было бы синхронным, то через 20 ч развития яйца, например, лягушки (начало гаструляции) зародыш бы состоял из 1 млн клеток, а через 2 суток – 25·1013 клеток. К моменту ранней хвостовой почки (к концу седьмых суток) количество клеток было более 1028, а размер зародыша оказался бы 5,2·1012 м3. И это было бы при диаметре клеток ≈ 10 мкм. В 1962 г. Уильям С. Буллоу в межклеточной среде обнаружил вещества, концентрация которых подавляет деление клеток. Они были названы кейлоны (англ. кеу – ключ, long – единственный), которые тормозят в окружающих клетках реакции митоза, скорость синтеза нуклеиновых кислот и белков, и при этом не обладают видовой специфичностью. Концентрация этих веществ зависит от числа клеток в данном объёме, а количество кейлона, вырабатываемое одной клеткой является величиной постоянной. Итак, дробление – строго координированный процесс, находящийся под генетическим контролем. Образование многоклеточности – первая и основная функция дробления. Другая функция – в увеличении ядерно-плазменного отношения. Видовые особенности этого процесса определяются двумя основными параметрами: 1) количеством и распределением желтка в цитоплазме; 2) присутствием в цитоплазме факторов, влияющих на ориентацию митотического веретена и время его образования. Если желток в яйце распределен неравномерно, то в области, содержащей меньше желтка (анимальный полюс), клеточные VII. Дробление 135 деления идут с большей скоростью, чем в противоположной богатой желтком области (вегетативный полюс). Ядро зиготы в этом случае сдвигается к анимальному полюсу. То есть можно сказать следующее – желток подавляет дробление. Правила зависимости между расположением желтка и положением ядер и веретен были сформулированы немецким ученым О. Гертвигом на основе правил, предложенных ранее ботаником Ю. Саксом для растительных меристем. Эти правила состоят в следующем: 1) клеточное ядро стремится расположиться в центре свободной от желтка цитоплазмы; 2) веретено клеточного деления стремится расположиться по направлению наибольшего протяжения свободной от желтка цитоплазмы. Перетяжки, разделяющие дробящуюся зиготу на всё более мелкие клетки (бластомеры), называют бороздами дробления. Борозды дробления делят на следующие виды: 1) меридианная (меридиональная) – проходит через анимально-вегетативную ось зиготы; 2) экваториальная, которая может быть смещена к анимальному полюсу из-за большого количества желтка в вегетативном полюсе, проходит по широте зиготы (поперек) или по экватору зиготы; 3) тангенциальная – проходит параллельно поверхности зиготы, так образуется несколько слоев клеток зародыша. У зигот, содержащих относительно мало желтка (изолецитальные, мезолецитальные яйца), в процессе деления борозда дробления проходит через всё яйцо, то говорят, что дробление полное, или голобластическое. Зиготы, содержащие много желтка (полилицетальные яйца) и по расположению желтка находящегося в вегетативной области (телолецитальные яйца), в процессе деления борозды дробления не проникают в богатую желтком область цитоплазмы, т. е. не проходят через все яйцо, в этом случае дробление неполное или меробластическое. Однако желток – только один из факторов, оказывающий влияние на характер дробления зиготы. Существуют также унаследованные особенности клеточных делений, которые накладываются на влияние желтка. Это можно увидеть на примере изолецитальных яиц, содержащих очень мало желтка. 136 Болотов А. В. Биология размножения и развития При отсутствии большого количества желтка наблюдается четыре типа дробления (рис. 27, 1–5): Рис. 27. Классификация типов дробления (по Голиченкову В. А., 2004): 1 – полное равномерное дробление (иглокожие, бесчерепные); 2 – полное неравномерное дробление (амфибии, осетровые рыбы); 3 – полное билатеральное дробление (асцидии); 4 –полное равномерное ротационное дробление (плацентарные млекопитающие); 5 – полное спиральное дробление (большинство моллюсков; кольчатые, плоские и круглые черви); 6, 7 – неполное дискоидальное дробление (6 – костистые рыбы; 7 – рептилии, птицы); 8 – неполное поверхностное дробление (насекомые) 1. Радиальное голобластическое дробление, которое может быть 2 видов: а) полное равномерное дробление (бесчерепные (ланцетник) и иглокожие). Олиголецитальные яйцеклетки (рис. 27, 1); б) полное неравномерное дробление (амфибии и осетровые). Мезолецитальные яйцеклетки) (рис. 27, 2). 2. Спиральное голобластическое дробление (большинство моллюсков, кольчатые, круглые и плоские черви) (рис. 27, 5). 3. Билатеральное голобластическое дробление (асцидии) (рис. 27, 3). VII. Дробление 137 4. Асинхронное, ротационное голобластическое дробление (плацентарные млекопитающие) (рис. 27, 4). При наличии большого количества желтка наблюдается два типа дробления (рис. 27, 6–8): 1) дискоидальное меробластическое дробление (рептилии, птицы, костистые рыбы) (рис. 27, 6–7); 2) поверхностное меробластическое дробление (насекомые) (рис. 27, 8). 1. Радиальное голобластическое дробление Полное равномерное дробление Указанный тип дробления встречается у иглокожих и бесчерепных (ланцетник). Их яйцеклетка – алецитальная или первично олиго- и изолецитальная, т. е. не содержит вовсе или содержит очень мало желтка (распределенного равномерно). Поэтому дробление полное (дробятся все части зиготы), равномерное (образуются клетки одинаковой величины) и синхронное (деления разных клеток зародыша происходят одновременно) (рис. 28, А–Е). Борозды дробления ориентированы параллельно или перпендикулярно анимально-вегетативной оси. После соединения пронуклеусов ось первого митотического веретена ориентируется перпендикулярно анимально-вегетативной оси яйца. Первая борозда дробления проходит через анимальный и вегетативный полюсы яйца, т. е. через анимально-вегетативную ось и ориентированы вдоль неё. Такое деление называют еще меридиональным, поскольку борозда деления проходит через два полюса подобно меридиану на глобусе (рис. 28, А). Это приводит к образованию двух одинаковых дочерних клеток. Плоскость второго деления тоже проходит через оба полюса, но перпендикулярно плоскости первого деления (рис. 28, Б). Таким образом, два первых деления дробления меридиональны, но взаимно перпендикулярны. Третье деление экваториальное, т. е. митотические веретёна в каждом бластомере располагаются параллельно анимально-вегетативной оси (рис. 28, В). Все образующиеся бластомеры одинаковы по размерам. Четвертое деление снова меридиональное и приводит к образованию двух ярусов по 8 клеток в каждом. Пятое – экваториальное. 138 Болотов А. В. Биология размножения и развития Рис. 28. Радиальное голобластическое равномерное дробление (А, Б, В, Г, Д, Е) (ланцетник) и образование бластулы (Ж) Дальше наблюдается чередование меридиональных и экваториальных делений. В результате образуется зародыш, у которого бластомеры располагаются горизонтальными рядами. На обоих полюсах зародыша бластомеры сближаются и возникает шар с полостью внутри. Такой шар называется бластулой, полость бластоцель. Если разделить зародыш по меридиану или по экватору, то образуются две зеркальные половинки. Такой тип симметрии присущ шару и носит название радиальной симметрией. Образовавшаяся бластула называется целобластула: ее стенка – бластодерма – составляет один слой одинаковых по размеру клеток, а полость – бластоцель – расположена в центре (рис. 28, Ж). По существу, это однослойный сферический пузырек, стенка которого везде одинакова по толщине. Полное неравномерное дробление Для амфибий и осетровых рыб также характерно радиальное голобластическое дробление, но с существенными модификациями (рис. 29). Яйцеклетка – умеренно телолецитальная: желтка много, и он в большей мере сосредоточен у вегетативного полюса яйцеклетки. Это придает дроблению некоторые особенности: дробление полное (дробятся все части зиготы), но при VII. Дробление 139 этом неравномерное (на вегетативном полюсе зародыша клетки крупнее), и асинхронное (клетки вегетативного полюса делятся медленней). Рис. 29. Полное неравномерное дробление (А, Б, В, Г, Д, Е, Ж) и образование амфибластулы (З) Первое и второе деление проходят так же, как и у иглокожих и бесчерепных (рис. 29, А, Б). Третье деление тоже экваториальное и отделяет один полюс от другого, плоскость этого деления смещена в анимальное полушарие, поэтому дробление получило характеристику неравномерное, ведущее к образованию микромеров в анимальном и макромеров в вегетативном полушариях (рис. 29, В, Г). На анимальном полюсе четыре более мелких бластомера – микромеры, на вегетативном – четыре более крупных – макромеры (рис. 29, Д). Затем деление опять идет в меридианных плоскостях, а потом опять в широтных и т. д. У амфибий первые четыре деления проходят синхронно, последующие 7 делений волнами, начинающимися на анимальном полюсе и заканчивющиеся на вегетативном (рис. 29, Ж). В результате подобного дробления образуется амфибластула (рис. 29, З). Ее особенности следующие: 1) анимальный полюс, называемый крышей, содержит мелкие клетки; 2) вегетативный полюс, или дно, состоит из крупных клеток; 140 Болотов А. В. Биология размножения и развития 3) область же между крышей и дном – краевая зона, клетки здесь – промежуточного размера; 4) бластодерма во всех перечисленных областях является многослойной; 5) бластоцель смещена к крыше. 2. Спиральное голобластическое дробление Спиральное дробление характерно для нескольких групп червей (кольчатых, круглых и плоских) и почти всех моллюсков, за исключением головоногих. Оно во многих отношениях отличается от радиального дробления. Во-первых, яйца не делятся параллельно или перпендикулярно анимально-вегетативной оси зиготы. Плоскости делений дробления ориентированы наклонно (под некоторым углом), что и приводит к «спиральному расположению дочерних бластомеров (рис. 30). Во-вторых, число контактов между клетками больше, чем при радиальном дроблении. Бластомеры принимают расположение, наиболее стабильное в термодинамическом отношении, и минимальным объёмом, подобно расположению контактирующих мыльных пузырей. Рис. 30. Спиральное дробление (А, Б) и образование стерробластулы (В) В-третьих, зародыш с этим типом дробления проходит меньшее число делений до начала гаструляции. Это позволило проследить судьбу каждой отдельной клетки бластулы. Когда сопоставили судьбу отдельных клеток зародышей кольчатых VII. Дробление 141 червей, плоских червей и моллюсков, то оказалось, что у клеток, занимающих одно и то же положение, судьба одинакова. В-четвертых, первые три деления дробления происходят синхронно. Первые два дробления почти меридиональны и приводят к образованию 4 крупных клеток – макромеров (A, B, C, D) (рис. 30, А). У многих видов эти бластомеры различаются по размерам, при этом самыми крупными обычно бывают D. При последующих делениях каждый макромер на своем анимальном полюсе отделяет маленький микромер (рис. 30, Б). Каждый последовательно появляющийся квартет микромеров смещается вправо или влево относительно сестринских макромеров, это и приводит к характерному расположению бластомеров по спирали. Возникающие впоследствии бластулы не имеют бластоцеля и носят название стерробластула (рис. 30, В). 3. Билатеральное голобластическое дробление Характерен для нематод, а также для многих низших хордовых, в том числе для асцидий, аппендикулярий. Характерной особенностью этого типа является раннее проявление билатеральной симметрии. Замечательная особенность этого типа дробления заключается в том, что плоскость первого деления устанавливает единственную плоскость симметрии зародыша (рис. 31, А). Каждое последующее деление ориентируется по отношении к этой плоскости симметрии так, что половина зародыша по одну сторону от первой борозды представляет зеркальное отражение половины зародыша по другую сторону. Второе деление, так же как и первое, – меридиональное, но в отличие от первого борозды дробления не проходят через центр яйца. В результате возникают два крупных передних бластомера (A, D) и два задних меньшего размера (рис. 31, Б). На каждой стороне теперь имеются один крупный и один небольшой бластомер. При последующих делениях различия в размерах и форме бластомеров только подчеркивают билатеральную симметрию зародыша (рис. 31, В). 142 Болотов А. В. Биология размножения и развития Рис. 31. Билатеральное голобластическое дробление (А, Б, В) и образование плакулы (бластула) (Г) На 32 клеточной стадии формируется бластула с небольшим бластоцелем и начинается гаструляция. Бластула у асцидий называется плакула (новолат. placula, от греч. plax – плоскость, пластинка), которая имеет вид двуслойной пластинки, образованной более или менее однородными клетками, между слоями которой есть небольшая полость (рис. 31, Г). 4. Асинхронное, ротационное голобластическое дробление Эмбрионы млекопитающих развиваются, как правило, внутриутробно. По этой причине яйцеклетка у них олиголецитальная (мало желтка) и изолецитальная (желток равномерно распределен). После оплодотворения зигота постепенно спускается по маточной трубе в полость матки. В этот период, в течение примерно трех дней, зигота проходит стадию клеточного деления, известную как дробление. При дроблении число клеток увеличивается, но общий их объем не меняется, так как каждая дочерняя клетка мельче, чем исходная. Дробление полное, но неравномерное и асинхронное. Дробление у млекопитающих отличается от всех других типов дробления по многим признакам: 1. Первая особенность заключается в относительно медленном темпе делений. 2. Своеобразное расположение бластомеров относительно друг друга. Первое деление – меридиональное. В ходе второго VII. Дробление 143 деления один бластомер делится меридионально, но перпендикулярно первому делению, а второй – экваториально. Поэтому такой тип дробления был назван чередующимся или ротационным (рис. 32, Б). Рис. 32. Сравнение радиального (А) и асинхронного, ротационного дробления млекопитающих (Б) и образование бластоцисты у млекопитающих (В). А – первое и второе деление радиального дробления; Б – первое и второе деление асинхронного, чередующегося дробления. В – образование бластоцисты: 1 – трофобласт; 2 – зародышевый узелок (эмбриобласт); 3 – бластоцель 3. Выраженная асинхронность раннего дробления. Бластомеры у млекопитающих не делятся все одновременно, поэтому у зародышей не происходит равномерного нарастания числа бластомеров. Число которых может быть нечетным. 4. Явление компактизации. Бластомеры на 8-й клеточной стадии расположены рыхло. Однако после третьего деления поведение бластомеров резко изменяется. Они внезапно сближаются и площадь контакта между клетками максимально увеличивается, и они образуют плотный клеточный шар – морулу. Эта компактная упаковка стабилизируется плотными контактами, которые образуются между клетками, расположенными на поверхности шара, и изолируют лежащие внутри клетки. Между клетками внутри шара образуются щелевые контакты, через которые могут молекулы и ионы диффундировать из клетки в клетку. 144 Болотов А. В. Биология размножения и развития 5. Образование бластулы, которая называется бластоциста (рис. 32, В). Первоначально морула не имеет внутренней полости, но в процессе кавитации наружные клетки, которые называются трофобластом секретируют в морулу жидкость, что и приводит к образованию полости бластулы. А внутренние клетки, которые называются внутренней клеточной массой располагается на одной стороне стенки полого шара. В течение первых суток оно происходит медленно. Первое деление завершается через 30 ч. За стадией двух бластомеров следует стадия трех бластомеров. Второе дробление наступает через 10 ч после первого и приводит к образованию четырехклеточной стадии. Через 40 ч образуются 4 клетки. Дробящийся зародыш постепенно перемещается по яйцеводу и в нижней его части достигает восьмиклеточной стадии (рис. 33). Примерно через 50–60 ч после оплодотворения достигается стадия морулы – шара из 16 и более клеток. Затем один из них делится, а второй по времени запаздывает с делением. Клетки бластомеры, которые делятся чаще – бледные и мелкие, а те, которые делятся реже – крупные и темные. Рис. 33. Схема развития зародышей млекопитающих от оплодотворения до имплантации (по Гилберту С., 1993) VII. Дробление 145 Клетки эмбриона, бластомеры, укладываются компактнее (происходит компактизация), дифференцируются на внутренние и наружные, различающиеся строением и будущей судьбой. Следующие деления приводят к образованию морулы в виде плотного комка клеток, на 3–4-е сутки начинается формирование бластоцисты, напоминающую полый пузырек, заполненный жидкостью. Ее трофобласт – тонкая стенка пузырька – образован вытянутыми крупными клетками, потомками наружных клеток морулы, а внутри находится небольшое скопление мелких клеток, асимметрично прикрепленных к стенке. Это – внутренняя клеточная масса (ВКМ), или эмбриобласт. На стадии бластоцисты зародыш попадает в матку, выходит из оболочки оплодотворения (zona pellucida) и прикрепляется к стенке матки с помощью трофобласта, который затем станет эмбриональной частью плаценты. 5. Дискоидальное меробластическое дробление Дробление резко телолецитальных яйцеклеток (рептилии, птицы, костистые рыбы) называется дискоидальным. Так происходит дробление у костистых рыб (рис. 34). Рис. 34. Дробление яйца у костистых рыб (А, Б, В, Г) и образование бластулы (Д), бл – бластомеры; ж – желток 146 Болотов А. В. Биология размножения и развития Вскоре после оплодотворения яйцеклетки основная масса цитоплазмы, которая ранее тонким равномерным слоем накрывала желток, перетекает к анимальному полюсу, где с самого начала было расположено ядро зиготы. Поскольку деления дробления не распространяются на богатую желтком цитоплазму, на ранних стадиях дробления клетки у своего основания не отделены друг от друга, т. е. не образуются отдельные клетки (рис. 34, А). В результате на анимальном полюсе тангенциальными делениями, которые проходят параллельно поверхности зиготы, образуется однослойная бластодерма (рис. 34, В, Г). Затем дробления идут перпендикулярно, так образуется несколько слоев клеток зародыша. Таким образом формируется сначала однослойная, а затем и многослойная группа бластомеров – дискобластула. Между бластодермой и желтком имеется пространство, называемое подзародышевой полостью или бластоцелью (рис. 34, Д). Дискоидальное дробление птиц (рис. 35) идет по сходному пути, с той главной разницей, что бластодиск, лежащий на желтке, значительно тоньше, нежели у костистых рыб, и поэтому должно пройти больше делений дробления, прежде чем дискоб ластула станет многослойной. К тому времени, когда птица Рис. 35. Дробление яйца у птиц (А, Б, В, Г) и образование бластулы (Д) VII. Дробление 147 откладывает яйцо, бластодерма содержит около 60 тысяч клеток и клетки начинают выселяться в подзародышевую полость, где образуется второй слой. Таким образом зародыш птицы имеет 2 слоя клеток: наружный – эпибласт и лежащий под ним гипобласт. Таким образом, в результате этого типа дробления образуется дискобластула (рис. 35, Д). Она включает три компонента: 1) плоский зародышевый диск, образующийся из апикальной части зиготы и состоящий из небольших клеток; 2) нераздробившийся желток; 3) бластоцель – узкая щель между диском и желтком. 6. Поверхностное меробластическое дробление Центролецитальным яйцам насекомых, очень богатых желтком, свойственен этот тип дробления. Из-за большого количества желтка, расположенного центрально, дробление ограничено только поверхностным слоем цитоплазмы. Рис. 36. Поверхностное дробление зародыша дрозофилы. 256 ядер – стадия сенцитиальной бластодермы; 512 ядер – формирование полярных клеток, начало обособления клеток бластодермы; 750 ядер – формирование клеток бластодермы; 320´ – стадия клеточной бластодермы, бластуляция; 340´ – начало формирования зародышевой полоски. Цифры над зародышем соответствуют числу минут; цифры внизу обозначают число ядер 148 Болотов А. В. Биология размножения и развития Одна из примечательных особенностей этого типа дробления заключается в том, что обособление клеток происходит только после многократного разделения ядер. На стадии 256 ядер, ядра с островками цитоплазмы (энергиды) мигрируют на периферию яйца, примерно на 81-й минуте, где продолжаются деления, но с убывающей скоростью. Теперь зародыш носит название синцитиальной бластодермы. После образования полярных клеток плазматическая мембрана яйца образует выпячивания, которые углубляются между ядрами и в конечном счете обособляется каждое из них в отдельной клетке. Так возникает клеточная бластодерма, спустя почти 5 часов, клетками которой однослойным покровом одевают богатую желтком сердцевину яйца. Так образуется бластула, называется перибластула, пузырёк, стенка которого состоит из одного слоя клеток, а полость заполнена нераздробившимся желтком (см. рис. 36). VIII. ГАСТРУЛЯЦИЯ Несмотря на большое разнообразие процессов гаструляции, можно дать вполне определенную классификацию гаструл и путей возникновения их из предыдущей стадии бластулы. К сожалению, еще не создана классификация, которая исходила бы из принципа эволюции, так как историческое происхождение разных типов гаструляции изучено недостаточно, и эмбриология имеет в этой области больше научных загадок, чем достоверных сведений. Некоторые способы гаструляции встречаются во всех типах животного мира. 1. Инвагинационная гаструла (рис. 37, А) образуется путем инвагинации (впячивания) части однослойной бластодермы (стенки бластулы) внутрь бластоцеля. Грубое подобие такого типа гаструляции – результат впячивания стенки резинового мячика при надавливании на него. Впячивание можно довести вплотную до внутренней стенки мячика (на противоположном от места вдавливания участке), или вдавливание можно сделать не столь значительным. Образуется двухслойный мешок, наружной стенкой которого является первичная эктодерма, а внутренней – первичная энтодерма. Впячивание образует архентерон (первичный кишечник), а отверстие, при помощи которого он сообщается с наружной средой, называется первичным ртом, или бластопором (рис. 37, А). Судьба бластопора у разных животных неодинакова. У многих животных первичный рот, соответственно развиваясь и дифференцируясь, превращается в дефинитивный рот взрослого организма. Эти животные называются первичноротыми (Protostomia). К первичноротым относятся черви, моллюски и членистоногие. Не менее обширна группа вторичноротых (Deuterostomia), у которых бластопор превращается или в анальное отверстие, или (у хордовых) в нервно-кишечный канал, находящийся в заднем конце эмбриона. Рот у таких животных возникает на переднем конце, на брюшной стенке, в результате особых формообразовательных процессов (впячивание эктодермы, прободение стенки 150 Болотов А. В. Биология размножения и развития средней кишки). К Deuterostomia относятся: щетинкочелюстные, плеченогие, иглокожие, кишечнодышащие и хордовые. Рис. 37. Разные способы гаструляции. A – инвагинационная гаструла; Б – иммиграционная гаструла; В – деляминационная гаструла; Г– эпиболическая гаструла Указанными признаками отнюдь не исчерпывается необходимость разделения животных на две группы: первично- и вторичноротые. В дальнейшем будут сообщены иные не менее важные признаки обеих групп. Очень многие вопросы являются спорными потому, что биология еще не знает истинных предков современных; нет единого мнения и о филогенетическом значении разного типа гаструл. Многие современные исследователи, присоединяясь к И. И. Мечникову, считают исходной формой гаструлы иммиграционную гаструлу и выводят все способы образования энтодермы из мультиполярной иммиграции и деляминации. VIII. Гаструляция 151 Возвращаясь к характеристике инвагинационной гаструлы, отметим, что в ней имеются две полости: в большей или меньшей степени сохраняется бластоцель (хотя в некоторых случаях эта полость почти нацело вытесняется впячиванием) и возникает новая полость – гастроцель (рис. 37, А), или гастральная полость или полость первичного кишечника (архентерон). 2. Иммиграционная гаструла (рис. 37, Б) характерна для многих кишечнополостных. Она развивается вследствие активного выселения части клеток стенки бластулы внутрь бластоцеля. Это выселение клеток в случаях униполярной иммиграции происходит лишь с одного полюса (вегетативного). Выселившиеся клетки образуют позднее внутренний слой гаструлы (энтодерму). Таким образом, как и при инвагинации, зародыш делается двухслойным. Так происходит гаструляция, например, у гидроидного полипа Glytia, гидромедузы Aequorea или у медузы Octorchis gegenbauri. Униполярная иммиграция свойственна почти всем гидромедузам, особенно тем, для которых характерна плавающая целобластула. Клетки бластодермы могут проникать в бластоцель не в каком-либо одном участке, а по всей поверхности зародыша, причем такая иммиграция может начинаться уже на стадии 16 бластомеров. Эта так называемая мультиполярная иммиграция свойственна, например, зародышу медузы Solmundella, что обнаружил впервые И. И. Мечников. Мультиполярная иммиграция, однако, редкое явление. У многих видов кишечнополостных, которым свойственна иммиграционная гаструла, происходит столь массовое активное выселение клеток бластодермы, что бластоцель полностью исчезает, так как заполняется плотной массой клеток. Как видим, у иммиграционной гаструлы нет бластопора и, значит, нет характерного для инвагинационной гаструлы сообщения гастроцеля с внешней средой. Можно лишь, проводя далеко идущую аналогию (оправданную представлением об общности животного мира), говорить о месте бластодермы, соответствующем бластопору, что, однако, доступно критике. В случае мультиполярной иммиграции еще меньше оснований говорить о бластопоре или даже указывать какой-либо соответствующий ему участок бластодермы. 152 Болотов А. В. Биология размножения и развития 3. Деляминационная гаструла (рис. 37, В), встречающаяся также у кишечнополостных, впервые описана И. И. Мечниковым. Морфогенетические процессы, сопровождающие этот редкий случай гаструляции, очень своеобразны и в типичном случае кажутся простыми. Яйца некоторых сцифомедуз, например Geryonidae, имеют различимые, концентрически расположенные участки цитоплазмы: один участок – плотная, зернистая эктоплазма и другой – ячеистого строения эндоплазма, в который заходит наружный эктоплазматический слой, содержащий полужидкий желток. Структура яйца такова, что оно может быть отнесено к центролецитальным яйцам. У таких яиц происходит относительно равномерное синхронное дробление, так что образуется зародыш, состоящий из 2, 4, 8, 16 и, наконец, 32 бластомеров, кажущихся одинаковыми по размерам и пространственному положению. Дальнейшие дробления бластомеров происходят параллельно поверхности зародыша, и в результате дроблений наружный слой бластомеров будет состоять из одной (бывшей) эктоплазмы, а внутренний – из эндоплазмы и частично из эктоплазмы. Происходит деляминация (расщепление) одного слоя клеток на два. В дальнейшем дробятся только «внутренние» бластомеры, притом опять-таки параллельно поверхности зародыша; возникают, таким образом, две легко различимые клетки: наружный, «эктоплазматический» бластомер и внутренний, «эндоплазматический». В результате такой своеобразной гаструляции зародыш оказывается шаром, состоящим из 64 плоских клеток, образующих наружный слой – эктодерму, и из 32 более высоких клеток внутреннего слоя – энтодермы. У животных, для которых характерна типичная морула (например, у полипа Clava squaniata), процесс гаструляции происходит так, что клетки, находящиеся на периферии, приобретают отличное от других клеток строение. Пласт первых клеток образует эктодерму, а внутренний пласт – энтодерму. Такой тип гаструляции можно рассматривать как один из вариантов деляминации. 4. Эпиболическая гаструла (рис. 37, Г) формируется у некоторых животных с резко выраженным телолецитальным строением яиц, например у Bonellia. Значительного размера медленно делящиеся бластомеры (макромеры), с большим содержанием желтка, не обнаруживают способности к каким-либо 153 VIII. Гаструляция перемещениям. Их обрастают (наползают на них) более быстро делящиеся мелкие, лежащие на поверхности клетки (микромеры). Выходит, что у зародышей таких животных бластопор отсутствует, архентерон не формируется. Только впоследствии, когда макромеры, делясь, становятся меньшего размера, образуется (по неясным биохимическим и физическим причинам) полость и формируется зачаток первичного кишечника. 5. Смешанный тип гаструляции. Типы гаструляции кажутся настолько различными, что их невозможно сравнивать. Несомненно, каждый тип гаструляции требует причинного анализа, но все же в некотором отношении разные способы гаструляции сравнимы. Так, своеобразный процесс эпиболии можно рассматривать как один из особенных вариантов инвагинации. Процессы деляминации можно уподоблять процессам иммиграции, конечно, с большими оговорками. Важно отметить далее, что гаструляция у многих животных происходит так, что имеют место и явления инвагинации, и явления эпиболии, и другие морфогенетические процессы. Такова, например, гаструляция у амфибий, пресмыкающихся, птиц, млекопитающих. Образование мезодермы У всех животных, за исключением кишечнополостных, в связи с гаструляцией (параллельно с ней или на следующем этапе, обусловленном гаструляцией) возникает и третий зародышевый пласт – мезодерма. Это совокупность клеточных элементов, залегающих между эктодермой и энтодермой, т. е. в бластоцеле. Таким образом, зародыш становится не двухслойным, а трехслойным. У высших позвоночных трехслойное строение зародышей возникает уже в процессе гаструляции, тогда как у низших хордовых и у всех других типов в результате собственно гаструляции образуется двухслойный зародыш. Вопросы о путях образования мезодермы у разных животных издавна интересовали и сравнительных анатомов, и эмбриологов. Если отвлечься от всех разнообразных деталей формирования мезодермы у разных животных, можно установить два принципиально разных пути возникновения ее: телобластический, свойственный Protosiomia, и энтероцельный, характерный для Deuterostomia. 154 Болотов А. В. Биология размножения и развития Рис. 38. Образование мезодермы. А – телобластический путь; Б, В, Г – энтероцельные пути образования мезодермы: собственно энтероцельный, деляминационный, полиферационный. 1 – эктодерма; 2 – эпигастральное впячивание с листком энтодермы; 3 – бластоцель; 4 – формирующаяся мезодерма; 5 – гасторцель У первичноротых во время гаструляции на границе между эктодермой и энтодермой, по бокам бластопора, уже имеются две большие клетки (или несколько таких клеток-телобластов), отделяющие от себя (вследствие делений) мелкие клетки. Таким образом, формируется средний пласт – мезодерма. Телобласты, давая новые и новые поколения клеток мезодермы, остаются на заднем конце зародыша. По этой причине такой способ образования мезодермы и называют телобластическим (от греч. telos – конец) (рис. 38, А). Энтероцельные способы (рис. 38, Б, В, Г) образования мезодермы 1. Собственно энтероцельный способ (рис. 38, Б). На боковых стенках гастрального впячивания образуются карманоподобные выступы, которые направлены внутрь бластоцеля. Эти выступы отшнуровываются от кишечника и отделяются от него в виде мешочков. Полость мешочков превращается в целóм, т. е. во вторичную полость тела, целомические мешки могут подразделяться на сегменты. VIII. Гаструляция 155 2. Деляминационный (рис. 38, В). Морфогенетические процессы, сопровождающие этот случай образования мезодермы очень редок. Встречается у сцифомедуз, например Geryonidae. В период гаструляции цитоплазма поляризуется, т. е. имеет различной плотности цитоплазму боковых стенок внутренней и наружной поверхности гастрального впячивания: зернистая эктоплазма и ячеистая эндоплазма. Затем боковые стенки, критически поляризованные, расщепляются и отделяются в бластоцель, где через некоторое время один клеточный пласт снова расщепляется и внутри образуется вторичная полость тела или целóм. 3. Полиферационный (рис. 38, Г). Этот тип образования мезодермы встречается у кишечнополостных и других животных в развитии. В период гаструляции в силу разных физикохимических процессов из боковых стенок гастрального впячивания начинают активно выселяться клетки внутрь бластоцеля. Выселившиеся клетки образуют позднее два слоя, между которыми развивается вторичная полость тела или целóм. IX. ОСОБЕННОСТИ РАЗВИТИЯ МЛЕКОПИТАЮЩИХ Предками млекопитающих являлись, вероятно, яйцекладущие рептилии. Наиболее примитивны из современных млекопитающих также яйцекладущие животные. В подклассе первозверей (Prototheria) яйцо утконоса Ornithorhynchus развивается вне материнского организма; у ехидны Echidna яйцо развивается в кожной складке на брюшной стороне тела, но питание от матери зародыш не получает. Яйца этих животных содержат большое количество желтка, дробление частичное, свойственное телолецитальным яйцам. Эмбриональное развитие во всех существенных чертах сходно с развитием рептилий. В подклассе сумчатых (Metatheria) зародыши также почти не получают питание от матери. У некоторых сумчатых развивается примитивная плацента, функционирующая непродолжительное время. Зародыши у сумчатых, например, рождаются недоразвитыми, очень маленькими (1–2 см) и донашиваются матерью в сумке, где устанавливается связь пищевода зародыша с соском матери: ее молоко впрыскивается в пищевод. Яйца у этих животных содержат небольшое количество желтка. У высших млекопитающих, в том числе и у человека, яйцо практически не содержит желточных включений и у него нет в достаточном количестве ни органических, ни неорганических веществ даже для начальных стадий развития. Все необходимое для своего развития зародыш получает с самого начала от материнского организма. Яйцевые клетки плацентарных животных микроскопического размера (100–300 мкм в диаметре), алецитального или изолецитального типа. Как происходила эволюция яиц, и каков характер развития млекопитающих по мере прогресса установления связи зародышей с материнским организмом, эмбриология не знает, хотя и высказаны важные предположения. Очень интересна и вполне аргументирована смелая гипотеза зоолога В. А. Фаусека, согласно которой эволюцию установления все более тесных связей зародыша с материнским организмом надо рассматривать в свете явлений паразитизма: у за- IX. Особенности развития млекопитающих 157 родыша, как у паразита, развиваются приспособления для использования материнского организма в качестве места обитания и как источника пищи. Для анализа причин характера дробления яиц млекопитающих недостаточно гипотез о дроблении. Казалось бы, раз яйца млекопитающих практически лишены желтка, их дробление должно быть таким же, как у изолецитальных яиц беспозвоночных и низших хордовых. На самом деле у многих высших млекопитающих дробление кажется неправильным, оно несинхронное, зародыш проходит, например, стадию трех, пяти бластомеров и т. д. Если учесть сложность организации млекопитающих, а также большие видовые особенности в дроблении, закономерно встречающиеся у разных видов млекопитающих, то надо признать, что говорить о дроблении яиц млекопитающих как о «неправильном» имеется еще меньше оснований, чем oб «анархическом» дроблении некоторых кишечнополостных. У большинства млекопитающих в результате дробления возникает морула, но у некоторых видов (например, у слоноземлеройки или хоботкового прыгунчика) уже на четырехбластомерной стадии появляется бластоцель. Эта стадия развития соответствует бластуле других позвоночных животных. Внутренняя масса бластомеров морулы отличается от поверхностнолежащих своей дальнейшей судьбой. Внутренняя группа бластомеров – эмбриобласт (рис. 39, А, 2) – это источник развития зародыша, его оболочек и желточного мешка, а поверхностно лежащие бластомеры являются трофобластом («питающим зачатком») (рис. 39, А, 1). Эти клетки участвуют в установлении связи зародыша с тканями матки. Морула млекопитающих превращается в бластоцистубластодермический пузырек, размеры которого у разных животных неодинаковы (у кролика около 7 мм в диаметре). Полость его наполнена жидкостью, вероятно, продуктом жизнедеятельности клеток зародыша. К одному из участков однослойной стенки бластоцисты прилегает скопление клеток эмбриобласта – зародышевый узелок (рис. 39, 3). Участок трофобласта, с которым соприкасается эмбриобласт, называют рауберовым слоем (рис. 39, 4). В ходе дальнейшего развития зародышевый узелок превращается в зародыше- 158 Болотов А. В. Биология размножения и развития вый щиток (рис. 39, 7). Начинаются процессы гаструляции. В зародышевом щитке обособляется энтодерма, в результате чего он становится двухслойным, состоящим из эпи- и гипобласта. В зародышевом щитке, как и у птиц, будут происходить процессы формирования первичной полоски и гензеновского узелка (рис. 42), т. е. тех же структур, с которыми связано образование системы осевых органов зародыша. На стадии гаструляции зародыш имплантируется в стенку матки. Это – сложный в морфологическом и физиологическом отношениях процесс. Зародыш внедряется в слизистую оболочку матки целиком или только отростками трофобласта. Выросты трофобласта называются первичными ворсинками. Впоследствии развиваются вторичные ворсинки с соединительной тканью и сосудами. Так формируется хорион – ворсинчатая оболочка, гомологом которой является серозная оболочка зародышей рептилий и птиц. Рис. 39. Разрез ранней (А) и поздней (Б) бластоциты и дифференцирующего эмбриобласта (В) (по К. Уоддингтону, 1957). 1 – трофобласт, 2 – эмбриобласт, 3 – зародышевый узелок, 4 – рауберов слой трофобласта, 5 – эктобласт (эпибласт), 6 – энтодерма (гипобласт), 7 – зародышевый щиток X. ПРОВИЗОРНЫЕ (ВНЕЗАРОДЫШЕВЫЕ) ОРГАНЫ Провизорные, или временные, органы, развивающиеся в процессе эмбриогенеза вне тела зародыша, выполняют многообразные функции, обеспечивающие рост и развитие самого зародыша. В связи с тем, что некоторые из этих органов окружают зародыш, распространено и другое название – зародышевые оболочки. К ним относят: желточный мешок, амнион. серозную оболочки, аллантоис, хорион, плаценту. В эволюции они появились не одновременно. Желточный мешок В ряду хордовых животных внезародышевые органы впервые появляются у рыб в виде желточного мешка, депонирующего желток, используемый зародышем в процессе развития. Его формирование начинается на стадии ранней гаструлы, когда во внутреннем листке можно выделить зародышевую (кишечную) энтодерму и расположенную по периферии диска внезародышевую желточную энтодерму. Своим свободным краем желточная энтодерма образует край обрастания, который начинает надвигаться на желток. После возникновения хордомезодермального зачатка между экто- и энтодермой прорастают париетальный и висцеральный листки мезодермы. Желток обрастает всеми четырьмя листками. Зародыш приподнимается над диском и отделяется от желтка туловищной складкой. При образовании туловищной складки зародышевая энтодерма, до того распластанная на желтке, сворачивается в кишечную трубку. Зародыш связан с желточным мешком полым канатиком – желточным стебельком. Желточный мешок рыб выполняет трофическую функцию. Другая функция мешка – кроветворная – заключается в образовании клеток крови в мезодерме стенки мешка. С выходом животных на сушу (у пресмыкающихся и птиц) в связи с развитием зародыша под скорлупой появляются новые зародышевые органы: амнион, серозная оболочка и аллантоис. 160 Болотов А. В. Биология размножения и развития Как и у рыб, у пресмыкающихся и птиц возникают туловищные складки, отделяющие зародыш от желточного мешка. Желточный мешок у них также выполняет трофическую и кроветворную функции. Амнион Позднее в эмбриогенезе пресмыкающихся и птиц за счет эктодермы и париетального листка мезодермы формируются амниотические складки, растущие в направлении дорсальной поверхности зародыша. По мере роста головного конца зародыша амниотические складки как бы надвигаются спереди на зародыш, причем он одновременно вдавливается в желток (рис. 40, А, Б). Нарастающие на зародыш складки смыкаются и оба листка – эктодерма и прилегающий к нему париетальный листок мезодермы срастаются с одноименными листками противоположной стороны. Из двух листков складок при этом образуются две оболочки – амниотическая, или водная, обращенная к зародышу, и серозная, наружная (рис. 40, В). Рис. 40. Схема развития внезародышевых органов у млекопитающих. А – сближение амниотических складок и начало образования аллантоиса; Б, В – сформированные внезародышевые органы; 1 – тело зародыша; 2 – туловищная складка; 3 – амниотические складки; 4 – полость амниона; 5 – желточный мешок; 6 – аллантоис; 7 – амниотическая оболочка; 8 – кишка; 9 – хорион Амниотическая оболочка на ранних стадиях отделена от тела зародыша узкой щелью, которая позднее превращается в за- X. Провизорные (внезародышевые) органы 161 полненную жидкостью амниотическую полость. Эта жидкость, вырабатываемая клетками эктодермы амниотической оболочки, обращенной в полость амниона, содержит белки, углеводы. Жидкая среда амниона обеспечивает условия для свободного развития зародыша, а также амортизации возможных сотрясений и ударов. Не следует путать с серозной оболочкой, покрывающей снаружи большинство внутренних органов. Серозная оболочка Серозная оболочка образуется одновременно с амниотической оболочкой у пресмыкающихся и птиц. Серозная оболочка участвует в снабжении эмбриона кислородом, что позволяет рассматривать ее как провизорный орган дыхания у яйцекладущих позвоночных. Аллантоис В отличие от амниона развитие аллантоиса начинается в каудальном отделе самого зародыша в виде выроста вентральной стенки задней кишки, образованного энтодермой и висцеральным листком мезодермы. Проксимальная часть аллантоиса располагается вдоль желточного стебелька, а дистальная, разрастаясь, врастает в щель, между амнионом и серозной оболочкой. Этот орган газообмена и выделения: по сосудам, образующимся в мезодерме аллантоиса. доставляется кислород; в аллантоис выделяются продукты обмена веществ зародыша. Сращенную серозную оболочку и аллантоис у птиц и пресмыкающихся часто называют хорионом, или хориоаллантоисом. У млекопитающих внезародышевые органы представлены хорионом, амнионом, желточным мешком, аллантоисом (рис. 40, 6) и плацентой. Хорион Хорион (рис. 40, 9), или ворсинчатая оболочка, развивается из трофобласта и внезародышевой мезодермы. Первоначально трофобласт представлен оболочкой с первичными ворсинками, за счет которых после имплантации зародыша устанавливается связь с материнским организмом. Со времени появления в эмбриобласте внезародышевой мезодермы (у человека – на 2–3-й неделе развития) она подрастает к трофобласту и образует вместе с ним вторичные эпителиомезенхимальные ворсинки. С это- 162 Болотов А. В. Биология размножения и развития го времени трофобласт превращается в хорион, или ворсинчатую оболочку. Внедряясь в слизистую оболочку матки, хорион образует вместе с ней плаценту. Плацента Эта связь обеспечивается формированием специального органа – плаценты. По строению плаценты делят на несколько типов (рис. 41). Плацента эпителиохориального типа, или полуплацента (рис. 41, А), характерна для некоторых видов сумчатых, свиньи, лошади и других млекопитающих. Ворсинки хориона, контактируя со слизистым эпителием матки и входя в его углубления, не разрушают материнские ткани. Маточные железы секретируют маточное молоко (эмбриотроф), которое всасывается ворсинками хориона. Таким образом, материнский организм обеспечивает зародыш питательными веществами и кислородом. При родах ворсинки хориона отъединяются от эпителия матки без разрушения ее. Десмохориальная плацента, или соединительно-тканная хориальная (рис. 41, Б) (жвачные животные), развивается так, что ворсинки внедряются в стенку матки, эпителий ее слизистой оболочки разрушается; ткани зародыша оказываются в контакте с соединительной тканью матки; однако более интимная связь между материнским организмом и зародышем не устанавливается и при этом типе плацентации. Область контакта зародыша с тканями матери осуществляется в районах котиледон, небольших площадок, разбросанных по поверхности обширного плодного пузыря, превосходящего в сотни раз объем самого зародыша. После родов в стенке матки остаются участки, лишенные эпителия. Последующая регенерация происходит очень быстро. Вазохориальная плацента, или эндотелиохориальная (рис. 41, В) (хищные), обеспечивает еще более тесную связь зародыша с матерью. При плацентации происходит не только разрушение слизистого эпителия матки, но и деструктивные процессы в соединительной ткани: трофобласт ворсинок хориона оказывается в непосредственном контакте с эндотелием сосудов. Этот способ плацентации обеспечивает более совершенное снабжение зародыша питательными веществами и кислородом. X. Провизорные (внезародышевые) органы 163 Гемохориальная плацента (рис. 41, Г) развивается у некоторых насекомоядных, летучих мышей, грызунов, обезьян и у человека. Благодаря плацентам этого типа устанавливается еще более интимная связь зародыша с материнским организмом. Совокупность очень сложных процессов (лизис эпителия матки, разрушение соединительной ткани и сосудов) приводят к тому, что трофобласт непосредственно соприкасается с кровью матери, ворсинки хориона «купаются» в материнской крови. Рис. 41. Гистологические виды плацент. А – эпителиохориальная; Б – десмохориальная; В – вазохориальная; Г – гемохориальная Строение гемохориального типа плацент варьирует у разных животных. У обезьян и человека ворсинки хориона сложно ветвятся, и тем самым создается огромная поверхность соприкосновения ворсинок с кровью. Поэтому такая плацента называется ворсинковой гемохориальной плацентой. Увеличение поверхности контакта трофобласта с кровью в других случаях (у грызунов, например) достигается путем сильного разветвления выпячиваний трофобласта, образующих сложный лабиринт каналов, по которым течет материнская кровь; поэтому такого типа плацента называется лабиринтной гемохориальной плацентой. Естественно, что при такой связи зародыша с матерью роды сопровождаются сильным кровотечением, отпадает эпителий слизистой оболочки матки. Стенка матки обладает хорошей регенерационной способностью и происходит быстрое восстановление её. XI. ОРГАНОГЕНЕЗ Органогенез – предмет исследования эмбриологии, гистологии, анатомии, физиологии и других наук, так как он является совокупностью процессов изменения формы, тканевого строения, возникновения и изменения биохимических и физиологических явлений и т. д. После завершения гаструляционных движений, начинается следующий этап в развитии зародыша (к концу 3-й недели зародыш вступает в пятую стадию развития) – органогенез. Происходит дифференцировка зародышевых листков, закладка и построение отдельных систем своих органов – нервной, пищеварительной и кровеносной. Хордовые животные обладают удлиненным телом с двусторонней симметрией. На брюшной стороне у переднего конца тела находится рот, а у заднего – анальное отверстие. 1. Развитие производных эктодермы У позвоночных органогенез начинается одновременно с формированием мезодермы, закладываются зачатки осевых органов на спинной стороне зародыша – хорда (осевой скелет) и нервная трубка. 1. Стадия нейрулы. У зародыша человека нейруляция происходит в течение 3-й недели после зачатия. На этой стадии длина зародыша составляет всего около 2 мм; он почти неотличим от зародыша лягушки или морского ежа, но уже окружен оболочками и прикреплен плацентой к матке. Плацента пока еще очень мала. Оболочки, соединяющие зародыш с плацентой, превращаются в пуповину, которая по мере развития становится толще и длиннее. Хорда возникает из энтодермы. Зачаток хорды представлен хордальным отростком, который вырастает из первичного узелка в виде тяжа клеток, проникающего под эктодерму. Зачатком нервной трубки является нервная пластинка, образующаяся из клеток эктодермы, прилежащих к хордальному отростку. Первичный узелок – на дорсальной поверхности зародышевого щитка образуется уплотненный тяж клеток – первичная по- XI. Органогенез 165 лоска (рис. 42), заканчивающаяся первичным узелком. Зародышевый узелок – это прилежащие друг к другу части эктодермального и энтодермального пузырьков. В последующие стадии зародышевого периода происходит: а) обособление тела зародыша от внезародышевых органов, б) дифференцировка мезодермы и других зародышевых листков, в) закладка зачатков разных органов, г) окончательно Рис. 42. Развитие млеко- формируются осевые органы. питающего на 14-й день Хорда наибольшего развития достигает на 4-й неделе зародышевого периода и занимает в теле зародыша центральное (осевое) положение. Позднее вокруг хорды закладываются позвонки, а сама хорда редуцируется. Остатки хорды преобразуются в студенистые ядра межпозвоночных дисков. По бокам хорды и нервной трубки лежит продольно сегментированная туловищная мускулатура. В первую очередь начинается нарастание эктодермы по краям нервной пластинки. Нервная трубка формируется из нервной пластинки (рис. 43, А, 1). Эта пластинка превращается в нервный желобок (рис. 43, Б, 2), а затем в нервную трубку (рис. 43, Г, 5), которая в передней и задней части зародыша некоторое время остается открытой. В процессе замыкания нервной трубки, т. е. в задней части тела эктодерма нарастает на бластопор, и отверстие нервной трубки закрывает ее так, что нервная трубка остается в сообщении с кишечной полостью. Канал, соединяющий нервную трубку с кишечной полостью, называется нервно-кишечным каналом, он отделяется от остальной эктодермы и погружается под нее. При этом часть клеток боковых краев нервного желобка отшнуровывается и образует ганглиозные пластинки или нервные гребешки (рис. 43, 4). Нервная трубка является зачатком головного и спинного мозга, а из ганглиозных пластинок развиваются узлы (ганглии) спинномозговых и черепных нервов. В области шеи и туловища образуется глубокая туловищная складка, которая и разделяет нервную трубку на переднюю и заднюю часть (спинной мозг). 166 Болотов А. В. Биология размножения и развития Рис. 43. Образование нервной трубки и нервного гребешка. 1 – нервная пластинка; 2 – нервный желобок; 3 – нервная складка; 4 – нервный гребешок; 5 – нервная трубка Из передней части нервной трубки формируется головной мозг и органы чувств. Канал нервной трубки превращается в центральный спинномозговой канал развившегося мозга. В разных участках переднего отдела нервной трубки благодаря неравномерному ее росту очень рано заметными в определенных участках становятся утолщения, образуются карманоподобные выпячивания, особенно значительные в участках будущих мозговых полушарий. Снаружи образуются в определенных местах неглубокие сжатия нервной трубки и обозначаются три мозговых пузыря: передний, средний, задний. У человека это происходит на 4-й неделе развития (рис. 44). А Б Рис. 44. Стадия трех мозговых пузырей: А – вид сбоку, Б – вид сверху XI. Органогенез 167 Рис. 45. Стадия пяти мозговых пузырей. А – вид сбоку, Б – вид сверху Передний мозговой пузырь в дальнейшем подразделяется на: зачаток большого мозга (концевого) и зачаток промежуточного мозга (рис. 45). Средний мозговой пузырь, развиваясь и дифференцируясь, превращается в средний мозг (рис. 45). Задний мозговой пузырь при дальнейшем развитии подразделяется на: зачаток мозжечка и моста и зачаток продолговатого мозга (рис. 45). Продолговатый мозг переходит в спинной (рис. 45). Верхняя стенка зачатка продолговатого мозга при дальнейшем развитии остается тонкой, и в ней не образуются нервные клетки. Большая часть верхней стенки врастает вместе с сосудистой оболочкой в полость 4-го желудочка мозга, образуя сосудистое сплетение 4-го желудочка. Продолговатый мозг образуется в результате разрастания и развития боковых и нижних стенок зачатка продолговатого мозга. На боковых стенках переднего мозгового пузыря (из боковых стенок промежуточного мозга) выпячиваются зачатки глаз – глазные пузыри (рис. 44). Утолщения боковых стенок промежуточного мозга образуют зрительные бугры. Дно промежуточного мозга формирует глубокое выпячивание – воронку мозга. Из ее нижнего конца возникает нейральная часть важнейшей железы внутренней секреции – гипофиза. 168 Болотов А. В. Биология размножения и развития Из стенки промежуточного мозга, расположенной сзади от воронки, образуется подбугровая область мозга – гипоталамус, а в области тонкой дорсальной стенки промежуточного мозга – эпифиз, или шишковидная железа. Уже на ранних стадиях развития разные отделы мозга отличаются друг от друга неравномерным утолщением стенок. В области переднего мозга разрастаются переднебоковые стенки, что приводит к образованию пары выступов- зачатков полушарий головного мозга, которые особенно сильно разрастаются у высших позвоночных, где они накрывают собой все находящиеся сзади отделы мозга, вплоть до мозжечка. Неравномерное разрастание их поверхности приводит к появлению глубоких борозд. Дорсальная часть нервной трубки вытягивается сильнее, чем вентральная, вследствие чего задний конец трубки выносится за бластопор. Затем происходит изгибание трубки на некотором расстоянии от бластопора. Вершина образующегося изгиба становится зачатком хвоста. Участок, лежащий между вершиной зачатка хвоста и бластопором, дифференцируется как мышцы хвостовой области. 2. Кожа. Большая часть эктодермы тела зародыша составляет кожную эктодерму, из которой развивается эпидермис кожи и его производные (перо, волосы, ногти, кожные и молочные железы). Кожа позвоночных развивается из двух зародышевых листков – эктодермы и мезодермы. Эмбриональная эктодерма сначала превращается в 2-слойный, а затем в многослойный эпителий – кожный эпидермис. Его внутренний, прилежащий к мезодерме слой (ростковый) в течение всей жизни организма сохраняет функции камбия: в нем происходят клеточные деления, и вновь образующиеся клетки перемещаются во внешние слои эпидермиса, где дифференцируются. У всех наземных позвоночных дифференцированные клетки синтезируют роговое вещество – белок кератин. В дальнейшем дифференцированные клетки заполняются кератином, уплощаются, теряют ядро и формируют поверхностный омертвевающий слой кожи. Мезодермальный слой кожи (дерма) образуется соединительно-тканными клетками. За счет деятельности клеток дермы формируются коллагеновые, эластические и ретикулярные волокна. XI. Органогенез 169 Рис. 46. Развитие волоса (А) и пера (Б) Зачатки пера и волоса представляют собой эпидермальные плакоды – линзовидные утолщения эпидермиса (рис. 46, А, Б). При развитии пера под плакодами возникают сгущения дермальных клеток, которые как бы приподнимают эпидермис, придавая ему вид бугорка, обращенного вершиной назад (рис. 46, Б). Деление клеток, окружающих зачаток пера приводит к образованию перьевой сумки из которой и вырастает перо (рис. 46, Б). Непрерывно образующиеся новые кератиноциты в воротничке, расположенном в основании сумки, вытесняют старые клетки вверх, в результате чего и образуется трубчатое перо. Из дермальных клеток формируется мякоть пера, а из эпидермиса – его поверхностный ороговевающий слой (рис. 46, Б). При развитии волоса эпидермальная плакода глубоко погружается в дерму, образуя волосяной узелок (рис. 46, А). Наружные слои узелка дают начало влагалищу волоса и сальным железам, а внутренние – собственно волосу. На дне волосяного узелка имеется камбиальная зона, поддерживающая рост волоса. Дно узелка вогнутое, и в это углубление вдается дермальный со- 170 Болотов А. В. Биология размножения и развития сочек, куда прорастают нервы и питающие волос кровеносные сосуды (рис. 46, А). 3. Компоненты органов слуха, обоняния. Органы обоняния позвоночных развиваются из обонятельных плакод – парных утолщений эктодермы в передней части головы. Эпителий обонятельных плакод (обонятельный эпителий) содержит нервно-чувствительные клетки, которые посредством аксонов (длинный отросток нервной клетки, который проводит нервные импульсы от тела нервной клетки) связаны с обонятельным отделом головного мозга. В результате разрастания обонятельного эпителия плакоды превращаются в обонятельные мешки, открывающиеся наружу обонятельными ямками. Орган слуха – в его образовании участвуют покровная эктодерма и головная мезенхима (рис. 47). Из покровной эктодермы формируется основная часть органа слуха – внутреннее ухо, развитие которого начинается с образования парных утолщений покровной эктодермы на уровне заднего мозга – слуховых плакод. Рис. 47. Закладка органа слуха Слуховые плакоды впоследствии выпячиваются и почти полностью отшнуровываются от эктодермы, образуя слуховые пузырьки. Каждый слуховой пузырек некоторое время связан с внешней средой узким эндолимфатическим каналом; при дальнейшем развитии у большинства позвоночных этот канал замыкается. Слуховой пузырек подразделяется на верхний и нижний отделы (см. рис. 47, 48). XI. Органогенез 171 Рис. 48. Развитие слухового пузырька. А – начальная стадия развития; Б – поздняя стадия развития В верхнем отделе образуются три полукруглых уплощения (рис. 48, А). Каждое из них позже прорывается посередине и превращается в полукружный канал (рис. 48, Б). Полукружные каналы представляют собой органы равновесия позвоночных. В нижней части слухового пузырька появляется мешковидное вздутие (рис. 48, А), а на его конце – слепой вырост, который у высших позвоночных удлиняется и закручивается в канал слуховой улитки (рис. 48, Б). В стенке этого канала развивается орган слуха – кортиев орган. Под воздействием самого слухового пузырька из окружающей мезенхимы формируется хрящевая слуховая капсула, которая повторяет форму внутреннего уха. Слуховая капсула формирует скелетный лабиринт, подобный внутреннему лабиринту внутреннего уха. Д. П. Филатов проводил следующий опыт: удалял слуховой пузырек у зародышей жабы. Хрящевая капсула слухового пузырька на оперированной стороне не развивается. При пересадке слухового пузырька в разные области головы и в другие области тела вокруг них формируется хрящевая капсула из любой мезенхимной ткани. Важнейшая часть органа слуха – 172 Болотов А. В. Биология размножения и развития барабанная полость, или полость среднего уха, возникает из полости жаберного кармана (рис. 51, Б). Барабанная полость разрастается и развивается далее; формирующиеся вначале вне ее слуховые косточки оказываются внутри барабанной полости, отделяясь от нее слизистой оболочкой среднего уха. Наружное ухо у млекопитающих и человека развивается из жаберного кармана. Его средний участок образует ямку ушной раковины, превращающуюся постепенно в полость ушной раковины. Возникает наружный слуховой проход. Мезодермального происхождения клеточный материал, находящийся на границе между барабанной полостью и полостью наружного уха, дифференцируется в барабанную перепонку. 4. Зубы. Зубы развиваются из общего эпителиального утолщения эктодермального происхождения и из мезенхимной ткани, в которую это утолщение вдается. Из эпителиального утолщения формируется тяж – зубная пластинка (рис. 49). Рис. 49. Стадии развития зуба На ее внутренней поверхности появляются колбовидные выросты, из которых возникают эмалевые органы (рис. 49). Навстречу каждому из них в виде зубного сосочка растет мезенхима. Эмалевый орган имеет форму колпачка и называется также эмалевым колпачком. Внутренний, прилежащий к мезенхиме слой колпачка состоит из цилиндрических эпителиальных клеток (амелобластов), выделяющих на своей поверхности белковые вещества, называемые зубной эмалью. XI. Органогенез 173 Мезенхимные клетки наружных сосочков, прилежащих к эмалевым колпачкам, дифференцируются, превращаясь в преодонтобласты. Эти клетки синтезируют и секретируют дентиновый коллаген, дентиновые фосфопротеиды, вначале формирующие предентин, который, минерализуясь, превращается в дентин. Из зубного сосочка развивается зубная мякоть, куда позже прорастают нервы и кровеносные сосуды. 5. Развитие рта. Начальный отдел пищеварительного тракта имеет эктодермальное происхождение. В месте контакта передней кишки с кожной эктодермой на последней образуется ротовая ямка. Отверстие ротовой ямки у зародыша млекопитающих сначала представляет дыру, по окружности которой образовалось пять выступов: верхний непарный – лобный; два парных верхних выступа – верхнечелюстные отростки; два нижних – нижнечелюстные, которые срастаясь между собой образуют нижний край ротового отверстия (рис. 50, Б, В). Рис. 50. Этапы формирования лица у зародыша человека (по Б. Пэттену): 1 – лобный отросток; 2 – носовые плакоды; 3 – ротовая бухта; 4 – верхнечелюстной отросток; 5 – нижнечелюстная дуга; 6 – гиоидная дуга 174 Болотов А. В. Биология размножения и развития В дальнейшем одновременно с формированием лобного отростка обонятельных плакод на нем возникает средний лобный и два боковых носовых отверстия (рис. 50, Б–Д). По мере роста верхнечелюстные отростки срастаются с боковыми носовыми и со средним лобным отростками, образуя верхний край ротового отверстия. На среднем лобном отростке возникает поперечный носовой валик – зачаток кончика носа (рис. 50, Д, Е). Отверстия обонятельных ямок преобразуются в наружные носовые отверстия. Боковые носовые отростки превращаются в крылья носа. Процесс образования лицевых компонентов оканчивается в постэмбриональный период. Первичная ротовая полость постепенно подразделяется на носовою и собственно ротовую. Верхняя стенка ротовой полости – первичное мягкое небо. Задние концы обонятельных ямок, достигая мягкого нёба, открываются в первичную ротовую полость с помощью образующихся отверстий – первичных хоан. Глоточная перепонка, являющаяся задней границей первичной ротовой полости, перфорируется, вследствие чего эта полость присоединяется к средней кишке. На внутренних сторонах обоих верхнечелюстных отростков образуются нёбные отростки, срастающиеся и разделяющие первичную ротовую полость на собственно ротовую и носовую, в которую открываются первичные хоаны. На внутренней поверхности среднего лобового отростка образуется носовая перегородка, разделяющая носовую полость на правую и левую. Края небных отростков срастаются между собой и с нижним краем носовой перегородки, вследствие чего нёбо приобретает окончательное строение. В дальнейшем происходит формирование носовых раковин из складок, возникающих на поверхности носовых полостей. Лицевые части формируются в течение 2-го месяца и уже на 6-й неделе хорошо обозначаются. Образуются глазные впадины, появляются веки, обособляются губы. Нос остается еще широким и плоским. Язык возникает в виде небольшого выступа нижней поверхности глоточной кишки между челюстной и подъязычной дужками. Из эктодермы также образуются: хрусталик и роговица глаза, слизистые оболочки полости рта и полости носа. XI. Органогенез 175 2. Развитие производных энтодермы 1. На 3–4-й неделе – обособление тела зародыша от внезародышевых органов (желточного мешка, амниона) происходит путем образования так называемой туловищной складки – глубокой борозды (находится в головном отделе). По мере углубления этой складки энтодерма зародыша втягивается внутрь его тела, а желточный мешок обособляется. Одновременно тело зародыша погружается в полость амниотического пузырька, внутри которого происходит его дальнейшее развитие. Полость амниотического пузырька постепенно увеличивается, стенка его приближается к хориону, а желточный мешок редуцируется. Эктодермальный (амниотический) и энтодермальный (желточный) пузырьки тесно связаны друг с другом. Амниотический пузырек образуется раньше чем желточный. Хорион – ворсистая оболочка зародыша. Тело зародыша из плоского щитка превращается в объемное, эктодерма покрывает зародыш со всех сторон, а энтодерма зародыша после отделения от желточного мешка, оказывается внутри тела зародыша, и представляет собой кишечную энтодерму. Она сворачивается в замкнутую на концах трубку – зачаток будущей кишки (первичную кишку). По мере развития зародыша первичная кишка подразделяется на: переднюю (головную), среднюю и заднюю. В начальном отделе передней кишки образуется глоточный аппарат, участвующий в формировании лица, органов полости рта и шейной области. Глоточный аппарат состоит из пяти пар глоточных карманов и такого же количества глоточных дуг и щелей. Из структур глоточного аппарата первыми появляются глоточные карманы (рис. 51, Б) – выпячивания энтодермы в области боковых стенок глоточного отдела первичной кишки. Четыре пары карманов появляются так же, как и глоточные дуги, по кранио-каудальному градиенту. Пятая пара глоточных карманов отсутствует или рудиментарна. Эмбриональный материал глоточных карманов служит источником для многих важных структур головы и шеи. Первая пара глоточных карманов расположена между первой и второй глоточными дугами. Наиболее важная структура, образующаяся из первой пары глоточных карманов, – 176 Болотов А. В. Биология размножения и развития слуховая труба (рис. 51, Б). Из материала второй пары глоточных карманов и частично стенки глотки формируются нёбные миндалины (рис. 51, Б). Третья и четвёртая пары глоточных карманов участвуют в закладке тимуса и паращитовидных желез (верхняя и нижняя) (рис. 51, Б). Рис. 51. Развитие первичной кишки. А – Жаберные карманы и щели у 5-недельного зародыша человека (по Carlson, 1981) и развитие легких. Б – Глоточные карманы и щели; В – Развитие органов пищеварительной системы В области передней кишки формируются углубления – 4 пары жаберных карманов, часть которых превращается в сквозные жаберные щели. У зародыша человека первая пара жаберных карманов превращается в евстахиевы трубы, соединяющие XI. Органогенез 177 полости среднего уха с ротовой полостью (рис. 51, А), барабанную полость и наружный слуховой проход. Вторая пара дает начало тонзиллярный синусу. Из других жаберных карманов развиваются железы внутренней секреции: третья пара дает начало парному зачатку зобной железы (тимусу) (рис. 51, А), четвертая – паращитовидным железам (рис. 51, А). Из кармана Ратке формируется зачаток аденогипофиза (рис. 51, А). Из энтодермы дна глотки в виде непарного выпячивания формируется щитовидная железа. На уровне четвертого жаберного кармана из стенки передней кишки образуется в виде выпячивания непарный вырост – зачаток органов дыхания (гортань, дыхательная трубка и легкие). Конец выпячивания, расширяясь и дифференцируясь, разветвляется на две ветви, вырастающие в сторону и назад, которые преобразовываются в бронхи и легкие (рис. 51, А). Срединная часть развивается в трахею. На верхнем конце выпячивание, преобразовавшись в трубку, остается связанным с кишкой; при этом передний расширенный участок ее дифференцируется в гортань. У птиц и млекопитающих в результате обильного ветвления бронхов, на их концах образуются многочисленные легочные пузырьки (альвеолы). Развитие легких происходит на ранних стадиях, а их функционирование – после рождения. На 8-й неделе закладывается хрящевой скелет дыхательной трубки. Передняя часть передней кишки также участвует в развитии полости рта, полости глотки, пищевода, большей части двенадцатиперстной кишки (duodenum), печени, поджелудочной железы, желудка (рис. 51, В). Средняя часть – источник формирования эпителия задней половины двенадцатиперстной кишки, части поджелудочной железы, а также всего остального кишечника, оканчивается слепым выростом. Задняя часть – в печеночный дивертикулум (полый слепой вырост). Эмбриональная (первичная) кишка вначале замкнута, в ее переднем и заднем концах имеются впячивания эктодермы – ротовая (будущая ротовая полость) и клоакальная (заднепроходная) ямки. На 3–4-й неделе прорывается передняя перепонка, на третьем месяце – задняя. 178 Болотов А. В. Биология размножения и развития Печень у всех позвоночных возникает из энтодермального эпителия брюшной стороны двенадцатиперстной кишки. Зачаток печени подразделяется на две части: переднее выпячивание, образующее собственно зачаток печени, и заднее – зачаток желчного пузыря (рис. 51, В). Параллельно развивается и поджелудочная железа из двух выпячиваний кишечной трубки (рис. 51, В). В дальнейшем оба зачатка сближаются и срастаются, открываясь единым протоком. 3. Развитие производных мезодермы Из мезодермы формируется – мышечная, соединительная, хрящевая, костная ткани и эмбриональная соединительная ткань – мезенхима, кровеносная система, выделительная система, почки, половые железы. В конце 3-й недели развития начинается дифференцировка мезодермы. У всех позвоночных имеются осевая (рис. 52, 6–8) и боковая мезодермы (рис. 52, 10–12, 14). Рис. 52. Формирование сомитов. 1 – хорда; 2 – нервная трубка; 3 – нервный канал; 4 – эктодерма; 5 – энтодерма; 6 – склеротом; 7 – дермотом; 8 – миотом; 9 – париетальный листок мезодермы; 10 – висцеральный листок мезодермы; 11 – вторичная полость тела (целóм); 12 – нефротом; 13 – дуга аорты; 14 – внеэмбриональная мезодерма (мезенхима); 15 – желточная энтодерма Осевая мезодерма расположена по бокам хорды и подразделяется на спинные сегменты – сомиты, число пар которых на 34й день развития достигает 43–44. Из клеток сомита образуется XI. Органогенез 179 три основные закладки (части): переднемедеальная – склеротом (рис. 52, 6), из которого развиваются кости и хрящи скелета; латеральнее расположен миотом (рис. 52, 8), из которого формируется поперечно-полосатая скелетная мускулатура; кнаружи лежит дерматом, (рис. 52, 7) из которого возникает собственно кожа. Боковая мезодерма – спланхнотомы (боковые пластинки являются продолжением мезодермы сомитов) представляет из себя более рыхлые клеточные массы, не подразделяющиеся на сомиты и не утолщающиеся (рис. 52, 9, 10, 14). Спланхнотомы образуют две пластинки: прилежащие к первичной кишке – медиальная (висцеральная, внутренняя) называется спланхноплеврой (стенка внутренностей); прилежащая к стенке тела зародыша латеральная (наружная) пластинка – соматоплеврой (стенка тела). Из данных пластинок развивается мезотелий серозных оболочек, а пространство между ними превращается в полости (брюшинную, плевральную и перикардиальную). Из мезенхимы спланхнотома образуются клетки крови, гладкая мышечная ткань, кровеносные и лимфатические сосуды, соединительная ткань, а также корковое вещество надпочечника и эпителий половых желез (яичек, яичников). На границе между сомитами и спланхнотомами из мезодермы образуются нефротомы (рис. 52, 12), из которых развивается эпителий почек и семявыносящих путей. 1. Развитие осевого скелета. Опорной системой тела в онтогенезе разных позвоночных служит хорда. На следующих этапах развиваются частично хрящевые структуры. У большинства позвоночных на более поздних стадиях онтогенеза хрящевой скелет дополняется и в большей своей части замещается костным скелетом. Источником для развития осевого скелета служит склеротом. Мезенхимные клетки, из которых состоят сомиты, размножаясь, заполняют пространство, окружающее хорду, покрывают ее и оказываются источником развития хрящевых тел и дуг позвонков. Мезенхима склеротома распространяется, покрывая развивающийся спинной мозг на всем его протяжении, хвостовые артерии и вены. С каждой стороны хорды развиваются треугольные массы, углы которых постепенно вытягиваются в спинном, брюшном и срединном направлениях – это зачатки 180 Болотов А. В. Биология размножения и развития невральных, поперечных и хордальных отростков позвонков (рис. 53, 1–3). Стороны хордальных отростков срастаются друг с другом, формируя позвонки. Вскоре происходит охрящевение позвонков. Заключенная в образующийся столб, хорда впоследствии почти целиком дегенерирует, точнее, редуцируется до межпозвоночных прослоек. Хрящевой, а позднее костный позвоночный столб замещает функционально хорду. Хрящевые ребра развиваются в спинных отделах межмышечных связок. Они растут в брюшном направлении, концы выросших хрящевых ребер сливаются друг с другом, образуя зачаток грудины. Череп образуется по бокам и впереди переднего конца хорды. Сначала развивается и дифференцируется соединительная ткань, окружающая головной мозг и развивающиеся органы слуха, зрения и обоняния. Этот перепончатый череп постепенно охрящевеет, а впоследствии происходит окостенение. Рис. 53. Поперечный разрез эмбриона человека 12 недель (коллекция Мичиганского университета). 1 – поперечный отросток; 2 – спинальный отросток прилежащего позвонка; 3 – латеральный центр окостенения; 4 – мышцы спины; 5 – реберный центр окостенения; 6 – головка ребра; 7 – твердая оболочка (dura mater); 8 – срединный центр окостенения в теле позвонка; 9 – подпаутинное пространство; 10 – мягкая оболочка (pia mater); 11 – бугорок ребра XI. Органогенез 181 2. Развитие сердца. Сердце человека начинает развиваться очень рано (на 17-й день внутриутробного развития) из двух мезенхимных закладок, которые превращаются в трубки неравномерной толщины (рис. 54, А). Эти трубки затем сливаются в непарное простое трубчатое сердце, расположенное в области шеи, которое кпереди переходит в примитивную луковицу сердца, а кзади – в расширенный венозный синус. В дальнейшем происходит изменение ее формы и она подразделяется на четыре основные части: 1) венозный синус, 2) предсердие, 3) желудочек, 4) артериальный конус (рис. 54, Б). Рис. 54. Последовательные стадии (А–Д) развития сердца (вид с вентральной стороны). 1 – к аортам, 2 – от вен, 3 – артериальный конус, 4 – желудочек, 5 – предсердие, 6 – венозный синус, 7 – аорта, 8 – легочная артерия, 9 – правый желудочек, 10 – левый желудочек Его передний отдел артериальный, задний – венозный. Быстрый рост фиксированного среднего отдела трубки приводит к тому, что сердце изгибается S-образно (рис. 54, В). В нем выделяют предсердие, венозный синус, желудочек и луковицу с артериальным стволом. На внешней поверхности сигмовидного сердца появляются предсердно-желудочковая борозда (будущая 182 Болотов А. В. Биология размножения и развития венечная борозда дефинитивного сердца) и луковичножелудочковая борозда, которая после слияния луковицы с артериальным стволом исчезает. Предсердие сообщается с желудочком узким предсердножелудочковым (ушковидным) каналом. В его стенках и у начала артериального ствола образуются валики эндокарда, из которых формируются атрио-вентрикулярные клапаны, клапаны аорты и легочного ствола. Общее предсердие быстро растет, охватывает сзади артериальный ствол, с которым к этому времени сливается примитивная луковица сердца. По обеим сторонам артериального ствола спереди видны два выпячивания – закладки правого и левого ушек. На 4-й неделе появляется межпредсердная перегородка, она растет вниз, разделяя предсердия. Верхняя часть этой перегородки прорывается, образуя межпредсердное (овальное) отверстие. На 8-й неделе начинают формироваться межжелудочковая перегородка и перегородка, разделяющая артериальный ствол на легочный ствол и аорту. Сердце становится четырехкамерным (рис. 54, Г, Д). Венозный синус сердца сужается, превращаясь вместе с редуцировавшейся левой общей кардинальной веной в венечный синус сердца, который впадает в правое предсердие. 3. Развитие выделительной системы. У губок и кишечнополостных нет выделительных органов; клетки у этих организмов могут освобождаться от ненужных продуктов обмена благодаря своим клеточным механизмам. Для низших червей, у которых нет кровеносной системы, забирающей продукты обмена из тканей и выносящей их к почкам, органами выделения служат – протонефридии (рис. 55, А). Это сильно разветвленные, проникающие во все участки тела трубочки, открывающиеся на поверхности кожи порами. Концы разветвления не имеют отверстий, а состоят из одной или нескольких клеток с ресничным аппаратом. Благодаря колебательным движениям длинных жгутиков ток жидкости направляется к выходному отверстию. У кольчатых червей, которым свойствен целóм и кровеносная система, имеются метанефридии-трубочки, воронкообразный конец которых открывается в полость тела, а другой конец – на поверхности кожи. Ресничный аппарат воронки обеспечивает направление тока жидкости из целомической полости во внешнюю среду. XI. Органогенез 183 Рис. 55. Строение и развитие выделительных систем животных. A – протонефридия кольчатого червя: 1 – соленоциты; 2 – выделительная трубка протонефридия; 3 – ядро; 4 – жгутик соленоцита. Б – схема метонефридии аннелиды. В, Г, Д – Схема развития мочеполовой системы у высших наземных позвоночных (В – исходная стадия; Г – мочеполовой аппарат самки; Д – мочеполовой аппарат самца): 1 – предпочка (пронефрос); 2 – первичная почка (мезонефрос); 3 – вторичная почка (метанефрос); 4 – гонады; 5 – яичник; 6 – семенник; 7 – мочевой пузырь; 8 – вольфов канал; 9 – мюллеров канал; 10 – прямая кишка; 11 – мочеточник; 12 – мочеиспускательный канал; 13 – матка; 14 – придаток яичника (остаток первичной почки); 15 – придаток семенника (видоизмененная первичная почка) 184 Болотов А. В. Биология размножения и развития Через метанефридии выводятся наружу и половые клетки (рис. 55, Б). У позвоночных развиваются почки – совокупность мочевыносящих, секретирующих канальцев. Канальцы почки позвоночных связаны с кровеносными сосудами в единое целое. У низших позвоночных (рыбы и амфибии) в зародышевом периоде развивается примитивно построенная головная почка или предпочка (пронефрос), сменяющаяся потом туловищной или первичной почкой (мезонефрос). У зародышей высших позвоночных (рептилий, птиц и млекопитающих) закладывается зачаточный, вскоре редуцирующийся пронефрос (рис. 55, В, Г, Д). Затем развивается и функционирует некоторое время первичная почка, которую можно назвать в связи с ее локализацией туловищной почкой; затем у ее задних границ образуется и функционирует окончательная наиболее совершенного строения вторичная почка (метанефрос) – это тазовая почка (рис. 55, В, Г, Д). Предпочка представляет объемный орган, состоящий из извитых канальцев, открывающихся с одной стороны в целóм воронкой с мерцательным эпителием, а с другой – в общий мочевыносящий канал – первичный мочеточник или вольфов канал (рис. 55, В–Д). На уровне предпочки формируются, свешиваясь в полость тела, сосудистые клубочки – гломусы, инфильтрующие из крови в целом продукты метаболизма и воду. Благодаря воронкам канальцев предпочки с их ресничным аппаратом моча – продукты метаболизма и вода – вбирается и выводится из тела во внешнюю среду первичным мочеточником, который развивается из собственного зачатка, закладывающегося у задних границ предпочки в виде плотного клеточного скопления. При образовании первичной почки устанавливается связь почечных канальцев с кровеносной системой. Главным отличием первичных почек от предпочек заключается в том, что продукты метаболизма поступают в экскреторные органы не из целóма, а непосредственно из крови. Первичная почка развивается из большого числа туловищных нефротомов, канальцы ее не образуют воронок и не имеют связи с целомом. И канальцы, и протоки у всех позвоночных – производные мезодермы, они развиваются из нефротомов. Вторичная почка – дефинитивная почка (у птиц и млекопитающих) развивается из нефротомов задних сегментов туловища и из эпителиального зачатка вторичного мочеточника. XI. Органогенез 185 У человека в начале 4-й недели концевые отделы вольфовых каналов утолщаются и на дорсальной стороне утолщений образуются небольшие выпячивания, которые врастают в нефрогенные массы клеток и превращаются в каналы – зачатки мочеточников, а возникающие на их концах расширения дают начало формированию почечных лоханок. Возникшие из стенок вольфовых каналов правый и левый мочеточники после разделения клоаки на прямую кишку и мочеполовой синус открываются в последний благодаря отверстиям вольфовых каналов. Когда происходит развитие и обособление от мочеполового синуса мочевого пузыря и мочеиспускательного канала, мочеточники обособляются от вольфовых каналов, на спинной стороне мочевого пузыря образуется самостоятельное отверстие мочеточника. 4. Развитие половой системы. Половые клетки возникают из первичных гоноцитов и позже заселяют половые железы. Герментативный эпителий – это соматическая ткань, образующая стенку половой железы. Сама железа на ранних стадиях представляет собой складку, вдающуюся в полость тела – так называемую половую складку. Эта складка постепенно заполняется окружающей мезенхимой, за счет которой развивается внутренняя (мозговая) часть железы. До определенной стадии развития половая железа имеет одинаковое для обоих полов строение. Затем под влиянием проникших в нее первичных половых клеток, а также в зависимости от гормонального баланса организма железа дифференцируется либо в семенник, либо в яичник. В эмбриогенезе позвоночных параллельно вольфовым каналам идут – мюллеровы каналы (рис. 55, В, Г, Д). У самцов они позже дегенерируют (рис. 55, Д), а у самок сохраняются и превращаются в яйцеводы (рис. 55, Г). Мужские и женские гонады позвоночных развиваются в задней половине тела из клеточного материала латеральной пластинки мезодермы. Зачаток гонад появляется в виде продольной пластинки. На поверхностях первичных почек образуются небольшие утолщения – половые валики. Они представляют собой компактную массу клеток, состоящую из нескольких слоев и выдающуюся в полость тела. 5. Развитие конечностей. На 5–8-й неделе у зародыша появляются зачатки вначале верхних, а затем нижних конечностей в виде кожных складок, в которые позднее врастают закладки 186 Болотов А. В. Биология размножения и развития костей, мышц, сосудов и нервов. Парные конечности позвоночных развиваются из мезенхимных клеток. У зародышей амфибий скопление мезенхимы происходит почти одновременно в районе будущих передних и задних конечностей. У рептилий, птиц и млекопитающих скопления мезенхимы происходят не только в районах развития конечностей, но и по всей длине в виде гребней. В этих областях эпидермис, покрывающий мезенхиму, несколько утолщается и выпячивается. Образуется почка конечностей, которая состоит из клеток мезодермального происхождения и клеток эпидермиса (рис. 56, А, Б). У амниот эктодерма верхушки конечности утолщается, образуя апикальный гребешок. По мере роста конечности меняется ее форма: апикальная часть расширяется и уплощается, зачаток конечности скручивается вокруг своей длинной оси. На апикальной поверхности возникают одновременно зачатки пальцев (см. рис. 56, А, Б). Рис. 56. Формирование пальцев на конечностях у лягушки (А) и человека (Б) Одновременно с внешней дифференцировкой конечности формируется ее внутренний скелет путем образования хрящей из сгущений мезенхимных клеток. Первым выделяется зачаток проксимального хряща – стилоподия, из которого в передней конечности разовьется плечевая кость, а в задней – бедренная. Затем образуются хрящи – зигоподия (локтевой и лучевой хрящи в передней конечности, большой и малый берцовые – в задней) и аутоподия (хрящи кисти или стопы и фаланг пальцев). Хрящи плечевого и тазового поясов формируются позже стило- XI. Органогенез 187 подиев, но раньше аутоподиев. В конечность прорастают кровеносные сосуды. К концу 8-й недели развития у зародыша человека имеются зачатки всех органов. Сформированы части тела: голова, туловище и зачатки конечностей; внешняя конфигурация приобретает черты, характерные для человека. Двухмесячный месячный зародыш человека принято называть плодом. Начиная с 3-го месяца развития, происходит дальнейший рост и дифференцировка органов и тканей. Также на 3-м месяце беременности женщина начинает ощущать движение плода. С этого времени и далее наиболее очевидным проявлением становится увеличение размеров плода. Однако все еще происходят и некоторые изменения. Нервная, а также кровеносная и дыхательная системы продолжают развиваться; плод еще не может выжить вне организма матери. Известен случай преждевременных родов, когда плод находился примерно на 20-й неделе развития; ребенок выжил, однако в течение многих недель он находился в особых условиях, обеспечивающих поддержание дыхания, питания и температуры тела. Органогенез завершается вылуплением из яйца или рождением детеныша. XII. РОСТ, РАЗВИТИЕ, ДИФФЕРЕНЦИРОВКА, МОРФОГЕНЕЗ, РОЛЬ ГЕНОВ Организм взрослого человека состоит примерно из 1015 или около одного квадриллиона – клеток, и каждая из них с самого начала формируется сама и занимает правильное положение среди других клеток. Каждая клетка, поэтому помогает формировать и конструировать по общему плану развития весь организм. Рост – процесс увеличения какого-либо качества со временем. Развитие – это упорядоченная последовательность прогрессивных изменений, приводящих к повышению уровня сложности. Форма того или иного органа или целого организма зависит от множества процессов, переплетенных в сложном взаимодействии. В конечном счете она зависит от генов, определяющих наследственные признаки, и факторов среды, которые создают условия для работы генов. В процессе развития можно наблюдать и анализировать явления дифференцировки, роста и морфогенеза. Дифференцировка – это комплекс изменений, приводящих к прогрессивному увеличению разнообразия структуры и функции клетки. Этот процесс зависит от многих причин, и в частности от химических веществ (если в воде имеются соли Li, то у животных (морской еж) меняется образование зародышевых листков). Дифференцировка может протекать как самодифференцировка, а она – важнейшая часть всякого развития. Морфогенез (от греч. morphê форма и genesis происхождение, или «формообразование») – это процесс возникновения новых структур и изменения их формы как в индивидуальном, так и в историческом, или эволюционном, развитии. Морфогенез, как рост и клеточная дифференцировка, относится к ациклическим процессам, т. е. не возвращающимся в прежнее состояние и по большей части необратимым. Он определяется последовательностью переключения экспрессии групп генов, однако эти гены пока не столь детально изучены. XII. Рост, развитие, дифференцировка, морфогенез, роль генов 189 Считается, что все клетки, произошедшие от зиготы практически одинаковы, но среди множества их генов, есть два, способные включать и выключать другие гены. Клетки, в своей массе включают и выключают активные и тормозные вещества, которые проникают в близлежащие клетки «включая» и «выключая» другие. Так они формируют ткани и органы зародыша. И по мере роста зародыша, увеличения его массы, меняется и его форма. Во время развития зародыша происходит дифференциация и перемещение его клеток с образованием тканей и органов. Рассмотрим на примере мухи-дрозофилы этот процесс. В развитии мухи происходит последовательная смена форм, значительно отличающихся друг от друга: яйцо, личинка, куколка и имаго (взрослая особь). Такое развитие называется развитием с метаморфозом (с греч. – изменение формы) (рис. 57). Рис. 57. Развитие дрозофилы: личинка, куколка и имаго Цитоплазма яйцеклетки не гомогенна, в ней асимметрично распределены различные биологически активные компоненты. У эмбриона уже определены оси тела, число и ориентация сегментов тела, из которых затем развиваются части тела взрослой мухи. Эти процесс контролируются различными наборами генов, которые называются морфогены. Они кодируют белки, ко- 190 Болотов А. В. Биология размножения и развития торые регулируют экспрессию других генов, отвечающих за формирование органов. Градиент концентрации белков-морфогенов определяет передне-заднюю и дорсо-вентральную (спинно-брюшную) оси тела. У дрозофилы в определении дорсо-вентральной оси тела участвуют 12 генов, главный из них – dorsal. Белок Dorsal сконцентрирован в цитоплазме на той стороне эмбриона, которая станет спинной, и проникает в ядро клеток на брюшной стороне, активируя группы генов, продукты которых необходимы для определения осей тела. Детерминация передне-задней оси тела контролируется другими генами. Один из них называется bicoid, его мРНК сконцентрирована в цитоплазме передней части яйца (фиксирована своим 3'-концом). В результате при трансляции возникает градиент концентрации белка Bicoid от переднего к заднему концу яйца. Градиент поддерживается продолжительным периодом синтеза белка и его коротким временем жизни. Bicoid, так же как и Dorsal, морфоген, он активирует гены, которые необходимы для развития головы и грудных структур. Его экспрессия ингибируется продуктом гена nanos, сконцентрированного на заднем конце эмбриона (рис. 58). Рис. 58. Последовательная работа генов, устанавливающая передне-заднюю, спинную и брюшные оси тела и ориентацию сегментов На следующем этапе включаются гены сегментации. Они контролируют дифференциацию эмбриона на индивидуальные сегменты (рис. 58). После оплодотворения транскрибируется около 25 генов сегментации, их экспрессия регулируется градиентами белков Bicoid и Nanos. XII. Рост, развитие, дифференцировка, морфогенез, роль генов 191 192 Болотов А. В. Биология размножения и развития 1. Гомеозисные гены После сегментации и установления ориентации сегментов активируются так называемые гомеозисные гены. Различные их наборы активируются специфическими соотношениями концентраций белков, упоминавшихся выше. Продукты гомеозисных генов активируют другие гены, которые определяют сегмент-специфичные особенности. Глаза в норме возникают только на головном сегменте, а ноги – только на грудных сегментах. Гомеозисные гены кодируют регуляторные белки, связывающиеся с ДНК. Каждый из них содержит кластер нуклеотидов, называемый гомеобоксом, который сходен во всех гомеотических генах. Он содержит 180 нуклеотидов и кодирует 60 аминокислот, функционирующих как ДНК-связывающий домен. Впервые мутации гомеозисных генов были идентифицированы в 1894 г., когда Уильям Бэтсон заметил, что у растений иногда части цветка появляются на неправильных местах. Гомеозисные гены как бы определяют адрес клетки в конкретном сегменте, сообщая клеткам, в каком районе они находятся. Когда они мутируют, клетки получают «ложный адрес» и ведут себя так, будто они находятся в другом месте эмбриона (рис. 59). Рис. 59. Нарушение в работе гомеозисных генов. Порядок расположения гомеозисных генов соответствует порядку контролируемых ими сегментов тела XII. Рост, развитие, дифференцировка, морфогенез, роль генов 193 После того, как были открыты и изучены гомеозисные-гены дрозофилы, сходные гены были найдены у всех других животных от нематоды до человека. У млекопитающих они называются НОХ-генами (гомеобокс-содержащими генами), и также кодируют белки, регулирующие транскрипции и определяющие структуры тела и их положение в передне-заднем направлении (рис. 60). Рис. 60. Гомология работы гомеозисных генов дрозофилы (геомео-гены) и млекопитающих (мышь и человек) НОХ-гены в определении передне-задней оси и сегентации тела Таким образом, в эмбриональном развитии исходный градиент-белков и мРНК в яйцеклетке стимулирует локальную экспрессию генов эмбриона, которая ведет к дальнейшей дифференциации генной экспрессии и определяет судьбу клеток развивающегося эмбриона. Процесс, в котором формируются конечности, ткани и органы, называется морфогенезом, и определяется последовательностью переключения экспрессии групп 194 Болотов А. В. Биология размножения и развития генов, однако эти гены пока не столь детально изучены. Морфогенез – буквально означает образование новой формы или принятие новой формы. Он связан с дифференциальным ростом, но чаще связан с организованным движением клеток и групп клеток. Морфогенез может идти при перераспределении клеток в пространстве и во времени. На ход нормального развития влияет взаимодействие частей зародыша, нарушение которого может привести к уродствам. Рис. 61. Реализация разного эпигенитического ландшафта при взаимодействии генов со средой В процессе эмбриогенеза осуществление записанной в генах программы развития происходит в конкретных условиях среды. Взаимодействие генов и среды можно описать на следующей модели (рис. 61). Эмбриональное развитие можно сравнить с шариком, катящимся по наклонной поверхности с разными желобками. Такое представление эмбрионального развития, на- XII. Рост, развитие, дифференцировка, морфогенез, роль генов 195 званное эпигенетическим ландшафтом, было предложено Кондратом Уоддингтоном. Действие окружающей среды может быть и вредным, то есть у организма под действием внешней среды возникают отклонения развития при совершенно нормальных генах. Для примера рассмотрим один случай (см. рис. 61). В 60-х годах в Германии было сильно разрекламировано новое снотворное под названием талидомид (см. рис. 61). Среди принимавших новое лекарство были беременные женщины. Спустя некоторое время было замечено, что в стране стало рождаться много детей с патологией конечностей. У них отсутствовали длинные кости конечностей, т. е. прямо от основания тела начинались кисти или ступни (см. рис. 61). Раньше такое заболевание встречалось один раз на несколько тысяч новорожденных, и вдруг такой всплеск. Начали проводиться исследования, и выяснилось, что причина в новом лекарстве. Как оказалось, талидомид имеет большое сродство к гуанину. Взаимодействуя с ДНК, он может приводить к функциональным нарушениям. Промотор гена, отвечающего за рост и развитие длинных конечностей, содержит большое количество гуанина, таким образом, талидомид нарушает работу этих генов, и зачатки костей длинных конечностей так и не начинают развиваться. Многие из этих детей не выжили, часть из тех, кто выжил, ведут жизнь инвалидов, но есть среди них люди, которые, несмотря на инвалидность, реализовали свои возможности. По мере развития происходит смена активно работающих генов. В разных частях зародыша функционируют разные гены, хотя есть гены, активные во всех клетках. Часть генов, активных в оогенезе не теряют своей активности и в эмбриональном развитии. Точнее – активной остается лишь часть генов. Некоторые генные системы, последовательно включающиеся в процессе развития, организованы по кластерному типу. Порядок расположения HOX-генов соответствует временной последовательности их активности в ходе развития. Подобного рода явления описаны и у некоторых эукариот (дрожжи). Колинеарность в расположении генов и контролируемых ими признаков. Этот удивительный феномен означает соответствие линейного расположения генов и линейной последовательности рас- 196 Болотов А. В. Биология размножения и развития пределения детерминируемых ими признаков в теле развивающегося зародыша. Об этом говорилось в главе о гомеозисных генах: признаки, которые гены контролируют (голова, грудь, брюхо), расположены точно в той же последовательности, что и соответствующие комплексы генов (ANT-C, ВХ-С, abd-A, AbdB). Иными словами, в определенном участке хромосомы как бы «нарисована» в молекулярных терминах – миниатюрная или мушка-дрозофила, или мышка, или человечек. Получается нечто похожее на то, о чем думали преформисты XVII в. Только они полагали, что в половой клетке присутствует миниатюрный человек в буквальном смысле этого слова, в действительности же его «образ записан», но в определенной структуре клетки, несущей наследственную информацию (хромосома), и на специфическом языке (молекулярно-генетическом). 2. Реализация программ морфогенеза Реализация программ морфогенеза происходит под воздействием комплекса генетических и негенетических (парагенетических) факторов. Генетические программы морфогенеза образованы двумя группами генов: 1) Гены, управляющие переключением: главные гены, «гены-господа». К ним относятся гены-регуляторы, продукты которых влияют на экспрессию других генов, и гомеозисные гены, продуцирующие морфогены – вещества, определяющие морфогенетические процессы. К морфогенам относятся как тканеспецифические вещества (например, гормоны), так и неспецифические низкомолекулярные соединения (ретиноивая кислота). 2) Гены, обеспечивающие переход от одного состояния (узла) к другому: исполняющие гены, «гены-рабы», продуктами которых являются ферменты, структурные белки. Историческая справка. Термины «гены-господа» («MasterGenes») и «гены-рабы» («Slaves-Genes») предложил шведский цитолог Ян-Эрик Эдстрем в начале 1960-х гг. Суперрегуляторные гены у дрозофилы открыл швейцарский эмбриолог и генетик Вальтер Геринг (начало 1990-х гг.). Термин «гомеозис» предложил У. Бэтсон в 1894 г. Под гомеозисом он понимал превращение одной части организма в другую. Гомеозис- XII. Рост, развитие, дифференцировка, морфогенез, роль генов 197 ные гены у дрозофилы открыли Эдвард Льюис (США) и Кристина Нюссляйн-Вольхардт и Эрик Вишхаус (Германия) (Нобелевская премия). Экспрессия всех генов контролируется разнообразными эффекторами. Часть из них закодирована в генотипе, часть – поступает в клетки извне или образуется в ходе метаболических реакций. Синтез эффекторов контролируется условиями внешней среды, например, белки «теплового шока», регулирующие процессы транскрипции, синтезируются у дрозофилы при температуре свыше 35°С, при воздействии антибиотика антимицина А, гидроксиламина, колхицина, хлорида аммония и других веществ. Регуляция экспрессии всех генов происходит на различных уровнях: 1. Регуляция на генном уровне происходит различным образом. 1.1. Модификация ДНК (например, замена цитозина или гуанина на метил-цитозин или метил-гуанин; метилирование оснований снижает активность генов). 1.2. Увеличение объема ДНК в клетке путем дифференциальной амплификации ДНК (например, многократное копирование генов рРНК) или за счет образования политенных хромосом. 1.3. Программированные количественные изменения ДНК (например, изменение ориентации промотора). 1.4. Сплайсинг ДНК (например, вырезание участков генов, кодирующих антитела). 1.5. Диминуция хроматина – необратимая утрата части генетического материала в соматических клетках некоторых организмов (инфузорий, аскарид, циклопов). 1.6. Изменение активности целых хромосом (например, инактивация одной из двух X–хромосом у самок млекопитающих). 1.7. Изменение последовательностей ДНК с помощью подвижных генетических элементов, например, транспозонов. 2. Регуляция на уровне транскрипции – путем регуляции транскрипции мРНК. Интенсивное функционирование отдельных генов или их блоков соответствует определенным этапам развития и дифференцировки. Регуляторами транскрипции у животных часто являются стероидные гормоны. 198 Болотов А. В. Биология размножения и развития 3. Регуляция на уровне сплайинга (посттрансляционной модификации мРНК) – обеспечивает возможность образования различных типов зрелой, функционально активной мРНК. Процессинг РНК регулируется с помощью рибозимов (катализаторов рибонуклеиновой природы) и ферментов матураз. Некоторые генетические заболевания человека (фенилкетонурия, некоторые гемоглобинопатии) обусловлены нарушением сплайсинга. 4. Регуляция на уровне трансляции – обусловлена различной активностью разных типов мРНК. 5. Регуляция на уровне посттрансляционной модификации белков – регулируется путем посттрансляционной модификацией белков (фосфорилированием, ацетилированием, расщеплением исходной полипептидной цепи на более мелкие фрагменты и т. д). Рассмотренные примеры свидетельствуют о многообразии способов реализации генетической информации путем регуляции активности самих генов либо их продуктов. Следует, однако, отметить, что для клетки наиболее экономична регуляция на уровне транскрипции, поскольку она препятствует образованию соответствующих мРНК и белков, когда клетка не испытывает в них потребности. Вместе с тем регуляция на уровне транскрипции идет сравнительно медленно, тогда как, например, активация белков путем расщепления молекул-предшественников хотя и неэкономична, но происходит очень быстро. Гомеозисные мутации. При нарушении структуры гомеозисных генов возникают гомеозисные мутации, которые изменяют порядок экспрессии исполняющих генов. Фенотипический эффект гомеозисных мутаций заключается в превращении одних органов в другие. Например, у мушки дрозофилы мутация группы генов bithorax, контролирующих развитие грудных и брюшных сегментов у дрозофилы, может приводить к появлению крылоподобных образований вместо галтеров. Мутации группы генов antennapedia выражаются в том, что у насекомых на месте антенн вырастают ножки. Мутации ophthalmoptera приводят к развитию крыла из имагинального диска глаза. Мутации proboscipedia приводят к развитию ноги или части антенны (в зависимости от температуры) вместо хоботка. У мутантов tumorous head ткани головы за- XII. Рост, развитие, дифференцировка, морфогенез, роль генов 199 мещаются другими типами тканей, включая структуры, характерные для гениталий. Основные свойства онтогенеза: 1. Исходная запрограммированность процессов. Наличие уникальной неизменной генетической программы развития, сформированной вследствие мейоза и оплодотворения. 2. Необратимость онтогенеза. При реализации генетической программы невозможен возврат к предыдущим стадиям. 3. Углубление специализации: по мере развития уменьшается вероятность смены траектории онтогенеза. 4. Адаптивный характер: поливариантность онтогенеза обеспечивает возможность приспособления к различным условиям. 5. Неравномерность темпов: скорость процессов роста и развития изменяется. 6. Целостность и преемственность отдельных этапов. Признаки, появляющиеся на более поздних стадиях, базируются на признаках, проявляющихся на ранних стадиях. 7. Наличие цикличности: существует цикличность старения и омоложения. 8. Наличие критических периодов, связанных с выбором пути в узловых точках (точках бифуркации) или с преодолением энергетических порогов. Условно можно выделить четыре группы генов: 1. Отвечает за образование ферментов энергетического обмена, за синтез больших молекул (белок, ДНК, РНК), за образование структур, общих для всех клеток. 2. Функционирует в одинаковых клетках какой-нибудь ткани (эти гены определяют синтез сократительных белков в мышечных тканях или коллагена – в опорных тканях). 3. Специфичные для определенного типа клеток. Специфичен синтез гормонов, медиаторов. 4. Регуляторные гены. Они включают и выключают активность генов 1-, 2- и 3-й групп. Одинаковые по внешнему виду клетки находятся в разных тканях, но они отличаются по активности генов. Так, например, формируется определенная форма хрящей в разных органах или определенная форма, структура мышц. Гены «лепят» эту форму в зависимости от места. XIII. ОНТОГЕНЕЗ 1. Особенности, свойства и виды онтогенеза Индивидуальное развитие особи называется онтогенезом (Э. Геккель, 1866). Особью, или индивидом (от лат. individuum – неделимый) называется неделимый далее организм (от лат. organizo и франц. organisme – устраиваю, придаю стройность). Главные существенные признаки особи – это её целостность, строгая взаимозависимость всех частей, органов и систем органов: разделить особь на части без потери морфофункциональной индивидуальности невозможно. Само выражение «особь» подразумевает обособленность: таким образом, особь обособлена, отделена от других подобных особей, она способна (хотя бы частично) к самостоятельному существованию. С эволюционной точки зрения, особью называется морфофизиологическая единица, происходящая от одного зачатка: от одной зиготы (при половом размножении), яйцеклетки (при партеногенезе), споры (при споровом размножении), почки или любого другого зачатка (при бесполом или вегетативном размножении). В дальнейшем зачаток, дающий начало новой особи, будем для краткости называть зиготой, поскольку именно при образовании зиготы возникают новые сочетания наследственных факторов, определяющих индивидуальность особи. Именно особь является объектом воздействия эволюционных факторов, в первую очередь – естественного отбора. Целостность и дискретность онтогенеза. Онтогенез особи начинается с момента её образования. Этим событием особи может быть прорастание споры, образование зиготы, начало дробления зиготы, возникновение особи тем или иным путем при вегетативном размножении (иногда начало онтогенеза относят к образованию исходных клеток, например, оогоний). В ходе онтогенеза происходят рост, дифференцировка и интеграция частей развивающегося организма. Онтогенез особи может завершиться её физической смертью или её воспроизведением (в частности, при размножении путем деления). XIII. Онтогенез 201 Каждый организм в период индивидуального развития представляет собой целостную систему, следовательно, и онтогенез – это целостный процесс, который не может быть разложен на простые составляющие части без потери качества. Однако существует морфологическая и функциональная дискретность онтогенеза, обусловленная дискретной генетической детерминацией. Реализация генотипа в онтогенезе изменчива и происходит приспособительно к конкретным условиям среды. Таким образом, генотип способен обеспечивать в определенных пределах изменчивость онтогенеза в зависимости от изменяющихся условий внешней среды. Степень возможной изменчивости в ходе реализации генотипа называется нормой реакции и выражается совокупностью возможных фенотипов при различных условиях среды. Это определяет так называемую онтогенетическую адаптацию, обеспечивающую выживание и репродукцию организмов иногда даже при значительных изменениях внешней среды. Необратимость онтогенеза. Онтогенез растений и животных состоит из качественно различных периодов: эмбриогенез, юность, зрелость и старость. Онтогенез многоклеточных организмов сопровождается рядом общих основных процессов: • рост – увеличение числа клеток и/или их объема (растяжение); • гистогенез – образование и дифференцировка тканей; • органогенез – образование органов и систем органов; • морфогенез – формирование внутренних и внешних морфологических признаков; • физиолого-биохимические преобразования. Все это происходит на основе биохимической, физиологической, генетической и морфологической дифференцировки клеток, тканей и органов. В ходе онтогенеза возникает ряд особенностей, обеспечивающих приспособление организма к окружающей среде. Онтогенез включает две группы процессов: морфогенез и воспроизведение (репродукцию). При соблюдении принципов дискретности и необратимости онтогенеза особь вначале должна использовать энергию для осуществления морфогенетических процессов, и лишь по достижении зрелости – для воспроизведения. 202 Болотов А. В. Биология размножения и развития Основные типы онтогенеза Существует множество основных типов онтогенеза и еще большее число производных типов, например: • Онтогенез организмов с бесполым размножением и/или при зиготном мейозе (прокариоты и некоторые низшие эукариоты). • Онтогенез организмов с чередованием ядерных фаз при споровом мейозе (большинство растений и грибов). • Онтогенез с чередованием полового и бесполого размножения без смены ядерных фаз. Чередование поколений с половым и бесполым размножением у Кишечнополостных называется метагенезом. Чередование партеногенетического и амфимиктического поколений у червей, некоторых членистоногих и низших хордовых называется гетерогонией. • Онтогенез с наличием личиночных и промежуточных стадий: от первично-личиночного анаморфоза до полного метаморфоза. • Онтогенез с утратой личиночных стадий и/или стадий бесполого размножения: пресноводные гидры, олигохеты, наземные и вторично-водные брюхоногие моллюски. • Онтогенез с утратой конечных стадий и размножением на ранних этапах онтогенеза; проявляется в виде педоморфозов (сохранении личиночных черт) и неотении (размножения на личиночной стадии). Особенности онтогенеза у животных У животных важную роль в регуляции онтогенетических процессов играют эндокринная и нервная системы. В онтогенезе высших животных выделяют следующие этапы (периоды) онтогенеза: 1) предзародышевый (преэмбриональный) – развитие половых клеток (гаметогенез) и оплодотворение; 2) зародышевый (эмбриональный) – развитие организма под защитой яйцевых и зародышевых оболочек или под защитой материнского организма; 3) послезародышевый (постэмбриональный) – до достижения половой зрелости; 4) взрослое состояние – размножение, забота о потомстве, старение и гибель. XIII. Онтогенез 203 В рамках эмбрионального периода различают следующие типы онтогенеза: 1) первично-личиночный – личинка способна к самостоятельному существованию (паренхимулы губок, планулы кишечнополостных, трохофоры полихет, головастики амфибий); 2) неличиночный (яйцекладный) – прохождение ранних этапов гисто- и морфогенеза под защитой яйцевых оболочек (представители губок, кишечнополостных, кольчатых червей, ракообразных и многие другие группы, утратившие первичноличиночные стадии) и зародышевых оболочек (насекомые с прямым развитием, яйцекладущие амниоты); 3) вторично-личиночный – характеризуется разнообразием вторичных типов личинок, например, свободноживущие (редко паразитические) личинки насекомых с полным превращением (личинки жуков, гусеницы бабочек и т.п.); в данном случае появляется особая стадия – стадия куколки; отдельно выделяются личинки-паразиты (например, у паразитических червей); 4) внутриутробный – зародыш развивается под защитой материнского организма; при этом различают яйцеживорождение (морфологических связей между зародышем и материнским организмом не возникает), истинное живорождение (у плацентарных млекопитающих) и множество промежуточных типов (например, у живородящих акул, у сумчатых млекопитающих). Смена типов эмбрионального развития повышает независимость гисто- и морфогенеза от внешней среды, способствует автономизации онтогенеза и возможности выхода в новую адаптивную зону. Особенности онтогенеза у высших растений. Для растений характерны жизненные циклы с чередованием полового и бесполого поколений со сменой ядерных фаз (гаплоидного гаметофита и диплоидного спорофита), а также вегетативное размножение каждого из названных поколений. В зависимости от особенностей жизненного цикла зачатком новой особи может считаться спора, зигота (или – при партеногенезе – яйцеклетка), семя или вегетативный зачаток (почка, более или менее видоизмененный побег и пр.). Наиболее полно разработано учение об онтогенезе у семенных растений. Целостность онтогенеза у этих организмов обеспечивается за счет образования и 204 Болотов А. В. Биология размножения и развития взаимодействия фитогормонов, а также за счет обмена метаболитами между органами и частями растений. В онтогенезе семенных растений выделяют следующие периоды: 1) предзародышевый (преэмбриональный) – развитие гаплоидных структур – микроспорогенез и образование пыльцевых зерен, мегаспорогенез и образование эндосперма с архегониями (у голосеменных) или зародышевого мешка (у покрытосеменных); опыление и оплодотворение; 2) зародышевый (эмбриональный) – развитие семени из семязачатка; 3) стадия проростка – проросток образуется при прорастании семени и существует за счет запасов питательных веществ; 4) ювенильная стадия – растение переходит к самостоятельному питанию; 5) имматурная стадия – происходит ветвление стебля, формируется корневая система; 6) виргинильная стадия – формируется общий облик взрослого растения (габитус); однако генеративные органы отсутствуют; 7) генеративная стадия – на этой стадии происходит семенное размножение: образуются генеративные органы: цветки, а затем семена и плоды; различают три этапа генеративной стадии: ранняя генеративная стадия, средняя и поздняя; 8) сенильная стадия – семенное размножение прекращается, и растение отмирает. Онтогенез растений в значительной степени зависит от условий внешней среды. В результате у них выработались защитные реакции (период покоя, фотопериодизм, термопериодизм), благодаря которым период активной жизнедеятельности приурочен к наиболее благоприятному времени года. Цикличность онтогенеза Онтогенез представляет собой циклический процесс. Внешне цикличность проявляется в виде повторения морфогенетических и/или физиолого-биохимических процессов. Особенно ярко эта закономерность проявляется у многолетних растений, онтогенез которых должен быть адаптирован к сезонным изменениям факторов среды. Отечественный ботаник Н.П. Кренке (1940) разработал теорию циклического старения и омоложения расте- 205 XIII. Онтогенез ний в онтогенезе. Согласно этой теории, развитие организма есть борьба и единство противоположных процессов в нем – старения и омоложения. Онтогенез определяется исходным потенциалом жизнеспособности, который обусловлен генетическими особенностями разных организмов; исходный потенциал жизнеспособности является эволюционно сложившимся признаком. Онтогенетическое старение выражается в циклическом понижении потенциала жизнеспособности этих частей, что неизбежно приводит к естественной смерти индивидуума. Омоложение есть новообразование и развитие молодых веществ и структур, а также задержка старения существующих элементов, но не возврат индивидуума или его частей к прошлому. Таким образом, цикличность вовсе не означает возврат на предыдущие этапы онтогенеза; онтогенез – это процесс необратимый. Представления Н.П. Кренке о цикличности онтогенеза широко используются в современной биологии индивидуального развития. 2. Онтогенез человека Полный цикл индивидуального развития делят на два периода: пренатальный (внутриутробный) и постнатальный (внеутробный). Основные периоды пренатального онтогенеза у человека Периоды пренатального онтогенеза (начальный, зародышевый и плодный). Начальный период охватывает первую неделю развития (дробление). В течение зародышевого периода, который длится 8 недель, происходит формирование органов и частей тела, свойственных взрослому человеку. В фетальный (плодный) период главным образом увеличиваются размеры и завершается органообразование. Скорость роста плода возрастает до 4–5 месяцев. После 6 месяцев скорость роста линейных размеров уменьшается. По-видимому, одна из причин замедления роста в конце внутриутробного периода – ограниченные размеры полости матки. Скорость роста близнецов замедляется в тот период, когда их общий вес становится равным весу одиночного 36-недельного плода. Основные периоды постнатального онтогенеза у человека. Различают 11 периодов жизни в постнатальном онтогенезе у человека. 206 Болотов А. В. Биология размножения и развития 1. Сразу поле рождения наступает период новорожденности. В течение 8–10 дней происходит вскармливание ребенка молозивом. Аккомодация не работает. Это критический период в развитии ребенка. Должны быть сформированы три важнейших рефлекса: сосательный, глотательный, хватательный. 2. Грудной период продолжается до 1 года. Начало его связывают с переходом к питанию зрелым молоком. Во время этого периода наблюдается наибольшая интенсивность роста по сравнению со всеми остальными периодами внеутробной жизни. Длина тела увеличивается в 1,5 раза, а вес утраивается. С четырех месяцев начинают прорезываться молочные зубы. Руки становятся пропорциональной величины. Начинает работать аккомодационный аппарат. Ребенок начинает осознанно говорить, ходить. Начинает формироваться собственная иммунная система, аккомодационная система глаз. 3. Период раннего детства длится от одного года до трех лет. К концу второго года жизни заканчивается прорезывание молочных зубов. С этого момента абсолютные и относительные величины годичных приростов размеров тела быстро уменьшаются. Этот период важен, так называемым психическим формированием, т. е. формируется модель будущего характера и поведения человека. 4. Период первого детства (от 4 до 7 лет). Наблюдается небольшое увеличение скорости роста (первый ростковый скачок). Этот скачок не свойственен всем детям. Начиная с шести лет, появляются первые постоянные зубы. Возраст от 1 года до 7 лет называют также периодом нейтрального детства, поскольку мальчики и девочки почти не отличаются друг от друга по размерам и форме тела. Однако уже в этот период у девочек больше количество жира. 5. Период второго детства (мальчики 8–12 лет, девочки 8–11 лет). В этот период выявляются половые различия в размерах и форме тела, а также начинается усиленный рост в длину. Ноги достигают пропорциональной величины. Темпы роста у девочек выше, чем у мальчиков, так как половое созревание у девочек начинается в среднем на два года раньше. Примерно в 10 лет девочки обгоняют мальчиков по длине, весу тела, ширине плеч (первый перекрест ростковых кривых). К 12–13 годам за- XIII. Онтогенез 207 канчивается смена зубов (кроме третьих моляров). В период второго детства начинают развиваться вторичные половые признаки. У девочек сначала формируется грудная железа (средний возраст у различных этнических групп 9–10 лет). Одновременно развиваются матка и влагалище. В конце периода второго детства появляются волосы на лобке, а потом в подмышечных впадинах. У мальчиков процесс полового созревания выражен в меньшей степени. Лишь к концу периода у них начинается ускоренный рост яичек, мошонки, а затем полового члена (на 0,7 см). 6. Подростковый период (пубертатный период, период полового созревания) – мальчики 13–16 лет, девочки 12–15 лет. В этот период наблюдается дальнейшее увеличение скоростей роста – пубертатный скачок, который касается всех размеров тела. Наибольшие прибавки по длине тела у девочек имеют место между 11 и 12 годами, по весу тела – между 12 и 13 годами, у мальчиков соответственно – между 13 и 14 годами и 14 и 15 годами. Особенно велики скорости роста большинства размеров у мальчиков, в результате чего в 13,5–14 лет они обгоняют девочек по длине тела (второй перекрест ростковых кривых). К концу периода размеры тела составляют 90–97 % своей окончательной величины. В этот период происходит перестройка основных физиологических систем организма. У мальчиков в этот период особенно интенсивно развивается мышечная система. К концу периода основные функциональные характеристики подростков приближаются к характеристикам взрослого организма. В подростковый период формируются вторичные половые признаки. У девочек продолжается развитие грудных желез, рост волос на лобке и в подмышечных впадинах. Наиболее четким показателем созревания женского организма является первая менструация (менархе). Она обычно начинается после того, как пройдет максимум скорости роста тотальных размеров тела. В 1970–1980 гг. возраст появления менархе составляет около 13 лет в большинстве европейских стран. В сельской местности на 6–10 месяцев позднее. В экстремальных условиях – позднее. У мальчиков продолжается рост яичек и полового члена, особенно интенсивно в возрасте 13–14 лет. К 13 годам происходит мутация голоса и появляются волосы на лобке; к 14 годам наблюдается пубертатное набухание сосков, появляются волосы в подмышеч- 208 Болотов А. В. Биология размножения и развития ных впадинах. К 15 годам начинается рост волос на верхней губе и подбородке. В 14–15 лет у мальчиков появляются первые поллюции. У мальчиков по сравнению с девочками более продолжителен предпубертатный период и сильно выражен пубертатный скачок. Различия в размерах тела между взрослыми мужчинами и женщинами в значительной степени зависят от времени наступления, продолжительности и интенсивности пубертатного скачка роста. До этого скачка различия в росте между мальчиками и девочками не превышают 2 %, а после него они достигают в среднем 8 %. 7. Юношеский возраст продолжается у юношей от 17 до 21 года, у девушек – от 16 до 20 лет. В этот период заканчивается в основном процесс роста и формирование организма. Все основные размерные признаки тела достигают дефинитивной (окончательной) величины. 8. Зрелый возраст делится на два периода. Первый период длится от 22 до 35 лет у мужчин и от 21 до 35 лет у женщин; второй от 36 до 60 и от 36 до 55 соответственно. В этом возрасте форма и размеры тела изменяются мало. В первый период продолжается рост позвоночного столба за счет отложения новых слоев костного вещества на верхних и нижних поверхностях позвонков. Этот рост незначителен 3–5 мм. Во второй период длина тела остается постоянной. Наблюдается увеличение жирового слоя, увеличение веса, стабильный уровень функциональных показателей. В конце второго периода развивается климакс, потеря репродуктивной способности. Развитие климакса связано с нейрогуморальной перестройкой в системе гипоталамус – гипофиз – половые органы. Развивается климакс именно в тот период, когда в половых клетках начинают нарастать повреждения генома, повышается риск передачи искаженной наследственной информации. Поэтому потерю репродуктивной способности в этот период можно рассматривать как адаптивное приспособление, препятствующее вырождению потомства. У мужчин нейрогуморальная перестройка происходит более длительное время, и они сохраняют репродуктивную функцию до преклонного возраста. 9. Пожилой возраст – 61–74 года (муж.), 56–74 года (жен.). 10. Старческий – 75–90 лет. 11. Долгожители – 90 лет и выше. XIII. Онтогенез 209 Три последних возраста составляют постдефинитивный период, для которого характерны инволюционные изменения в строении организма. В этот период происходит уменьшение роста, падение веса тела, снижение функциональных показателей, изменение покровов, осанки и общее одряхление организма. На характер, течение и сроки возрастных периодов существенное влияние оказывает половая принадлежность. О глубине половых различий свидетельствует различие хромосомных наборов мужчин и женщин. Фактор пола локализован в специальных Х- и Y-хромосомах. Женщины имеют две одинаковые Х-хромосомы, а мужчины две неодинаковые ХУ-хромосомы. Эти различия генетически предопределяют биологические возможности различных полов. Предполагают, что с добавочной Х-хромосомой связана большая надежность работы генетического аппарата у женщин, их жизнеспособность, большая продолжительность жизни (табл.). Женщины менее подвержены повреждениям сердечно-сосудистой системы, лучше переносят стресс, но более подвержены гормонзависимым опухолям (матка, молочные железы). При описании основных морфологических особенностей человека в различные возрастные периоды используют средние показатели. Однако индивидуальные различия в процессах роста и развития могут варьировать в широких пределах. Это послужило основанием для введения такого понятия как биологический возраст, или возраст развития. Биологический возраст может быть равен паспортному, а может быть и нет. Если наблюдается опережение – акселерированный возраст, если наблюдается замедление – ретардированный возраст. Основными критериями биологического возраста считаются: 1. Зрелость, определяемая по степени развития вторичных половых признаков – может быть использован только в период полового созревания. Имеются половые формулы для девочек и для мальчиков. Половая формула для девочек : Ma (развитие грудных желез) – 0-4б, Ax (подмышечное оволошение) – 0-4б, Р (лобковое оволошение) – 0-4б, Ме (первая менструация) – +или –. Половое формула для мальчиков – Ax – 0-4б, Р – 0-4б, С (набухание сосков) – 0-3б, G(увеличение гениталий, яичек и полового члена) – 0-12б, Ро (первые эякуляции(семяизвержения) 210 Болотов А. В. Биология размножения и развития или поллюции) + или –, L( ломка голоса) – +или –. Опережение и запаздывание на 2–3 года требует медицинского контроля. У девочек наблюдаются жироотложения, а у мальчиков образования мышц. Таблица Продолжительность жизни мужчин и женщин в разных странах по данным ВОЗ, 2009 г. Место Страна 1 2 3 3 3 12 12 12 12 12 12 22 22 22 22 28 28 28 28 34 34 46 80 104 120 120 125 192 192 193 194 Сан-Марино Япония Австралия Адорра Швейцария Испания Италия Монако Франция Исландия Сингапур Канада Новая Зеландия Норвегия Швеция Австрия Нидерланды Греция Германия Великобритания Финляндия США Китай Белорусь Украина Россия Индия Зимбабве Афганистан Замбия ЧАД Продолжительность жизни мужчины женщины общая 82 85 83,5 80 86 83 80 84 82 79 85 82 80 84 82 78 85 81,5 79 84 81,5 78 85 81,5 78 85 81,5 80 83 81,5 79 84 81,5 79 83 81 79 83 81 79 83 81 79 83 81 78 83 80,5 78 83 80,5 78 83 80,5 78 83 80,5 78 82 80 77 83 80 76 81 78,5 72 76 74 64 76 70 62 74 68 62 74 68 63 66 64,5 47 50 48,5 47 50 48,5 46 50 48 47 48 47,5 XIII. Онтогенез 211 2. Скелетная зрелость служит хорошим показателем биологического возраста для всех периодов. Костный возраст определяется по стадиям оссификации скелета. Учитывается время и последовательность появление точек окостенения, а также сроки наступления синостозов. Для определения делают ренген кисти, используют метод уайтхауса (берется 20 косточек, по степени замещения хряща костной тканью, оценивается по баллам замещения, а потом по целой величине находится биологический возраст). Шов черепа между клиновидной и затылочной костью – если полностью зарос, то есть 18 лет. После 18 (у девушек) и 22 (у парней) до 35 лет костная система находится в стабильном состоянии, поэтому только после 35 лет начинают снова определять возраст по скелету (рентген кисти, плотность и т. д., а также наличие костных шипиков). Самый информативный, самый объективный и адекватный критерий. 3. Зубная зрелость определяется путем подсчета числа прорезавшихся зубов и сопоставление его с существующими стандартами (в качестве показателя может использоваться только до 13–14 лет). Учитываются сроки прорезывания молочных и постоянных зубов, стёртость зубов определяется по прорезыванию зубов, поэтому будет работать в определенных возрастах: в период прорезывания молочных (6 месяцев – 2 года), самые первые прорезываются нижние резцы, потом два верхних и т. д. I (резцы) – 2 (нижний), C (клыки) – 1, М (моляры) – 2. В молочной смене 20 зубов. Постоянные зубы (в 5–6 лет до 13–14 лет) – I–2, С- 1, Р- 2, М- 2+1. 4. Показатели зрелости отдельных физиологических систем организма на основании возрастных изменений микроструктур различных органов или по биоэлектрической активности клеток. Например, такие показатели: величина и плотность расположения нейронов, длина аксонов, степень миелинизации, синаптогенез, возможно, стадиоспецифичные антигенные факторы, активность ацетилхолинэстеразы, содержание ДНК, содержание пигмента липофусцин в перикарионах нейронов, сенильные бляшки в нейропиле, атрофия дендритного древа, исчезновение дендритных шипиков, уменьшение размеров и потеря нейронов. 5. Морфологическая и психологическая зрелость. Морфологическая зрелость оценивается на основании развития опорно- 212 Болотов А. В. Биология размножения и развития двигательного аппарата ─ мышечной силы, статической выносливости, частоты и координации движений. С морфологической и физиологической зрелостью тесно связана школьная зрелость, под которой подразумевают степень психофизиологической и морфологической зрелости, достаточную для начала школьного обучения. Оценка морфологической зрелости основана на изменении пропорций тела, происходящем от того, что замедляется рост головы и шеи, но ускоряется рост конечностей. Психологическую зрелость сложно определять, определяют обычно возраст детей. Определение биологического возраста в старших возрастах после 35 лет осуществляется с использованием комплексных психофизиологических показателей с применением компьютерных технологий. 3. Основные закономерности роста и развития 1. Эндогенность. Рост и развитие организма совершаются по внутренним, присущим самому организму и запечатленным в наследственной программе законам. Рост необходим для реализации естественной потребности организма в достижении взрослого состояния, когда делается возможным продолжение рода. Внешние факторы лишь сказываются на темпах роста и развития. 2. Необратимость. Человек не может вернуться к тем особенностям строения, которые были у него в детстве или младенчестве. 3. Цикличность. Существуют периоды активации и торможения роста: а) от рождения до года; 6) 6–7 лет (полуростовый скачок); в) 11–14 лет (ростовый, или пубертатный скачок роста). Были обнаружены периоды вытяжения (активация роста) и округления (задержка роста с увеличением массы). Увеличение длины тела происходит в основном в летние месяцы, нарастание веса – осенью. 4. Постепенность. Человек в своем развитии проходит ряд этапов, совершающихся последовательно один за другим. Пропустить какой-либо из этих этапов при нормальном развитии организм не может. 5. Синхронность. Процессы роста и старения совершаются относительно одновременно в разных органах и системах тела. Правило синхронности нарушается при ускорении роста и старения. Вопросы для аттестации по предмету 1. Предмет и задачи биологии индивидуального развития. Краткая история развития и современные направления. Теория преформизма и эпигенеза. 2. Половое и бесполое размножение. Детерминация пола, основы. 3. Морфологические особенности органов размножения у животных и человека. Особенности половых клеток. 4. Основные этапы эмбрионального развития. 5. Гаметогенез. Сперматогенез и Оогенез: сходство и различие процессов. 6. Профаза мейоза. Блок мейоза в оогенезе. 7. Спермиогенез, стадии. 8. Сперматозоид, строение и функция. Капацитация. Эякулят. Количество и состав спермы. 9. Нарушения сперматогенеза. Факторы, влияющие на сперматогенез. 10. Половое созревание животных. Строение и функция половых органов самцов. 11. Строение и функция половых органов самок. 12. Овуляция. Овуляторный и ановуляторный цикл у женщин. Эволюция гаметогенеза. 13. Регуляция полового цикла у женщин. Развитие желтого тела. 14. Образование фолликулов и виды фолликулов. 15. Яйцеклетка, строение. Оболочки яйцеклетки: первичная, вторичная и третичная. Понятие о полярности яйцеклетки: анимальновегетативная ось клетки. Классификация яйцеклеток по количеству и характеру распределения желтка. 16. Осеменение, типы. Виды оплодотворения. 17. Оплодотворение, его фазы. Факторы, способствующие и препятствующие оплодотворению. 18. Дробление. Типы дробления. Зависимость типа дробления от вида яйцеклеток. Скорость и темпы дробления. 19. Радиальное голобластическое дробление. 20. Спиральное голобластическое дробление. 21. Билатеральное голобластическое дробление. 22. Асинхронное голобластическое дробление. Особенности дробления у млекопитающих. 23. Поверхностное меробластическое дробление. 24. Дискоидальное меробластическое дробление. 25. Бластодерма и бластоцель. Эмбриобласт, трофобласт. 26. Гаструляция, сущность процесса. Основные способы гаструляции. 27. Инвагинационная гаструляция. 28. Эпиболический тип гаструляции. 214 Болотов А. В. Биология размножения и развития 29. Деляминационный тип гаструляции. 30. Иммиграционный тип гаструляции. 31. Особенности гаструляции у млекопитающих. 32. Образование мезодермы. Телобластический и Энтероцельный пути. 33. Внезародышевые органы хордовых, их функции. 34. Гистологические типы плацент: эпителиохориальная, десмохориальная, вазохориальная, гемохориальная. 35. Дифференцировка, рост, морфогенез. 36. Управление процессом развития генами. 37. Дифференцировка эктодермы. Дифференцировка энтодермы. Дифференцировка мезодермы. 38. Развитие хорды. Развитие нервной системы. 39. Развитие зуба, пера, волоса. 40. Развитие сомитов. 41. Развитие спланхнотома. 42. Развитие пищеварительной системы. 43. Развитие легких. 44. Развитие печени и желчного пузыря. 45. Развитие органов обоняния. 46. Развитие органа слуха и вестибулярного анализатора. 47. Развитие осевого скелета. 48. Развитие сердца. 49. Развитие выделительной системы. 50. Онтогенез. Онтогенез человека. Периоды пренатального онтогенеза. 51. Начальный период развития у человека и его особенности. 52. Зародышевый период развития у человека и его особенности. 53. Плодный период. Особенности периода. 54. Основные периоды постнатального онтогенеза у человека. 55. Период новорождености, его значение. 56. Особенности грудного периода развития у человека. 57. Особенности периода раннего детства. 58. Особенности периода первого детства у человека. 59. Особенности развития человека в период второго детства. 60. Особенности подросткового периода у человека. Особенности развития мальчиков и девочек. 61. Особенности юношеского периода в развитии человека. 62. Характеристики зрелого возраста у человека. 63. Пожилой и старческий периоды, долгожительство. 64. Основные закономерности роста и развития. Список использованной литературы 1. Алмазов И. В. Атлас по гистологии и эмбриологии / И. В. Алмазов, Л. С. Сутулов. – М. : Медицина. – 1978. – 543 с. 2. World Health Organization. WHO laboratory manual for the examination and processing of human semen. – 5th ed. – World Health Organization, 2010. – 271 p. 3. Безруких М. М. Возрастная физиология: (Физиология развития ребенка) : учеб. пособие для студ. вузов / М. М. Безруких, В. Д. Сонькин, Д. А. Фарбер. – М. : Академия, 2003. – 415 с. 4. Белоусов Л. В. Основы общей эмбриологии : учебник / Л. В. Белоусов. – 3-е изд. переработ., и доп. – М. : Изд-во Моск. ун-та ; Наука, 2005. – 368 с. 5. Воробьева О. А. Влияние морфологии сперматозоидов на частоту оплодотворения и нарушения развития эмбрионов в программе ЭКО / О. А. Воробьева, О. А. Леонтьева, B. C. Корсак // Пробл. репрод. – 1998. – № 1. – С. 14–18. 6. Всемирная Организация Здравоохранения (ВОЗ). Руководство по лабораторному исследованию спермы человека и взаимодействию спермы с цервикальной слизью : пер. с англ. – Тбилиси, 1988. 7. Гилберт С. Биология развития : в 3 т. : пер. с англ. / С. Гилберт. – М. : Мир, 1993. – Т. 1. – 228 с. 8. Гилберт С. Биология развития : в 3 т. : пер. с англ. / С. Гилберт. – М. : Мир, 1994. – Т. 2. – 235 с. 9. Гилберт С. Биология развития : в 3 т. : пер. с англ. / С. Гилберт. – М. : Мир, 1995. – Т. 3. – 352 с. 10. Гистология, цитология и эмбриология : учебник / Ю. И. Афанасьев [и др.] ; под ред. Ю. И. Афанасьева, С. Л. Кузнецова, Н. А. Юриной. – 6-е изд., перераб. и доп. – М. : Медицина, 2004. – 768 с. 11. Голиченков В. А. Эмбриология / В. А. Голиченков, Е. А. Иванов, Е. Н. Никерясова. – М. : Академия, 2004. – 224 с. 12. Голиченков В. А. Практикум по эмбриологии / В. А. Голиченков, М. Л. Семенова. – М. : Academia, 2004. – 208 с. 13. Джелдубаева Э. Р. Биология индивидуального развития : курс лекций / Э. Р. Джелдубаева. – Семфирополь, 2008. – 210 с. 14. Дондуа А. К. Биология развития : в 2 т. / A. K. Дондуа. – СПб. : Изд-во СПбГУ, 2004. – Т. 1. – 244 с. 15. Дондуа А. К. Биология развития : в 2 т. / A. K. Дондуа. – СПб. : Изд-во СПбГУ, 2005. – Т. 2. – 188 с. 16. Корочкин Л. И. Биология индивидуального развития (генетический аспект) : учебник / Л. И. Корочкин. – М. : Изд-во МГУ, 2002. – 263 с. 216 Болотов А. В. Биология размножения и развития 17. Кузнецов С. Л. Лекции по гистологии, цитологии и эмбриологии : учеб. пособие / С. Л. Кузнецов, М. К. Пугачев. – М. : Мед. информ. агентство, 2004. – 432 с. 18. Курило Л. Ф. Руководство по проведению исследования и оценке эякулята человека. ВОЗ, пятое издание, 2010: что нового? / Л. Ф. Курило, Н. П. Макарова // Андрология и генитальная хирургия. – 2010. – № 4. – С. 10–13. 19. Лабораторные занятия по курсу гистологии, цитологии и эмбриологии : учеб. пособие / Ю. И. Афанасьев [и др.] ; под ред. Ю. И. Афранасьева, А. Н. Яцковского. – М. : Медицина, 2004. – 328 с. 20. Леонтьева О. А. Сравнительный анализ морфологии сперматозоидов человека: нативный эякулят – прогрессивно подвижная фракция / О. А. Воробьева, О. А. Леонтьева // Пробл. Репрод. – 1999. – № 3. – С. 29-36. 21. Медицинская биология : учебник / под ред. В. П. Пишака, Ю. И. Бажоры. – Винница : Нова Книга, 2004. – 656 с. 22. Практикум по эмбриологии : учеб. пособие / В. А. Голиченков [и др.] ; под ред. В. А. Голиченкова, М. Л. Семеновой. – М. : Академия, 2004. – 208 с. 23. Реунов А.А. Сперматогенез многоклеточных животных / А. А. Реунов ; отв. ред. В. Л. Касьянов ; Ин-т биологии моря. – М.: Наука, 2005. – 123 с. 24. Ролдугина Н. П. Практикум по цитологии, гистологии и эмбриологии : учеб. пособие / Н. П. Ролдугина, В. Е. Никитченко, В. В. Яглов. – М. : КолосС, 2004. – 215 с. 25. Рост и развитие ребенка / В. В. Юрьев, А. С. Симаходский, Н. Н. Воронович, М. М. Хомич. – 2-е изд., перераб. и доп. – СПб. : Питер, 2003. – 260 с. 26. Руководство ВОЗ по лабораторному исследованию эякулята человека и взаимодействия сперматозоидов с цервикальной слизью : пер. с англ. – 4-е изд. – М. : МедПресс, 2001. – 144 с. 27. Станек И. Эмбриология человека / И. Станек. – Братислава : Веда, 1997. – 440 с. 28. Тегако Л. Антропология : учеб. пособие / Л. Тегако, Е. Клементинский. – М. : Новое знание, 2004. – 400 с. 29. Токин Б. П. Общая эмбриология : учебник / Б. П. Токин. – М., 1987. – 480 с. Рекомендуемая литература Основная 1. Гистология, цитология и эмбриология : учебник / Ю.И.Афанасьев [и др.] ; под ред. Ю. И. Афанасьева, С. Л. Кузнецова, Н. А. Юриной. – 6-е изд., перераб. и доп. – М. : Медицина, 2004. – 768 с. 2. Голиченков В. А. Эмбриология / В. А. Голиченков, Е. А. Иванов, Е. Н. Никерясова. – М. : Академия, 2004. – 224 с. 3. Корочкин Л. И. Биология индивидуального развития (генетический аспект) : учебник / Л. И. Корочкин. – М. : Изд-во МГУ, 2002. – 263 с. Дополнительная 1. Алмазов И. В. Атлас по гистологии и эмбриологии / И. В. Алмазов, Л. С. Сутулов. – М. : Медицина. – 1978. – 543 с. 2. Безруких М. М. Возрастная физиология: (Физиология развития ребенка) : учеб. пособие для студ. вузов / М. М. Безруких, В. Д. Сонькин, Д. А. Фарбер. – М. : Академия, 2003. – 415 с. 3. Белоусов Л. В. Основы общей эмбриологии : учебник / Л. В. Белоусов. – 3-е изд. переработ., и доп. – М. : Изд-во Моск. ун-та ; Наука, 2005. – 368 с. 4. Биология : учебник. В 2 кн. / В. Н. Ярыгин [и др.] ; ред. В. Н. Ярыгин. – 7-е изд., стер. – М. : Высш. шк., 2005. – Кн.1. – 432 с. 5. Быков В. Л. Цитология и общая гистология: функциональная морфология клеток и тканей человека / В. Л. Быков. – СПб. : Сотис, 1999. 520 с. 6. Гилберт С. Биология развития : в 3 т. : пер. с англ. / С. Гилберт. – М. : Мир, 1993. – Т. 1. – 228 с. 7. Гилберт С. Биология развития : в 3 т. : пер. с англ. / С. Гилберт. – М. : Мир, 1994. – Т. 2. – 235 с. 8. Гилберт С. Биология развития : в 3 т. : пер. с англ. / С. Гилберт. – М. : Мир, 1995. – Т. 3. – 352 с. 9. Кузнецов С. Л. Лекции по гистологии, цитологии и эмбриологии : учеб. пособие / С. Л. Кузнецов, М. К. Пугачев. – М. : Мед. информ. агентство, 2004. – 432 с. 10. Лабораторные занятия по курсу гистологии, цитологии и эмбриологии : учеб. пособие / Ю. И. Афанасьев [и др.] ; под ред. Ю. И. Афранасьева, А. Н. Яцковского. – М. : Медицина, 2004. – 328 с. 11. Молдавская А. А. Васкуляризация производных пищеварительной трубки на этапах онтогенеза [Электронный ресурс] / А. А. Молдавская. – М. : Изд-во Академия Естествознания, 2007. – Режим доступа: http://www.rae.ru/ru/publishing/mono06.html 218 Болотов А. В. Биология размножения и развития 12. Практикум по эмбриологии : учеб. пособие / В. А. Голиченков [и др.] ; под ред. В. А. Голиченкова, М. Л. Семеновой. – М. : Академия, 2004. – 208 с. 13. Ролдугина Н. П. Практикум по цитологии, гистологии и эмбриологии / Н. П. Ролдугина, В. Е. Никитченко, В. В. Яглов. – М. : КолосС, 2004. – 216 с. 14. Тейлор Д.. Биология. В 3 т. / Д. Тейлор, Н. Грин, У. Стаут ; ред. Р. Сопер ; пер. с англ. Ю. Л. Амченкова, И. В. Еланской, Н. О. Фоминой. – 2002. – Т. 3. – 452 с. 15. Юшканцева С. И. Гистология, цитология и эмбриология. Краткий атлас : учеб. пособие [Электронный ресурс] / С. И. Юшканцева, В. Л. Быков. – СПб : Изд-во П-2, 2006. – URL: Режим доступа: http://www.medliter.ru/?page=get&id=013616 Учебное издание Болотов Андрей Валентинович БИОЛОГИЯ РАЗМНОЖЕНИЯ И РАЗВИТИЯ Раздел. БИОЛОГИЯ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ Редактор Э. А. Невзорова Дизайн обложки: А. В. Болотов Темплан 2011. Поз. 105 Подписано в печать 28.12.11. Формат 60х90 1/16. Уч.-изд. л. 7,8. Усл. печ. л. 12,7. Тираж 50 экз. Заказ 150. ИЗДАТЕЛЬСТВО ИГУ 664003, Иркутск, бульвар Гагарина, 36