УЧ ЕБН А Я Л И Т ЕРА Т У РА
Д ля студентов медицинских институтов
Биология
ПОД Р ЕД А К Ц И ЕЙ
В. Н. Я Р Ы Г И Н А
Допущено Главным управлением учебных
заведений Министерства здравоохранения
С С С Р в качестве учебника для студентов
медицинских институтов
М осква. «Медицина». 1984
Б Б К 28.0
Б 63
У Д К 57(075.8)
\
Ю. К . Богоявленский, Т. Н. Улиссова, И. М. Яровая, В. Н. Ярыгин
Биология / Под ред. В. Н. Ярыгина. — М .: Медицина, 1984, 560 с., ил.
В . Н. Я р ы г и н
рогова.
— проф., зав. кафедрой биологии II М О Л Г М И им. Н. И. П
В учебнике освещены основные разделы биологии и ее задачи в системе медг
пинского образования — общебиологические закономерности жизнедеятельност
человека, человек и биосфера, биологические аспекты .экологии человека, био­
тические основы паразитизма. Материал учебника изложен с учетом достиж
биологической науки, показана взаимосвязь отдельных ее областей.
Учебник соответствует программе, утвержденной Министерством здравоохран
ния С С С Р , и предназначен для студентов медицинских институтов.
Рисунков 232. таблиц 27.
Р е ц е н з е н т ы : Г. В. Хомулло — проф., зав. кафедрой биологии КалининскогЪ
медицинского института; И. С. Б а ж е н о в
стовского медицинского института.
проф., зав. кафедрой биологии Р о ­
,* •
.' о г
чгвЧ
Юрий Константинович Богоявленский, Татьяна Николаевна Улиссова,
Ирина М оисеевна Яровая, Владимир Никитич Ярыгин
БИ О Л О ГИ Я
З а в . редакцией С. Д. Крылов. Р е д ак то р И. В. Войтехсгва
Х уд о ж естве н н ы й р едактор Н. И. Синякова -Оформление х уд о ж н и ка В. С. Сергеевой
Техн и ческий редактор С. П. Танцева- Ко р р екто р //. С. Филиппова
И Б № 8571
Сдано в набор ОС 01.84. П о дп исано к п ечати 30.08.84. Т-02653. Ф о р м а т б ум аги 6 0 х 9 0 ’ /|в- Б у м а г а офс.
Г а р н и т у р а « Т а й м с» . П е ч а т ь о ф сетн ая. Уел . неч. л. 37,0. Уел. кр.-отт. 40,0. Уч.-изо л 42.15
Т и р а ж 1501
(1-ый завод 1— 50 ООО) экз. З а к а з /*> 56. Ц е н а I р. 80 к
О рдена Трудового К р асн о го Зн ам ен и и зд ател ь ств Ь «М ед и ц и н а» ,103062 /М осква, Петр овер и гски й
Я р о с л а вск и й иол игр аф ко м б и нат СоюзполТТГ’РТфпрома*' при Го суд а р ствен н о м к о м и т е т е . С
И зд ательств, полиграф ии и кни ж но й тор го вли. 150014, Я р о с л а в л ь , у л . С вободы , 97.
Учебная библиот“ « *
2001000000— 246
Б — ---------------- 16— 84
0 3 9 (0 1 )-8 4
Рлз. кед. ии-та
1
\
\
\
© Издательство «Медицина», Москва, 19
П РЕД И С Л О ВИ Е
Биологии в естественнонаучной и мировоззренческой подготовке
..рача принадлежит ведущая роль. Будучи фундаментальной дисципли­
ной, она раскрывает закономерности развития жизни. Многие частные
вопросы биологии имеют прикладной характер и учитываю тся при
решении актуальных проблем профилактической и лечебной медицины,
большое внимание в учебнике уделено объекту врачебной деятельно­
сти — человеку. Человек является закономерным результатом процесса
>>{*горического развития органического мира и одновременно неЪ(*^емлемой частью природы планеты. Поэтому в учебнике важное
1)1Ссто занимает изложение общебиологических предпосылок его
жизнедеятельности, обсуждается роль человечества в биосфере.
Широко представлен также материал, освещающий проблемы генети­
ки, онтогенеза, популяционной генетики, экологии.
Авторы стремились отразить современное состояние тех отраслей
биологии, идеи и факты которых привлечены ими при написании
учебника. Они испытываю т чувство глубокой благодарности к колле­
гам, труды котопых б ти использованы в процессе работы.
Современная (
,
|К .
эояще~ ч здание. Каждый год увеличи­
вается число В1 . ..
'
шчные аспекты важнейших теоретиче­
ских положений остаютс. „ .мметом дискуссий специалистов. Многие
фундаментальные концепции, остававшиеся незыблемыми на протяже­
нии десятилетий, пересматриваются. В этих условиях авторам нередко
приходилось делать выбор между различными точками зрения.
При подготовке настоящего учебника использован опыт преподава­
ния биологии на кафедрах II Московского медицинского института
им. Н . И. Пирогова,
1 Московского
медицинского
института
им. И . М . Сеченова, Московского медицинского стоматологического
института им. Н . А . Семашко. Дружеское участие и помощь сотрудни­
цей кафедр мы приняли с благодарностью. Искреннюю благодарность
авторы приносят заведующим кафедрами биологии Калининского и
, отовского медицинских институтов профессорам Г . В. Хомулло и
! I. С. .Баженову за полезные советы при обсуждении рукописи учебника.
Авторы признательны за все критические замечания и пожелания.
ВВЕДЕНИИ
ч
Термин « б и о л о г и я » (от греч.
жизнь, 1о§о5 — на у к г'
введен в начале X IX столетия независимо Ж . Б. Ламарком и Г . Тревиранусом для обозначения науки о жизни как особом явлении природы.
'~ТТнастоящее время его используют и в ином смысле, относя к группам
организмов, вплоть до вида (биология микроорганизмов, биология
северного оленя), биоценозам (биология арктического бассейна),
отдельным живым структурам (биология клетки). В данном курсе этот
термин употребляется прежде всего во втором своем значении
(биология человека, биология возбудителей паразитарных заболеваний
людей). Однако при рассмотрении биологических предпосылок разви
тия, жизнедеятельности и экологии человека неизбежно затрагиваются
общие вопросы сущности жизни, уровней ее организации, механизмов
сохранения жизни во времени и эволюции. Наряду с физикой, химией,
математикой биология относится к естественным наукам, предмет
изучения которых — природа. Особенность предмета биологии в меди
цинском институте заключается в том,
■
центре внимания
находится человек. Закономерности же развития человека как личности
и человеческого общества в целом изучаются социальными (обще­
ственными) науками. Указывая на то, что сущность людей социальна,
К. Маркс подчеркивал, что естествознание включит в себя науку о
человеке в такой же мере, в какой наука о человеке включит в себя есте­
ствознание. Обобщенная картина жизнедеятельности человека склады­
вается в результате изучения биологии совместно с общественными
дисциплинами.
В процессе поступательного развития и по мере обогащения новых:..'
фактами биология преобразовалась в комплекс наук, исследующих
закономерности, свойственные живым существам, с разных сторон
Некоторые из этих наук представлены в медицинских институт; *
самостоятельными дисциплинами — анатомией, физиологией, гис’ . ‘
логией, биохимией, микробиологией. Особенность современной био.
гии заключается в утверждении принципа единства главных механизме
жизнеобеспечения, осознании роли эволюционного процесса в суцц
ствовании и изменениях органического мира, который включает и
человека, признании первостепенной важности экологических зак томерностей с распространением их на человека. Э то объясняет и де
необходимым преподавание будущим врачам наряду с антрополог и
скими дисциплинами, названными выше, специального курса биол л ии.
Биология относится к ведущим отраслям естествознания. Вы .••эк»-:
уровень ее развития, как это подчеркивается в Программе К П С С , сл.
жит необходимым условием подъема и повышения эффективное.,,
медицины.
Г л а в а
I
ч » \И Т И Я ЬИ О Л О ГИ И
Т 1 А П Ы РА, ЗЛ
>
И (учение мира жнпых существ началось олновременно с образова­
нием человеческою общества. Ьиология как наука о закономерностях
жи щи прошла определенный путь, на отдельных этапах которого
возникали новые способы познания биологических явлений и меха11И З М О В .
Практические запросы людей еще на заре возникновения
человеческого общества стимулировали классифицирование живых
форм.
Гораздо
позже
возникла
идея
единства
о р I а и и ч е с к о I о м и р а. Значение ее для медицины заключается
в том, ч ю она указывает на универсальность биологических
закономерностей, распространение их на весь органический мир,
включая человека. В известном смысле история современной биологии
представляет собой цепь фундаментальных открытий и обобщений,
обосновывающих згу идею. Важным аргументом в пользу единства
все! о живого послужила к л е т о ч н а я
т е о р и я Т. Шванна и
М. Шлейдена (1839). О ткрытие клеток, доказательство их гомологии,
уяснение того, что все организмы состоят из клеток и продуктов их
жизнедеятельности, дало толчок плодотворному изучению фунда­
ментальных закономерностей строения, функционирования и развития
живых существ.
Идея единства ор| эпического мира, вытекающая из того факта,
что клетка является своего рода общим знаменателем живого, полу­
чила подкрепление в исследованиях б и о х и м и ч е с к и х
(м е ­
таболических)
основ
физиологии
клеток.
Наиболее
демонстративны
достижения
молекулярной
б и о л о г и и. Она приобрела положение самостоятельного направле­
нии биологической науки в пятидесятые годы нашего столетия.
Хронологически эго было связано с описанием Д ж . Уотсоном и
Ф . Криком (1953) макромолекулярной структуры Д Н К . Молекулярная
биология концентрирует внимание на связи процессов жизнедеятельносги с биологическими макромолекулами и прежде всего на законо­
мерностях хранения, использования и передачи в клетках наслед­
ственной информации. Молекулярно-биологические исследования от­
крыли физико-химические механизмы, которые обусловливают такие
свойства живого, как специфичность и структурированность биологиче­
ских объектов, воспроизводимость клеток и организмов в ряду
поколений, а также показали универсальность этих механизмов, их
приложимость к существам разных типов организации.
Основные з а к о н ы
наследственности
были уста­
новлены в прошлом (Г . Мендель, А. Вейсман) и начале текущего
(Г . де-Фриз, Г. Морпш) столетии. Значение указанных законов для
утверждении идеи единства ор!анического мира состоит в том, что они
вскрыли всеобщий механизм передачи от особи к особи и перераспреде­
ления в пределах вида наследственной информации. Этим были созданы
предпосылки к вскрытию биологической сущности полового размноже­
ния, индивидуального развития, смены поколений.
Клеточная теория, положения молекулярной биологии, законк
наследственности обосновывают идею единства современного органи­
ческого мира. То, что живое представляет собой единство в историчес­
ком плане, вытекает из существа т е о р и и э в о л ю ц и и , создан­
ной Ч . Дарвиным (1858), и получившей дальнейшее развитие в трудах
А . Н . Северцева, Н . И . Вавилова, Р. А . Фишера, С. С. Четверикова,
С. Райта, И . И. Шмальгаузена. Эволюционная теория объясняет един­
ство органического мира общностью происхождения. Она описывает
универсальный механизм, посредством которого возникает разнообра­
зие живых форм, приспособленных к среде обитания и отличающихся
прогрессивно повышающимся уровнем структурно-физиологической
организации. Теория эволюции обращает внимание на генетическое
родство живых форм, которое заключается в преемственности
фундаментальных механизмов развития и жизнедеятельности. Она
указывает на условность грани между живой и неживой природой.
Результаты исследований молекулярного и атомарного состава тканей
живых существ, синтез в искусственных условиях соединений,
свойственных только живому, доказали возможность в истории Земли
перехода от неживого к живому. Эволюционная теория поставила на
научную основу проблему антропогенеза и указала на единство природы
и человека. Принцип клеточной организации, молекулярные механизмы
жизнедеятельности, генетические законы распространяются на челове­
ка. Эволюционная теория обосновывает представление о биологиче­
ском наследстве людей и описывает механизмы его формирования.
История свидетельствует о том, что каждый крупный шаг на пути
познания фундаментальных законов жизни сказывался на состоянии
медицины, приводил к изменениям в оценке существа и механизмов
развития патологических процессов. Соответственно пересматривались
принципы и практика лечебной и профилактической медицины. Так,
исходя из положений клеточной теории и способствуя ее упрочению,
Р. Вирхов создал концепцию целлюлярной патологии (1858), которая на
долгое время определила главные пути развития медицины. Использо­
вав генетико-биохимический подход в изучении болезней человека, врач
А. Гаррод (1908) заложил основы молекулярной патологии, дал ключ к
пониманию таких вопросов практической медицины как различная
восприимчивость людей к болезням и вариабельность (индивиду­
альность) реакций на лекарства. Успехи общей и экспериментальной
генетики, достигнутые к концу 20-х — началу 30-х годов, стимулирова­
ли исследования по генетике человека. В результате возник новый
раздел патологии — наследственные заболевания, появилась особая
служба практического здравоохранения — медико-генетические кон­
сультации. Молекулярная биология порождает принципиально новые
подходы к лечению заболеваний и пороков развития, которые, зависят
от дефективности генов. Н а основе этих открытий возникла новая
наука — генетическая инженерия.
6
История биологии, кроме доказательств влияния биологических
открытий и теорий на состояние медицины, знакомит с мировоззренче­
скими проблемами современной науки о человеке.
1.1. М Е Т О Д О Л О Г И Ч Е С К И Е В О П Р О С Ы
БИ О Л О ГИ И
В биологии существует два основных пути познания явлений и
механизмов. Один из них заключается в расчленении сложных
биологических процессов и объектов на составляющие части и анализе
каждой составляющей части в отдельности. Указанный путь получил
название р е д у к ц и о н и з м а . Концентрация внимания на вычленя­
емых в целостном организме тканях, клетках, отдельных клеточных
органеллах, метаболических циклах, биополимерах обогатило биоло­
гию конкретными фактами, позволило подойти к пониманию субкле­
точных и физико-химических основ жизнедеятельности. Второй путь
познания — и н т е г р а т и з м — заключается в поиске обобщенных
характеристик биологического процесса или объекта с выделением черт,
свойственных ему как целому. В названных путях познания отражена
важная мировоззренческая проблема соотношения целого и его частей.
На отдельных этапах развития биологии удельный вес в биологических
исследованиях редукционизма и интегратизма менялся. Переход от
простого наблюдения к широкому экспериментированию способствовал
укреплению редукционизма. П о мере накопления фактов возникла
потребность в обобщениях. Примером этому служит клеточная теория.
Как обобщение, описывающее отношение клетки к организму и живой
природе в целом, она была сформулирована в 1839 г., тогда как первое
профессионально выполненное описание клетки сделано Р. Гуком еще в
1665 г.
История биологии содержит многие доказательства тому, что для
познания процессов жизнедеятельности и развития организма необхо­
димо единство анализа и синтеза, редукционизма и интегратизма в
соответствии с диалектико-материалистическим решением проблемы
части и целого.
Познание биологических закономерностей происходит как путем
изучения проявлений жизни конкретными науками, так и путем
разработки общих теорий. Такие теории, как правило, отражают
господство в тот или иной исторический период определенной
философской системы. На протяжении своей истории биология не­
изменно была ареной борьбы идеалистического и материалистического
мировоззрений. Принципиальное разногласие между идеализмом и ма­
териализмом заключается в понимании отношения между материей и
сознанием. Идеализм утверждает первичность сознания, некоего
духовного начала и подчинения ему материального. Материализм
утверждает первичность материального мира, рассматривая сознание
как свойство высокоорганизованной материи.
В биологии идеализм представлен в и т а л и з м о м
(от
лат. — У15, УЙаНз — жизненная сила). Витализм признает наличие
особой, непознаваемой опытным путем духовной сущности, от которой
зависит исключительность свойств жизни. Распространению витализма
7
способствовало то, что долгое время науке не были известны движущие
силы эволюции, которая создает особей, удивительно приспособленных
к среде обитания, и органы, в деталях отвечающие выполняемым
функциям. Витализм был подвергнут критике французскими материа­
листами X V III века. Развивая взгляды ученого и мыслителя первой
половины X V II века Р . Декарта, врач и философ Ж . О. Ламетри создал
учение о «человеке-машине». Он писал: «Если все может бы ть
объяснено тем, что открывает в мозгу анатомия и физиология, к чему
мне создавать фиктивную субстанцию? Если я отождествляю душу с
органами тела, то потому, что все меня убеждает в этом»1.
Взгляды французских материалистов X V III века формировались под
влиянием успехов точных наук и, прежде всего, механики. Идея
развития органического мира была им, как правило, чужда. Э то
объясняет их определенную ограниченность и непоследовательность.
Философские взгляды материалистов X V III—X I X веков в биоло­
гии представлены м е х а н и ц и з м о м . Ём у свойственно отрицание
качественного своеобразия живого. Биологические закономерности он
сводит к физическим явлениям и химическим превращениям. Некоторые
представители механицизма . все разнообразные проявления жизни
объясняли наличием единого Универсального биологического принципа.
философской основой современной биологии служит д и а л е к ­
тический
м а т е р и а л и з м — ученйе, созданное К . Марк­
сом и Ф . Энгельсом во второй половине X IX века. Диалектиче­
ский Материализм глубоко и последовательно отражает всеобщие
законы материального Мира, вскрывает своеобразие и закономерности
переходов различных форм движения материи. Фундамент диалектиче­
ского материализма составляет идея развития. Ф . ЭнгельЬ подчеркивал
решающее значение для утверждения Принципов диалектического
материализма великих открытий в области естественных наук: закона
сохранения материи и энергии, клеточной теории, эволюционной
теории Ч . Дарвина.
Конкретные философские системы, возникающие в то т или иной
исторический период, оказавшись несостоятельными, в последующем
не повторяются. Э то относится и к теоретическим построениям
виталистов и механицистов X V II— X IX веков. Остается, однако,
опасность возрождения этих взглядов в иных формах. Особенно это
относится к механицизму. Во-первых, механистический материализм
создает видимость простого и рационального объяснения биологиче­
ских явлений. При этом он порождает обманчивую перспективу
проникнуть в самое их существо. Во-вторых, дифференциация
естественных наук, широкое и плодотворное использование в совре­
менных биологических исследованиях редукционистского пути познания
способствует появлению выводов, в которых характеристики сложных
процессов сводятся к свойствам простых элементов. Успехи молеку­
лярной биологии, например, обострили проблему отношения высшей
(биологической) и низших, (химической, физической) форм движения
1 Цит. по Бляхеру Л . Я- Курс общей биологии. 3-е изд., М .— Л .: Медгиз. 1938,
с. 13.
8
материи, в неправильной оценке которых и заключается ошибка
механицистов. Диалектический материализм отрицает понимание
отношения высших и низших форм как главного и побочного. Физико­
химические процессы нельзя рассматривать как побочное содержание
жизнедеятельности организмов. Они являю тся именно фундаменталь­
ными процессами, а биологические — производными, возникшими на
их основе и отличающиеся качественным своеобразием. Характеризуя
соотношение химии и биологии, Ф . Энгельс писал, что «... химия
подводит к органической жизни, и она продвинулась достаточно далеко
вперед, чтобы гарантировать нам, что она одна объяснит нам
диалектический переход к организму»'. В дополнение к этому он
отмечает, что «... форма движения в органическом теле отличается от
механической, физической, химической, содержа их в себе в снятом
виде»2. Таким образом, Ф . Энгельс подчеркивает не факт сосуществова
ния или соподчинения, а своеобразие связи низших форм движения с
высшей, которая возникла в результате их развития и на их основе. Для
нашей планеты выделяют три основных этапа развития материи — не­
органический, биологический и социальный, каждый из которых
начинается с новой формы движения материи. С появлением высшей
формы движения отмечается ускорение развития материи. При этом
низшие формы движения сохраняются, но перестают играть ведущую
роль в дальнейшем прогрессе. Так обстояло дело в переходные моменты
истории земли, каковыми
были
появление жизни и
чело­
века.
Мировоззренческие вопросы, рассмотренные выше, касаются
отношения биологии и наук о неживой природе. Еще большее значение
в познании человека имеет методологическая проблема отношения
биологии и общественных наук. Некоторые биологи и социологи
пытаются распространить действие биологических закономерностей на
социальную сферу жизни современных людей, вывести из этих
закономерностей настоящее и будущее человека. Теории такого рода
составляют содержание антропосоциологии и социал-дарвинизма. Они
сводят проблему развития общества к конкуренции расовых типов,
выделяют избранные расы, в частности европейские, считая их высшим
продуктом естественного отбора. Антропосоциология и социалдарвинизм реакционны по своей сути и служат идеологической основой
расизма и фашизма. В своих псевдонаучных построениях авторы
указанных теорий прибегают к аналогиям из области дарвинизма. При
этом они игнорируют ограничения в отношении распространения
закономерностей низшей формы движения материи (биологической) на
высшую форму (социальную). Протестуя против необоснованного
расширения сферы действия биологических законов, К . А . Тимирязев
писал: «Останавливаясь на отношении дарвинизма к человеку, мы
должны различать две точки зрения: отношение к его прошлому и к его
настоящему. Дарвин никогда не предлагал своего учения в качестве
кодекса для поведения человека в его настоящем; оно должно было
1 М аркс К .. Энгельс Ф . Соч. 2-е изд., т. 20, с. 564.
2 М аркс К .. Энгельс Ф . Соч. 2-е изд., т. 20, с. 597.
9
служить только ключом для объяснения его темного прошлого»’.
Биологизаторские тенденции в оценке природы человека несколько
оживились в настоящее время. Э то связано с развитием молекулярной
биологии и генетики, которые открыли перед медициной перспективу
генетической инженерии, возможности целенаправленно и зм ен яв
содержание наследственной информации.
Лидирующей отраслью современной биологии, наряду с молеку­
лярной биологией, является экология. Экологические исследования
имеют выраженную практическую направленность. Поэтому их пер­
спективность для решения вопросов медицинской паразитологии и эпи­
демиологии была оценена быстро. П о результатам систематического
исследования биологии и экологии червей — паразитов человека —
ипреждевсего их жизненных циклов К . И . Скрябин поставил на повестку
дня задачу девастации (полного уничтожения) гельминтов. Экологиче­
ской по своей сути является теория природной очаговости заболеваний,
разработанная Е. П . Павловским, которая указывает пути высоко­
эффективных профилактических мероприятий. Лидирующее положение
экологии зависит однако не от этого важного, но частного момента, а
от широкого проникновения экологических представлений в научное
познание мира в целом. Особая роль в развитии этих представлений
принадлежит В . И. Вернадскому, который создал современное учение о
биосфере, и В . Н . Сукачеву, сформулировавшему понятие о биогеоцено­
зах как элементарных единицах биологического круговорота.
В условиях научно-технической революции традиционный взгляд на
место в природе человека пересматривается. Причиной этому является
переход биосферы планеты в ноосферу. Э то состояние требует
осознанного регулирования взаимоотношений между развивающимся
обществом и окружающей средой. Н а повестку дня поставлен вопрос о
том, чтобы не только биосфера включалась в общественное производ­
ство, что практикуется человечеством на протяжении всей его истории,
но и производство соизмерялось с законами развития биосферы.
Экологические
проблемы
приобрели глобальный характер, что
делает неизбежным процесс экологизации науки. Экологические пред­
ставления становятся доминирующими и в медицине.
Знакомство с историей биологии как особой отрасли естествознания
свидетельствует о той большой роли, которую биологические знания
играют в прогрессе медицины. Э та роль обусловливается наличием в
человеке б и о л о г и ч е с к о г о
наследства
в виде фунда­
ментальных физико-химических, клеточных и системных механизмов
жизнедеятельности и развития. Она вытекает из факта включенности
человека в многообразные экологические связи. Столь же очевидно
значение биологических знаний в формировании мировоззрения врача.
Они обосновывают идею единства природы и человека, раскрывают на
конкретных материалах соотношение низших и высшей форм
движения материи. Э ти знания показывают возможности редукцио­
низма и интегратизма как ведущих путей познания живых объектов.
Биологические знания являются важным элементом того фундамента,
на котором строится обобщенное представление о человеке.
1 Тимирязев К . А. Исторический метод в биологии.
М., 1922, с. 159.
Р а з д е л
I
Б И О Л О Г И Ч Е С К И Е ЗАКО НО МЕРНО СТИ
Ж ИЗНЕДЕЯТЕЛЬНОСТИ ЧЕЛОВЕКА
Глава
2
О Б Щ А Я Х А Р А К Т Е Р И С Т И К А Ж И В О ГО
2.1. С В О Й С Т В А И О П Р Е Д Е Л Е Н И Е Ж И З Н И
Ж и з н ь в ее конкретных проявлениях отличается большим
многообразием. Можно однако выделить совокупность свойств,
которые присуши всем живым существам и отличают их от тел
неживой природы. Для живых объектов1 характерна особая Форма
взаимодействия с окружающей средой — о б м е н в е щ е с т в ! Основу его составляю т взаимосвязанные и ^^лансированные тппиеггы
а с с и м и п я п я и (анаболизм) и д и с с и м и л я ц и и
(катабоТшзм). Э ти процессы направлены на обновление структур организма, а
также на обеспечение различных сторон его жизнедеятельности
необходимыми питательными веществами и энергией. Обязательным
обмена веществ служит поступление извне определенных
IX соединений, т. е. существование организма как открытой
процессы ассимиляции и диссимиляции представлены объединенными в м е т а б о л и ч е с к и е
каскады
и
циклы
хим и ч ескими
реакциями,
которые
строго
у п о р я д о ч е н ы в о в р е м е н и и п р о с т р а н с т в е . Пока­
зательны расчеты для микоплазм — микроорганизмов, занимающих по
размерам промежуточное положение между вирусами и типичными
бактериями. Наиболее мелкие из них превосходят по диаметру атом
водорода всего в 1000 раз, но их метаболический аппарат включает при­
мерно 100 реакций (в клетке человека их более 10 000). Согласованное
протекание большого количества реакций в малом объеме достигается
путем закономерного распределения отдельных звеньев обмена веществ
в протоплазме — с т р у к т у р и р о в а н н о с т и , что является обя­
зательным свойством живых объектов.
Для выяснения связей между структурированностью, обменом
веществ и существованием живых форм как открытых систем следует
обратиться к понятию э н т р о п и и .
В соответствии с первым началом термодинамики при химических и
физических превращениях энергия не исчезает и не образуется вновь, а
1 Под живым объектом (формой) понимается дискретная единица, наделенная
совокупностью свойств, отличающих живое от неживого, — организм.
2 Система (от греч. 5 уз(ета — целое, составленное из частей, соединение) —
множество элементов, находящихся в закономерных отношениях и связях друг с дру­
гом и образующих единство, целостность. Основные биологические категории (клет­
ка, организм, популяция, биогеоценоз) отличаются системной организацией.
II
переходит из одной формы в друз ую ( закон сохранения гнергии).
Поэтому теоретически каждый процесс мо1 бы протекать одинаково
легко в прямом и в обратном направлениях. В природе же разнообраз­
ные процессы самопроизвольно, без воздействий со стороны окружаю­
щей среды (извне) идут всегда в одном направлении: теплота переходи I
от более теплого объекта к мслее теплому, частицы в раст воре переме­
щаются из зоны высокой концентрации в зону низкой концентрации и
т .д . Мерой необратимости природных процессов служит энтропия.
Закономерности изменения энтропии описываются вторым началом
термодинамики, согласно которому в энергетически изолированной
системе, если в ней происходят неравновесные процессы, количество
энтропии изменяется в одну сторону, становясь максимальным но
достижении состояния равновесия. Статистическое ист олкование вто­
рого начала термодинамики (принцип Больцмана) говорит, что
равновесное состояние является наиболее вероятным в естественных
условиях и характеризуется наименьшей упорядоченностью элементов
системы. Поэтому энтропию рассматривают и как меру упорядоченно­
сти (структурированности) природных систем. Живой организм сохра­
няет структурированность на протяжении жизни, противостоя все­
общей тенденции к возрастанию энтропии, что становится возможным
благодаря постоянному притоку энергии извне, которая поддерживает
состояние равновесия. Живой организм в отличие от тел неживой приро­
ды представляет собой энергетически открытую систему, т. е. активно
организует поступление энергии за счет обмена веществ с окружающей
средой. Э то свойство обусловлено приспособленностью организма к
условиям обитания. Перекачивая из внешней среды энерг ию и за счет
этого понижая энтропию внутри себя, живые существа повышают
энтропию окружающей среды.
Благодаря обмену веществ организм выполняет работу, направлен­
ную на объекты внешней среды. Результатом ее является,
в частности, отыскание и поглощение пищи, что обеспечивает приток и
усвоение необходимых для жизнедеятельности веществ. Часть субстра­
тов и энергии, высвобождаемых в результате обмена веществ,
используется на постоянное самообновление. структур ор| анизма,
которые в процессе жизнедеятельное!и- и взаимодействия со средой
обитания подвергаются неблагоприятным изменениям. Э ти игменения
в целом повышают энтропию, тогда как самообновление препятствует
этому процессу. Постоянная работа по поддержанию в самих себе
энтропии на достаточно низком уровне служит важным свойством
живых форм.
В результате самообновления воссоздаются структуры, в точности
соответствующие разрушаемым. Э т о происходит благодаря наличию в
живых объектах и н ф о р м а ц и и (определенного знания), которая
создавалась в процессе эволюции вида и содержится в наследственном
веществе— Д Н К . Испольювание биологической информации состав­
ляет еще одно свойство живых форм. Воплощение информации в
структуру живого объекта происходит в процессе характерного для
живых форм и н д и в и д у а л ь н о г о р а з в и т и я , в ходе которого
наблюдается и такое свойство, как с п о с о б н о с т ь к р о с г у.
12
В структуры живых организмов входят уникальные по своим
свойствам химические соединения - биополимеры, не имеющие анало­
гий в неживой природе. Речь идет о б е л к а х - ф е р м е н т а х ,
благодаря которым биохимические реакции протекают в требуемом
направлении, с достаточными скоростями, при обычных условиях
температуры и давления. Ферменты обладают специфичностью дейс­
твия и катализируют превращения веществ определенного химическо­
го строении или даже отдельног о вещества. Специфичность ферментов,
равно как и белков, не выполняющих каталитических функций, зависит
от постоянства их первичной структуры — определенной последова­
тельности аминокислотных остатков в молекуле полимера. Белки
клеш к постоянно обновляются, вместо разрушающихся молекул синте­
зируются новые. Отдельные белковые молекулы воспроизводятся
таким образом всякий раз заново, но сохраняют постоянство своей
структуры. Источником информации служат н у к л е и н о в ы е
к и с л о т ы-. Такое отношение между белками и нуклеиновыми
кислотами соответствует двум формам существования биологической
информации, отражением которых на уровне организма служит
наличие у него г е н о т и п а и ф е н о т и п а .
Обмен веществ в зависимости от состояния внешней среды
способен к и з м е н е н и я м
приспособительного
ха­
р а к т е р а . Адекватная реакция организма как целого на состояние
окружающей, а также внутренней среды зависит от механизмов
регистрации соответствующих изменений, анализа поступающих дан­
ных, выработки решений о содержании и интенсивности ответа. Это
свойство роднит живые объекты с кибернетическими устройствами,
которые подчиняются законам передачи и переработки информации.
Термин информация употребляется здесь в широком смысле. Биологи­
ческая информация, о которой речь шла выше, количественно и
качественно эквивалентна наследственной информации, в которой
сконцентрирован многовековой опыт эволюционного развития. Инфор­
мация в кибернетическом смысле включает и личный опыт конкретного
организма. В биологии способность живого объекта воспринимать
действие внешних факторов обозначается термином раздражимость, а
осущесгвлять ответную реакцию — возбудимость. Еще одним важным
свойством живых форм является с п о с о б н о с т ь к р а з м н о ­
ж е н и гд, т. е. к воспроизведению себе подобных по типу обмена
веществ и принципиальным чертам структурно-функциональной
организации.
Совокупность свойств, перечисленных выше, обнаруживается в
отдельных живых объектах (особях). Есть также свойства, распростра­
няющиеся на область жизни в целом. Они отражают универсальные
принципы ее существования во времени и пространстве. Одно из таких
свойств — в к л ю ч е н н о с т ь о р г а н и з м о в в п р о ц е с с э вол ю ц и и. Благодаря этому жизнь как особое явление материального
мира сохраняется на протяжении вот уже 3 млрд. лет. Второе такое
свойство — с у щ е с т в о в а н и е о т д е л ь н ы х о р г а н и з м о в л и ш ь
в о в з а и м о д е-й с т в и и с д р у г и м и в с о с т а в е о с о б ы х
с о о б щ е с т в — б и о ц е н о з о в . В зависимости от подходов,
13
используемых авторами, существуют общие, сущностные, описа­
тельные формулировки. Всеобщим является определение жизни как
особой формы движения материи, которая возникла на определенном
этапе ее развития. Важное в практическом отношении следствие из
этого определения заключается в наделении живых объектов специфиче­
скими качествами, не сводимыми к закономерностям физики и химии.
В сущностных определениях жизни на первый план выдвигаются
отдельные свойства живых объектов. Ж изнь определяли как «питание,
рост и одряхление» (Аристотель), «стойкое единообразие процессов при
различии внешних влияний» (Тревиранус), «совокупность функций,
сопротивляющихся смерти» (Биш а), «химическую функцию» (Лаву­
азье), «сложный химический процесс» (И . П . Павлов), систему по
достижению низкоэнтропийных состояний. Классическое определение
жизни дано Ф . Энгельсом. Он пишет: «Ж изнь есть способ существования белковых тел. и этот способ существования- состоит посвоеТйу
существу в постоянном самообновлении химических составных частей
тел>Л.~И~'далее:'ЗТ6 такой способ, «существенным моментом которого
является постоянный обмен веществ с окружающей их внешней
природой, причем с прекращением этого обмена веществ прекращается
и жизнь, что приводит к разложению белка»2. В определении
Ф . Энгельса охарактеризован главный субстрат жизни — белок, важ­
нейшее условие его сохранения во времени — самообновление и необхо­
димая предпосылка к выполнению этого условия — обмен веществ на
основе взаимодействия с внешней средой.
В практическом отношении полезны описательные определения.
Одно из них, например, характеризует жизнь как макромолекулярную
систему, которой свойственны иерархическая3 организация, способ­
ность к самовоспроизведению, обмен веществ, тщательно регулируе­
мый поток энергии и которая представляет собой распространяющееся
ядро упорядоченности в менее упорядоченной Вселенной.
2.2. У Р О В Н И О Р Г А Н И З А Ц И И Ж И З Н И
Ж ивая природа является неоднородной, целостной системой,
которой свойственна и е р а р х и ч е с к а я о р г а н и з а ц и я . В ней
можно выделить ряд у р о в н е й . В практическом отношении полезно
и широко используется классификация уровней в соответствии с объек­
тами исследования биологических и антропобиологических наук.
Нередко при определении уровней исходят из основных методов,
используемых той или иной наукой — наблюдение невооруженным
глазом, с помощью светооптического микроскопа, электронного
микроскопа и т. д. Э ти два подхода дают сходные картины
иерархической организации живой природы (табл. 1).
В курсе биологии человек рассматривается в его взаимосвязях с
другими элементами органического мира, современное состояние
1 М аркс К .. Энгельс Ф Соч., 2-е изд., т. 20, с. 82.
2 М аркс К .. Энгельс Ф . Соч., 2-е изд., т. 20, с. 616.
3 Иерархия (от греч. Ыегов — священный и агсНе - власть) — расположение
частей или элементов целого в порядке от высшего к низшему.
14
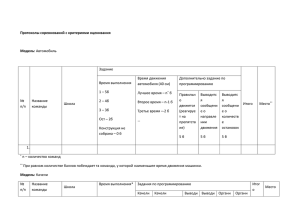
Т а б л и ц а I. Уровни организации и исследования, выделяемые в многоклеточ­
ном организме (по Э. де Робертису, В. Новинскому, Ф . Саэсу, 1967, с изменениями)
Ти п и чн ы е р азм еры
и зуч ае м о го о б ъ е к та
0,1 мм (100 мкм) и
более
100— 10 мкм
10— 0,2 мкм
(200 нм)
200— 1 нм
Менее! нм
О б ъ е к т и зучен и я
Органы,
организм
У р о ве н ь ор ган изаци и
(п о о б ъ е к т у исследо­
вания)
Органный, организменный
Тканевой
Клеточный
Ткани
Клетки
(эукариотические
и прокариотиче­
ские)
Клеточные
компо­ Субклеточный
ненты
Молекулы биополи­ М ак р о м о л е к у л я р ный
меров
Уровень
и сследования
(п о методу исследо­
вания)
Анатомический
Г истологический
Цитологический
Ультраструктурный
Физико-химический
(молекулярный)
которого является результатом эволюции. Механизмы эволюции
охватывают весь диапазон явлений жизни — от макромолекулярных до
биосферных. Главные события происходят на молекулярно-генетиче­
ском, клеточном, организменном, популяционно-видовом, биогеоценотическом уровнях организации живой природы1. Биологическое наслед­
ство людей формировалось в ходе исторического развития органическо­
го мира, поэтому его содержание отражает процессы и результаты
эволюционных преобразований на перечисленных уровнях. Н а каждом
из этих уровней выделяются элементарная единица (основной элемент)
е д и н и ц а — это
и элементарные явления. Э л е м е н т а р н а я
структура, закономерные изменения котогоЕ. обозначаемые как
э л е м е н т а р н ы е я в л е н и я , составляют на соответствующем
уровне содержание эволюционного процесса.
Элементарной
единицей
на
_
молекулярно­
г е н е т и ч е с к о м у р о в н е служит ^ен!— фрагмент молекулы
нуклеиновой кислоты, в котором записан определенный в качественном
и количественном отношении объем биологической информации. Эле^
ментарное явление заключается в процессе)конвариантной редуплика­
ции Али самовоспроизведении с изменениями генов. Путем редуплика"Щ ш Д Н К происходит копирование генов и заключенной в них
биологической информации, что обеспечивает преемственность и сох­
ранность (консерватизм) свойств организмов в ряду поколений.
Редупликация, таким образом, является ценовой наследственности. В
силу ограниченной стабильности молекул или ошибок синтеза в Д Н К
происходят изменения, которые искажают информацию генов. В
последующих циклах редупликации эти изменения воспроизводятся в
молекулах-копиях и переходят к особям дочерних поколений. Указанные
изменения возникают и : тиражируются закономерно, что делает
редупликацию информационных макромолекул конвариантной, т. е.
происходящей с некоторыми искажениями. В генетике последние
1 Приводится по Н. И. Тимофееву— Ресовскому и др. (1977). Клеточный уровень
выделен дополнительно.
15
получили название истинных мутаций. Конвариантность редупликации
служит основой мутационной изменчивости.
Биологическая информация молекул Д Н К непосредственно в про­
цессах жизнедеятельности не участвует. В действующую форму она
переходит в процессе биосинтеза белка, который происходит при
наличии специальных структур, субстратов и энергии. Указанный
процесс осуществляется на к л е т о ч н о_м у р о в н е организации
живой природы, элементарной единицей которого служит кпеткя
Элементарные явления представлены реакциями' клеточного метабо­
лизма. Благодаря деятельности клетки поступающие извне вещества
превращаются в субстраты и энергию, которые утилизируются в
процессе биосинтеза белков в соответствии с существующей информа­
цией. Таким образом на клеточном уровне сопрягаются механизмы
передачи информации и превращения веществ и энергии. Элементарные
явления на этом уровне создают энергетическую и вещественную основу
жизни на других уровнях.
Элементарной единицей о р г а н и з м е н н о г о
уровня
служит особь, которая рассматривается в развитии от момента
'зарождения до прекращения существования в качестве живой системы,
что
позволяет
назвать
этот
уровень
также
онтогене­
т и ч е с к и м . Закономерные изменения организма в индивидуаль­
ном развитии составляют элементарные явления. В ходе онтогенеза, в
результате реализации наследственной информации в определенных
условиях внешней среды формируется фенотип организмов данного
биологического вида.
Элементарной
единицей
популяционно-видового
у р о в н я служит популяция — совокупность особей одного вида.
Объединение особей в популяцию происходит на основе общности
генофонда. Популяция, в силу возможности межпопуляционных
скрещиваний, представляет собой открытую генетическую систему.
Действие элементарных эволюционных факторов приводит к эволюционно значимым изменения^ генофонда популяции, что и принимается
за элементарное явление на этом уровне.
Особи одного вида населяют территорию с известными абиотиче­
скими показателями (климат, химизм почв, гидрологические условия) и
взаимодействуют с организмами других видов. В процессе совместного
исторического развития организмов разных систематических групп
образуются динамичные, устойчивые во времени сообщества —
биогеоценозы,
которые
служат
элементарными
единицами
б и о г е о ц е н о т и ч . е с к о г о у р о в н я . Видовой состав, а так­
же характеристики местообитания для отдельных биогеоценозов
обеспечивают вещественно-энергетические круговороты, которые пред­
ставляют на рассматриваемом уровне элементарные явления. Ведущая
роль в этих круговоротах принадлежит живым организмам. Биоце­
ноз — это открытая в вещественном и энергетическом плане система.
Благодаря этому биогеоценозы объединяются в единый комплекс —
область распространения жизни или биосферу.
Рассмотренные уровни отражаю т общую структуру эволюционного
процесса, закономерным результатом которого является человек.
16
Элементарные единицы и явления различных уровней организации
жизни распространяются и на людей с некоторыми отклонениями в
силу их социальной сущности.
Глава
3
Б И О Л О ГИ Я К Л Е Т К И
К л е т к а является основной единицей биологической активности.
Она способна к самовоспроизведению в среде, не содержащей других
живых систем. Э т о наименьшая по объему структура, которой присуща
вся совокупность свойств жизни и которая может в подходящих
условиях поддерживать эти свойства в самой себе, а также передаватьих в ряду поколений. В природе планеты клетка играет роль
элементарной
структурно-функциональной
и генртчи^ ской ел иницы и составляет основу
жизнедеятельности и развития
всех
живы х
ф о р м . Клетка обеспечивает обмен веществ, размножение, наслед­
ственность, изменчивость, раздражимость, обусловливая тем самым
присущие органическому миру качества единства, непрерывности,
разнообразия.
Занимая по отношению к миру живых существ положение
элементарной еййницы, клетка обличается сложной структурой. При
этом определенные общие черты обнаруживаются во всех без
исключения клетках, характеризуя наиболее существенные стороны
клеточной организации как таковой.
3.1. П Р И Н Ц И П Ы С Т Р У К Т У Р Н О - Ф У Н К Ц И О Н А Л Ь Н О Й
О РГА Н И ЗА Ц И И КЛ ЕТ К И
В природе существует значительное разнообразие конкретных
клеточных форм. Вместе с тем число основных типов клеточной
организации ограниченно. Выделяют прокариотический и эукариотиче­
ский типы с подразделением второго на подтип клеток простейших
организмов и подтип клеток многоклеточных (рис. 1).
К летки п р о к а р и о т и ч е с к о г о
т и п а имеют особенно
малые размеры — не более 0,5— 3,0 мкм в диаметре. У них нет
морфологически обособленного ядра, так как ядерный материал в виде
Д Н К не отграничен от цитоплазмы оболочкой. В клетке отсутствует
развитая система мембран. Генетический аппарат образован един­
ственной кольцевой хромосомой, которая лишена 5СН1жныл ~бёл:
'ков — гистонов1.
— ——-----1 Гистоны являю тся белками клеточных ядер. Благодаря значительному количе­
ству основных аминокислот аргинина и лизина они имеют щелочной характер и пред­
ставлены пятью главными фракциями — Н 1, Ь}2а, Н2Ь, Н З и Н4, различающимися
содержанием аргинина, лизина, глицина. В хромосомах эукариотических клеток
гистоны, соединяясь с Д Н К , подавляют процесс считывания биологической инфор­
мации. Различия прокариотических и эукариотических клеток по содержанию гистонон указы ваю т на оа^ру,1С механизмы регуляции функций наследственного м-<териала.
"
У
Рис. 1. Типичные черты структурной организации клеток.
д —
п р о к а р и о ти че ск а я; б
-
э ук а р и о т и ч е ск а я р а с т и т е л ь н а я ;
в
животная.
/ У
У прокариот отсутствует клеточный центр. Для них не типичны
внутриклеточные перемещения цитоплазмы и амебоидное движение
Время, необходимое для образования двух дочерних клеток из
материнской (время генерации), сравнительно мало и исчисляется
десятками минут. Прокариотические клетки не делятся митозом.
К этому типу клеток относятся бактерии и сине-зеленые водоросли.
Эукариотический
тип клеточной организации пред­
ставлен двумя подтипами. Особенностью организмов п р о с ­
т е й ш и х (рис. 2) является то, что они, исключая колониаль­
ные формы, в структурном отношении представляют собой клетку,
в физиологическом — полноценную особь. В связи с этим в клетках
некоторых простейших имеются миниатюрные образования, выполня­
ющие на клеточном уровне функции органов, аппаратов и систем
органов многоклеточного организма (цитостом, цитофарингс и поро­
шица, сократительные вакуоли, генеративное и вегетативное ядра
инфузорий).
Высокая упорядоченность внутреннего содержимого эукариотиче­
ской клетки достигается путем к о м п а р т м е н т а л и з а ц и и- ее
объема — подразделения на «ячейки», отличающиеся деталями химиче­
ского (ферментного) состава.
Компартментализация (рис. 3) способствует пространственному
разделению веществ или процессов в клетке. Отдельный компартмент
представлен органеллой (лизосома) или ее частью (пространство,
I ограниченное внутренней мембраной митохондрии). Важная роль в
; осуществлении компартментализации принадлежит биологическим
1мембранам, которые выполняют ряд функций: отграничивающую
гТбарьёрную), регуляции и обеспечения избирательнОТТфошщаемости
веществ, образования поверхностей раздела между водной (гидро'фщщнойХ1 Г "негодной (гидроФо5ной)Фазами с размещением на „этих
поверхностях определенных ферментных комплексов. Благодаря прич-> сутствию липидов (жировых веществ) мембраны формируют гидро18
Рис. 2. Структурная организация
одноклеточного организма (инфузо­
ри я).
I
- ген еративное ядро; 2
- ве гетати вно е
ядро, 3 — цитостом с ц итоф ариш сом; 4
п ищ евар и тел ьн ы е
ва ку о л и ;
5 - порош ица;
(? - со кр а ти те л ьн ы е ваюуоли; 7
ги ал о ­
п л а зм а ; 8 — реснички.
фобную внутриклеточную фазу как компартмент для химических
реакций в неводной среде. Молекулярный состав мембран, набор
соединений и ионов, адсорбированных на их поверхностях неодинаков, с
что обусловливает их функциональную специализацию* Включение в )
мембрану молекул рецепторов делает ее восприимчивой к биологиче- ‘
ски активным соединениям, например гормонам.
Известно несколько схем взаимоотношений в мембране основных '
химических компонентов — белков и липидов, а также других веществ, <
адсорбируемых на мембранной поверхности.
В настоящее время большей популярностью пользуется точка/_.
зрения, согласно которой мембрана составлена из бимолекулярного! !!
слоя липидов. Гидрофобные участки их молекул повернуты друг к \другу, а гидрофильные находятся на поверхности слоя. Разнообразные )•
белковые молекулы встроены в этот слой или адсорбированы на его / '
поверхностях (рис. 4).
Благодаря компартментализации клеточного объема в эукариотиче­
ской клетке наблюдается разделение функций между разными структу­
рами. Одновременно различные органеллы закономерно взаимодей­
ствую т друг с другом.
Клетки м н о г о к л е т о ч н ы х
организмов (растительные и
животные) имеют оболочку. Плазмолемма (клеточная оболочка)
животных клеток образована мембраной, покрьТтой~~снаружТГ~СЛОем~
гликокдликсатодщвиой 10—20 нм. Основными компонентами глйкокаликса служат.комплексы полисахаридов с белками (гликопротеиды) и
жирами (гликолипиды). Плазмолемма выполняет отграничивающую и
барьерную функцию. Благодаря свойству избирательной проницаемо­
сти она регулирует состав внутренней среды клетки. Наличие в
плазмолемме молекул-рецепторов, специфически распознающих биоло­
гически активные вещества, например гормоны, обеспечивает целесо­
образную реакцию клетки на изменение ее окружения или состояния
организма.
В клетке выделяют ядро и цитоплазму. В ядре, наряду с оболочкой
и ядерным соком, обнаруживаются ядрышко и хроматин. Функцио­
нальное значение я д е р н о й о б о п о ч к и заключается в обособлег
нии наследственного материала (хромосом) эукариотической клетки от
цитоплазмы с присущими ей многообразны ми метаболическими реак­
циями, а также оегУлянии двухстппонниу взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных
бкОЛОядерным (перинуклеарным) пространством, которое может со-
общаться с канальцами цитоплазмати­
ческой сети. Она пронизана порами диа­
метром около, 120 нм. Пора или поровый
комплекс имеет определенную организа­
цию, что указывает на сложный характер
регуляций
ядерно-цитоплазматич&ских
транспортов. Количество пор зависит
от функционального состояния клетки —
чем выше активность наследственного
материала, тем больше их число. В об­
ласти поры начинается так называемая
плотная пластинка — белковый слой,
подстилающий на всем протяжении
внутреннюю мембрану ядерной оболоч­
ки и выполняющей в отношении ядра
скелетную функцию. Благодаря этому
ядро сохраняет свою форму при разруше­
нии ядерных мембран, но сохранении
плотной пластинки. Предполагают, что
закономерная связь хромосом с вещест­
вом этой пластинки способствует упорядоченнрму
расположению хромосом в
Рис. 3.
Компартмснтализацни
ядре.^р.
объема клетки с помощью мемб­
ран.
Основу я д е р н о г о
с о к а или
I
ядро, 2
ш е р о х о в ата я ц ии ш л .ю
матрикса
составляют
белки,
включая
м а ти че ск а и с е ть : Л
I р а н и ю р пп.! и п и т
п л а зм ати че ск и й п узы р е к . 4
и .ы ы и и
фибриллярные. Ядерный сок представ­
ча ты й ко м п л е кс. 5
гр а н у л » се к р ет».
ляет внутреннюю среду ядра и, по-види­
6
м итохондрия, 7
л и зосом а
мому, играет немалую роль в обеспе­
чении нормального функционирования
наследственного материала. Наличие в нем фибриллярных белков
указывает на выполнение им опорной функции.
Я д р ы ш к о представляет собой структуру, в которой происхо­
дит синтез и созревание рибосомных Р Н К (р Р Н К ), а также соединение
их с белками. Гены р Р Н К занимают определенные участки одной или
нескольких хромосом (у человека 13— 15 и 21—22 пары) — ядрышковые
организаторы, в связи с которыми и образуется ядрышко. В последнем
с помощью электронного микроскопа выявляю тся нитчатый и зерни­
стый компоненты. Нитчатый (фибриллярный) компонент представлен
комплексами белка и гигантских молекул РНК-предшественниц более
мелких молекул зрелых р Р Й К . В процессе созревания фибриллы
преобразуются в рибонуклеопротеидные зерна (гранулы), которыми
представлен зернистый компонент. Х р о м а т и н является интерфаз­
ной формой существования хромосом клетки. Его организация
рассматривается в 3.2.1.
Ц и т о п л а з м а представлена основным веществом (матрикс,
гиалоплазма), в котором распределены включения и органеллы.
Основное
вещество
цитоплазмы
заполняет прос­
транство между плазмолеммон, ядерной оболочкой, органеллами и
другими структурами. Даже электронный микроскоп не выявляет в нем
20
Рис. 4. Молекулярная ор| .пш пщия Оно
Ю1 ичегкой мембраны.
I
01 1
Г>ИМ . Г К \ . Я|ЖЬП
11.Ц1И ф«>Г«|н>. ПШНДОК,
'/
Гнмки
с
какой-либо внутренней организации. Оно представлено множеством
растворенных в воде органических и неорганических веществ, включая
ферменты и другие белки. В основном веществе цитоплазмы сконцен­
трированы предшественники и промежуточные продукты многйх
биохимических циклов. В нем происходит гликолиз, которому при­
надлежит важная роль в образовании потока энергии.
В к л ю ч е й и я м и (рис. 5) называют относительно непостоякггйёГкомпоненты цитоплазмы, которые служат запасными пита­
тельными веществами (крахмал, гликоген), продуктами, подлежащими
выделению из” клетки (гранулы секрета), балластными веществами
(некоторые пигменты). О р г а н е л л ы — это постоянные структуры
цитоплазмы, выполняющие в клетке определенные функции.
Выделяют органеллы общего значения и специальные. Последние
встречаются в большинстве клеток, но в значительном количестве
присутствуют только в клетках, специализированных к выполнению
определенной функции. К ним относятся микроворсинки всасывающей
поверхности эпителиальных клеток кишечника, реснички эпителия
трахеи и бронхов. Такие специальные органеллы, как синаптические
пузырьки, транспортирующие медиаторы-переносчики нервного воз­
буждения с одного нейроцита на другой или клетку рабочего органа, а
также миофибриллы, обеспечивающие акт сокращения мышцы,
имеются лишь в клетках определенной функциональной специализации.
Детальное рассмотрение специальных органелл входит в задачу курса
гистологии.
К органеллам общего значения относят элементы канальцевой и
вакуолярной системы цитоплазмы в виде шероховатой и гладкой
цитоплазматической сети, пластинчатый комплекс, митохондрии,
рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и
микротрубочки, центриоли клеточного центра. В растительных клетках
выделяют также хлоропласты, осуществляющие фотосинтез.
К а н а л ь ц е в а я и в а к у о л я р н а я с и с т е м а образова­
на сообщающимися или изолированными трубчатыми и уплощенными
цистернами, ограниченными мембраной и распространяющимися но
всей цитоплазме клетки. Нередко цистерны имеют пузыревидные
расширения. В этой системе выделяют шероховатую и гладкую
цитоплазматическую сеть (см. рис. 3). Особенность строения шерохова­
той сети состоит в прикреплении к мембранам полисом. В силу этого ее
функцией являе гея синтез определенных белков, например, секретируемых клет ками желез. Плотно упакованные слои цистерны шероховатой
сети представляют собой участки наиболее активного белкового
21
к »А
’•
•V
‘
О ж . чв-. л—’
г ’ с
,* "Л \ :
^ ^
V %л <«^
• ч .
* ;;;:■
л ■»
-V Д |
*г
»
*•I ?*
•
« •*
4
**
л Ч
*
,V
Рис. 5. Цитоплазматические включения эукариотической клетки,
к
п игм ент лип оф усц ин , б
1 лико ген, в
л ип и д н ая к а п л я ; г
гр анул ы секрета
синтеза и называются эргастоплазмой. Мембраны гладкой цитоплаз­
матической сети лишены полисом. В ней происходят некоторые стадии
обмена углеводов, жиров и других веществ небелковой природы.
Предполагают, что в участках гладкой сети начинается процесс
образования всех внутриклеточных мембран. По канальцам происходит
транспорт веществ.
Р и б о с о м а — это округлая рибонуклеопротеидная частица
диаметром 20— 30 нм. Она состоит из малой и большой субъединиц,
объединение которых происходит в присутствии информационной Р Н К
(и Р Н К ). Одна молекула и Р Н К нередко объединяет несколько рибосом|
наподобие нитки бус. Такая структура называется п о л и с о м о й .
Полисомы свободно располагаются в основном веществе цитоплазмы'
или прикрепляются к мембранам шероховатой цитоплазматической
сети. В обоих случаях они служат местом синтеза белка. При этом на
свободных полисомах образуются белки, используемые в жизнедея­
тельности самой клетки, а на прикрепленных — белки, функционирую­
щие вне тела клетки.
П ластинчатый
к о м п л е к с образован совокупностью
диктиосом числом от нескольких сотен до нескольких тысяч на клетку.
22
Диктиосома (рис. 6,а) прелставлеиа стопкой из 3— 12 уплощенных
дискообразных цистерн, от краев которых отшнуровываются пу­
зырьки. Локальные расширения цистерн приводят к образованию
вакуолей. В дифференцированных клетках позвоночных животных
диктиосомы обычно собраны в околоядерной зоне цитоплазмы.
В пластинчатом комплексе образуются секреторные пузырьки или
вакуоли, содержимое которых представлено так называемыми экспор­
тируемыми белками и другими соединениями, подлежащими выводу из
клетки. При этом просекрет, поступающий в дикгиосому из мест
синтеза, подвергается в ней некоторым химическим преобразованиям.
Он также обособляется (сегрегируется) в виде «порций», которые здесь
же приобретают оболочку из мембраны. В пластинчатом комплексе
образуются первичные лизосомы. В цистернах дикгиосом синтезиру­
ются полисахариды, образуются комплексы этих соединений с белками
(гликопротеиды) и жирами (гликолипиды), которые затем можно
обнаружить в гликокаликсе плазмолеммы.
М и т о х о н д р и и (рис. 6,г) — это структуры округлой или
палочковидной формы длиной обычно от 1,0 до 5,0 мкм, присутствую­
щие в большинстве клеток в количестве 150— 1500 экземпляров.
Оболочка митохондрии состоит из двух мембран, отличающихся по
химическому составу, набору ферментов и функциям. Внутренняя
мембрана образует впячивания листовидной (кристы) или трубчатой
(тубулы) формы. Пространство, отграниченное внутренней мембраной,
заполнено матриксом органеллы, в котором с помощью электронного
микроскопа обнаруживаются гранулы диаметром 20— 50 нм, накапли­
вающие ионы кальция и магния, частицы углеводов, например
гликогена. В матриксе же находится собственный аппарат биосинтеза
белка. Он представлен 2—6 копиями кольцевой, лишенной гистонов
молекулы Д Н К , рибосомами, транспортными Р Н К (г Р Н К ), фермента­
ми транскрипции и трансляции наследственной информации. По
основным показателям, таким как размеры и внутренняя структура
рибосом, организация собственного наследственного материала (Д Н К )
этот аппарат сходен с таковым у прокариот и отличается от аппарата
биосинтеза белка цитоплазмы эукариотической клетки. Э то говорит в
пользу симбиотической гипотезы происхождения последней. Гены
собственной Д Н К митохондрии кодируют последовательности нуклео­
тидов митохондриальных рибосомных и транспортных Р Н К , а также
первичную структуру некоторых белков, главным образом внутренней
мембраны органеллы. Последовательность аминокислот большинства
белков митохондрии закодирована в Д Н К хромосом клеточного ядра и
образуется вне органеллы в цитоплазме. Главная функция митохондрий
состоит в извлечении из органических веществ энергии путем их
окисления и накоплении энергии в биологически утилизируемой форме
в молекулах аденозинтрифосфорной кислоты (А Т Ф ). В осуществлении
энергетической функции участвую т все структурные компоненты
митохондрии, но ведущая роль принадлежит внутренней мембране.
В ней размешены комплексы ферментов транспорта электронов
(дыхательная цепь), дегидрогеназы, катализирующие окисление суб­
стратов дыхания, ферменты, сопрягающие процесс транспорта элек23
К"Ч
Г /
- V
О
,1-
* $
&
Ы
/ >
'*
<
.•
У *
•?
••->*' ^
'
,
.
«"
;а,>
- л . т
1
А
Л- «а .
■'
—
•
к
.
> ■
'
I
-У-Х
'V
V
'*,;<.
<-1д ' гV
,*«Г.
.
1*4.3 *
*
.
V.
*»
'«»
«’ ' V* Л - ”'•& Л ч \ <
Г
,
- » : ? *
-••/ ? л
.':**• •
.'е
с 5'.-*-^
Тчг * >
•
'
--' « л Ч
- >*
Г
V»
% Ч; ' ^
. •X
*
Рис. 6. 06щсклрточн1.1с органеллы. Объяснение я тексте
•1
тронов, сопровождающийся выделением энергии, с процессом синтеза
А Т Ф . Побочными функциями митохондрий являются синтез стеро­
идных гормонов, некоторых аминокислот (глутаминовой).
I
Лизосрмы
(рис. 6, 6) представляют собой пузырьки диа­
метром до 2 мкм, которые содержат набор ферментов кислых
гидролаз, катализирующих гидролитическое (в водной среде) расщепле­
ние нуклеиновых кислот, белков, жиров, углеводов. Они имеют
оболочку из одной мембраны, покрытой иногда снаружи волокнистым
слоем белка. Функция лизосом — внутриклеточное переваривание
различных химических соединений и структур. Первичными лизосомами называют неактивные органеллы, вторичными — органеллы, в ко­
торых происходит процесс переваривания. Вторичные лизосомы
образуются из первичных. Они подразделяются на гетеролизосомы
(фаголизосомы) и аутолизосомы (цитолизосомы) (рис. 6, в). В первых
переваривается материал, поступающий в клетку извне путем пиноцитоза или фагоцитоза, во вторых разрушаются собственные структуры
клетки. Вторичные лизосомы, в которых процесс переваривания
завершен, называют остаточными тельцами. В них отсутствую т
гидролазы и содержится непереваренный материал.
Сборную группу органелл составляют м и к р о т е л ь ц а . Э то
ограниченные одной мембраной пузырьки диаметром 0,1— 1,5 мкм с
мелкозернистым матриксом и нередко кристаллоидными или аморф­
ными белковыми включениями. К этой группе относятся пероксисомы. Они содержат ферменты оксидазы, катализирующие образова­
ние перекиси водорода, которая затем разрушается под действием
фермента пероксидазы. Эти реакции используются в различных мета­
болических циклах, например в обмене мочевой кислоты в клетках
печени и почек.
К органеллам общего значения относятся некоторые постоянные
структуры цитоплазмы, лишенные мембран. М и к р о т р у б о ч к и
(рис. 6, д) — трубчатые образования различной длины диаметром
24 нм, которые обнаруживаются в свободном состоянии в цитоплазме
или как структурные элементы центриолей, митотического веретена,
жгутиков и ресничек. Свободные микротрубочки и микротрубочки
жгутиков, ресничек и центриолей имеют разную устойчивость к
разрушающим воздействиям. В свободном состоянии микротрубочки
выполняют опорную функцию, определяют направления перемещения
внутри
клетки
пузырьков
и
других
структур.
Микроф и л а м е н т а м и (рис. 6,е) называют длинные, тонкие образова­
ния, обнаруживаемые по всей цитоплазме, но нередко концентрирую­
щиеся под плазмолеммой и вблизи ядерной оболочки. По-видимому,
существует
несколько
разных
классов
микрофиламентов.
Микрофиламенты из сократительного белка актина обусловливают ток
цитоплазмы, например в растительных клетках вокруг центральной
вакуоли, внутриклеточные перемещения пузырьков, хлоропластов,
ядер, амебоидное движение, деление клеточных тел перетяжкой.
Для животных клеток, делящихся митозом, части клеток растений,
грибов и водорослей характерен к л е т о ч н ы й ц е н т р , важным
элементом которого являю тся центриоли Центриоль имеет вид полого
25
цилиндра диаметром около 150 нм и длиной 300— 500 нм. Ее стенка
образована 27 микротрубочками, сгруппированными в 9 триплетов.
В функцию центриолей входит образование нитей митотического
веретена. Они поляризуют процесс деления клетки, обеспечивая
закономерное расхождение хроматид (дочерних хромосом) в анафазе
митоза.
Жизнедеятельность клетки как единицы биологической активности
обеспечивается совокупностью взаимосвязанных, приуроченных к опре­
деленным внутриклеточным структурам, упорядоченных во времени и
пространстве метаболических процессов. Э ти процессы образуют три
потока — информации, энергии и вещества.
.
) (М/
3.1.1. Поток информации
Благодаря наличию
потока
информации
клетка,
используя многовековой эволюционный опыт предков, создает органи­
зацию, соответствующую критериям живого, сохраняет и поддерживает эту организацию во времени, несмотря на меняющиеся условия
внешней среды, передает ее в ряду поколений. В потоке информации
участвуют ядро (Д Н к,, хромосом), макромолекулы, переносящие
информацию в цитоплазму (и Р Н К ), цитоплазматический аппарат
транскрипции (рибосомьЪ и полисом ы ,-тРН К, ферменты активации
аминокислот). На завершающем этапе этого потока полипептиды,
син!езированные на полисомах, приобретают третичную и четвертич­
ную структуру и используются в качестве катализаторов или структур­
ных блоков (рис. 7). Кроме ядерного генома, основного по объему
заключенной информации, в эукариотических клетках функционируют
также геномы митохондрий, а в зеленых растениях и хлоропластов.
И з приведенной схемы видно, что в рассматриваемом потоке
происходит перенос информации с Д Н К на белок. Ч то представляют
собой коды, с.помощью которых записана информация в Д Н К и белке?
Каков механизм перекодирования?
К о д и р о в а н и е заключается в записи определенных сведений
при помощи специальных символов с целью придать информации
компактность, обеспечить ее использование неоднократно и по частям,
создать удобства при транспортировке. Типичный пример кодирова­
ния — фиксация человеческой мысли в виде письменного текста.
В процессе кодирования путем сочетаний си м в о л о в составляют
к о д о в ы е г р у п п ы , служащие для обозначения существенного
элемента информации. Весь объем сообщения представлен опреде­
ленной последовательностью кодовых групп. Совокупность символов
составляет алфавит, а совокупность кодовых групп словарь кода.
Символами кода Д Н К служат дезоксирибонуклеотиды, различаю­
щиеся по азотистому основанию (адениловое, гуаниловое, тимидиловое, цитидиловое), поэтому алфавит четырехбуквенный. Кодовой
группой служит кодон — участок молекулы Д Н К , состоящий из трех
нуклеотидов. Э то делает код триплетным. Информация записывается в
линейном порядке по длине молекулы Д Н К в виде последовательности
кодонов. Код Д Н К неперекрывающийся, так как каждый нуклеотид
26
С1юц«а пцунтуры
у
Рис. 7. Поток биологической информации в клетке.
входит в один кодон. Он не имеет запятых и в пределах блока информа­
ции, соответствующего, например, одному полипептиду, кодоны сле­
дуют друг за другом без перерывов.
Символом кода белка служат аминокислоты. Они же соответству­
ют и кодовым группам. Информация также записывается в линейном
порядке по длине молекулы полипептида в виде последовательности
аминокислот.
Сопоставление участка молекулы Д Н К как начального пункта и
отвечающего ему по содержанию полипептида как завершающего
пункта потока информации указывает на к о л л и н е а р н о с т ь
к о д о в Д Н К и б е л к а : кодоны следуют в том же порядке, что и
остатки аминокислот, кодируемых ими.
Положение конкретного аминокислотного остатка в молекуле
полипептада может бы ть обозначено в Д Н К при помощи одного из
нескольких кодонов-синонимов, что свидетельствует о в ы р о жденности
кода
Д Н К . Указанное свойство вытекает из
соотношения объемов словарей кодов Д Н К и белка. Сочетанием по три
из четырех возможных дезоксирибонуклеотидов образуются 64
различных кодона, тогда как в состав белка входит 20 аминокислот.
Вырожденность кода Д Н К носит регулярный характер: большая часть
11
Риг. к. I 1|МШС1 Г 1ММ К|>\ И^ЮЙ Ми
И *К \ . 11<
1
1ф Г I пи I I
РНК
пн мм |»1Ч1К
I
V) гI И •И \ 1,1 .1 1 1 1 '.
К . 1.1 1*11 К
»1 г1
||.
Г»
Г |>\ 1'а1.Г,|
• .1111» »■! |1Ч 1*М. Ч
- ■!■
1 \ 1 М1 <
•
* "
||>|
"
Ф|
1 :ш
.1*. .
I | 11.1 .
V* I I I
| [)>,< ИП'Л
*
||||Гн>|
!|\1
1
||'|1 I! Ч» II I 'А I*МК ! |!|1«,*1|Ч‘.М|
11и.||| ■■
1.1ИЧ 11]>» '<I .-И I* |»'Н (,»■
ИI I II I■-.1
|1.ч\|«-||.|Ч
||||».1| М» 1.|1>и I * I I К
I»
I '11111111
■К .1/Ь I •' II ■!>! 1 1»« ИИ | ! I .1 )
\
X
о-’
информации приходится на первые два нуклеотида кодона. Каждой
аминокислоте соответствует не более двух таких начальных дуплета,
тогда как число кодонов-синонимов может доходить до шести
(например для аргинина). Вырожденность кода и информационная
неравнозначность нуклеотидов в кодоне влияют на фенотипическое
выражение точковых мутаций* Действительно, наряду с изменениями,
приводящими к замене одного аминокислотного остатка другим,
возможны «безмолвные» мутации, если изменение переводит кодон в
синоним. Х о тя замена кодона синонимом не нарушает последова­
тельности аминокислот в полипептиде, она может повлиять на
скорость его синтеза. Три кодона из 64, названные бессмысленными,
не кодируют аминокислот Они служат терминаторами и обозначакгг—'тотт<у прокрашен ИИ считывания информации. К о д
Д Н К
универсален
в том смысле, что он тождествен у всех
организмов. Единичные факты, не согласующиеся с таким заключение
ем, касаются детален пунктуации (например, обозначения начала
считывания у кишечной палочки и в клетке млекопитающего) и
считывания бессмысленных кодонов.
Перекодирование информации происходит
в
процессе
биосинтеза
белка.
На первом этапе,
обозначаемом как т р а н с к р и п ц и я , исходная информация Д Н К
считывается путем синтеза рибонуклеиновых кислот. Последние
комплементарны лишь одной из полинуклеотидных цепей Д Н К , место
тимина в них занимает близкое к нему азотистое основание — урацил.
В эукариотической клетке этот этап осуществляется в ядре, а также
независимо в митохондриях и хлоропластах. В результате тран­
скрипции образуется несколько разновидностей Р Н К , при этом и РН К
приобретает информацию о последовательности аминокислот в поли­
пептидах, а р Р Н К и т Р Н К обеспечивают перенос информации с и Р Н К
на полипептиды.
Особенность транскрипции с ядерной Д Н К эукариотической клетки
заключается в образовании первоначально большего количества Р Н К ,
чем то, которое затем примет в синтезе полипептидов непосред­
ственное участие. Избыточная Р Н К , природа и функции которой не
ясны, разрушается в ходе преобразования ( п р о ц е с с и н г а ) Р Н К
(рис. 8) перед транспортом ее из ядра в цитоплазму.
28
»
Считывание информации и РН К с переносом ее на белок (этап
т р а н с л я ц и и ) происходит в цитоплазме. Центральная роль здесь
принадлежит различным т Р Н К , которых в клетке имеется несколько
десятков. Каждый образец т Р Н К способен присоединять определенную
аминокислоту в активированном состоянии (обогащенную энергией).
В результате активации аминокислоты и присоединения ее к т Р Н К
образуется комплекс «аминоацил-тРНК». Благодаря наличию антико­
дона — последовательности из трех нуклеотидов, комплементарных
нуклеотидам кодона данной аминокислоты — т Р Н К узнает место этой
аминокислоты в полипептиде в соответствии с последовательностью
кодонов и Р Н К . Так как перенос информации на белок осуществляется
не с Д Н К , а с и Р Н К , колоны определенных аминокислот обозначают в
соответствии с нуклеотидным составом Р Н К (табл. 2). Таким образом,
именно т Р Н К считывает информацию с и Р Н К .
Т а б л и ц а 2.
Генетический код, универсальный для всех организмов
е *,
• Х 'э
*
В р ^ р а я~Л*у>Кк»
н 'н т '
у
II
УЦУ
У
УГУ
У АЦ
УГЦ
ц
УЦА
УАА
У ГА
А
УЦГ
УАГ
УГГ
ИДУ
ЦАУ
Р
Гер
УУА
1
Цис
Тир
уцц
УУЦ
Г
У АУ
Ф ен
У
, г
А
Ц
УУУ
а
' \с
Л ей
УУГ
ЦУУ
Лей
Ц.УЦ
Ц
ЦАЦ
цгц
ДНА
Ц АА
Ц УГ
ииг
1ГАГ
ЛУУ
АЦУ
ААУ
Ц УА
ь
А
Ц ГГ
Г
Сер
ц
А НА
АЛА
А ГА
А
АДГ
ЛАГ
А ГГ
Г
ГУ У
П (У
ГЛ У
ГГ.У
.У
ГУЦ
гцц
ГА Ц
АЦЦ
Арг
Л из
АУТ
Г
Мст
ГУ Г
П р и м I* ч а и и о
Асп
ГЦ А
гиг
П о д чер кн у I N кодоны-гор чи н а ю р ы
ц
ГГЦ
Гли
Ала
Вал
ГГ А
ГА Л
Глу
ГАД
я
У
А ГУ
Тре
ГУ А
а
ц
А ГЦ
Иле
А УА
В
т
ААЦ
АУЦ
К
У
Г.пи
А
V
е
ЦТ А
Асп
Г)
Г
Арг
Про
Иле
Я
Ц ГУ
Гис
НИИ
Три
ГГ
Г
А
Г
б
V
К
в1
к
аК
//
Рис. 9. Рибосомный цикл синтеза белка. Объяснение в тексте.
Сборка молекул полипептида происходит на рибосоме, которая
обеспечивает требуемое расположение участников процесса трансляции:
и Р Н К , комплексов «аминоацил-тРНК» и «тРНК-строящ ийся поли­
пептид». Представление о функции рибосом дает рибосомный цикл
синтеза белка (рис. 9).
Функционирующая рибосома состоит из большой и малой
субъединиц и молекулы и Р Н К (а). В одном из двух ее активных учас­
тков — пептидальном ( I) происходит наращивание полипептида, а к
другому — аминоацильному ( II) прикрепляются т Р Н К с активиро­
ванными аминокислотами. Комплекс «аминоацил-тРНК», прибывший
первым, инициирует считывание и занимает участок I (б). В участке II
фиксируется второй аналогичный комплекс, соответствующий пер­
вому смысловому коду и Р Н К (в). После образования между амино­
кислотами пептидной связи т Р Н К участка I высвобождается (г).
Н а ее место в виде комплекса с двумя аминокислотными остатками
перемещается т Р Н К , занимающая участок I I (д). К участку II при­
соединяется очередной комплекс «аминоацил-тРНК», отвечающий
следующему смысловому кодону и Р Н К . Описанный цикл повторяется,
пока не будет достигнут терминирующий кодон и Р Н К (У А А , У А Г или
У Г А ), по отношению к которому т Р Н К не существует. Н а этой стадии
рибосома распадается на субъединицы с высвобождением и Р Н К и
полипептида.
зо
3.1.2. Поток энергии
П о т о к э н е р г и и у представителей разных групп организмов
представлен
внутриклеточными
механизмами
энергообеспече­
ния — брожением, фото- или хемосинтезом, дыханием.
Центральная роль в биоэнергетике клеток животных принадлежит
дыхательному обмену. Он включает реакции расщепления низкокало­
рийного органического «топлива» в виде глюкозы, жирных кислот,
аминокислот и использования выделяемой энергии для синтеза
высококалорийного клеточного «топлива» в виде А Т Ф . А Т Ф и другие
соединения, богатые энергией в биологически утилизируемой форме,
называются макрозпгическими. Энергия А Т Ф , непосредственно или
•будучи перенесенной на другие макроэргические соединения, например
[креатинфосфа-ц используемый в мышцах, в разнообразных процессах
преобразуется в тот или иной вид работы — химическую (синтезы^,
осмотическую (поддержание градиентов веществ), электрическую,
механическую, регуляторную. Представление о потоке энергии в
животной клетке дает рис. 10.
Среди органелл такой клетки особое место в дыхательном обмене
принадлежит митохондриям, с внутренней мембраной которых связаны
ферменты дыхательной цепи, а также матриксу цитоплазмы, в котором ,
протекает процесс безкислородного расщепления глюкозы— анаэроб­
ный гликолиз. Из преобразователей энергии химических связей А Т Ф в
работу наиболее изучена механохимическая система поперечнополоса­
той мышцы. Она состоит из сократительных белков и фермента,
расщепляющего макроэргические соединения с высвобождением энер­
гии.
31
Белон
Н рахмал
Ж иры
Рис. П . Взаимосвязь внутриклеточного
обмена белков, жиров л углеводов через
цикл Кребса.
Особенностью потока «энер­
гии растительной клетки служит
фотосинтез — механизм пре­
образования энергии солнечного
света в энергию химических
связей органических веществ.
Механизмы энергообеспече­
ния клетки высокоэффективны.
Коэффициенты полезного дей­
ствия
хлоропласта
достига­
ю т 2 5 % , а митохондрии — 45—
6 0% , существенно превосходя
аналогичный показатель паровой
машины (8 % ) или двигателя
внутреннего сгорания (17%).
3 .1 .3 . П О Т О К
_
вещества
г
Реакции дыхательного обме­
на не только поставляют энер­
гию, но и снабжают клетку строительными блоками для синтеза разно­
образных молекул. Ими служат многие продукты расщепления пище­
вых веществ. Особая роль в этом принадлежит центральному звену
дыхательного обмена — ц и к л у
Кребса,
осуществляемому в
митохондриях. Через этот цикл проходит путь углеродных атомов
(углеродных скелетов) большинства соединений, служащих промежу­
точными продуктами синтеза химических компонентов клетки, а также
переключение метаболизма клетки с одного преобладающего пути
на другой, например, с углеводного на жировой. Таким образом,
дыхательный обмен одновременно составляет ведущее звено потока
веществ, объединяющего метаболические пути расщепления и синтеза
углеводов, белков, жиров, нуклеиновых кислот (рис. 11).
3.1.4. Другие внутриклеточные механизмы
Потоки информации, энергии и вещества осуществляются непре­
рывно и составляют необходимое условце существования клетки как
живой системы.
Кроме структур и процессов, прямо включенных в названные
потоки, в клетке функционируют механизмы, которые, хотя и могут
быть названы дополнительными, так же являю тся жизненно необходи­
мыми. Так, лизосомы, воздействуя ферментами на пиноцитированный
или аутофагированный материал, обеспечивают гидролитическое
расщепление макромолекул до низкомолекулярных соединений. Они же?
разрушают структуры, утратившие свое функциональное значение?
Пероксисомы ликвидируют возникающие в клетке перекиси, токсичные,
для живой протоплазмы. Организация внутриклеточных транспортных
потоков обусловливается активностью микротрубочек и микрофиб­
рилл.
32
3.1.5. Клетка как целостность.
Коллоидная система протоплазмы
Принципы структурно-функциональной организации клетки, изло­
женные выше, в основном сформулированы по результатам применения
в биологических исследованиях редукционистского подхода. Однако,
взаимодействуя с окружающей средой и отвечая на регуляторные
стимулы, к л е т к а
ведет
себя
как
целостность.
Об этом свидетельствует стереотипность реакций разных видов
клеток на действие разнообразных по природе раздражителей, вызыва­
ющих переход клетки в возбужденное состояние1.
Важная роль в функциональном объединении структурных
компонентов и компартментов клетки принадлежит свойствам живой
протоплазмы2. В целом ее принято рассматривать как особую полифазную коллоидную систему. О т банальных коллоидных систем
б и о к о л л о и д отличается сложностью дисперсной фазы, основу
которой составляют макромолекулы, присутствующие в составе
плотных, микроскопически видимых структур (органелл) или в диспер­
гированном состоянии, близком к растворам или рыхлым 'сете­
образным структурам типа гелей. Будучи коллоидным раствором в
физико-химическом смысле, биоколлоид благодаря наличию липидов и
крупных частиц проявляет одновременно свойства эмульсий и суспен­
зии. На обширных поверхностях макромолекул адсорбируются разно­
образные примеси, что вызывает изменение агрегатного состояния
протоплазмы. При этом совершается работа, в результате которой
осуществляются различные внутриклеточные превращения — сборка
мембран или микротрубочек из субъединиц , выброс из клетки секрета,
изменение конфигурации белковых молекул, приводящее к торможению
или усилению ферментативной активности. Особенностью биоколлоида
является также то, что в физиологических условиях переходы
протоплазмы из одного агрегатного состояния в другое обратимы
благодаря наличию особого ферментативного механизма. Свойства
биоколлоида обеспечивают клетке способность при наличии энергии в
^.,,ет на действие стимулов многократно совершать работу.
Представления о биоколлоиде соответствую т интегративному
подходу в анализе клетки и рисуют ее в виде сгустка живого вещества,
своеобразной упорядоченной совокупности молекул, макромолекул и
надмолекулярных комплексов, объединяемых в единое целое возможно­
стью взаимопереходов.
3.2. З А К О Н О М Е Р Н О С Т И С У Щ Е С Т В О В А Н И Я К Л Е Т К И
(
|
|
ВО В Р Е М Е Н И
Закономерные изменения структурно-функциональных характериггик клетки во времени составляют содержание ее жизненного цикла
клеточного цикла). К л е т о ч н ы й ц и к л — это период сущестйо1 Под клеточным возбуждением понимают обратимые изменения протоплазмы,
служащ ие пусковым механизмом и основой той или иной клеточной деятельности.
2 Протоплазма — содержимое живой клетки, включающее ее ядро и цитоплазму.
33
ч»
Рис. 12. Жизненный цикл клетки многоклеточного организма.
М — м итоз;
О , — п р есин тети чески й
период,
0 2 — п о стси н тети ч е ски й
период;
8 - си н тети ческий ,
период; К, и Р 2 — периоды покоя к л еточн ого ц икл а; I — м и тоти чески й цикл; 2
переход к ле тки
н диф ф еренцированное со сто ян и е ; 3 — ги бель к ле тки . 2с — к о л и ч е ств о Д Н К в диплоидиом наборе хромо­
сом; 4с — удвоенное к о л и че ств о Д Н К
вания клетки от момента ее образования путем деления материнской
клетки до собственного деления или смерти.
Обязательным
компонентом
клеточного
цикла
является
митотический
(пролиферативный)
ц и к л — ком­
плекс взаимосвязанных и детерминированных хронологически собы­
тий, происходящих в процессе подготовки клетки к пелению и на
протяжении самого деления. Кроме того, в жизненный цикл
включается п е р и о д в ы п о л н е н и я к л е т к о й многоклеточ­
ного
организма . с п е ц и а л ь н ы х
функций,
а
также
п е р и о д ы п о к о я . В периоды покоя ближайшая судьба клетки
неопределенна: она может либо начать подготовку к митозу, либо стать
на путь специализации (рис. 12).
Морфология и фазы процесса разделения ядерного вещества
материнской клетки между дочерними (собственно митоз) известны со
второй половины прошлого столетия (И . Д. Чистяков, В» Флемминг,
Э . Страсбургер). Представления о митотическом цикле оформились
после 1953 г., когда было установлено, что предмитотический синтез
Д Н К происходит в интерфазе и отделен во времени от начала и оконча­
ния митоза. В митотическом цикле выделены четыре перио­
да — м и т о з
(М ), а такж е п о с т м и т о т и ч е с к и й
(О,),
с и н т е т и ч е с к и й (5) и п р е д м и т о т и ч е с к и й (С 2) перио­
ды интерфазы. Продолжительность митотического цикла для боль­
шинства клеток составляет от 10 до 50 ч. Длительность цикла
регулируется путем изменения продолжительности всех его периодов.
У млекопитающих длительность М составляет 1— 1,5 ч, С 2— 2— 5 ч,
5 — 6— 10 ч.
Биологическое значение митотического цикла состоит в том, что он
обеспечивает преемственность хромосом в ряду клеточных поколений,
образование клеток, равноценных по объему и содержанию наслед­
ственной информации. Таким образом цикл служит универсальным
механизмом воспроизведения клеточной организации эукариотического
типа.
Главные события митотического цикла заключаются в следующем:
1) р е д у п л и к а ц и я
(самоупоение) наследственного материала
материнской клетки; 2) р а в н о м е р н о е р а с п р е д е л е н и е
этого материала~между дочерними клетками. При этом закономерно
изменяются химическая и морфологическая организация хромо­
сом — ядерных структур, в которых сосредоточено более 90%
наследственного материала эукариотической клетки.
3.2.1. Химическая и структурная организация хромосом
Хромосомы во взаимодействии с внехромосомными механизмами
обеспечивают: 1) хранение наследственной информации: 2) использо­
вание этой' информации для создания. -и~доддержани»- «легочной
организации; 3) регуляцию считы вания наследственной информации;
4) самоудвоение генетического материала; 5) передачу. его от
материнской клетки дочерним.
"“ Главные химические компоненты хромосом представлены Д Н К ,
основными (гистоновые) и кислыми (негистоновые) белками, на долю
которых п р и х о д и т с я соответственно 40% и около 2 0 % . В хромосомах
содержатся Д 1 К , липиды, полисахариды, идны мщщцтрв.
В молекулах Д Н К закодирована наследственная информация, что
делает их ведущим функциональным компонентом хромосом^ Д Н К
эукариотических клеток представлена следующими фракциями:
( I ) уникальные нуйеотйдные постедовательн6сти;(5Р повторы определе^ о й последовательности (о т 102 до Ю 4— 10 копий);<35> повторы
(10 — 10 копий). У н и к а л ь н ы е п о с л е д о в а т е л ь н о с т и
присутствую т в гаплоидном наборе в единственном— экземпляре— п
образуют д ^ с^ 1 м од ^ а^ ^ 5^ р |ё^ ю 3 3 5 ет5 ^ слё^ ствё^ рсо~Ш тф й ал& т. е. основную массу структурных* и регуляторньВГ^гЪнов.
К
фракции
со
средним
числом
повторов
относятся некоторые структурные гены, например кодирующие после­
довательность аминокислот в молекулах гистонов или нуклеотидов в
р Р Н К и т Р Н К . По расчетам в клетке человека находится не менее 450
генов р Р Н К . Наличие повторов повышает количество единиц тран­
скрипции определенной информации и, возможно, служит фактором
защиты генов, жизненно важных для всех клеток, против мутаций.
В эту же группу входит и часть пегупятпрньтх генпи ф р а к ц и я
многократно повторяющихся последователь­
н о с т е й образована нетранскрибируемой сателлитной (спутничной)
Д Щ К . Роль ее в физиологии наследственного материала неизвестна.
О на^возможно, выполняет функцию спейсеров, т. е. фрагментов,
разделяющих структурные и регуляторные гены, или обусловливает
взаимоузнавание гомологичных хромосом. В Д Н К хромосом человека,
по ориентировочным данным, уникальные последовательности состав­
ляю т более 56%, многократно повторяющиеся — 12%, среднеповторя­
ющиеся — 8 % .
2*
35
10 нм
I
Г к
Н2В
,п с
Н2А
Н
Н4
Рис. 13. Интерфазная и митотическая
структурной организации хромосом,
формы
а
хроматин в и нтер ф азн ы х ядр ах, б
вкл ю чен и е в ядра
р ад и о а к т и в н о ю пр ед ш ествен н ика си н те за Д Н К , в си н тети че
еком периоде м и тоти ческо го ц икл а, в
к л е т ка в м етаф азе
м итоза.
Г истон
Н1
-Д Н Н
Рис. 14. Пуклеосомная организация элементар
ной нити хромосомы.
Г и с т о н ы как уже говорилось, представлены пятью главными
фракциями и выполняют структурную и регуляторную роль. Число
фракций н е г и с т о н о в ы х б е л к о в превышает 100. Среди них
ферменты синтеза и процессинга Р Н К , редупликации и репарации Д Н К .
Кислые белки хромосом выполняют также структурную и регуля­
торную роль. Р Н К хромосом представлена отчасти продуктами
транскрипции, еще не покинувшими место синтеза. Некоторым
фракциям свойственна регуляторная функция. Регуляторная функция
компонентов хромосом заключается в «запрещении» или «разрешении»
считывания информации с молекулы Д Н К .
Осуществление хромосомой отдельных функций приурочено к фазам
и периодам митотического цикла и сопровождается изменением ее
морфологии. Различают м и т о т и ч е с к у ю
и интерфаз­
ную формы
структурной организации хромосом, взанмопереходящие друг в друга в митотическом цикле. Первая свойственна перио­
ду
митоза,
когда
под
микроскопом
хромосомы
видны
как отдельные интенсивно окрашенные, плотные тельца. Вторая
соответствует хроматину ядер интерфазных клеток, который виден под
микроскопом как совокупность более или менее рыхло расположенных
нитчатых образований и глыбок (рис. 13).
'^/Элементарной структурой хромосомы, различимой с помощью
электронного микроскопа, является нить диаметром 10— 13 нм',
представляющая собой комплекс Д Н К и гистоновых белков (нуклеоги1 I нм
36
10
мкм
10 ' мм.
г
г
стон). Толщина нити зависит от располагающихся по ее длине
телец — нуклеосом. Диаметр межнуклеосомных участков менее 1,5 нм,
что совпадает с толщиной биоспирали Д Н К (рис. 14). Ядра телец
образованы 8-ю молекулами гистонов 4-х разных типов — Н2а, Н2Ъ,
НЗ и Н4. Они служат основой, на которую «накручены» фрагменты
Д Н К длиной примерно в 200 пар нуклеотидов. Гистон Н1 «сшивает»
витки Д Н К . Функциональное значение нуклеосом неясно. Есть данные,
что транскрибируемые фрагменты Д Н К , кодирующие р Р Н К , не имеют
нуклеосомной структуры. В отношении других генов есть указания, что
при транскрипции нуклеосомная структура утрачивается. Закручивание
молекул Д Н К на гистоновые тельца уменьшает длину биоспирали
Д Н К в 7 раз, т. е. служит целям упаковки наследственного материала.
Данные микроскопического и электронно-микроскопического изуче­
ния хроматина и митотических хромосом дают следующую схему
структурной организации хромосомы. Биспираль Д Н К диаметром
1.5 нм в результате скручивания и присоединения белка преобразуется в
нуклеогистоновый комплекс с нуклеосомной структурой. Он имеет вид
нити диаметром 10— 13 нм. При дальнейшем скручивании и присоеди­
нении белков возникает нить диаметром 20— 25 нм. Она обнаруживает­
ся с помощью электронного микроскопа как в интерфазных, гак и в ми­
тотических хромосомах. В результате дальнейшего скручивания этой
нити, происходящего многократно и дополняемого складыванием,
образуются митотические хромосомы. Э т а схема носит предвари­
тельный характер, она объединяет области интереса цитогенетика
медико-генетической консультации (микроморфология митотических
хромосом) и специалиста по функциональной организации хромосомы
на ультраструктурном и молекулярном уровнях.
Реорганизация нити нуклеогистона с образованием более компак­
тной структуры называется с п и р а л и з а ц и е й
(конден­
сацией),
процесс, обратный описанному, — д е с п и р а л и з а ц и е й ( д е к о н д е н с а ц и е й ) . Благодаря спирализации дос­
тигается плотная упаковка наследственного материала, что важно
при перемещениях хромосом в процессе митоза. О плотности упаковки
свидетельствуют следующие цифры. Ядро соматической диплоидной
клетки человека содержит около 6 пг Д Н К , что соответствует нити
нуклеогистона длиной почти 2 м. Совокупная же длина всех хромосом
клетки человека в метафазе митоза равна 150 мкм. Биоспираль из 100 г
Д Н К человека, если ее вы тянуть в одну нить, покроет расстояние
2.5 х 10ю км, что более чем в 100 раз превосходит расстояние от Земли
до Солнца.
Изложенные сведения об укладке нити нуклеогистона согласуются с
Генетическими представлениями о непрерывности и линейности распо­
ложения генов по длине хромосом. Они соответствую т допущению, что
каждая хромосома содержит одну двойную спираль Д Н К . В особых,
так называемых п о л и т е н н ы х х р о м о с о м а х клеток насеко­
мых одновременно присутствует несколько двойных спиралей Д Н К .
Так как они уложены «бок в бок», такая конструкция совместима с
принципом линейного и непрерывного размещения генов.
Для изучения кариогипа особое значение имеют митотические
37
метафазные
хромосомы
ТрисГТ5Г"Т)ни образованы
двумя х р о м а т и д а м и .
Ц ентром ера
Последние являются дочер­
ними хромосомами, кото­
Г етер ох ро м ати чесн и н у ч а с т о н
рые в процессе митоза
разойдутся в дочерние клет­
ки. Хроматиды соединены
в области п е р в и ч н о й
Х р ом атм да
перетяжки (центро­
меры. кинетохора),
к которой прикрепляются
нити
веретена деления.
Фрагменты, на которые
первичная перетяжка делит
хромосому,
называются
М атринс
п л е ч а м и , а концы хро­
мосомы — т е л о м е р а м и.
В зависимости от ^ '
Я д ры ш ко
положения первичной пере- уЧ '
тяжки различают м е т а В тори чн ая
ц е н - Т Р и ч е с к и е (рав­
п ер етя ж ка
ноплечие), с у б м е т ац е н т р и ч е с к и е (уме­
ренно неравноплечие), г. кроцентрические и
субакроцентричес к и е (выраженно неравРис. 15. М етафазная хромосома
ноплечие) хромосомы. У че­
ловека метацентрическими
являются хромосомы 1 и 3 пар, Х-хромосома, субметацентрическими
2,6— 12, 16—20 пары, акроцентрическими и субакроцентрическими —
4— 5, 13— 15, 21—22 пары и V-хромосома. При использовании некото­
рых методов приготовления препаратов в хромосомах видны полухроматиды, однако вопрос об их присутствии в клетке нельзя считать
решенным. Возможно они являются результатом воздействия на
вещество хромосомы материала, используемого для приготовления
препарата. Некоторые хромосомы имеют в т о р и ч н ы е п е р е т я ж к и . Они возникают в участках неполной конденсациихроматина,
например, в околоцентромерных участках длинного плеча 1, 9 и 16 хро­
мосом человека. Вторичные перетяжки отделяют концевые участки
коротких плеч 13— 15, 21—22 хромосом человека в виде спутников.
В области вторичных перетяжек некоторых хромосом располагаются
ядрышковые
организаторы.
Они содержат гены,
кодирующие р Р Н К , и служат местом образования ядрышка. Описан­
ные структурные особенности используют для идентификации
хромосом.
Х отя для интерфазных хромосом в целом свойственно деспирализованное состояние, степень спирализации отдельных фрагментов
Т о чка прикрепле
н и я нити в е р е т е н а
38
/
Уу
/у /
варьирует. Выделяют эухроматин, структурный гетерохроматин и
факультативный гетерохроматин. г Э у х Р о м а т и й
образован
участками хромосом, которые десгшрализируются в конце митоза.
В интерфазных ядрах — это слабо окрашивающиеся нитчатые струк­
туры. В области эухроматина располагаются структурные гены,
т р у к т у р II ы й т е т е р о х р о м а г и И ) отличается высокоспирализованным состоянием, которое сохраняется на протяжении
всего митотического цикла. Он занимает постоянные участки, сходные
в гомологических хромосомах. Обычно это фрагменты, прилегающие
к области центромеры, а также расположенные на свободных концах
(теломерах) хромосом. Э то т вид гетерохроматина структурных генов,
по-видимому, не содержит и функция его не ясна. В каждой хромосоме
свой порядок расположения эу- и гетерохроматиновых участков. Это
используется для идентификации отдельных хромосом в цитогене­
тических
исследованиях
человека.
('Факультативный)
.г е т е П-р-х п о м а г и нД образуется при спиралПзации одной из двух"
гомологических хромосом. Типичный пример — генетически неактив­
ная Х-хромосома соматических клеток женских особей млекопитающих
и человека (тельца полового хроматина). Функциональная роль факультативной гетерохроматизации заключается в компенсации (снижении) дозы определенных генов, |
(3.2??)) Изменения наследственного материала
в митотическом цикле
По двум главным событиям митотического цикла в нем выделяют
<Я.$ Р е п р о д у к т и в н у ю и р а з д е л и т е л ь н у ю фазы, соответствующие интерфазе и митозу классической цитологии.
В начальный отреюк интерфазы — в постмитотический (пресинтетический или С ,) период, велел за митозом восстанавливаются черты
организации интерфазной клетки, завершается формирование ядрышка.
Из цитоплазмы в ядро поступает значительное (до 9 0 % ) количество
белка. В цитоплазме, параллельно реорганизации ультраструктуры,
интенсифицируется синтез белка, что способствует увеличению массы
клетки. Если дочерней клетке предстоит вступить в митотический цикл,
синтезы приобретают направленный характер: образуется запас
предшественников Д Н К , ферменты, катализирующие реакцию ре­
дупликации, синтезируется белок, включающий эту реакцию. Таким
образом, осуществляются процессы подготовки следующего периода
ингерфазы — синтетического.
В синтетическом или 5-периоде удваивается количество наслед­
ственного материала клетки. За малыми исключениями редуплика­
ция1 Д Н К осуществляется полуконсервативным способом. Он заклю­
чается в расхождении биспирали Д Н К на 2 молекулы с последующим
синтезом возле каждой из них комплементарной молекулы. В результа­
те возникают 2 идентичные биспирали. Молекулы Д Н К , компле­
ментарные материнским, образуются фрагментами по длине хромосо1 Ин*<да улове пне ЛИК *<означают термином репликации, оставляя термин
редупликация для обозначения удвоения хромосом
39
мы, асинхронно в разных участках одной хромосомы, а также в
разных хромосомах. Такие фрагменты составляю т единицы репликации
или р е п л и к о н ы . Затем фрагменты новообразованной Д Н К
«сшиваются» в одну макромолекулу. В клетке млекопитающего
содержится более 20 ООО репликонов, причем количество их меняется в
онтогенезе. Участки хромосом, в которых начинается синтез Д Н К ,
называют точками инициации. Возможно ими являются точки
прикрепления интерфазных хромосом к внутренней мембране ядерной
оболочки. Можно думать, что Д Н К отдельных фракций, о которых
речь шла выше, редуплицируется в строго определенной части 5-пери­
ода, например большая часть генов р Р Н К удваивает Д Н К в начале его.
Редупликация запускается поступающим в ядро из цитоплазмы
сигналом, природа которого не выяснена. Синтезу Д Н К в репликоне
предшествует синтез Р Н К . Скорость редупликации составляет
0,2—2 мкм/мин. Она возрастает к концу синтетического периода.
В клетке, прошедшей 5-период интерфазы, хромосомы содержат
удвоенное количество наследственного материала. Наряду с Д Н К в
синтетическом периоде интенсивно образуется Р Н К и белок, а количе­
ство гистонов точно удваивается. Около 10% Д Н К животной клетки
находится в митохондриях. Незначительная часть митохондриальной
Д Н К редуплицируется в синтетичедком, тогда как основная — в
постсинтетическом периоде интерфазы.
Отрезок времени от окончания синтетического периода до начала
митоза .занимает предмитотический (постсинтетический) или С 2 -пери­
од интерфазы. Он характеризуется интенсивным синтезом Р Н К и
особенно белка. В итоге завершается удвоение массы цитоплазмы по
сравнению с началом интерфазы, что необходимо для перехода клетки
в период митоза. Часть образуемых белков (тубулины) используется в
дальнейшем для построения микротрубочек веретена деления. Синтети­
ческий и предмитотический (постсинтетический) периоды связаны с
митозом непосредственно. Э то позволяет выделить их в особый период
интерфазы — препрофазу.
М итоз делят на 4 фазы: профазу, метафазу, анафазу и телофазу.
Наряду с реорганизацией морфологии и упорядоченными перемеще­
ниями хромосом обязательным для разделительной фазы цикла
является построение м и т о т и ч е с к о г о а п п а р а т а . Он состо­
ит из системы микротрубочек, которые образуют ахроматиновое
веретено или веретено деления, и структур, поляризующих митоз1 .
В клетках млекопитающих и человека роль поляризующих структур
выполняют центриоли.
Митотический аппарат обеспечивает направленное перемещение
дочерних хромосом в анафазе. Для этого необходим контакт
микротрубочек со специализированными участками хромосом —
центромерами (кинетохорами). При разрушении центромер в экспери­
менте расхождения хромосом к полюсам клетки не происходит.
Природа сил, обеспечивающих расхождение, не известна. Предположи­
тельно в основе функционирования митотического аппарата лежит
1 Ряд авторов включают в митотический аппарат такж е хромосомы.
40
механохимическая система, аналогичная обнаруженной в поперечнопо­
лосатой мышце. Она включает сократимые белки и фермент,
катализирующий расщепление А Т Ф для обеспечения процесса энергией.
Данные генетики и цитологии указываю т на с о х р а н е н и е
структурной индивидуальности хромосом в
к л е т о ч н о м ц и к л е . Есть свидетельства в пользу упорядоченно­
го размещения хромосом в объеме интерфазного ядра. Особенности
взаиморасположения хромосом могут иметь функциональное значение.
Так, пространственная близость в клетках человека хромосом 13, 14, 15,
21 и 22 пар, содержащих гены р Р Н К , объясняется, по-видимому, их
участием в формировании ядрышка и образования рибосом. Есть
указания на то, что в клетках разных типов расположение хромосом не­
одинаково.
Н а основе митотического цикла возник ряд механизмов, с помощью
которых в том или ином органе количество наследственного материала
может бы ть увеличено при сохранении постоянства числа клеток. Так,
удвоение Д Н К иногда не сопровождается разделением цитоплазмы.
Поскольку механизм такого удвоения идентичен механизму редуплика­
ции Д Н К в митотическом цикле и оно сопровождается кратным
увеличением числа хромосом, это явление получило название
э н д о м и т о з а . С генетической точки зрения результат эндомитоза
следует рассматривать как геномную соматическую мутацию, о чем
будет сказано ниже. П о л и т е н и я
заключается в увеличении
содержания Д Н К в отдельных хромосомах при сохранении их
пиплоидного числа. И эндомитоз, и политения приводят к образованию
шолиплоидных клетокТ] отличающихся кратным увеличением объема
Наследственного материала. Таким образом, в таких клетках в отличие
от диплоидных гены повторены более чем 2 раза. Пропорционально
увеличению генов увеличивается масса клеток, что, в свою очередь,
Г пбвышает функциональные возможности органа. /
Наряду с непрямым делением или митозом :уществует прямое
деление клеток — а м и т о з . Последний заключается в разделении
ядра путем перетяжки без сложной перестройки наследственного
материала в виде конденсации хромосом и их точного распределения
между дочерними клетками с помощью митотического аппарата.
Предполагают, хотя это строго не доказано, что вслед за ядром делится
цитоплазма. Обычно амитоз рассматривают как неполноценное
деление, происходящее вследствие отсутствия механизма закономерно­
го и точного разделения Д Н К . Измерения показали, что на двуядерной
стадии предполагаемого амитотического деления ядра содержат, как
правило, равное и кратное гаплоидному набору хромосом количество
Д Н К . Э то снимает вопрос об их генетической неравнозначности.
Исследование с помощью современных методов показало, что все
двуядерные клетки в печени мелких грызунов появляются в результате
незавершенного митоза. Бытовавш ая ранее точка зрения, согласно
которой двуядерные печеночные клетки образуются вследствие амитотической перешнуровки ядер, не подтверждается. Применение совре­
менных методов исследования в отношении клеток других органов и
тканей даст ответ, существуют ли митоз и амитоз как 2 самостоятель­
ных способа клеточного размножения.
41
за
V
-
►
*
» \ ? 1 ,
>* ч
^
А ^ / ' * 1с
V
4
С
IX II хк ■и и
№ и кй ик йй и м
1
2
3
4
Ю
МЧ ЛЛ ЙЙ
13
14
■
=КК
**
С
Л Л
л А
21
22
19
л
5
11
12
Е XX лл лл
15
20
Рис. 16. Хромосомные наборы мужчины ( I )
классификации.
16
17
18
й
А
X
У
и женщины ( I I )
согласно денверской
а — м е т а ф а зн а я п л а сти н к а ; 6 — кариотип; в — идиограм м а; А — С : группы ауто со м ; Х ,У — п ол овы е хромо­
со м ы ; 1— 22: отд ельн ы е ауто сом ы . М е та ц е н тр и ч е ск а я (в, I ) , су б м е та ц е н тр и че ск а я (в , 4— 5 ), акроцентрич е с к а я (в , 21— 22) хромосомы.
3.3. П О Н Я Т И Е О К А Р И О Т И П Е
Исследованиями цитологов установлен факт специфичности хромо­
сомного набора клеток организмов одного вида. Специфичность
проявляется в постоянстве числа хромосом, их относительных
размеров, формы, деталей строения. Хромосомный комплекс клеток
конкретного вида растений и животных с присущими ему морфологиче­
скими особенностями, называется к а р и о т и п о м . Важнейшим
показателем кариотипа служит число хромосом (табл. 3).
42
7
• • *• • • < V
/Г'*'*
+ ЛЛ.***
у*1*
*
ХШ0€т
-
.
* *
^
в »
у
й й «к ■к «а
1
М
6
°
2
3
XX №
7
8
XX АА хь
13
14
16
4
XX XX IX XX
9
10
*
с
6
20
АА
АА
21
22
11
17
16
8
!
10-11
12
12
ХА х « X*
> XX як
19
5
18
т
1
16
1
1
17® 18
II
М
19 20
X X
Рис. 16. Продолжение.
Д ля соматических клеток многоклеточных организмов характерен
диплоидный
( у д в о е н н ы й ) хромосомный набор. В нем
каждая хромосома имеет парного себе гомологичного партнера,
повторяющего в деталях размеры и особенности ее морфологии. Таким
образом, в хромосомном наборе соматических клеток выделяют
г о м о л о г и ч н ы е (из одной пары) и н е г о м о л о г и ч н ы е
(из разных пар) хромосомы.
Половые клетки отличаются вдвое меньшим — г а п л о и д н ы м
числом хромосом. Хромосомный набор гамет включает по одному
партнеру каждой гомологичной пары.
Хромосомному комплексу свойственны половые различия. Наборы
43
хромосом самца и самки отличаются по одной паре. Поскольку эти
хромосомы участвуют в определении пола организмов, они называ­
ю тся п о л о в ы м и (гетерохромосомами). Остальные пары пред­
ставлены а у т о с о м а м и
и неразличимы по своей структуре у
самца и самки.
Таблица
организмов
3.
Число
хромосом
соматических
Виды
Малярийный плазмодий (Р1актос1ш т т а !а п а е )
Гидра (Н убга уи1^апк)
Таракан (В1а1а опеп1аПк)
Комнатная муха (Микса ботекИ са)
Сазан (С урп п ик сагрю)
Окунь (Регса ПиуйайПк)
Зеленая лягуш ка (Р а п а екси1еп1а)
Голубь (С о 1 и тЬа ЙУ1а)
Кролик (Ьерик с и т е Ы и к )
Шимпанзе (АпН'орорНЬесик)
Человек (Н о т о кар 1епк)
клеток
некоторых
видов
Ч исло хромосом
2
32
48
12
104
28
26
80
44
48
46
Исследование кариотипа человека проводится путем микроскопирования препаратов метафазных хромосом (рис. 16). Для такого
исследования необходимо иметь большое ь-оличргтво кпеток. делящихся
м н т т л м ^ тп м у условию удовлетворяет, например, пунктат костного
мозга. Ч аше прибегают в непрямому способу получения интенсивно
размножающихся клеток. Клетки соединительной ткани кожи, различ­
ных тканей абортированных эмбрионов или околоплодной жидкости,
лимфоциты периферической крови выращивают вне организма в куль­
туре ткани. Высокой митотической активности лимфоидных клеток
добиваются, воздействуя на них особым веществом растительного
происхождения — фитогемаглютинином (Ф Г А ). Делящиеся клетки
обрабатывают алкалоидом колхицином, который, разрушай щ щ ю ц зу—
бочки веретена деления, вызывает торможение митоза в|метафазе7чем
способствует накоплению метафазных клеток. В условТТЯх обработки
колхицином облегчается тахже разброс хромосом по плоскости
предметного стекла в процессе приготовления гистологических препа­
ратов. Затем на клетки в культуре действуют гипотоническим
раствором, чтобы добиться лучшего разобщения х р о м о с о м . После
фиксации в растворе, содержащем уксусную кислоту и этиловый
адквтшш^_взвесь клеток наносят на предметное стекло и высушивают.
В результате обработки клеток колхицином, гипотоническим раство­
ром и высушивания получают достаточный разброс хромосом мета­
фазных пластинок. Э то позволяет проводить индивидуальный анализ
их размеров и морфологии. Полученные препараты окрашивают
такими кпягитепями, кяк я чур основной фуксин, орсеин, дающими
сплошное о к р а ш и и я н и р в е щ е с т в а х р о м о с о м . Д л я изучения отбирают
цельные метафазные пластинки без налегающих друг на друга
хромосом (см. рис. 16,а). Последние должны иметь среднюю степень
44
конденсации. Для индивидуальной идентификации хромосом использу­
ю т следующие признаки: размер, положение первичной перетяжки,
наличие вторичных перетяжек и спутников. Результат представляется в
виде идиограммы, на которой хромосомы располагаются в порядке
убывания размеров (см. рис. 16, б, в).
В случае применения метода, описанного выше, идентификацию
хромосом
человека
проводят
согласно
денверской
к л а с с и ф и к а ц и и 1960 г. (табл. 4).
■
Таблица
центромеры
4.
Классификация хромосом по размеру и расположению
Г р у п п а хромосом
А (I)
В (II)
С (III)
О ( IV )
Е (V )
Р ( V I)
С (V II)
Х-хромоеома
(относится
пе I I I )
У-хромосома
к
Н о м ер по
кар и оти п у
Х а р а к те р и с ти к а хромосом
1, 2, 3
4,5, 6— 12
13— 15
16— 18
19 20
21— 22
1и 3 — почти метацентрические; 2 — круп­
ная субметацентрическая
Крупные субметацентрические
Средние
Средние акроцентрические
Мелкие субметацентрические
Самые мелкие метацентрические
„
акроцентрические
,
груп­
23
23
М елкая акроцентрическая
Недостатком денверской классификации является то, что разграни­
чение гомологичных пар внутри группы хромосом встречает зачастую
непреодолимые трудности.
В настоящее время используются дифференциальные методы
окрашивания метафазных хромосом с избирательным выявлением их
отдельных фрагментов. Топография окрашиваемых участков по длине
хромосомы зависит от локализации определенных фракций Д Н К ,
например сателлитной, распределения участков структурного гете­
рохроматина и ряда других факторов.
Применяют 4 основных метода дифференциальной окраски: О, С,
К и С. Все они вы являю т закономерную линейную неоднородность
фрагментов по длине метафазных хромосом. Характер окрашивания
специфичен для каждой не гомологичной хромосомы, что дает их
точную идентификацию (рис. 17). Постоянство локализации окрашива­
емых фрагментов позволяет составить «химические» карты хромосом.
Сопоставление этих карт с генетическими используется для расшифров­
ки функционально-генетических особенностей различных районов хро­
мосом. Н а основе избирательной окраски в 1971 г. в Париже были
разработаны
карты
линейной
дифференциров а н н о е т и ~х р о м о с б~М человека тгтТредложена "система- их
обозначения!
— ---------Латинскими буквами р и д обозначаются, соответственно, короткое
и длинное плечо хромосомы. О т центромеры к теломере по имеющимся
45
Рис. 17. Схематическое изображение расположения локусов в хромосомах человека
при их дифференциальной окраске.
отчетливым морфологическим указателям (маркерам) в каждом плече
выделяют районы, обозначаемые арабскими цифрами. В пределах
районов идентифицируют сегменты — регулярные участки, отличаю­
щиеся по интенсификации окраски. Они также обозначаются арабскими
цифрами. Так, символ \р22 означает 2-й сегмент 2-го района короткого
плеча хромосомы 1.
Дифференциальная окраска позволяет построить цитологические
карты «групп сцепления» с точной локализацией генов и их комплексов
(«пучков») в соответствующей хромосоме. Так, для Х-хромосомы
46
Р и с . 18. К а р т а Х - х р о м о с о м ы ч е л о в е к а .
С п р а в а п о к азан о р асп ол о ж ен ие генов: а
сц епленных с л о к ус а м и Iр уп п ы
крони Х ^ ; б
I
си н те за некоторых ф ер м ентов; д
цветовой слепоты
и гем оф илии А , с л е в а : р
кор о ткое п лечо хромосом ы;
- длинное плечо
хромосом ы, 1,2
сегм енты п лечей хромосом.
0
Ч
человека известны 96 локусов, некоторые из которых картированы.
Имею тся «пучки» сцепленных генов, концентрирующихся вокруг
локусов цветовой слепоты, группы крови Х<у и др. (рис. 18). К настоя­
щему времени установлена хромосомная локализация более 200 генов
.. человека.
3.4. К Л Е Т О Ч Н А Я Т Е О Р И Я
'1
/
К л е т о ч н а я т е о р и я служит фундаментальным обобщени­
ем биологии. Она устанавливает связь всех проявлений жизни на Земле с
клеткой, отмечает универсальность принципов клеточной организации,
определяет клетку одновременно и как целостную самостоятельную
единицу биологической активности, и как составную часть существ
высшего порядка — многоклеточных растительных и животных
организмов.
Клеточная теория сформулирована немецким исследователем —
зоологом Т. Шванном (1_8^9). В своих теоретических построениях
Т. Шванн широко использовал работы ботаника М . Ш лейдена, которо­
го по праву считают соавтором клетонной^Зёоршй Исходя из
предположения об общей природе или гомологичности растительных и
животных клеток, доказываемой одинаковым механизмом их возник­
новения, Т. Шванн обобщил многочисленные данные в виде теории,
согласно которой клетки являю тся структурной и функциональной
основой живых существ. В конце прошлого столетия клеточная теория
получила дальнейшее развитие в трудах патолога Р . Вирхова. Он сделал
важный вывод о том, что клетки образуются лишь путем деления предсуществующих клеток. И м также создана вызвавшая критику
концепция «клеточного государства». Согласно последней многокле­
точный организм состоит из относительно самостоятельных единиц
(клеток), поставленных в своей жизнедеятельности в тесную зависи­
мость друг от друга. Указанная концепция принижала значение
интеграции клеток в единую систему организма. Она не воспринимала
организм как целостность.
Клеточная теория в современном виде включает тпи главных
положения. П е р в о е п о л о ж е н и е соотносит клетку с живой
природой планеты в целом. Оно утверждает, что жизнь, какие бы
сложные или простые формы она ни принимала, в ее структурном,
функциональном и генетическом отношении обеспечивается в конечном
47
итоге только клеткой. Выдающаяся роль клетки как первоисточника
жизни обусловливается тем, что она является биологической единицей
извлечения из внешней среды, превращения и использования организма­
ми энергии и веществ. В клетке сохраняется и используется биологиче­
ская информация.
Согласно в т о р о м у п о л о ж е н и ю в настоящих условиях
новые клетки возникают только путем деления предсуществуюших
клеток. В обосновании клеточной природы жизни на Земле тезису о
единообразии путей возникновения клеток принадлежит особое место.
Он был использован М . Шлейденом и Т. Шванном для доказательства
гомологии разных типов клеток’. Современная биология расширила
круг доказательств этому положению. Независимо от индивидуальных
морфофункциональных особенностей все клетки одинаковым образом:
1) хранят биологическую информацию; 2) редуплицируют наследствен­
ный материал с целью его передачи в ряду поколений; 3) используют
информацию для осуществления своих функций на основе синтеза
определенных белков-ферментов; 4) хранят и переносят энергию;
5) превращают энергию в работу; 6) регулируют обмен веществ.
Т р е т ь е п о л о ж е н и е клеточной теории соотносит клетку с
многоклеточными формами, для которых характерен принцип цело­
стности и системной организации. Для системы свойственно наличие
новых качеств, благодаря взаимному влиянию и взаимодействию
единиц, составляющих эту систему. Структурно-функциональными
единицами многоклеточных существ являются клетки. Вместе с тем
многоклеточный организм характеризуется рядом специфических
свойств, которые нельзя свести к свойствам и качествам отдельной
клетки. В третьем положении клеточной теории мы встречаемся с уже
знакомой проблемой соотношения части и целого, редукционизма и
интегратизма. Системный подход как научное направление использу­
ется в биологических исследованиях с начала текущего столетия.
1
3.5. Э В О Л Ю Ц И Я
КЛЕТКИ
, I Прокариотические клетки появились на Земле ранее эукариотиче1 ских. По данным палеонтологии первые существовали 3,0—
3,5 млрд. лет тому назад, тогда как ископаемые останки вторых
встречаются в породах, датируемых не ранее чем 1,0— 1,4 млрд. лет.
Э то обстоятельство, а также сходство основных биохимических
процессов заставляет думать о происхождении эукариотической клетки
от прокариотической предковой формы.
Согласно с и м б и о т и ч е с к о й г и п о т е з е (рис. 19), попу­
лярной в настоящее время, корпускулярные органеллы эукариотической
клетки, имеющие собственный геном, характеризуются независимым
происхождением и ведут начало от прокариотических клеток-симби1 Авторы клеточной теории, утверждая верное положение о единообразии пути
возникновения клеток, непосредственный механизм их образования представляли
неверно. М., Шлейден считал, что молодые клетки возникают путем конденсации сли­
зистого вещества первоначально в ядро с дальнейшим наслоением и отграничением
цитоплазмы. Т., Ш ванн разделял эту точку зрения.
48
Рис. 19. Происхождение эукариотической клетки согласно симбиотической
инвагинационной ( I I ) гипотезам.
(I)
и
а — Д Н К прокар и оти ческой к л е т ки ; б — ядро э ук ар и о ти ч е ск о й к л е т ки ; в — митохонд рия; г — хлоропласт;
д — ж г у т и к ; 1 — ан аэр об н ы й прокар и от ( к л е т к а - х о з я и н ); 2 — аэр об н ы й прокар и от (п р е зум л ти ви а я
м и т о х о н д р и я); 3 — аэр обн ы й прокар и от (кле тка- р о д о н ачал ьн и ц а со гл асн о ги п отезе I I ) ; 4 — сине-зеленая
водо ро сль (п р е зум п ти вн ы й х л о р о п л а с т); 5 — сп и рохетоподобная б а к те р и я (п р е зум п ти вн ы й ж г у т и к ); 6 —
п р окар и оты , имеющ ие митохондрии; 7 — прим ити вн ы й э ук ар и о т со ж гу ти к о м ; 8 — ж и в о т н а я к л е т к а со
ж г у ти к о м ; 9 — р аст и т е л ь н а я к л е т к а ; 10 — инваги н ац ии клеточн ой оболо чки, д авш и е ядро и митохондрии;
11 — прим ити вн ы й эук ар и о т; 12 — вв о р а чи ва н и е клеточн ой об ол о чки, д а в ш е е х л ор о п ласт; 13 — р а ст и т е л ь ­
н а я к л е тка .
онтов. Предположительно клеткой-хозяином служил анаэробный
прокариот, способный лишь к амебоидному движению. Митохондрии и
жгутики произошли путем изменений проникших в клетку-хозяина
симбионтов— аэробных прокариотов и бактерий, напоминающих
современных спирохет. Хлоропласты клеток зеленых растений возник­
ли из симбионтов — прокариотических клеток Сйне-дедецш водо­
рослей. Особое зг*»" 1■
и**п ;, имелр ^приобретение кде
оцессе
1г
49
эволюции жгутиков с базальными тельцами, близкими к центриолям.
Это позволило интенсифицировать двигательную активность и создать
механизм митоза.
Трудным является вопрос о происхождении ядра. Первоначально
объем информации в геномах клетки-хозяина, с одной стороны, и
симбионтов — презумптивных1 митохондрий, центриолей и хлоропластов, с другой, был, по-видимому, сопоставим. В дальнейшем могла
произойти утрата геномами симбионтов части генетических функций с
перемещением блоков генов в геном клетки-хозяина. Э то сопровожда­
лось ростом объекта ядерного генома, централизацией генетической
регуляции функций клетки и потерей митохондриями и другими
корпускулярными органеллами их автономности. Нельзя однако
исключить, что ядро также представляет собой производное внутрикле­
точного симбионта. Происхождение внутриплазматических мембран
шероховатой и гладкой цитоплазматической сети, пластинчатого
комплекса и везикуло-вакуолярных структур объясняют эволюционны­
ми преобразованиями наружной мембраны ядерной оболочки. Симбио­
тическая гипотеза не объясняет ряд фактов. Так, белок бациллин, из
которого состоят реснички и жгутики современных прокариот, отличен
от белка тубулина эукариот. У бактерий не обнаружено структур с
типичным для жгутиков, ресничек, базальных телец или центриолей
эукариотической клетки составом микротрубочек «9 + 2» или «9 + 0».
Инвагинационная
гипотеза
(см. рис. 19) про­
исхождения эукариотической клетки исходит из того, что предковый
формой был аэробный прокариот. Он содержал несколько геномов,
каждый из которых прикреплялся к клеточной оболочке. Корпуску­
лярные органеллы, имеющие Д Н К , а также ядро, возникли в результате
впячивания и отшнуровки фрагментов оболочки вместе с геномом с
последующей функциональной специализацией в ядро, митохондрии,
хлоропласты, усложнением ядерного генома, развитием цитоплазмати­
ческих мембран. Э т а гипотеза удовлетворительно объясняет наличие
2 мембран в оболочке ядра, митохондрий и хлоропластов. Она
встречается с трудностями в объяснении различий в деталях процессе
биосинтеза белка в корпускулярных органеллах и цитоплазме эукарио­
тической клетки. В митохондриях и хлоропластах этот процесс в
точности соответствует таковому в современных прокариотических
клетках. Симбиотическая и инвагинационная гипотезы не исчерпывают
все точки зрения на происхождение эукариотического типа клеточной
организации.
Эволюционные возможности эукариотического типа клеточной
организации превосходят таковые у прокариот. Указанную особенность
объясняют прежде всего увеличением размеров ядерного генома.
Количество генов в геноме бактерии и клетки человека соотносится как
1:100— 1000. Известное значение мргли иметь наличие в клетке двух
аллелей каждого гена (диплоидность), многократное повторение
некоторых генов. Все это расширяет масштабы мутационной изменчи-
1 О т лат. ргаекитрПо — предположение, наметка.
. 50
вости, совместимой с выживанием, и способствует образованию
резерва наследственной изменчивости.
При переходе к эукариотическому типу усложнился также механизм
регуляции жизнедеятельности клетки. Н а уровне генетического матери­
ала это проявилось в увеличении относительного числа регуляторных
генов, замене единичных «голых» молекул Д Н К ' множественными
хромосомами, в которых Д Н К комплексируется с белками. В итоге
оказалось возможным считывать информацию по частям с разных
блоков генов в разных клетках и в разное время. В бактериальной клет­
ке, например, одновременно транскрибируется 80— 100% информации
генома. В клетках человека в зависимости от органа считывается от
8— 10% (печень, почка, селезенка) до 44% (головной мозг). Использова­
нию информации по частям принадлежит исключительная роль в
образовании многоклеточных живых форм. Определенное значение для
перехода к многоклеточности имело, по-видимому, и приобретение
клетками эукариотического типа эластичной оболочки, что позволяет
образовывать стабильные комплексы клеток. В условиях усложнения
генетического аппарата эукариот важным эволюционным приобретени­
ем стал митоз как механизм воспроизведения в поколениях генетически
сходных клеток. Возникший путем эволюции митоза мейотический
процесс интенсифицировал комбинативную изменчивость. Благодаря
отмеченным особенностям за, 1 млрд. лет своего существования
эукариотическая клетка обеспечила развитие разнообразных живых
форм от простейших до млекопитающих и человека.
Глава
4
РАЗМНОЖЕНИЕ
Продолжительность жизни особи меньше продолжительности
существования вида, к которому она принадлежит. Поэтому история
видов — это история сменяющихся поколений организмов. Очередное
(дочернее) поколение образуется в результате размножения особей
предшествующего (родительского, материнского) поколения. Спо­
собность к размножению является неотъемлемым свойством живых
форк/!1С его помощью сохраняются во времени биологические виды и
жизнь как таковая. Различия, закономерно наблюдаемые в фенотипах
особей разных поколений, делают возможным естественный отбор и
видообразование. Размножение возникло в ходе исторического разви­
тия органического мира вместе с клеткой. В процессе биологического
размножения решение задачи увеличения числа особей сочетается с
обеспечением развития в каждом поколении организмов определенного
вида, т. е. воспроизведением себе подобных^Последнее зависит от
того, что в процессе размножения происходит передача в ряду
поколений наследственного материала (Д Н К ), т. е. определенной по
объему и содержанию биологической информации.
1 Учитывая особенности структурной организации, для обозначения наследст­
венного материала прокариот предложен специальный термин «генофор».
4.1. К Л А С С И Ф И К А Ц И Я С П О С О Б О В Р А З М Н О Ж Е Н И Я
Способы размножения разнообразны. По
классификации
выделяют
бесполое
р а з м н о ж е н и е (табл. 5).
Таблица
5.
О бщ ая
широко принятой
и
половое
характеристика бесполого и полового размножения
С п особ р азм н о ж ен и я
П о к а за т е л и
Обычно две особи
Одна особь
Родители
Родители продуцируют половые
Клеточные источни­ Многоклеточные:
одна или несколько сомати­
ки
наследствен­
клетки (гаметы), специализи­
ческих (телесных) клеток
ного
материала
рованные к выполнению функ­
для развития по
родителя;
ции размножения. К аж д ы й ро­
томна
дитель представлен в потомке
простейшие:
исходно одной клеткой
клетка-организм как це­
лое
Потомство
Генетически точные копии ро­ Генетически отличны от к а ж ­
дого из родителей
дителей, т. е. составляют
клон организмов (при от­
сутствии соматических м у­
таций)
Основной
клеточ­ М итоз
Мейоз
ный
механизм
развития
Эволюционное зна
чение
Усиливает роль стабилизи­ Способствует генетическому раз­
нообразию особей вида, созда­
рующей функции естествен­
ет предпосылки к освоению
ного отбора, способствует
разнообразных условий обита­
сохранению
наибольшей
ния, дает эволюдионные пер­
приспособленности в неме
спективы
няющихся условиях обита­
ния
Типичные формы бесполого размножения приведены на схеме 1.
Деление
надвое
приводит к возникновению из одного
родительского организма двух дочерних и преобладает у прокариот и
простейших, но встречается и у многоклеточных: п р о д о л ь н о е у
медуз, п о п е р е ч н о е
у кольчецов. М н о ж е с т в е н н о е
д е л е н и е ( ш и з о г о н и я ) одноклеточных характерно для некото­
рых паразитов человека (малярийный плазмодий). При размножении
п о ч к о в а н и е м потомок формируется первоначально как вырост
на теле родителя. Ф р а г м е н т а ц и я заключается в распаде тела
многоклеточного организма на части, которые далее превращаются в
полноценные особи. Она описана у плоских червей, иглокожих. У видов,
размножающихся с помощью спор, дочерний организм развивается из
специализированной клетки — споры, которая содержит наслед­
ственный материал одной родительской особи. При бесполом размно­
жении дочерняя особь возникает из одной клетки (деление, шизогония,
спорообразование) или группы клеток
материнского организма.
52
С х ем а
1
Бесполое размножение
Группой клеток
(полицитогенное вегетативное)
Одной клеткой
(моноцитогенное)
I
Деление ро­
дителя надвое
Образование
Множественное деспор
ление родителя (шизо­
гония)
Почкование
или нерав­
номерное
деление
Прокариоты,
одноклеточ­
ные эукариоты
(саркодовые)
Одноклеточ­
ные эукариоты
(жгутиконос­
цы. споровики)
1
Однокле­
точные эука­
риоты (со­
сущие ин­
фузории)
Среди мно­
гоклеточ­
ных осо­
бенно рас­
тения
I----
■Образование
почек, стеб­
левых и кор­
невых клуб­
ней, луковиц
Растения
I
1
Упорядочен­
ное деление
Неупоря­
доченное
деление
(фрагмен­
тация)
Почкова-
В эмбрио­
нальном раз­
витии (полиэмбриония)
Радиально­
симметрич­
ное — медузы,
морские звез­
ды (продоль- •
ное), кольча­
тые черви (по­
перечное)
Ресничные,
ленточные
черви
Губки, ки­
шечнопо­
лостные,
кольчатые
черви
Некоторые
млекопитаю­
щие (броне­
носцы);
встречается
у человека
В последнем случае размножение называют в е г е т а т и в н ы м .
Оно распространено среди растений. Бесполое размножение встреча­
ется у животных на сравнительно низких уровнях структурно­
физиологической организации, к которым принадлежат многие парази­
ты человека. У паразитирующих форм бесполое размножение не только
обусловливает увеличение численности вида, но и является средством
расселения, помогает пережить неблагоприятные условия.
В процессе развития жизни бесполое размножение возникло раньше
полового, которое существует уже более 3 млрд. лет, и обнаруживается
в жизненных циклах всех основных групп организмов] Универсальность
полового размножения объясняется тем, что юно обеспечивает
значительное генетическое разнообразие и, следовательно, фенотипиче­
скую изменчивость потомства. Эти м создаются большие эволюци­
онные и экологические перспективы. Предполагают, что в процессе
эволюции половому размножению предшествовало размножение спо­
рами.
и цГЬснове полового размножения лежит п о л о в о й п р о ц е с с ,
суть которого сводится к объединению в наследственном материале для
развития потомка генетической информации из двух разных источни­
ков — родителей. Представление о половом процессе дает процесс
к о н ъ ю г а ц и и инфузорий. Он заключается во временном соедине­
нии двух особей с целью обмена (рекомбинации) наследственным
материалом, в результате чего появляются особи,, генетически
отличные от родительских организмов. В дальнейшем они осуществля­
ю т бесполое размножение. Поскольку количество инфузорий в ре­
зультате конъюгации остается неизменным, говорить о размножении в
прямом смысле нет основания. У Ргоюгоа половой процесс осуществля­
ется тагже в виде к о п у л я ц и и которая заключается в слиянии
двух особей в одну, объединении и рекомбинации наследственного
материала. На определенном этапе эволюции у многоклеточных
организмов половой процесс как способ обмена генетической информа­
цией между особями в пределах вида оказался связанным с размножени­
ем.
Для осуществления полового размножения особи родителей
вырабатывают г а м е т ы — клетки, специализированные к обеспече­
нию генеративной функции. Слияние материнской и отцовской гамет
приводит к возникновению з и г о т ы — клетки, представляющей
собой дочернюю особь на наиболее ранней стадии развития.
У некоторых организмов зигота образуется в результате
объединения гамет, морфологически неотличимых. В таких случаях
говорят об и з о г а м и и . У большинства видов по структурным и
функциональным
признакам
половые
клетки
делятся
на
м а т е р и н с к и е ( я й ц е к л е т к и ) и о т ц о в с к и е (спер­
м а т о з о и д ы ). К ак правило, яйцеклетки и сперматозоиды
вырабатываются разными организмами — женскими и мужскими,
самками и самцами. Они отличаются друг от друга по ряду признаков.
В подразделении гамет на яйцеклетки и сперматозоиды, а особей на
самок
и
самцов
заключается
явление
полового
д и м о р ф и з м а (рис. 20, 21). Наличие его в природе отражает
54
с
\
Рис.20. Половой диморфизм на уровне половых клеток.
л
2
яй ц е к л е т к а
I
ц и то п л азм а; 2
ш е й ка ; 3
головка
ядро, 3
хроматин ядра
б
сп ер м атозо иды
!
ж гути к
различия в задачах, решаемых в процессе полового размножения
мужской или женской гаметой, самцом или самкой.
Образование гамет обоих видов в одном организме, имеющем и
мужскую
и
женскую
половые
железы,
называется
г е р м а ф р о д и т и з м о м ' . Г ермафродитизм характерен для неко­
торых паразитов человека, например плоских и ленточных червей.
Несмотря на продукцию гермафродитом и мужских и женских гамет,
самооплодотворение для них не типично, что связано в несо- адении
времени созревания яйцеклеток и сперматозоидов. Истинный гермаф
родитизм описан у человека. Чаще он развивается в результате
нарушения эмбриогенеза при одинаковом наборе половых хромосом —
X X или X V во всех соматических клетках. У некоторых людей
гермафродитов обнаружен мозаицизм по половым хромосомам. Одни
соматические клетки имеют пару X X , другие — Х У (см. табл. 12).
Хотя оплодотворение представляет характерный признак полового
размножения| дочерний организм иногда развивается из неоплодотворенной яйцеклетки. Э то явление называется п а р т е н о г е н е з о м
Источником наследственного материала для развития потомка в этом
случае обычно служит Д Н К яйцеклетки (гиногенез). Реже наблюдается
андрогене!— развитие потомка из клетки с цитоплазмой ооцига и
ядром сперматозоида. Ядро женской гаметы в случае андрогеыеза
погибает. Партеногенез представляет модификацию полового размно­
жения в процессе эволюции определенных видов животных. У пчел,
например, он используется как механизм генотипического определения
пола: женские особи (рабочие пчелы и царицы) развиваются из
'О т
истинного
гермафродитизма,
о котором здесь идет речь,
следует отличать г е р м а ф р о д и т и з м л о ж н ы й , заключающийся в сочетании
в одной особи наружных половых органов и вторичных половых признаков обоих
полов при наличии половых желез одного типа
мужского или женского.
55
Рис. 21. Половой диморфизм у людей на организменном уровне.
Х а р а к т е р н ы р а зл и ч и я по: 1 — к ар и о ти п у и гл ав н о м у п оловом у горм ону; 2 — ст р ук ту р е волос и х а р а кте р у
овол о сен и я; 3 — стр оени ю го р тан и ; 4 — р азв и ти ю м олочны х ж е л е з ; 5 — р азв и ти ю м ус к у л а т у р ы ; 6 —
строению половых органов: 7 — распределению ж ир о во й тк а и и ; 8 — п о к а за т е л ям роста длинных т р у б ч а ­
ты х костей.
оплодотворенных яйцеклеток, а мужские (трутни) — партеногенетически. Партеногенез включен в жизненные циклы многих паразитов. Он
обеспечивает рост численности особей в условиях, затрудняющих
встречу партнера противоположного пола. Имею тся указания на
возможность девственного развития у человека. В яичниках девушек,
56
погибших при случайных обстоятельствах, в отсутствии предшествую­
щего осеменения находили зародыши на ранних этапах дробления.
Наблюдения завершенного эмбриогенеза с партеногенетическим разви­
тием в отношении человека отсутствую т. При партеногенезе,/как и
типичном половом размножении, развиваю тся особи с диплоидными
соматическими клетками. Восстановление диплоидного набора хромо­
сом происходит обычно путем слияния ооцита и редукционного тельца
во втором делении мейоза.
\
У некоторых видов закономерно наблюдается п о л и э м бр и о н и я — бесполое размножение зародыша на ранних ста­
диях эмбриогенеза животных, размножающихся половым способом.
Полиэмбриония типична для броненосцев и заклю чается в разделении
на стадии бластулы клеточного материала первоначально 1 зародыша
между 4— 8 зародышами, из которых развиваю тся полноценные особи.
В результате полиэмбрионии у человека рождаю тся однояйцевые
близнецы.
4.2. С М Е Н А П О К О Л Е Н И Й
Больш инство организмов, размножающихся бесполым путем,
способно к половому размножению. При этом обычно ряд поколений с
бесполым размножением сменяется поколением особей, размножаю­
щихся с помощью гамет или же осуществляющих половой процесс.
Смена
(чередование)
половых
и
бесполых
п о к о л е н и й происходит у разных видов с разной периодичностью,
регулярно или же через неодинаковые промежутки времени.
П е р в и ч н о е ч е р е д о в а н и е п о к о л е н и й заключается
в смене полового размножения спорообразованием. Оно наблюдается у
представителей классов споровиков, жгутиконосцев, некоторых расте­
ний и отражает сохранение в филогенезе соответствую щ их организмов
как более древней (бесполой), так и более прогрессивной (половой) форм
размножения. В т о р и ч н о е ч е р е д о в а н и е п о к о л е н и й
заклю чается в переходе на некоторых стадиях жизненного цикла к
бесполому или партеногенетическому размножению животных, осво­
ивших половое размножение. Оно распространено у кишечнополос­
тных, членистоногих.
Включение в цикл развития организмов, размножающихся
преимущественно бесполым путем, полового поколения, активизируя
комбинативную изменчивость, способствует преодолению генетическо­
го однообразия потомков, расширяет эволюционные перспективы
группы.
4.3. П О Л О В Ы Е К Л Е Т К И
П о сравнению с другими клетками функция гамет уникальна. Они
обеспечивают передачу наследственной информации между особями
разных поколений, чем сохраняют особенности проявлений жизни во
времени. Длительное время половые и соматические клетки противопо­
ставляли друг другу, наделяя лиш ь первые всей полнотой свойств
жизни, проносимых ими через поколения. В настоящее время экспери­
К л е тк и эпителия кишки
го л овасти ка— источник
-ч. н аслед ственного
Л
материала
Я й ц екл етка с убитым
УФ-лучами ядром —
источник цитоплазмы
I
Рис. 22. Развитие шпорцевой лягуш ки из лишенного ядра яйца, в которое переса­
дили ядро клетки кишечного эпителия.
ментально доказана возможность развития полноценного организма на
основе наследственной информации ядра дифференцированной сомати­
ческой клетки, например кишечного эпителия (рис. 22).
Гаметы
представляют собой одно из многих направлений
дифференцировки/#Зеток развивающегося организма. Они образуют
«клеточную линию», специализированную к выполнению репродуктив­
ной функции. Предполагают, что клетки указанной линии возникают из
бластомеров, имеющих на вегетативном полюсе цитоплазму особого
рода — так называемую з а р о д ы ш е в у ю
п л а з м у ’ , бога­
тую Р Н К .
В сравнении с другими линиями соматических клеток (эпители­
альные, нервные, мышечные) гаметы характеризуются рядом отличий.
Важнейшая из них — г а п л о и д н ы й н а б о р х р о м о с о м в
ядрах, что обеспечивает . воспроизведение в зиготе типичного для
организмов данного вида диплоидного числа хромосом.
Действительно, оплодотворение сперматозоидом яйцеклетки, ядра
которой содержат 23 хромосомы, обусловливает формирование зиготы
с 46 хромосомами, что типично для соматических клеток человека.
Гаметы отличаются нетипичным для других клеток значением
ядерно-цитоплазматического
отношения.
У яйцеклеток оно снижено благодаря увеличенному объему ци­
топлазмы, в которой размешен питательный материал (желток) для
развития зародыша. У сперматозоида благодаря малому количеству
1 Термин «зародышевая плазма» был ранее использован А. Вейсманом, но в
совершенно ином смысле — для обозначения наследственного вещества ядра клетки
(хромосом).
58
V
цитоплазмы ядерно-цитоплазматическое отношение высокое. Э то
находится в соответствии с главной функциональной задачей мужской
гаметы — транспортировкой наследственного материала к яйцеклетке.
Половые клетки отличаются н и з к и м у р о в н е м о б м е н н ы х
п р о ц е с с о в , близким к состоянию анабиоза. Мужские гаметы н е
вступают
в митотический
ц и к л . У яйцеклеток эта
способность восстанавливается при оплодотворении или действии
фактора, активирующего партеногенез.
По ряду признаков женские и мужские гаметы отличаются друг от
друга, что связано с различными функциями яйцеклетки и сперматозои­
да в процессе размножения. Яйцеклетки имеют оболочки, которые
выполняют защитную функцию, обеспечивают требуемый уровень
обмена веществ, препятствуют проникновению в яйцеклетку более
одного спермия, способствуют внедрению (имплантации) зародыша в
стенку матки у плацентарных животных, поддерживают форму
зародыша. Для яйцеклетки характрна ц и т о п л а з м а т и ч е с к а я
с е г р е г а ц и я . После оплодотворения (у асцидий уже через 5 мин) в
еще не дробящемся яйце происходит закономерное перераспределение
цитоплазмы. В дальнейшем цитоплазма разного состава распределя­
ется по клеткам разных тканевых зачатков. По-видимому, способность
бластомеров развиваться в определенном направлении зависит от
наследования ими веществ, концентрирующихся в разных участках
цитоплазмы яйцеклетки.
Сперматозоид имеет а п п а р а т д в и ж е н и я в виде жгутика.
В семенной жидкости мужская гамета человека развивает скорость
до 5 см /ч1. Яйцеклетка, лишенная аппарата активного движения,
преодолевает расстояние до полости матки, равное примерно 10 см, за
4—7 сут. Сперматозоиды некоторых видов животных имеют акросомный аппарат, выбрасывающий при контакте с яйцеклеткой длинную
нить. Он обеспечивает проникновение ядра мужской гаметы в ци­
топлазму яйцеклетки. Описаны и другие приспособления, способствую­
щие оплодотворению.
4.4. Г А М Е Т О Г Е Н Е З
Г а м е т о г е н е з — процесс
образования
яйцеклеток
(о о г е н е з) и сперматозоидов ( с п е р м а т о г е н е з ) — подразде­
ляется на ряд стадий (рис. 23).
• ‘
Диплоидные клетки, из которых образуются гаметы, называются
сперматогониями
и
овогониями.
Стадия
р а з м н о ж е н и я сводится к серии последовательных митотических
делений сперматогоний и овогоний, в результате чего их количество
существенно возрастает. Сперматогонии размножаются на протяжении
всего периода половой зрелости мужской особи. Размножение овогоний
приурочено главным образом к периоду эмбриогенеза. В женском
организме этот процесс наиболее интенсивно протекает между 3-м и 7-м
1 Если учесть соотношение преодолеваемого расстояния и длины движущегося
объекта, то при названной, скорости сперматозоид человека перемещается в 1,5 раза
быстрее, чем пловец олимпийского ранга.
59
Сперм атогонии0
Сперм атоц ит
I п оряд н а
Сперм атоцит
II п о р я д н а
Сперм атид ы
Зрелы е
спер м атозо ид ы
а
б
Рнс. 23. Схема гаметогенеза:
а — сп ер м атоген ез; б — овоген ез, п — ко л и че ство хромосомных набо р ов; с д укц ионны е те л ьц а .
к о л и че ств о Д Н К ; Р Т — ре­
месяцами эмбриогенеза и завершается на 3-м году жизни. Н а
с т а д и и р о с т а происходит увеличение клеточных размеров и
превращение
мужских
и
женских
половых
клеток
в
с п е р м а т о ц и т ы и о в о ц и т ы I п о р я д к а , причем послед­
ние достигают больших размеров, чем первые. Основными событиями
с т а д и и с р з р е в а н и я являю тся два последовательных деле­
ния — р ел_у к ц и _9 н н
и^
н и о н н о -е. представляющее
со б о й " обычный митоз. ПосЬе первого деления образуются
с п е р м а т о ц и т ы и о в о ц и т ы I I п о р я д к а , имеющие реду­
цированное вдвое число хромосом, а после второго — сперматиды и
зрелая яйцеклетка. В результате делений на стадии созревания каждый
сперматоцит I I порядка дает 4 с п е р м а т и д ы , тогда как каждый
овоцит I I порядка — 1 полноценную яйцеклетку и З р е д у к ц и онных
тельца,
которые
в
размножении
не
участ­
вуют.
Благодаря этому в женской
гамете концентрируется
максимальное количество питательного материала желтка. Процесс
сперматогенеза завершается с т а д и е й ф о р м и р о в а н и я или
спермиогенеза.
Ядра сперматид уплотняются вследствие
гиперспирализации хромосом, которые становятся функционально
инертными. Пластинчатый комплекс перемещается к одному из
полюсов ядра, образуя акросому, играющую большую роль в опло­
дотворении. Центриоли занимают место у противоположного полюса
ядра, причем от одной из них отрастает жгутик, у основания которого в
виде спирального чехлика концентрируются митохондрии. Н а этой
стадии почти вся цитоплазма сперматиды отторгается, так что головка
зрелого сперматозоида практически лишена ее.
Центральным событием гаметогенеза является м е й о з. Э то
способ клеточного размножения, обусловливающий возникновение гап­
лоидных клеток. В мейозе наблюдается также перераспределение
(рекомбинация) наследственного материала между гомологичными
60
хромосомами, устанавливаются разные соотношения хромосом отцов­
ского и материнского происхождения в гаплоидных наборах гамет.
Мейоз состоит из двух последовательных делений, которым
предшествует о д н о к р а т н а я р е д у п л и к а ц и я Д Н К . Важ ­
ная особенность профазы первого мейотического деления заключается в
синапсисе
или пространственном объединении гомологичных
хромосом, образовании между ними
хиазм
и
обмене
гомологичными
(взаимосоответствуюшими)
у ч а с т к а м и . Своеобразие анафазы первого деления мейоза состоит
в случайном расхождении гомологов из каждой пары хромосом.
Рассмотрим последовательность событий мейоза в процессе овогенеза у
человека.
^ Цитогенетический результат мейоза, который заключается в
^образовании
гаплоидных
клеток
и
^ ' р е к о м б и н а ц и и н а с л е д с т в е н н о г о м а т е р и а л а , за( висит от особенностей первого деления. В профазе этого деления
выделяют ряд стадий (рис. 24). В л е п т о т е н_е—(а) происходит
спирализация хромосом диплоидного набора ооцита в плотные,
' интенсивно окрашиваемые тельца. К концу названной стадии степень
спирализации снижается. Хромосомы, образующие в совокупности
клубок, имеют вид нитей с утолщениями по длине. В последующей
стадии — з и г о т е н е (б) — гомологичные хромосомы спирализуются, сближаются по длине и образуют пары. Происходит синапсис
хромосом.
Число
пар
равно
гаплоидному — 23.
В стадии
п а х и т е н ы (в) в ооцитах видны попарно расположенные гомоло­
гичные хромосомы или б и в а л е н т ы , причем каждая хромосома
образована двумя хроматидами. Таким образом, комплекс из пары
гомологичных хромосом (бивалент) представлен четырьмя морфологи­
ческими структурами (хроматидами). Поэтому его называют
т е т р а д о й . На этой стадии отмечается дальнейшая спирализация и
особенно тесный контакт между хроматидами. Видимо, в пахитене
осуществляется к р о с с и н г о в е р —
реципрокный
(взаимный)
обмен генетическим материалом между спаренными хромосомами.
В дипл о х ш
(г) т омологичные хромосомы частично деспирали5уются и несколько отходят друг от друга. Вместе с тем они сохраняют
взаимосвязь при помощи мостиков — хиазм, которые служат структур­
ным выражением кроссинговера, имевшего место в предыдущую
стадию (рис. 25).
Особенностью мейоза в оогенезе является наличие специальной
\|. стадии — д и к т и о т е н ы , отсутствующей в сперматогенезе. Н а
этой стадии, достигаемой у человека еще в эмбриогенезе, хромосомы,
• приняв особую морфологическую форму «ламповых щеток»1 , прекращают какие-либо дальнейшие структурные изменения на многие годы.
‘ -По достижении женским организмом репродуктивного возраста под
влиянием лютеинизирующего гормона гипофиза, как правило, один
ооцит ежемесячно возобновляет мейоз. Он проходит завершающую
1 Особая форма хромосом, вы являем ая на известной стадии мейоза и соответ­
ствую щ ая периоду интенсивного синтеза Р Н К на определенных генах.
61
с1
/./ Л / ' / / / У
“/
■>л
Рнс. 25. Бивалент иа стадии диплотены.
з
I
—
хр ом атида; 2 —
хи азм ы ;
3 —- центромеры.
стадию профазы первого деления — д и а к и н е з (см. рис. 24, д). На
этой стадии хромосомы высоко спирализованы, ядерная оболочка и
ядрышко исчезают, образуется веретено деления. В м е т а ф а з е (е)
п е р в о г о д е л е н и я гомологичные хромосомы, сгруппированные
в биваленты, располагаются в плоскости экватора клетки. В этой фазе
может б у т ь определено число хиазм. В среднем в ооците женщины их
насчитывается 42— 50. Существует мнение, что у человека кроссинговер
происходит в оогенезе чаще, чем в сперматогенезе, однако это
предположение
требует
подтверждения.
В а н а ф а з е (ж)
п е р в о г о д е л е н и я к полюсам расходятся не дочерние хроматиды гомологичных хромосом, как в митозе, а непосредственно гомоло­
гичные хромосомы. Таким образом, на полюсах клетки избирается по
одной из гомологичных хромосом каждой пары. Так как они состоят из
двух хроматид, их называют д и а д а м и . В процессе цитотомии
в т е л о ф а з е (з) п е р в о г о
деления
образуется первое
редукционное (направительное, полярное) тельце. Оно содержит пол­
ный гаплоидный комплект хромосом, но погибает. Вторая клетка,
также гаплоидная, продолжает оогенез.
И н т е р ф а з а (и) между двумя делениями редуцирована. Второе
деление мейоза фактически начинается с м е т а ф а з ы (к). Хромосо­
мы на этой стадии оогенеза представлены диадами. Э то деление
завершается после о в у л я ц и и, т. е. выхода ооцита из яичника в
женские половые пути, и для его завершения необходимо, чтобы
произошло
оплодотворение
клетки
сперматозоидом.
В а н а ф а з е (л) в т о р о г о
деления
к полюсам клетки
расходятся дочерние хроматиды, из которых на более ранних стадиях
состоят диады. В последующей т е л о ф а з е (м) отделяется второе
редукционное тельце, которое погибает. Гаплоидные наборы хромосом
яйцеклетки и оплодотворившего ее сперматозоида объединяются в
диплоидном наборе хромосом зиготы.
Таким образом, рекомбинация генетического материала между
гомологичными хромосомами происходит в пахитене, а рекомбинация
отцовских и материнских хромосом — в анафазе первого деления
мейоза. В анафазе этого деления образуются гаплоидные клетки.
Последние стадии оогенеза воспроизводятся и вне организма
женщины в искусственной питательной среде. Э то позволило осуще­
ствить зачатие человека «в пробирке». Перед овуляцией хирургическим
путем яйцо извлекается из яичника и переносится в среду со сперматозо­
идами. Возникающая в результате оплодотворения зигота, будучи
помещена в подходящую среду, осуществляет дробления. Н а стадии
8— 16 бластомеров зародыш переносится в матку женщины-реципи­
ента. Число успешных опытов такого рода в последнее время
возрастает.
В сперматогенезе происходят сходные цитогенетические события —
рекомбинация генетического материала путем кроссинговера и незави­
симого расхождения гомологичных хромосом в анафазе первого
деления, а также образование гаплоидных клеток. Так как сперматоге­
нез начинается в организме, достигшем половой зрелости, закономерно
отсутствие стадии диктиотены. Вместе с тем хорошо выражен период
63
Половозрелые
° с°™
уГ
Диплофаза
,ЧИСЛС
о,
х р о м о с о м 2п)
Гап лоф аза
(ч и с л о
хромосом п)
'
\
^
3 и г0 Т а
Гам етоген ез
Ч
/
\
*
Гаметы
Рис. 26. Ф азы жизненного цикла многоклеточных животных.
ф ормирования, когда м уж ская
половая кл етка приобретает
СВОЮ ОКОНЧаТеЛЬНуЮ струкТУРУ
ГамеТОГвНеЗ
ОТЛИЧаеТСЯ
высокой
производительностью . За время половой жизни
мужчина продуцирует не ме« е е Л З О м л р Д . СПерматОЗОИДОВ.'
Н й ПЯТОМ МССЯЦС ЭМОрИОГСНС-
за в зачатке женской половой
железы насчитывается 6 00(1000 клеток-предшественниц яйцеклеток.
К началу репродуктивного периода в яичниках обнаруживается
примерно 100 000 ооцитов. О т момента полового созревания до
прекращения гаметогенеза в яичниках созревает (500ц—4(Х) ооцитов.
4.5. Ч Е Р Е Д О В А Н И Е Г А П Л О И Д Н О Й
И ДИПЛОИДНОЙ
Ф А ЗЫ Ж И ЗН ЕН Н О ГО Ц И КЛ А
Организмы с половым размножением в результате мейоза образуют
гаплоидные гаметы. В момент оплодотворения благодаря слиянию
двух таких клеток возникает диплоидная зигота. Многократные
митотические деления зиготы и ее потомков приводят к увеличению
числа диплоидных клеток, которые строят тело организма в процессе
его развития. По достижении последним половой зрелости возобновля­
ется процесс образования гаплоидных гамет. Таким образом, в жизнен­
ных циклах организмов, размножающихся половым способом, выделя­
ются 2 фазы — г а п л о и д н а я
и диплоидная
(рис. 26)..
Относительная продолжительность этих фаз варьирует у представите­
лей различных групп живых существ. У простейших и грибов
преобладает первая, у высших растений и животных — вторая фаза.
Удлинение диплофазы в историческом развитии объясняется пре­
имуществами диплоидного состояния перед гаплоидным. Благодаря
гетерозиготности и рецессивности в диплоидном состоянии укрыва­
ю тся от
естественного отбора, сохраняются и накапливаются
разнообразные аллели. Э то повышает объем генетической информации
в генофондах популяций и видов, ведет к образованию резерва
наследственной изменчивости, от которого зависят эволюционные
перспективы. В то же время у гетерозигот вредные рецессивные аллели,
не оказывая влияния на развитие фенотипа, не снижают жизнеспо­
собность.
4.6. П У Т И П Р И О Б Р Е Т Е Н И Я О Р Г А Н И З М А М И
БИ О Л О ГИ ЧЕС КО Й ИНФО РМ АЦ ИИ
Благодаря генетической рекомбинации, которая закономерно
происходит в процессе гаметогенеза и при оплодотворении,
п о л о в о е р а з м н о ж е н и е представляет собой эволюционно
обусловленный механизм обмена наследственной информацией между
организмами одного биологического вида. Некоторые факты из
64
области зоологии и особенно вирусологии и микробиологии указывают
на то, что имеются пути приобретения биологической информации и от
организмов других видов. Э т а информация воспроизводится в фенотипе
организма и обусловливает развитие признаков, незакодированных в
наследственном материале родителей. Так, в клетках пищеварительно­
го дивертикула брюхоногого моллюска (Е1ума ушсНх) сохраняются
хлоропласты поедаемой водоросли (Сосйшп йга^Пе), в результате чего
моллюск приобретает способность к фотосинтезу. Стрекательные
капсулы гидроидных полипов, которые поедаются некоторыми турбелляриями, не перевариваются, а перемещаются в эпителиальный пласт и
используются червем в качестве орудия защиты. В классической
зоологии такие примеры получили название к л е п т о г е н е з а или
эволюции путем воровства.
Явление т р а н с д у к ц и и заключается в том, что в генетиче­
ский материал клетки-хозяина (бактериальной или эукариотической)
встраивается нуклеиновая кислота вируса с фрагментом генома другой
клетки. Превносимая таким путем биологическая информация вслед­
ствие редупликации чужеродной Д Н К может передаваться в ряду
клеточных поколений, а также воздействовать на состояние генетиче­
ской системы клетки-хозяина, изменяя, например, частоту му­
тирования отдельных генов. Чужеродная Д Н К может присутство­
вать в клетке в виде . плазмид и эписом-фрагментов нуклеи­
новой кислоты, лишенных, в отличие от Вирусных частиц,
белковых чехлов. П л а з м и д ы .самостоятельны по отношению к
хромосомам клетки-хозяина, а э п и с о м ы могут встраиваться в них.
Биологическая информация плазмид и эписом, проявляясь в фенотипе,
дает широкий круг признаков, включая устойчивость к антибиотикам.
Примеры проникновения в организм действующей биологической
информации брганизмов из других таксонов, прежде всего вирусов,
описаны у высших животных и человека. Так, сотрудники, длительно
работавшие в онкологических лабораториях с вирусной опухолью
кроликов — папилломой Шопа, имеют, как правило, пониженное
содержание в плазме крови аминокислоты аргинина. Объясняется это
тем, что вирус папилломы, которым «заражены» такие люди, несет ген
синтеза аргиназы, катализирующей обмен аргинина. Будучи активным,
этот ген обусловливает образование дополнительных молекул фермен­
та. Описанный факт относится к широкой области так называемого
«вирусоносительства» или «латентных вирусных инфекций». Вирус,
присутствуя в клетках организма длительное время и не вызывая
собственно патологических изменений, приводит к развитию некоторых
фенотипических признаков. У человека известен генетический дефект,
который проявляется в серьезных нарушениях развития и зависит от
недостатка аргиназы. Открытие способа контролируемого «зараже­
ния» таких людей вирусом папилломы Шопа могло бы нормализовать
их фенотип.
Половой процесс служит универсальным механизмом обмена
генетической информацией между особями в пределах вида. Рассмот­
ренные факты свидетельствуют о том, что количество и содержание
информации, используемой различными организмами для развития и
§1
ж изнедеятельности, не всегда ограничивается то й информацией,
которая б ы л а ими приобретена о т родителей. С ущ е ств у ю т дополни­
тельные п ути приобретения биологической информации. Видоспеци­
фичная информация, получаемая в процессе размножения, обеспечивает
развитие особей определенного морфофизиологического типа. Д о ­
полнительная информация по биологическому содержанию в значи­
тельной степени случайна и нередко нарушает реализацию собственной
генетической информации хозяина. В связи с эти м в эволюции возникли
м е х а н и з м ы
з а щ и т ы
от
п р о н и к н о в е н и я
чу­
ж ер о д н о го
н а с л е д с т в е н н о г о
м а т е р н а л а. П р и ­
мером м о ж е т с л у ж и ть интерферон — белок, вы р аб аты ваем ы й клетка­
ми млекопитаю щ их и птнц в о тв ет иа внедрение вирусов и представляю ­
щий собой фактор неспецифического противовирусного им м унитета.
Г л а в а
5
ГЕН ЕТ И КА
5.1.
В В Е Д Е Н И Е . В ГЕ Н ЕТ И К У
I
В последовательных поколениях возникаю т особи, подобные друг
I другу по морфологическим, физиологическим, биологическим и другим
I признакам, ч то обусловливается
н а с л е д с т в е н н о с т ь ю —
| фундаментальной характеристикой ж ивых форм, под которой понима­
ю т их свойство обеспечивать стр уктур н ую и функциональную преем­
ственность между поколениями. П оскольку структурны е, функцио­
нальные н иные признаки организма ф ормирую тся в процессе
онтогенеза, наследственность является та к ж е механизмом передачи в
ряду поколений способности к процессу индивидуального развития,
типичному для особей конкретного биологического вида. К а ж д ы й вид
организмов сохраняет в ряду поколений характерные черты строения и
физиологии: утка вы во д и т у т я т , пшеница воспроизводит пшеницу,
человек рождает человека. Особо важ ная ро ль в обеспечении свойства
структурно-функциональной преемственности м еж д у поколениями при­
надлежит хромосомам. С оответственно это м у в качестве главной
формы вы д ел яю т хром осомную или ядерную наследственность. Пере­
дача-потомку некоторых признаков и свойств происходит при помощи
наследственных задатков цитоплазм ы . Э т о дает право говорить о
цитоплазматической или внеядерной наследственности. Принципи­
альных различий м ежду механизмами хромосомной и цитоплазм атиче­
ской наследственности не сущ ествует — обе формы о сновы ваю тся на
передаче в ряду поколений дискретных единиц наследственности генов.
О рганизм ы дочернего и родительского поколений, как правило, не
бываю т
то ч н ы м и
копиями
друг
друга
вследствие
и зI м е н ч и в о с т и , которая, как и наследственность, с л у ж и т фунда­
ментальной характеристикой живого. И зм ен чи вость проявляется в из­
менении от особи к особи нли м ежду особями разных поколений наслед­
ственных задатков (генов), их сочетаний, индивидуального развития.
Закономерности няспепстненнпстн и изменчивости, биологические
механизмы, их обеспечивающие, изучает г е н е т и Т Г а .
66
Рнс. 27. И зображ ение на печатке записи о разведении лош адей в Д вур ечье 6000 лет
назад
Н аследственность и изм енчивость тесно связаны с размножением и
индивидуальным развитием и с л у ж а т необходимыми предпосылками
процесса эволюции. Благодаря изменчивости сущ ествует разнообразие
ж ивых форм и, следовательно, возм о ж н ость освоения различных сред
обитания, «всю д ность ж изни» (В. И . Вернадский). Наследственность
сохраняет эволюционный о п ы т биологического вида в поколениях.
П ер вые генетичегкнг лррад.тавплния формировались в _связи с. _
сельскохозяйственной и медицинской деятельностью людей. Историче­
ские докум енты свидстельствую т7‘чТ0 у ж е 6000 л е т назад в ж и вотн о­
водстве составлялись родословные (рис. 27). Наблю дения о наследуе­
м ы х патологических признаках, например повыш енной кровоточивости
у лип м уж ского пола, о траж ены в религиозных документах, в частно­
сти, в Т алм уде (4— 5 век до н. э.). Развитие племенного дела и семено­
водства во второй половине Х Т Х века, опубликование Ч . Д арвином
учения о происхождении видов стали стим улам и к изучению явлений
наследственности и изменчивости. Соверш ается ряд о ткр ы ти й , за­
нявших в последующем важ ное место в системе генетических знаний.
Так, о п и е ^ а в т СЯ непрямпр_^гтеленне соматических клеток, обращается
внимание на особые ядерные стр уктур ы хром осомы , устанавливается
постоянство их числа и индивидуальных морфологических черт в
клетках- организмов одного вида; ~а та кж е редукция числа хромосом
вдвое при образовании половых клеток. В ]855Т-. Г . Мендель олу&лико-__
вал работу « О п ы т ы нал растительны м и ^ ибрилами», в которой
сформулировал ряд ф ундаментальных генетических законов.
“ Официальной датой рождения генетики счи та ю т ПХХТТ., когда б ы л и
3*
67
Г Мендель (1822
18841
Т . Г , М орган (1866
1945)
опубликованы данные Г . де Ф риза,
К . Корренса и К . Черм ака, фактиче­
ски переоткрывших закономерности
наследования признаков, установлен­
ные Г . Менделем, и сделавшие их до­
стоянием пауки. П ервы е десятилетия
X X века оказались исключительно
плодотворны м и в развитии основных
положений и направлений генетики.
Б ы л о сформулировано представление
о мутациях (Г . де Ф ри з), популяциях
и чи сты х линиях организмов (В . Иогаиисен), хромосомная теория наслед­
ственности (Т . Г . М орган), о тк р ы т
закон гомологичных рядов (Н . И . В а ­
вилов), получены данные о то м , ч то
рентгеновские лучи в ы з ы в а ю т наслед­
ственные изменения ( Г . С . Филиппов,
Г . М еллер). В ы сказы вается предполо­
Н . И . В ав и л о в (1887 1943)
жение о то м , ч т о химическую основу
гена со ставл яю т биологические макро­
№
молекулы (Н . К . Ко льц ов), указы вается на связь между генами и
ферментами (А Гаррод). Б ы л а н ачата разработка основ генетики
популяций организмов (Г . Гарди, В . Вайиберг, С . С . Четвериков).
Устанавливается наследственная природа и проводится клинический
анализ некоторых наследственных заболеваний. Разр аб аты вается
методика
медико-генетического
консультирования
населения
(С . Н . Давиденков).
68
Н. К
Кольцов (1872- 1940)
С С Ч етвериков (1880
1959)
Реш аю щ ее значение для развития
генетики на настоящем этапе и м ею т
о ткр ы ти е «вещ ества наследственно­
ст и » — Д Н К , расшифровка биологи­
ческого кода, описание механизма
биосинтеза белка.
5.1.1.
О с н о в н ы е направления
ге н е т и к и че л о века
Исторически интерес медицины к
генетике формировался первоначально
в связи с наблюдениями за наследуе­
м ы м и патологическими признаками.
В о в т о р о й половине X I X века английский биолог Ф . Га л ь то н выделил -Н ас л е д с т в е н н о с т ь
человек а как сам остоятельны й предмет ис­
следования. О н Ж е предложил - ряд
Специальных методов генетического
С . Н 1акиденкоь (1880 1961)
анализа — генеалогический, б л изнецовый,
статистический.
Изучение
закономерностей
наследования
норМалБных-тгтгатологических признаков и сейчас занимает ведущее
место в генетике человека. П ри этом предметом непосредственного
изучения с л у ж а т как качественные (дискретные), так н количественные
показатели организма. Д олгое время маркерами в исследованиях
генетических закономерностей б ы л и морфологические или клинические
признаки. Обнаружение взаимосвязи м ежду генами и ферментами
А
привело к созданию б и о х и м и ч е с к о й и м о л е к у л я р н о й
г е н е т и к и . Тот факт, что белок является первичным продуктом
функциональной активности гена, обусловило интенсивное развитие
этого направления в наши дни. Аналогично объясняется прогресс
и м м у н о г е н е т и к и , изучающей генетические основы иммунных
реакций организма человека. Выяснение первичного биохимического
нарушения, приводящего через ряд промежуточных этапов к наслед­
ственному заболеванию, облегчает поиск путей коррекции соответству­
ющих клинических проявлений. Так, заболевание фенилкетонурия,
обусловленное недостаточным синтезом фермента фенилаланингидроксипазы и, следовательно, нарушенным обменом аминокислоты фенила­
ланина, поддается лечению, если из пищи исклю чить эту амино­
кислоту.
Наряду с н а с л е д с т в е н н ы м и б о л е з н я м и выявлены
заболевания
с
наследственным
предрас­
положением
(сахарный
диабет,
язвенная
и
гиперто­
ническая болезни, некоторые формы психических болезней). Изучение
соотносительной роли генетических факторов и факторов среды в
развитии заболеваний с наследственным предрасположением пред­
ставляет собой один из ведущих разделов м е д и ц и н с к о й
г е н е т и к и 1.
Наследственные болезни и заболевания с генетической предрасполо­
женностью зависят от наличия неблагоприятных аллелей генов или их
сочетаний. П о п у л я ц и о н н а я г е н е т и к а изучает распределе­
ние аллелей отдельных генов в группах людей, закономерности
изменения этого распределения во времени и по территории, причины
неравномерного распределения аллелей. Э то позволяет прогнозировать
число некоторых наследственных заболеваний в поколениях и целе­
направленно планировать медицинские мероприятия. Так, аллель,
обусловливающий развитие фенилкетонурии, о которой шла речь выше,
встречается существенно чаще в генотипах ирландцев и шотландцев,
чем англичан. При этом предки многих семей, например из юговосточной Англии, в которых наблюдаются больные фенилкетону­
рией, — также выходцы из Ирландии й Шотландии. Больш ая или
меньшая заболеваемость болезнью с наследственным предрасположе­
нием (степень риска) в различных группах людей также может зависеть
от неравномерного распределения в популяциях тех или иных аллелей.
К развитию ишемической болезни сердца предрасполагает, например,
повышенное содержание в крови холестерина. Э то т признак контроли­
руется доминантным аллелем определенного гена. У лиц с названным
аллелем рано (до 30 лет) появляю тся приступы стенокардии, к 50-ти
годам у них развивается ишемическая болезнь сердца и около половины
подобных больных к 60-ти годам погибает.
1 Медицинская генетика — частный раздел генетики человека (антропогене­
ти ка), изучающий закономерности развития наследственных заболеваний, а такж е
разрабатывающий методы их диагностики и коррекции. Т ак как генетические меха­
низмы наследственных болезней и нормальных свойств одинаковы, ряд авторов счи
тают выделение этого раздела условным (М . Е. Л обаш ев).
70
В эукариотических клетках гены распределены между хромосомами.
Разработка методов хромосомного анализа, изучение структурно­
функциональных характеристик хромосом, их картирование по присут­
ствию генов, выяснение роли отдельных хромосом в индивидуальном
развитии составляю т задачи ц и т о г е н е т и к и . Достижения цито­
генетики
использую тся
для
диагностики
и
изучения
хромосомных
б о л е з н е й , которые представляют собой
пороки развития вследствие изменений в клетках числа хромосомных
наборов, количества хромосом или их структуры. Табл. 6 дает
представление о зависимости частоты самопроизвольных выкидышей и
мертворождений от хромосомных изменений.
Таблица
6. Частота хромосомных аномалий среди спонтанных абортов,
мертворожденных и живорожденных
Х р ом осо м н ы е аном алии
Г руппа
Спонтанные аборты
Мертворожденные
Ж и ворожден ные
Ч и сл о обследованны х
аб с. чи сл о
%
4 209
1775
42,2
283
17
6,0
28 828
202
0,7
Изменения в генетическом материале могут возникнуть под
воздействием факторов окружающей среды, которые необходимо
исследовать на мутагенное действие. Важными разделами этого
направления являю тся р а д и а ц и о н н а я г е н е т и к а и исследо­
вания по мутагенному действию химических соединений. Значение
последнего раздела видно из того факта, что около 10% веществ,
ежегодно поступающих в обращение в виде средств борьбы с насекомы­
ми, промышленных соединений, лекарств, пищевых добавок, способны
вы зы вать в клетках человека мутации.
Индивидуальные и групповые особенности реакций людей,
различающихся генетически, на терапевтические воздействия изучает
ф а р м а к о г е н е т и к а . Она вскрывает наследственные факторы
изменчивости, эффективности и выраженности побочных действий
лекарств у разных лиц.
Наряду с проблемами генетики человека существенный вопрос для
медицины
представляют
генетические
вопросы
биологии
возбудителей
инфекционных
з а б о л е в а н и й — вирусов, бактерий. Важное место занимают
проблемы изменчивости и разнообразия ш таммов (разновидностей)
возбудителей, выработки иммунитета, устойчивости к антибиотикам и
другим лекарственным препаратам. Изучение генетики микроорга­
низмов имеет большое значение в микробиологической промышленно­
сти для организации производства биохимических продуктов путем
синтеза их подходящими ш таммами бактерий.
71
5.1.2. Генетика человека и евгенические программы
Наследственность и изменчивость зависят от наличия, перераспре­
деления и изменений биологической информации, которую вид накопил
в процессе эволюции и которая отличается большим разнообразием.
Наряду с генами, обеспечивающими оптимальное выполнение функций
и высокий уровень жизнеспособности в целом, имеются гены,
снижающие жизнеспособность или даже вызывающ ие гибель орга­
низма. В последней четверти X IX века Ф . Гальтон поставил вопрос о
развитии особой науки — е в г е н и к и
(о т греч. ей — хороший,
Зепеы$ — род, происхождение). Ее задачей должно было стать
улучшение человеческого рода путем повышения в генотипе количества
полезных аллелей (особенно генов талантливости, гениальности) и
снижения доли вредных. Достичь этого предполагалось систематиче­
ским избирательным размножением одаренных людей и ограничением
репродукции асоциальных индивидуумов, например, преступников.
Такое размножение должно регламентироваться законодательными
актами.
Попытки воплощения евгенической идеи на практике имели место в
странах Западной Европы и Северной Америки в первые десятилетия
текущего столетия. Они показали бесперспективность и опасность
решения социальных задач, используя биологические механизмы.
Законы об ограничении браков, деторождения, о проведении принуди­
тельной стерилизации, принятые в двадцати ш татах С Ш А , вызвали
бурные протесты и через 1—2 года были фактически упразднены.
Концепция «расовой гигиены» использовалась нацистами фашистской
Германии в качестве оправдания их человеконенавистнической полити­
ки, решительно осужденной человечеством. Евгенические программы
полностью дискредитировали евгенику как науку, что явилось следстви­
ем ошибочности теоретических и философских принципов, из которого
исходили основоположники евгеники, известного примитивизма генети­
ческих представлений того времени, игнорирования качественного
своеобразия социальной формы движения материи.
Сейчас очевидно, что выполнение евгенических программ на основе
избирательного размножения по организационным соображениям
нереально даже в той части, в какой они направлены на снижение доли
вредных генов. П о мнению популяционных генетиков каждый человек в
среднем несет 2— 4 летальных рецессивных аллеля, которые приводят в
гомозиготном состоянии к смерти индивидуума. Д ля того чтобы эти
аллели не пропустить в следующее поколение, необходимо обеспечить
участие в размножении лишь одного из каждых 55 мужчин, предлагая
ему выбор партнера всего из 2 % женской популяции. При сохранении
возможности вступить в брак всем членам популяции для достижения
названного результата требуется ограничить деторождение одной
супружеской парой на каждые 2980 пар.
При увеличении числа
нежелательных аллелей до 10 (4 летальных и 6 полулетальных)
последняя цифра возрастает до 485 млн. супружеских пар.
Интерес к евгенике способствовал разработке некоторых вопросов
антропогенетики. В целом же способы и результаты • реализации
72
евгенических программ бросили тень на генетику человека как таковую ,
что существенно затормозило развитие этой важнейшей области науки
о наследственности и изменчивости.
Современная генетика, молекулярная биология и медицина
располагают средствами манипулирования с наследственным материа­
лом, намного превосходящими по своим возможностям ограничение
браков. Э то искусственное осеменение и зачатие «в пробирке» с
последующим перемещением зародыша в м атку женщины-реципиента,
отбор зародышей на ранних стадиях развития, генетическая инженерия,
пересадка ядра соматической клетки в цитоплазму яйцеклетки с га­
рантией развития полноценной особи. Важно, однако, понимать, что
биологические способы воздействия с целью улучшения человеческого
общества неприемлемы, какую бы конкретную форму они ни
принимали. В этом смысле возврат к евгенике невозможен. Н о генетика
и медицина не могут бы ть освобождены от ответственности за
здоровье потомства. Действительно, в настоящее время в мире
4— 5 % детей рождаются с наследственными нарушениями, 10—
20% детской смертности обусловлено наследственной патологией,
25— 30% коек в детских больницах заняты пациентами с наследственны­
ми болезнями. Расходы общества на содержание только больных
болезнью Дауна равны затратам на борьбу с гриппом. В клинике
кожных болезней описано около 250 наследственных заболеваний,
глазных болезней — более 200, нервных болезней — около 200. Общее
количество известных наследственных болезней превыш ает 1500.
В этом вопросе современная медицина ориентируется прежде всего на
профилактику наследственных болезней путем медико-генетического
консультирования, обнаружения гетерозиготных носителей небла­
гоприятных рецессивных аллелей, советов супружеским парам с повы­
шенным риском рождения больного ребенка, разъяснения нежела­
тельности поздних браков и деторождении (табл. 7).
Т а б л и ц а 7. Вероятность рождения ребенка с болезнью
Д аун а у матерей разного возраста (Англия, 1954)
В о з р а с т ж ен щ и н ы , годы
15— 19
20— 24
25— 29
30— 34
35— 39
40— 44
45 и более
Частота
болезни Д а у н а среди
ных (н а 10 000)
з
4
4
11
33
124
312
н о во р о ж д е н ­
.
Этой же цели служ ат медико-просветительные и социальные
мероприятия. К ним относятся разъяснение нежелательности и запре­
щение близкородственных браков, законодательные акты , направлен­
ные против загрязнения окружающей среды мутагенами или ужесточа­
ющие контроль на мутагенное действие химических соединений
73
промышленного и бытового назначения. Генетика и медицина в борьбе
за здоровье людей в каждом поколении учиты ваю т, что существенное
влияние на проявление положительных и отрицательных наслед­
ственных свойств, оказывает среда, в которой происходит развитие
человека. Исходя из этого принципа, Н . К . Кольцов (1929) выделил
новое направление в практической генетике человека — е в ф ен и к у — науку о благоприятном проявлении наследственных задатков.
5.2. Н А С Л Е Д С Т В Е Н Н О С Т Ь
5.2.1. Структурно-функциональные уровни организации
наследственного материала эукариот
Наследственность и изменчивость обеспечивают: 1) индивидуальное
(дискретное) наследование и изменение отдельных признаков; 2) вос­
произведение в особях каждого поколения всего комплекса морфо­
функциональных характеристик организмов конкретного биологическо­
го вида; 3) перераспределение у видов с половым размножением в
процессе воспроизведения наследственных задатков, в результате чего
потомок имеет сочетание признаков, отличное от их сочетания у
родителей. Закономерности наследования и изменчивости признаков и
их совокупностей вы текаю т из принципов структурно-функциональной
организации генетического материала. Различаю т три уровня организа­
ции наследственного материала эукариотических организмов: генный,
хромосомный и геномный (уровень генотипа).
Элементарной структурой г е н н о г о у р о в н я служит г е н .
Передача генов от родителей потомку необходима для развития у него
определенных признаков. Х о тя известно несколько форм биологической
изменчивости, только нарушение структуры генов изменяет смысл
наследственной информации, в соответствии с которой формируются
конкретные признаки и свойства. Благодаря наличию генного уровня
возможно индивидуальное, раздельное (дискретное) и независимое
наследование и изменения отдельных признаков.
Гены клеток эукариот распределены группами по
хро­
мосомам.
Э то структуры клеточного ядра, которым свойст­
венна индивидуальность и способность к самовоспроизведению с
сохранением в ряду поколений индивидуальных черт строения. Наличие
хромосом обусловливает выделение х р о м о с о м н о г о у р о в н я
организации наследственного материала. Размещение генов в хромосо­
мах влияет на соотносительное наследование признаков, делает
возможным воздействия на функцию гена со стороны его ближайшего
генетического окружения — соседних генов (о сцепленном наследовании
и эффекте положения см. 5.2.4. и 5.2.5.). Хромосомная организация
наследственного материала служит необходимым условием перераспре­
деления наследственных задатков родителей в потомках при половом
размножении.
Несмотря на распределение по разным хромосомам, вся со­
вокупность генов в функциональном отношении ведет себя как целое,
образуя единую систему, представляющую г е н о м н ы й (г е н о ­
типический)
уровень
организации
наследственного
74
материала. Н а этом уровне происходит широкое взаимодействие и
взаимовлияние наследственных задатков, локализующихся как в одной,
так и в разных хромосомах. Итогом является взаимосоответствие
генетической информации разных наследственных задатков и, следова­
тельно, сбалансированное по времени, месту и интенсивности развитие
признаков в процессе онтогенеза. Функциональная активность генов,
режим репликации и мутационных изменений наследственного матери­
ала также зависят от характеристик генотипа организма или клетки в
целом. Об этом свидетельствует, например, относительность свойства
доминантности.
5.2.2. Ген и его свойства
В процессе жизнедеятельности организма наследственный материал
функционирует, редуплицируется, испытывает изменения мутационно­
го или рекомбинационного характера. Классики генетики предполагали,
что элементарной единицей этих процессов служит одна и та же
структура — г е н . В их представлениях он соответствовал наслед­
ственным задаткам, откры ты м Г . Менделем. Выход генетики на
молекулярный уровень исследования лишил ген положения единицы
редупликации, мутирования и рекомбинации. В настоящее время ген
рассматривается как
единица
функционирования
н а с л е д с т в е н н о г б" м а т е р и а л а. ^ то означает, что переда­
чки 1 енгш и 'рТО7 Ягеёточных или организменных поколений достигается
материальная
структурно-функциональная
п р е е м с т в е н н о с т ь , наследование потомками признаков роди­
телей.
Здесь целесообразно дать определение понятию «п р и з н а к». Под
признаком понимают единицу морфологичеткпй, физиологической,
'биохимической, иммунологической, клинической и любой другой
Секретности организму, т. е. любое отдельное качество или свойство,
по которому одну особь можно отличить от другой. Морфофизиологи­
ческие и клинические признаки более сложны по сравнению с биохими­
ческими или иммунологическими, которые сводятся к отдельным
белкам. Сложность морфологических и клинических признаков зависит
от того, что они генетически опосредуются через синтез нескольких
ферментов, иммунопротеинов и т . д. Лиш ь по осуществлении ряда
биохимических реакций, катализируемых этими ферментами, в орга­
низме появляется соответствующ ий признак. Примером этому служит
повышенная чувствительность к солнечным лучам и опухоли кожи, что
типично для клиники наследственного заболевания пигментной
ксеродермы. Долгое время считали, что названные признаки определя­
ю тся самостоятельными генами, однако в настоящее время выяснено,
что наследуется дефект фермента биохимической системы восстановле­
ния молекулы Д Н К от поражений ультрафиолетовым облучением.
Сходные уточнения первичных звеньев, определяемых генетически
непосредственно, будут, по-видимому, сделаны в отношении большин­
ства, если не всех сложных признаков.
П о химической природе ген представляет фрагмент молекулы Д Н К ,
75
который содержит информацию о последовательности аминокислот в
определенном полипептиде или нуклеотидов в рибонуклеиновых
кислотах (рибосомных, транспортных)1 . В современной генетике
указанный фрагмент Д Н К обозначается термином «цистрон».
Ген как единица функционирования наследственного материала
имеет
ряд
свойств.
Одно
из
них — д и с к р е т н о с т ь
д е й с т в и я , т. е. развитие различных признаков контролируется
разными генами, локализация которых в хромосомах не совпадает. Ген
как
дискретная
единица
наследственности
отличается
с т а б и л ь н о с т ь ю (постоянством) — при отсутствии мутации он
пепрлается в ряду поколений в неизменном виде.
|/ ( Действие генов с п е ц и ф и ч н о в том смысле, что каждый из
" них обусловливает развитие определенного признака или их группы.
‘Способность отдельных генов обеспечивать развитие одновременно
уДч - нискольких признаков называется п л е й о т р о д и е й или плей'
отрбпным эффектом. Примером плейотропного эффекта гена у челове­
ка служит синдром Марфана. Х о тя это наследственное заболевание
зависит о т присутствия в генотипе одного измененного гена, оно
характеризуется в типичных случаях триадой признаков: подвывихом
хрусталика глаза, аневризмой аорты, изменениями опорно-двиГУтельного аппарата в випе «паучьих пальцев», деформированной грудной
клеткой, высоким сводом стопы. Все перечисленные признаки являю тся
сложными. По-видимому, в основе их лежит один и то т же дефект раз­
вития соединительной ткани.
Так как продуктом функции гена наиболее часто является белокфермент, выраженность плейотропного эффекта зависит от оаспоастраненности в организме биохимической реакции, которую катализи­
рует фермент, синтезируемый под генетическим контролем данного
' гена. Распространенность поражений в организме в случае наследственного заболевания тем больше, чем выраженнее плейотропный
эффект измененного гена. Примеры участия в развитии одного
сложного признака сразу нескольких генов будут рассмотрены ниже.
Больш инство генов сущ ествует в виде двух или большего числа
альтернативных
(взаимоисключающ их) в а р и а н т о в —
а л л е л е й . Все аллели данного гена локализую тся в одной и той же
хромосоме, в определенном ее участке, именуемом л о к у с о м.
Альтернативный характер аллельных генов заклю чается в том , что
одновременно в соответствую щ ем локусе хромосомы может нахо­
диться лиш ь один из всех возможных аллелей конкретного гена. С ф ия
аллелей возникает путем мутаций в одном и том же локусе. В силу это­
го аллели отличаю тся друг от друга содержанием наследственной
информации о признаке, развитие которого контролирует ген.
Показателен пример с гемоглобинами человека, которых к настоящему
времени идентифицировано более 100. Белковая часть нормального
гемоглобина взрослых людей (Н ЬА ) представлена а- и Р-полипептидами, каждый из которых повторен дважды, а-Полипептид образо1 Современная молекулярно-биологическая концепция тонкой структуры
изложена ниже (см. 5.2.10).
76
гена
ван 141, а р-полипептид — 146 аминокислотными остатками. Более
80 вариантов гемоглобина отличаю тся друг от друга одним амино­
кислотным остатком в а- или р-полипептиде, что указывает на
мутационную природу аллелей, контролирующих их синтез. НЬ8
отличается о т нормального НЬА наличием в шестом положении
2 полипептида валина, а Н Ъ С — лизина вместо остатка глутаминовой
кислоты. Э ти минимальные отличия меняю т свойства гемоглобина
настолько, что образование в организме исключительно НЪ8 или НЪС
% \ приводит к заболеванию анемией. Генетико-биохимический анализ
показал, что из трех названных гемоглобинов одновременно в орга­
низме человека образуется только два. Обнаружены следующие
варианты: НЪ8 /НЪ8 (серповидноклеточная анемия) или НЪС/НЪС
(анемия в более легкой форме), а такж е НЬА/Н Ъ8, Н ЬА /Н ЬС
^
(клинически здоровые люди) и НЬЗ/НЪС (анемия). Э то доказывает, что
^ синтез Н ЬА , НЬ8 и Н ЪС контролируется генами, занимающими один и
то т же локус, т. е. аллельными.
,4Г '
В приведенном примере присутствие в организме человека двух
*' разных аллелей приводит к образованию обоих вариантов р-поли­
пептида гемоглобина: Н ЬА и НЪ8, НЪА и НЪС, НЪ8 и НЪС. Аллели, в
* одинаковой мере функционально активные в случае их совместного
4, » присутствия в генотипе, называю тся
кодоминантными.
Аллель, функциональная активность которого не зависит от наличш Гв
% организме
другого
аллеля
данной
серии,
называется
д ом й н а' й т н ы м! Аллель, обеспечивающий развитие признака
» лишь в отсутствии других аллелей данного гена, обозначают как
р е ц е с с и в н ы й. Так, пигментация радужной оболочки глаза,
кожных покровов, волос человека зависит от образования черного
пигмента меланина. Соответствую щ ий ген представлен двумя аллеля­
ми. Первый (доминантный), присутствуя даже в единственном экзем­
пляре, проявляет себя в обязательном порядке, вы зывая синтез
§ меланина. Второй аллель (рецессивный) исключает возможность
синтеза меланина (аллель-альбинизма) и проявляет себя лиш ь в отсут^ ствие первого. Одной из Ф о р м взаимодействия аллельных генов/
является н е п о л н о е д о м и н и р о в а н и е , которое заклю ч а е тс я^ / ;
■"ч в ослаблении действия доминантного аллеля в присутствии рецессивного. Так, активность Фермента Фенилаланингидроксилазы у но^
сителей одновременно нормального и аномального (рецессивного)
аллелей выше, чем у больных фенилкетонурией, имеющих два
аномальных аллеля, но ниже, чем у носителей двух нормальных
аллелей. Неполное доминирование отражает собой, по-видимому,
дозированность действия доминантных аллелей.
Доминантность и рецессивность относительны. Так, развитие
кожной складки над верхним веком (так называемое третье веко или
эпикант) контролируется аллелем, проявляющим свойство доми­
нантности у представителей монголоидной расы и рецессивности — у
бушменов и готтентотов. Аллель роста волос на голове ведет себя как
доминантный у женщин и рецессивный у мужчин. При массированном
введении женщине в лечебных целях мужского полового гормона этот
аллель нередко начинает проявлять свойство рецессивности, что
77
Л *
< лУ1>-»л ’
«
- *
^
/* „ > * ч
-к У
Ч>
Л
О
Ч
,.
с* * * 4
,.\ *
Сп КК в1)Й У
КХ XXМ XX8» «8 88
I
С
6
О
АЛ ЛЛ ал
13
Р
* х
Ш
14
ХК
20
15
е
Ю
||
12
*х
кк
хл
16
17
18
8
6 Л А
21
ЛЛ
22
II
а
Рис. 28. Хромосомный набор больной с синдромом Шерешевского
сомия по паре половых хромосом),
а
Тернера (моно-
к ар и оти п .
приводит к нежелательному побочному эффекту в виде облысения.
Последний пример показывает, что свойства доминантности и ре
цессивности зависят от условий функционирования аллелей. Поэтому
названные свойства правильнее относить не к аллелям непосредственно,
а к определяемым ими признакам. В табл. 8 перечислены некоторые
78
доминантные и рецессивные признаки
человека.
Поскольку каждому гену соответ­
ствует строго определенный локус в од­
ной из хромосом, понятно, что в гапло­
идном хромосомном наборе содержится
по одному, а в диплоидном — по два
аллеля одной серии. В последнем случае
при наличии двух одинаковых аллей
говорят о гомозиготном состоянии орга­
низма, а при наличии разных аллелей —
о гетерозиготном. Сказанное относится к
генам, расположенным в аутосомах. Так
как генный состав половых хромосом
X и V совпадает лишь в небольшой своей
части, для организма мужчины по
многим генам Х-хромосомы и некото­
рым генам У-хромосомы создается гемизиготное состояние. В этом случае даже
в диплоидном хромосомном наборе ген
представлен одним аллелем.
О т аллелей зависит генетический
полиморфизм, наследственное разнооб­
разие особей определенного вида. Благо­
даря множественному аллелизму генов
создается резерв наследственной измен­
чивости. определяющий эволюционные
перспективы.
Действие гена
дозировано.
28 Прид()ЛЖени,
Присутствуя в клетках организма в
й фен.™.
одном экземпляре, аллель обеспечивает
развитие соответствую щ его признака до известного количественного
предела. Параллельно увеличению «доз» аллеля нарастает количество
признака. Так, синтез витамина А в клетках эндосперма кукурузы контролируется доминантным аллелем V. Поскольку рассматриваемые клет­
ки содержат тройной набор хромосом, в них возможны следующие ва­
рианты количества «доз» аллеля У: У У У , У У у , Ууу, ууу. Содержание
витамина А в эндосперме перечисленных растений составляет, соответ­
ственно, 7,50; 5,00; 2,25; 0,05 единиц. Некоторые хромосомные болезни
человека возникают вследствие изменения «дозы» генов определенных
хромосом. К развитию синдрома Шерешевского — Тернера, например,
ведет утрата клетками организма одной из пары половых хромосом
(рис. 28), тогда как при болезни Дауна имеет место увеличение до трех
«доз» генов 21 хромосомы (рис. 29). Описаны также утрата или
увеличение части хромосомы. К примеру, синдром «кошачьего крика»
развивается в случае потери фрагмента короткого плеча хромо­
сомы 5.
Определенная «дозировка» генов в онтогенезе необходима для
нормального развития. В этом убеждает обязательная функциональная
79
П 5^> И
3»
6
а*
7
XX
Ц
6Л
ПК
8
9
XX
12
11
10
ПК
&
х
АА
№
13
14
15
XX
XX
Лй
16
17
18
КX
XX
19
20
м
А Л Л
21
А
*
22
А
У
Рис 29. Хромосомный
набор при
болезни
Д ауна
(трисомия по
21 паре аутосом).
а — к ар и оти п ; С
фенотип.
Таблица
8. Доминантные и рецессивные признаки человека
Д о м и н антны е
Признаки
Глаза
Цвет глаз
Разрез глаз
Тип глаз
Острота зрения
Верхнее веко
Ямочки на щеках
Уши
Подбородок
Выступающие зубы
и челюсти
Ш ель между резцами
Волосы
Поседение волос
Облысение
Белая прядь волос над
лбом
Рост волос по средней
линии лба
Мохнатые брови
Н и ж н яя губа
Способность
загибать
язык назад
Способность
свертывать
язык трубочкой
Зубы при рождении
Кожа
Цвет кожи
Веснушки
Кисть
Преобладающая рука
Узоры на коже пальцев
Антигены системы
АВО
Голос (у женщин)
Голос (у мужчин)
Абсолютный
музыкальный слух
Наследственная глухота
Рец есси вн ы е
Большие
Карие
Прямой
Монголоидный
Близорукость
Нависаю щ ее
(зп и кан т)
Есть
Широкие
Длинный
Прямой
Широкий
Имеются
Маленькие
Г олубые
Косой
Европеоидный
Нормальное
Нормальное
Нет
Узкие
Короткий
Отступающий назад
Узкий н острый
Отсутствуют
Есть
С мелкими завитками
Ж есткие, прямые, «ежик*
Вьющиеся
В возрасте 25 лет
У мужчин
Имеется
Нет
Вью щ иеся, волнистые
Прямые, мягкие
Волнистые или прямые
После 40 лет
У женщин
Отсутствует
Есть
Нет
Есть
Толстая и отвисающая
Е сть
Нет
Нормальная
Нет
Есть
Нет
Имеются
Толстая
Смуглый
Есть
С 6 или с 7 пальцами
П равая
Эллиптические
А, В
Отсутствуют
Тонкая
Белый
Нет
С. 5 пальцами
Л евая
Циркулярные
0
Сопрано
Бас
Имеется
А тьт
Тенор
Отсутствует
Отсутствует
Имеется
инактивация комплекта генов одной из Х-хромосом в клетках женского
организма, которая происходит после 16 сут. внутриутробного разви­
тия путем ее гетерохроматизации и преврешения в тельце полового
хроматина (рис. 30).
81
О
Хромосомы
клето н
п о ло вы х
н ле то к
О
+
ге р м е н а т и в н о й
н л е тн и
XX
х
ХУ
половы х
кл е ток
( м о з а и к а п о а к ти в н о й
X хром осом е)
Рис. 30. Функциональная инактивация одной из пары Х-хромосом в онтогенезе
женщины (подчеркнута Х-хромосома, полученная от матери).
5.2.3. Закономерности наследования,
установленные Г. Менделем
Выдающийся вклад Г . Менделя в науку состоит в Экспери­
ментальном доказательстве наличия единиц наследственности (наслед­
ственных задатков, генов) и описании их важнейших свойств —
дискретности, стабильности, специфичности, аллельного состояния.
Разработав и применив в опытах на растениях гибридологический
метод, Г . Мендель, анализируя результаты моно- и дигибридных
скрещиваний гороха, пришел к заключению, что: 1) развитие признаков
зависит от передачи потомкам наследственных задатков; 2) указанные
задатки передаются в ряду поколений, не утрачивая своей индиви­
дуальности, т. е. характеризуются постоянством; 3) наследственные
задатки, контролирующие развитие конкретного признака, парные —
один из них переходит к потомку от материнского, второй — от
отцовского организма; в функциональном отношении эти задатки
проявляют свойства доминантности и рецессивности; 4) в пропессе
образования половых клеток парные наследственные задатки попадают
в разные гаметы (закон чистоты гам ет); восстановление пар таких
задатков происходит в результате оплодотворения; 5) материнский и
отцовский организмы в равной мере участвую т в передаче своих
наследственных задатков потомку.
Э ти положения отражаю т общие принципы организации наслед­
ственного материала: 1) дискретное определение развития наследуемых
признаков; 2) относительная стабильность наследственных единиц;
3) аллельное состояние наследственных единиц.
И з этих принципов вы текаю т правила (законы ) наследования,
сформулированные Г . Менделем: 1) единообразие признака у гибридов
первого поколения; 2) расщепление альтернативных вариантов призна­
ка среди особей второго поколения; 3) независимое комбинирование
признаков родителей в потомках. Генетические закономерности,
впервые откры ты е Г . Менделем, описываю т правила независимого
наследования, в основе которого лежит наличие дискретных единиц
наследственности генов.
Признаки, наследование которых следует закономерностям,
установленным Г . Менделем, называю тся менделируюшими. Некото­
рые менделирующие признаки человека перечислены в табл. 8.
Общее количество их, известных к 1975 г., составило более 2300. Все
менделирующие
признаки
дискретны
и
контролируются
м о н о г е н н о, т. е. одним геном.
5.2.4. Хромосома. Положения хромосомной
теории наследственности
Х о тя результаты опытов Г . Менделя свидетельствую т о передаче в
ряду поколений отдельных, обособленных единиц наследственности,
они ничего не говорят об их физической природе и расположении в
половых или соматических клетках. Вскоре после повторного открытия
ь в 1900 г. закономерностей Г . Менделя был отмечен ^раллёлйЗм^ в
наследовании признаков при моно-, ди- и полигибридных скрещиваниях
и поведении хромосом клеточного ядра в процессе гаметогенеза и при
оплодотворении (рис. 31). В анафазе 1-го мейотического деления число
хромосом редуцируется вдвое. При расхождении к полюсам хромосомы
разных гомологичных пар перемещаются независимо друг по отноше­
нию к другу и, следовательно, комбинируются в дочерних клетках
случайным образом. В результате оплодотворения хромосомы яйце­
клетки и сперматозоида объединяются в удвоенном наборе хромо­
сом зиготы. И з указанного параллелизма следует, что: 1) развитие
альтернативных вариантов признака обусловливается аллельными
генами, локализующимися в гомологичных хромосомах; 2) гаметы,
имея лишь по одной хромосоме из каждой гомологичной пары, несут по
одному аллелю соответствую щ его гена; 3) гены, контролирующие
развитие признаков, наследуемых независимо, размещ аются в негомо­
логичных хромосомах. Э ти заключения убеждают в хромосомной
локализации генов.
Наряду с признаками, наследуемыми независимо, обнаружены
признаки, наследуемые совместно (сцепленно). Экспериментальное
исследование этого явления, проведенное Т. Г . Морганом и его группой
(1910— 1916), подтвердило хромосомную локализацию генов и легло в
основу х р о м о с о м н о й т е о р и и н а с л е д с т в е н н о с т и .
В работах на плодовой мушке было установлено, что гены по признаку
совместной их передачи потомкам подразделяются на 4 группы. Число
таких « г р у п п с ц е п л е н и я » равно количеству хромосом в гапло83
Р га м е ты
Рис. 31. Схема независимого распределений признаков при дигибридном скрещи­
вании.
А , В , а, в — д о м инантны е и рецессивны е аллели , к о н тр о ли р ую щ и е р азв и ти е д вух п р и зн ак о в;
ты - п оловые к ле тки родителей; Р , — гибриды 1-го п окол е н и я; Р 2
гнбрнды 2-го п околения.
Р
га м е ­
’, ' с*У
Р-/-6 1-1/цЛ
>а тх1>м: к/Л'а-14- •с
я
./ Ь
I I
Н е н р о с с о в е р ы В 3%
Н р о с с о в е р ы 17%
Рис. 32. Сцепленное наследование генов окраски тела и состояния крыльев у пло­
довой мухи.
Г е н серого ц вета те л а ( В ) д о м инирует над геном черного ц вета те л а ( Ь ) . ген н орм альной длины кр ы л ье в
( V ) — над геном коротких к р ы л ь е в (V ). В н V н а х о д ятся в одной хромосоме.
а — полное сцепление п р и зн ак о в всле д стви е о т с у т с т в и я пер екр еста хромосом у сам ц о в дрозоф илы: Р Р —
са м к а с е р а я с н ор м альн ы м и к р ы л ья м и ( В В У \ ) скр е щ е н а с черным кор о ткокр ы л ы м сам цом ( Ь Ь у у ) ; Р х —
серый сам е ц с нор м альн ы м и к р ы л ьям и ( В й У у ) скр ещ ен с черной кор о ткокр ы л ой сам к о й (Ъ Ъ у у ); Р3 — по­
с к о л ь к у у сам ц а не происходит кр оссин говер ^, р о ж д а ю тс я д ва ва р и а н та пото м ко в: 5 0 % — черных ко р о тко ­
к р ы л ы х н 5 0 % — серых с нор м альн ы м и к р ы л ья м и ; б
- неполное (ч а с т и ч н о е ) сцепление п р и зн аков
всле д стви е пер екр еста хромосом *у сам о к др о зоф илы : Р Р — с а м к а с е р а я с нор м альн ы м и к р ы л ьям и
( В В У У ) ск р ещ ен а с черным к ор о ткокр ы л ы м сам цом ( Ь Ь у у ) ; Р , — се р а я с а м к а с нор м ал ьн ы м и к р ы л ьям и
( В Ь У у ) скр ещ ен а с черным к ор о ткокр ы л ы м самцом ( Ь Ь у у ) . Р 2 — п о ск о л ьк у у сам к н происходит кр оссин ­
говер гомологичных хромосом, о б р а зу е тс я четы р е ти п а гам ет и р о ж д а е т с я четы р е ва р и а н та пото м ко в:
н екр оссоверы — се р ы е с н ор м альн ы м и кр ы л ьям и ( В Ь У у ) н чер н ы е ко р о тко кр ы л ы е ( Ь Ь у у ) , кр оссовер ы —
чер н ы е с н ор м альн ы м и кр ы л ьям и ( Ь Ь У у ) , сер ы е к о р о ткокр ы л ы е ( В Ь у у ).
вдном наборе. М ожно было заклю чить, что развитие признаков,
которые наследуются сцепленно, контролируется генами одной
хромосомы. Э то т вывод обосновывается также данными следующих
наблюдений. Скрещивание серой мухи (В )1с нормальными крыльями (V )
и черной мухи (Ъ) с зачаточными крыльями (в) дает в первом поколении
серых гибридов с нормальными крыльями (ВЬ/У г). При скрещивании
самца-гибрида 1-го поколения (ВЬ/ Г г) с черной самкой с зачаточными
крыльями
рождаются особи 2 видов, аналогичных исходным
родительским формам, причем в равном количестве (рис. 32, а).
1 Буквам и латинского алфавита в генетике обозначают аллели гена: заглав­
ной — доминантный, строчной — рецессивный.
85
Полученные в проведенных скрещиваниях данные нельзя объяснить
независимым наследованием признаков. Рассматриваемые совместно
результаты обоих скрещиваний убеждают в том , что развитие
анализируемых_признаков конт ролируется разными генами^и сцеплен­
ное..наследование этих признаков объясняется локализацией генов в
одной хромосоме.
""С кр ещ и ван и е в сходных опытах самки-гибрида 1-го поколения с
черным самцом с зачаточными крыльями дает четыре варианта
особей
(рис. 32, б). Они имеют сочетания признаков, которые
образуются и при независимом наследовании, но количественное
соотношение гибридов другое. Такой результат указывает на частичное
сцсплецие-наследуемых признаков, а нарушения снепления объясняю тся
кроссинговером
Сравнение результатов 2-го (гибрид Г ,
в
«Скрещивании
представлен
самкой)
и
1-го
(гибрид Г ,
в
скрещивании представлен самцом) опытов свидетельствует о возмож­
ности как частично сцепленного, так и полностью сцепленного
наследования одной пары признаков. Э то противоречие объясняется
особенностью биологии плодовой мушки, которая заклю чается в отсут­
ствии кроссинговера в гаметогенезе самцов. В примере, рассмотренном
'"вы ш е, число псэтойкои С кзмененным~сб5Тношением признаков (такие
особи называю тся кроссоверами) и, следовательно, контролирующих
их аллелей, составляет 17%. Э та цифра указывает на относительное
число гамет, в процессе образования которых произошел кроссинговер
(кроссоверные гам еты). Другие сочетания генов плодовой мушки
отличаю тся иной частотой кроссинговера. При постоянных условиях
(температура, возраст самки и др.) для каждой пары генов одной
хромосомы это значение постоянно. Так, по генам белых глаз и желтой
окраски тела число кроссоверных гамет у дрозофилы составляет 1,5%,
а по генам желтой окраски тела и зачаточных крыльев — 4 7 % . При
этом сила сцепления генов обратно пропорциональна расстоянию между^ними в хромосоме ( п р а в и л о и л и з а к о н М о р 1г~а н 4)гг
'
~
~
Постоянство процента кроссинговера между генами используется
как показатель относительного расстояния между ними и порядка
взаиморасположения при составлении генетических карт хромосом. За
единицу расстояния между генами принята м о р г а н и д а. Она
соответствует дистанции, при которой кпоссинговеп... прпчгуппнт
в 1% гамет. Процент кроссинговера для разных пар генов колеблется
от долей единицы до пятидесяти, не превышая, однако, последнюю
цифру. При расстоянии в 50 морганид и более признаки наследуются
независимо, несмотря на то, что гены локализую тся в одной
хромосоме.
Основные положения хромосомной теории наследственности,
сформулированной Т. Г . Морганом, заклю чаю тся в следующем.
1. Гены располагаются в хромосомах; различные хромосомы
содержат неодинаковое число генов; набор генов каждой из негомоло­
гичных хромосом уникален.
2. Аллельные гены занимают определенные и идентичные локусы
гомологичных хромосом.
86
3. В хромосоме гены располагаются в определенной последова­
тельности по ее длине в линейном порядке.
4. Гены одной хромосомы образуют группу сцепления, благодаря
чему имеет место сцепленное наследование некоторых признаков; сила
сцепления находится в обратной зависимости от расстояния между
генами.
5. Каж ды й биологический вид характеризуется специфичным
набором хромосом — кариотипом.
Генетические закономерности, описываемые хромосомной теорией
наследственности, соответствую т правилам сцепленного наследования
признаков и вы текаю т из факта хромосомной локализации генов.
,2.5. Генетическая система организма (генотип, геном)
Несмотря на дискретное генетическое определение отдельных
признаков, в индивидуальном развитии воссоздается сбалансированный
комплекс признаков и свойств, соответствую щ ий типу морфофункцио­
нальной организации конкретного биологического вида. Закономерно
возникают плазмодий малярийный, кедр ливанский, аскарида человече­
ская, слон индийский, человек разумный. Э то достигается вследствие
интеграции дискретных в структурном отношении единиц наслед­
ственности в целостную в функциональном плане систему —
г е н о т и п ( г е н о м )1. Такая интеграция находит отражение в
разнообразных взаимодействиях генов в процессе их функционирова­
ния.
Обычно генотип определяют как совокупность всех генов (более
точно аллелей) организма. С учетом факта интеграции генотип
представляется системой определенным обпазом взаимолейгтнуюпрт
_генов._Генные взаимодействия происходят на нескольких уровнях:
непосредственно в генетическом материале клеток, между и РН К и
образующимися полипептидами в процессе биосинтеза белка, между
белками-ферментами одного метаболического цикла.
Взаимодействие генов на уровне продуктов функциональной
активности (Р Н К или полипептидов) лежит в основе развития сложных
признаков. Рассмотрим в качестве примера синдром Морриса.
У больных, кариотип которых вклю чает половые хромосомы X и V ,
отмечается недоразвитие вторичных половых признаков мужского
пола, которое зависит от продукции и взаимодействия на известной
стадии онтогенеза двух факторов — мужского полового гормона и
белка-рецептора, встраивающ егося в клеточную оболочку и делающего
клетки чувствительными к гормону. Синтез указанных факторов
контролируется разными генами. У лиц с синдромом Морриса мужской
половой гормон образуется своевременно и в требуемом количестве, но
не синтезируется белок-рецептор. Таким образом, нормальное развитие
сложного признака комплекса мужских вторичных половых признаков
контролируется двумя генами, -которые взаимодействуют на уровне
продуктов их функциональной активности.
1 Термином «генотип» обозначают совокупность аллелей (генов) диплоидного
набора хромосом, а термином «геном» — гаплоидного.
87
В настоящее время для больш инства признаков нельзя указать
точно уровень взаимодействия тех генов, которые контролируют их
развитие. Учиты вая интерес практического врача прежде всего к
закономерностям наследования признаков, ниже приводятся формы
взаимодействия генов, которые изменяют наследование определенным
образом. При этом уровень взаимодействия генов не оговаривается.
Выделяю т
взаимодействие
аллельных
и
н е а л л е л ь н ы х г е н о в . Основные формы взаимодействия ал­
лельных генов рассмотрены выш е. Они обусловливают доминантное,
рецессивное, кодоминантное наследование признаков, явление неполно­
го доминирования. При перечисленных формах доминирования ре­
зультаты взаимодействия генов проявляю тся во всех соматических
клетках организма* При
такой
форме взаимодействия
как
а л л е л ь н о е и с к л ю ч е н и е в части клеток организма, гетеро­
зиготного по данному локусу, активен один аллель, тогда как в других
клетках другой. В качестве примера рассмотрим генетический контроль
синтеза иммуноглобулинов — белков плазмы крови, которые обеспечи­
ваю т в организме человека реакции иммунологической защ иты. Они
состоят из «тяж елы х» и «легких» полипептидных цепей, которые
синтезируются под генетическим контролем трех разных групп
неаллельНЫХ генов. И «тяж елы е», и «легкие» полипептиды образуются
плазматическими клетками. При этом отдельные плазматические
клетки синтезируют лиш ь по одному из возможных вариантов
«тяжелых» и «легких» полипептидов глобулинов. Аллельное исключе­
ние увеличивает разнообразие признаков многоклеточного организма
при идентичности генотипов соматических клеток. Механизм этого
явления окончательно не установлен. Другим примером аллельного
исключения является генетическая инактивация одной из Х-хромосом
женских особей. В мировой литературе описаны лиш ь единичные случаи
заболевания женщин гемофилией. Вместе с ТСм матери — гетерози­
готные носители аллеля гемофилии — передают его половине своих
дочерей, которые нормальный аллель получаю т с Х-хромосомой отца.
Случайный характер инактивации путем гетерохроматизации приво­
дит к выключению из функции в одних клетках материнской, а в дру­
гих — отцовской Х-хромосомы. Таким образом, всегда остаю тся
клетки, которые несут нормальный аллель синтеза антигемофилического фактора в активном состоянии.
Одной из форм взаимодействия неаллельных генов является
\л«э ф ф е к т п о л о ж е н и я » . В нем участвую т гены одной хромосо­
мы, занимающие близлежащие локусы. Он проявляется в изменчивости
функциональной активности гена в зависимости о т того, какой аллель
находится в соседнем локусе. Так, эритроцитарные белки-антигены
системы групп крови «резус»’ (ЛА) синтезируются
в организме
человека под контролем трех генов. Последние, судя по выраженному
сцеплению между ними, располагаются в хромосоме на близком
расстоянии друг от друга. Каж ды й из них имеет доминантный и
■' Сейчас известно 14 систем групп крови человека, из которых наиболее извест­
на АВО.
Рис. 33. Эпистаз в насле
довании окраски плодов у
тыкв. Объяснение в тек­
сте.
тга
<
Г аметы С ? АаВЬ
АВ
АЬ
аВ
аЬ
<2 А В А А В В А АВЬ А а В В АаВЬ
2
ф
□
белые
АЬ ААВЬ ААЬЬ АаВЬ АаЬЬ
аВ
Я аЬь
I—
А аВВ АаВЬ ;ааВВ /ааВЬ/
АаВЬ
АаЬЬ СааВЬ- <ааЬЬ>
рецессивный аллели — В , С , Е к а, с, е. Индивидуумы СОЕ/сБе и
СИе/сОЕ генетически идентичны. Тем не менее у лиц с первой комбина­
цией аллелей образуется много антигена Е и мало антигена С , у лиц со
второй комбинацией аллелей наблюдается обратная картина, т. е. со­
седство аллеля С с аллелем Е снижает его функциональную активность.
«Эффект положения» служит частным случаем большого класса
генных взаимодействий, заключающихся в м о д у л и р о в а н и и
ф у н к ц и и г е н о в д р у г и м и г е н а м и . К этом у классу вза­
имодействия неаллельных генов относится также э п и с т а з,—
подавление одного гена другим. Если эпистатическим действием
обладает доминантный аллель, говорят о доминантном эпистазе. При
рецессивном эпистазе такое действие проявляю т рецессивные аллели в
гомозиготном состоянии. Примером зпистаза служит наследование й ,ила/[
окраски плодов у некоторых ты кв. Указанный признак контролируется^
геном, представленным двумя аллелями: доминантным — ж е л то й Д ^ '^
окраски (В ) и рецессивным — зеленой окраски (Ь). Если в гейотипрД] <растения присутствует доминантный аллель (А ) другого гена, то ъ_У-~°
присутствии любого из аллелей первого гена развивается белая окраска.
Рассмотрим с помощью решетки Пеннета результаты скрещивания
растений, дигетерозиготных по генам А к В (рис. 33). В отличие от
обычного дигибридного скрещивания в данном случае наблюдается
расщеплейие гибридов по признаку окраски плодов в отношении 12
(б ел ы е): 3 (ж е л ты е ): 1 (зеленые). Э то объясняется эпистатическим
действием аллеля А (белая окраска) на аллели В (ж елтая окраска) и Ъ
(зеленая окраска).
Рецессивным
эпистазом
объясняется
так
называемый
б о м б е й с к и й ф е н о м е н — необычное наследование антигенов
системы групп крови АВО. В зависимости От наличия в организме
определенных эритроцитарных антигенов все люди разделены на
4 группы (табл. 9).
Таблица
Г р у п п а крови
I
II
III
IV
9.
Группа крови системы АВО
Эр н тр о ц и тар и ы й антигеи
0
.
А
В
АВ
А л ле л и , п р и су тс тв ую щ и е в генотипе
ЕЕ
ЕЕ
ЕЕ
ЕЕ
или
или
ЕЕ
ЕЕ
Синтез антигенов контролируется аллельными генами / Л, I й и / °.
Аллели /А и /в кодоминантны. Необходимо подчеркнуть, что рассмат­
риваемый ген имеет узкую норму реакции, в связи с чем его наличие в
организме неизбежно вы зывает синтез контролируемого им антигена
В приводимой родословной, иллюстрирующей «бомбейский феномен»,
обращает внимание женщина, унаследовавшая от матери аллель / в, но
имеющая 0 группу крови. Н а рис. 34 она отмечена стрелкой1 (расшиф­
ровка символов, применяемых при составлении родословных,приведена
на рис. 35). Активность аллеля /в в данном случае блокируется редким
рецессивным аллелем гена х в гомозиготном состоянии, который
оказывает эпистатическое действие.
При эпистазе модулирующее действие заключается в подавлении
одними генами функции других генов. Гены , оказывающие такой
эффект, называю тся и н г и б и т о р а м и или с у п р е с с о р а м и .
Гены, усиливающие функции других генов, называю тся интенсификаторами.
Ещ е одной формой взаимодействия неаллельных генов является
к о м п л е м е н т а р н о с т ь . Она заклю чается в том, что развитие
признака тр ебует. наличия в генотипе доминантных аллелей двух
определенных генов. В присутствии доминантного аллеля лиш ь одного
из них признак не воспроизводится.Так, мыши «агути», составляющие
большинство в природных популяциях, имеют серую окраску шерстно­
го покрова. Такая окраска развивается вследствие взаимодействия
доминантных аллелей двух генов, один из которых (А ) контролирует
синтез пигментд. Рецессивные гомозиготы по этому локусу являю тся
альбиносами. Другой ген (В ) обеспечивает скопление пигмента преиму­
щественно в основании и на конце волоса. Рецессивные гомозиготы по
этому локусу характеризуются сплошным разделением пигмента по
длине волоса. Воспользуемся решеткой Пеннета и рассмотрим
скрещивание мыш ей, дигетерозиготных по генам А и В (рис. 36).
Расщепление гибридов по признаку окраски шерстного покрова
происходит в отношении 9 («агути ») : 7 (3 с черной ш ерстью + 4 с
белой ш ерстью ). Таким образом, мыши «агути» действительно имеют
генотипы А —/В —.В скрещиваниях мышей, гетерозиготных по рассмат­
риваемым генам, возникают также «черные» и «белые» мыши с
генотипами А —/ЬЪ и аа/В— Отбор в природных условиях благоприят­
ствует окраске «агути», которая является покровительственной.
У человека комплементарным действием обладают гены пигмента­
ции волос. Ген М контролирует образование черного пигмента
меланина и представлен тремя аллелями М Вк, М в" и М ва. Первый
аллель обусловливает синтез значительных, второй — промежуточных
и третий — малых количеств пигмента. Последний аллель нельзя
путать с аллелем альбинизма, локализующимся в другом локусе.
У гомозигот по аллелю альбинизма пигмент вообще не образуется.
Доминантный аллель (Я 2) второго гена контролирует образование
красного пигмента. Сочетания аллелей названных генов даю т весь
1 Здесь и далее римскими цифрами обозначены последовательные поколения,
арабскими — потомки одного поколения.
90
□
1
О
о
10
О тО
А ,В
Хх
I
1
©
оо
Хх
Рис. 34. Родословная, иллюстри
рующая наследование «бомбей­
ского феномена». Объяснение
тексте.
ш
б^Ъс^Ь
9
©
13
14
□ Уо'
X
Рис. 35. Символы, используемые при составлении родословных.
1 — лицо мужского пола; 2 — лицо женского пола; 3 — пол неизвестен; 4 — брак; 5 — родственный
брак; 6 — сибсы (братья, сестры ); 7 — монозиготные близнецы; 8 — дизиготные близнецы; 9 — выки­
дыш; 10 — аборт; 11 — мертворожденный, 12 — бездетный брак, 13 — гетерозиготная носительница аллеля
в Х-хромосоме; 14 — умершие; 15 — пробанд (лнио, в родословной которого впервые обнаружен
признак).
спектр окрасок волос, наблюдаемых у человека. Наряду с этим при не­
которых комбинациях их появляю тся дополнительные признаки, такие
как лоснящиеся или глянцевитые волосы (рис. 37).
Результатом рассмотренных форм взаимодействия аллельных и
неаллельных генов служ ат стереотипные варианты наследования
признаков, которые легко вы являю тся гибридологическим методом.
Появляется все больше доказательств том у, что кроме таких форм
взаимодействия генов имеется зависимость функциональных проявле­
ний любого гена от особенностей генотипа в целом. По-видимому,
каждый ген обладает в той или иной мере свойством плейотропии, т. е.,
кроме генетического контроля развития определенного признака, он
оказывает модулирующее влияние на другие гены и сам испытывает
аналогичные влияния со стороны последних.
^
Ш ироту генных взаимодействий необходимо учиты вать при анализе
% .генетических явлений. Показателем зависимости функционирования
5 Унаследственных задатков от характеристик генотипа служит экспрес­
сивность и пенетрантность генов. Э к с п р е с с и в н о с т к з я к т о ч я ется в изменчивости количественного выражения признака у разных
особей — носителей соответствую щ его аллеля. И з рис. 38 видно, что
большинство организмов-носителей имеет среднюю степень развития
признака. В направлении меньших и больших значений относительное
.
число особей снижается, р о д п е н е т р а н т н о с т ь ю п о н и м я ю т
!
Способность гена обеспечить развитие признака до такой степени, к л г п я
.егоудЗеТся оонарУжить с ппмпшып имеющихся методов обследования
(рис. 38 1. Пенетрантность измеряется процентом организмов, имеющих
91
Гам еты
ссоо
<
АВ
АВ
АаВЬ
АЬ
А А В В ААВЬ
аВ
аЬ
АаВВ
АаВЬ
АЬ
А А В Ь 'А А Ь Ь / А а В Ь 'А а Ь Ь '
аВ
АаВВ
аЬ
О
агути
черны е
Рис. 36. Комплементарное
действие генов в наследо­
вании окраски шерстного
покрова мышей. О бъясне­
ние в тексте.
V,
АаВЬ
белые
;ааВЬ<
ГааВ В V
VVVV
А а В Ь /АаЬЬ- Й ш ВЬ ; >ааЬЬ<
ч/УУУУ
М еланин:
Н р а с н ы й п и гм ен т
в, =-
МЬЬМЬ«
Блондин с
р ы ж е в а ты м
о ттен ком
Р г\ К О
К ,К ,
Яркорыжий
С ве тл ы й
ш атен
Ш атен с
р ы ж е в а ты м
Темно­
рыж ий
М Ь Ь М Ы< м Ь « м Ь «
Ш атен
Н а ш та н о в ы й
Т ем н о-р ы ж и й
мЬ»мЬк
,мЬкмЬк
К2к2
Темны й
ш а те н
Т е м н ы й ш ате н "Темны й
(лоснящ и еся
ш ате н
волосы )
«Доминирование:
К.К,
Брю нет
Брю нет
(лосн ящ и еся
(во л о сы
с глянцем)
волосы )
Брю нет
(во л о сы
с глянцем)
М ь^ > М*Эуу> Р 2 >
Рис. 37. Комплементарное
действие генов у человека.
Объяснение в тексте.
Рис. 38. Экспрессивность
и пенетрантность генов.
Степень проявления (экспрессивность)
а — часть популяцнн, в которой
признак не обнаруживается; б —
группа лнц нз данной популяции
с выраженностью признака на
грани разрешающей способно­
сти методов его обнаружения;
в — группа лиц из данной по­
пуляции. имеющих признак, вы­
раженный в разной степени.
в Фенотипе признак, от общего количества обследованных носителей
соответствующ его аллеля. Экспрессивность и пенетрантность отража­
ю т зависимость Ф у н к ц и и гена от особенностей генотипа и проявляются
^процессе развития признака. Следовательно, в основе этих генетиче­
ских явлений может лежать колебание активности самих генов,
х а р а к т е р взаимодействия п р о д у к т о в генной активности, особое сочета­
ние условий среды в онтогенезе организмаГ 1
5.2.6. Функционально-генетическая классификация генов
Сущ ествует несколько классификаций генов. Некоторые из них,
например подразделенние на аллельные и неаллельные гены, были
приведены ранее. К другим мы обратимся в дальнейшем (например,
выделение летальных и полулетальных генов). Характеристики гена как
единицы функции наследственного материала и системный принцип
организации генотипа отражены в функционально-генетической класси­
фикации наследственных задатков, приведенной на схеме 2.
Сх е ма 2
Гены
.
Г
1. Структурные:
а) кодирующие аминокислотные
последовательности структур­
ных (коллаген) и фермента­
тивных белков;
б) кодирующие аминокислотные
последовательности
белков,
функционирующих во всех
клетках (например, рибосомных, гистонов);
в ) кодирующие последователь­
ность нуклеотидов в молеку­
лах р Р Н К и т Р Н К .
1
1
II. Модуляторы:
а) ингибиторы
или
су­
прессоры (см. «эффект
положения» или эпи­
стаз);
б) интенсификаторы (н а­
пример,
гены-мутаторы, повышающие час­
тоту мутаций у соответ­
ствующей особи);
в) модификаторы
комплементарные
ны).
•
I
III. Регуляторы
(см.
ге­
С т р у к т у р н ы м и называю тся гены, контролирующие разви­
тие конкретных признаков. Продуктом первичной активности гена
является либо и Р Н К и далее полипептид, либо р Р Н К и т Р Н К . Таким
образом, структурные гены содержат информацию об аминокислотных
или нуклеотидных последовательностях макромолекул. Структурные
гены трех подгрупп, приведенных в классификации, отличаю тся
степенью плейотропного действия, причем выраженная плейотропия
отличает гены второй и третьей подгрупп, которые активно
функционируют во всех клетках. При их мутациях наблюдаются
разнообразные и обширные нарушения развития организма. Неслучай­
но поэтому эти гены представлены в генотипе в количестве нескольких
десятков копий и образованы среднеповторяющимися последова­
тельностями Д Н К .
Г ен ы -м о д у л я т о р ы
смещ ают в ту или иную сторону
процесс развития признака или другие генетические явления, например
93
частоту мутирования структурных генов. Ч асть структурных генов
выполняет одновременно и роль модуляторов (см . пример «эффекта
положения»). Другие гены-модуляторы, по-видимому, лишены какихлибо иных генетических функций. Появление таких генов в эволюции
имело большое значение. Благодаря плейотропному действию многие
структурные гены, наряду с благоприятным и необходимым для
нормального развития организма действием, оказываю т и нежела­
тельные эффекты, которые снижаю т жизнеспособность особи. Небла­
гоприятное действие их ослабляется генами-модуляторами.
К регуляторным относятся гены, координирующие активность
структурных генов, контролирующие время включения различных
локусов в процессе индивидуального развития в зависимости от типа
клеток многоклеточного организма, а такж е от состояния среды.
5.2.7. Полигенная наследственность
И з изложенного выше следует, что наследование многих признаков
зависит о т передачи потомку дискретных единиц-генов, т. е. является
м о н о г е н н ы м . Однако еще Г . Мендель обнаружил, что изменчи­
вость окраски цветов гороха можно объяснить лишь одновременным
контролем этого признака несколькими наследственными задатка­
ми.
Позже
бы л
описан
феномен
полигенной
на­
следственности
или п о л и м е р и и . Он заклю чается в
том, что два или более генов в виде своих доминантных аллелей содру­
жественно и в одинаковой степени контролирую т развитие одного
признака.
Такие
гены
назы ваю тся
множественными,
п о л и м е р н ы м и или п о л и г е н а м и и обозначаются одной
буквой латинского алфавита с цифровыми индексами. Полигенное
наследование наблюдается при скрещивании гибридов 1-го поколения,
полученных от скрещивания пшениц с красными и белыми зернами
(рис. 39). Указанный признак контролируется двумя генами, доми­
нантные аллели которых (>1, и А 2) обеспечивают синтез пигмента.
Гибриды 2-го поколения расщепляются на растения с пигментиро­
ванными и непигментированными зернами в отношении 15:1. При этом
пигментированные зерна отличаю тся у разных гибридов по интенсив­
ности окраски, которая варьирует от темно-красной через промежу­
точные оттенки до бледно-розовой. П о интенсивности красной окраски
зерна разных гибридов образую т непрерывный ряд. Максимальная
интенсивность окраски наблюдается у доминантных дигомозигот
(А,А*4 ^2), а минимальная — у растений, в генотипе которых имеется
один из доминантных аллелей рассматриваемых генов — А , а, а2 а2
или аха,А 2а 2.
Полимерия представляет форму взаимодействия неаллельных
генов, важной особенностью ее является аддитивность (суммирование)
влияния на развитие признака. Благодаря этом у полигены представля­
ю т эффективный механизм генетического контроля развития количе­
ственных признаков. При моногенном наследовании признака возмож­
ны 3 варианта «дозы» гена в генотипе — А А , А а и аа. При полигенном
определении в генотипе диплоидного организма «собирается» 4 и более
94
«доз». Суммирование дей­
ствия и варьирование коли­
чества доминантных алле­
лей полигенов обеспечивает
непрерывное изменение ко­
личественного признака.
Рассмотрим следующий
пример (рис. 40). Пигмен­
тация кожных покровов
зависит от образования в
организме человека мела­
нина
и контролируется
моногенно. Доминантный
аллель обусловливает син­
тез пигмента особыми клет­
ками, тогда как рецессив­
ные гомозиготы являю тся
альбиносами. Генетический
контроль количества обра­
зуемого меланина осущест­
вляется независимо четырь­
мя полимерными генами1.
При наличии доминантных
аллелей этих генов синтези­
руется много пигмента, а
рецессивных — мало. Не; альбиносы как африканцы,
| так и представители кавказ!'1 ской (европеоидной) расы
имеют в генотипах аллели
: синтеза меланина, поэтому
кожа и тех и других пиг;Ментирована.
О. Интенсивность пигмен­
тации зависит от коли­
чества образуемого пигмен­
та. Она максимальна у
африканцев, которые явля­
ю тся доминантными тетра­
гомозиготами, и минималь­
на у европеоидов — рецес­
сивных тетрагомозигот по
соответствую щ им
локу­
сам. В связи с вышеизло­
женным в браках африкан-
1 В литературе есть указа­
нии на то, что этот признак конт­
ролирует 3, 5 и более локусов.
Исходные
К р асн ое зер н о
родители
А ] А 1А 2 А 2
Гибриды 1-го
поколения
•
Белое зерно
а1 а | а 2 а2
'
К р а с н о е зерно
А , а , А 2 а2
Гаметы гибридов
А ^А 2
а1 ^ 2
А |а2
а 15 2
----------- Л ^
^
А 1А 2 А] А |А 2А 2,А|А| А'2а2 А |а|А ^ А 2 А|*а| А 2 а2
9
А 1а 2 А1АТА2 ? 5 А |А] а2 а2 А 1
А 2 а2 6.1а1а 2 а 2
***
а 1А2
А1а 1А"2^2 А) а |А 2 &2 а|"а 1А 2 А"2 д| а, А 2 а 2
а, а2
А] а | А 2 а 2 3| в] А 2 а 2
а |А 2 а 2 Э|й| а 2 а 2
Пигментированные
□
Непигментированные
Рис. 39. Полигенное наследование интенсивности
окраски зерен пшеницы. Объяснение в тексте.
Генотипы
светлый—Р ( Р| Р 2 р 2 р З р З р 4 р4
темный—Р (Р 1 Р 2 Р 2 Р 3 Р 3 Р 4 Р 4
м у л а т - Р |р |Р 2 Р 2 Р з Р 3 Р 4 Р 4
Р | Р | Р 2 Р 2 Р З р З Р 4 р 4 * Р |р |Р 2 Р 2 Р 3 Р 3 Р 4 Р 4
число Р - 8
7
число р —О
1 2
6
5
4
3
2
1 0
3
4
5
6
7
8
Фенотип
темный —*
пром еж уточны й—
светлый
Рис. 40. Полигенное наследование интенсивности
пигментации кожи человека. Объяснение в тек­
сте.
Р 1—♦ — доминантные, Р| 4 — рецессивные аллели генов,
контролирующих интенсивность пигментации кожн.
95
цев и представителей европеоидной расы рождаю тся м улаты — тетрагетерозиготы по полигенам синтеза меланина, имеющие по сравнению с
родителями промежуточные значения интенсивности пигментации
кожных покровов.
Благодаря независимому комбинированию аллелей четырех поли­
мерных генов количество доминантных аллелей в генотипах детей в
браках мулатов варьирует в диапазоне от 0 до 8. В таких браках
рождаются дети с пигментацией кожи о т максимально светлой до
максимально темной. При этом вероятность появления крайних
вариантов пигментации наименьшая, а варианта, близкого к родитель­
скому, — наибольшая.
В приведенном примере обращает внимание то, что развитие
некоторых признаков находится под двойным генетическим контролем.
Воспроизведение в потомке определенного свойства, качественной
характеристики организма контролируется моногенно, тогда как
степень развития этого свойства (количественная характеристика)
контролируется полигенно. Таким образом, моно- и поли генная
наследственность дополняют друг друга. Подобно генам менделирующих признаков полигены включены в единую систему генотипа и
участвую т в разнообразных генных взаимодействиях.
Полигенное наследование отличается следующими типичными
чертами:
1) изменчивость количественных признаков зависит от варьирования
числа доминантных аллелей полимерных генов у отдельных особей;
2) мерой изменчивости таких признаков является амплитуда
варьирования;
3) пределы колебаний степени выраженности количественного
признака находятся под генетическим контролем;
4) амплитуда колебаний тем значительней, чем больше локусов
образую т полиген.
Полимерное наследование характерно для таких признаков
человека, как рост, масса тела, телосложение, умственные способности.
Полигенно в типичных случаях наследуются количественные
признаки. Имею тся примеры полигенного наследования формально
качественных признаков, например формы стручка у пастушьей сумки.
Определенная форма биологической структуры возникает обычно
вследствие неравномерного роста ее частей. Возможно, что наблюдае­
мая форма стручка возникает благодаря полигенному генетическому
контролю количественных показателей роста.
5.2.8. Наследственность и наследование.
Типы и варианты наследования признаков
Термины «наследственность» и «наследование» не являю тся
синонимами. Под н а с л е д с т в е н н о с т ь ю
понимается сово­
купность генетических механизмов, которые обеспечивают структурно­
функциональную преемственность организмов в ряду поколений. Э ти
механизмы вы текаю т из принципов организации наследственного
96
материала на генном, хромосомном и геномном уровнях, а также
закономерностей поведения генов в процессе гаметогенеза и размноже­
ния.
Под
наследованием
понимают
процесс
вос­
произведения
в последовательных
поколениях
общего
плана
структурно-функциональной организации и отдельных признаков у
особей конкретного биологического вида. Наследование — это
внешнее выражение
наследственности
в
том
смысле,
что
механизмы
наследственности обусловливают правила
(законо­
мерности) наследования. Э то хорошо иллю стрирую т результаты
исследований
Г . Менделя,
который
в
своих
опытах
на­
блюдал внешние проявления генетической связи между растениями
разных поколений и сформулировал три правила независимого
наследования признаков. О т правил он пришел к формулировке законов
наследственности, в которых отразил закономерности организации и
динамики генетического материала — дискретность и аллельное
состояние генов, генетическая «чистота» гамет и т. д. Правильное
отношение к понятиям «наследственность» и «наследование» особенно
важно для врача, так как в повседневной практике он первоначально
встречается с проявлениями наследования. Обратив внимание на
определенный признак пациента, врач сравнивает его состояние с
таковым у родителей и детей больного. Н а этапе диагноза
наследственного заболевания и прогноза развития нежелательного
признака в потомстве врач обращается к механизмам наследственности.
Выделяю т несколько основных типов наследования признаков,
каждый из которых имеет варианты.
Учиты вая особенности генетического контроля качественных и
количественных признаков, в отношении отдельных признаков различа­
ю т моногенное и полигенное наследование (схема 3, а), о которых шла
Схема ^
а
Наследование
1....
......
Моногенное
4
Полигенное
1
4
4
Сцепленное с половыми
хромосомами
Аутосомное
Г
Доминантное
1
Рецессивное
4 .
X-сцепленное
4
V-сцеп ленное
1
Рецессинное
Доминантное
6.
Наследование
4-------------------------Независимое
Частично сцепленное
4 — 56
,
Полностью сцепленное
97
с
к(
2
О
с
[
2
С
<
речь ранее. Моногенное насле­
дование бывает аутосомным
или сцепленным с полом.
Аутосомное и сцепленное с
Х-хромосомой наследование
может бы ть доминантным и
рецессивным. Названные ти­
пы и варианты соответствую т
правилам наследования от­
дельно взятых менделирующих признаков. Закономерно­
сти соотносительного насле­
дования нескольких признаков
описываются такими типами,
как независимое, частично
сцепленное и полностью сцеп­
ленное
наследование
(схе­
ма 3, б).
Типы и варианты наследо­
вания, свойственные другим*
организмам, обнаружены и у
человека и могут бы ть проил­
люстрированы родословными
людей. А у т о с о м н о д о м и н а н т н ы й вариант
наследования обусловливает­
ся передачей в ряду поколений
доминантного аллеля гена,
локализующегося в аутосоме,
для которого характерны оп­
ределенные черты, проявляю­
щиеся в родословных:
1) при достаточном числе
потомков признак обнаружи­
вается в каждом поколении;
2) редкий признак наследу­
ется
примерно
половиной
детей;
3) потомки мужского и
женского пола наследуют при­
знак одинаково часто;
4) оба родителя в равной
мере передают признак детям.
Отмеченные черты характери­
зую т, например, наследование
признака «шерстистых волос»
появившихся несколько поко­
лений тому назад в одной из
норвежских семей (рис. 41).
А у т о с о м н о - р е ц е с с и в н ы й вариант наследования обус­
ловливается передачей в ряду поколений рецессивного аллеля гена,
локализующегося в аутосоме. Типичные черты этого варианта,
выявляемые путем анализа родословных, сводятся к следующему:
1) даже при достаточном числе потомков признак может
отсутствовать в поколении детей, но появляется в поколении внуков;
2) признак может развиться у детей при отсутствии его у обоих ро­
дителей; среди детей, рождаемых в таких семьях, признак обнаружива­
ется в 25% случаев;
3) признак наследуется всеми детьми, если оба родителя имеют его;
4) признак развивается у 50% детей, если он обнаруживается у одно­
го супруга;
5) признак наследуется потомками мужского и женского пола
одинаково часто.
Перечисленные особенности проявляю тся в исследовании альби­
низма (рис. 42).
•собенностью аутосомно-доминантного наследования при не­
полном доминировании и кодоминировании служ ит менее выраженное
развитие признака у потомков — гетерозигот по сравнению с доми­
нантными гомозиготами. Так, изменения скелета у лиц с доминантным
аллелем брахидактилии (короткопалость) более выражены у гомозигот,
чем у гетерозигот. У людей, гетерозиготных по локусу синтеза р-полипептида гемоглобина (НЪА/НЫГ), содержание нормального гемоглобина
составляет 55—7 5 % .
[Особенности вариантов наследования признаков, с п е п л е н н ы х
с п о л о м ' , отражаю т факт локализации соответствую щ их генов в
■1ЮЛ0ШХ Хромосомах X и V . Напомним, что эти хромосомы за редким
исключением не имеют гомологичных л о к у с о в (р и с . 4 3 ). п о э т о м у по
больш инству генов Х-хромосомы клетки мужского организма гемизиготны___
Вариант наследования при локализации гена в У - х р о м о с о м е
(голандрическое наследование) отличается п р я м о й передачей признака
от отца всем сыновьям и соответствую щ им обнаружением этого
признака в каждом поколении у потомков мужского пола. Такое
наследование иллюстрирует родословная, приведенная на рис. 44.
В настоящее время неизвестны признаки, для которых голандрическое
наследование было бы доказано безоговорочно. М ожно думать, что так
наследуется признак «волосатых ушей» (гипертрихоз края ушной
раковины), который проявляется в том , что в юношеском возрасте на
ушной раковине вырастает пучок длинных жестких волос.« Описана
родословная, согласно которой мужчины пяти поколений имели этот
признак.
Особенности варианта наследования, сцепленного с полом, при
локализации гена в Х-х р о м о с о м е зависят пт гр»«мчпг<*гог»гти
соответствую щ их локусов у мужчин и функционально-генетической
1 Наследование, сцепленное с полом, необходимо отличать от наследования,
ограниченного полом. Последнее обусловливается генами аутосом, но возможность
развития признака зависит от пола организма. Примером такого признака служит
тембр голоса: баритон и бяс наблюдаются только у мужчин.
4*
99
1
(УсУфффбасУфсУфбб
Рис. 42. Аутосомно-рецессивное наследование признака альбинизма.
I
9-
О б щ а я цветовая с л е п ота—
14-
П игментная н с е р о д е р м а —
2834-
Пигментный р етини т-------Геморрагичесний
диатез
•9
14
- 28
■34
Рис. 43. Схема гомологичных и негомологичных локусов половых хромосом человека.
а — Х-хром осома: за ш тр и х о в а н ы л о к ус ы , о т с у т с т в у ю ш и е в хромосоме V (кр асн о - зел е н а я сле п о та, гемоф и­
л и я, атр оф ия зр и те л ьн ого и е р в а );
б — V хромосом а: за ш тр и х о в а н ы л о к ус ы , о т с у тс т в у ю щ и е в Х-хромосоме (пер епо нки м еж д у п ал ьц ам и , «в о л о с а ты е у ш и » ), Н е за ш тр и х о ва н н ы е у ч а с т к и хромосом со о тв е т­
с т в у ю т го м о логи чн ы м л о к усам Ц и ф р ы у к а з ы в а ю т р ассто ян и е данного л о к у с а от л о к у с а , определяю щ его
пол ( в м о р га н и д ах ).
Рис. 44. Гипотетическая родословная наследования признака, сцепленного с У-хромосомой.
и н а к т и в я п им ппнлй и з Х-уррмосом у женшин. Е сли признак р е ч е нием,—
^УГот вариант наследования характеризуется следующими ""чертами:
П 'м уж чины наследуют этот признак чащ е^лем женщины; /2| девочки
'теюледуют такой признак только от о тц а;(3 )\ браках, где о'&а супруга
лишены признака, могут родиться дети,Ччмеющие его, при этом он
наследуется 50% сы н о вей ^ ) прослеживается тенденция к чередованию
поколений с больш им или мёйьшим числом мужчин, имеющих признак.
Отмеченные особенности рецессивного наследования, сцепленного с Xхромосомой, иллюстрирует родословная по гемофилии (рис. 45). Если
признак доминантный, вариант Х-сцепленного наследования отличает
эщие черты:/! ) женщины наследуют признак чаще, чем мужчины;
100
Г ерцог
Саксон-НобургГ отский
Э д уард
герцог Кентский
(1 7 6 7 - 1 8 2 0 )
^
А льб ер т[Э
I Елена
| Виктория
\(ж ена
Г ] Э д уард VII
''им пера-Ч -1Э дуард VII
тора Фрил
риха)
)идк_
Г
Г
I Герцог [Георг
□ В и н д - р VI <
зорский
I
ПринцО
Елизавета I
VIII
гесс^ кскаяА у^
__________
1 Кайзер П Георг V
Зильгельм II Т
VII
Виктория
(1819—1901)
Т
О
Г)
Т
I |«
Фридрих
■
[Генриха)
т
I|
I
I_.__г
Ирина
® (ж ен а □
V
Принц
М аунттбетон Сигизмунд
Бирманский
П русский
1 ген р и х
I
]А лексан -Л
I ' |
|[
О
Ш ®
Ш
©
®
®
й
" м"
а”
Й
^
г а И)
Николая II)
1
Алексей
ГГраф
пяг
I Леопольд
~
______
*
I
I1
V
1
^
|В и к то - |Леопольд1
|Леополь;
□ □
□ ®
ф Евге-И
Евгиея- И
□
[Алиса I
'
|Атлонская
(1 )^ -1
Руперт, Альфонс'
с*иол- виконт
С мит Трематон
Беатри
■
М орис
Т иия
|(жена Альф онса XIII)
О
|
5 ” !
Гон~
Гонсалес
I) |Ь © © □ © © 6 - 0
М аргарет
Хуан Софья
Карлос
Филипп
йбЕГЬ
Рис. 45. Наследование рецессивного
Виктории.*-
I
фЕ-у? т=и
5
признака, сцепленного с
Х-хромосомой
(гемофилия). Родословная английской королевы
Рис. 46. Наследование доминантного признака, сцепленного с Х-хромосомой (рахит,
не поддающийся лечению витамином б ) .
/ 2} если признак есть лишь у супруги, его наследуют либо все дети (м ать
^гомозиготна), либо половина детей безотносительно к их полу (м ать
гетерозиготна); (3 )\сл и признак наблюдается лиш ь у супруга, он
наследуется всеми"' детьми женского пола. Описанные особенности
имеют место в наследовании таких признаков, как «рахит, не
поддающийся лечению витамином Б » и «коричневая эмаль зубов»
(рис. 46).
В табл. 10 приведено абсолютное число менделирующих признаков
человека, для которых к 1975 г. был установлен вариант наследования.
В скобках указано количество признаков, для которых вариант
наследования предполагается. Значительное их число объясняется
объективными трудностями выявления типа и варианта наследования.
Характерные для каждого варианта черты воспроизводятся в полном
объеме и достаточно отчетливо лиш ь в типичных случаях.
Таблица
10.
Менделирующие
Ф е н о ти п ы
признаки
у
человека
Ч и сл о опи санны х ф енотипов со о тв е т­
с тв у ю щ е го в ар иан та н аследования
Аутосомно-доминантные
Аутосомно-рецессивные
Сцепленные с Х-хромосомой
Всего .
583
(+
466
( + 4 8 !)
93
1142
635)
( + 78)
(+ 1 1 9 4 )
2336
Неполное доминирование, меняющ аяся экспрессивность и непблная
пенетрантность аллеля, зависимость развития признака о т гормо­
нального фона, возраста организма и действие факторов окружающей
среды способны затуш евать картину.
Дополнительные трудности в определение варианта наследования
вносят г е н о к о п и и — случаи, когда одно и то же состояние
признака развивается под контролем разных генов. Так, фенилкетонурия возникает при дефиците синтеза как фенилаланингидроксилазы, так
102
•л
А
П
;
\
\
\
(АВу
(аВ}
2 5 /.
2 5 /,
25%
+
+
+
['аь']
Г*4]
(О )
I
и
/
,
\
1
I
тип
тип
©
50/
Юз:
+
+
+
(аь''
ГаЬ4]
('аь']
50,
В
Р е н о м - Р о д и те л ь с к и е Р е к о м ­
бинант
типы
бинант
©
(дв;
25%
г
1 1 1 1 1
АаВ Ь
АаЬЬ
ааВЬ
ааЫ)
АаВЬ
ааЬЬ
ааВЬ
25/
25/
25%
25 /
50%
50/
Ю%
40/,
40)..
©
10/
+
в
1 I
["аЬ4)
I
АаВЬ
ааЬЬ
АаЬЬ
407
40%
10%
Рис. 47. Схема независимого, полностью и частично сцепленного наследования
признаков при дигибридном скрещивании.
а
н езави си м о е, б
п олно стью , в
ч а сти ч н о сцепленное наслед ован ие п р и зн ак о в А и В ; I - гены
пр изн аков А и В р а сп о л а г а ю т ся в негом ологичны х ( а ) хромосомах или на разном р асстоян и и в одной
хромосоме (б , в ) ; 2
ча с то та о б р а зо ва н и я гам ет с р азн ы м со ста во м ал л ел е й ( в п р о ц е н тах ); 3 — ан ал и ­
зи р ую щ е е ск р е щ и ва н и е с о соб ью , о б р азую щ е й один тип гамет- 4
соотн о ш ени е потом ков с разным и
генотипам и, р о ж д а ю щ и х с я в р е зу л ь та т е ан ал и зи р у ю щ его скр е щ и ва н и я.
и других ферментов, например дегидроптеридинредуктазы и, возмож­
но, дегидрофолатредуктазы.
Н а схеме 3,6 дана классификация основных типов соотносительного
наследования признаков.
Н е з а в и с и м о е н а с л е д о в а н и е отличается случайным
комбинированием признаков родителей у потомков. Оно свойственно
признакам, которые контролируются генами из негомологичных
хромосом, а также находящимися в одной хромосоме на расстоянии
более 50 морганид. У человека независимо наследуются, например,
способность ощ ущ ать вкус фенилтиомочевины и выделение в слюну
эритроцитарных антигенов системы
групп крови
АВО. При
полностью
сцепленном
наследовании
соответ­
ствующ ие признаки родителя развиваются у потомка всегда одним
блоком. Так наследуются признаки, контролируемые генами из одной
хромосомы, которые расположены настолько близко, что это
исключает кроссинговер. Полностью сцепленное наследование наблю­
дается для таких признаков, как синтез полипептидов Р- и 5гемоглобина человека. При ч а с т и ч н о м
сцепленном
н а с л е д о в а н и и соответствую щ ие признаки родителей воспро­
изводятся у части потомков совместно, а у части — независимо. Такое
наследование наблюдается для генов одной группы сцепления,
расстояние между которыми в хромосоме допускает регулярный
кроссинговер. Так, у человека гены, контролирующие выделение в
слюну антигенов системы групп
крови АВО и системы ЬщЬегап,
расположены в одной хромосоме на оасстоянии 15 морганид. Э ти
признаки наследуются по частично сцепленному типу.
103
Рис. 47 поясняет
хромосомные механизмы независимого,
полностью сцепленного и частично сцепленного соотносительного
наследования признаков.
5.2.9. Фенотип организма.
Факторы, определяющие развитие фенотипа
Совокупность признаков и свойств особи составляет ее
ф е н о т и п . Фенотип складывается в процессе индивидуального раз­
вития. Он соответствует тому типу структурно-функциональной
организации, который свойствен данному биологическому виду.
Фенотип развивается в соответствии с наследственной информацией,
которая содержится в генотипе. При этом отдельные гены обусловли­
ваю т лиш ь возможность развития признаков. Э та возможность
осущ ествляется при наличии подходящих условий внешней среды.
В н е ш н я я с р е д а вклю чает всю совокупность негенетических
(т. е. не связанных непосредственно с наследственным материалом)
факторов, действующих на организм в процессе его развития и
жизнедеятельности. В зависимости от изменений внешней среды
состояние сложных признаков варьирует от организма к организму.
Такие вариации называю тся м о д и ф и к а ц и я м и . Они имеют
приспособительное значение, а диапазон модификаций каждого призна­
ка находится под генетическим контролем. Так, пределы изменения
количества эритроцитов в периферической крови человека в зависимо­
сти от величины парциального давления кислорода в воздухе
ограничены генетически. То или иное значение количества красных
кровяных клеток в пределах возможных колебаний зависит от вы соты
местности над уровнем моря.
Взаимодействие генов и факторов окружающей среды составляет
основу развития как отдельных признаков, так и фенотипа в целом. Э то
нашло отражение в таком генетическом понятии, как « н о р м а
р е а к ц и и » — специфический способ реагирования организма на
изменения внешней среды. Она зависит от видовых характеристик и
индивидуальных особенностей генотипа. По-другому норму реакции
определяют как весь спектр путей развития, которые возможны у
носителя конкретного генотипа в любой среде, совместимой с жизнью .
По отношению к разным признакам «норма реакции» бывает узкой и
широкой. В первом случае одинаковое состояние признака возникает в
широком спектре колебаний факторов среды. Во втором — признак
отличается значительной изменчивостью в зависимости от параметров
внешней среды. В качестве примера приведем соответственно систему
групп крови АВО и рост индивидуума. Рис. 48 дает представление о
диапазоне варьирования степени развития признаков с узкрй и широкой
нормой реакции в зависимости от генотипа.
Сходные состояния некоторых признаков возникаю т у одних особей
благодаря наличию в генотипе определенного аллеля, а у других — в
результате особого сочетания внешних факторов. Изменения фенотипа,
сходные с изменениями генетической природы, но вызванные фактора­
ми внешней среды, называю тся ф е н о к о п и я м и . Так, у женщин,
I
104
Степень развития признака
С тепень развития признака
Рис. 48. Степень развития признака у гомо- и гетерозиготны* организмов.
а — узкая, б — широкая норма реакции гена; аа и А А — гомозиготы; Аа — гетерозигота.
перенесших на ранних сроках беременности краснуху, нередко рожда­
ю тся дети с врожденной катарактой (помутнение хрусталика), не
отличимой о т наследственной катаракты.
* К основным факторам, от которых зависит фенотип организма,
относятся гены с присущими им свойствами, разного рода генные
взаимодействия и параметры внешней среды, в которой осущ ествляется
развитие. Проиллюстрируем действие этих факторов на примере
развития п р и з н а к а п о л а 1. У раздельнополых организмов среди
новорожденных соотношение числа особей мужского и женского пола
близко 1:1 (табл. 11).
Т а б л и ц а 11.
Соотношение полов* у некоторых раздельнополых организмов
прн рождении и в последующие возрастные периоды (для человека)
Вид организма, возрастной период
Голубь
Курица
Утка
Осел
Лош адь
Человек:
новорожденные
дети
юноши
50 лет
80 лет
Н а 100 особей ренского пола приходится
особей мужского пола
100
98
100
98
104
106
103
100
85
50
* Разли чаю т п е р в и ч н о е
соотношение
полов,
определ немое генетически иа уровне
зигот. В т о р и ч н о е с о о т н о ш е н и е п о л о в регистрируется на момент рождения, а т р е т и ч н о е —
в период половой зрелости. Н а уровне зигот соотношение мужского и женского вариантов у человека
составляет 125— 135: 100.
У людей в среднем на каждые 100 девочек рождается 106 мальчиков.
Э то соотношение закономерно изменяется в последующие возрастные
периоды. Предположительно, отношение особей по полу, близкое к
единице, обеспечивает максимальную вероятность встречи самки и
самца, передачу более разнообразной наследственной информации
потомкам, поддержание оптимальной численности организмов в попу1 В данном случае целесообразно рассматривать пол как отдельный, хотя и
сложный признак.
105
Рис. 49. Латеральный гинандроморф плодовой мухи, гетерозиготной по локусу
окраски глаз, размещающемуся в X хромосоме. Л ев ая половина тела женского
гипа, правая
мужского; левый глаз красный (генотип X X ) , правый
белый
(генотип X V )
ляциях. Некоторые биологические виды характеризуются в репро­
дуктивном возрасте существенно измененными значениями этого
отношения, что зависит от особенностей их экологии. Известны,
например, безсамцовые линии божьих коровок. Причина, по-видимому,
заключается в биологической несовместимости цитоплазмы яйцеклетки
и генома спермия, если он содержит У-хромосому.
Закономерное воспроизведение из поколения в поколение расщепле­
ния в отношении 1:1 указывает на генетический контроль развития
признака пола. Наблюдаемое расщепление типично для скрещивания
типа А ах аа, при котором один родитель гетерозиготный, а другой —
гомозиготный. Наблюдения цитологов, обнаруживших наряду с аутосомами половые хромосомы, вскрыли хромосомный м е х а н и з м
генотипического
определения
пола,
который
объясняет такое расщепление.
У животных откры ты 4 основных варианта этого механизма:
1) один из полов имеет пару разных половых хромосом (Х У ), а
второй — одну половую хромосому (Х0)\ 2) один из полов имеет
одинаковые хромосомы (X X ), тогда как другой — разные (А Т );
3) развитие по женскому или мужскому типу зависит о т соотношения
количества Х-хромосом и числа наборов аутосом (А ); комбина­
ция X X + 2А дает женский фенотип, Х У + 2 4 — мужской; 4) один пол
(самки) первично диплоидный, а противоположный ему — первично
гаплоидный
)0 6
Ч
■
* *
ч*
«С
&
.V
V V * *« .
и * *\**
ч *
а
И
1
И
л
ИХ
2
в Н КХ
3
4
5
с XX XX хх хх хх хх кя
6
Рис. 50. Несоответствие
фенотипу
генотипу
по
признаку пола Механизм
развития неизвестен.
а
фенотип (м у ж с к о й ); б
нотип 46, X X (ж е н с к и й )
7
8
9
10
12
II А
АА
Ал
Дд
I «
дж
13
14
15
16
17
18
Я ■
Ж■
« А
А А
ЖА
19
20
21
22
XX
ге­
/
11
Первый варилн*г хромосомного механизма определения пола описан
у клопов рода РпЛепог, второй — у многих видов животных и человека,
третий — у плодовой мушки дрозофилы, четвертый — у некоторых
насекомых, например пчел. У парамеций обнаружен моногенный
механизм определения пола.
Эффективность генетического контроля развития пола подтвержда­
ется наличием в природе г и н а н д р о м о р ф о в . Э то животные, у
которых одна часть тела, включая половые железы, женского, а
другая — мужского типа. Н а рис. 49 изображена плодовая мушка,
являющ аяся латеральным гинандроморфом. Такие особи возникают в
случае утраты клетками правой половины тела одной Х-хромосомы в
107
процессе индивидуального развития, что
сдвигает соотношение количества Х-хромосом и наборов аутосом в сторону м уж­
ского типа. Клетки левой половины тела
сохраняют обе Х-хромосомы. Известны,
однако, примеры отклонения фенотипа
организма в сторону противоположного
пола, несмотря на комбинацию половых
хромосом (рис. 50, 51). Лиш ь у части
индивидуумов с истинным гермафроди­
тизмом обнаруживается мозаициз’м по
прловым хромосомам (габл. 12).
Описаны случаи закономерной смены
пола особи в жизненном цикле. Так, брю­
хоногий моллюск СгерккЛа р1апа после­
довательно проходит стадии самца,
гермафродита и самки. Если развитие
происходит в отсутствии самок, первая
ртадия укорачивается. Тихоокеанские
рыбки ТаЬго1с1е5 сНткйаШх обитаю т груп­
пами из нескольких самок и одного самца.
Если самец погибает, то у наиболее ак­
тивной самки (и -самки) происходит пол­
ная реверсия пола, включающ ая развитие
функционально полноценных семенни­
Рис. 51. Несоответствие фсиоти
ков. Наблюдения из практики животно­
па генотипу по признаку пола
водства и результаты некоторых экспе­
при синдроме Морриса,
риментов
показываю т, что расхождение
я
фенотип ( ж е н с к и й ),
фенотипа и генотипа по признаку пола
зависит от условий, в которых происхо­
дит онтогенез. Типичен пример фри-мартинов. Замечено, что у коров
в разнополых двойнях нередко рождаются нормальный бычок и
фенотипически интерсексуальная телочка (фри-мартин). В эмбрио­
генезе между близнецами часто устанавливается перекрестное крово­
обращение. М ожно думать, что вещества из организма бычка
попадают в кровоток телочки и вы зы ваю т маскулинизирующее
действие.
О пыт по переопределению пола с помощью гормонов был
произведен на аквариумных рыбках медаках (Огугтах (айрех), осоТаблица
12.
Генетическая
с истинным гермафродитизмом
Г е н е т и ч е с к а я к о н сти туц и я
46, XV
46. XX
46. X V /46, X X (мозаичная)
Всего
108
конституция
Ко ли чество случаев
У
6
2
17
лиц
м
«и
■ Л1
V 1
Ц>
Щ > ~?
в©\
|1|1
2
бН
\ )< 1
■1Л
3
4
V)
О0
) 0(1 и
О С)
8
9
10
пП
5
(\П
II
12
О ои
•О
ю
14
\Л
}с.
‘
А А
1А
Ю
1Э
/
17
1
П
о!'
1Я
(О
\>
г Х,\
л
19
У
Я
20
0X
. '.
21
X X
22
й *
Х У
1’ис. 51. Продолжение.
О
генотип 40
XV
(М У Ж С К О Й )
бенность которых заклю чается в локализации доминантного аллеля
красной окраски тела (К ) в хромосоме V , а рецессивного г — в
гомологичном локусе хромосомы X (рис. 52). В отсутствие кроссинговера, который происходит редко, у медак из поколения в поколение
воспроизводятся белые самки (Х г Х г ) и красные самцы (Х г V " ).
Выклю нувш имся малькам в течение 8 мес скармливали женский
половой гормон, в результате чего все поколение оказалось самками.
109
Примерно половина из них
были красными, т. е. имели
/ Х гх
X
мужской генотип (Х г V я ).
Наряду
с типичными вторич­
9
ными половыми признаками
у таких самок были яичники, и
в скрещиваниях с нормальны­
/Х'ХГ
ми самцами они давали по­
томство. При этом наблюда­
О пыг
лось необычное расщепление
по признаку пола — на одну
самку приходилось три самца,
что объяснялось выработкой
яйцеклеток с хромосомой V я.
В таких скрещиваниях появля­
лись и необычные по генетйческой конструкции самцы —
У ЯУ Я, которые скрещивались
с
нормальными
самками
(Х г Х г). При этом все потом­
ство было представлено сам­
цами (Х гУ к).
Пример с переопределени­
X
' Х ГХ Г
ем пола у ЬаЬго1йе5 сНткНаШк,
о чем шла речь выш е, под­
тверждает роль гормонов, в
частности
надпочечниковых
желез, в развитии признака
пола в природных условиях.
Рис. 52. Переопределение фенотипичных при­
У этих океанских рыб муж­
знаков пола у медак.
ской фенотип возникает у осо­
би, которая в сообществе себе
подобных испытывает наименьшее стрессовое напряжение. Этим
объясняется, что именно а-самка, занимающая в иерархической струк­
туре группы 2-ю ступень, заменяет погибшего самца.
Рассмотренные примеры говорят о влиянии на процесс формирова­
ния фенотипа условий развития организма и факторов внешней среды.
Синдром Морриса убеждает в зависимости этого процесса такж е от
генных взаимодействий. Такие люди, несмотря на мужской гено­
тип Х У , имеют в целом женский фенотип (см. рис. 51). Правда, у них
отсутствую т менструации и наблюдаются некоторые характерологиче­
ские и физические черты, присущие в большей мере мужскому полу
(реш ительность, предприимчивость, выносливость к физическим наг­
рузкам). Поначалу эмбриогенез лиц с синдромом Морриса соответству­
ет мужской генотипической конституции зиготы: закладка половой
железы развивается в семенник, клетки которого продуцируют мужской
половой гормон. Н о в силу гомозиготности по рецессивному аллелю Гйп
((тапхГогтег) в таких зародышах не образуется белок-рецептор,
который делает клетки развивающихся тканей и органов чувствиИ о н тр оль
9
9
но
тельными к гормону. В отсутствие взаимодействия продуктов активно­
сти различных генов — гормона и рецептора развитие по мужскому
варианту прекращается.
Развитие признака пола происходит в два этапа. Н а первом" этапе в
момент оплодотворения осущ ествляется о п р е д е л е н и е п о л а
будущего организма при помощи генотипического хромосомного
механизма. Н а втором этапе в ходе онтогенеза происходит
дифференциация
п о л а , т. е. формирование конкретного
фенотипа с учетом особенностей генотипа и условий развития. Н а этом
этапе вклю чаю тся такие факторы, как генные взаимодействия и
влияния внешней среды. При этом генотип дает общее направление
развития и определяет пределы модификаций признаков.
У человека, как и у других организмов, зигота потенциально бисек­
суальна. Главны м фактором, сдвигающим фенотип в мужскую сторону,
является У-хромосома. Выбор направления происходит на 6— 10 нед
эмбриогенеза. Если в указанный срок зародыш, имеющий У-хромосому,
не начал развиваться в мужском направлении, в дальнейшем он
приобретает женские вторичные половые признаки. Ш ирота «нормы
реакции» признака пола варьирует у представителей разных видов, чем
объясняется то, что возможность переопределения пола во взрослом
состоянии ограничена узким кругом животных.
5.2.10. Молекулярно-биологические представления
о строении и функционировании генов
М олекулярная биология как самостоятельное направление сложи­
лось в последние десятилетия. К настоящ ему времени ее идеи проника­
ли во все отрасли науки о жизни и определяют главные тенденции раз­
вития теоретической, экспериментальной и прикладной биологии.
Молекулярная биология складывалась в ходе исследований физико­
химических свойств и биологической роли нуклеиновых кислот и
белков. Основы ее заложены работами по генетике вирусов и фагов,
химической природе наследственного материала, механизму биосинтеза
белка, биологическому коду, закономерностям ультраструктурной
Таблица
13.
Состав
азотистых
оснований Д Н К
из разных
А зо ти сты е о сно ван и я
О р ган и зм
источников
+Т
А
г
Г+ Ц
Т
Ц
1,01
0,99
1,02
1,00
0,93
1,00
1,04
1,00
1,00
0,95
1,02
1,00
1,07
1,09
1,01
1,12
А
И ст о ч н и к Д Н К
Вилочковая железа
Геленок
Эритроциты
Курица
Сперма
Лосось
Все ткани
Морской краб
К л е то ч н а я
ДНК
Эвглена
Хлороиласты
Эвглена
Киш ечная палочка
Ф аг Т2
А
т
Г
Ц
29.0
28,8
29,7
47,3
22,6
38,2
24,7
32.5
28,5
29,2
29,1
47,3
24,4
38,1
23,6
32,5
21,2
20,5
20,8
2,7
27,7
12,3
26,0
18,2
21,2
21,5
20,4
2,7
25,8
11,3
25,7
16,8
1,351,39
1,42
8,75
0,88
3,23
0,93
1,88
111
организации клетки. В связи с
этим м о л е к у л я р н у ю
биологию
можно опреде­
лить как область изучения зако­
номерностей структуры и изме­
нений информационных макро­
молекул и участия их в фунда­
ментальных процессах жизне­
деятельности.
В области генетики молеку­
лярная биология вскрыла хими­
ческую природу вещества наслед­
ственности, показала физико­
химические предпосылки хране­
ния в клетке информации и точ­
ного копирования ее для переда­
чи в ряде поколений. Д Н К боль­
шинства биологических объектов
(о т млекопитающих до бакте­
Г уани н
риофага) содержит равные коли­
Ц и тсзи н
чества нуклеотидов с пуриновы­
Аденин
ми (аденин, гуанин) и пиримиди­
Тимин
новыми (тимин, цитозин) азоти­
Рис. 53. Реализация принципа комплемен­
стыми основаниями (табл. 13).
тарности в процессе редупликации ДНКЭ то означает, что объединение
молекул Д Н К в двойную спи­
раль осущ ествляется закономерно, в соответствии с п р и н ц и п о м
к о м п л е м е н т а р н о с т и — адениловый нуклеотид связывается
с тимидиловым нуклеотидом, а гуаниловый с цитидиловым (рис. 53).
Такая конструкция делает возможным п о л у к о н с е р в а т и в н ы й с п о с о б р е д у п л и к а ц и и Д Н К . Вместе с тем вдоль
биспирали Д Н К пары А — Т и Г — Ц располагаются случайным обра­
зом — А + Т ^ Г + Ц . Следовательно, п у т е м н е з а в и с и м о ­
го к о м б и н и р о в а н и я
нуклеотидов,
различающихся
по азотистому основанию, по длине молекул Д Н К удается записать
разнообразную информацию, объем которой пропорционален коли; честву нуклеиновой кислоты в клетке.
Согласно молекулярно-биологическим представлениям ген как
единица Функционирования наследственного материала характеризу­
ется "слож ны м строением. Многие детали тонкой структуры гена
остаю тся неизвестными. Вместе с тем успехи современной науки в этой
области достаточно велики, чтобы можно было нарисовать принципи/альную модель функционирующего гена.
Функциональная активность гена заклю чается в синтезе на молекуле
Д Н К молекул Р Н К или т р а н с к р и п ц и и
(переписывании)
биологической информации с целью ее использования для образования
белка. Единицы транскрипции ( т р а н с к р и п т о н ы ) превыш аю т по
размерам структурные гены (рис. 54). Согласно одной из моделей
транскиптона в
клетках эукариот
он
состоит
из
н е и н112
V
А к ц е п т о р н а я или
н еи н ф о р м ати вн ая
зо н а
П р о м о»тор
то р
С т |р у к т у р н а я или
инн ф о р м а т и в н а я
и
зо н а
А кц еп то р ны е
лонусы
V I х*
С тр уктур ны е
ге н ы
/*Л
-
*
Р И Н —п о л и м е р а з а
Регулято рны е
белки
Д еград ац ия
I
П еренос
в ц итоплазм у
Рис. 54. Схема регуляции транскрипции в эукариотической клетке. Объяснение в
тексте.
формативной
(акцепторной)
и
информатив­
ной
зоны.
Последняя
образована
структурными
генами
(ц и с т р о н а м и), которые разделены вставками Д Н К — с п е й с е рами^
не
несущими
информации
об
аминокислотных^
последовательностях белков. Неинформативная зона начинается
г е н о м - п р о м о т о р о м (р), к которому присоединяется фермент
РНК-полимераза, катализирующ ая реакцию ДНК-зависимого образо­
вания рибонуклеиновых кислот. Далее следуют акцепторные гены или
г е н ы - о п е р а т о р ы (о, , а 2 и т. д .), связывающ ие регуляторные
белки ( г г2 и т. д .), изменения которых «откры ваю т» Д Н К структур­
ных генов (8,, 82 и т. д.) для считывания информации. Н а транскрипто­
не синтезируется одна больш ая
молекула Р Н К .
Благодаря
п р о ц е с с и н г у неинформативная ее часть разруш ается, а инфор­
мативная расщепляется на фрагменты, соответствую щ ие отдельным
структурным генам. Э ти фрагменты в виде и Р Н К для синтеза
конкретных полипептидов транспортируются в цитоплазму. Согласно
приведенной модели в транскрипте находится несколько структурных
генов. Группа этих генов образует функциональный блок «“ называется
о п е р о н о м . Функциональное единство оперонов зависит о т наличия
генов-операторов, которые воспринимают сигналы из метаболического
аппарата цитоплазмы и активирую т структурные гены.
Природа сигналов, регулирующих функцию генов, изучена у
прокариот. Э то белки, синтез которых контролируется особыми
г е н а м и - р е г у л я т о р а м и , действующими на гены-операторы.
Активация структурных генов посредством генов-регуляторов и опера­
торов представлена на схеме (рис. 55). В обычных условиях ген-регу113
I
Рис. 55. Схема регуляции транскрипции структурных генов прокариотической клетки
по типу индукции.
лятор активен и в клетке протекает синтез белка-репрессора, который
связывается с геном-оператором и блокирует его. Э то выклю чает из
функции весь оперон.
Включение оперона происходит, если в цитоплазму проникают
молекулы субстрата, для переваривания которого требуется во­
зобновление синтеза соответствующ его фермента. Субстрат присоеди­
няется к репрессору и лишает его способности блокировать геноператор. В этом случае информация со структурного гена считывается
и требуемый фермент образуется. В описанном примере субстрат
играет роль и н д у к т о р а (побудителя) синтеза «своего» фермента.
Последний запускает биохимическую реакцию, в которой используется
данный субстрат. По мере снижения его концентрации освобождаются
молекулы репрессора, которые блокируют активность гена-оператора,
что приводит к выключению оперона. У бактерий описана система
регуляции, переводящая активные структурные гены в неактивное
состояние в зависимости о т концентрации в цитоплазме конечного
продукта определенной биохимической реакции (рис. 56). При этом под
генетическим контролем гена-регулятора образуется неактивная форма
репрессора гена-оператора. Репрессор активируется в результате
взаимодействия с конечным продуктом данной биохимической реакции
и, блокируя ген-оператор, выклю чает соответствую щ ий оперон. Синтез
фермента; катализирующего образование вещества, активирующего
репрессор, прекращается. Описанные системы регуляции функции
структурных генов носят приспособительный характер. В первом
примере синтез фермента запускается поступлением в клетку субстрата
соответствующ ей реакции, во втором — образование фермента прекра­
щается, как только исчезает потребность в синтезе определенного
вещества.
114
Рис. 56. Схема регуляции транскрипции структурных генов прокариотической клетки
по типу репрессии.
Принципы регуляции генной активности у эукариот, по-видимому,
сходны с таковыми у бактерий. Вместе с тем появление ядерной оболоч­
ки, усложнение генных взаимодействий в условиях диплоидности,
необходимость тонкой корреляции генетических функций отдельных
клеток многоклеточного организма повлекли за собой при переходе к
эукариотическому типу клеточной организации усложнение регуля­
торно-генетических механизмов, генетико-биохимические и кибернети­
ческие основы которых во многом еще не выяснены. Можно
предположить также, что в эволюции увеличилось число геновоператоров. Индукторами транскрипции многих структурных генов
эукариот служ ат гормоны. Предполагается наличие
генови н т е г р а т о р о в , включающих в ответ на стимул одновремен­
но «батареи генов». Генетическая система высш их организмов отли­
чается, по-видимому, большой гибкостью реакций на действие негене­
тических факторов. В подтверждение этого допущения рассмотрим
ряд факторов. Так, некоторые структурные гены животных не явля­
ю тся непрерывными последовательностями кодонов, а составлены
из фрагментов, которые прерываются неинформативными участка­
ми Д Н К . Ген Р-полипептида гемоглобина мыши,например, прерывает­
ся вставкой из 550 пар нуклеотидов. Соответствую щ ий этой вставке
участок отсутствует в зрелой глобиновой и Р Н К , что говорит о его
разрушении в ходе процессинга первичной транскрибированной Р Н К
с воссоединением информационных фрагментов и Р Н К . Информацион­
ные участки таких генов получили название э к з о н о в, «молча­
щие» — и н т р о н о в ,
а процесс воссоединения информационных
фрагментов
и РН К — с п л а й с и н г а
(сплавления). Количест115
^
во Д Н К в области нитронов в 5— 10 раз выше, чем в области экзонов.
Предполагается, что сплайсинг служит механизмом образования
некоторых генов в момент их функциональной активности, т. е. на
уровне и РН К .
Известны
также
«блуждающие»
структурные
г е н ы , положение которых в хромосоме меняется в зависимости от
фазы жизненного цикла. Так, «тяж елы е» и «легкие» полипептиды
иммуноглобулинов состоят из константного (С ) и вариабельного (У )
участков, синтез которых контролируется сцепленными, но разными
генами. В зрелых плазматических клетках эти гены разделены нетранскрибируемой вставкой длиной в 1000 пар нуклеотидов. В клетках
эмбрионов названная вставка во много раз длиннее. Таким образом в
процессе клеточной дифференцировки изменяется взаиморасположение
генов. Исследование механизмов регуляции генной активности и генных
взаимодействий у эукариот представляет важнейшую область совре­
менной молекулярной биологии и генетики.
5.2.11. Цитоплазматическая наследственность
Благодаря работам А . Вейсмана и Т . Моргана теорию наслед­
ственности эукариотических организмов называю т хромосомной. Эти м
подчеркивается факт размещения наследственного материала в хромо­
сомах клеточного ядра. По мере развития генетики накапливались
данные, необъяснимые с точки зрения исключительно ядерной
локализации генов и свидетельствовавшие о возможности прямого
участия
в
явлениях
наследственности
цитоплазмы.
Ц и т о п л а з м а т и ч е с к а я н а с л е д с т в е н н о с т ь обеспе­
чивается генами, локализованными вне ядра клетки. Ей соответствует
особый тип одностороннего наследования по материнской линии, при
кйтором’ признак передается через цитоплазму яйцеклетки. Сово­
купность
наследственных
задатков
цитоплазмы
называется
п л а з м о й о м , а сами задатки — п л а з м а г е н а м и . П о мате­
ринскому типу наследуется устойчивость к стрептомицину у хламидо­
монад, направление завитка раковины улиток, пятнистость листьев и
мужская стерильность некоторых растений. Уникальной остается
родословная, согласно которой в семьях трех поколений родилось
72 девочки и ни одного мальчика. Э то может бы ть объяснено
цитоплазматической наследственностью, хотя допустимы и другие
объяснения.
Плазмагены разнородны по своей природе. Их можно разделить на
две группы: 1) гены ДНК-содержащих органелл клетки (митохондрии,
хлоропласты); 2) инфекционные агенты или симбионты клетки (вирусы,
плазмиды, эписомы). Плазмагены обоих групп сходны по своим
Свойствам с ядерными генами и осущ ествляю т генетический контроль
синтеза ряда важных ферментов, а, следовательно, и развития
некоторых сложных признаков. Они способны к редупликации и
случайным, устойчивым, передающимся в ряду поколений изменени­
ям — точковым мутациям. В качестве примера рассмотрим плазмаге- .
т а и о р д ^ содержит 4— 5 кольцевых
молекул Д Н К , каждая длиной примерно в 15 ООО пар нуклеотидов. За
счет собственной генетической информации в митохондриях образу­
ю тся т Р Н К , рибонуклеиновые кислоты и белки рибосом, некоторые
ферменты аэробного энергетического обмена и структурные белки.
Д Н К митохондрий редуплицируется, вслед за чем происходит деление
исходной органеллы на две дочерние. Предположительно мутациями
митохондриальных генов объясняю тся такие пороки развития человека,
как 5 р та Ы йёа (раздвоенный позвоночный столб), сращение нижних
конечностей.
Генетический контроль структуры и функции митохондрий
плазмагены обеспечивают во взаимодействии с генами хромосом ядра.
Простой расчет показывает, что объем собственной наследственной
информации митохондрии недостаточен для воспроизведения всей
совокупности рибонуклеиновых кислот и белков органеллы. Многие
белки, особенно ферментативные, вклю чаю тся в структуру мито­
хондрии, будучи синтезированы в цитоплазме на и Р Н К , поступившей из
ядра. Описано явление генокопирования по ядерным и цитоплазматиче­
ским генам. Так, к мужской стерильности растений приводят в одних
случаях мутации ядерных генов, а в других — плазмагенов.
5.2.12. Генетический анализ.
Методы генетических исследований человека
И зучая наследственность и изменчивость, генетика использует
разнообразные методы. Специфическим для генетического анализа
является ( Г и б р и д о л о г и ч е с к и й
м е т о д , разработанный
Г . Менделем. О т обычных скрещиваний растений и животных с
последующим наблюдением за потомством этот метод отличается
следующим: 1) практикуется целенаправленный подбор родителей,
различающихся по одной, двум, трем и т . д. парам контрастных
(альтернативных) стабильных признаков; 2) в каждом поколениш ает
ведется отдельно по каждой такой паре; 3) этот учет носит
количественный характер и проводится на гибридах ряда последова­
тельных поколений; ^ осущ ествляется индивидуальная оценка потом­
ств а от каждого родителя в ряду поколений. '
Эффективное применение гибридологического метода зависит от
выбора подходящего объекта генетического анализа, который должен
легко скрещ иваться в условиях эксперимента, быстро достигать
состояния половой зрелости, давать большое число потомков, иметь
малое количество групп сцепления. Наследуемые признаки, взяты е для
анализа, должны, по возможности, иметь узкую «норму реакции», т. е.
бы ть устойчивыми к модифицирующим действиям внешней среды.
Именно этими качествами обладают бактериофаги, кишечная палочка,
некоторые самоопыляющиеся растения (в опытах Г . Менделя —
горох), кукуруза, гриб нейроспора, плодовая муш ка дрозофила, мыш ь.
Эффективность гибридологического метода в познании закономерно­
стей наследования и законов наследственности иллюстрируется ре­
зультатами опытов Г . Менделя по моно- и дигибридному скрещиванию
и Т . Г . Моргана по сцепленному наследованию. В практике генетиче117
ского анализа широко исс к р е ^ и _в а н и е, парт­
нерами в котором служат
гетерозиготный гибрид 1-го
поколения (А а) и один из
исходных
гомозиготных
родителей (А А или аа).
Особый интерес представ­
ляет вариант Р,А а х Раа
или ачн а л и з и р у ю ш е е
с к р е шТТваТГие ( р и с . 77).
В этом случае характер рас­
щепления потомства по фе­
нотипу указывает на ассор­
тимент гамет гибридного
организма, что дает воз­
можность проверить гетерозиготность
особей по
Рис. 57. Схема возвратного анализирующего
изучаемой
паре
генов в
скрещивания гибрида Р, с рецессивной родитель­
любом поколении Р ь Р2 ...
ской формой.
I — родители с пурпурной
( А а ) и б е л ой (а а )
окраской
Ри
цветка; 2 — га м е ты родителей ; 3 — ра сщ е п л е н и е п о то м к о в
Ч е л о в е к к а к о1
по г е н о т и п у и ф е н о т и п у .
(е к г г е н е т и ч е с к и
ШГ
с с л е д о в а н и й^) имеет и недостатки, и достс
человеческом обществе невозможны целенаправленны й подбор роди­
тельских пяР н здцдер м ш 1 хад щ щ браки. В настоящее время большин­
ство семей имеет не более 2—3 детей, что недостаточно для
корректного статистического анализа распределения признаков в потомстве. Продолжительность жизни одного поколения людей равна
25— 30 годам, "а Тмйнймальный пёриод жизни до наступления половой
зрелости — 13— 15 лет1 . Таким образом, генетик “Не может наблюдать
Непосредственно более' 1—2 поколений. У человека много «групп
сцепления» (23 против 4 у дрозофилы). Л ю д Д отдичаех ..большое
(Щствённоё разноббрЗЖег~~Если допустить,' что каждая пара
гомологичных хромосом содержит всего по паре аллелей, то и тогда
благодаря независимому комбинированию хромосом в мейозе гетерози­
готные особи могут дать 8 388 608 разных типов гам ет. Однако
количество локусов только в Х-хромосоме составляет 92. Суммарное
число генов в геноме человека оценивается величинами 104- 10*.
Генетическое разнообразие и значительная вариабильность экологиче­
ских и социальных условий жизни приводят к большой фенотипической
изменчивости людей. Несмотря на трудности в настоящее время,
человек служит объектом интенсивных генетических исследований.
Преимущество человека как объекта исследования перед другими
живыми организмами заклю чается в высоком уровне изученности его
1 П о времени наступления
Западной Европы и С Ш А .
первой
менструации среди населения
ряд5'
фенотипа — анатомических, физиологических, биохимических, имк до­
логических, поведенческих и прочих признаков.
Для изучения-генетики человека применяют специальные методы.
В основуГг е й е а л о г и ч е с к о г о м е т о д а ) положено составление_д анализ родословныхгтю ставление р о д о с л о в н о й начинается с
п р о б а н д а (пропозита) — человека, попавшего в поле зрения
генетика или врача благодаря наличию признака, состояние которого у
родственников по отцовской и материнской линиям целесообразно
выяснить. В современном генеалогическом анализе широко применя­
ю тся статистические методы. Генеалогический анализ позволяет:
(^ установить наследственную природу признака: 2) определить-тип и
вариант наследования; 3) оценить пенетрантность аллеля; 4) изучать
сцепление генов и их локализацию в хромосомах. Он используется
также для оценки интенсивности м утационного процесса и изучения
взаимодействия генов у человека. Гёнеалогический~метод находит
широкоеТфименение в медицинской практике, для уточнения генетиче­
ской природы патологического состояния и при составлении прогнозов
здоровья потомков. Примеры родословных с разными типами и
вариантами наследования приведены в 5.2.8.
. Изучение генетических закономерностей на близнецах выполняется
(бГл~и~з н е ц о в ы м м е т о д о м т) Рождение двойней происходит
примерно в Г %
случаев от общего числа родов. М о н о зиготными
(однояйцевыми,
идентичными)
называю т
близнецов,
развившихся
из
одной
зиготы.
Онг
всегда
однополые
и
отличаю тся
большим
сходством
фи­
зических признаков (рис. 58). Важной особенностью таких детей
служит идентичность генотипов и, следовательно, той биологической
информации, на основе которой осущ ествляется их онтогенез.
Д изиготны ми (двуяйцевыми, неидентичными)
называю т близнецов, развившихся в одной беременности из различных
зигот. Они бы ваю т разнополыми, имеют в среднем 50% одинаковых
генов и похожи друг на друга не более братьев и сестер, рожденных в
разное время (рис. 59). О т последних их, однако, отличает большая
общность условий внутриутробного и постнатального развития, что
также учитывается в генетических исследованиях. О т общего количе
ства двоен дизиготные составляю т в среднем % — % . Соотношение
рождений моно- и дизиготных близнецов варьирует в различных
популяциях людей (табл. 14).
Т а б л и ц а 14. Коэффициенты м о н о - (т ) и дизиготности
частоту рождения одно- и двуяйцевых близнецов
т
По пуляц ия
<
1
По пуляц ия
(с1),
отражающие
ш
|
||
- -4
л
Русские
( X V I I I X IX
(X X век)
Финны ( X IX век)
(X X век)
Французы
Голландцы
Австрийцы
Индийцы
век)
6,6
3,6
3,1 ' 4,4
16,6
3,1
11,7
3,6
3,7
7,1
3,7
8,1
3,4
7,5
2,3
11,0
Африканские племена:
зулусы
свази
йоруба
ибо
Японцы
Китайцы
Малайцы
7,0
4,0
5,0
10,0
5,3
6,1
3,0
21,0
24,0
49,0
23,0
2,?
2,1
2,2
Рис. 58. Фенотипическое сходство
между однояйцевыми близнецами
Рис. 59. Фенотипические различия
между двуяйцевыми близнецами.
Разнополая двойня, в которой сест­
ра имеет нормальную пигментацию,
а брат — альбинос.
Диагностику монозиготности проводят путем сравнения детей по
совокупности внешних признаков (форма носа, губ, ушных раковин,
пигментация кожи, волос), моногенным иммунологическим маркерам
(эритроцитарные антигены групп крови АВО, резус), приживлению
кожного трансплантата. Близнецовый метод используется прежде всего
для оценки соотносительной роли наследственности и среды_в развитии
120
признака. _ При этом сопоставляется состояние признака в парах
монозиготных близнецов, дизиготных близнецов, в парах близнецов и
остальной популяции. Таким образом вы ясняю тся закономерности
развития, соответственно, на основе одинаковых генотипов при
ыезначихельных_разлиниях Внешней среды,'разных генотипов в близких
__ условиях окружающей среды, одинаковых или разных генотипов в
сходных или различающихся условиях. Иногда монозиготных близне­
ц о в разлучаю т вскоре после рождения, и на таких парах изучают
закономерности развития на основе одинакового генотипа в су­
щественно различающихся условиях экологической и социальной
среды.
Для суждения о роли генотипа или условий развития сравнивают
к а н к о р д а-д-д^л о с т и ( и л и д и с к о р д а н т н о с т и ) парт­
неров по качественным признакам или величины дисперсии количе­
ственных признаков. Пара близнецов является конкордантной, если
исследуемый признак обнаруживается у обоих партнеров, и дискордантной, если признак присутствует лишь у одного из близнецов. Чем выше
Т а б л и ц а 15. Конкордантность моно- и дизиготных близнецов по некоторым
нормальным признакам и заболеваниям
К п н ко р д ан тн о е ть
П ризнак
(%)
и.ш з а б с ы и а н и е
монозиготные
Цвет глаз
волос
кожи
Форма иоса
„
ушей
Папиллярные линии
Н ачало менструаций (различие
в месяцах)
Косолапость
З а яч ь я губа
Врожденный вывих бедра
Туберкулез
Бронхиальная астма
Сахарный диабет
Корь
Коклюш
д и эиготны е
9 9 ,5
2 8 ,0
9 7 ,0
2 3 ,0
1 0 0 ,0
4 5 ,0
1 0 0 ,0
3 0 ,0
9 8 ,0
2 0 ,0
9 2 ,0
4 0 ,0
3
3 5 ,0
13
3 2 ,0
3 ,0
3 3 ,0
5 ,0
4 1 ,0
3 ,0
3 7 ,0
1 5 ,0
4 7 ,0
2 4 ,0
6 5 ,0
1 8 ,0
9 8 ,0
9 4 ,0
9 7 ,0
9 3 ,0
конкордантность, тем больш ую роль в развитии признака играет
наследственность (табл. 15). Совпадение значений конкордантности в
парах моно- и дизиготных близнецов свидетельствует о преобладаю­
щем значении в развитии признака факторов среды. Такая ситуация
кладывается в отношении некоторых инфекционных болезней,
заражение которыми зависит от контакта с возбудителем (см. корь и
коклюш в табл. 16).
Значения конкордантности в парах моно- и дизиготных близнецов
используют в расчетах показателя влияния наследственности на
121
фенотип. Он служит более строгим критерием, чем непосредственно
конкордантность При выраженной изменчивости качественного приз­
нака, например из-за меняющейся пенетрантности гена, показатели
конкордантности искажаю тся. Состояние количественного признака в
парах близнецов оценивается величиной дисперсии, которая служит
статистическим показателем пределов вариабельности этого признака в
разных группах людей. Меньшая вариабельность указывает на большую
генетическую обусловленность, Используя данные близнецового мето­
да в оценке соотносительной роли наследственности и среды в форми­
ровании признаков, необходимо у ч и т ыват наложный характер процесса _
развития фенотипа. Так, известны монозиготные близнецы-сестры с
нормальным кариотипом. Одна из девочек была здоровой носительни­
цей аллеля гемофилии, а вторая болела гемофилией. Дискордантность
по рассматриваемому признаку возникла у сестер, видимо, потому,
что у больной генетическая инактивация затронула преимущественно
отцовские Х-хромосомы.________ .— ------ ^
О Т и т1 Т Г?тгТ Т ггГч е с к и й м е т о
заклю чается в изучении
хромосом лри помощи микроскопа. Чащ е объектом микроекоинрования служат митотические (метафазные), реже мейотические (профазные
и мегафазные) хромосомы. В интерфазных ядрах соматических клеток
женщин легко определяется генетически инактивированная Х-хромосома (половой хроматин или тельце Бара). В последнее время с помошью
люминесцентной микроскопии идентифицируется У-хромосома клеток
м урчит.
Применяемые гистологические методы дают сплошную или диффе­
ренциальную окраску вещества хромосом, что позволяет провести их
групповую или индивидуальную идентификацию. С помощью цитоге­
нетического метода осущ ествляю т: 1) изучение кариотипов организма;
2) уточнение числа хромосомных наборов, количества и морфологии
отдельных хромосом для диагностики хромосомных болезней; 3) сос­
тавление генетических карт хромосом; 4) изучение геномных и хромо­
сомных мутаций. Технически наиболее просто диагностировать измене­
ние числа X хромосом. Для этого подсчитывают количество телец
полового хроматина в ядрах лейкоцитов периферической крови
чти эпителиальных клеток слизистой оболочки ротовой полости
(рис. 60)
Методы
генетики
соматических
клеток
образуют отдельную группу. Их объединяет сходство объекта
исследования, каковым служат клетки органов и тканей, выращиваемые
на питательной среде вне организма. В процессе культивирования
проводят клонирование, т. е. размножение потомков единственной
клетки, выделенной из общей клеточной массы. Такие потомки имеют
идентичные генотипы. Подбирая питательные среды, проводят се­
лекцию (отбор) клеток, отличающихся определенными признаками.
В условиях культуры удается получить клетки-гибриды, в которых
совмещены геномы разных особей одного вида или организмов разных
пцппч
Сущ ествую т гибриды клеток человека и мыш и, крысы, морской
свинки, обезьяны. В генетических исследованиях человека эта группа
122
\\
Н е 1 поло!
го хром ат
О д н о тел
полового
хроматинг
(в)
\\\\
М аленько
тельце пс
левого хр
матина
\\\\ >
тельца п о ­
лового
хр ом ати н а
Рис. 60. Соответствие количества телец «полового хроматина» в соматических клст
ках генетической конституции организма по Х-хромосоме.
<1
ж зависимость количества телец от числа X-хромосом
I
методов позволяет осущ ествлять: 1) анализ сцепления и локализации
генов; 2) изучение первичных продуктов активности отдельных генов;
3) вскрытие механизмов взаимодействия генов и путей регуляции
генной активности; 4) изучение генных мутаций. Э ти методы нашли
применение в медицинской практике. Путем исследования культур
клеток околоплодной жидкости еще до рождения ребенка диагностиру­
ю т более шестидесяти наследственных заболеваний.
Особенности распределения наследственных признаков и контроли­
рующих их генов в разных группах людей изучаю тся при помощи
п о п у л я ц и о н н о - с т а т и с т и ч е с к о г о м е т о д а . Указан­
ный метод позволяет: I) оценить вероятность рождения лиц с опреде­
ленным фенотипом в данной группе населения или в близкородственных
браках; 2) рассчитать частоту носительства в гетерозиготном состоя­
нии рецессивных аллелей. В основе метола лежит закон Харди —
Вейнберга, который будет рассмотрен ниже.
В последние годы расширяется круг методов, в основном
б и о х и м и ч е с к и х , которые позволяю т диагностировать гетеро­
зиготное носительство нежелательного аллеля. Э то возможно в тех
случаях, когда интересующий врача-генетика аллель проявляет свойс­
тво кодоминантности или же имеет место лозовый эффект и выра­
женность признака доминантной гомозиготы и гетерозиготы раз­
лична. Так, при электрофоретическом анализе гемоглобина гетеро­
зигот Н ЬА /Н Ь5 обнаруживается как нормальный, так и аномальный
белок (рис. 61).
Гетерозиготные носители рецессивного аллеля фенилкетонурии реагируют на введение фенилаланина более сильным повышением
концентрации аминокислоты в плазме крови, чем нормальные
гомозиготы. Э ти методы широко используются в медико-генетическом
консультировании для определения вероятности рождения ребенка с
наследственным заболеванием.
В разделе экспериментальной патологии, изучающем наслед123
Рис. 61. Обра ю нш жс пор
мильного и аномального гг
М О | лобина I стсро 1ИГОТНМИ по
локусу ссрпонидноклсточно
сти эритроцитов.
ственные болезни, применяется метод моделирования на животных,
наследственно обусловленных патологических признаков человека.
Теоретическую основу его составляет закон гомологичных рядов
наследственной изменчивости Н . И . Вавилова. У животных обнаруже­
ны мутации, приводящие к развитию гемофилии, сахарного диабета. Н а
рис. 62 показаны организмы разных классов и типов — рецессивные
гомозиготы по аллелю альбинизма.
124
Рис. 62. Продолжение.
6
рыбы, в
5.3.
н асеком ы е
и зм е н чи в о ст ь
Благодаря наследственности достигается единообразие плана
строения, механизмов развития и жизнеобеспечения организмов одного
вида. Разнообразие деталей строения и физиологических отправлений
особей, наблюдаемое на фоне указанного единообразия, зависит от
изменчивости.
125
Изменчивость
-ц
Генотипическая
Фенотипическая
Модификационная
1
I--
Соматическая
Г енеративная
Случайная
Мутационная
Комбинативная
Геномные мутации
Изменение числа
геномов
Изменение числа
отдельных хромосом
Комбинативная
Хромосомные мутации
I------
Межхромосомные
— ^“ 1
Мутационная
Генные мутации
1
Внутрихромосомные
1
Полиплоидия
Моносомии
Три- и полисомии
Транслокации (с из­
менением групп
сцепления)
Инверсии (без изме­
нения групп сцепле­
ния)
Дупликации, делеции, дефишенси (с из­
менением групп
сцепления)
Рис. 63. Кролики горностаевой породы.
а — фенотип о б ы чн ы й , б
при пониж енной тем пер атур е на спине и кр естц е вы р осл а те м н а я ш ер сть
Изменчивость является результатом различных процессов. Некото­
рые из них происходят в наследственном материале (генотипе). Другие
ограничиваются фенотипом. В соответствии с этим выделяю т изменчи­
вость г е н о т и п и ч е с к у ю и ф е н о т и п и ч е с к у ю . Феноти­
пическая изменчивость подразделяется н а м о д и ф и к а ц и о н н у ю
и с л у ч а й н у ю ; а генотипическая, в зависимости от природы
изменяющихся клеток, — на
генеративную
и
сома­
тическую
с выделением в них
мутационной
и
комбинативной
изменчивости.
В классической
генетике
М у Т а Т Д Г П называется скачкообразное изменение наслепственно
"обусловленного признака. М утации возникают внезапно, без пере­
ходных состояний по сравнению с исходной формой признака. Они
устойчивы во времени и происходят применительно к одному признаку
в различных направлениях. Одна и та же мутация может возникать пов­
торно. М утации классифицируют в зависимости о т уровня организации
наследственного материала, на котором происходит Изменение,
проявляющееся затем на уровне признака. Так, выделяю т генные,
хромосомные и геномные мутации. Отдельные виды геномных и
хромосомных мутаций разграничивают, исходя из характера измене­
ния — отклонения, в числе геномов, количестве или морфологии
хромосом. Основные типы и формы изменчивости представлены на
схеме 4.
5.3.1. Фенотипическая изменчивость
Ф е н о т и п и ч е с к а я и з м е н ч и в о с т ь охватывает изме­
нения состояния непосредственно признаков, которые происходят под
влиянием условий развития или факторов внешней среды. Наслед­
ственный материал при этом в изменения не вовлекается.
Если фенотипические изменения не выходят за пределы «нормы
реакции» и вызываю тся известным фактором внешней среды, их
называю т м о д и ф и к а ц и я м и . Так, сущ ествует порода кроликов
русских горностаевых, которые имеют белый мех с черными пятнами
на кончике морды, ушах, лапах, хвосте. Если на спине такого кролика
выбрить участок и поместить животное на некоторое время на холод,
то оголенное место зарастает темной шерстью (рис. 63). Н а участках,
127
занятых обычно темной ш ерстью, при повышении температуры сверх
определенного предела вырастает белая ш ерсть. При выращивании в
обычном температурном режиме кроликов, рожденных о т родителей с
измененным описанным способом распределением темной шерсти,
воспроизводится исходный горностаевый тип окраски. Таким образом,
регистрируемая изменчивость затрагивает лиш ь фенотип особи и обус­
ловливается воздействием конкретного фактора среды — темпера­
турного. Э т о т фактор изменяет функцию гена, контролирующего
синтез пигмента. Последнее подтверждается тем , что кролики-альбино­
сы н^ рёаггфуют на изменение температуры появлением участков
темной шерсти. Модификационная изменчивость в естественных усло­
виях обитания имеет приспособительное значение. Так, у обычного для
прибрежной зоны рек и озер растения стрелолиста строение листа
соответствует месту произрастания — на суше, с частичным или
полным погружением в воду.
Фенотипические изменения называю т с л у ч а й н ы м и , если они
появляю тся в результате совместного действия на организм многих
факторов внешней среды, причем интенсивность действия каждого из
них сопоставима и незначительна (Б . Л . Астауров). Закономерности
фенотипической изменчивости важны для медицинской практики, так
как знание их помогает оценить значение отдельных факторов среды
или всей совокупности условий, в которых протекает онтогенез, для
развития фенотипа. Модификационной изменчивостью объясняется
феномен фенокопирования. В качестве примера рассмотрим гипоспадию, — порок развития мужской половой системы. Он проявляется в
смещении наружного отверстия мочеиспускательного канала с головки
полового члена к его основанию. Причиной гипоспадии могут бы ть
изменения в генетическом материале или отклонения в ходе эмбриогене­
за. В последнем случае изменение представляет собой фенокопию.
Тактика врача в отношении больных гипоспадией в обоих случаях
одинакова — производится хирургическое исправление дефекта. Н о для
решения вопроса о возможности рождения в семье страдавшего
гипоспадией больного ребенка необходимо разграничение случаев
заболевания генотипической и фенотипической природы.
5.3.2. Генные мутации
В классической генетике г е н н о й
(точковой,
истин­
ной) м у т а ц и е й
называется неразличимое с помощью мик­
роскопа изменение внутренней организации хромосомы на уровне
отдельных генов, которое обнаруживается по появлению в потомстве
измененного признака. Современная генетика подчеркивает, что генные
мутации заклю чаю тся в изменении химической структуры генов.
Происходит нарушение последовательности нуклеотидов в Д Н К , что
ведет к искажению наследственной информации в соответствующих
фрагментах молекул нуклеиновой кислоты. Конкретно генные мутации
являю тся заменами, вставками, выпадениями и потерями пар нуклеоти­
дов (рис. 64). Наименьший участок молекулы Д Н К , изменение которого
приводит к мутации, называется м у т о н о м . Он равен одной паре
нуклеотидов.
128
Существует несколько класси­
' ' I I I ' ! I I [ | 'Г'''т 1’
Л
7 I \ 11 Л Г Ц 1
Л !Д; I I А !
фикаций генных мутаций. С п о н ­
Т Л Л Ц Г 1 АГ ЦТ Г I Л т ц Г Л
т а н н о й ( с а м о п р о и з в о л ьI I ! ! 1 I I 1 1 I1 1 =I 1 .
н о й) называю т мутацию, кото­
рая происходит вне прямой связи
1
1II ; I .
I I I I ■' I I I 1 I !
с каким-либо физическим или хими­
А Т Т Г;Т ,Л Т Ц Г А Ц Ц Т А Г ЦТ
2 и
к н 'П ■ш! и и ш ш 1 ч1 и1 н >
•Н1 ш и
ческим фактором внешней среды.
Т
А А Ц1А-Т А Г Ц Г Г Г Л Т Ц I А
I
1
I
1:
1
11
I
I
|
I
I
[| I I
Если мутации вызываю тся наме­
ренно, воздействием на организм
— I I |I I г I I
I ! I : I I 1 г 1"т г г факторами известной природы, они
А т т Г и Л Т и
Г;т:л Ц Ц
Г А Г
Ч
II •
II I " III III III III
III III 1111 'II III III 1*1 III III " I
называю тся
индуцирован­
г л
ц I
: . I I
ными.
Агент, индуцируюший
Щмутации, называю т м у т а г е ­
*
ном.
— Г I П 'Т Т
А Т Т Г Ц А Т А Г Ц Т А Ц
Природа мутагенов разнообраз­
А II II II ш |н п и и |н >11 II н ш
А Т Ц Г А Т Г
Т Л А Ц Г Т
на — это
физические
факторы,
11111
химические соединения, в том числе
*
вещества, используемые в сельском
хозяйстве в качестве гербицидов Р ис. 64. Главные классы генных мута­
(гидразид малеиновой кислоты) и ций.
ор м ал ьн ая п осл е д овате л ьн о сть н ук л ео ти ­
пестицидов (Д Д Т ), в медицине в дI ов, 2н—
зам е н а п ар ы « Ц - Г » на пару « А — Т » ,
качестве лекарственных препаратов 3 - в с т а в к а пары « Т - А »; 4 - потеря блока
из ш ести пар оснований.
(производные тиазинового ряда,
нитрофураны), в различных произ­
водствах (афоксид— импрегатор текстильных тканей, формальдегид —
производство искусственных смол, гидроксиламин — производство
бумаги), в пищевой промышленности (натрий бисульфит — консервант
вин). Установлено мутагенное действие некоторых биологических
объектов — вирусов, простейших, гельминтов при проникновении их в
организм человека. Соответственно различают физические, химические,
фармакологические, промышленные, биологические мутагены. Среди
физических мутагенов сильное мутагенное действие оказываю т ионизи­
рующее ( р а д и а ц и о н н ы й м у т а г е н е з ) и ультрафиолетовое
излучение. Отдельные виды мутагенов вы зы ваю т специфические
повреждения Д Н К . Ультрафиолетовое облучение, активируя пирими­
диновые азотистые основания, способствует образованию ти.миновых
димеров « —Т — Т — ». Эффект азотистой кислоты объясняется, повидимому, дезаминированием (отщеплением аминогруппы) аденина,
гуанина и цитозина. Химические аналоги азотистых оснований,
например бромурацил, заменяю т в процессе синтеза нормальные
основания и приводят к ошибкам редупликации.
В результате доминантных и рецессивных мутаций в фенотипе
появляю тся доминантные и рецессивные измененные признаки (см.
табл. 9). Д о м и н а н т н ы е м у т а ц и и проявляются в фенотипе
уже в 1-м поколении. Р е ц е с с и в н ы е м у т а ц и и укры ты в
гетерозиготах от действия естественного отбора, поэтому они
накапливаются в генофондах видов в больш ем количестве.
В силу системного принципа организации генотипа лишь в редких
случаях мутации, возникаюшие заново, являю тся по своему физиологи­
а
5 — 56
т
а
г
ц
а
т
а
г ц
г
а
129
ц
т
ческому действию б л а г о п р и я т н ы м и
или н е й т р а л ь ­
н ы м и , повышающими или хотя бы не снижающими жизне­
способности их носителей — м у т а н т о в . Чащ е наблюдаются
неблагоприятные
мутации, среди которых выделяю т
л е т а л ь н ы е и п о л у л е т а л ь н ы е 1. Первые приводят орга­
низм к гибели на ранних этапах индивидуального развития, например на
стадии зиготы. В случае полулегальной мутации эмбриогенез может
бы ть завершен, однако у такого организма жизнеспособность настоль­
ко снижена, что он погибает до достижения половой зрелости и не
участвует в репродуктивном процессе, а следовательно, не передает
своих генов потомкам. Даже выраженный неблагоприятный эффект
рецессивных летальных и полулегальных мутаций не препятствует их
сохранению в генофондах видов в скрытом гетерозиготном состоянии.
В зависимости от функционально-генетической характеристики мути­
ровавших генов мутантные аллели оказываю т разное плейотропное
действие.
Показателем интенсивности мутационного процесса служит частота
мутирования, которую рассчитываю т в среднем на геном или отдель­
но для конкретных локусов. С р е д н . я я
частота
му­
т и р о в а н и я сопоставима у широкого круга живых существ (о т
бактерий до человека) и не зависит о т уровня и типа морфофизиологиче­
ской организации. Она равна К Г 4 — 10“® мутации на 1 локус за
поколение. Продолжительность жизни одного поколения людей
составляет 25— 30 лет. Принимая у человека среднюю частоту
мутирования равной 1 х 10-6 , а количество локусов 105— 10е , можно
видеть, что на каждый гаплоидный набор генов за поколение возникает
от одной до десяти новых мутаций. Ч а с т о т а м у т и р о в а н и я
отдельных
генов
варьирует более широко. О т частоты
мутаций в конкретном локусе зависит относительное количество гамет,
несущих соответствующ ий мутантный аллель (табл. 16).
Таблица
16.
Частота мутаций у человека
З а б о л е ва н и е , в ы зы в ае м о е м утац ией
Эпилойя
Недоразвитие радужной оболочки
(аниридия)
Талассемия
Серповидноклеточная анемия
Альбинизм
Ихтиоз
О бш ая цветовая слепота
Гемофилия (данные по Англии)
(данные по Дании)
Ч а с т о т а м утац ии
Ч и с л о м утантн ы х
половых клет«Ук (н а
1 млн. га м е т)
4,8 X I О-6
4— 5
1,2x10 5
4,0 X 10_ 4
1,0 X I О "2
2 .8Х10-5
1,1 X 10-*
2,8 X 10-5
2,0 X Ю ' 5
3 .2x10 5
12
400
10 000
28
11
28
20
32
1 Д анная классификация, как и деление на спонтанные и индуцированные мута­
ции, распространяется на хромосомные и геномные мутации.
130
Рис. 65. Генная мутация по типу замены,
индуцируемая мутагеном 5-бромурацилом.
Н а интенсивность мутирования влияет генетическая конституция
особи в целом. К примеру, частота мутации Я г — г ' у кукурузы
колумбийской и корнельской линий различается в 3 раза и составля­
ет 6,2 и 18,2 на 104 гамет. Описаны а л л е л и - м у т а т о р ы ,
присутствие которых в геноме изменяет, иногда на несколько порядков,
частоту мутирования в других локусах. Частота мутирования некото­
рых генов изменяется в зависимости от нахождения их в яйцеклетке или
сперматозоиде. Так, сопоставление числа рождающихся женщин и
мужчин, гетерозиготных и гемизиготных по аллелю гемофилии,
позволяет заклю чить, что названная мутация случается в 10 раз чаще в
мужских гаметах. П о локусу мышечной дистрофии Дюшена, который
также локализуется в Х-хромосоме, частота мутирования в гаметах
обоих типов одинакова. Н а интенсивность мутирования влияет
состояние организма. Вероятность присутствия в сперматозоиде аллеля
синдрома Марфана, например, растет с возрастом мужчины. В опытах
на животных показан рост интенсивности мутационного процесса с
повышением температуры. Э то т вывод не подтверждается для
человека. Так, не установлено различий в среднем уровне мутирования
генов сперматозоидов и яйцеклеток, хотя последние благодаря
положению яичников в глубине тканей развиваю тся при более высокой
температуре.
Молекулярные механизмы генных мутаций окончательно не
выяснены. Предположительно изменения Д Н К служ ат следствием
ошибок в ходе таких внутриклеточных процессов, как редупликация,
рекомбинация и репарация (восстановление после повреждений)
наследственного материала.
Роль о ш и б о к р е д у п л и к а ц и и в возникновении генных
мутаций иллюстрирует следующий пример. Допустим, что в процессе
синтеза в молекулу Д Н К вместо обычного азотистого основания
тимина вклю чается его химический аналог бромурацил (Б У ), что
приводит к образованию пары « А — Б У » . В отличие от тимина
бромурацил имеет большее сродство не к аденину, а к гуанину, что в
следующей редупликации Д Н К клетки даст пару « Б У —Г ». Благодаря
этому в очередном редупликационном синтезе появляется пара « Г — Ц »
(рис. 65). Таким образом вместо исходной пары «А — Т » в определенном
участке молекулы Д Н К образуется пара « Г — Ц », что соответствует
мутации по типу замены.
О
появлении
генных
мутаций
вследствие
ошибок
рекомбинации
наследственного материала свидетельствует
меньшее в 10—20 раз количество мутаций в клетках дрожжей при
бесполом размножении, чем при половом, когда происходит мейоз.
5*
131
Репарация
наследственного
материала
заключается в ферментативном разрушении измененного участка
молекулы Д Н К с восстановлением на этом отрезке последовательности
нуклеотидов, комплементарной фрагменту неповрежденной молекулы
Д Н К . В некотором проценте случаев фермент ошибочно разрушает
фрагмент нормальной молекулы Д Н К , комплементарный измененно­
му. В результате последующего синтеза образуется м утантная двойная
спираль.
Так как молекула Д Н К представлена двойной спиралью,
образование генной мутации проходит в два этапа. Н а первом этапе
изменение захватывает лишь одну молекулу биспирали. Э то состояние
называется м о л е к у л я р н о й г е т е р о з и г о т н о с т ь ю или
п о т е н ц и а л ь н о й м у т а ц и е й . Переход в и с т и н н у ю
мутацию
происходит путем распространения изменения на
гомологичный локус комплементарной молекулы. В результате дости­
гается состояние м о л е к у л я р н о й
гомозиготности.
Распространение изменения на обе молекулы двойной спирали приводит
к том у, что мутация наследуется всеми потомками мутировавшей
клетки. Согласно распространенной гипотезе переход в состояние
молекулярной гомозиготности
является результатом
ошибки
р е п а р а ц и и . Альтернативное объяснение заклю чается в том , что в
ближайшем цикле редупликации образую тся полностью м утантная и
нормальная биспирали Д Н К , которые при делении расходятся в разные
дочерние клетки. В последующих циклах редупликации и деления одна
клеточная линия погибает вследствие различной жизнеспособности
мутантных и нормальных клеток.
5.3.3. Биологические антимутационные механизмы
В результате толковых мутаций изменяется смысл биологической
информации, записанной в генах. Последствия этого м огут бы ть
двоякого рода. В мало меняющихся условиях обитания новая информа­
ция обычно снижает выживаемость вида. При резкой смене условий
обитания, освоении видом новой экологической ниши наличие
разнообразной информации в генофонде полезно. Характерен пример с
бабочками
березовыми
пяденицами
(ВЫ оп
ЪеШ1апа). Исходная популяция бабочек на Британских островах сос­
тояла из светлых особей, темные особи — м е л а н и с т ы — встре­
чались в единичных экземплярах. К началу текущего столетия в про­
мышленных районах Англии меланисты составляли уже более 99%
популяции, что было связано с загрязнением лесов копотью . Н а фоне
закопченных стволов деревьев темная окраека стала защитной.
В незагрязненных областях светлые формы сохраняются в большин­
стве. Темная окраска бабочек развивается в присутствии доминантного
аллеля С (сагЬопапа), а светлая — рецессивного с. Наличие двух аллелей
по локусу синтеза меланина, таким образом, расширяет зону расселения
березовых пядениц.
Интенсивность мутационного процесса поддерживается на уровне,
при котором возникновение и сохранение новой биологической
132
информации не вызывает катастрофического снижения жизнеспособно­
сти вида. Важная роль в ограничении неблагоприятных последствий
генных мутаций принадлежит е с т е с т в е н н ы м а н т и м у т а ционным
барьерам,
различающимся по своей при­
роде. Так, парность хромосом в диплоидных кариотипах эукарио­
тических организмов, благодаря феномену гетерозиготности, препя­
тствует непременному проявлению в фенотипе рецессивных м ута­
ций. Формой защ иты против точковых мутаций служат повторы
(экстракопии) некоторых генов, например кодирующих р Р Н К , т Р Н К ,
гистоновые белки хроматина. Интенсивность мутационного процесса
снижается благодаря таким свойствам биологического кода, как
триплетность и вырожденность. Протяженность в три азотистых
основания минимизирует число перестановок внутри кодона. При
вырожденности кода мутация в виде замены основания может дать
кодон-синоним, так что нормальная последовательность аминокислот в
соответствующ ем полипептиде не изменяется. Так, Р-полипептид
гемоглобина человека кодируется фрагментом Д Н К из 438 нуклеоти­
дов. Около 25% замен одного из них не вносит искажений в аминокис­
лотную последовательность, а 2— 3 % приводят к образованию
укороченного полипептида в связи с преждевременной терминацией
синтеза и РН К . Оставшиеся 73% приводят к замене одного
аминокислотного остатка. Изменения разных нуклеотидов триплета
неравнозначны. Наименее эффективны замены третьего нуклеотида,
64% которых происходит без искажения смысла. Наиболее эффективны
замены второго нуклеотида, которые вы зы ваю т мутации в 100%.
Ограничение неблагоприятных последствий точковых мутаций достига­
ется также благодаря неравнозначному функциональному действию
различных замен аминокислот. Если новая и заменяемая аминокислоты
сходны по основным свойствам, изменения в третичной структуре и
биологических свойствах белка незначительны. Так, мутантные
гемоглобины 5 и С человека отличаю тся от нормального замещением
глутаминовой кислоты в 6-ом положении на валин и лизин. Первая
замена резко изменяет свойства гемоглобина и приводит к развитию
тяжелой серповидноклеточной анемии. При второй замене развивается
малокровие легкой степени. Одна из причин этого заключается в том,
что глутаминовая кислота и лизин гидрофильные аминокислоты, а
валин гидрофобная. Рассмотренные антимутационные барьеры ограни­
чиваю т проявления мутации в фенотипе особей.
Репарация
или
коррекция
молекулярных
н а р у ш е н и й с т р у к т у р ы Д Н К приводит к устранению из
наследственного материала клетки измененного участка. Молекулярные
нарушения Д Н К , вызываемые различными физическими и химическими
факторами, устраняю тся разными путями. Более универсальной
является система « р е п а р а ц и и п у т е м в ы р е з а н и я», кото­
рая состоит из следующих этапов (рис. 66): а) разрыв молекулы Д Н К в
месте повреждения с сохранением целостности комплементарной
молекулы на этом участке; б) удаление (вырезание) зоны повреждения с
расширением дефекта в обе стороны; в) синтез фрагмента Д Н К
(«заплаты ») на месте разрушенного; г) восстановление непрерывности
133
Зона повреждения
Насечна эндонуклеазой
репарируемой молекулы
г
ТI" I 1"ТI I I I I I I I I п I I I ГI I I I днк.
Описанным
спосо­
бом из молекулы Д Н К
устраняю тся, например,
Т А Г Т Ц Ц Д Г а а т г ц а т ц ц ц т т г а г г
пиримидиновые
димеры
Л1
(Т
Т
,
Т
Ц
,
Ц
Ц
),
которые
„Состригание“
I
энд онуклеазам и
/ V
I \ ч
образуются под действи­
чччч У а л ч\ ч \\ уЧ\\
ем УФ-облучения. Э та
I I И
Г
\>
I I I I I I I I I I1 1 1 1 1
система репарации функ­
А Т Ц А Г Г Т
А Ц Г Т А Г Г Г
ААЦТЦЦ
ционирует в клетках че­
' II II III II III III II
II III III II II III III III II II
ловека, а ее генетический
Т А Г Т Ц Ц А Г А А Т
Г Ц А Т Ц Ц Ц Т Т
ГАТТ
дефект, заключающийся
в нарушении первого,
ДНК- п о л и м е р а з а
этапа, приводит к забо­
- с и н т е з ------леванию
пигментной
ксеродермой.
У таких
А Т Ц
А Ц Г Т А Г Г
Г А А Ц Т Ц Ц
пациентов часто наблю­
даются изменения Д Н К
Т А
А А Т Г Ц А Т Ц Ц Ц Т Т Г А Г Г
и хромосом при дейст­
вии солнечных лучей.
В клетках эукариот об­
Сшивание лигазой
наружены две разные
системы «репарации пу­
А Т Ц А Г Г Т Ц Т
Т А Ц Г Т А Г Г Г А А Ц Т Ц Ц
тем вырезания», кото­
рые устраняю т дефекты
Т А Г Т Ц Ц А Г А А Т Г Ц А Т Ц Ц Ц Т Т
Г А Г Г
различной природы, от­
I I I I I I I 1 I I 1 I I I I I II I I I I I I
личаю тся протяженно­
стью
разрушаемого
Рис. 66. Этапы репарации Д Н К путем вырезания.
фрагмента Д Н К и дли­
Объяснение в тексте.
тельностью процесса ре­
параций — 1—24 ч и
100 нуклеотидов, 5 мин — 2 ч и 3— 4 нуклеотида соответственно. У эу­
кариот найдена такж е система « п о с т р е п л и к а т и в н о й р е п а ­
р а ц и и», которая функционирует в синтетическом периоде митоти­
ческого цикла. В процессе предмитотического синтеза участки моле­
кулы Д Н К имеющие, например, тимидиновые димеры — Т — Т — ,
не редуплицируются и на их месте образуются бреши. Недостающие
фрагменты достраиваются с использованием в качестве матрицы нор­
мальной молекулы Д Н К . Э то позволяет «обойти» поврежденный уча­
сток и избежать наследования первичного мутационного изменения
дочерними клетками.
Эффективность репарации и других антимутационных барьеров с
достаточной точностью не определена. Она зависит о т природы
повреждений Д Н К и состояния клеток. Так, в клетках почек сирийского
хомячка удаляется до 70% измененных нуклеотидов. Длительное
культивирование этих клеток вне организма подавляет репаративные
процессы.
а
т
ц
а
г
г
т
ц
т
т
а
ц
г
т
а
г
г
г
а
а
ц
т
ц
ц
I I 1 I I I I I I I I II I I I I I I I
I I II I I I I I I I I I I
I I I I I I II I II I I I I
134
5.3.4. Хромосомные мутации
Хромосомные
мутации,
хромосомные
пе­
р е с т р о й к и или а б е р р а ц и и заклю чаю тся в изменении ко­
личества или перераспределении наследственного материала хромо­
сом. Их рассматриваю т как рекомбинации генов, имеющие в отличие
от мейотической рекомбинации нерегулярный характер. Некоторые
виды таких мутаций диагностируются по изменению морфологии
метафазных хромосом с помощью микроскопа. Перестройки подраз­
деляют
на
внутрихромосомные
и
меж хромосомные.
В н у т р и х р о м о с о м н ы е п е р е с т р о й к и (рис. 67) заклю­
чаю тся в утрате части хромосомы
(д е л е ц и я),
удвоении
или умножении некоторых ее участков ( д у п л и к а ц и я ) ,
пово­
роте фрагмента хромосомы на 180° с изменением последователь­
ности расположения генов ( и н в е р с и я ) . Дупликации и делеции при­
водят к изменению дозы генов, поэтому их рассматривают также как
частичные три- или моносомии по соответствую щ им хромосомам.
При м е ж х р о м о с о м н ы х п е р е с т р о й к а х негомологич­
ные хромосомы обмениваются участками, т. е. происходит
т р а н с л о к а ц и я наследственного материала. Часты комбиниро­
ванные нарушения морфологии хромосом, например сочетание делеции
и транслокации. Некоторые ввды хромосомных аберраций приводят к
изменению генного состава групп сцепления (делеции, дупликации,
транслокации), другие (инверсии) — нет. Во втором случае отклонения
в фенотипе м утанта обусловливаются эффектом положения, а частота
хромосомных перестроек зависит от влияния внешних факторов и
состояния организма. Механизмы возникновения разных видов мутаций
различны, однако во всех случаях имеет место разрыв хромосомы с
возможным соединением фрагментов в последующем. К хромосомным
мутациям приводит первичное изменение физико-химического состоя­
ния вещества хромосомы.
При индукции хромосомных аберраций ионизирующим излучением
• повреждения хромосом, вопреки теоретическим ожиданиям, распреде­
ляю тся неслучайным образом. Наблюдается предпочтительная локали­
зация разрывов в гетерохроматиновых участках. Избирательное
распределение мутаций в еще большей степени наблюдается при
химическом мутагенезе.
Хромосомные мутации изменяют дозу некоторых генов, вы зы ваю т
135
перераспределение генов между группами сцепления, меняю т локализа­
цию их в группе сцепления. Эти м они наруш ают генный баланс клеток
организма, в результате чего происходят отклонения в соматическом
(физическом) и психическом развитии особи. Как правило, изменения
распространяются на несколько систем органов. У человека наиболее
часто отмечаю тся пороки развития головного мозга, опорно-двига­
тельной, сердечно-сосудистой, мочеполовой систем. Наблюдается
умственная отсталость разной степени выраженности в сочетании с
физическим недоразвитием. Отклонения со стороны половой системы
функционально проявляются в первичном бесплодии мужчин и женшин,
привычном невынашивании беременности. При доношении беременноТаблица
хромосомы 7
17.
Признаки
частичных
трисомий
по
разным
участкам
У ч а с т о к хромосомы 7
П р и зн аки
7
Низкая масса тела при рождении
Задержка роста
Выступающий затылок или лоб
Микроцефалия
Глазная щель
+
_
+
—
31
4+
Узкая
+
Плоская
+
—
+
—
—
+
Низко
Таблица
мосомам
посажены
Большие
+ нал и чи е п р и зн ака, — его о тсу тст ви е
18. Обнаружение формы анеуплодий у человека по разным хро­
П олисом ии
0
0
0
0
0
0
0
0
0
0
0
0
Пр им ечание:
136
21 (и л и 22) — 7
+
Прим ечание:
1
2
3
4
5
6
7
8
9
10
11
12
7
+
—
горизонтальная,
узкая
—
Нормальная
Эпикант
Переносица
Ретрогнатизм
Микрогнатизм
Волчья пасть
Гипотония
Уши
•V* хромо­
сомы
31,7
Трисом ии
0
п
п
0
0
0
п
11
п
п
0
0
0
М оносом ии
М
п
0
п
0
0
0
0
0
0
0
0
0
не о б н ар уж е н о ан ом ал и й; 11
хромо­
со м ы
13
14
15
16
17
18
19
20
21
22
X
V
11(1.1ИСОМ ИИ
Трисомии
М оносом ии
0
0
0
0
0
0
0
0
0
0
п .м
п
п ,м
п
п
п
0
Г1,М
0
п
п .м
п
п .м
п .м
0
0
0
0
0
0
0
0
п
п
Г1.М
п .м
п о л н а я ф орм а; М - м о заи чн ая форма.
II
III
IV
Рис. 68. Схема образования гамет с несбалансированным числом хромосом.
I
- исхо дная к л е т ка
с л и ш н е й хромосомой;
6 — тр и со м н ая.
в процессе гам е то ге н еза ; I I га м е ты : а — с н ех ваткой хромосомы; 6
III
нор м альн ы е гам еты второго р од ителя: I V — зи го ты : а — моносомнаи,
сти у таких женщин рождаются дети с небольшой массой тела. Тяжесть
фенотипических нарушений коррелирует с количеством утраченного или
дополнительного наследственного материала. Конкретное выражение
изменений фенотипа зависит от локализации мутации в той или иной
хромосоме или ее участке (табл. 17). М утации по аутосомам, как
правило, приводят к более неблагоприятным последствиям, чем по
половым хромосомам. Влияние одной и той же мутации на
жизнеспособность особи варьирует. Общее число описанных хромо­
сомных мутаций у человека превышает 700. Распределение некоторых
видов аббераций по отдельным хромосомам с полной или мозаичной
формой изменения фенотипа показано в табл. 18.
5.3.5. Геномные мутации
К г е н о м н ы м м у т а ц и я м относят г а п л о и д и ю , п о ­
липлоидию и анеуплоидию.
Анеуплоидией называю т
изменение количества отдельных хромосом — отсутствие
(м он о с о м и я)
или
наличие
дополнительных
(т р и с о м и я,
т е т р а с о м и я , в общем случае п о л и с о м и я ) хромосом, т. е.
несбалансированный хромосомный набор. Клетки с измененным
числом хромосом появляю тся вследствие нарушений в процессе митоза
или мейоза, в связи с чем различают митотическую и мейотическую.
анеуплоидию. Образование гамет с несбалансированным числом
хромосом иллюстрирует рис. 68. Если дочерний организм развивается
из зиготы, в образовании которой участвовала половая клетка с изме­
ненным хромосомным набором, мутация распространяется на все
клетки потомка, что приводит к полной форме изменения фенотипа.
Если мутация возникла вследствие нарушения митотического деления
бластомеров на стадии дробления, ее наследует лишь часть клеток
организма. В этом случае наблюдается мозаичная форма изменения
фенотипа. У человека, по-видимому, лишь моносомия по паре половых
хромосом с кариотипом Х0 совместима с жизнью в постнатальном
периоде. Известны субъекты с три-, тетра и пентасомией по X137
хромосоме, трисомией по У-хромосоме. Совместимые с жизнью
трисомии аутосом наиболее часто затрагиваю т хромосомы 8, 13,
18 и 21, которые отличаю тся о т других аутосом групп С, В , Е и С
относительно высоким содержанием гетерохроматина. Неблагопри­
ятные последствия изменения числа хромосом варьирую т у разных
особей. Так, 2/3 организмов с трисомией 21 погибает в эмбриональном
периоде онтогенеза, тогда как */3 переживает момент рождения и
образует группу больных синдромом Дауна. До рождения доживает
один из 40—50 эмбрионов с моносомией Х 0 (синдром Шерешевского — Тернера). Распределение анеуплодий по парам хромосом дано в
табл. 18.
Уменьшение и увеличение числа хромосомных наборов соматиче­
ских клеток по сравнению с диплоидным обозначают соответственно
как гаплоидию и полиплоидию. Г а п л о и д н ы е о р г а н и з м ы в
естественных условиях обитания обнаруживаются в основном среди
растений, в том числе высших (дурман, пшеница, кукуруза). Так как
клетки таких организмов имеют по одной хромосоме каждой
гомологичной пары, все рецессивные аллели проявляю тся в фенотипе,
чем объясняется сниженная жизнеспособность гаплоидов. Гаплоидные
растения отличаю тся от диплоидных меньшими размерами, что
связано с пониженной вдвое дозой генов. У гаплоидных организмов
нарушается процесс мейоза, что делает их почти бесплодными.
В условиях воздействия на икру резких колебаний температуры в
эксперименте получали гаплоидных тритонов, которые отличались
сниженной жизнеспособностью.
П о л и п л о и д н ы е о р г а н и з м ы часто встречаю тся среди
растений, что объясняется распространенностью в растительном мире
вегетативного размножения и партеногенеза. Так, более трети всех
покрытосемянных являю тся полиплоидными. В природе . среди
животных, не размножающихся партеногенетически, полиплоид­
ные особи редки. В эксперименте полиплоиды получены у насеко­
мых (тутовы й шелкопряд), амфибий (тритон), птиц (индюк), млекопи­
тающих (м ы ш ь, кролик). Различаю т а в т о п о л и п л о и д и ю и
а л л о п о л и п л о и д и ю . В первом случае отмечается умножение
хромосомных наборов одного биологического вида. Во втором случае в
организме объединяются геномы разных видов. К развитию автопо­
липлоидов приводят отклонения о т нормального хода митоза, мейоза,
первого деления зиготы (рис. 69).
Митотическая
пол и- >
плоидизация
происходит вследствие незавершения деления
клетки цитотомией. Все потомки такой клетки имеют увеличен­
ное вдвое число хромосомных наборов и являю тся тетраплоидным.
Так как часть клеток организма остается диплоидной, возникает
мозаичная форма полиплоида. Если увеличение числа хромосомных
наборов по аналогичному механизму происходит при первом делении
зиготы ( з и г о т и ч е с к а я п о л и п л о и д и з а ц и я ) , то тетраплоидными оказываю тся все соматические клетки, что соответствует
полной форме полиплоидного организма. При нерасхождении
хромосом в первом делении мейоза образуются гаметы с нередуциро­
ванным, диплоидным хромосомным набором. Участие такой гаметы в
138
оплодотворении дает триплоидный организм. При
этой разновидности авто­
полиплоидии, называемой
м е й о т и ч е с к о й из­
менение числа хромосом­
ных .наборов распространя­
ется также на все клетки
организма. Аллополиплоидные организмы образу­
ю тся путем м е ж в и д о ­
вой
(отдаленной)
г и б р и д и з а ц и и . Ти­
пичным примером может
служ ить гибрид редьки и
капусты Г . Д . Карпеченко.
В клетках этого гибрида со­
браны хромосомные набо­
ры обоих родителей. Поли­
плоидные организмы
в Рис. 69. Схема полиплоидизации.
сравнении с диплоидными I — м и т о т и ч е ск а я; 2 — з и го ти ч е с к а я : 3 — м ей отическаи, а —
исходные диплоидные к л е т ки ; б — га м еты ; в ‘ — зи готы ;
имеют, как правило, боль­ г — д — со м ати че ск и е клетки о р ган и зм а.
шие размеры благодаря
увеличению размеров клеток.
Из-за нарушения мейоза в условиях увеличенного количества
хромосом у многих полиплоидов снижена плодовитость. Ряд полипло­
идных форм растений имеют повышенную устойчивость к неблагопри­
ятны м факторам внешней среды, что объясняется меньшей вероятно­
стью проявления в фенотипе рецессивных аллелей. Наряду с этим для
автойолиплоидов характерны нарушения онтогенеза. Так, курица,
соматические клетки которой при двух половых хромосомах имели три
набора аутосом, при нормальной жизнеспособности отличалась
недоразвитием правой половой железы и мозаичным строением
(овотестис) левой. У млекопитающих отклонения в развитии более
выражены. У мышей, например, большинство триплоидных эмбрионов
погибает в первой половине беременности. Так как автополиплоиды
имеют сбалансированный набор хромосом, отклонения в развитии
объясняю тся изменением дозы генов.
П о л и п л о и д н ы е з а р о д ы ш и ч е л о в е к а обнаружены
при изучении материала, полученного при выкидышах. Описаны
единичные случая рождения три- и тетраплоидных людей. Продолжи­
тельность жизни новорожденных с полной триплоидией варьировала
от 15 мин до 7 сут. Нарушения фенотипа при этом разнообразны:
отмечается общее соматическое недоразвитие, сращение I I I — IV
пальцев кисти и стоп, дефект межжелудочковой перегородки сердца
(в
случаев), уродства мочеполовой и нервной систем. Мозаичная
диплоидно-триплоидная форма обнаружена у жизнеспособных детей
9— 10 лет.
139
Рис. 70. Соматическая мутация у плодовой мухи
Сектор белою цвета (1) на красном глазе (2)
Рис. 71. Односторонняя гинекомастия (увеличенная
молочная железа) у 14 летнего мальчика
5.3.6. Соматические мутации
М утации, возникающие в телесных клетках на той или иной ста­
дии индивидуального развития организма, называю т
сома­
тическими.
Различаю т генные, хромосомные и геномные со­
матические мутации. Механизмы развития и частота возникновения
соматических мутаций сходны с аналогичными показателями генера­
тивных мутаций. Соматическая мутация наследуется потомками
только той клетки, в которой она произошла, поэтому распространение
ее в организме всегда мозаично. Чем раньше в онтогенезе появляется
такая мутация, тем больше мутантных клеток образуется в результате
последующих делений и тем значительнее объем изменений в фенотипе.
При сохранении мутантными клетками пространственной близости
измененный участок отчетливо выделяется на фоне органа с нормаль­
ным клеточным составом (рис. 70). У видов, размножающихся
исключительно половым путем, соматические мутации в ряду
поколений организмов не передаются. Вегетативное размножение
открывает такую возможность и позволяет использовать соматические
мутации в селекции, например растений. Так И . В. Мичуриным был
создан сорт Антоновка 600-граммовая. Соматической мутацией по тийу
анеуплоцдии, заключающейся в утрате У-хромосомы, можно объяс­
нить мозаичный фенотип мальчика (рис. 71).
У животных и человека в определенных органах, например печени,
закономерно имеются полиплоидные клетки, число которых увеличива­
ется с возрастом. Учиты вая более крупные размеры таких клеток,
избирательная
соматическая
полиплоидия
представляет собой полезное явление. Она способствует увеличению
функциональных возможностей органа в условиях, когда достичь этого
путем интенсификации пролиферации клеток невозможно.
Изменение числа хромосом и их морфологии типично для клеток
злокачественных новообразований животных и человека. Одинаковые
140
изменения наблюдаются в
опухолях разной природы.
Некоторые новообразова
ния однако имеют специфи
ческие нарушения кариотипа. Так, при хрониче­
ском миелоидном лейко
зе — опухолевом разраста­
нии кроветворной ткани
костного мозга — в изме­
ненных клетках регистри­
руется делеция длинного
плеча хромосомы 22 с
транслокацией отделивше­
гося фрагмента на хромо­
сому 9. Для острой формы
этого заболевания типич­
на, хотя и не обязательна,
транслокация наследствен­
ного материала между 8 и
21 хромосомами.
сГ
1<г
Рис. 72. Клеточные механизмы комбинативный
изменчивости.
I
га м е гы и сх о д н о й п а р ы родителей ; 2
ге н о тип п отом
ка Р 1, 3
га м е ты потом ка Р , , 4
ген отип ы п о то м к о в
5.3.7. Комбинативная изменчивость
К о м б и н а т и в н о й называю т изменчивость, которая зависит
от перекомбинации аллелей в генотипах потомков по сравнению^ с
'генотипами родителей. В отличие от перераспределения наследственно­
го материала нерегулярного типа при хромосомных абберациях такая
рекомбинация у организмов, размножающихся половым способом,
происходит закономерно.
Сущ ествует
три
механизма
комбинативной
изменчивости
(рис. 72). Два из них связаны с мейозом. Благодаря обме­
ну генетическим материалом между гомологичными хромосомами при
кроссинговере аллельный состав хромосом половых клеток потомка
изменяется в сравнении с таковым гамет родителей. В связи с неизве­
стным характером расхождения отцовских и материнских хромосом
каждой гомологичной пары в анафазе редукционного деления в хромо­
сомных наборах гамет потомка представлены одновременно хромосо­
мы, унаследованные от обоих родителей, причем в разных сочетаниях
(на рис. 72, см. 1, 3). Третий механизм комбинативной изменчивости
связан с оплодотворением. Благодаря случайному подбору гамет,
участвующих в оплодотворении, создается разнообразие сочетаний
хромосом и, следовательно, аллельного состава зигот (см. рис. 72, 4).
Явление комбинативной изменчивости объясняет, почему в
детях и внуках сочетаю тся признаки родственников по отцовской или
материнской линии.
141
5.3.8. Медико-генетический аспект брака
Ж ивы е сущ ества, вклю чая человека, являю тся
генети­
ческими
наследниками
своих предков, что проявля­
ется в наличии в генотипах родственников определенной доли общих
аллелей. Вероятность наличия у каждых из двух индивидуумов в
известном локусе аллелей, полученных от общего предка и идентичных
по происхождению, называю т к о э ф ф и ц и е н т о м р о д с т в а .
Он выражается в долях единицы. Так, применительно к человеку, для
родителей и детей, братьев и сестер, исключая монозиготных
близнецов, этот коэффициент составляет 1/2. Значения его для других
степеней родства приведены в табл. 19.
Таблица
родства
19.
Д оля
общих
аллелей
Степень родства
Родитель, ребенок, сибс (брат, сестра)
Монозиготные близнецы
Дизиготные
»
Дед, внук, дядя, тетя, племянник, племянница
Двоюродные сибсы (братья, сестры)
Троюродные
у
людей
разной
степени
кровного
Д оля общих аллелей
7.
Уо
У/
74
/32
Понятие родства в том смысле, в каком оно используется здесь, нес­
колько неопределенно. В эволюционном смысле, например, все люди на
известном этапе существования человечества имели общих предков и,
следовательно, состоят в родстве. Об этом свидетельствую т простые
расчеты. Ж ивущ ие ныне люди имеют по два родителя, четыре пра­
родителя, восемь прапрародителей. Таким образом п поколений том у
назад каждый из них имел 2" предков. Примем, что в среднем за столе­
тие сменяется четыре поколения. Тогда 1000 лет том у назад у любых
двух индивидуумов наших дней, не состоящих в родстве друг с другом,
ймелось бы 240 предков. Известно, однако, что в X веке население пла­
неты не превышало 200 млн. человек. В сравнительно недалеком
прошлом, однако, человечество было представлено совокупностью
относительно малочисленных групп, в пределах которых и заключались
браки. Лю ди каждой группы имели более высокий показатель
генетического родства, чем индивидуумы из разных групп.
У человека аллельный состав генотипов потомков зависит от
с и с т е м ы б р а к о в , практикуемой конкретной группой людей на
протяжении длительного периода времени. При
неизби­
р а т е л ь н ы х б р а к а х ( п а н м и к с и и ) сущ ествует равная ве­
роятность вступления в брак любого индивидуума одного пола с лю­
бым индивидуумом противоположного пола из числа лиц, входящих
в данную группу. Если индивидуумы, состоящие в родстве, вступаю т
в брак чаще, чем при случайном подборе брачных пар, говорят об
и н б р и д и н г е , если реже — об а у т б р и д и н г е . Сущ ествую т
142
также системы
положи­
Самооплодотворение
тельного
и отрица­
100
тельного
ассорта90
тивного
(избирательного)
образования *
брачных пар. В первом случае | 8 0
отмечается более частое, а во °
70
втором — более редкое вступ- §
ление в брак индивидуумов, I 6 0
сходных по определенным
50
фенотипическим признакам.
Так, браки, между глухонемы0
2
4
6
8 ю 12 и 15
М И , а также Л Ю Д Ь М И , С Х О Д Н Ы Поколения, инбридинга
ми по росту или умственному
развитию ,наблюдаются Чаще, рис 7 3 Изменение гомозиготности потомков
чем М О Ж Н О было бы ожидать в ряду поколений при разной степени генетипри панмиксии. Напротив, ческого родства скрещиваемых особей,
рыжеволосые мужчины и жен­
щины избегают вступать в
брак друг с другом. Генетические последствия разных систем браков
заклю чаю тся в изменении уровня гетерозиготности потомков. На
рис. 73 показано изменение уровня гомозиготности потомства в ряду
поколения под влиянием скрещиваний при различной степени родства
родителей. При самооплодотворении, являющ емся наиболее тесной
формой инбридинга, относительная доля гетерозигот в следующем
поколении снижается вдвое, что обусловливает высокую скорость
перехода в гомозиготное состояние. При снижении степени генетиче­
ского родства родителей указанный процесс резко замедляется. Так,
шесть поколений самооплодотворения оказывается более эффектив­
ным, чем семнадцать поколений браков типа «брат-сестра» или «роди­
тель-потомок». Сходным с инбридингом, но более Мягким действием
обладает положительное ассортативное скрещивание.
Величину гомозиготизации организма вы ражаю т через
к оэффициент
инбридинга
(Е ),
который
показывает
вероятность наличия в определенном локусе хромосом родителей
аллелей, идентичных по происхождению. Другими словами, этот
коэффициент отражает долю локусов в генотипе потомка конкретной
пары родителей, по которым он гомозиготен:
Р =
2
(1 + Рг),
где и и и '— число поколений, считая от общего предка до родителей ин­
дивидуума; Р г — коэффициент инбридинга для общего предка.
Если предок неинбреден, то Р 2 =0, а Г = (^ - )и+и' +|. Так, двою­
родные сибсы отделены от своих прародителей двумя поколения­
ми. Тогда, по возможным линиям родства (от деда или от бабки), если
сами прародители неинбредны, для потомства двоюродных сибсов
коэффициент инбридинга равен:
(общ ий предок
(общ ий предок
ж ен щ и н а)
143
Коэффициенты инбридинга лиц, рождаемых в браках «родитель —
потомок», равен /« , а «дядя (тетя) — племянница (племянник)» — /в.
В группах людей относительно редко встречаю тся так называемые
и н ц е с т н ы е б р а к и (запретные) между родственниками первой
степени родства («отец — дочь», «брат — сестра»). В истории че­
ловечества столь тесная форма инбридинга как система допускалась
в отдельных случаях, например в династии Птоломеев, правивших в
Египте в 305— 30 годах до н. э. В племени эрнодан с Малабарского
берега полуострова Индостан старшая дочь обычно становилась второй
женой отца. В настоящее время браки такого рода запрещены почти
повсеместно. Во многих государствах запрещены браки «племянник
(племянница) — тетя (дядя)». В С Ш А более чем в трети ш татов
запрещены браки между двоюродными сибсами. Частота браков
последнего типа в большинстве западноевропейских стран ниже 1 % .
В высокогорных деревнях Швейцарии, еврейских общинах многих
городов Германии частота браков между двоюродными родственника­
ми составляет 6— 12%. Частота родственных браков по Самаркандской
области в среднем равна 11,6% (табл. 20). Высокая частота близкород­
ственных браков типична для малых по размерам групп людей,
изолированных в силу географических (высокогорье), экономических,
религиозных (секта самаритян), национальных и расовых особенностей
уклада жизни или предрассудков.
Медицинское значение различных систем браков обусловливается
тем, что в условиях инбредного размножения при близкородственных
браках среди потомков растет число гомозигот, в том числе и по
локусам вредных рецессивных аллелей. Длительно практикуемый ин­
бридинг у растений и животных часто, хотя и не всегда вызывает сни­
жение жизнеспособности и плодовитости. Предполагают, что такая
«инбредная
д е п р е с с и я » является следствием утраты по
многим локусам гетерозиготности. О сниженной жизнеспособности
потомства родителей-родственников говорит увеличенная детская
смертность в таких семьях (табл. 21). В них повыш ается также
вероятность наследственных заболеваний, передающихся по аутосомно-рецессивному типу. Вместе с тем известны примеры длительно
Таблица
20.
Структура браков (Самаркандская область)
Ти п ы браков
По родству:
Всего обследовано
неизвестные случаи
супруги не состоят в родстве
супруги состоят в родстве:
дядя — племянница
Племянник — тетя
двоюродные сибсы
троюродные сибсы
дальние родственники
инцестиый брак
144
С ельское
н а се л е н и е
4988
362
4026
600
48
14
199
72
265
2
Городское
н а се л е н и е
1998
142
1646
210
4
4
129
21
52
0
И то го
6986
504
4862
810
52
18
328
93
317
2
Таблица
21.
Детская смертность в потомстве родителей, состоящих и
несостоящих в родстве, на примере трех местностей во Франции (н а 1000 случаев
рождения)
М е стн о сть
Мод о НО
родителей
М е р тв о
рожден н ые
Ум ер ш ие и во зр асте
П
Морибан
Ф и нистер
Л у ар
Нет
Есть
Нет
Есть
Нет
Есть
21
50
21
28
19
26
2 мес
23
43
13
31
21
30
2
12 м сс
39
38
25
50
19
24
о/ |ода
и ниш е
55
96
70
96
39
56
существующих малочисленных групп людей, которые не испытываю т
вредных последствий инбридинга.
Аутбридинг поддерживает высокий уровень гетерозиготности, что
иногда приводит к повышению жизнеспособности. Х о тя это строго не
доказано, увеличение роста жителей Европы в текущем столетии
(явление акселерации) может бы ть отчасти связано с преодолением
ограничений в заключении браков.
5.3.9. Генетическая инженерия
Формы генотипической изменчивости, рассмотренные выше,
вы зы ваю т изменение содержания или перераспределение наслед­
ственной информации в процессе развития организмов в естественных
условия^. В настоящее время возникло новое направление молеку­
лярной биологии н генетики — г е н с-л: д_й..е.С-К.а,А .. ...и н ж .е н ер и я, которая занимается направленным изменением биологической
ш т ормяци^ кДеток или^' организмов для получения живых сущ еств с
заданными фенотипическими характеристиками. Задачи генетической
инженерии разнообразны, что объясняет разные уровни ее примене­
ния — организменный, клеточный, генный.
Представление об о р г а н и з м е н н о м у р о в н е п р и м е ­
н е н и я г е н е т и ч е с к о й и н ж е н е р и и дает пример а л л оф е н н ы х ж и в о т н ы х . Тела их состоят из генотипически разных
тканей, развившихся из клеток нескольких родителей, искусст­
венно объединенных в данном потомке (рис. 74). Так, зародыши мы­
шей на стадии восьми бластомеров, извлеченные из двух и более
матерей, в пробирке разбиваются на отдельные бластомеры. Путем
комбинации бластомеров создается комплексны# эмбрион, который на
стадии гаструлы вводится в м атку мыши-воспитательницы. Рожден­
ный аллофенный мышонок объединяет в фенотипе черты всех родите­
лей, но вместе с тем отличается некоторыми уникальными свойствами.
Интересен, например, факт нормального развития и жизнедеятельности
аллофенного потомка, составленного из клеток родителей, ткани
которых во взрослом состоянии иммунологически не совместимы.
145
Д р об ящ и еся
яйца
П роназа
37°С
Бластула
Рис. 74. Получение аллофеиных мышей путем образования смешанной бластулы,
а — процедуры, о с ущ е ств л яе м ы е в пробирке; б — введение смеш анной б л а сту л ы в м а т к у приемной м а ­
тери н р ож дение аллоф енного п отом ка.
П утем объединения цитоплазмы яйцеклетки и ядерного материала
соматической клетки с последующим развитием полноценного орга­
низма создаются генетически тождественные особи даже у млекопита­
ющих. В селекции это открывает возможность прямой передачи
потомкам особо ценного генотипа в обход явления комбинативной
изменчивости.
На к л е т о ч н о м
уровне применения
генети­
ческой
инженерии
путем соматической ги б р и д и зац и и по­
лучаю т гибриды, совмещающие в одной клетке генотипы орга­
низмов разных биологических видов. Особенность гибридов «че­
ловек-мышь», например, заклю чается в постепенном удалении из
клетки человеческих хромосом. Наблюдения за изменениями клеточно­
го фенотипа после потери отдельной хромосомы уточняю т генный
состав групп сцепления человека.
Эффективным путем управления наследственностью на генном
уровне является г е н н а я и н ж е н е р и я . Она объединяет в себе
методы получения отдельных генов и введения их в геномы других
организмов с целью изменить фенотип последних. Основные этапы
процесса изменения генотипа и фенотипа в этом случае заклю чается в
следующем: 1) выделение гена из клетки-донора или искусственный его
синтез; 2) присоединение гена к молекуле Д Н К (вектору), способной
ввести его в клетку-реципиент; 3) включение гена в геном клеткиреципиента; 4) активация гена и проявление информации в фенотипе
клетки-реципиента путем ее транскрипции и трасляции. Роль векторов
выполняю т фаги, вирусы, плазмиды, эписомы, митохондриальная
Д Н К.
Достижения генной инженерии использую тся в нескольких
направлениях. Б и о т е х н о л о г и ч е с к о е
направление
146
заключается в получении микроорганизмов, которые благодаря
внедрению соответствующ их генов синтезируют биологически актив­
ные вещества. Так, получены ш таммы микроба кишечной палочки,
продуцирующие соматостатин и инсулин человека — гормоны,
необходимые для лечения сахарного диабета. Селекционеры путем
пересадки генов азобактерий пытаю тся получить растения, фиксирую­
щие азот непосредственно из воздуха. Некоторые перспективы
открываю тся и в области медицины. Введение в организм соответству­
ющих генов при дефективности собственных устранит наследственно
обусловленные нарушения обмена веществ.
Генная инженерия служит мощным орудием изменения наслед­
ственности живых организмов на благо человека. Н о безответ­
ственность в исследованиях такого рода таит опасность глобальной
катастрофы в связи с появлением патогенных свойств у микроорга­
низмов, в обычных условиях безвредных для человека. На Междуна­
родной конференции в г. Асиломаре (С Ш А ) в 1975 г. выработаны тре­
бования, соблюдение которых снижает вероятность неблагоприятных
последствий генной инженерии. Так, в качестве клеток-реципиентов
предлагается использовать ш там м ы «саморазрушающихся бактерий»,
которые при температуре человеческого тела погибают.
5.3.10. Медико-генетическое консультирование
Механизмам наследственности принадлежит ведущая роль в про­
цессах развития и жизнедеятельности живых организмов, поэтому
изменения в наследственном материале нередко приводят к нарушению
здоровья человека — возникновению наследственных болезней. В зави­
симости от уровня, на котором произошло изменение, выделя­
ю т генные и хромосомные болезни. В основе г е н н ы х
на­
следственных
болезней
наиболее часто лежат не­
благоприятные по фенотипическим проявлениям мутации структур­
ных генов. Группа х р о м о с о м н ы х б о л е з н е й представлена
патологическими состояниями, причиной которых служат хромо­
сомные или геномные мутации. К наследственной патологии от­
носят
также
болезни
с
наследственным
п р е д р а с п о л о ж е н и е м . В и х развитии решающее значение
имеет определенное сочетание генетических и внешних факторов.
В диагностике наследственных болезней широко и эффективно
используются данные клинических лабораторных исследований. Иногда
для постановки и уточнения диагноза таких заболеваний приходится
прибегать к специальным генетическим методам. Особенностями
многих генных и хромосомных болезней служ ат возможность воспро­
изведения их в потомках, а также гетерозиготное носительство
неблагоприятных рецессивных аллелей клинически здоровыми индиви­
дуумами. В обоих случаях встает задача прогнозирования здоровья
потомства, которая решается на данных специального генетического
обследования.
Помощ ь лечащему врачу в постановке диагноза наследственного
заболевания, определение прогноза здоровья будущего или родившегося
147
ребенка проводится врачом-генетиком в рамках специализированной
медицинской помощи — м е д и к о - г е н е т и ч е с к о г о
кон­
сультирования.
Медико-генетическое консультирование вклю чает три главных
этапа. Н а первом этапе проводится уточнение диагноза с использовани­
ем специальных генетических методов. Выполняется подробное генеа­
логическое обследование и составляется родословная, что позволяет
сделать заключение о типе и варианте наследования патологического
состояния. С помощью биохимико-генетических методов устанавлива­
ю тся генетически обусловленные изменения обмена вещ еств. Э ти
данные ценны тем, что они нередко вы являю т непосредственную
причину патологического состояния в виде дефекта синтеза опреде­
ленного фермента. При наличии клинических проявлений хромосомной
болезни показано применение цитогенетического метода. При подозре­
нии на заболевание, связанное с нарушением синтеза иммуноглобули­
нов, проводится иммуногенетическое обследование, включающее ана­
лиз глобулиновой фракции белков крови и тестов на функциональное
состояние разных типов лимфоцитов.
Диагностика гетерозиготного носительства рецессивных аллелей
основана на выявлении фенотипических различий между гомо- и
гетерозиготными индивидуумами, главным образом по количеству
первичного продукта генной активности. У лиц, гетерозиготных по
аллелю фенилкетонурии, например, концентрация фенилаланина, в кро­
ви, особенно на фоне введения в организм аминокислоты извне, выше,
чем у нормальных гомозигот. Гетерозиготы по аллелю серповидноклеточности отличаю тся наличием в эритроцитах, наряду с нормальным,
аномального гемоглобина, что устанавливается биохимически (см.
рис. 61).
Все более широкое применение в целях прогноза здоровья ожи­
даемого ребенка приобретает
пренатальная
(дородо­
вая)
диагностика.
Метод
ультразвукового
скеннирования
или
ультрасонографии
основан на регистрации пропускания тканями плода ультра­
звуковых волн. С его помощью определяются анатомические
отклонения и нарушения пропорционального развития частей
организма. В отличие от рентгенографии этот метод безопасен для
матери и плода. Перспективен метод а м н и о ц е н т е з а ,
за­
ключающийся в заборе шприцем, обычно на 15— 16-й неделе бере­
менности, околоплодной жидкости с содержащимися в ней спущенными
клетками плода. Далее жидкость и клетки, прямо или после
культивирования на питательной среде, подвергают цитологическим,
цитогенетическим, биохимико-генетическим, иммуногенетическим ис­
следованиям. В случае обнаружения тяжелой патологии производится
операция искусственного прерывания беременности.
Н а втором этапе медико-генетического консультирования определя­
ется прогноз потомства, который основывается на данных о типе и
варианте наследования патологического состояния, результатах прена­
тальной диагностики.
Третий этап медико-генетического консультирования состоит в
148
формулировании заключения и объяснении заинтересованным лицам в
доступной форме смысла генетического риска. Основная задача врачагенетика на этом этапе заключается в консультативной помощи семье
при принятии решения о деторождении.
Для оказания специализированной генетической помощи организу­
ю тся медико-генетические консультации, выполняющие четыре глав­
ные функции: 1) определение прогноза в семьях с наследственной
патологией; 2) разъяснение величины риска и помощь в выборе
решения; 3) уточнение диагноза наследственного заболевания для леча­
щего врача; 4) пропаганда медико-генетических знаний.
Согласно ориентировочным расчетам в консультациях врачагенетика нуждаются 5—25% всех семей.
Глава
6
Б И О Л О ГИ Я РА ЗВ И Т И Я
6.1. О П Р Е Д Е Л Е Н И Е И Т И П Ы
И Н Д И ВИ Д У А Л ЬН О ГО РА ЗВИ ТИ Я
Во времени жизнь организована как смена поколений организмов.
Организмы каждого поколения осущ ествляю т закономерный процесс
развития или ж и з н е н н ы й
ц и к л . Наиболее демонстративен
жизненный цикл многоклеточных растений и животных, размножаю­
щихся половым способом, который начинается одной клеткой —
зиготой. Совершающиеся в определенной'последовательности преобра­
зования клеток, образующихся в результате деления зиготы и ее
потомков, обусловливают рост организма, выделение в нем клеток
разных направлений специализации и частей, различающихся строением
и выполняемыми функциями, и наконец, достижение состояния
зрелости. Зрелый организм выполняет главную биологическую зада­
чу — воспроизведение особей следующего поколения. В дальнейшем
организм стареет, что проявляется в снижении уровня его жизнедея­
тельности. Жизненный цикл завершается смертью . Жизненные циклы
некоторых одноклеточных эукариот и микроорганизмов нередко
исчерпываются клеточным циклом. Их усложнение связано с возмож­
ностью образования цист или спор, включением стадии полового
размножения. Переходной формой между циклами одноклеточных и
многоклеточных организмов служит жизненный цикл некоторых
колониальных простейших, например У о 1у о х . В отличие от однокле­
точных у них происходит стабильное выделение в развитии линий
генеративных и соматических клеток, однако не наблюдается
разнообразия морфофункциональных специализаций соматических кле­
ток. У многих простейших и низших многоклеточных циклы отлича­
ю тся высокой степенью сложности.
Совокупность взаимосвязанных и детерминиппванных хронологических событий, закономерно совершающихся в процессе осуществления
организмом жизненного цикла, обозначают терминами « о н т »
г е н е з» или « и н д и в и д у а л ь н о е р а з в и т и е » ,
Различаю т два главных типа индивидуального развития —
;н е п п я м о е (с м е т а м о р ф о з о м ) и п р я м о е . Первый из
названных типов- характеризуется наличием особой вставочной фор­
мы — л и ч и н более или менее отличной о т зрелой особи по
строению тела и ведущей активный образ жизни. Некоторые личинки
имеют органы захвата и переработки пищи, тогда как другие снабжены
лишь органами расселения. Последнее типично для личинок паразитов
(мирацидий и циркария сосальщиков, корацидий широкого лентеца).
Совокупность процессов, в результате которых происходит переход от
личиночной к взрослой форме, называется м е т а м о р ф о з о м . Он
заключается в изменении внешнего вида и строения животного и
достижении им половозрелого состояния. Нецрямой тип индивиду­
ального развития свойствен видам, откладывающ им яйца с относи­
тельно малы м количеством^ж елтка.
При прямом развитии зародышевый период заканчивается
рождением молодой формы, имеющей общий план строения, набор
органов и систем, характерный для зрелого состояния, но отличающ ей­
ся меньшими размерами, функциональной и структурной незрелостью
органов и систем. Э то т тип развития присущ животным, откладываю ­
щим яйца с высоким содержанием желтка.
Характерные
особенности
имеет
ти п
развития
п л а ц е н т а р н ы х м л е к о п и т а ю щ и х и ч е л о в е к а . Он
является вариантом прямого развития, но отличается тем, что
непосредственно по окончании зародышевого периода после рождения
новый организм не способен к самостоятельному образу жизни, так как
нуждается в специфическом питании — секрете определенных желез
материнского организма (молоко).
Изменения в индивидуальном развитии проявляю тся на разных
уровнях организации особи — генетическом, молекулярно-биохимиче­
ском, клеточном, тканевом, органном, системном. Исследования
индивидуального развития проводятся с участием специалистов многих
отраслей биологической науки — генетиков, биохимиков, морфоло­
гов, эмбриологов, молекулярных биологов. Усиление роли междисцип­
линарных исследований онтогенеза, наметившееся в начале текущего
столетия, привело к возникновению самостоятельной области науки о
живом — б и о л о г и и р а з в и т и я . Она изучает наследственные,
молекулярные, структурные основы, а также механизмы регуляций
онтогенетических изменений на всех этапах жизненного цикла особи.
Основу процесса индивидуального развития составляет наслед­
ственная информация, получаемая потомками от родителей. Д оста­
точно, однако, сравнить, например, человека на начальной, однокле­
точной стадии онтогенеза и во взрослом состоянии, чтобы прийти, к
заключению о том , что в ходе развития объем информации,
воспроизведенный в структурах и метаболизме организма, возрастает.
Об этом свидетельствует, в частности, большее разнообразие химиче­
ских соединений, их неслучайное распределение в органах, наличие
самих органов и многое другое, что мы наблюдаем у взрослой особи и
не обнаруживаем в зиготе. Накопление информации в процессе развития
служит важной чертой онтогенеза и свидетельствует о его системном
150
характере. Первичная наследственная информация зиготы играет роль
инструкции, в соответствии с которой при активном регулирующем
влиянии факторов окружающей среды в развивающемся организме
последовательно образуются и закономерно взаимодействуют друг с
другом молекулы и структуры разных уровней сложности. С учетом
этого замечания онтогенез можно определить как процесс реализации
потомкам наследственной^инФорм'йиии родителей в определенных
условиях окружающей ср ед ы .' ЭТо 'определение подчеркивает, что
генетические закономерности играют важную роль в индивидуальном
развитии, но не исчерпывают всего его содержания.
Кроме зародышевого развития, роста, старения биология развития
изучает также молекулярно-генетические, клеточные и системные
механизмы регенерации — совокупности процессов, обусловливающих
восстановление структур, снашиваемых в процессе жизнедеятельности
организма или утрачиваемых вследствие травм ы.
6.2. П Е Р И О Д И З А Ц И Я О Н Т О Г Е Н Е З А
Онтогенез представляет собой непрерывный процесс развития
особи. Однако для удобства изучения, а также в связи с тем , что на от­
дельных этапах его происходит смена преобладающих молекулярных,
клеточных и системных механизмов и характера отношений организма
с окружающей средой онтогенез многоклеточных организмов подразде­
ляю т на периоды и стадии. Предложено несколько схем периодизации
индивидуального развития. В соответствии с одной из них, имеющей
широкое распространение, выделяю т э м б р и о н а л ь н ы й
и
п о с т э м б р и о н а л ь н ы й периоды. У плацентарных животных
и человека выделяю т д о р о д о в ы й ( а н т е н а т а л ь н ы й ) и
п о с л е р о д о в ы й ( п о с т н а т а л ь н Ы й ) п е р и о д ы . Пер­
вы й” охватывает развитие до рождения особи и происходит под
покровом яйцевых оболочек, а у плацентарных в материнском
организме. В этот период факторы окружающей среды оказываю т на
развивающийся организм опосредованное действие. После рождения, с
началом постнатального периода принципиальным образом меняются
условия существования организма. Он начинает самостоятельную
жизнь, вступая в непосредственное взаимодействие с окружающей
средой.
Названные периоды онтогенеза подразделяются на стадии,
различающиеся по конкретному содержанию изменений. У животных,
размножающихся половым способом, эмбриональный период пред­
ставлен Следующими стадиями: одноклеточной (зигатдТ дробления
(образование однослойного зародыша бластулы ), гаструляции (образо­
вание трехслойного зародыша), гисто- и органогенезов (образование
тканей и органов). В первые _8,нед развивающийся организм человека
называю т эмбрионом или зародышем] что соответствует прохождению
им зародышевой стадии. С 9-й недели начинается плодная стадия
развития. Организм приобретает характерные наружные формы, в нем
обособляются закладки органов. Н а этой стадии он называется плоде*'
В постнатальном периоде при прямом типе развития выделя!
ранний и поздний постнатальный онтогенез. При этом к раннему
постнатальному онтогенезу относят отрезок жизни до приобретения
черт структурно-функциональной и репродуктивной зрелости, а к поз­
днему — отрезок жизни, соответствующ ий зрелому состоянию и старе­
нию организма. Дальнейшее подразделение наиболее детально прово­
дится для человека. Оно обосновывается результатами исследования
возрастной физиологии и медицины. Так, в^раннем постнатальном
онтогенезе человека выделяю т} периоды новорожденное™, 9грудного
возраста,Дош кольного иЦшкольного возраста, ^полового созревания
(пубертатный). Выделение их способствует оптимальному решению
практических задач педиатрии, так как ранний постнатальный
онтогенез характеризуется относительно быстрой сменой функцио­
нальных показателей различных органов и систем организма.
Соответственно меняются требования к характеру питания, гигиениче­
скому режиму, а также выносливость по отношению к температурным,
физическим, эмоциональным нагрузкам.
Схема периодизации онтогенеза, которой мы будем придерживаться
в дальнейшем, вытекает из сущ ества генетических механизмов
индивидуального развития, которое рассматривается как процесс
реализации наследственной информации, обусловливающий достиже­
ние состояния зрелости и участие организма в репродукции. В этой
схеме, отражающей общебиологические закономерности, выделяю т
д о р е п р о д у к т и в н ы й,,; з р е л ы й ( а к т и в н ы й р е п р одуктивныЙ)
и
пострепродуктивный
пеои,6 д ы. П ервый из них, начинаясь с момента образования зиготы, огра­
ничивается достижением половой зрелости и может бы ть назван
также п е р и о д о м р а з в и т и я д е ф и н и т и в н о г о ф е н от иТГа, второй —
периодом
с т а б и лГь н о г о
фу нк { ц и о н и р о в а н"и я о р г а н о в
й с и с т е м ^ третий — гГё^
р и о д ом
старения
организма.
Одним из главных
критериев выделения периодов согласно приводимой схеме является
участие организма в репродукции, что создает трудности с установле­
нием точных границ периодов. В частности, у млекопитающих и
человека состояние половой зрелости достигается развивающимся
организмом нередко раньше, чем он реально получает возможность
активно участвовать в размножении. Репродуктивный и пострепро­
дуктивный периоды онтогенеза женщины разграничены достаточно
отчетливо (менопауза). Стареющий мужчина сохраняет способность к
воспроизведению потомства, однако активность его в этом отношении
снижается. Соответственно снижается доля участия в формировании
состава генофонда следующего поколения. В силу социальной сущности
биологический критерий зрелости, используемый в рассматриваемой
схеме, в отношении человека дополняется показателями эффективности
обучения, трудовой деятельности, творческой активности людей в
разные возрастные периоды.
Дорепродуктивный период включает эмбриональное развитие и
ранний постнатальный онтогенез, выделяемые согласно первой схеме
периодизации. Х о тя акт рождения принципиально изменяет характер
отношений между организмом и внешней средой, в раннем постна152
тальном периоде в сравнении с эмбриональным периодом главное
направление развития сохраняется. В частности, продолжаются формо­
образовательные процессы, рост организма, происходят изменения
клеточного состава и межтканевых отношений в различных органах.
Однако если в эмбриональном периоде формообразовательные про­
цессы доминируют, то в раннем постнатальном онтогенезе происходит
смена этих процессов обычными формами жизнедеятельности,
характерными для каждого органа во взрослом состоянии.
В последнее время появились основания для выделения в индивиду­
альном
развитии
предзародышевого
(предэмбрионального)
п е р и о д а , который соответствует гаметогенезу. Такое выделение обосновывается тем, что в дополне­
ние к выработке питательного материала зародыша желтка в оогенезе синтезируются и сохраняются в цитоплазме ооцитов до начала
развития некоторые биологически важные макромолекулы, например
информационные Р Н К , контролирующие ранние стадии эмбриогенеза.
6.3. П Р Е Д Э М Б Р И О Н А Л Ь Н Ы Й
П ЕРИ О Д
П редэмбриональный
период
или п р о г е н е з
заключается в образовании половых клеток. Яйцеклетки и сперматозоиды развиваю тся из п е р в и ч н ы х
половых
клеток,
которые обособляются на ранних стадиях эмбриогенеза. У аскариды и
веслоногого рака циклопа это происходит после первых делений
дробления, у моллюсков, насекомых, высших позвоночных — несколь­
ко позже, но до наступления стадии гаструлы.
' Области зародыша, в которых обнаруживаются первичные половые
клетки, различаются у разных групп организма. У млекопитающих, как
и у других позвоночных, первичные половые клетки образуются во
внезародышевой мезодерме, отличающейся ранним развитием крове­
носных сосудов и повышенной в сравнении с другими областями
зародыша концентрацией питательных веществ. Впоследствии они
мигрируют к месту своей окончательной локализации в развивающуюся
половую железу — яичник или семенник.
Образование первичных половых клеток, по-видимому, связано с
наличием особой « п о л о в о й ц и т о п л а з м ы », располагающей*
ся обычно на вегетативном полюсе яйцеклетки. У многих животных
участок цитоплазмы яйца, ответственный за выделение первичных
половых клеток, отличается пигментацией или содержанием гранул,
названных п о л о в ы м и д е т е р м и н а н т а м и . В яйцеклетках
двукрылых и лягушки эти гранулы видны еще до начала развития.
В последующем они попадают в цитоплазму первичных половых
клеток. Последние нередко отличаю тся от других клеток зародыша
большей "величиной, хорошо заметным ядром, некоторыми цитохими­
ческими особенностями, например более высокой активностью фермен­
та щелочной фосфатазы. Использование цитохимических методов
маркировки позволило проследить возникновение и миграцию первич­
ных половых клеток у человека (рис. 75). Они появляю тся в стенке жел­
точного мешка спустя 24 сут после оплодотворения. У 4-недельного
153
Рис. 75. Первичные половые клетки человека.
I - в желточном мешке; 2 - в кровеносных сосулах; о — в Брыжейке; 4 — в половой железе.
зародыша эти клетки обнаружи­
ваю тся в брыжейке задней киш­
ки, а спустя неделю — в зачатке
половой железы.
Вплоть до недавнего времени
было неясно, служ ат ли первич­
ные половые клетки единствен­
ным источником
оконча­
тельных
(дефинитив­
ных) п о л о в ы х к л е т о к ,
или же они стимулируют превра­
щение в гаметы клеток так назы­
ваемого зачаткового эпителия
половой железы. Сейчас можно
считать доказанным, что все дефинитивные гаметы происходят
И3
перВИЧНЫХ ПОЛОВЫХ КЛвТОК.
^
________
Образование окончательных
половых клеток из первичных
происходит в процессе гаметогенеза. Последовательность событий и
механизмы, приводящие к возникновению гаплоидных гамет, описаны
в главе 4. Наряду с этим в ходе оогенеза наблюдается ряд изменений,
которые непосредственно связаны с обеспечением развития нового
организма на ранних этапах, следующих за оплодотворением. Э ти
изменения целесообразно рассмотреть в настоящей главе.
V
Развитие женских половых клеток происходит в яичниках (рис. 76).
Цервоначально половые клетки занимаю т поверхностный слой яичника,
называемый герминативным.~”Ёщ е в эмбриогенезе у млекопитающих
они окружаю тся' так называемыми
фолликулярными
к л е т к а м и , что приводит к образованию п е р в и ч н ы х и л и
п р и м о р д и а л ь н ы х ф о л л и к у л о в . Функция фолликуляр­
ных клеток сводится, в основном, к обеспечению притока в яйцеклетку
питательных веществ — белков, жиров, аминокислот. Синтез этих
веществ осущ ествляется клетками вне яичника, например в печени.
Можно сказать, что в питании яйцеклетки принимает участие весь
организм. Благодаря ргымножению клеток стенка фолликула стано­
вится многослойной, а сам он при этом увеличивается в размерах.
В процессе дальнейшего роста фолликула фолликулярные клетки
внутри слоя
постепенно
расходятся, а образующаяся полость
заполняется
жидкостью.
В результате
возникает
зрелый
ф о л л и к у л или пузырчатый яичниковый фолликул, содержащий
яйцеклетку, способную к оплодотворению. В яичнике женщин, достиг­
ших половозрелого возраста, ежемесячно созревает обычно один
фолликул, в результате разрыва стенки которого яйцеклетка, окру­
женная слоем фолликулярных клеток, покидает яичник и попадает в
яйцевод. Разры в зрелого фолликула с выходом яйцеклетки называется
о. в у л я ц и е й. После овуляции фолликул сокращается, и его полость
заполняется клетками, содержащими в цитоплазме желтый пигмент.
Образуется ж е л т о е т е л о ,
функционирующее как временная
154
Р и с . 76. Развитие яйцеклеток в яичнике женщины.
с кий Пф о ° ИЧНЫе *п ^,и м о ^’ д и ал ьн ь,0* Ф о л л и к ул ы ; 2 -
р а ст ущ и е ф о л л и к у л ы ; 3 — зр е л ы й ф о л л и к у л ; 4 — о в у л я ц и я ; 5 —
ж е л т о е тело; 6 — белое тело; 7 -
атретиче
железа внутренней секреции. В том случае, если оплодотворения не
происходит, желтое тело подвергается обратному развитию и замеща­
ется рубцом из соединительной ткани. При состоявш ейся беременности
желтое тело разрастается и активно функционирует в течение первой
половины беременности. Совокупность этих процессов составляет
содержание о в а р и а л ь н о г о ц и к л а . Их последовательность
регулируется гормонами. Три гонадотропных, т. е. действующих на
функцию половых желез, гормона образуется в гипофизе. И з них
фолликулостимулирующий гормон (Ф С Г ) вы зы вает рост фолликулов,
лютеинизирующий гормон (Л Г ), совместно с Ф С Г вызывает созревание
фолликула, овуляцию и образование желтого тела. Лю теотропный
гормон (Л Т Г ) способствует поддержанию желтого тела в активном
состояний. П о принципу обратной связи на гипофиз действую т два
гормона яичника — эстроген (синтезируется клетками растущих фолли­
кулов) и прогестерон “(вырабатывается клетками желтого тела).
Эстроген стимулирует секрецию гипофизом Л Г и понижает секрецию
Ф С Г , подготавливая овуляцию и образование желтого тела. Когда
желтое тело подвергается обратному развитию , уменьшается образова­
ние прогестерона, что стимулирует синтез гипофизом Ф С Г , рост
очередного фолликула, образование повышенных количеств эстрогена.
Таким образом закономерная смена действия гормонов обусловливает
правильный ход овариального цикла. В случае беременности, когда
желтое тело сохраняется, роста очередного фолликула не происходит, а
регуляторные функции переходят от гормонов яичника к гормонам
плаценты. Секреция гипофизом Ф С Г и Л Г регулируется активностью
нервных клеток гипоталамической области головного мозга, которая
зависит от концентрации в организме гормонов яичника.
К изменениям, наблюдаемым в оогенезе и связанным с обеспечени­
ем ранних этапов развития зародыша, относятся амплификация генов,
интенсивное образование р Р Н К и и Р Н К «впрок» для использования в
биосинтезе белков на стадиях зиготы, дробления, бластулы , ранней
гаструлы, а также создание запаса питательного материала желтка.
А м п л и ф и к а ц и я — это образование многочисленных копий
генов, кодирующих нуклеотидные последовательности р Р Н К . В ре­
зультате амплификации названных генов количество Д Н К в ооците
может увеличиваться более чем в 100 раз по сравнению с гаплоидным
количеством. Н а этих копиях, которые из ядра выходят в цитоплазму,
осущ ествляется синтез р Р Н К . Множественное копирование соответ­
ствующих генов позволяет в короткие сроки осущ ествить синтез
достаточного для обеспечения ранних стадий эмбриогенеза количества
р Р Н К . В ооцитах шпорцевой лягуш ки, например, благодаря амплифи­
кации указанные сроки сокращ аются до 3—6 месяцев. Если бы число
генов сохранилось таким, как и в соматических клетках, процесс синтеза
р Р Н К растянулся бы на 500 лет.
В оогенезе с и н т е з и р у ю т с я « в п р о к » некоторые и РН К .
Это происходит на хромосомах, приобретающих вид «ламповых
щ еток», о которых уже упоминалось (см . 4.4). Примерно % и Р Н К ,
образованной таким образом, используется в качестве матриц при
синтезе белков не ооцитом, а клетками развивающегося зародыша.
156
Амплификация генов р Р Н К , накопление р Р Н К и и Р Н К для
биосинтеза белков в теле зародыша происходят в профазе первого
деления мейоза вплоть до диплотены. Н а этом отрезке оогенеза
наблюдается относительно небольшое увеличение размеров яйцеклетки,
благодаря чему он получил название периода малого роста.
х/ Важнейшим событием предэмбрионального периода является
образование
и
накопление
в
цитоплазме
яйцеклетки
питательных
веществ,
расходуемых
после оплодотворения, — желтка, гликогена, жира. Ж елток — это
гранулы или различной формы пластинки кристаллизованных белковых
веществ. Х о тя точное место образования желтка в клетке окончательно
не установлено, отложение его в ооците, по-видимому, связано с
мембранами цитоплазматической сети и(или) пластинчатого комплек­
са, а такж е митохондриями.
Отложение питательных веществ сопровождается увеличением
размеров яйцеклетки (период большого роста). Э то приводит к умень­
шению показателя ядерно-цитоплазматического отношения. П о за­
вершении периодов роста размеры яйцеклеток увеличиваются, напри­
мер, у дрозофилы в 90 ООО, у лягуш ки в 64 ООО, у мыш и в 40 раз. Про­
должительность роста также варьирует. У плодовой мухи он занимает
примерно 3 сут, у человека — около 30 лет.
6.3.1. Морфофункциональная организация
зрелых половых клеток
Ооцит представляет собой клетку, специализированную к выполне­
нию функции размножения (см . 4.3), что проявляется в особенностях
его строения, которые формируются в предэмбриональном периоде.
Овулировавшее яйцо по морфологическим показателям сходно с
другими клетками. Н а первый взгляд в нем не обнаруживается ничего,
что предопределяло бы сложное строение будущего организма.
В процессе развития яйцеклетки, однако, проявляю тся некоторые
специфические черты организации (рис. 77). Так, выделяется поверхно­
стный сократимый к о р т и к а л ь н ы й с л о й цитоплазмы, кото­
рый располагается непосредственно под клеточной мембраной и имеет
толщ ину 2— 3 мкм. У больш инства видов животных в кортикальном
слое содержатся гранулы полисахаридной природы диаметром от долей
микрометра до 2 мкм, а также некоторое количество пигмента.
С разрыва кортикальных гранул начинаю тся, по-видимому, изменения
цитоплазмы яйцеклетки, активируемые контактом со сперматозоидом
в ходе оплодотворения. Изнутри кортикальный слой четко отграничен
от остальной цитоплазмы, снаружи он связан с плазматической
мембраной клетки.
Кортикальный слой играет важную формообразовательную роль на
начальных стадиях эмбриогенеза. Его изменения непосредственно вслед
за оплодотворением защ ищ ают яйцо от проникновения сверхчисленных
сперматозоидов. Кортикальный слой участвует в закономерном
распределении веществ в цитоплазме яйцеклетки в ходе ооплазматической сегрегации (см. ниже). Его структурой определяется положение
157
Рис. 77. Организация
зрелого
яйца.
I — ядр о; 2
ц и то п л азм а; 3
ко р ти к ал ьн ы й
слой; 4
- ц и т о п л а зм а ти че ск а я м ем б р а н а ; 5
б л е с т я щ а я зо н а; 6 — ф о л л и к у л яр н ы е 'клетки .
Рис. 78. Дифференцировка цитоплазмы
яйца (карты презумптивных органов).
а -— низш ие хордовые (а с ц и д н я ); б — рыбы,
в — ам ф ибии ; г — рептилии и пти ц ы ; I — э к т о ­
дерма; 2 — н ер вная п л а сти н ка ; 3 — хорда; 4 к и ш е ч н а я энтод ерм а; 5 — м езодерма.
сагиттальной плоскости и полярность яйца — главные факторы прос­
транственной упорядоченности развития. Механизмы формообразова­
тельных влияний кортикального слоя не выяснены.
В оогенезе у больш инства животных в связи с процессом отложения
желтка вы является п о л я р н о с т ь я й ц е к л е т о к . Противопо­
ложные полюсы называю т вегетативным и анимальным. Воображае­
мая линия, соединяющая эти полю сы, представляет главную ось яйца.
Она ориентирует в пространстве основные изменения, происходящие в
развитии. Анимально-вегетативная поляризация характеризует яйце­
клетки всех животных, но у разных представителей выражена в
неодинаковой степени.
Полярность яйца проявляется в эксцентричном расположении ядра,
которое смещается в область анимального полюса, а такж е в особенно­
стях локализации цитоплазматических включений. Так, во многих яйцах
количество желтка заметно возрастает от анимального полюса к
вегетативному. Иногда полюсы существенно различаю тся по пигмента­
ции. Например, у амфибий анимальная часть яйца интенсивно
окрашена, а вегетативная — почти лишена пигмента.
Яйца некоторых видов животных еще до оплодотворения
приобретают билатеральную симметрию. У млекопитающих она
заметна благодаря асимметричному распределению в цитоплазме
яйцеклеток жира. Э та симметрия, однако, еще лабильна и может бы ть
после оплодотворения перестроена.
158
Рис. 79. Развитие и строение сперматозоидов человека.
а — р азви ти е в стен ке к а н а л ьц а сем енника: I — сперматогонии; 2 — сперм ато ц нты 1-го п о р яд ка; 3 — сп ер м атоц н ты 2-го п о р яд к а; 4 — спер м атнд ы ;
5
сп е р м атозо и д ы ; 6 — п о д д е р ж и ваю щ ая к ле тка (С е р т о л и ); 7
- соедини­
т е л ь н а я т к а н ь ; б — зр елы й сп ер м атозо ид : 1 — го л о вк а; 2 — ш е й ка ; 3 — ср ед­
ний отдел; 4 — хвост.
Специфическая организация яйца проявляется и в особенностях
химизма разных участков цитоплазмы. Благодаря этом у уже на уровне
яйцеклетки можно представить карту презумптивных зачатков частей
будущего организма (рис. 78). У различных видов с разной степенью
выраженности в процессе созревания яйца непосредственно перед
оплодотворением происходят перемещения цитоплазмы. Э то т процесс,
называемый о о п л а з м а т и ч е с к о й с е г р е г а ц и е й , приво- /\
дат к том у, что в разных участках яйца состав цитоплазмы становится К У
различным. Так наблюдается перемещение и увеличение концентрации
гликогена и Р Н К на анимальном полюсе. Аскорбиновая кислота,
локализация которой в яйцеклетках коррелирует с размещением
пластинчатого комплекса, концентрируется главным образом по
экватору.
Закономерное смещение и избирательное распределение ци­
топлазмы, начинаясь еще в неоплодотворенном яйце, продолжается, а
нередко становится особенно заметным после проникновения в яйцек­
летку сперматозоида. Э то т процесс в форме неслучайного перераспре159
деления цитоплазмы между бластомерами происходит и далее на
ранних стадиях эмбриогенеза.
Кроме плазмолеммы, большинство яйцеклеток имеет одну или
несколько дополнительных оболочек. «Голы е» яйцеклетки описаны
только у губок и кишечнополостных. В зависимости от происхождения
различают первичные, вторичные и третичные оболочки.
П е р в и ч н ы е о б о л о ч к и особенно хорошо выражены у
позвоночных. Они возникают в результате выделения ооцитом и,
возможно, фолликулярными клетками веществ, которые образуют
слой,контактирующ ий непосредственно с плазматической мембраной
яйцеклетки. Электронно-микроскопические данные свидетельствую т об
общности строения и процесса формирования первичной оболочки яиц
различных позвоночных. П о мере образования эта оболочка прониэы
вается отростками фолликулярных клеток. Н а поверхности ооцита, в
свою очередь, возникает большое количестзо микроворсинок, вдаю­
щихся во внутреннюю часть первичной оболочки. Образование
отростков фолликулярными клетками и микроворсинок ооцитом
служит приспособлением, увеличивающим транспорт в ооцит пита
тельных веществ. Зрелый ооцит, выходяший из яичника при овуляции,
сохраняет свою первичную оболочку.
Вторичные
оболочки
имеются не у всех яиц. Они
распространены у насекомых и образуются выделениями фоллику­
лярных клеток яичника. Во вторичных оболочках яиц многих насекомых
обнаруживается м и к р о п и л е — канал для проникновения внутрь
яйцеклетки сперматозоида.
Третичные
оболочки
разнообразны по строению,
химическому составу и физиологическому значению. Они образуются
при прохождении яйца по яйцеводу 'за счет секреторной деятельности
специальных желез и могут иметь сложное строение. Так, у птиц они
представлены белковой подскорлуповой пергаментной оболочкой и
скорлупой. У акуловых рыб третичная рболочка имеет роговой
компонент и выполняет роль «лю льки» для зародыша.
Строение вторичных и третичных яйцевых оболочек животных
разных видов соответствует тем условиям, в которых происходит
развитие зародыша. Так, оболочки яиц многих рыб и амфибий, у
которых развитие происходит в водной среде, имеют студенистую
консистенцию, яйца рептилий и птиц одеты кожистыми и скорлуповыми оболочками. В силу внутриутробного развития зародышей яйца
млекопитающих имеют первичную оболочку, которая получила
название б л е с т я щ е й — гопа реИисЙа.
Яйцеклетка и сперматозоид по своим структурным показателям
являю тся высокоспециализированнымй клетками. Черты специализа­
ции мужской половой клетки, как и женской, приобретаются в ходе
гаметогенеза.
Развитие сперматозоидов происходит в извитых канальцах
семенника. Стенки этих канальцев состоят из соединительнотканной
основы и слоя фолликулярных (сертолиевых) клеток. Последние
образуют углубления, в которых слоями размещены мужские половые
клетки на различных стадиях сперматогенеза (рис. 79, а).
160
В связи с тем что сперматогенез начинается неодновременно по
длине канальца и в разных канальцах, то он распространяется как бы
волнообразно. Последовательные стадии сперматогенеза образуют
с п е р м а т о г е н н ы й цикл.
З р е л ы й с п е р м а т о з о и д состоит из четырех главных
частей — головки, шейки, средней части, хвоста (рис. 79, б).
Головка представлена почти исключительно ядром, которое
окружено лиш ь тонким слоем цитоплазмы. В последней обнаружива­
ется так называемая а к р о с о м а , которая образуется в связи с
пластинчатым комплексом сперматозоида и состоит из компактной
массы над ядром и мембраны. Ей принадлежит большая роль в
обеспечении проникновения сперматозоида в яйцо при оплодотворении.
В шейке сперматозоида расположены две центриоли — прокси­
мальная и дистальная, занимающие положение соответственно ближе и
дальше от ядра. Дистальная центриолъ участвует в образовании осевой
нити хвоста, а проксимальная — веретена деления оплодотворенного
яйца.
Средняя часть сперматозоида почти полностью занята митохондри­
ями. Последние расположены в виде изолированных компактных
скоплений или окружаю т дистальную цекгриоль и начало осевой нити
хвоста. Они обеспечивают энергией двигательную активность мужских
половых клеток.
Х вост или жгутик служит органом движения. В основе его лежит
осевая нить, окруженная небольшим количеством цитоплазмы и плаз­
матической мембраной'. У млекопитающих осевая нить окружена
дополнительными, более тонкими нитями, а у некоторых видов
животных — ундулируюшей мембраной.
6.4. Э М Б Р И О Н А Л Ь Н О Е Р А З В И Т И Е
6.4.1. Оплодотворение
О п л о д о т в о р е н и е заклю чается в соединении сперматозои­
да с яйцом. С него начинается эмбриональный период индивидуального
развития. Важнейшими этапами процесса оплодотворения являю тся:
1) проникновение сперматозоида в яйцо; 2) активация в яйце разно-,
,образных синтетических процессов; 3) слияние ядер яйца и сперматозо1 ида с восстановлением диплоидного набора хромосом.
Д ля тогочтобы оплодотворение произошло, необходимо сближение
женских и мужских половых клеток. Оно достигается благодаря
о с е м е н е н и ю сперматозоидами самца яйцеклеток самки. При­
способления, обеспечивающие осеменение, разнообразны у разных
видов животных. У большинства водных животных осеменение на­
ружное, т. е. половые клетки выделяю тся особями обоего пола во
внешнюю среду. При внутреннем осеменении при помощи специальных
копулятивных органов сперматозоиды вводятся в половые пути самки.
Необходимым условием успешного осеменения как наружного, так и
внутреннего служит согласованное поведение самки и самца в брачный
период.
Продолжительность жизни и способность к оплодотворению у яиц и
6— 56
161
сперматозоидов ограничены. Яйца больш инства беспозвоночных и вод­
ных позвоночных должны бы ть оплодотворены почти сразу же после
овуляции. У больш инства млекопитающих яйцо сохраняет способность
к оплодотворению на протяжении 24 ч, а у человека 12—24 ч после
овуляции.
Сперматозоиды сохраняют оплодотворяющую способность в поло­
вых путях самки также в течение нескольких часов. Однако у таких
животных, как пчелы, летучие мыши мужские гаметы в половых путях
самок остаю тся жизнеспособными и активными на протяжении года и
более.
Продолжительность жизни сперматозоидов и сохранение ими
оплодотворяющей способности зависит от ряда внешних факторов
(температура, освещенность, концентрация углекислого газа, водо­
родных ионов в среде, в которой находятся гам еты).
Оплодотворение происходит лиш ь при определенной концентрации
сперматозоидов в семенной жидкости. Так, обычно в 1 мл эйякулята
мужчины содержится около 350 млн. сперматозоидов. Если количество
их падает ниже 60 млн. в 1 мл, оплодотворяющая способность спермы
резко снижается.
В практике животноводства и птицеводства большое значение имеет
разработка способов консервации спермы элитных производителей с
сохранением сперматозоидами оплодотворяющей способности. Для
целей искусственного осеменения сперму таких производителей собира­
ю т в особый резервуар и медленно охлаждают примерно до 4 ,5 °С .
В таком виде она может бы ть использована для оплодотворения в
течение недели. При некоторых условиях (добавление глицерина)
сперматозоиды переносят быстрое замораживание до температу­
ры —79— 196°С с сохранением оплодотворяющей способности до
6 лет. Сперма, используемая для искусственного осеменения, как
правило, разводится в несколько раз, что позволяет осеменить семенной
жидкостью элитного самца большее, чем в естественных условиях число
самок. С помощью этого приема в течение редуктивного сезона от
одного племенного быка или барана удается получить до 2500 по­
томков.
В условиях осеменения непосредственный контакт сперматозоида и
яйцеклетки, с которого начинается процесс оплодотворения, обеспечи­
вается, с одной стороны, избыток мужских гамет, а с другой — специ­
альными механизмами дистантного взаимодействия половых клеток.
Долгое время была распространена точка зрения, согласно которой
в основе движения сперматозоида к
яйцу лежит
явление
х е м о т а к с и с а ? Направленное перемещение мужской гаметы по
градиенту концентрации определенного вещества обнаружено лишь в
исключительных случаях. Так, у некоторых папоротникообразных
сперматозоиды движутся по градиенту концентрации яблочной кис­
лоты.
Правилом, однако, служит выделение яйцеклеткой и сперматозои­
дом особых вешеств, регулирующих двигательную активность мужских
гамет. Э ти вещества названы соответственно г и н о г а м о н а м и и
а н д р о г а м о н а м и . Г иногамоны I , представляющие собой низ-
Рис. 80. Последовательные стадии взаимодействия сперматозоида и яйца прн оплодо­
творении.
а — приближение сперматозоида к яйцу; б, а
соединение мембран половых клеток и образование
бугорка оплодотворения; г — перемещение ядра сперматозоида в цитоплазму яйца.
комолекуля^ные небелковые соединения, повышаю т вероятность контакта гамет, стимулируя подвижность сперматозоидов. А ндрогамоны I имеющие сходную химическую природу, являю тся антагонистами
гиногамонов I . В отличие от последних гиногамоны II (фертилизины,
изоагглютинины), будучи по химической структуре белками или
гликопротеидами и обладая видовой специфичностью, блокирую т
двигательную активность мужских гамет своего вида путем их
склеивания в процессе иммунной реакции агглютинации в результате
соединения с белками-андрогамонами II (антифертилизинами), встро­
енными в оболочку сперматозоида. Биологический смысл дистантных
взаимодействий, модифицирующих подвижность сперматозоидов, зак­
лю чается в повышении вероятности их контакта с яйцом, а взаимодей­
ствий, ограничивающих подвижность мужских гамет, — в предохране­
нии яйца от проникновения избы тка сперматозоидов. Возможно, что
реакцией гамет на основе взаимодействия гино- и андрогармонов II
достигается прикрепление оплодотворяющего сперматозоида к по­
верхности яйцеклетки. Непосредственный контакт сперматозоида и
яйца вы зы вает в обеих клетках закономерные изменения, обусловлива­
ющие проникновение ядра и центриоли мужской гаметы внутрь
яйцеклетки и активацию метаболизма последней. Указанные изменения
со стороны сперматозоида получили название акросомной реакции, а со
стороны яйцеклетки — кортикальной реакции. Последовательные ста­
дии контактных взаимодействий яйца и сперматозоида при оплодотво­
рении показаны на рис. 80.
У млекопитающих и человека оплодотворяющий сперматозоид
преодолевает несколько рядов фолликулярных клеток, примыкающих к
яйцу, блестящ ую оболочку и плазматическую мембрану.
Проникновению сперматозоида в яйцо способствует фермент
гиалуронидаза и другие биологически активные соединения (спермолизи6*
163
ны), которые повыш аю т проницаемость основного межклеточного
вещества, разрыхляя его. Ферменты выделяю тся акросомой в процессе
а к р о с о м н о й р е а к ц и и . Сущ ность ее заклю чается в следую­
щем. В момент контакта с яйцеклеткой на вершине головки спермия
плазматическая мембрана и прилежащая к ней часть мембраны
акросомного пузырька растворяю тся. Благодаря выделению из акро­
сомной гранулы ферментов происходит растворение прилежащего
участка яйцевой оболочки. Акросомная мембрана выпячивается наружу
и образует вырост в виде полой трубочки. Последняя удлиняется,
проходит через яйцевые оболочки и вступает в контакт с плазматиче­
ской мембраной яйца. В области такого контакта возникает выпячива­
ние или бугорок оплодотворения, вслед за чем плазматические
мембраны обеих гамет сливаю тся и начинается объединение их
содержимого. С этого момента сперматозоид и яйцо представляют
собой единую клетку — зиготу. Акросомная реакция протекает исклю­
чительно быстро. У морских звезд, например, она начинается спустя
всего 1 с от момента контакта гамет, а образование акросомного
выроста происходит в основном в первые 10 с и полностью завершается
за 60 с.
При общем сходстве акросомной реакции у различных видов
животных она имеет свои особенности. Так, у кольчатых червей
Образуется не один, а несколько выростов акросомной мембраны.
У многих беспозвоночных и рыб навстречу яйцеклетке выбрасывается
акросомная нить. В таких случаях акросома хорошо развита. У многих
насекомых, осетровых и костистых рыб контактные взаимодействия
гамет происходят при наличии плотных яйцевых оболочек и спермий
достигает поверхности яйца в особых участках по специальному каналу
(микропиле), не встречая особых препятствий. При этом акросома
утрачивает свое значение и редуцируется.
Акросомная реакция у млекопитающих отличается от таковой у
беспозвоночных и низших позвоночных тем , что она протекает без
образования акросомного выроста. После контакта с яйцом плазмати­
ческая и акросомная мембраны на вершине головки сперматозоида
слипаются во многих местах. В точках слипания возникаю т небольшие
отверстия, через которые в окружающ ую среду выделяю тся заклю ­
ченные в акросоме ферменты. Последние разрыхляю т зоны контактов
между фолликулярными клетками яйценосного бугорка и материал
блестящий оболочки, в результате чего сперматозоид получает доступ к
собственной оболочке яйца. Оплодотворяющая способность спермато­
зоидов млекопитающих повыш ается благодаря воздействию на них
среды женских половых путей. О птимальная продолжительность
воздействия различна для разных видов и составляет примерно 1,5 ч для
овцы, 3 ч для крысы. 5 ч для кролика, около 7 ч для человека.
Активация яйца или к о р т и к а л ь н а я р е а к ц и я , развиваю­
щаяся в результате контакта со сперматозоидом, имеет морфологиче­
ские и биохимические проявления.
Видимым при малых увеличениях микроскопа проявлением
активации, которое часто используется как ее критерий, служат
изменения поверхностного кортикального слоя ооилазмы и образование
164
оболочки оплодотворения. Э та оболочка, называемая также желточ­
ной, возникает путем отслаивания от поверхности яйца. Под нею
формируется перивителлиновое (околожелточное) пространство, в ко­
торое изливается содержимое гранул кортикального слоя цитоплазмы
яйцеклетки. Оболочка оплодотворения защищает яйцо от проникнове­
ния сверхчисленных спермиев, т. е. обеспечивает «блок полиспермии».
Ж идкость, скапливающаяся в перивителлиновом пространстве, служит
специфичной средой, в которой протекает развитие зародыша до того
момента, когда он покидает яйцевые оболочки.
Биохимически активация яйца сводится к усилению его метаболиче­
ской активности. Отмечается существенное увеличение потребления
кислорода, начинается интенсивный синтез белка.
Обычно сперматозоид втягивается в ооплазму целиком, но у
некоторых видов хвостовой отдел остается снаружи и отбрасывается.
Дальнейшая судьба отдельных частей сперматозоида в цитоплазме яйца
различна — одни органически вклю чаю тся в развитие оплодотворенно­
го яйца, другие дегенерируют. Так, дистальная центриоль, лежащ ая в
основании ж гутика, дегенерирует, тогда как проксимальная участвует в
формировании митотического веретена первого деления дробления.
Однако в случае партеногенетического развития яиц также образуется
веретено деления, причем нормального строения. Э то говорит о том,
что центриоли веретена дробления могут возникать независимо от
центриолей спермия. Митохондрии средней части спермия набухают и
распадаются.
Ядро сперматозоида, продвигаясь к месту встречи с женским ядром,
преобразуется в м у ж с к о й п р о н у к л е у с . Его хроматин, ранее
плотно упакованный, разрыхляется, ядерная оболочка растворяется.
Преобразования и перемещения мужского пронуклеуса, как правило,
происходят одновременно с завершением второго деления созревания
яйца, за исключением тех случаев, когда к моменту проникновения
сперматозоида женский п р о н у к л е у с уже сформирован. Если же
сперматозоид входит в яйцо на ранних стадиях мейоза, то он не изменя­
ется вплоть до наступления второго деления созревания. По-видимому,
именно на этой стадии оогенеза в цитоплазме яйцеклетки появляется
фактор, вызывающ ий преобразование ядра спермия. У морских ежей и
некоторых червей наблюдается объединение непосредственно пронукле­
усов с образованием ядра зиготы — с и н к а р и о н а , чаще, однако,
объединение отцовского и материнского хромосомных наборов про­
исходит минуя фазу общего ядра, путем образования единой
экваториальной (метафазной) пластинки первого деления зиготы.
Закономерное распределение цитоплазмы, отличающ ейся по
определенным показателям в объеме ооцита, намечается у представите­
лей некоторых видов еще до оплодотворения (см. рис. 78). У большин­
ства видов животных проникновение сперматозоида в яйцо резко
усиливает процесс ооплазматической сегрегации. Последний заключа­
ется в перемещении, отмешивании различных составных частей
ооплазмы. В результате намечаются основные черты пространствен­
ной организации эмбриона, происходит пространственная «разметка»
дальнейшего развития — делений дробления, перемещения клеточного
165
материала в ходе гаструляции. Наглядна ооплазматическая сегрегация у
амфибий. Сперматозоид, перемещаясь внутрь яйца, увлекает часть.,
гранул кортикального слоя, что вы зы вает перемещение в область
вхождения мужской гаметы гранул из других участков. В результате
против места проникновения сперматозоида, в плоскости его вхождения
в яйцо, в которой указанный отток более выражен, на границе вегета­
тивного и анимального полушарий образуется участок разряжения
серого оттенка, имеющий серповидную форму — с е р ы й
серп.
Значение последнего состоит в том , что он намечает плоскость
двусторонней симметрии. М атериал серого серпа в дальнейшем
используется для построения дорсальной губы бластопора — «первичД
ного организатора» формирования комплекса осевых органов зароды-/
ша (см . 6.5.1).
Х о тя для оплодотворения яйцеклетки необходим один сперматозо­
ид ( м о н о с п е р м й я),
у некоторых видов птиц, рептилий,
акуловых рыб в цитоплазму яйца проникает одновременно несколько
мужских гамет ( п о л и с п е р м и я ) . Ядра сверхчисленных спермато­
зоидов могут некоторое время делиться, однако, в конце концов,
неизбежно гибнут.
6.4.2. Дробление. Бластула
С т а д и я д р о б л е н и я эмбриогенеза заклю чается в повторяющихся делениях клеток зародыша. Превращения ядерного материала в
делениях дробления полностью соответствую т тому, что наблюдается
в ходе обычного митоза — за удвоением количества Д Н К следуез ее
точное распределение между дочерними клетками. Отличительной
чертой делений дробления служит отсутствие роста новообразующихся
клеток перед очередным делением. Благодаря этому объем тела
зародыша не изменяется.
О тсутствие клеточного роста между делениями дробления
выражается в особенностях жизненного цикла клеток развивающегося
зародыша на рассматриваемой стадии. И з него фактически полностью
выпадает постмитотический (О ,) период интенфазы, а период предмитотического синтеза Д Н К (8) начинается в телофазе предшествующего
митотического цикла. Клетки, образующиеся в процессе дробления,
называю тся б л а с т о м е р а м и .
Биологическое значение изменений на стадии дробления сводится к
следующему. Благодаря повторяющ имся циклам редупликации и разде­
ления Д Н К происходит размножение (редупликация) генотипа зиготы. В
результате цитотомии, сопровождающей деление ядер, зародыш из
одноклеточного превращается в многоклеточный. Так как деления
дробления не сопровождаются ростом бластомеров, они способствую т
повышению величины ядерно-цитоплазматического отношения, сни­
женного в период большого роста ооцита, до значений, типичных для
соматических клеток. М ногоклеточность, возникающ ая в ходе дробле­
ния, сопровождается образованием и прогрессивным расширением
площади межклеточных контактов. Э то обстоятельство представля­
ется важным в связи с большим значением контактного взаимодействия
166
|
/щ
Ти п я и ц
V
клеток в процессе эмбриогенеЛ ан ц етн ик
Л ягуш ка
П тица
•за. Одна из особенностей цито­
томии в делениях дробления
заключается в том, что на­
правления борозд дробления
зависят от неоднородности
внутренней организации от­
дельных участков цитоплаз­
Д робление
мы. Э то приводит к тому, что
на этой стадии усиливается
процесс ооплазматической се­
грегации. К концу стадии
дробления
устанавливается
определенная мозаика бласто­
меров, отличающихся в той
или иной мере по структуре
цитоплазмы и направлениям
дальнейшего развития.
Разным группам живот­
ных свойствен свой режим
дробления, который зависит
от количества и распределения
в яйцеклетках желтка (рис. 81),
Бластула
формы и перемещений бласто­
меров.
( У ланцетника и амфибий
при умеренном количестве
желтка в яйце борозды дроб­
ления разделяют его полно­
стью . Такое дробление называетСЯ П О Л Н Ы М ИЛИ Г 0_-___ Рис. 81. Типы яиц и соответствующие им типы
л о б л а с т и ч е с к и м . В ре- дробления.
Зультате при достаточно рав­
номерном распределении желтка в цитоплазме (гомолецитальные
яйца ланцетника) образуются бластомеры равной величины. При
неравномерном распределении желтка (телолецитальные яйца амфи­
бий) бластомеры оказываю тся разного размера. Соответственно этому)
показателю выделяют дробление р а в н о м е р н о е и н е р а в н о ­
м е р н о е . Мелкие бдастомеры~втннкакЗТ в ооласти анимального
полюса [ м и к р о м е р ы ), крупные — в области вегетативного
полюса зародыша ( м а к р~о~м е р ы).,
У животных, яйца которых содержат большое количество желтка,
последний при дроблении не делится. Такое дробление называется
н е п о л н ы м или м е р о б л а с т и ч е с к и м. В зависимости от
рйЯбУюженйя экёлтка дробление может оы ть д и с к о и д а л ь н ы м
и п о в е р х н о с т н ы м . В перром случае желток концентрируется
на вегетативном полюсе и составляет значительную часть яйца, а в
образовании клеток зародыша принимает участие лишь цитоплазма
анимального полюса яйцеклетки (костистые рыбы, пресмыкающиеся,
167
птицы). Во втором случае желток расположен в виде пояса внутри яйца
(центролецитальные яйца насекомых и некоторых других беспозво­
ночных). При этом первые деления ядра, находящегося первоначально в
центре яйца, который свободен от желтка, проходят без разделения
цитоплазмы. Затем участки центральной цитоплазмы с ядрами
перемещаются к поверхности, где сливаю тся со слоем периферической
цитоплазмы, который после этого делится на клеточные территории,
соответствующ ие отдельным бластомерам. Процесс деления на массу
желтка не распространяется.
Влияние распределения желтка на характер дробления подтвержда­
ется экспериментально. Так, при центрифугировании оплодотворенных
яиц лягушки желток смещается к вегетативному полюсу, что изменяет
тип дробления с полного неравномерного на дискоидальное.
Влияние распределения желтка на тип дробления опосредуется
зависимостью от этого показателя локализации в клетке ядра и
ориентации веретена деления. Э та зависимость описывается правилами
Сакса—Гертвига, согласно которым: 1) ядро клетки стремится занять
место в центре свободной от желтка области цитоплазмы; 2) длинная
ось веретена деления ориентируется по направлению наибольшей
протяженности свободной от желтка цитоплазмы.
Н а различиях в характере взаиморасположения бластомеров
основана еще одна классификация типов дробления. Р а д и а л ь н о е
дробление наблюдается у губок, иглокожих, хордовых. В этом Случае
плоскости цитотомии клеток в последующих делениях проходят
перпендикулярно друг другу. В итоге бластомеры располагаются друг
над другом широтными рядами симметрично относительно любой
плоскости зародыша, которую можно провести через анимально-.
вегетативную ось.
Среди моллюсков, ресничных и кольчатых червей распространено
спиральное
дробление, при котором в последовательных
делениях происходит закономерное по направлению смещение новообразующихся клеток относительно анимально-вегетативной оси яйца.
Плоскости клеточных делений в этом случае проходят под углом к
названной оси, а также к экватору. При этом бластомеры располагают­
ся не правильными широтными рядами, а более или менее чередуются
друг с другом. Спиральное дробление на определенно)! стадии развития'
переходит в б и л а т е р а л ь н о е дробление, которое в качестве
самостоятельного встречается у некоторых червей, например аскариды.
Первая борозда дробления в этом случае проходит не меридионально, а
в плоскости экватора яйца. Спиральный и билатеральный типы
дробления обусловливаются закономерными поворотами бластомеров
соответственно в анафазе или интерфазе клеточных циклов.
У низших беспозвоночных, в частности паразитических плоских
червей, наблюдается а н а р х и ч е с к о е
дробление, когда слабо
связанные друг с другом бластомеры формируют неправильной формы
цепочки.
Спиральное и билатеральное дробление называю т д е т е р ­
минированным.
Эти м
подчеркивается ранняя жесткая
детерминация (предопределение) дальнейшего развития бластомеров
168
уже после первых делений. Радиальное дробление называю т
р е г у л я ц и о н н ы м . П уть развития бластомеров в этом случае
определяется однозначно на более поздних стадиях эмбриогенеза.
Яйцам каждого вида животных свойствен определенный ритм
дробления. Так, у золотой рыбки (Кхсех аи гаш т) дробление происходит
регулярно каждые 20 мин. У лягушки первые дробления совершаются
приблизительно каждые 60 мин. Дробление у млекопитающих осуще­
ствляется медленными темпами — интервал между смежными делени­
ями нередко достигает 10 и более часов. По мере дробления меняются
временные показатели отдельных фаз клеточного цикла бластомеров.
В качестве общей тенденции отмечается удлинение продолжительности
клеточного цикла.
^
Деление бластомеров бывает с и н х р о н н ы м
и -н-е-син­
х р о н н ы м . У большинства видов оно происходит несинхронно с
самого начала развития, у других становится таковым уже
после первых делений. Если бы режим делений, характерный для начала
стадии дробления сохранялся достаточно долго, то уже через 20 ч
эмбрионального развития зародыш лягуш ки, например, состоял бы из
1 млн. клеток. Через 2 сут от начала дробления он имел бы
25 х 1013 клеток, а через 3 сут зародыш достиг бы размеров
5,2x10м м 3.
1 в^^Яробление у представителей разных групп животных имеет свои
особенности, однако оно завершается образованием близкой по
сто ен и ю структуры — б л а с т у л ы .
/ Бластула состоит из слоя клеток — б л а с т о д е р м ы ,
ограничиваюшего полость — б л а с т о ц е л ь .
Бластула формиру­
ется, начиная с ранних этапов дробления благодаря расхождению
бластомеров. Возникающ ая при этом полость заполняется жидкостью.
Бластодерма представляет собой пласт клеток, в связи с чем
образование бластулы нередко обозначают как процесс эпителизации ее
стенки, благодаря которому устанавливаю тся более тесные межкле­
точные контакты и создается основа для скоординированных переме­
щений бластомеров в дальнейшем развитии.
Строение бластул у разных видов животных во многом зависит от
типа дробления. При равномерном дроблении, напримёр у ланцетника,
бластула принимает вид однослойного пузырька с большим бластоцелем.
Ее
называю т
типичной
бластулой
или
целобластулой.
В случае телолецитальных яиц (амфибии) пласт бластодермы
Ж построен из клеток разных размеров —^м елких на анимальном и
1 крупных на вегетативном полюсах. Такое строение типично для
1амфибластулы.
С
Дискоидальное дробление (птицы) приводит к формированию
71 и с к о б л а с т у л ы ,
полость которой имеет вид сплющенной
щели, подчас очень узкой, находящейся под зародышевым диском.
При поверхностном дроблении центральная часть зародыша
заполнена желтком, а бластодерма состоит из одного слоя клеток.
Бластулу такого строения называю т п е р и б л а с т у л б й .
Бластула с небольшим, центрально расположенным бластоцелем
169
называется с т е р р о б л а с т у л о й . Она встречается у моллюсков,
некоторых кишечнополостных, червей. Н а определенном этапе своего
развития к стерробластуле по строению приближается бластула
млекопитающих.
Известны кишечнополостные, у которых в бластуле отсутствует
бластоцель. Зародыш имеет вид плотного комка клеток и называется
м о р у л о й.
В процессе образования бластулы происходят важные изменения
состояния клеток зародыша. Наблюдается десинхронизация клеточных
делений, удлиняется продолжительность клеточного цикла, в послед­
нем появляется постмитотический период. У рыб и амфибий лиш ь по
достижении стадии бластулы начинается транскрипция ядерного
генома бластомеров, т. е. использование в жизнедеятельности зароды­
ша собственной наследственной информации. Э то проявляется в
активации синтеза Р Н К в ядрах. У млекопитающих заметный синтез
Р Н К наблюдается со стадии двух бластомеров.
6.4.3. Гаструляция
У представителей большинства групп животных бластула имеет вид
полого шара, поэтому ее называют однослойным зародышем. На
следующей стадии эмбриогенеза происходит расчленение клеточного
материала на два (кишечнополостные и, возможно, губки) или три
зародышевых пласта или листка. Указанный процесс называется
г а с т р у л я ц и е й , а зародыш на этой стадии г а с т р у л о й .
Первоначально образуются н а р у ж н ы й ( э к т о д е р м а ) и
в н у т р е н н и й ( э н т о д е р м а ) листки. Позже возникает третий,
с р е д н и й зародышевый листок ( м е з о д е р м а ) , располагающий­
ся в теле зародыша между эктодермой и энтодермой.
ХЗародышевые
л и с т к и — это динамичные скопления
клеток, закономерно образующиеся в эмбриогенезе многоклеточных
животных, которые характеризуются определенным пространствен­
ным взаиморасположением. Превращения клеточного материала заро­
дышевых листков на основе широких взаимодействий в рамках
целостного организма зародыша приводят в дальнейшем развитии квозникновению необходимых органов и систем органов.
Выделяю т четыре основных способа образования наружного и
внутреннего листков. Часто, однако, наблюдается комбинация несколь­
ких способов гаструляции (рис. 82).
V Достаточно
широко
в
природе
представлен
способ
и н в а г и н а ц и и , который заклю чается в том , что определенный
участок бластодермы, сохраняя структуру пласта, вворачивается
внутрь бластоцеля. Таким способом гаструляция проходит, например, у
ланцетника, для которого характерно полное, почти равномерное
дробление, дающее близкие по размерам бластомеры на анимальном и
вегетативном полюсах. Гаструляция начинается с движения внутрь
бластомеров вегетативного полюса. По мере того как они продвига­
ю тся по направлению к анимальному полюсу, бластоцель уменьшается
вплоть до полного исчезновения на завершающем этапе. В итоге
170
Рис. 82. Типы гаструл.
а — и н ваги н ац и о и и ая; 6 — э п и б о л и ч е ск а я; в
2 — эн тод ер м а; 3 — б ласто ц ел ь.
им м и гр ац и он ная;
I
делнм ииац иоииан,
|
эктодерма,
образуется двухслойный мешок, наружной стенкой которого служит
первичная эктодерма, а внутренней — первичная энтодерма. Последняя
выстилает
полость
первичной
кишки
или
гастроцель.
Отверстие, при помощи которого эта полость
сообщается с наружной средой, называется п е р в и ч н ы м р т о м
или б л а с т о п о р о м . Края бластопора образую т его губы.
Судьба бластопора в развитии у представителей разных групп
животных различна. У первичноротых он превращается в дефини­
тивный (окончательный) рот взрослого организма. У вторичноротых
он нередко дает анальное отверстие. Ро т у таких животных возникает
на переднем конце тела.
При полном неравномерном дроблении (амфибии) макромеры
вегетативного полюса практически не способны к перемещениям.
В этом случае зародыш превращается в двухслойный путем
^ э п и б о л и и — обрастания макромеров быстро делящимися микро­
мерами анимального полюса. У зародышей таких животных первона­
чально бластопор не образуется и отсутствует гастроцель. Впослед­
ствии, когда макромеры, делясь, приобретают меньшие размеры,
образуется зачаток первичной кишки.
^
Распространенным способом гаструляции у кишечнополостных
является и м м и г р а ц и я — вычисление части клеток стенки блас­
тулы внутрь бластоцеля. В эволюционном плане это, видимо, наиболее
древний способ образования эктодермы и энтодермы. Выселение клеток
происходит у разных видов животных по всей поверхности, а также на
одном или двух полюсах бластулы. Поэтом у различают мультиполярную , униполярную или биполярную иммиграцию.
У кишечнополостных с таким способом гаструляции нередко
выселение бластомеров в бластоцель происходит настолько активно,
что она фактически исчезает, будучи заполнена клетками.
V
Д е л я м и н а ц и я как способ гаструляции встречается у кишечно­
полостных, имеющих бластулу в виде морулы. В этом случае клетки,
находящиеся снаружи, преобразуются в пласт. Оставшиеся клетки,
занимающие более внутреннее положение, формируют энтодерму. При
таком способе гаструляции клеточные перемещения минимальны.
Четы ре способа гаструляции, описанные выш е, редко встречаю тся в
чистом виде. Так описано сочетание инвагинации с эпиболией
171
(амфибии), деляминации с иммиграцией (иглокожие). М о ж н о поэтому
назвать п яты й способ гаструляции — с м е ш а н н ы й .
Закладка третьего зародышевого листка происходит двумя
главными способами: телобластическим и энтероцельным.
Т е л о б л а с т и ч е с к и й способ характерен для первичноротых
ж ивотны х, особенно с вы р аж енн ы м спиральны м типом дробления.
В ы ш е отм ечалось, ч то прн это м типе дробления очень рано вы явл яется
п уть дальнейшего развития отдельных бластомеров. У моллю сков,
например, для которых типичен телобластический способ, мезодерма
образуется путем серии последовательных делений двух клеток
(телобластов или м езобластов), симметрично расположенных в поло­
сти бластоцеля в районе губ бластопора. Сам и эти клетки яв л яю тся по­
том ками единственного, то чно идентифицируемого бластомера, полу­
чившего обозначение 4с1.
Кр о м е м оллю сков, закладка мезодермы телобластическим спосо­
бом в его типичном варианте происходит у ко льчаты х червей,
некоторых ракообразных. Н апр оти в, у больш инства представителей
типа членистоногих телобластический способ представлен в затуш е­
ванном виде.
Э н т е р о ц е л ь н ы й
способ образования мезодермы типичен
для иглокожих. Будучи представлен в д остаточно ясной форме у
ланцетника, у больш инства других хордовых э то т способ обнаружива­
ется в стертой форме.
В случае энтероцельного способа клеточны й материал будущей
мезодермы первоначально входит в состав стенки первичной кишки,
т. е. участка бластодермы, впячивающ егося в бластоцель прн образова
нин наружного и внутреннего зародыш евых листков. Л и ш ь позже
мезодерма выделяется в качестве сам остоятельной закладки путем
симметричного выпячивания в полость бластоцеля определенных
участков стенки первичной киш ки с последующей их отш нуровкой.
В ходе дальнейших превращений мезодермальных закладок,
образующ ихся телобластически, в них возн и каю т полые участки,
представляющие
собой
целом
или
в т о р и ч н у ю
п о л о с т ь тела.
Целом при энгероцельном способе формиру­
ется как полость о тш нуровы ваю ш ихся мезодермальных пузырьков,
т. е. является частью бывш ей полости первичной кишки.
Первоначально третий зародыш евый л исток представляет собой
массу
о тр о стча ты х
клеток.
Эта
та к
называемая
н е д и ф ф е р е н ц и р о в а н н а я м е з о д е р м а в дальнейшем
развитии подразделяется в переднезаднем направлении на отдельные
участки — с о м и т ы . И з различных зон со м и то в на стадии гнсто- и
органогенезов образую тся определенные производные, о чем будет
сказано ниже.
В процессе гаструляции, благодаря закономерному перераспределе­
нию клеточного материала возникаю т закладки будущих органов.
Современные исследования показали, ч т о на этой стадии в зародыш е
впервые удается обн аруж и ть в достаточно больш их концентрациях
белкн, специфичные для некоторых направлений клеточной дифференцировки взрослого организма. Т ак
показано начало синтеза
гемоглобина и белка м ыш ечной ткани миоглобина.
172
Рис. 83. Г«аетр\'пяция у лягушки
414 ПМ нр1>ш*ч-и I
серпнвшния Оорогдка. б
6-|асюиора, I
бластоцель
ерпоянияая бор«
.
'
.Ор. ■ивапня г ■три-» 1И, в .
ипличь г<и.тиу ш ( | а с т р и ц с I
4
замиканш.
же.ш >I
6.4.4. Га стр у л яц и я у представителей некоторых классов
ПОЗВОНОЧНЫХ ЖИВ01НЫХ
Перечисленные в ы ш е способы закладки зародыш евых листков
о тр а ж а ю т главные направления преобразований клеточного материала
в процессе гаструляции В конкретном выражении у представителей
различных групп ж и вотн ы х названный процесс о тл ичается значи
тельн ы м разнообразием деталей. Подробное освещение этих деталей
выходит за рамки настоящего курса. Вм есте с те м для уяснения значе­
ния изменений, происходящих на стадии гаструляции, с которой
совпадает начало образования тканеспецифичных белков и которая
продолжается непосредственно в стадию гисто- и органогенезов,
полезно ознаком иться с ходом гаструляции у некоторых представите­
лей животного мира, у наиболее хорошо изученных в это м отношении
амфибий (низшие позвоночные, анамниа), птиц (вы сш ие позвоночные,
ам ниота), а та кж е высш их млекопитаю щ их, к ко то р ы м принадлежит и
человек.
Гастр уляц и я у амфибий. Э т а стадия представляет собой сложны й
комплекс клеточны х перемещений. Х о тя основны ми способами образо­
вания экто- и энтодермы у названных ж и вотн ы х сч и та ю т эпиболню и
инвагинацию, гаструляция у них начинается, по-видимому, с незначи­
тельного погружения некоторых из богатых ж ел тко м макромеров
вегетативного полю са вн утрь тела зародыш а. В связи с ограниченно­
ст ью указанных перемещений они получили название предгаструляци
онных.
Собственно гаструляция начинается в области серого серпа
(см. 6.4.1), где возникает щель, представляю щ ая собой закладку
бластопора, и приним аю щ ую позже вид серповидной бороздки
(рис. 83). В результате погружения клеток в области бороздки образу­
ется полость, которая дает начало первичной кишке. Зачаток
бластопора располагается на заднем конце зародыша. Верхняя кромка
бластопора обращена к спинной стороне, поэтом у она получила
название д о р с а л ь н о й г у б ы .
Н а это м этапе ведущую ро ль в процессе гаструляции начинает иг­
рать эпиболия. Кл ето ч н ы й материал как бы наползает, смещ аясь в
173
анимально-вегетативном направлении, и подворачивается через дор­
сальную губу бластопора в н утр ь бластоцеля. П ри это м источником
клеточного м атериала, проходящего через дорсальную губу, служ ат
бластомеры кры ш и бластоцеля в области анимального полушария,
которые интенсивно р азм н ож аю тся. В процессе гаструляции наблюда­
ется та кж е увеличение площади поверхности кры ш и полости б ластул ы
за счет вклинивания м ежду бластомерами, заним аю щ им и наружное
положение, клеток, поднимающихся из более глубоких слоев. Э т о
приводит к изменению соотношения плошадей поверхности зародыша,
занимаемых анимальны м и и вегетативным и бластомерами, в пользу
первых.
Бластопор, разрастаясь, охваты вает вегетативную зону кольцом.
Вегетати вн ы е бластом еры в просвете бластопора о бразую т так
н азы ваем ую ж е л т о ч н у ю
п р о^б к у. Н а это м этапе, кроме
дорсальной губы бластопора, вы д ел яю т та кж е в е н т р а л ь н у ю
(бр ю ш н ую ) и б о к о в ы е губы , через которые вн утр ь бластоцеля
подворачивается сравнительно малое количество клеточного м ате­
риала.
К л е тк и , сместивш иеся через бластопор внутрь, сначала о бразую т
сплош ную массу, которая оттесн яет бластоцель вентрально, суще­
ственно ум еньш ая ее размеры. Д алее в.результате процесса инвагинации
происходит распределение клеток по внутренней поверхности б ласто ­
дермы, что приводит к возникновению п е р в и ч н о й
кишки,
полость которой называется г а с т р о ц е л ь .
В следую щ ую фазу из стенки первичной киш ки выделяется
клеточный материал мезодермы и хорды, т. е. образование третьего
зародышевого листка происходит энтероцельн ы м способом.
Гастр уляц и я у птиц. Эмбриогенез высш их позвоночных (рептилии,
птицы, м лекопитаю щ ие), составляю щ их группу ам ниот, характеризу­
ется особенностями, о траж аю щ и м и освоение ими наземной среды
обитания. Рассм отри м эти особенности на примере эмбрионального
развития птиц.
К а к у всех ам ниот, у птнц клеточны й материал на ранних стади­
ях развития разделяется на две части:
з а р о д ы ш е в у ю
н
в н е з а р о д ы ш е в у ю . П ервая в дальнейшем используется для
построения тканей непосредственно организма, вторая — эмбрио­
нальных провизорных органов.
П о завершении стадии дробления зародыш птицы представлен
распластанным на ж елтке б л а с т о д и с к о м — несколькими слоя­
ми бластомеров, причем клетки наиболее вентрального слоя тесно
ко н та кти р ую т с ж елтко м . Ц ентральная зона бластодиска или
з а р о д ы ш е в ы й
ш и то к
является собственно зародышем.
С краев он окаймлен приподнятым над ж ел тко м валиком. Далее о т
центра бластодиска к периферии находится внезародышевая часть,
плотно прилегающая к ж елтку.
С та д и ю гаструляции у птнц можно разделить иа два этапа. Сначала
из клеточного материала отделяется г и п о б л а с т , ко то ры й позже
преобразуется во в н е з а р о д ы ш е в у ю
энтод ерм у.
Со­
ответственно современным представлениям э т о т э т 'п гаструляции
174
происходит, видимо, путем множественной иммиграции клеток, начиная-е-будушего заднего края зародышевого ш и тка. Р яд эмбриоло­
гов впло т ь до настоящего времени рассм атриваю т способ выделения
грпобласта, как лепяминацию.
Н а втором этапе клеточный материал бластодиска, оставш ийся
после выделения гипобласта и н азываем ый
эп и бл асто м ,
подразделяется на зачагки хорды, мезодермы, зародышевой энто­
дермы. О ставш аяся после этого часть зпибласта со ответствует эк­
тодерме.
В начале преобразования клеточного м атериала эпибласта на­
блю д аю тся сло ж ны е перемещения клеток, имеющие итогом схож­
дение их к заднему концу бластодиска, где в силу этого образуется
клеточное сгущение в виде п е р в и ч н о й п о л о с к и . Указанная
структура является Д и н а м и ч н ы ^ <Щроточным>> образованием. Кл етк и
эпибласта «вл и в аю тся» в нее спереди и с боков, о/Шако в иёй не задер­
ж и ваю тся, а перемещаются в полость между эпн- и гн поб ластом , рыхло
'распределяясь д о их поверТРгости. Благодаря различиям в интенсивно­
сти миграции клеток в первичной полоске на переднем конце образуется
углубление — п е р в и ч н а я я м э т а , а по средней линии — ж ело­
бок. Первичная п о л о ска _с_ж ел о б ко м называется
п ервичной
6 о р о з д к о й. Кпереди о т первичной ям ки возникает скопление
*■'клеток — п е р в и~ч~(Т ы й ~ у У е_л сП Г
■
Через переднюю часть первичной бороздки и первичный узелок
клеточны е потоки направляю тся преимущественно кпереди, а через
заднюю часть первичной полоски — кзади о т первичного узелка. Э т о
важное о бстоятельство, та к как различна судьба клеточного материала,
выселяющ егося из эпибласта двумя названными потокам и. Через
переднюю часть первичной бороздки и узелок последовательно
м игрирую т клетки, формирующие закладки головной и осевой (со­
м и т ы ) мезодермы, хорды. Ч а с т ь клеток, образующ их мезодерму,
перемещается через центральную область первичной бороздки, а клетки
внезародышевой мезодермы через задню ю ее часть.
В фазе завершения процесса гаструляции первичная бороздка
прогрессивно укорачивается, а первичный узелок смещ ается кзади и
впоследствии образует анальное отверстие.
Рассм атривая стадию гаструляции эмбриогенеза в сравнительном
плане, м ож но сказать, ч то второй этап гаструляции птиц гомологичен
гаструляции амфибий. П ри это м первичная бороздка соответствует
бластопору, а первичный узелок — его дорсальной губе.
В ы ш е рассмотрены изменения клеточного материала зародышевого
диска. В это время внезародыш евые порции гипо- и эпибласта
распространяются по поверхности желтка, что приводит к возникно­
вению ж е л т о ч н о г о м е ш к а . В обрастании ж ел тка уча с тв у ю т
все три зародыш евых листка, однако мезодерма, в связи с более поз­
дним ее образованием, несколько отстает. П ри это м боковая мезодерма
подразделяется на париетальный (пристеночный) и висцеральный
(внутренностны й) листки. Собственно стенка желточного меш ка
образуется внезародышевой ж елточной энтодермой и висцеральным
листком мезодермы. Эктодер м а и париетальная мезодерма примут
175
участие в формировании стенок другого провизорного органа — амнио­
на, а т а к ж е серозной оболочки (хор'иона), ограничивающ ей простран­
ство, названное э к з о ц е л о м о м . В экзоцелом врастает алланто­
ис — вы р о ст задней кишки зародыша.
Гаструляц ия у высших [плацентарных] млекопитаю щ их. Особенно­
сти эмбрионального развития этих ж и вотн ы х стан о вятся зам етн ы м и
уже на стадии дробления. В связи с алецитальностью яйцеклеток оно у
названных ж и вотн ы х полное, однако его нельзя отнести строго ни к
радиальному, ни к спиральному типу. П рактически с самого начала
процесс деления является асинхронным. В отличие о т низших позво­
ночных, уж е на стадии 2— 4 бластом еров активируется собственный
генетический аппарат. В результате дробления первоначально образу­
ется плотная структура — морула нли стерробластула. Н а ранних
стадиях (у м ы ш и на стадии 16 клеток) в такой стерробластуле
определяется группа внутренних, более тем ны х, и наружных, светлых,
бластомеров, которы е со о тветствую т зародышевой и внезародышевой
частям развиваю шегося организма. П рактически сразу ж е (у м ы ш и на
стадии 32 бластомеров) зародыш приобретает полость, заполненную
ж ид костью , и превращается в пузырек — бластоцисту. С тен ка такого
п узы рька — троф област — построена из клеток внезародышевой час­
ти; располагающаяся на одном из ее полю сов плотная группа
клеток — зародыш евый узелок (эмбрдобласт) со ответствует непосред
ственно зародыш у.
Собственно гаструляция начинается с обособления слоя клеток
эмбриобласта, обращенного в полость бластоцисгы , — энтодермы
(гипобласта). К л е тк и краевой зоны этого зачатка распространяются по
внутренней поверхности троф областа, ограничивая полость ж ел то чн о ­
го мешка, ко то ры й у плацентарных млекопитаю щ их рудиментарен.
В части зародышевого узелка, оставш ейся после отделения
гипобласта, процесс образования закладок 1 ародыш евых л истков во
многом сходен с та ко вы м у других ам ниот, например птиц. В ходе
преобразований клеточного материала зародыш евого щ и тка формиру­
ю тся первичная полоска, первичная бороздка, первичный узелок.
Определенные особенности сущ ествую т в образовании закладки
такого провизорного органа как амнион. У больш инства видов
млекопитаю щ их амниотическая полость возникает путем расхождения
клеток эмбриобласта, а не путем замы кания туловищ ны х складок,
формируемых внезародышевой эктодермой н париетальной мезо­
дермой.
Т ак же как у низших ам ниот, часть клеток, выселяясь через первич­
ную бороздку в пространство между троф областами и стенкой
желточного м еш ка, дает внезародыш евую мезодерму. Последнее,
наряду с троф областом, принадлежит больш ая роль в образовании
хориона — провизорного органа, обеспечивающего ко н та кт зародыш а
с материнским организмом и эффективное получение им питательных
веществ, кислорода. Особенность прим атов состоит в том , что
внезародышевая мезодерма у них выделяется очень рано, еше до
обособления зародышевого щ и тка в результате отделения гипо­
бласта.
176
6.4.5. О бразование тканей и органов
Процесс формирования систем органов в эмбриональном развитии
называется
о рганогенезом .
В построении любого органа
участвует несколько тканей, клеточны е и неклеточные компоненты
которых о бразую тся непосредственно в ходе доссоздания структуры
органов П оэтом у стадия органогенеза является и стадией гистогенеза.
Процесс органогенеза интенсивно изучался у позвоночных н особенно
млекопитающих, что объясняется потребностями медицины н ветери­
нарии.
В рассматриваемой стадии м ож но вы д ели ть две фазы. Первая,
называем ая н е й р у л я ц и е й ,
со стоит в образовании комплекса
осевых органов — нервной трубки, хорды. Вто р а я фаза заклю чается в
построении остальны х органов, приобретении различным и участкам и
тела типичной для них формы и черт внутренней организации,
установлении определенных пропорций. Отличительной чертой первой
"ф азы органогенеза сл уж и т то , что в морфологические перестройки,
сопровождающ ие формирование центральной нервной си стем ы , вовле­
кается почти весь зародыш. Разви ти е других органов представляет
собой пространственно ограниченные процессы.
Зародыш на стадии неируляцин называется
нейрулой.
М атериал, используемый на построение нервной систем ы у позво­
ночных ж ивотны х, представляет собой дласх^клеток эктодермы над
-зачатком хорды и сом итам и (и с Й р о э к т о д е р м а). Сначала в
этой области происходит уплошение клеточного пласта, что приводит к
образованию н е р в н о й п л а с т и н к и , края которой приподни­
м аясь, ф ормирую т н е р в н ы е
валики.
За счет перемещения
клеток по средней линии нервной пластинки возникает углубление. Н а
этом этапе нейруляцин закладка нервной систем ы называется
н ер в н ы м
ж е л о б к о м . Одновременно нервная пластника как
бы складывается по средней линии, а несколько позже ее края
см ы ка ю тся. В результате этих процессов возникает
нервная
т р у б к а с п олостью — н е в р о ц е л е м. С м ы кан и е валиков про­
исходит сначала в средней, а затем в задней части нервного желобка.
В последнюю очередь это происходит в головной части, которая
превосходит другие области по ширине. Передний, расширенный отдел
в дальнейшем развитии образует головной мозг, остальная часть
нервной трубки — спинной мозг.
Нервная трубка отделяется о т эктодермального пласта, который
над ней смыкается.
В процессе нейруляции часть клеток ие попадает в состав нервной
трубки, а образует скопление вдоль нее — г а н г л и о з н у ю
п л а с т и н к у
или н е р в н ы й
гребень.
М атериал гангли­
озной пластинки дает много производных, среди которых нервные
клетки спинномозговых и сигматических нервных узлов, клетки
мозгового вещества надпочечников, пигментные клетки эпидермиса
кожи, волос, перьев, хрящевые и костные клетки.
Образование хорды по времени со ответствует сам ы м ранним
•талам нейруляции н происходит путем обособления по средней линии
177
клеточного м атериала нз зачатка, обшего с энтодермой и мезо­
дермой — стенки первичной киш ки (см. 6.4.3).
П о бокам о т хорды располагается еще один компонент комплекса
осевых органов — мезодерма сом итов. г
^
Разви ти е отдельных органов вклю чает прогрессивное изменение их
формы, состава тканей, структурно-химические преобразования клеток,
появление и закономерную смену биохимических и физиологических
характеристик. Э л ем ен ты частной, эмбриологии, изучающей конкрет­
ные органогенезы, вклю чены в курсы анатомии, гистологии, физиоло­
гии. М ы остановим ся на рассмотрении производных, которы е образует
каждый из зародыш евых листков, и общих закономерностях органоге­
неза.
И з материала наружного зародышевого листка (эктодермы)
развиваю тся эпидермис кожи и его производные (перо, волосы, ногти,
кож н ы е и м олочные железы ), компоненты органов зрения, слуха,
обоняния, эпителий ротовой полости, эм аль зубов.
П роизводным и энтодермы яв л яю тся эпителий
желудка и ки­
шечника, клетки печени, секретирующ ие клетки поджелудочной,
киш ечных н желудочных желез. Передний отдел эмбриональной кишки
образует эпителий легких и воздухоносных путей, а та к ж е секретируюшие отделы передней и средней доли гипофиза, шитовидной и паращитовидной желез.
К началу стадии органогенезов мезодерма представлена сомитами,
занимающими положение сбоку о т хорды и переходяшими в сегмен­
т н у ю или со м и тн ую ножку, а та кж е боковой пластинкой с висце­
ральны м (спланхноплевра) и париетальны м (соматоплевра) листками.
К л ето ч н ы й материал сом итов распределяется м еж ду несколькими
зачатками. Н ар уж н ая область сом ита, прилегающая к эктодерме,
представлена д е р м о т о м о м
и образует соединительнотканный
компонент кож и (дерму). Внутренняя часть с к л е р о т о м участвует
в образовании скелетных структур, в частности тел позвонков. М еж ду
дермотомом и склеротомом располагается
м и о т о м — зачаток
поперечнополосатой м ускулатуры .
В
области сомитной ножки располагается зачаток органов
мочевыделения (н е ф р о т о м). М атериал сплахно- и сомагоплевры
используется в развитии половой (гонотом ), сердечно-сосудистой и
лимфатической систем, плевры, брю ш н н ы , перикарда.
Е щ е до подразделения зачатка мезодермы на со м и ты из него
вы селяю тся клетки, характеризующиеся наличием отростков. К ним
присоединяется та кж е некоторое количество клеток, выселяю щ ихся из
других зароды ш евых листков, в частности эктодермы. Совокупность
указанных клеток образует
мезенхиму.
И з этого зачатка
развиваю тся все виды соединительной ткани, гладкая м ускулатура,
кровеносная и лимфатическая система.
Классификация органов по их происхождению в эмбриогенезе из
определенного зародышевого листка довольно распространена, однако
носит условный характер, та к как в построении лю бого органа или
части организма приним аю т участие производные нескольких зароды­
ш евых листков.
178
6.4.6. П ровизорные органы |ародышей позвоночных
В эмбриональном развитии позвоночных и прежде всего амниот
б о л ьш ую роль и граю т п р о в и з о р н ы е
органы,
которы е
функционируют у зародыш а и о тсу тс тв у ю т во взрослом состоянии.
К ним отн осятся ж елточн ы й мешок и та к н азываем ые зародыш евые
оболочки — аминон, хорион и аллантоис (рис. 84).
Образование ж е л т о ч н о г о м е ш к а описано при рассмотре­
нии гаструляции у птиц (см . 6.4.4). Ж е л то ч н ы й м еш ок вы п о лн яет ряд
179
важнейших функций. В о внезародышевой мезенхиме возникаю т скопле­
ния клеток — кровяные^ТГстрОвки"! К л т п г " с ' периферии островка
преобразую тся в пласт и ограничиваю т просвет будущего кровеносного
сосуда. К л е тк и кровяного островка, располагающиеся во внутренней
его зоне, превращ аю тся в кровяные клетки. Объединение сосудов
приводит к формированию сосудистой сети. Ч а с т ь сосудов входят в
складки, образуемые на внутренней поверхности стенки ж елточного
меш ка, которые проникаю т глубоко в массу ж елтка, что создает
благоприятны е условия для всасывания продуктов расщепления
питательных веществ в кровеносное русло. Через специальные сосуды
кровеносная сеть желточного меш ка соединяется с сердечно-сосудистой
системой зародыша. Благодаря описанному строению ж ел то чн ы й
м еш ок о сущ ествляет функции питания, дыхания, выделения, кроветво­
рения.
В образовании ам ниона и хориона участвует в незародышевая
эктод ерм адпгаруж ны п пигггГк дц|здродышёвоД мезодермы . ТГервонЗчаЖ нсвбтгйзИ Головного конца зародыш а о бразую тся складки, которые
распространяю тся спереди назад, сб л и ж аю тся и, наконец, см ы ка ю тся
над телом зародыш а по средней линии. Последний оказы вается
заклю ченным в две оболочки — ближ айш ая к нему называется
амниотической,
следующая
сеоозной
или
*Орионом.
А м ­
н и о т и ч е с к а я о б о л о ч к а ограничивает п ол о с т ь
а м ­
ниона,
заполненную
а м н и о т и ч е с к о й
ж и д к о с ть ю ,
омы ваю щ ей теперь зародыш со всех сторон, и благодаря ко то ­
рой зародыш развивается в водной среде, что предохраняет его
о т механических травмирующ их воздействий, прилипания к обо­
дом кям
Благодаря с е р о з н о й о б о л о ч к е снаружи о т ам ниотиче­
ской
формируется
еще
одна
полость,
называемая
в н ез а р о д ы ш е в ы м
ц ело м о м
или
экзои ело м ом .
Е щ е один провизорный орган — а л л а н т о и с ,
образуясь как
выросг^задней киш ки, врастает в экзоцелом и в конце концов заполняет
ее целиком. Гл авн а я функция аллантоиса состоит в то м , ч то он является
зарод ы ш евы м органом выделения. В нем скапливаю тся продукты
распада, образующ иеся в ходе обмена веществ в теле зародыш а.
Т ак, стенка аллантоиса, в которой развивается богатая сеть
кровеносных сосудов, плотно прилегает к серозной оболочке, посуш еству срастаясь с ней ( х о р и о н - а л л а н т о и с ) .
Последнее
обстоятельство создает необходимые условия для газообмена, т . е. в ы ­
полнения хорион-аллантоисом дыхательной функции. Серозная обо­
лочка совместно с аллантоисом принимает, по-видимому, участие в
утилизации в процессах развития веществ белковой оболочки яйца.
Своеобразие процесса эмбрионального развития высших млекопи­
таю щ их распространяется и на провизорные органы.
У млекопитаю щ их, та к ж е как у рептилий и птиц, о бразую тся амни­
он, аллантоис, ж елточны й мешок. Последний в связи с алецитальнос т ью яйцеклеток существенной роли в литании зароды ш а не играет.
Е щ е одна особенность м лекопитаю щ их со стои т в то м , ч то у
б ольш инства из них амниотическая полость формируется не путем
180
Ри с . К5. О б о л о ч к и ы р и д ы ш я чс-
см ы кания складок, образуем ы х внезародышевой эктодермой (троф областом), а путем расхождения клеток зародыш евого узелка.
Оболочки зародыша человека показаны на рис. 85 и 86.
Бо льш ое значение в установлении связи зародыш а с м атеринским
организмом при внутриутробном развитии принадлежит изменениям
трофобласта. П о его наружной поверхности о бразую тся много­
численные в ы р о сты — п е р в и ч н ы е
ворсины.
Последние
превращ аются во в т о р и ч н ы е в о р с и н ы после того, как в них
врастает виезародышевая мезенхима, в которой интенсивно образую тся
181
Рис. 87. Типы плацент.
а _ элителиохорнальная; б — соедиинтелыюткаино-хориальиая: в — эндотелнохориальная; г — гемохориальиая; I — эпителий хорноиа; 2 — эпителий матки; 3 — к^рвеиосные сосуды хориона; 4 — крове­
носные сосуды матки; 5 — материнская кровь.
кровеносные сосуды. Возникает в о р с и н ч а т а я о б о л о ч к а
или х о р и о н , гомологичная серозной оболочке рептилий и птиц.
Ворсины хориона образую т зародышевую часть п л а ц е н т ы
( д е т с к о е м е с т о ) — органа, при помощи которого зародыш
получает из организма матери все необходимое для развития.
Материнская часть плаценты представлена участком стенки м атки, с
которым связаны ворсины хориона. Процесс внедрения зародыша в
стенку матки называется и м п л а н т а ц и е й .
В зависимости от уровня контакта ворсины и слизистой оболочки
матки различаю т несколько типов плацент (рис. 87).
В плаценте э п и т е л и о х о р и а л ь н о г о
т и п а ( полуп л а ц е н т е ) , характерной для свиньи, лошади, бегемота, верблюда,
китообразных, ворсины хориона погружены в складки слизистой
оболочки матки без нарушения целостности эпителия, выстилающ его
ее просвет.
Д ля жвачных типична д е с м о х о р и а л ь н а я (соединительно­
тканнохориальная) плацента. В этом случае целостность эпителиаль­
ной выстилки полости матки нарушается. Ворсины хориона погружены
в соединительную ткань стенки органа, однако прямой контакт с крове­
носными сосудами отсутствует.
В* плаценте э н д о т е л и о х о р и а л ь н о г о т и п а ворсины
хориона вступаю т в непосредственный контакт с эндотелием —
клетками, ограничивающими просвет сосудов стенки матки.
Г е м о х о р и о н а л ь н а я плацента приматов и человека обес­
печивает ^ иболее тесную связь зародыша с организмом матери.
В этом случае ворсины хориона омываю тся кровью , изливающейся из
сосудов в силу нарушения целости их стенок в лакуны, которые образу­
ю тся в стенке матки в области детского места.
6.5. М Е Х А Н И З М Ы О Н Т О Г Е Н Е З А Н А К Л Е Т О Ч Н О М
И Н АД КЛЕТО ЧН О М УРО ВН ЯХ
В условиях нормального развития преобразования отдельных частей
и организма в целом строго согласованы по срокам, месту и объему
изменений.
Процесс, в результате которого относительно однородный ма­
териал зародыша в ходе эмбриогенеза преобразуется в устойчивые
элементы, разнородные по морфологии, биохимическим показателям,
функциям и т. д., называю т д и ф ф е р е н ц и а ц и е й
(д и фф е р е н ц и р о в к о й ) . Э то т процесс распространяется на все глав­
ные структурные уровни организма. Так, возникновение разнооб­
разия типов клеток соответствует явлению цитологической дифференцировки, а образование разных тканей — гистологической.
В процессе дифференцировки клеточный материал эмбриональных
Л закладок преобразуется в определенный элемент взрослого организма.
Напомним, что мезодермальные сомиты подразделяются на дерматом,
склеротом и миотом. Первый из названных зачатков дает соедини­
тельнотканные клетки (фибробласты) дермы кожи, второй — клетки
хряща (хондроциты), третий — поперечнополосатые мыш ечные во­
локна. Следовательно, конечный результат развития отдельных
эмбриональных закладок предопределен или детерминирован. Для
обозначения предопределенности, необратимости дифференцировки в
эмбриогенезе используют термин « д е т е р м и н а ц и я » . Некоторые
эмбриологи отождествляю т с понятием детерминации совокупность
факторов, обусловливающих закономерный характер формообразова­
тельных процессов, или же воздействие одной части зародыша на
другие его части, побуждающие эти последние к прохождению в подхо­
дящих условиях фрагмента пути своего нормального развития.
Степень предопределенности путей дальнейшего развития конкрет­
ной части зародыша в ходе эмбриогенеза меняется. Так, опыты по
разделению, сращиванию или перемешиванию бластомеров на стадии
дробления показали, что у видов с радиальным (регуляционным) ти­
пом дробления бластомеры нескольких поколений, если их изолиро­
вать
и
поместить
в
подходящие
условия,
проявляю т
тотипотентность
(равнонаследственность), т . е. развива­
ю тся в полноценный организм. У многих гидромедуз, например, клетки
тотипотентны до стадии 32 бластомеров. Равнонаследственность
клеток обнаружена у кроликов на стадиях 2 и 4 бластомеров. У мышей
нормальный ход развития воспроизводится как из отдельных бластоме­
ров, так и из нескольких объединенных зародышей. В последнем случае
кратного увеличения структур и частей не наблюдалось. За равно­
наследственность и тотипотентность клеток зародышей человека до
стадии 2— 4 бластомеров говорят случаи рождения двух, трех, четырех
однояйцевых близнецов. Одна двойня приходится примерно на 80,
тройня на 6400 и четверня на 512 ООО родов.
*
Н а стадии бластулы клетки уже не проявляю т свойства тотипотентности, однако способны к переопределению ( т р а н с ­
детерминации)
пути дальнейшего развития. О пы ты , вы­
полненные на зародышах амфибий, показали, что перекрестные
пересадки участков экто- или мезодермы приводят к изменению
направления дальнейшего развития в соответствии с новым положе­
нием.
Результаты аналогичных пересадок на стадии гаструлы свиде­
тельствую т об утрате способности клеточного материала к трансде183
а
б
в
г
Рис. 88. Первичная эмбриональная индукция у амфибий.
а — м атер и ал донора (у ч а с т о к дорсальной губы б л а с т о п о р а ); б — т р а н сп л а н та ц и я м ате р и ал а донора иа
б р ю ш н у ю стор он у зароды ш а-рец ипиента: в — об р азо ван и е второй нервной тр у б к и ; г — обр азо ван и е
второго зар о д ы ш а .
терминации
(см .
6.5.1).
Таким
образом,
лабильная
детерминация
путей развития, наблюдаемая на ранних
стадиях, сменяется с т а б и л ь н о й д е т е р м и н а ц и е й .
П о мере прохождения зародышем последовательных стадий
эмбриогенеза происходит ограничение или даже утрата отдельными
зачатками потенций к изменениям в нескольких направлениях, т . е.
имеет место к а н а л и з а ц и я р а з в и т и я .
Н а любой из стадий эмбриогенеза зародыш представляет собой
целостность, что обусловливает взаимосвязь и взаимосогласованность
развития отдельных его частей. Дифференциация и детерминация
осущ ествляю тся на фоне и н т е г р а ц и и всех проявлений развития в
единый сбалансированный процесс. Изменение потенции отдельны^
частей к развитию происходит не случайно, а в зависимости от
положения этих частей в преобразующемся зародыше в ходе
эмбриогенеза в целом. Такая интеграция делает результаты развития •
определенными не только в деталях, но и в целом.
6.5.1. Эмбриональная индукция
Больш ое значение в упорядочении хода эмбриогенеза принадлежит
эмбриональной
индукции.
Начало принципиальному
изучению этого явления положил опыт Г . Шпемана и Г . Мангольд,
результаты которого были опубликованы в 1924 г. В нем дорсальная
губа бластопора, подлежащая в нормальных условиях эктодерме,
развивающейся в структуры нервной системы, из зародыша гребенча­
того (непигментированного) тритона на стадии ранней гаструлы
вырезалась и пересаживалась под эктодерму брюшной стороны,
дающую в дальнейшем эпидермис кожи зародыша примерно той же
стадии пазвития
обыкновенного
(пигментированного) тритона
(рис. 88). В итоге на брюшной стороне зародыша- реципиента возника­
ли сначала нервная трубка и другие компоненты комплекса осевых
органов — хорда, сомиты, а затем формировался дополнительный
зародыш. Наблюдения за распределением пигментированных и непигментированных клеток показали, что ткани дополнительного
зародыша формируются почти исключительно из клеточного материа­
ла реципиента.
Приведенные данные убедительно доказываю т, что в ходе
эмбриогенеза некоторые части зародыша выполняю т роль и н184
Рис. 89. Индукция хрусталика глаза.
I — передний м озг; 2
зкто д ер м а, 3 -- глазн ой п у зы р ь; 4
6 — х р у ста л и к ; 7 — роговиц а; 8 — с е т ч а т к а .
глазной б о к а л ; 5 — з а ч а т о к х р у ста л и к а;
дукторов
или
организаторов
(по терминологии
Г . Ш пемана), намечающих пути развития других частей. Явление
эмбриональной индукции состоит в побуждении к развитию в опреде­
ленном направлении одних структур зародыша в результате воздей­
ствия на них других структур, возникающих на более ранних стадиях.
Отдельные примеры индукционных воздействий ограниченного
характера, например образование хрусталика из эктодермы под
действием зачатка глаза (рис. 89), были известны и ранее. Значение
результатов опыта Г . Шпемана и Г . Мангольд состоит в установлении
факта п е р в и ч н о й э м б р и о н а л ь н о й и н д у к ц и и , т. е.
первого шага в цепи последовательных ( в т о р и ч н ы х ,
тре­
т и ч н ы х ) индукционных процессов в дальнейшем развитии.
Дорсальная губа бластопора, представляющая по своим потенциям
хордо-мезодермальный зачаток, является первичным индуктором и ор­
ганизатором у амфибий. У рыб ему соответствует дорсальный край
бластодиска, у птиц — первичный узелок.
Зачаток бластопора у ам ф и б и й в о зн и к а е т в области серого серпа.
Если небольшой участок кортикального слоя цитоплазмы яйцеклетки
лягуш ки из области названной структуры пересадить на брюшную
сторону другого зародыша, то у последнего индуцируется дополни­
тельная нервная система. Можно предположить, что клеточный
материал дорсальной губы бластопора наследует свойства первичного
организатора, которые были каким-то образом запрограммированы
еще на уровне яйца.
Многочисленными исследованиями, выполненными в 20—30-х го­
дах текущего столетия, показано, что в условиях эксперимента
индукцию развития эктодермы в направлении нервной системы вы зы ­
ваю т многие факторы — вы тяж ки из разных органов беспозвоночных и
позвоночных животных, тканей растений, неорганические вещества.
Наряду с этим было установлено, что сущ ествую т «специфические
индукторы», т . е. вещества, оказывающие индуцирующее действие в
ничтожных концентрациях, и различающиеся по конечному результату
своего действия. Так, экстракт из печени млекопитающих индуцирует
главным образом мозговые структуры , а из костного мозга — мезодермальные. При совместном воздействии обоих индукторов формиро­
вался зародыш почти нормального вида. В тканях куриных зародышей
185
высокоактивные индукторы относятся к классу белков или нуклеопротеинов.
В
развитии
многих
зачатков
вы являю тся
цепи
по­
следовательных
индукций.
Так, описана индукция
глазным бокалом хрусталика, хрусталиком .и даже взрослым глазом
роговицы. Продолговатый мозг индуцирует развитие слухового
пузырька, а последний — хрящевую капсулу. В отличие от первичной
эмбриональной индукции, результатом которой служит образование
дополнительного зародыша, примеры, описанные выше, относятся к
тканевому и органному уровню структурной организации. В основе
таких межорганных и межтканевых индукций лежат, по-видимому, не
химические, а контактные воздействия одних клеток на другие.
Важ ны м обстоятельством служит то , что в нормальном развитии
индуктор оказывает соответствующ ее действие лишь в отношении
зачатков, которые характеризуются восприимчивостью. Способность
эмбрионального зачатка к восприятию индукционного стимула называ­
ется к о м п е т е н ц и е й . Таким образом, индукционные процессы в
эмбриогенезе происходят благодаря приобретению одними частями
свойств индукторов, а другими — свойства компетентности.
В парах элементов «индуктор — компетентный зачаток» содержа­
ние изменений, провоцируемых индуктором, зависит от внутренних
потенций зачатка. Так, зачаток бедра задней конечности цыпленка
пересаживали под эпителий зачатка конечного (дистального) отдела
крыл».
И з трансплантата под влиянием эпителия, в норме индуцирующего
конечный отдел крыла, из презуптивного материала бедра сформирова-,
лись дистальные структуры , из ноги — стопа, фаланги пальцев.
Современные исследования показали, что действие индуктора не
воспринимается одиночными клетками, причем клетки в трехмерном
скоплении изменяются быстрее, чем будучи распластаны тонким слоем.
Чем больше масса индуцируемого зачатка, тем активнее в нем
происходит дифференцировка частей.
Такие характеристики эмбриогенеза, как тотипотентность частей
зародыша на достаточно ранних стадиях, прогрессивное ограничение
путей развития зачатков, явление нарастающей дифференциации, о
которых ш ла речь выше, хорошо согласуются с наличием цепей
индукционных процессов. При этом закономерная смена индукторов .и
состояний компетентности могут служ ить инструментом детерминации
последовательных этапов развития: от значительных (например,
формирование комплекса осевых органов) до ограниченных объе­
мом органа или клеточной группы.
Наблюдения показываю т, что зачаток почти любого органа
проходит в своем развитии две фазы. В фазе з а в и с и м о й
д и ф ф е р е н ц и р о в к и его судьба во многом зависит от действия
индукторов и внешнего окружения. С определенного момента зачаток
вступает в фазу н е з а в и с и м о й д и ф ф е р е н ц и р о в к и и
осущ ествляет закономерный цикл преобразований даже при изменении
внешних условий. Трансплантация зачатка в нетипичное окружение
186
в 1-й фазе приведет к т р а н с д и ф ф е р е н ц и р о в к е , во 2-й — не
вызовет изменения пути развития.
Представления о смене организаторов и состояния компетенции
зачатков как факторах детерминации последовательных этапов разви­
тия структур не противоречит положению о том , что на любой стадии
организм является целостностью, а не мозаикой органов и частей.
Целостность обусловливается системой связей между отдельными
элементами зародыша, характеристики которой закономерно изменя­
ю тся. Лиш ь условно можно говорить об одних частях зародыша как об
индукторах, а о других — как о реагирующих элементах. В процессе
развития, вклю чаясь в разные системы связей, «индукторы и реакторы»
(по терминологии И . И . Ш мальгаузена) постоянно меняются ролями.
Ф акторы , обусловливающие закономерный характер итога развития в
целом и на отдельных этапах, возникают по мере дифференцировки
зародыша благодаря взаимодействию результатов этой дифференци­
ровки.
Типы взаимозависимостей, наблюдаемых в индивидуальном
развитии (онтогенетические корреляции) рассматриваю тся в 7.12.5.
6.5.2. Изменения клеток и клеточных комплексов
в эмбриональном развитии
В ходе эмбрионального развития изменения наблюдаются на всех
уровнях структурной организации зародыша.
К типичным преобразованиям клеточных комплексов относятся
м е с т н ы е у т о л щ е н и я или о б о с о б л е н и я пластов, о бразование складок,
объединение
или р а с п а д
клеточных масс,
сгущения
клеток
или
их
кон­
ц е н т р а ц и я вокруг каких-либо структур.
В пользу участия указанных преобразований в формообразова­
тельных процессах говорят многие
факты. Так, сгущение ме­
зенхимных клеток предшествует образованию зародышевых крове­
носных сосудов, хряща, кости или мышечной ткани. Клетки из зачатка
глаза развиваю тся в сетчатку, находясь в трехмерном скоплении, и в
пигментный эпителий, будучи распластанными. Собственно, это и
происходит в результате превращения глазного пузыря в глазной бокал
при нормальном развитии.
Механизмы, обусловливающие зависимость формообразователь­
ных процессов о т преобразований клеточных комплексов, пока не
выяснены. Они м огут способствовать или затруднять диффузию
веществ, изменять характер контактных взаимодействий.
Важная роль в эмбриогенезе принадлежит пролиферации, перемеще­
ниям, избирательной сортировке, дифференцировке, гибели клеток
зародыша.
К л е т о ч н о е р а з м н о дк е н и е (пролиферация) лежит в ос­
нове развития всех органов. Благодаря ему достигается определенная
масса тканей. В отдельных зачатках делящиеся клетки м огут распола­
гаться без видимого порядка или концентрироваться в особых
м а т р и ч н ы х з о н а х . Последнее наблюдается, например, в за­
187
чатке центральной нервной системы, где митозы происходят только в
клеточном слое, прилежащем к нейроцелю. В отдельных случаях от
матричной зоны зависит дальнейшая судьба клетки. Так, в развиваю­
щемся головном мозге млекопитающих к концу эмбрионального
периода образуются в дополнение к первичной матричной зоне,
упомянутой выш е, вторичные матричные зоны, в частности в мозжеч­
ке. В таких зонах образую тся только мелкие короткоотростчатые
нервные клетки, обеспечивающие межнейрональные связи внутри
отдельных нервных центров.
Активные к л е т о ч н ы е п е р е м е щ е н и я в эмбриогенезе
обеспечиваются, главным образом, благодаря механизму амебоидного
движения. Траектория задается, по-видимому, особенностями рельефа
поверхности, по которой клетка движется, путем так называемой
контактной ориентировки.
Некоторые типы клеток
перемещаются по градиенту концентрации химических веществ
( х е м о т а к с и с ) , однако этот механизм ориентации встречается
редко.
Двигательная активность служ ит целям доставки клеточного
материала в нужную область зародыша. Так, клетки, выселяющ иеся из
ганглиозной пластинки, мигрируют в зоны развития чувствительных и
вегетативных нервных узлов, надпочечников, хрящей жаберных дуг и их
производных. В отдельных случаях контакты по ходу движения
определяют конечную судьбу клеток. Например, для развития
симпатических нервных клеток необходим контакт с определенным
участком мезодермы во время движения клеток-предшественниц
из ганглиозной пластинки в область локализации симпатических
узлов.
Явление и з б и р а т е л ь н о й с о р т и р о в к и состоит в вы ­
делении и объединении клеток одного зачатка из смеси, содержащей
клетки разных зачатков. Оно распространяется на клеточный материал
как зародышевых листков, так и отдельных органов. Процесс
объединения сходных клеток носит, по-видимому, вероятностный
характер. М ежклеточные контакты образуются случайно, однако свя­
зи однотипных клеток отличаю тся большей устойчивостью . Значение избирательной сортировки в развитии заклю чается в окончательном
упорядочении положения клеток в клеточных комплексах. Злокаче­
ственно перерожденные клетки не отделяю тся из смеси с нормальными
клетками, что может лежать в основе их прорастания в различные тка­
ни и органы.
Д и ф ф е р е н ц и р о в к е к л е т о к принадлежит в процессе
развития особая роль. Благодаря ей возникаю т устойчивые типы
клеток, специализированные к выполнению разных функций. Диффе­
ренцированные клетки нередко характеризуются сложной морфологией
(нервные клетки, палочки и колбочки сетчатки). Фундаментальной
чертой, однако, по которой клеточные типы отличаю тся друг от друга,
служит образование ими т и п о с п е ц и ф и ч н ы х б е л к о в , а
также веществ иной химической природы, например углеводной. Так, в
мышцах синтезируется сократительный белок миозин, в фоторецепто­
рах — зрительный белок опсин, в эритроцитах — гемоглобин. О т
188
наличия этих белков зависит функциональная специализация клеток
многоклеточного организма.
Наряду с типоспецифичными белками клетки, принадлежащие
одной ткани, синтезируют некоторое число общих для них
тканеспецифичных
белков. Все клетки, независимо от
тканевой и органной принадлежности, образуют к л е т о ч н ы е
белки, о б е с п е ч и в а ю щ и е п р о я в л е н и я ж и зн е д е ­
я т е л ь н о с т и , о б щ и е для всех клеток.
В настоящее время не вы зы вает сомнения, что синтез отдельных
белков контролируется определенными генами, а полный набор генов,
за редким исключением (например, безъядерные эритроциты млекопи­
тающ их), идентичен в клетках разных типов. Генетические ос­
новы
клеточной
дифференцировки
объясняет
гипотеза
д и ф ф е р е н ц и а л ь н о й а к т и в н о с т и г е н о в . Согласно
ей различия в спектре белков, образуемых дифференцируемыми
клетками, отражаю т различия в наборе активных генов. В клетках
любого направления специализации выделяю тся как бы три группы
активных генов: 1) контролирующие фундаментальные процессы жиз­
недеятельности клеток (например, энергетические) и активные во всех
живых клетках; 2) обусловливающие сходные черты клеток одной
ткани (например, возбудимость мембраны нервных клеток, наличие у
них устойчивых отростков); 3) контролирующие черты, специфичные
для клеток конкретного типа, активность которых лежит в основе,
например, морфологических и цитофизиологических различий груше­
видных нейронов мозжечка, гагантопирамидальных нейронов коры
большого мозга.
Главны й механизм клеточной дифференцировки, согласно совре­
менным представлениям, сводится к
избирательному
блокированию — деблокированию
или и н а к т и ­
в а ц и и — а к т и в а ц и и отдельных генов или их групп. Способы
достижения перечисленных состояний генов интенсивно изучаю тся,
однако все еще далеки от понимания. Больш ая роль принадлежит, повидимому, изменению физико-химических свойств хроматина, которое
во многом зависит от характера взаимодействия Д Н К , гистоновых н
негистоновых белков.
,
В ходе дифференцировки клеток зачатка зародыша наблюдается
последовательная смена активных генов. Удобным объектом для
демонстрации этого явления служ ат гигантские политенные хромосомы
клеток некоторых органов насекомых, например дрозофилы. Зоны
таких хромосом, содержащие активные гены, на которых интенсивно
синтезируется Р Н К , выглядят как вздутия. В зависимости от стадии
развития и тканевой принадлежности клетки распределение пуффов по
длине хромосомы варьирует (рис. 90).
П о мере дифференцировки число активных генов в клетке, повидимому, прогрессивно снижается. Так, из 40 ты с. генов генома
морского ежа на стадии бластулы активно примерно 30 ты с., гаструлы
и личинки — 12— 15, у взрослых животных — 3— 5 ты с. генов.
Регуляция генной активности клеток зародыша в процессе развития
достаточно сложна и во многом неясна. Регуляторные влияния
189
оказываю т вещества цито­
плазмы
яйцеклетки
(см.
6.3.1), характер межклеточ­
ных химических и контактных
взаимодействий (см . 6.5.1),
гормоны.
В эмбриональном разви­
тии многих органов и частей
организма наблюдается з акономерная
(запро­
г р а м м и р о в а н н а я )
г и б е л ь к л е т о к исход­
ного зачатка. Некоторые при­
меры указываю т, что она яв­
ляется инструментом прида­
ния зачатку окончательной
формы. Так, гибель клеток
служит разъединению закла­
Рис. 90. Пуффы хромосомы.
док
фаланг пальцев у птиц и
а — н еа кти вн ы й у ч а с т о к хромосом ы; б — у ч а с т о к хр о м о­
со м ы с д в у м я пуф ф ами.
млекопитающих. В развиваю­
щихся зачатках плеча и пред­
плечья курицы зоны закономерно гибнущих клеток размещены в опре­
деленных местах.
В других случаях роль клеточной гибели менее понятна* Например,
на известной стадии развития многих центров головного мозга
млекопитающих закономерно погибает более половины клеток. П р и.
этом среди гибнущих клеток есть и такие, отростки которых уже
установили контакт с органом-мишенью.
6.5.3. Морфогенез
В эмбриогенезе осущ ествляю тся интенсивные ф о р м о о б р а ­
зовательные
(морфогенетические)
процессы.
Благодаря им организм в целом и его отдельные части, с одной сторо­
ны, приобретают форму, типичную для взрослого состояния, а с дру­
гой — форма конкретных зачатков закономерно изменяется в ходе
развития1.
Примером морфогенеза служ ит процесс ф о р м и р о в а н и я
п а р н ы х к о н е ч н о с т е й позвоночных (рис. 91).
Первым
идентифицируемым
морфологическим
проявлением
этого процесса является образование почки конечности. Ее формиру­
ет сгущение мезенхимных клеток, мигрировавших из париетального
листка боковой пластинки мезодермы, покрытое утолщенным слоем
эпидермиса. Довольно рано вы является ориентация зачатка конечности
1 Исторически термин «морфогенез» был предложен дли обозначения преобразо­
ваний формы структур надклеточного уровня. В настоящее время его используют
такж е в отношении формообразовательных процессов на клеточном, субклеточном и
доклеточном (сборка вирусов н фагов) уровнях. Здесь этот термин употребляется в
первоначальном, более узком значении.
190
Рис. 91.. Формирование (морфогенез) конечности эмбриона человека.
а — п о чк а верхней кон ечн ости (д ли н а зар о д ы ш а 12 м м ); б — к и с ть и предплечье (д ли н а зар о д ы ш а
15 м м ); в — и зм енение ф ормы з а ч а т к а р у к и (д ли н а за р о д ы ш а 17 м м ); г — к и с ть и предплечье
(длина зар о д ы ш а 20 м м ); д — к и с т ь и Предплечье (д ли н а зар о д ы ш а 25 м м ); е — кисти плода (д л и ­
на 52 м м ).
относительно главных осей тела зародыша. У амфибий, например,
почки передних конечностей направлены вершинами вниз и назад.
Морфогенетические преобразования зачатка заклю чаю тся первона­
чально в его уплощении в виде «лопатки» или «пластинки», неровности
краев которой соответствую т будущим пальцам. Внутри зачатка
образую тся уплотнения мезенхимных клеток, которые, дифференциру­
ясь, приобретают способность к синтезу вещества хряща. Хрящ евые
закладки в дальнейшем преобразуются в скелетные элементы. Н а этом
этапе формируются основные отделы конечностей — плечевой и тазо­
вый пояса, плечо или бедро, элементы предплечья, голени, кисти,
стопы. К ак уже отмечалось, в преобразованиях хрящевых закладок
конечностей известную роль играет закономерная гибель части клеток.
Сущ ествует определенная временная последовательность образова­
ния закладок разных отделов конечности. В целом проксимальные
части скелета возникаю т раньше дистальных. В процессе формирования
закладок основных отделов наблюдаются закономерные перемещения
клеточных комплексов в виде сгибания, вращения. М ыш цы образуются
из материала нижних краев миотомов, врастающего (амфибии) или
мигрирующего (высшие позвоночные) в зачаток конечности.
В развитии конечности важное место принадлежит межтканевым
индукционным взаимодействиям. С одной стороны, мезенхимные клет­
ки, воздействуя на эпидермис, побуждают его к утолщению, с дру­
гой — именно эпидермис обусловливает преобразование формы за­
чатка — в случае удаления его может подавляться расчленение
мезенхимы на хрящи. Так, при пересадке эпителиального покрова
191
а
б
Рис. 92. Формирование (морфогенез) лица.
а — эбриои длиной 3,5 м м. I — лобны й отр о сто к; 2 — н о со вая п лако д а: 3
р о то в ая п л а сти н ка ; 4 — вер х­
нечелю стной о тр осток; 5 — н и ж н е ч е л ю стн а я д у га; б — эмбрион длиной 6.5 мм: I
лобны й о тр осток.
2 — н осо вая я м к а ; 3 — рот; 4 — верхнечелю стной о тр осток; 5 — н и ж н еч е л ю стн ая д уга; 6 - п о д ъ язы чн а я
(ги о и д и а я) д у га; в
эмбрион длиной 9 мм. г
эмбрион длиной 12 м м: I - средненосовой отр о­
сто к ; 2 — боковой носовой отр осток; 3
носогиоидная бор озд ка: 4
верхнечелю стной отр осток;
5
н и ж н я я челю сУ ь.
зачатка крыла на зачаток ноги курицы вместо ноги возникает
крылоподобная структура. Д ля выяснения общих закономерностей
морфогенеза интересны результаты опытов по развитию добавочных
конечностей у хвостатых амфибий при подсадке им под кожу между
зачатками передней и задней конечности инородных тканей, например
слухового пузырька. Развитие передней или задней добавочной
конечности зависит от того, подсаживается ли ткань-стимулятор ближе
к передней или задней собственной конечности. Приведенные результа­
ты позволяю т предположить наличие различающихся по морфогенети­
ческим потенциям «территорий передней и задней конечностей»,
локализация которых в развивающемся организме точно определена.
Сложный характер преобразований сопровождает развитие лица и
ротовой полости.
Развитие
лица
и ротовой
полости
(рис. 92)
начинается образованием в месте контакта передней кишки и экто­
дермы ротовой ямки. Ч асть дорсальной стенки ротовой ямки развива­
ется в зачаток передней доли гипофиза, который'соединяется с зачатком
задней доли гипофиза. Последняя является производным промежу­
точного мозга. Вход в ротовую яму у зародышей млекопитающих
сначала представляет отверстие, ограниченное пятью выступами:
верхним непарным (лобный), двумя парными верхними (верхнечелю­
стные отростки) ’ и двумя парными нижними (нижнечелюстные
отростки). Последние, срастаясь, образуют нижний край ротового
отверстия. В формировании верхнего края ротового отверстия участву­
ю т верхнечелюстные отростки, которые срастаю тся с боковыми
192
носовыми и со средним лобным отростками, возникающими в ре­
зультате преобразования лобного вы ступа. Н а среднем лобном
отростке образуется поперечный носовой валик — зачаток кончика
носа. Наружные носовые отверстия возникаю т из обонятельных ямок, а
крылья носа — из боковых носовых отростков. Задняя стенка пер­
вичной ротовой полости прободается и соединяется с кишкой. Н а
внутренних поверхностях верхнечелюстных отростков появляю тся
небные отростки. Срастаясь, они разделяют первичную ротовую
полость на собственно ротовую и носовую. В это же время из
материала среднего лобного отростка образуется носовая перегородка.
Края небных отростков срастаю тся между собой и нижним краем
носовой перегородки, вследствие чего небо приобретает окончательное
строение.
Зубы являю тся производными эктодермы (эм аль) и мезенхимы
(пульпа, дентин).
Я зы к образуется первоначально как выступ нижней стенки кишки
между челюстной (мандибулярной) и подъязычной (гиоидной) дуж­
ками.
6.5.4. Факторы интеграции процессов развития
Развитие зачатка конечности, лица, ротовой полости убедительно
показывает, что видное место в морфогенетических процессах
принадлежит преобразованиям отдельных клеток и клеточных ком­
плексов. Главны е типы таких преобразований были рассмотрены ранее
(см . 6.5.2). О тметим только, что для выполнения ими формообразова­
тельных функций клеточные изменения должны характеризоваться
избирательностью в смысле места, времени проявления и степени
выраженности.
Описательная и экспериментальная эмбриология дает много
примеров том у, что в процессе развития в зависимости от зачатка и ста­
дии наблюдается именно избирательная пролиферация, миграция,
дифференцировка, сортировка, гибель клеток. Важнейшая задача
современной биологии развития состоит в том , чтобы вскры ть природу
факторов пространственно-временнбй упорядоченности изменений кле­
точного материала зародыша, обусловливающей закономерный мор­
фогенетический результат.
Ранние представления об индивидуальном развитии отражены в
двух взаимоисключающих концепциях — преформизме и эпигенезе.
Напомним, что сторонники концепции п р е ф о р м и з м а счита­
ли, что структура будущего организма во всех деталях представлена
(предобразована, преформирована) в половых клетках родителей.
Согласно их воззрениям, процесс развития заклю чается в росте частей
зародыша без их новообразования. Укреплению позиций преформизма
на рубеже X V II— X V III веков способствовали успехи микроскопической
анатомии, открывш ей совершенно новый мир микроскопических
структур. Однако в силу ограниченности технических возможностей
первых микроскопов в то время -нередко допускались ошибки в
интерпретации получаемых данных.
7— 56
193
Концепция э п и г е н е з а исходит из принципа постепенности
развития, в ходе которого происходит усложнение структуры орга­
низма. Серьезным экспериментальным обоснованием эпигенеза стали
результаты наблюдений К . Ф . Вольфа (1734— 1794) над развитием
зародыша курицы. Установив, что зачатки кишечника и нервной
системы первоначально представлены пластами и лиш ь затем приобре­
таю т форму трубок, он показал процесс формообразования. Важный
вклад в укрепление позиций эпигенеза внесли результаты исследований
К . М . Бэра (1792— 1876), который обнаружил единство плана строения
зародышей разных позвоночных, правильно описал яйцеклетку млеко­
питающих и человека, дал доказательства преемственности последова­
тельных стадий развития и усложнения организации зародыша.
Вместе с тем он не разделял представлений о том , что закладки на
ранних стадиях эмбриогенеза бесструктурны. По К . М . Бэру, индиви­
дуальное развитие не является ни предобразованием, ни новообразова­
нием, но преобразованием структур.
Полемика между преформистами и эпигенетиками вновь оживилась
в конце X IX века. Согласно теории н е о п р е ф о р м и з м а , вырази­
телем которой является В . Р у (1850— 1924), считается, что в яйце
находится не зародыш как таковой, но каждая часть будущего
взрослого организма представлена соответствую щ им участком яйца,
причем каждый участок может развиться лиш ь в определенную часть
этого организма, таким образом, между участками яйца и взрослого
организма сущ ествует взаимно однозначное соответствие, и эти участки
обладают одинаковой степенью упорядоченной сложности. Согласно
теории «зародышевой плазмы», в трехмерной архитектуре оплодотво­
ренного яйца заключен эскиз упорядоченной сложности взрослого
организма.
О пыты Г . Дриша (1867— 1945) на морских ежах, показавшие
возможность развития целого организма из отдельных бластомеров,
позволили
ему
принять
противоположную
теорию
( н е о э п и г е н е з а ) . Признание этой теории привело Дриша к вита­
лизму. Он считал, что пространственное разнообразие и упорядо­
ченность в развивающемся организме происходит под действием
особого нематериального фактора — энтелехии.
Как преформизм, так и эпигенез, даже с учетом фактов, установлен­
ных генетикой и молекулярной биологией, не отраж аю т действи­
тельной сути процесса развития. Конечно, в эмбриогенезе осущ ествля­
ется эволюционно обусловленная программа развития, характерная для
организмов определенного биологического вида. Э т а программа
отражена в структурах оплодотворенной яйцеклетки. Последнее,
однако, нельзя понимать так, что гены служат зачатками признаков или
что все признаки взрослого организма в той или иной форме непосред­
ственно представлены в зиготе. Вместе с тем на каждом из очередных
этапов развития организм приобретает новые качественные характери­
стики. Э то не означает, что в ходе эмбриогенеза происходит
новообразование структур вне связи с общим ходом развития. В плане
достижения конечного морфогенетического результата одинаково
важны наследственная предопределенность пути развития, свойства
194
генотипа как системы взаимодействующих генов и условия, в которых
развитие происходит. Ярким примером этом у служ ит, в частности,
формирование признаков пола (см . 5.2.9).
Развитие сложной формы, закономерной внутренней архитектуры
органов и частей многоклеточного организма происходит в результате
согласованных изменений большого числа клеток. Возникает вопрос о
природе м е х а н и з м о в
интеграции
изменений на кле­
точном и тканевом уровнях, составляющих основу преобразования
органов и организма в целом в нормальном развитии. То, что эти ме­
ханизмы не выводятся из поведения отдельных клеток, следует,
например, из такого опыта. П утем определенных воздействий можно
зачаток головного мозга тритона превратить в смесь клеток. Со
временем в подходящих условиях за счет избирательной сортировки
клеток восстанавливаются с достаточной степенью приближения к
нормальным зачатки глаз, внутреннего уха и др., которые, однако,
расположены в совершенном беспорядке.
П о отношению к отдельным закладкам зародыш в целом является
ка^бы средой, в которой происходят их преобразования. Д ля обозначе­
ния зависимости последних от свойств этой среды эмбриологи ввели
понятие «поля», заимствовав его из физики.
Концепция « м о р ф о г е н е т и ч е с к о г о п о л я » как фактора
пространственно-временного упорядочения и интеграции процессов
развития в эмбриогенезе разрабатывалась в отечественной биологии
А . Г . Гурвичем (1874— 1954), Н . К . Кольцовым (1872— 1940). Наличие
поля означает, что в развивающейся структуре действует простой ряд
сил, правильно распределяющихся в пространстве. Таким образом,
зародыш воспринимается как силовое поле, в пределах которого
обнаруживаются разности потенциалов. Природа последних, по
Н . К . Кольцову, разнообразна. Они м огут бы ть электрическими,
температурными, гравитационными. Суммарное поле зародыша обра­
зуется путем наложения полей отдельных клеток и их комплексов.
Переход к очередной стадии эмбриогенеза сопровождается перераспре­
делением разностей потенциалов и изменением результирующего поля,
что влияет на формообразовательные процессы. Слабым звеном
рассматриваемой концепции является вопрос о материальной природе
носителей поля, оказывающих влияние на морфогенез конкретных
закладок, например передней и задней конечности позвоночных.
Гипотеза «морфогенетического поля» представляет собой1теорети­
ческое построение, которое не противоречит главным фундаменталь­
ным откры тиям экспериментальной эмбриологии, в частности, таким,
как явление эмбриональной индукции. В свете гипотезы «поля» инте­
ресны данные американского биолога Ч . М . Чайлда (1869— 1954), поло­
женные им в основу т е о р и и
физиологических
или
« а к с и а л ь н ы х » ( о с е в ы х ) г р а д и е н т о в , согласно которой
сущ ествуют закономерные различия в уровне обмена веществ,
тестируемом по напряженности окислительно-восстановительных про­
цессов и некоторым другим показателям, в разных зонах зародыша,
т. е. своего рода градиенты метаболической или физиологической
активности. Так, в яйцеклетке лягушки вы является отчетливый
7*
*
195
а н и м а л ь н о - в е г е т а т и в н ы й г р а д и е н т , сохраняющий­
ся и после оплодотворения. В зиготе высокая активность метаболиче­
ских процессов обнаруживается также вдоль спинной стороны яйца к
области серого серпа. Д ля бластулы, кроме анимально-вегетативного,
обнаруживается д о р с о в е н т р а л ь н ы й г р а д и е н т с зоной
высокой
активности
в
области
дорсальной
губы
б л а с т о п о р а . В процессе гаструляции уровень активности в обла­
сти бластопора повыш ается, а в области анимального полюса падает.
С началом нейруляции зона высокой физиологической активности
переходит в область з а ч а т к а
х в о с т а . П о мере развития
зародыша система градиентов усложняется. Нарушения естественных
градиентов внешними воздействиями приводят к выраженным откло­
нениям в развитии.
Следует отм етить, что физиологические или метаболические
градиенты совпадают с основной осью и плоскостью симметрии, заро­
дыша, а полюсы активности — с зонами эмбриональных организато­
ров. Э то допускает предположение об их упорядочивающей роли в
эмбриогенезе. Изменение системы градиентов в ходе развития может
отраж ать преобразование зародыша как целого во времени.
6.5.5. Влияние внешней среды на развитие организма
Н а любом этапе онтогенеза организм сущ ествует в единстве с
окружающей средой. Эмбриогенез в этом отношении не является
исключением.
Норма отношений организмов определенного биологического вида
и среды обитания складывается в процессе эволюции. Развитие одних
видов происходит в широком диапазоне условий, тогда как другие виды
могут развиваться при ограниченных колебаниях параметров среды.
Для организмов любого вида сущ ествую т минимум, оптимум и макси­
мум необходимых условий развития.
По замечанию К . А . Тимирязева, каждый организм в процессе
своего развития предъявляет к среде определенные «требования». *
Последние в зависимости от стадии онтогенеза изменяются, причем их
использование организмом связано с его перестройкой. Так^у зароды­
шей костистых рыб первичным органом дыхания служ ит сосудистая
сеть стенки желточного мешка, у личинок сомовых, ведущих
придонный образ жизни, эту функцию выполняет грудной плавник.
Во взрослом состоянии функция дыхания переходит к жабрам.
В связи с задачей управления эмбриогенезом особый интерес
представляет действие на зародыш не необычных по природе и
чрезмерных по силе факторов, а колебания естественно встречающихся
факторов (температура, влажность, атмосферное давление, излучения,
газовый состав среды).
Так, в зависимости о т температуры процессы развития замедляются
или интенсифицируются. Например, яйца лягуш ки из одной клади за
один и то т же период при 20° С развиваю тся в зародыши с сформиро­
ванным хвостом, а при 15°С — достигают лишь стадии нейрулы.
Предполагают, что большое значение для нормального хода эмбриоге­
196
неза имеет температурный градиент, создаваемый наседкой в яйце:
температура верхней половины яйца на 2— 3 °С выш е, чем нижней.
Роль газового состава среды, в которой осущ ествляется развитие,
иллюстрирует пример с аскаридой. При прекращении доступа к
эмбриону кислорода развитие прекращается.
Общим правилом служит то, что под действием света из сине-фио­
летовой части спектра эмбриональное развитие многих видов животных
ускоряется, а из красной — замедляется.
Х о тя на любой стадии зародыш развивается лиш ь при сохранении
связи с внешней средой, в эволюции фактически каждого вида
выработались приспособления, обеспечивающие то т или иной уровень
автономности эмбриона по отношению
к
условиям
среды.
А в т о н о м и з а ц и я онтогенеза, освобождение от прямого дейс­
твия факторов среды на развивающийся организм соответствует
прогрессивному направлению эволюции (см. 7.8.3). Приспособления,
обусловливающие такое освобождение, разнообразны и проявляются
уже в строении яйцеклеток. Наибольшей эффективности они достигают
у высших позвоночных животных, особенно у млекопитающих.
В процессе эмбриогенеза меняю тся не только «требования»
зародыша к среде, но и его реакция на внешние воздействия. В частно­
сти, закономерно наблюдаются к р и т и ч е с к и е
периоды,
которые отличаю тся большой восприимчивостью эмбриона к дейс­
твию внешних факторов. Такая повышенная чувствительность обнару­
живается по отношению к факторам различной природы, т. е. она
лишена специфичности.
Положение критических периодов на временной шкале эмбриогенеза
соответствует поворотным моментам развития — началу гаструляции
у низших позвоночных, имплантации бластоцисты в слизистую обо­
лочку стенки матки у млекопитающих. Вне критических периодов
процесс развития характеризуется устойчивостью .
6.5.6. Нарушение эмбриогенеза
Нарушения естественного хода эмбриогенеза приводят к формиро­
ванию у р о д с т в или п о р о к о в р а з в и т и я . Пороками разви­
тия называю т стойкие отклонения в строении органа (рис. 93) или
целого организма (рис. 94), приводящие к функциональным расстрой­
ствам. Врожденные пороки встречаю тся у животных разных типов
структурно-функциональной организации, вклю чая человека. Причины,
механизмы и пути профилактики их изучает наука т е р а т о л о г и я ,
которая представляет собой пограничную область между эмбриоло­
гией, генетикой и медициной.
П о данным ВО З пороки развития имеют место у 1— 2 % людей. Не­
которые из них особенно часты . Так, дивертикул подвздошной кишки и
удвоение мочеточников обнаруживаются примерно у 4 % новорож­
денных.
Пороки развития заклю чаю тся в отсутствии органа или его части
( а п л а з и я ) , недоразвитии органов ( г и п о п л а з и я ) , уменьшении
массы тела ( г и п о т р о ф и я ) , непропорциональном увеличении
197
Рис. 93. Нарушения процесса формирования органов и частей тела.
а — н еза р ащ е и и е верхней гу б ы и твердого неба.
массы органа ( г и п е р т р о ф и я ) ,
увеличении
длины
тела
( г и г а н т и з м ), нетипичной локализации группы клеток или органа в
организме ( г е т е р о т о п и я , э к т о п и я ) , нарушении дифференци­
ровки тканей ( г е т е р о п л а з и я ) , сужении ( с т е н о з ) или отсут­
ствии (а т р е з и я) канала или отверстия, сохранении эмбриональных
структур ( п е р с и с т и р о в а н и е ) .
В зависимости от причины, приведшей к неправильному развитию ,
различают пороки генетической, экзогенной (внешней) и комбиниро­
ванной (мультифакториальной) природы.
В основе п о р о к о в г е н е т и ч е с к о й п р и р о д ы лежат
мутационные изменения наследственного материала (см . 5.3).
Экзогенные
пороки
возникаю т в связи с действием на
зародыш повреждающих внешних факторов. Некоторые из та­
ких пороков являю тся фенокопиями определенных генетических по­
роков.
Причиной м у л ь т и ф а к т о р и а л ь н ы х
п о р о к о в слу­
жит неблагоприятное сочетанное действие на процесс развития генети­
ческих и экзогенных факторов. Воздействия, приводящие к порокам,
называю тся т е р а т о г е н н ы м и .
Непосредственной мишенью действия неблагоприятных факторов
могут бы ть половые клетки, а такж е зародыш на разных стадиях
198
Рис. 93. П род олж ение.
б — ан ом али и р а зв и т и я конечностей.
ж
и
Рис. 94. Двойные уроды.
а. б, в — ср ас тан и е го л ово к; г. д. е — ср астан и е грудных к л е т о к ; ж . з. и — ср ас тан и е т а зо в ы х о б л астей .
эмбриогенеза. В первом случае говорят о г а м е т о п а т и я х , во
втором — об э м б р и о п а т и я х .
П е р в и ч н ы е п о р о к и возникаю т под действием терато­
генных факторов, в т о р и ч н ы е являю тся осложнениями первичных
пороков. Так, водянка головного мозга представляет собой вторичный
порок и развивается как осложнение первичного порока — атрезии
водопровода мозга.
Выраженность тератогенного действия неблагоприятных факторов
200
изменяется в зависимости от стадии эмбриогенеза. Ранимость
зародыша возрастает в критические периоды.
К ак уже отмечалось, у млекопитающих и человека критические
периоды с наибольшей чувствительностью к действию патогенных
агентов соответствую т стадиям имплантации эмбриона и развития
плаценты. Так, у крыс при действии наркотических средств, эфира,
рентгеновских лучей первый максимум смертности зародышей прихо­
дился на 4-й день развития, т. е. предимплантационный период.
Второй максимум смертности наблюдается на 10— 12-й день эмбриоге­
неза, т. е. на критический период развития плаценты. Все отклонения от
нормального развития зародыша в это время связаны с нарушением
плаценты. Преобладающей формой нарушений являю тся расстройства
кровообращения.
Характерной особенностью повреждений зародышевого развития
служит их однотипность, проявляющ аяся в том , что действие
различных тератогенных факторов приводит к сходным отклонениям.
Однотипность аномалий объясняется общ ностью молекулярных основ
нарушения роста, дифференцировки и морфогенеза.
6.5.7. Внутриутробное развитие человека
Пренатальное развитие человека можно разделить на- начальный
период (1-я неделя развития), зародышевый период (2— 8-я неделя
развития) и плодный период, завершающийся рождением ребенка.
Возраст плода при рождении составляет 266 ± 7 дней.
После овуляции примерно на 14-й йень с момента начала последнего
менструального цикла яйцеклетка попадает в маточную трубу, где
спустя 24 ч происходит оплодотворение. В течение первых 3 сут
развития оплодотворенное яйцо перемещается по трубе, проходя при
этом стадию дробления. Н а 4-й день образуется бластоциста (см . рис.
95), которая на 6-й день после овуляции прикрепляется к поверхности
слизистой оболочки стенки матки, а на 7-й день — имплантируется в
слизистую оболочку. Н а этой стадии на вентральной стороне
зародышевого диска различима энтодерма в виде тонкого слоя клеток,
появляется полость амниона.
В это время питание зародыша зависит о т вещ еств, проникающих иэ
слизистой оболочки путем диффузии. Н а
14-й день начинается
образование'плаценты и в то же время образуется желточный мешок.
Примерно на 1б-й день появляется первичная полоска и начинается
формирование зародышевой мезодермы. С 16-го по 21-й день образу­
ется нервная пластинка, которая превращается в нервную трубку.
Одновременно происходит сегментация мезодермы на сомиты, а во
внутризародышевой мезодерме образуются полости перикарда и внутризародышевый целом. Возникаю т закладки сердца, щитовидной
железы, пупочных сосудов. Питание зародыша все в большей степени
зависит от веществ, поступающих из крови матери.
Зародышевый период характеризуется быстрым ростом (рис. 96),
дифференцировкой зачатков основных органов и формированием
внешних признаков. Зародыш в возрасте 3 нед имеет длину около 2 мм.
201
В нем насчитывается 10 сомитов,
нервная трубка замкнута почти на
всем протяжении вплоть до задне­
го мозгового пузыря и остается от­
крытой лиш ь на переднем и заднем
концах. В незамкнутой части нерв
ной трубки намечаются передний,
средний и задний мозговые пузыри.
Хорда еще не полностью отде­
лена от лежащей под ней энтодер­
мы. Передняя, средняя и задняя киш­
ка различимы. О т задней кишки от­
ходит аллантоис. Пупочные вены
и артерии следуют за аллантоисом
по ножке зародыша в плаценту.
К этому сроку сердце начинает соРис. 95. Зародыш человека
настадии
крагцаться. Между двумя слоями
бластоцнсты
боковой
пластинки
мезодермы
I зародышевый узелок: 2 трофоблает
образуется целом. Устанавливается замкнутое кровообращение
через желточный мешок и ножку зародыша. В ножке сомита обособ­
ляю тся скопления клеток, представляющие собой зачатки канальцев
предпочки.
Между 21-м и 32-м днями после овуляции длина зародыша составля­
ет 3,5 мм, число сомитов достигает 25 пар. В это время завершаются
ранние стадии развития сердца и устанавливается односторонний ток
крови через него. Хорош о видны зачатки рук, зачатки ног только начи­
нают развиваться, о т ч е т л и в о различима хвостовая почка. Появляю тся
3 пары висцеральных дуг. В нефротоме развиваются канальцы первич­
ной почки.
В возрасте около 31 дня после овуляции зародыш достигает 7,5 мм в
длину. На этой стадии хорошо различимы зачатки рук и ног. Каждый
зачаток руки можно разделить на кистевой и плечевой сегменты
В зачатке ноги дифференцировка выражена слабее.
В возрасте около 6 нед зародыш достигает в длину приблизительно
12 мм. На этой стадии можно различить 5 первичных отделов мозга.
Особенно больших размеров достигают зачатки переднего и промежу­
точного мозга, формируются тимус, паращитовидные железы. Идет
гистогенез пищеварительного тракта. В это время отчетливо выражена
половая дифференцировка гонад.
С 5-й и до конца 8-й недели (рис. 97) развитие человеческого зароды­
ша характеризуется ростом и дифференцировкой органов. Восьмине­
дельный зародыш имеет длину около 40 мм и массу около 5 г. В период
между 6-й и 8-й неделями общий вид зародыша изменяется, намечаются
черты лица и он начинает обнаруживать больше сходства с человеком.
Обособляется шея, голова становится круглой, возникают зачатки
наружного уха и носа. Глаза смешаются с боковых поверхностей
кпереди и сближаю тся, появляю тся веки. Конечности стано­
вятся длиннее, четко намечаются и оформляются все их отделы.
202
,
П о сл е д н яя
мен стр уац и я
Я й ц ен л ети а
Возраст после оплодотворения, нед
— 0,15 мм
Рис. 97. Плод человека.
а
8 нед; б
е - 16 нед.
9 нед, в
10 нед; г
11 лед; д
12 нед,
Рис. 96. Истинные размеры эмбрионов че­
ловека в зависимости от срока развития.
Хорош о развиваю тся пальцы, особенно на руках. Х во ст почти неза­
метен.
В переднем отделе мозга начинав ся рост больших полушарий.
Происходит дальнейшая дифференцировка секреторных компонентов
вторичной почки. Первичная почка в значительной степени дегенериру­
ет, но на протяжении части зародышевого периода, по-видимому,
функционируют обе почки. Начинается образование надпочечников.
К 8-й неделе завершается дифференцировка зачатков гонад в
мужские или женские половые железы, например в первых уже
отчетливо видны семенные канальцы.
К концу 8-й недели завершается зародышевый период развития.
Практически все основные структуры и системы органов дифференци­
рованы. Плодный период, начинающийся с 9-й недели, характеризу­
ется ростом структур, их дальнейшей дифференцировкой и началом
функционирования.
Родившийся ребенок некоторое время связан с находящейся в матке
плацентой при помощи пупочного канатика. Акушер, помогающий
при родах, обычно перерезает канатик, предварительно наложив на него
зажим, однако, если зажим не наложен, то пупочные кровеносные
сосуды сами сокращ аются, прерывая кровообращение между ново­
рожденным и плацентой. Пупочные сосуды лишены иннервации, но .•
просвет их быстро закрывается в результате сокращения мышечных
стенок в ответ на действие механических, химических и температурных
раздражителей внешней среды. Зарастание пупочных сосудов происхо­
дит в течение нескольких недель, а функционировать они перестают уже
через несколько минут после рождения.
Глубокие и резкие изменения со стороны сердечно-сосудистой и
дыхательной систем происходят после рождения. В то т момент, когда
ток крови по пупочным сосудам прекращается, плацента перестает
выполнять дыхательную и выделительную функции.
Первый вдох — необходимое условие начала постнатальной жизни.
Дыхание стимулируется действием недостатка кислорода и избытка углекислоты в крови на дыхательные центры головного мозга.
6.6. П О С Т Э М Б Р И О Н А Л Ь Н О Е Р А З В И Т И Е .
РО С Т О Р Г А Н И З М А
После выхода организма из яйцевых оболочек начинается
п о с т а м б р и о н а л ь н ре
(постнатальное)
разви­
тие.
Начальная его стадия или р а н н и й
п о с т н атальд ы й о н . т о г е н е з у животных с прямым развитием характеризу­
ется интенсивным ростом органов и частей организма, установлением
дефинитивных пропорций тела (рис. 98), переходом функциональных
систем ца режим взрослого организма.
Рассмотрим более подробно процесс роста. Р о с т проявляется в
прогрессивном увеличении массы и размеров организма. Х отя
названные показатели могут изменяться благодаря накоплению не­
органических веществ (рост скелета) или воды (увеличение массы
204
Рис. 98. Пропорции тела человека в эмбриогенезе и после рождения.
растения), рост в эмбриональном и раннем постнатальном развитии
происходит прежде всего вследствие прироста органического вещ ества
(протоплазмы).
У беспозвоночных рост нередко обусловливается увеличением
размеров клеток, сопровождающимся их пояиплоидизацией (см . 5.3.6).
Таким образом растут личинки насекомых, некоторые круглые черви.
Более распространен пролиферационный рост, в основе которого лежит
клеточное деление. Если каждая из образующихся в результате деления
дочерних клеток вновь вступает в митотический цикл, число клеток
возрастает в геометрической прогрессии:
Л *-2».
где N — количество клеток; п — номер очередного деления.
Ситуация, близкая к описанной, наблюдается в эмбриональном и
раннем постнатальном периодах индивидуального развития.
В других случаях одна из дочерних клеток делится, а вторая диффе­
ренцируется, что дает линейное нарастание числа клеток:
N„ — 2/1.
Э то , например, характерно для эпидермиса кожи. Пролиферация
здесь приурочена к внутренним клеточным слоям эпителиального
пласта, откуда дифференцирующиеся клетки смещ аются в поверхно­
стные слои, где ороговевают и гибнут.
В процессе индивидуального развития показатели роста меняются.
У многих животных рост приурочен к
определенным
стадиям
онтогенеза. Такой тип роста называется о г р а н и ч е н н ы м или
о п р е д е л е н н ы м . В период активного роста скорость последнего
пропорциональна массе организма:
1Г = /*',
205
где \У— скорость роста единицы массы (скорость удельного роста);
к — коэффициент скорости роста; I — время.
Однако, начиная с определенного этапа, скорость роста, как
правило, снижается. Монотонное снижение скорости ровта у человека
прерывается в возрасте 13— 15 лет, соответствую щ ем периоду полового
созревания — п у б е р т а т н ы й с к а ч о к р о с т а .
Прирост в большей степени выражен в эмбриональных и мало­
дифференцированных тканях. Так, в первые 10— 12 сут после рождения
у морской свинки ежедневно прирост массы тела составляет 5 % ,
начиная с -17— 18 сут — 4 % , а с 45 с у т — 1%. П оэтом у рост
больш инства организмов описывается кривой 5-образной формы.
С ущ ествую т организмы, например рыбы, которые растут на
протяжении всей жизни. Н о и у этих животных по достижении полово­
зрелого состояния скорость роста уменьшается. Такой тип роста
называю т н е о г р а н и ч е н н ы м или н е о п р е д е л е н н ы м . *
Рост целого организма представляет собой интегральную величину
и слагается из множества частных проявлений роста. Пространственная
организация роста отличается сложностью , но носит закономерный
характер. Для каждой стадии онтогенеза можно выделить свои
градиенты роста разных частей организма. О т нее в значительной мере
зависят видовые особенности формы организмов.
-— Показатели роста, с одной стороны, к о н т р о л и р у ю т с я
г е н е т и ч е с к и , а с другой — з а в и с я т
от условий, в
которых происходит развитие. Довольно просто устанавливается
зависимость роста от питания. В «голодные» годы наблюдается
задержка роста. Э то заключение иллю стрирую т, например, данные по
динамике массы и длины тела школьников г. Ш тутгарта за период с
1911 по 1953 г. Если с 1920 по 1940 г. отмечалось увеличение названных
показателей, то в посление годы второй мировой войны выявилась
обратная тенденция. Перемена условий питания в лучш ую сторону
обычно сопровождается интенсификацией роста, и организм как бы
«наверстывает упущенное». Показано влияние на рост и развитие
людей в дорепродуктивной стадии постнатального онтогенеза всего
комплекса социально-экономических факторов, окружающих ребенка,
включая жилищ ные условия, долю расходов на ребенка в семейном
бюджете.
О наличии генетического контроля роста говорит прежде всего то т
факт, что размеры организма служ ат видовым признаком. Для людей
показаны определенные половые и расовые отличия в реакции на
изменение условий жизни, влияющих на развитие. Так, рост девочек
более устойчив к недоеданию, чем рост мальчиков. Длительное
голодание в детстве приводит у европейца к уменьшению длины тела
без нарушения пропорций. У японца в описанных условиях происходит
относительное удлинение туловищ а и укорочение ног, а у афри­
канца — наоборот.
П ути генетического контроля роста разнообразны. Важное значение
в опосредовании влияния генотипа принадлежит гормонам, особенно
вырабатываемым гипофизом, щитовидной и половыми железами.
Больш ая заслуга в разработке вопросов влияния эндокринных желез на
206
рост и развитие организма в постнатальном периоде принадлежит
М . М . Завадовскому (1891— 1957)'.
В
последнее
время обнаружен особый класс химических
соединений — регуляторов клеточной пролиферации и роста, среди
которых привлекают внимание к е й л о н ы . Последние являю тся
естественными тканеспецифичными ингибиторами митотического де­
ления. Белковое вещество ф а к т о р р о с т а н е р в о в специфиче­
ски интенсифицирует пролиферацию клеток-предшественниц нейронов
спинномозговых и симпатических узлов, а такж е рост зрелых
симпатических нервных клеток. Обнаружены такж е факторы роста
эпидермиса, кроветворной ткани.
6.7. С Т А Р О С Т Ь И С Т А Р Е Н И Е .
С М ЕРТ Ь КА К БИ О Л О ГИ ЧЕС КО Е Я В Л ЕН И Е
С т а р о с т ь - представляет собой закономерную стадию индиви­
дуального развития, по достижении которой организм приобретает
определенные изменения во внешнем виде и физическом состоянии.
С тарость наступает в пострепродуктивном периоде онтогенеза,
однако начало угасания репродуктивной функции или даже ее полная
утрата не могут служ ить нижней границей старости. Так, менопауза у
женщины определяет окончание репродуктивного периода ее жизни.
Вместе с тем к моменту достижения менопаузы больш инство внешних и
внутренних признаков далеко не достигают уровня типичного для
старых людей. С другой стороны, многие изменения, выраженные в
старости, начинаются задолго до снижения репродуктивной активно­
сти. Э то относится как к физическим признакам (поседение волос), так и
к функциям отдельных органов. Например, у мужчин снижение
секреции мужских половых гормонов гонадами и повышение секреции
гонадотропных гормонов гипофиза, что характерно для старого
организма, начинается вскоре после 25 лет.
Наблюдения в лабораторных условиях показали, что яйцекладка
может снижать продолжительность жизни. У плодовых мух, например, %
девственные самки живут дольше самок, участвующ их в спаривании.
Аналогичная закономерность обнаружена в отношении мышей.
Интересны в этом плане данные, касающиеся человека. Одинокие в
целом ж ивут меньше, чем вступившие в брак, однако овдовевшие и
разведенные характеризуются, в среднем, меньшей продолжительно­
стью жизни, чем одинокие.
Различаю т х р о н о л о г и ч е с к и й и б и о л о г и ч е с к и й
( ф и з и о л о г и ч е с к и й ) в о з р а с т . Согласно современной клас­
сификации, основанной на анализе средних показателей состояния
организма, людей, хронологический возраст которых достиг 60—74 лет,
называю т пожилыми, 75— 89 лет — старыми, свыш е 90 лет — долго­
жителями. Точное определение биологического возраста затруднено
' М. ,М . Завадовский был инициатором создания кафедр биологии в медицин­
ских вузах и первым профессором кафедр общей биологии I и I I Московских меди­
цинских институтов.
207
тем, что отдельные признаки старости появляю тся в разном
хронологическом возрасте и характеризуются различной скоростью
нарастания. Кроме того, изменения даже одного параметра, коррелиру­
ющего с возрастом, подвержены значительным половым и индивиду­
альным колебаниям. Так, если исходить из такого показателя, как
упругость кожи, то один и то т же биологический возраст достигается
женщиной примерно в 30 лет, а мужчиной в 80. С целью определения
биологического возраста, что необходимо для суждения о скорости
старения, пы таю тся использовать « б а т а р е и т е с т о в » — сово­
купность многих характеристик, закономерно изменяющихся в процессе
жизни. Основу таких «батарей» составят, по-видимому, сложные
функциональные показатели, зависящие о т согласованной деятельности
нескольких систем организма. В этом убеждают результаты поисков
тестов старения. Например, скорость прохождения нервного импульса,
которая зависит от состояния нервного волокна, снижается в интервале*
20— 90 лет на 10% , тогда как жизненная емкость легких, определяемая
координированной работой нервной и мышечной систем, — на 50% .
Состояние старости в биологическом понимании достигается
благодаря изменениям, составляю щ им содержание п р о ц е с с а
с т а р е н и я . Э ти изменения распространяются на все функцио­
нальные системы и обнаруживаются на любом из уровней структурной '
организации особи — молекулярном, субклеточном, клеточном, ткане-"
вом, органном. Суммарный результат многочисленных частных
проявлений старения на уровне целостного организма заклю чается в
нарастающем с возрастом снижении жизнеспособности особи, умень­
шении эффективности адаптационных, гомеостатических механизмов.
Показано, например, что молодые крысы после погружения в ледяную
воду на 3 мин восстанавливаю т температуру тела примерно за 1 ч.
Ж ивотны м среднего возраста на это требуется 1,5 ч, а старым около
2 ч. В целом это приводит к прогрессивному повышению в процессе
старения вероятности смерти. Таким образом, биологический смысл
старения заклю чается в том , что оно делает неизбежной с м е р т ь 1/
Последняя же представляет собой универсальный механизм ограничен
ния определенным пределом участия многоклеточных организ­
мов* в репродукции себе подобных. Без смерти нё было бы смены
поколений — одной из главных предпосылок эволюционного процесса
(см. главу 4).
К снижению биологических возможностей организма с возрастом
приводят изменения отнюдь не каждого показателя. У человека и
1 Наступлению биологической смерти нередко предшествует состояние к л и н и ­
ч е с к о й с м е р т и , в котором клетки и ткани сохраняют достаточный уровень
жизнеспособности, чтобы организм с помощью определенных воздействий мог быть
возвращен к жизни ( р е а н и м а ц и я ) .
Вопрос о потенциальном бессмертии и отсутствии старения у простейших,
поставленный на заре экспериментальной геронтологии, требует дальнейшей разра­
ботки. Наблюдения над амебами, размножающимися бесполым способом, показали,
что выживание клеточной линии, образованной исходно одной клеткой (клона кле­
ток), зависит от условий культивирования. Клон инфузорий, для которых типична
смена бесполого размножения и полового процесса, постепенно утрачивал способ­
ность к последнему и вымирал.
208
многих высших позвоночных в процессе жизни приобретается опыт,
умение избегать потенциально опасных ситуаций. Интересна в этом
плане и система иммунитета. Х о тя в целом ее эффективность после
достижения состояния зрелости снижается, благодаря «иммунологи­
ческой памяти» по отношению к некоторым инфекциям старые
животные могут оказаться более устойчивыми, чем молодые.
Скорость нарастания и выраженность изменений в процессе
старения находятся под генетическим контролем и зависят о т условий, в
которых происходило предшествующее развитие особи.
В пользу г е н е т и ч е с к о г о
контроля
старения
говорит то, что максимальная продолжительность жизни является
видовым признаком.
У человека выявлена положительная корреляция между длительно­
стью жизни потомков и родителей, особенно матери. Величины
продолжительности жизни у однояйцевых близнецов более близки, чем
у двуяйцевых. Попарные различия составили в среднем 14,5 года для
первых и 18,7 года для вторых. Описаны наследственные болезни с
ранним появлением изменений, обычно наблюдаемых у старых людей.
Так, при синдроме Хатчинсона—Гипфорда (инфантильная прогерия или
преждевременное старение в детском возрасте) уж е на первом году
жизни отмечается задержка роста, рано начинается облысение, на коже
появляю тся морщины, развивается атеросклероз. Половая зрелость,
как правило, не достигается, а смерть наступает в возрасте до 30 лет.
Указанный синдром характеризуется аутосомно-рецессивным типом
наследования.
В лабораторных условиях получены инбредные линии плодовой
мухи и мыш и, существенно различающиеся по средней и максимальной
продолжительности жизни. Гибриды 1-го поколения от скрещивания
короткоживущих инбредных родителей ж ивут гораздо дольше. Мухи
Е>го8орЫ1а те1апова$1ег, гомозиготные по аллелю зачаточных крыльев,
имеют меньшую продолжительность жизни, чем мухи дикого типа.
Потомки от скрещивания м утантов с мухами дикого типа по
рассматриваемому показателю отличаю тся единообразием и близки к
последним. Среди гибридов 2-го поколения от скрещивания таких
потомков между собой наблюдается расщепление по признаку
продолжительности жизни в отношении 3 (дикий тип) к 1 (мутантный
тип). Описанный опы т указывает на прямой генетический контроль
продолжительности жизни плодовых мух со стороны локуса, контроли­
рующего развитие крыльев. Первичное звено биохимических нарушений
в данном случае не установлено. К ак у дрозофилы, так и у мыш и описа­
но много примеров влияния отдельных генов на продолжительность
жизни животных.
О пы ты , имеющие цель вы яснить в л и я н и е н а п р о ц е с с
с т а р е н и я у с л о в и й ж и з н и , дали в целом положительный
ответ, однако вскрыли противоречивый характер этих влияний.
Средняя продолжительность жизни увеличивается у некоторых беспоз­
воночных (планария, дафния, коловратки) и позвоночных животных
при ограничении пишевого рациона. В одном из опытов крысятамотъемыш ам скармливали лиш ь 46% обычного рациона. Такие
209
животные жили дольше контрольных крыс и у них позже развивались
болезни, типичные для состояния старости, например дегенерация
скелетных мыш и. Вместе с тем увеличение продолжительности жизни в
описываемом опыте происходило вследствие удлинения периода до
наступления половой зрелости (ювенильного) при сохранении обычной
длительности периода зрелости. Вводя животным преднизолон (синте­
тический аналог адренокортикостероидов — гормонов коры надпо­
чечников), получали двукратное увеличение продолжительности жизни
короткоживущих инбредных мышей и более позднее развитие
некоторых биохимических и физиологических изменений, связанных с ч
возрастом. Одновременно происходила задержка роста на 2 5 % . При
введении преднизолона мыш ам долгоживущей линии увеличения
продолжительности жизни не отмечалось. В опытах на плодовых
мухах продолжительность жизни увеличивали, выращивая личинки при
повышенной плотности популяции или в условиях более высокой
температуры. В первом случае имаго отличались меньшими, а во
втором — большими по сравнению с контролем размерами тела.
В обоих случаях, однако, происходило замедление развития на
доимагинальных стадиях. Общий вывод о влиянии условий жизни
заключается в том , что факторы, замедляющие развитие, способству­
ю т увеличению продолжительности жизни. С этим выводом согласу­
ю тся наблюдения о существовании положительной корреляции между
‘
продолжительностью жизни и длительностью периодов беременности и
достижения половозрелого состояния (табл. 21).
Таблица
21.
Зависимость продолжительности жизни от длительности
периодов беременности и достижения половой зрелости для различных млеко­
питающих
М акси м альная про­
д о лж и те л ьн ость
ж и з н и , м ес
О р га н и з м
Человек
Индийский слон
Лош адь
Шимпанзе
Бурый медведь
Д омаш няя собака
Домашний крупный
гатый скот
Макака-резус
Кошка
Свинья
Саймири
Овца
Коза
Серая белка
Европейский кролик
М орская свинка
Домашний кролик
Золотистый хомячок
Мышь
210
Д л и те л ьн о сть
беременн ости,
м ес
В о з р а с т наступ лен ия
половой зрелости ,
мес
1380
9
144
840
21
156
744
11
12
534
8
120
442
7
72
408
2
7
360
9
348
5 .5
36
336
2
15
324
4
4
252
5
36
240
5
7
216
5
7
180
1 .5
12
156
1
12
90
2
2
56
0 ,7
2
48
0 ,5
2
42
0 ,7
1 ,5
ро­
6
Возраст
М ужнины
9 0 и выше
8 5 -9 0
8 0 -8 5
7 5 -8 0
7 0 -7 5
6 0 -6 5
5 5 -6 0
5 0 —55
4 5 -5 0
4 0 -4 5
3 5 -4 0
3 0 -3 5
2 5 -3 0
2 0 -2 5
1 5 -2 0
10-15
5 -1 0
5
Ж енщ ины
В озр аст
\
1
У
- й
т Ц
—
,1
-
/
л
'/ А
'/ /
,1
А
У
У / У/ УУ
У 'у У//,
■
1
'//а У/а.
у .
У ' 1,
у
У/У А А А,
у
А/Л
А '' Л
т
[■
у
ш У/А У/ ■ / У//,
У/а у / А/У
у Уа/, А/А
1
/У/ -АА ц
У/
V- У/7,
Миллионы 4
'
3
2
1
1
У / ■' У / , А
У/
У/
0
1 2
3
М уж чины
-
4
Миллионы
Ж енщ ины
-
—
9 0 и выше
8 5 -9 0
8 0 -8 5
7 5 -8 0
7 0 -7 5
6 5 - 7 0 ____
6 0 —6 5
5 5 -6 0
5 0 -5 5
.4 5 -5 0
4 0 -4 5
3 5 -4 0
3 0 -3 5
2 5 -3 0
2 0 -2 5
1 5 -2 0
10-15
5 -1 0
5
УУ
— -
[/ .,
-
]
Л /уУ '
Ул
1
А
У/У
'/ ; ;
уУ'У"
\
к
:
/7 .
>, .
_
1
’-
У
Г- •
у/,' У у
1/У/, ;
//<
'У А
1
,1
1
')
'/у
4
3
2
1 0
1 2
3
4
Рис. 99. Возрастной состав населения (X X столетие)
а
-- в н а ч а л е ; А
в середине сто л е тн я.
Учиты вая сложный характер влияния генетических и средовых
факторов на процесс старения, нелегко ответить на вопрос о том , как
долго может ж ить человек. Разные авторы называю т величины от 70 до
200 лет. По-видимому, истинная величина б и о л о г и ч е с к о й
п р о д о л ж и т е л ь н о с т и ж и з н и укладывается в эти пределы.
Если в расчетах исходить из соотношения длительности дорепродуктивного периода онтогенеза и продолжительности жизни, типично­
го для млекопитающих (5— 8-кратное превышение вторым показате­
лем первого), и принять длительность дорепродуктивного периода
человека за 20—25 лет, то биологическая продолжительность жизни
превышает 100 лет и даже приближается к 150— 200 годам. Если же
основываться на статистическом анализе показателей смертности в
разные возрастные периоды, то интересующая нас величина находится
в области 90 лет.
Улучшение социально-гигиенических условий жизни, качества
питания, успехи медицины обусловили существенный подъем в эконо­
мически развйтых странах средней продолжительности жизни в теку­
щем столетии (рис. 99). В настоящее время средняя продолжительность
жизни в экономически развитых странах составляет 71,1, а в развиваю­
щихся странах — 52,2 года. При этом женщины ж ивут в среднем на
3— 5 лет дольше.
6.7.1. Изменения органов и систем организма млекопитающих
и человека в процессе старения
Рассмотрим кратко изменения, происходящие в различных органах
и функциональных системах человека по окончании репродуктивного
периода (рис. 100).
К ак правило, после 50 лет у человека возникаю т стойкие внешние
проявления с т а р е н и я к ож и. П оявляю тся рубцы, пиг­
ментные пятна, бородавки и ро­
динки, а также морщины, кото­
рые образуются из-за потери
подкожного жира. Уменьш ается
число потовых желез, что делает
кожу более сухой и шершавой.
Происходят структурные изме­
нения коллагена кожи, приводя­
щие к снижению ее эластичности.
Кож а становится дряблой. Дру­
3 0 4 0 5 0 6 0 70 8 0 9 0
Годы
гие характерные внешние призна­
В о зр а с т
ки — это выпадение иЛи поседеРис. 100. Возрастные изменения жизненн и е ВОЛОС Н а ГОЛОВе. Поседение
в популяции начинается в возра­
ных функций человека.
сте менее 30 лет и не зависит от
исходного цвета волос.
Изменения в с и с т е м е п и щ е в а р е н и я
заклю чаю тся в
потере зубов, снижении уровня пищеварительных соков.
Начиная примерно с 40-летнего возраста, отмечается прогрессивный
подъем а р т е р и а л ь н о г о д а в л е н и я . Закономерные измене­
ния наблюдаются в стенках сосудов: в них откладывается холестерин и
происходят другие превращения, снижающие эластичность и искажаю­
щие реакции на различные стим улы. К ак правило, развивается
разрастание соединительной ткани.
В старости снижается интенсивность фильтрации в почечных
клубочках, а такж е обратное всасывание веществ из ф ильтрата в
почечных канальцах. Функциональные изменения в п о ч к а х про­
исходят не изолированно, а в тесной связи с изменениями в других сис­
темах, особенно сердечно-сосудистой и эндокринной.
Функциональные расстройства в д ы х а т е л ь н о й с и с т е м е
являю тся типичными для состояния старости. После 40-летнего
возраста наблюдается уменьшение жизненной емкости легких.
Изменения со стороны м ы ш ц носят распространенный характер.
Они регистрируются в гладкой, скелетной мускулатуре и сердечной
мышце. С возрастом снижается сила сокращений, быстрее наступает
утомление (например, скелетных мыш ц). М ыш цы становятся расслаб­
ленными и вялыми.
В приведенных выш е примерах такие показатели, как упругость
кожи и частичное поседение волос изменяются с возрастом довольно
монотонно. В отношении других (уровень артериального давления,
жизненная емкость легких, сила сокращения поперечнополосатой
мыш цы) изменения носят п о р о г о в ы й х а р а к т е р и начина­
ю тся примерно в возрасте 35—45 лет.
Сложный характер носят возрастные изменения о р г а н о в
э н д о к р и н н о й с и с т е м ы . Синтез гормонов гонадами, а также
щитовидной железой, надпочечниками снижается, тогда как некоторые
гормоны гипофиза, например гонадотропные, образую тся в большем
212
3
5
7
9
5
10
Эм бр и он ал ьны й
период, мес
15
20
25
30
35
40
90
П о стн атал ьн ы й в о з р а с т , годы
Рис. 101. Возрастные изменения тимуса человека.
количестве, при этом изменяются не только показатели синтеза
гормонов, но и чувствительность к ним клеток органов-мишеней.
В отношении гормонов шитовидной железы, коркового и мозгового
вещества надпочечников, инсулина этот показатель у старых людей
повышен.
С возрастом заметно изменяются и м м у н н ы е р е а к ц и и
организма. За гуморальный иммунитет ответственны В-лимфоциты,
образующиеся у млекопитающих, по-видимому, в костном мозге и
вырабатывающ ие антитела, специфичные по отношению к носителям
чужеродной генетической информации — антигенам. Образование Тлимфоцитов связано с вилочковой железой (тим усом ). Они ответ­
ственны за клеточный иммунитет, например за отторжение трансплан­
тата. Некоторые Т-лимфоциты участвую т в реакциях гуморального
иммунитета.
К старости отмечается ослабление реакций как гуморального, так и
клеточного иммунитета. Функциональные возможности иммунной
системы закладываются в связи с функционированием тимуса, который
подвергается обратному развитию по достижении зрелого возраста
(рис. 101). Нарушение хода развития этой системы путем удаления
тимуса, например у мышей, приводит к сокращению продолжительно­
сти жизни животных.
К старости, по-видимому, складываю тся условия, когда имм> окомпетентные клетки ошибочно вы рабатываю т антитела против
собственных антигенов организма. Такими ошибочно реагирующими
лимфоцитами могут бы ть мутантные клетки лимфоидной системы.
К ак известно, число мутантных клеток растет с возрастом. С другой
стороны, соматические мутации в клетках различных органов и тканей
могут настолько изменять их антигенные свойства, что на них
происходит выработка антител. Таким образом, старение сопровожда­
ется нарастанием аутоиммунных реакций.
Особенность н е р в н о й
системы
высших позвоночных
заклю чается в том , что предшественники нервных клеток прекращают
213
деление в конце эмбрионального или в крайнем случае в самом начале
постэмбрионального периода. Благодаря этому количество нервных
клеток в течение жизни организма может только снижаться.
Определение истинных масш табов гибели нервных клеток от рождения
до старости затруднено из-за несовершенства методов анализа.
В отдельных исследованиях показано, что за 100 лет жизни количество
грушевидных нейронов в мозжечке людей уменьшается на 2 5 % . Есть
данные о том , что у очень старых мышей в целом головном мозге нас­
читывается 2 млн. нервных клеток по сравнению с 5 млн. в головном
мозге молодых животных. В пользу гибели нервных клеток с возрастом
говорят наблюдения о снижении концентрации нервных окончаний,
а также нейромедиаторов в различных органах у старых животных и
человека. С труктура нервных клеток к старости закономерно изменя­
ется. Так, в их цитоплазме накапливается пигмент липофусцин (см . рис.
5, а), количество которого хорошо коррелирует с возрастом.
К функциональным изменениям, которые развиваются в процессе
старения нервной системы, относятся снижение памяти, нарушения
двигательной координации и др.
В старости наблюдается снижение функций практически всех
о р г а н о в ч у в с т в . Уменьш ается способность глаза к аккомода­
ции, так как слабеют глазодвигательные мышцы и изменяется
структура вещества хрусталика. Э то ведет к пресмиопии или старческой
дальнозоркости. У старых людей хрусталик часто мутнеет — развива­
ется катаракта. О строта зрения падает.
Снижается чувствительность органа слуха особенно к звуковым
волнам высокой частоты . Старый человек хуже различает запах и вкус.
У него нарушается чувство равновесия.
Таким образом в процессе старения изменяются функции всех
органов, однако зти изменения развиваю тся в организме неравномерно.
Более того, в одном и том же органе старого человека обнаруживаются
клетки с разной степенью выраженности изменений или вовсе без них.
6.7.2. Молекулярные и клеточные проявлений старения
Старение сопровождается выраженными молекулярными изменени­
ями структуры и функции генетического аппарата клеток разных тканей
и органов. Отмечается снижение содержания Д Н К и Р Н К , однако
химический состав Д Н К существенно не меняется. Изменяю тся физико­
химические свойства белков хроматина клеточных ядер, увеличивается
прочность связывания гистонов с Д Н К . Последнее может проявляться в
репрессии некоторой части генома.
При старении повреждаются все основные молекулярно-генетиче­
ские процессы — транскрипция и трансляция наследственной информа­
ции, репликация и репарация Д Н К . Э ти повреждения м огут бы ть
следствием неизбежных ошибок в ходе синтеза и преобразований
макромолекул (см . 5.3.2) или ж е бы ть результатом первичных
мутационных изменений. Например, синтез белков, контролируемый
мутантными генами, обусловливает дефектность их ферментативной
или иной функции.
214
Накопление в старости дефектов макромолекул сказывается на
функции клеток. Молекулярные изменения, совместимые с жизнью
клеток, могут тем не менее существенно повреждать их функцию и
снизить их вклад в физиологические процессы на тканевом, органном и
организменном уровнях.
Следует отм етить, что дифференцированные клетки разных
направлений специализации старею т в деталях по-разному. В нервных
клетках и поперечнополосатых мышцах, являющихся долгоживущими,
типично накопление липофусцина. В короткоживущих эритроцитах
этого не наблюдается и на первый план вы ступаю т изменения
плазматической мембраны.
Закономерный характер старения отдельных типов клеток и разную
продолжительность их жизни трудно объяснить с позиций накопления
случайных ошибок. Ряд исследователей связываю т старение с програм­
мой дифференцировки клеток. Предполагается, что дифференциро­
ванные клетки, осуществив до конца программу
специализации,
утрачиваю т способность возобновлять некоторые синтезы предшеству­
ющих этапов развития, что обрекает их на гибель.
Особо следует остановиться на изменении в процессе старения
способности клеток многоклеточного организма к делению. Показано,
что в условиях нормального развития потомки каждой клетки проходят
ограниченное число клеточных циклов. Так, фибробласты, синтезиру­
ющие коллаген, осущ ествляю т 50— 60 клеточных циклов, т. е. про­
должительность жизни клеточного клона ограничена. Неограниченной
способностью к пролиферации характеризуются лишь клетки с изме­
ненным наследственным аппаратом, например раковые.
6.7.3. Механизмы старения
Несколько сотен гипотез высказано относительно механизма
старения. Многие из них не выдержали проверку временем и пред­
ставляю^ в настоящее время исторический интерес. К ним относятся
гипотезы, объясняющие старение израсходованием особого ядерного
вещества, страхом смерти, утратой некоторых веществ, имеющихся
у каждого индивидуума в ограниченной дозе в момент оплодотворения,
самоотравлением продуктами жизнедеятельности. Гипотезы , сохраня­
ющие свое научное значение в наши дни, можно объединить в несколько
групп.
Согласно с т о х а с т и ч е с к и м гипотезам в основе старения
лежит накопление «ошибок» и повреждений, случайно (стохастически)
возникающих в процессе жизнедеятельности индивидуума на разных
уровнях его структурной организации. Сторонники различных вари­
антов таких гипотез подчеркивают первостепенную роль повреждения в
( генетическом аппарате (гипотеза накопления соматических мутаций)
или же относят первичные повреждения к широкому спектру
биологических молекул, вклю чая Р Н К , ядерные и цитоплазматические
белки, липиды клеточных мембран (свободнорадикальная гипотеза,
гипотеза повреждения структуры ферментов). Разного рода поврежде­
ния возникаю т вследствие «ош ибок» в ходе естественных внутрикле215
'
точных процессов или в результате действия внешних факторов.
Первичные изменения'на молекулярном уровне преобразуются в фун­
кциональные нарушения на более высоких структурных уровнях.
Согласно п р о г р а м м н ы м гипотезам старение детерминиро­
вано генетически, т . е. информация о начале и содержании его
представлена в геноме клеток. Одна из таких гипотез исходит из
признания связи между старением и клеточной дифференцировкой
(см . 6.7.2). В пользу запрограммированности старения говорит наличие
в природе видов, у которых вслед за размножением бурно нарастаю т
изменения, приводящие животных к гибели. Типичным примером
в этом отношении служат тихоокеанские лососи (горбуш а, нерка),
погибающие после нереста. Пусковой механизм в этом случае связан с
режимом секреции половых гормонов. Кастрированная горбуша не
нерестится и живет в 2— 3 раза дольше.
Программные гипотезы старения основываю тся на допущении, что
в организме функционируют своеобразные «часы », в соответствии с
которыми осущ ествляю тся возрастные изменения, механизм таких
часов точно не установлен. Предположительно, в его основе могут
лежать запрограммированное число делений в клоне клеток (см . 6.7.2)
или особенности функционирования в онтогенезе определенного
органа, например тимуса, который подвергается ранней возрастной
инволюции.
Генетические программы развития организмов представляют собой
результат эволюционного процесса. Э то распространяется и на
старение, если оно запрограммировано. Н а первый взгляд есте­
ственный отбор должен благоприятствовать увеличению продолжи­
тельности жизни, в связи с чем эволюционная обусловленность старе­
ния как механизма, создающего неизбежность смерти, кажется нело­
гичной. Противоречие преодолевается, если допустить, что старение
представляет собой косвенное, хотя и неизбежное следствие есте­
ственного отбора. Последний действует в направлении большей
приспособленности организма на ранних этапах онтогенеза, отодвигая
время проявления неблагоприятных аллелей. Учиты вая широкое
распространение явления плейотропии, легко представить, что один и
тот же ген может контролировать наряду с полезными признаками
(например, повышение плодовитости) такж е и вредные. Подобные гены
сохраняются и распространяются в генофондах популяций, если на
время нейтрализуются их отрицательные эффекты. Последнее достига­
ется, например, при помощи генов-модификаторов (см . 5.2.6).
Характерен пример с таким наследственным заболеванием, как
хорея Геттингтона (пляска святого В и тта), типичным симптомом
которого служит сильный тремор (дрожание) головы и конечностей.
Симптомы его проявляются обычно в возрасте 35— 39 лет, причем у
мужчин позднее, чем у женщин. Наблюдаемое различие в сроках
проявления болезни объясняю т тем, что у мужчин, имеющих в
сравнении с женщинами больш ую продолжительность репродуктивного
периода, давление отбора против соответствующ его признака угасает с
возрастом более медленно.
Рассмотренная гипотеза, объясняющ ая эволюцию старения,
216
привлекательна также тем , что она отвечает на вопрос о множествен­
ных проявлениях старческих изменений одновременно на всех структур­
ных уровнях организации, т. е. практически во всех тканях, органах и
функциональных системах. Действительно, отодвигая в онтогенезе
время проявления вредных генетических эффектов, естественный отбор
должен был их синхронизировать.
Х о тя проблема старения интересует науку давно, целенаправленные
фундаментальные исследования в этой области начаты сравнительно
недавно. Главны м стимулом работ в этом направлении служат
настоящ ая демографическая ситуация и прогноз изменения возрастной
структуры населения планеты на ближайшее будущее. Задача биологии
и медицины в такой ситуации состоит в поисках путей предупреждения
снижения функциональной активности и поддержания здоровья
человека, перешагнувшего возрастной рубеж 55—60 лет.
6.8. РЕГЕНЕРАЦИЯ
Основным свойством жизни является обмен веществ, который,
осущ ествляясь непосредственно на молекулярном уровне, представляет
собой основу постоянного обновления в процессе жизнедеятельности
структурных элементов организма. В условиях патологии или воздей­
ствия травмирующих факторов разрушительные процессы в клетках,
тканях и органах нередко интенсифицируются, однако и в этом случае
наблюдается та или иная степень восстановления структур. Сово­
купность процессов, направленных на восстановление снашиваемых
или
разрушаемых
биологических структур
называется
о егП г е т т а Т Г и е й Г
Восстановление структур, снашиваемых в процессе нормальной
Держание их морфофункциональных показателей,** обозначается как
ф и з и 6 л о г и ч е с к а я р е г е н е р я н п я Т Т я к в крови человека
циркулирует 25 •10 °
эритроцитов. Ежедневно замешается около 1%
этого количества, а каждые четыре месяца происходит полная смена
эритроцитов. Восстановление с т р у к т у р , разрушенных или утраченных
вследсцщ е.. да10Л^ических__^ме.н^НИЙ 1Ш 1 „. Зрад м ы , называется
репаративной регенепанией.
Проявления регенерации классифицируют такж е, исходя из уровня
структурной организации, на котором осущ ествляется восстановление.
В н у т р и к л е т о ч н а я р е г е н е р а ц .и я охаатьшает процессы
росстановления макромолекул и клеточных органедд,_Увеличение числа
органелл цитоплазмы достигается, в зависимости от их строения,
интенсификацией образования и сборки элементарных структурных
единиц (микротрубочки) или же путем деления сохранившихся органелл
(митохондрии). После выраженных изменений на восстановление
ультраструктурной организации клетки у млекопитающих требуется в
среднем примерно 4—7 сут.
1 Е с ть все основания включить в круг регенераторных процессов такж е восста-"
новление отдельных макромолекул, например Д Н К (см. 5.3.3). Ликцидацию молеку­
лярных повреждений обычно обозначают термином «репарация».
217
а
в
б
Рис. 102. Схема регенерационного процесса после удаления конечности у аксолотля,
а — н анесение т р а вм ы ; 6 — о б р азо ван и е б л а сте м ы ; в
м ис; 2 — дерма. 3 — м ы ш ц ы ; 4 — ко сть.
—
р еге н е р и р овавш ая к о н еч н о сть;
I
-
эпидер­
Восстановление на н а д к л е т о ч н ы х у р_од н я х (тканевый,
оргД1Ш Ш г_0 1 Я!Ш нш ленньЩ ^обётёчиваётся путем новообразования
клеток, внутриклеточной регенерации или сочетания обоих названных
механизмов.
У человека новообразование (пролиферация) клеток составляет ос­
нову физиологической и репаративной регенерации эпидермиса кожи,
эпителиального пласта кишки, кроветворной ткани. В нервной системе
в связи с утратой нервными клетками способности к пролиферации
восстановительные процессы осущ ествляю тся, по-видимому, исключи­
тельно путем интенсификации функций предшествующих клеток на
основе увеличения количества внутриклеточных структур (внутриклеточняг регенерация). В печени, почках, легких клетки сохраняют
способность к пролиферации, которая к том у же усиливается в случае
травмы органа. Вместе с тем в восстановительных процессах в
названных органах происходит и внутриклеточная регенерация.
Различаю т
несколько
способов
регенерации.
При
эпи.м о р ф..С1Д^е процесс регенерации идет от раневой поверхности.
Результатом его является восстановление недостающей части органа,
например конечности тритона в ее типичной форме (рис. 102).
В известном смысле эпиморфоз заклю чается как бы в достраивании
части до целого. При таком способе старые (культя) и новые(ретен?рат)
ткайй' разграничены достаточно отчетливо.
При м о р ф^ихл-а-к-е^ьсе оставш аяся после ампутации часть
органа перестраивается, превращаясь в целый орган, но м еньших
размеров. Д ля этого способа характерно не дополнение до целого
(надстройка), а перестройка—образование нового целого из остатков
старого. Разграничить культю и регенерат при этом не удается. Такой
способ наблюдается, например, в случае регенерации конечности
таракана.
Своеобразный
способ
регенерации
в
виде
регене­
рационной
гипертрофии
характерен
для
восста­
новления внутренних органов. В этом случае вслед за травмой
раневая поверхность заживает рубцом, удаленный участок не отрастает
и форма органа не восстанавливается. Вместе с тем благодаря
увеличению числа клеток и их размеров масса органа и его функцио­
нальный потенциал приближаются к исходным величинам. Таким
путем у млекопитающих регенерируют печень, легкие, почки, надпо­
чечник, поджелудочная, слюнные, щитовидная железы.
218
Рис. 103. Атипичная регенерация.
а — р егенерац ия у си к а вм е сто гл аза у
вм е сто кон ечн ости у ящ ериц ы.
р ак а; б
регенерация хвоста
Рис. 104. Различия в способности к регенерации у разных
животных.
а — п л а и а р н я; б — а к со л о тл ь ; в — л я г у ш к а .
В зависимости от способа регенерации клеточные изменения
сводятся к пролиферации, дифферендировке, увеличению размеров
клеток (гипертрофииУГТ1оследнее является морфологическим эквива­
лентом внутриклеточной регенерации.
Регенерация различается не только по масш табам и способам, но и
по конечному результату. Различие между исходным и регенериро219
вавшим органом может бы ть не только количественным, но и
качественным. В одних случаях регенерирует орган, который был
удален или разрушен — г о м о м о р ф о з, в иных на месте удаленно­
го органа развивается другой — г е т е р о м о р ф о з . Атипичная ре­
генерация встречается в основйом 'у беСПйзвбночных. Так, у рако­
образных вместо глаза возможна регенерация усика, а у пресмыкаю­
щихся — вместо конечноёти хвоста (рис. 103).
Объем и темпы регенераторного процесса в определенной степени
зависят о т условий, в которых этот процесс протекает. Стимулирующее
влияние на регенераций сказы ваю т гормоны некоторых эндокринных
желез, например щитовидной, гипофиза. Регенерация патологически
измененных органов имеет свои закономерности. Стимуляция
регенераторного процесса в таких случаях может приводить к обрати­
мости патологических изменений.
Эволю ция регенерационной способности. Физиологическая регене­
рация представляет собой процесс, свойственный всем ж ивым орга­
низмам.
М асш табы и способы репаративной регенерации существенно
варьирую т у представителей групп животных, различающихся система­
тическим положением (рис. 104). В ходе эволюции отдельных групп
организмов повышалась роль одних способов регенерации на фоне
снижения роли других. Изменялись и масш табы регенерации. Так,
амфибии обладают большей способностью к восстановлению типичной
структуры органов, чем круглоротые. У губок, кишечнополостных и
червей регенерация нередко осущ ествляется в полном объеме — из
части восстанавливается целый организм. Ряд биологов выделяю т это
явление в самостоятельное и называю т с о м а т и ч е с к и м
э м б р и о г е н е з о м , рассматривая его как вариант вегетативного
размножения. У червя планарии, например, целый организм восста­
навливается из 1/10 части исходного, а у гидры — из 1/ 200.
У членистоногих и моллюсков наблюдается регенерация отдельных
органов. Низшие представители хордовых способны восстанавливать
целый организм из его части (асш ш ии). Позвоночные в целом имеют
суженный масш таб регенерации путем эпиморфоза. Правда, представи­
тели амфибий и рептилий м огут восстанавливать отдельные органы,
например конечности, хвост. П тицы и млекопитающие восстанавлива­
ю т кожу, кости, мыш цы, внутренние органы. Восстановление способом
регенерационной гипертрофии, например, позволяет компенсировать
потерю 4/ 5 печени.
6.8.1. Трансплантация
Пересадка органов или тканей от одного животного другому
называю т т р а н с п л а н т а ц и е й . Организм, служащий источни­
ком пересаживаемого материала, называется д о н о р о м , организм,
которому пересаживают материал, — р е ц и п и е н т о м . Трансплан­
тируемый материал называется т р а н с п л а н т а т о м .
В зависимости от видовой принадлежности донора и реци­
пиента различаю т а л л о и гетеротрансплантацию
220
( к с е н о т р а н с п л а н т а ц и ю ). В первом случае реципиент полу­
чает трансплантат о т особи того же вида, во втором — от особи
другого. А у т о т р а н с п л а н т а ц и е й
называется пересадка
тканей в пределах одного организма.
В силу наличия механизмов иммунологической защ иты у высших
позвоночных и человека ксенотрансплантация заканчивается рассасыва­
нием трансплантата, а аллотрансплантация проходит успешно лишь у
однояйцевых близнецов, которые генетически идентичны. Применение
фармакологических препаратов, подавляющих иммунные реакции,
повыш ает вероятность успеха в случае аллотрансплантации органов.
Г V ШШ
6.9. Г О М Е О С Т А З В И Н Д И В И Д У А Л Ь Н О М Р А З В И Т И И
6.9.1. Понятие о гомеостазе. Роль нервной
и эндокринной систем в обеспечении гомеостаза организма
Ж ивой организм, будучи в энергетическом и вещественном плане
открытой системой, на любом этапе индивидуального развития
сущ ествует в единстве со средой обитания. При этом , несмотря на
определенные, иногда значительные колебания характеристик среды он
сохраняет себя во времени и пространстве как отдельную биологиче­
скую единицу, отличающ ую ся постоянством морфологии, основных
функциональных и поведенческих характеристик, физико-химических
параметров клеток, тканевой жидкости, крови. Такое положение
создается благодаря приспособлению (адаптации) организма к услови­
ям жизни. В ответ на изменения этих условий, если они по степени вы­
раженности совместимы с жизнью , он реагирует собственными
изменениями. Биологический смысл последних заклю чается в устране­
нии или компенсации отклонений со стороны биохимических, функцио­
нальных, структурных и прочих показателей и восстановлении
должного уровня жизнеспособности. Результатом таких изменений
служит не только сохранение в определенных пределах постоянства
внутренней среды, но и поддержание целостности организма.
Свойство живых форм поддерживать постоянство своей внутренней
среды, а также главные черты присущей ему организаций Жёсмотря
на 'йШ ёнчивость параметров окружающей среды
называется
г о~мё о с т а з о м 1.
~
Рсн б ву гомеостаза составляю т механизмы, сложившиеся в процессе
эволюции и” 'поэтом у закрепленные генетически. Эффективность
механизмов гомеостаза Н5 многом' бпрёНеляется генотипами особей,
разнообразие которых в пределах генофонда вида объясняет индивиду­
альные особенности уровня структурно-функциональной стабильности
конкретных организмов, различия их нормы реакции на одно и то же
изменение окружающей среды.
'■Термин «гомеостаз» был предложен первоначально для обозначения свой­
ства постоянства параметров жидких сред организма. В настоящее время наметилась
тенденция к более широкому использованию этого понятия, распространению его на
такие характеристики, как генетическая конституция, структурные показатели и др.
22)
Так как генотип представляет собой систему генов, сбалансиро­
ванную по содержанию разнообразной биологической информации,
появление в организме чужеродной генетической информации оказыва­
ет, как правило, на жизнеспособность неблагоприятное действие.
В связи с этим важная роль в поддержании гомеостаза в целом
принадлежит механизмам, ограждающим организм от проникновения и
вмеш ательства в процессы жизнедеятельности такой информации.
Носителями чужеродной наследственной информации м огут бы ть
как внешние агенты (бактерии и их токсины, вирусы, клетки и ткани
других организмов), так и собственные мутировавшие соматические
клетки. Поддержание генетического постоянства внутренней среды
организма или состояния г е н е т и ч е с к о г о
гомеостаза
осущ ествляется при помоши неспецифических и специфич«.киХ"ХДИ!с:
твующих строго против конкретного чужеродного агента) защ итных
механизмов. К первым относятся, например, барьерные свойства кожи
и слизистых оболочек, антимикробные свойства лизоцима слю ны,
фагоцитоз. Вторы е представлены механизмами клеточного и гумо­
рального иммунитета, аллергическими реакциями.
Больш ое значение в обеспечении гомеостаза принадлежит также
механизмам, поддерживающим постоянство и целостность морфоло­
гической организации. Они составляю т основы с т р у к т у р н о г о
г о м е о с т а з а , и действуют на разных уровнях — молекулярном,
клеточном, тканевом, органном. Универсальными эффективными
механизмами такого рода являю тся физиологическая и репаративная
регенерация (см . 6.8).
Не менее важное значение в обеспечении гомеостаза принадлежит
способности
организма
поддерживать
постоянство
х и м и ч е с к о г о с о с т а в а и свойств жидкой, внеклеточной его
части внутренней среды организма (кровь, лимфа, тканевая жидкость,
омывающ ая клетки). Механизмы, при помоши которых решается
указанная задача, разнообразны. Не вдаваясь в подробности, которые
рассматриваю тся в курсах нормальной и патологической физиологии,
отметим, что сохранение в оптимальных пределах, например,
кислотно-основного состояния крови зависит от уровня метаболических
процессов в печени, почках, мышцах, газообмена в легких, буферных
свойств гемоглобина и др.
Х о тя в целях удобства изучения и выделяю т различные виды
гомеостаза (генетический, структурный, системы крови, газовый и др.),
в действительности механизмы, их обеспечивающие, функционируют в
тесной взаимосвязи друг с другом, причем некоторые из них являю тся
общими для нескольких видов гомеостаза. Так, поддержание постоян­
ства физико-химических свойств крови требует в дополнение к сохране­
нию кислотно-основного состояния, регуляции газообмена, а также
выделения в кровь и выведения из нее осмотически активных веществ.
Взаимодействие разных видов гомеостаза отражает системный
характер организации живых форм. Объединение частных гомеостати­
ческих механизмов клеток и органов в целостную приспособительную
реакцию организма достигается благодаря функционированию регуля­
торных интегрирующих систем — нервной и эндокринной. Нервная и
222
эндокринная регуляция различается по скорости ответа и "Времени
сохранения его по отношению к моменту действия раздражителя. Как
правило, относительно быстрые изменения состояния организма
обеспечиваются нервной системой. Гормональные влияния распростра­
няю тся на клетки и органы медленнее, но и сохраняются обычно более
длительное время.
•—«^Примером генерализованного ответа организма на необычные по
силе или продолжительности воздействия со стороны окружающей
среды, развертывающегося на основе тесного взаимодействия нервных
и эндокринных механизмов регуляции, может служ ить состояние
с т р е с с а (стресс-реакция), которое развивается в организме при
неблагоприятных жизненных условиях, когда возникает угроза наруше­
ния гомеостаза. Комплекс реакций в названном состоянии лишен
'специфичности,' т. е. в своих главных чертах не зависит от природы
стрессора-агента, спровоцировавшего эти реакции. При стрессе наблю­
дается изменение состояния больш инства систем организма (м ы ­
шечной, дыхательной, сердечно-сосудистой, пищеварительной), орга­
нов чувств, уровня кровяного давления, клеточного состава крови.
В своем конкретном выражении эти изменения соответствую т отдель­
ным видам гомеостаза. И тог указанных изменений заклю чается прежде
всего в повышении обшей сопротивляемости организма по отношению
к неблагоприятным Факторам! Так, искусственно поддерживая высокий
уровень стероидных гормонов, выделяемых в естественных условиях
при стресс-реакции, в эксперименте удавалось в значительной степени
снизить смертность при тяжелых отравлениях пестицидами и некото­
рыми медикаментами, избежать нарушения свертывания крови при
введении такого препарата, как фининдион, предотвратить омертвение
тканей надпочечников в результате воздействия канцерогенными
углеводородами или повреждения тканей сердца при введении избытка
широко используемого в кардиологических клиниках препарата дигитоксина.
В определенных ситуациях состояние стресса путем быстрой
мобилизации и концентрации сил организма способствует решению
таких жизненноважных задач, как спасение бегством или отражение
нападения врага, т . е. выступает как защ итная реакция в узком смысле.
Независимо от результата, достигаемого'благодаря стресс-реакции,
первичное ее звено заклю чается во взаимосвязанных и взаимообусловливаемых нейроэндокринных изменениях, которые собственно
и придают реакции в целом общеорганизменный характер. При так
называемом «соматическом стрессе», с помощью которого решается,
например, задача повышения обшей сопротивляемости организма, цепь
событий представляется следующим образом.раздражитель активиру­
ет синтез особого класса гормональных веществ — релизинг-факторов
(о т англ. ге1еа5е — высвобождать, выделять), которые образуются
секретирующими нервными клетками подбугровой (гипоталамической)
области головного мозга. Функция релизинг-факторов заклю чается в
том , >что они, воздействуя на клетки передней доли гипофиза,
усиливаю т образование ими тропных гормонов, стимулирующих ак­
тивность других эндокринных желез, например щитовидной, надпо-
Мечниковой. Одни из названных релизинг-факторов способствует повы­
шенному образованию гипофизом адренокортикотропного гормона
(А К Т Г ), который, в свою очередь, усиливает синтез стероидных
гормонов клетками коркового вещества надпочечников. Последние,
воздействуя на клетки различных органов, изменяю т их функцио­
нальное состояние в направлении повышения защитного потенциала
организма.
В случае «психического стресса», развивающегося, например, при
болевом воздействии, цепь событий, соответствуя на многих этапах
выш е описанной, вклю чает дополнительно изменения функционального
состояния коры больших полушарий, лимбической системы головного
мозга, симпатической нервной системы, клеток мозгового вещества
надпочечников с выбросом в кровь значительных количеств адреналина.
По сходному пути развиваю тся, по-видимому, события при стрессреакциях людей на эмоциональные воздействия. Повышение содержа­
ния в организме биологически активных катехоламинов, к которым
относятся адреналин и медиатор симпатических нервных клеток — иорадренапин, зарегистрировано у пилотов и космонавтов во время
полетов, спортсменов в ходе соревнований, студентов на экзаменах.
Примечательно, что состояние стресса у человека может возникать в
ответ иа положительные эмоциональные стимулы.
Гомеостатические механизмы, активируемые в состоянии стресса,
способны противостоять действию неблагоприятных внешних факто­
ров до определенного предела. В развитии стресс-реакции выделяю т
три стадии: 1) мобилизации защ итных механизмов или тревогй; 2)
повышения сопротивляемости организма; 3) истощения защитных
механизмов. Первые две стадии соответствую т сохранению состояния
гомеостаза, третья стадия наступает при чрезмерных по силе или
продолжительности воздействиях и заклю чается, по сущ еству, в срыве
механизмов гомеостаза и развитии патологических изменений.
6.9.2. Кибернетические основы гомеостаза
В наличии гомеостатических механизмов находит отражение то т
факт, что живой организм в функциональном плане и отношениях с
окружающей средой представляет собой сложную саморегулирующую­
ся систему. Принципы существования и развития таких систем изучает
кибернетика.
В соответствии с кибернетическими идеями, глубоко проникшими в
последнее время в биологию, организм следует рассматривать как
совокупность неслучайных элементов, закономерным образом взаимо­
действующих друг с другом. Благодаря определенности набора и
структурно-функциональных связей элементов изменения системы
(организма) на конкретные воздействия извне носят закономерный
характер и определяются законом поведения этой системы. Указанный
закон задает такой порядок или алгоритм изменения элементов,
который обусловливает достижение системой (организмом) известной
цели, например, сохранения постоянства внутренней среды или общих
черт структурной организации.
224
Принцип построения кибернетической системы воспроизведен на
схеме:
Система (2) —
совокупность элементов, удовлетворяю­
щих определенному закону поведения:
(5)
(Вход) . (1)
Управляющий
блок
^
Исполняющий
блок
Выход
(3 )
*
г_________________________________________
О братная связь (4)
Применительно к ж ивым системам переменные на входе системы
обозначают терминами причина, стимул, раздражитель, переменные на
выходе — ответ, эффект, реакция.
В основе функционирования кибернетических систем лежат
процессы передачи и обработки в соответствии с заданным алгоритмом
информации, поступающей на входе (1). При этом в поведение системы
(2) вносятся коррективы, характер которых вы является путем регистра­
ции во времени отклонений переменных на выходе (3) в зависимости от
переменных на входе. Информация, используемая для коррекции
поведения системы, подается на управляющий блок последней по
каналам обратной связи (4). Различаю т положительные и отрица­
тельные обратные связи. Регуляция состояния системы по каналу
положительной обратной связи приводит к изменениям, усиливающим
зависимость реакции от действия раздражителя, по каналам отрица­
тельной связи — к изменениям, уменьшающим эту зависимость.
Отрицательная обратная связь таким образом способствует восста­
новлению исходных параметров в случае их отклонения в результате
действия внешних факторов, поэтому именно эта связь лежит в основе
регуляции гомеостатических реакций.
Н а /йсеме, приведенной выш е, стрелками (5) показано влияние не
только управляющей части системы на исполнительную, но и обратная
зависимость управляющих механизмов от состояния исполнительной
части. Такая взаимозависимость легко прослеживается в живом
организме. Физиологические параметры дыхательной и сердечно­
сосудистой систем, например, находятся под постоянным контролем со
стороны нервной системы. Вместе с тем для полноценного функциони­
рования нервных клеток необходим достаточный уровень снабжения
головного мозга кислородом.
Общие принципы автоматического регулирования и управления,
разрабатываемые кибернетикой, приложимы к лю бым системам,
существующ им в неживой и живой природе. О тличительная черта
биологических систем, в частности организма, заклю чается в особой их
сложности. Интересно вспомнить, что в процессе прогрессивной
эволюции в ряду живых форм степень сложности организма возрастает
(см . 7.8.3). Э то не удивительно, так как следствием усложнения служит
повышение уровня надежности, устойчивости системы по отношению к
изменениям окружающей среды. Устойчивость, находящая свое
конкретное выражение в свойстве гомеостаза, обусловливается дубли­
рованием механизмов поддержания постоянства внутренней среды на
разных уровнях. Таким образом в природе воспроизведен кибернетиче­
ский принцип многоконтурности регуляции состояния системы.
К главным контурам регуляции постоянства внутренней среды
организма относятся в первую очередь клеточные и тканевые
гомеостатические механизмы, в управлении которыми значительная
роль принадлежит генетическим факторам, местным рефлекторным
(нервным) влияниям, химическим и контактным межклеточным взаи­
модействиям. В связи с этим им свойственна высокая степень
автоматизма. Вклад клеточного и тканевого уровней в гомеостаз
организма осущ ествляется путем изменений показателей внутрикле­
точных синтезов, соотношения процессов пролиферации и клеточной
дифференцировки.
Ещ е один из главных контуров регуляции соответствует уровню
функциональных систем организма. Регуляторный компонент здесь
представлен сегментарными центрами спинного мозга и ядрами ствола
головного мозга, гормонами желез внутренней секреции. К этому
уровню относятся механизмы, обеспечивающие отдельные виды гомео­
стаза.
В следующий контур регуляции гомеостаза включены высшие
отделы центральной нервной системы, в частности кора больших
полушарий.
Наличие нескольких контуров регуляции постоянства внутренней
среды позволяет, во-первых, в зависимости о т жизненных условий
ограничить приспособительные изменения какой-либо одной системой
или же осущ ествить генерализованный ответ. Во-вторых, в этом случае
появляется возможность не только достичь состояния равновесия
с окружающей средой (за счет гомеостатических механизмов первых
двух уровней), но и активно преодолевать неблагоприятные условия,
избегая или устраняя их (условнорефлекторная деятельность).
6.9.3. Гомеостатические механизмы в процессе индивидуального
развития
Будучи необходимой предпосылкой совместимых с жизнью
отношений организма и окружающей среды, состояние гомеостаза
поддерживается на любой стадии индивидуального развития особи.
Вместе с тем в процессе онтогенеза меняются как сам организм, так и
условия его существования. Соответственно наблюдается смена
преобладающих гомеостатических механизмов.
Эмбриональное развитие многоклеточных животных протекает, как
правило, при более или менее выраженной изоляции от непосред­
ственного действия факторов внешней среды. Способы такой изоляции %
различны. У яйцекладущих наземных позвоночных изоляция связана с
образованием третичных яйцевых оболочек, у млекопитающих — с пе­
реходом к внутриутробному развитию . У представителей названной
группы животных в ходе эмбриогенеза образую тся провизорные
органы, такие как амнион, аллантоис. Благодаря этим органам
226
создается как бы собственная среда развития зародыша, опосредующая
и смягчающ ая внешние воздействия.
Важ ная особенность эмбриогенеза состоит в закономерном
чередовании периодов большей и меньшей устойчивости развития
к действию внешних по отношению к зародышу факторов. В периоды
относительной неустойчивости, называемые также критическими
(см . 6.5.5), даже слабые стимулы вы зы ваю т существенные изменения
хода формообразовательных процессов. Напротив, в периоды устойчи­
вости сильные воздействия скорее приводят к гибели зародыша, нежели
к отклонениям в развитии. Чередование периодов устойчивости и
неустойчивости, отмеченное выше, обусловливает специфику гомеоста­
за организма в эмбриогенезе. Наличие критических периодов учитыва­
ется в акушерской практике при составлении рекомендаций беременным
женщинам.
К ак уже отмечалось, в момент рождения происходит радикальная
смена образа жизни организма. Н а протяжении раннего постнатального периода, параллельно функциональному созреванию различных
систем организма, соответственно новым жизненным условиям вклю­
чаю тся эффективные гомеостатические механизмы. В этот период,
например, активно образуются клетки, обеспечивающие иммунологиче­
ский надзор — реакции клеточного (Т-лимфоциты) и гуморального (Влимфоциты) иммунитета. В ближайший после рождения отрезок жизни
(перинатальный период) развивается толерантность (нечувствитель­
ность) клеток, реагирующих на носителей чужеродной генетической
информации, к собственным клеткам и антигенам организма.
Состояния функциональной зрелости достигают главные регуляторные
системы — нервная и эндокринная.
Наибольшей надежности механизмы гомеостаза достигаю т в зре­
лом возрастном периоде онтогенеза. Старение организма сопровожда­
ется снижением эффективности механизмов генетического, структурно­
го и некоторых других видов гомеостаза, регуляторных влияний
нервной [й эндокринной систем. Э то обстоятельство в сочетании с
уменьшением резерва приспособительных возможностей больш инства
систем организма служит причиной падения в рассматриваемый
период онтогенеза общей сопротивляемости организма действию неб­
лагоприятных факторов окружающей среды.
6.9.4. Биологические ритмы
В эволюции выработалась способность организмов ориентиро­
ваться во времени, которая позволяет согласовывать скорость и нап­
равление главных физиологических процессов с закономерными
и прежде всего циклическими изменениями условий обитания. Меха­
низмы, лежащие в основе указанной способности, объединяют под
общим термином « б и о л о г и ч е с к и е ч а с ы ». Внешним проявле­
нием функционирования таких часов служ ат ритмические колебания
функций организма — б и о л р г и ч е с к и е р и т м ы Гр б л а сть био­
логии, изучающ ая закономерности временной организации живых
систем, называется х р о н о б и о л о г и е й .
Циклические изменения характеризуют различные процессы на
клеточном, тканевом, органном и организменном структурных уров­
нях. Так, с определенной периодичностью изменяется содержание
гликогена в клетках печени, количество клеток, редуплицирующих Д Н К
или делящихся митозом, происходит вы лет имаго из куколок у
плодовых мух или свечение одноклеточной водоросли Сопуаи1ах,
обусловливающее свечение морской воды. Многочисленны примеры
таких изменений у растений: поднимание и опускание листьев или
движение лепестков в зависимости от времени суток, опорожнение спор
из спорангиев у грибов и водорослей.
Биологические ритмы различаю тся продолжительностью цикла.
О к о л о ч а с о в ы е р и т м ы характеризуют временную организа­
цию некоторых внутриклеточных метаболических процессов, например
синтез и выделение белкового секрета клетками некоторых желез. Их
изучение начато сравнительно недавно. Изменения растений и
животных в связи со сменой времен года, издавна привлекавшие
с годовой
внимание людей, являю тся примером р и т м о в
периодичностью.
Интенсивно изучаю тся с у т о ч н ы е ( ц и р к а д н ы е ) р и т м ы ,
которые заклю чаю тся в закономерных изменениях физиологических
показателей организма в зависимости о т времени суток.
Суточные ритмы многих физиологических процессов являю тся
э н д о г е н н ы м и, т. е. определяются механизмами, действующими
в самом организме. В пользу этого говорит, например, сохранение
ритма, зависящего о т фотопериодичности, даже после помещения
организма в условия постоянного освещения. Так, мыш и, сущ ествуя
в течение нескольких поколений при постоянном освещении, по
возвращении в условия чередования света и тем ноты, воспроизводили
нормальную суточную периодичность двигательной активности.
Суточные ритмы реагируют на действие внешних факторов, прежде
всего чередование света и тем ноты, высоких и низких температур.
При этом изменяется положение фаз ритмических изменений. У челове­
ка, например, при переходе к образу жизни, противоположному
обычному (бодрствование ночью , сон днем), через 9— 10 сут наблюда­
ется смена фаз ритма колебаний температуры тела. Внешние факторы
способствую т выявлению эндогенных суточных ритмов путем синхро­
низации ритмических изменений отдельных клеток или особей.
Например, в популяциях плодовых мух, выдерживаемых в постоянных
условиях освещения, регистрируется непериодический вы лет имаго из
куколок. После воздействия светом благодаря синхронизации процесс
становится периодическим. Таким образом, внешние факторы могут
служить указателем времени.
Средняя длина периодов суточных ритмов у растений варьирует от
22 до 28 ч, у животных в большинстве случаев этот показатель уклады­
вается в пределы 23—25 ч. Сущ ествую т определенные индивидуальные
колебания длины периодов. При постоянных условиях длительность
цикла активности у четырех мышей составила в одном из опытов от
25,0 до 25,4 ч.
Эндогенные суточные ритмы ограничивают осуществление
228
иных функций определенным временем суток. Э то имеет большое
приспособительное значение, так как приводит организм в состояние
«готовности» по отношению к ожидаемым условиям среды в опреде­
ленное время. Так, вечерние прыжки лососей, требующие соответству­
ющего энергетического подкрепления, совпадают с максимумом
активности поедаемых насекомых. Благодаря эндогенному ритму
организмы сохраняют экологически целесообразную ориентировку во
времени суток, несмотря на периодическое выключение внешних
указателей времени, например в связи с непогодой.
Хронобиология представляет собой интенсивно развивающуюся
область науки, однако до сих пор нет отчетливого понимания
механизма биологических часов или способов сопряжения эндогенных
ритмов и циклических изменений внешних факторов. М ежду тем
познание указанного механизма имеет большое значение, например для
выбора оптимального режима активности человека. Так, ночная работа
в режиме «12-часовая смена, 24-часовой отдых» менее благоприятна,
чем многонедельная ночная работа, укладывающ аяся в суточный ритм.
Данные о суточном ритме клеточной пролиферации использую тся при
выборе времени назначения лекарств, действующих на делящиеся
клетки, например в онкологических клиниках.
Глава
7
В О П РО С Ы
эволюции
Эволюционное учение представляет собой фундаментальное научное
обобщение. Вот уже более ста лет оно оказывает решающее влияние на
состояние всех отраслей биологии. Под эволюцией понимают необра­
тимый, постепенный, закономерный процесс исторического развития
(филогенеза) природы. Эволю ция заключается в образовании приспо­
соблений, возникновении и исчезновении биологических видов, преобра­
зовании биогеоценозов и биосферы, при этом на отдельных ее этапах
появляю тся группы живых существ с ' более прогрессивными по
сравнению с предшественниками чертами структурно-функциональной
организации. В результате биологической эволюции на Земле сложи­
лись предпосылки к появлению социального существа — человека.
Ведущая роль в эволюции принадлежит процессу видообразования или
микроэволюции. Одновременно выделяю т «надвидовую» эволюцию
или макроэволюцию, представляющую собой историческое развитие
групп надвидового ранга — родов, семейств, отрядов. Историческое
развитие на уровне классов и типов иногда обозначают как
мегаэволюцию.
7.1. О Ч Е Р К П О И С Т О Р И И Э В О Л Ю Ц И О Н Н О Й Т Е О Р И И
7.1.1. Эволюционные представления в древнем мире
Представления о возникновении разнообразия живых форм в
результате саморазвития природы уходят корнями в глубь веков.
В древнеиндийских текстах Аюрвед, датируемых несколькими тысяче229
лети ям и до н. э ., указывается, что
человек произошел о т обезьяны . За
два ты сячел етия до н. э. в К и т а е суще­
ствовали учения, допускающие превра­
щения одних организмов в другие пу­
те м их постепенных изменений. М ы с л ь
о развитии всего многообразия неоду­
ш евленных и одушевленных природ­
ных объектов путем превращения
органического числа элементарных
начал неоднократно высказы валась
м ы слителям и древней Греции. Гера­
кл и т (конец IV — начало V века
до и. э.) рассматривал движение как
основу мира. Эм педокл ( V век до н. э)
считал, ч то все сущ ествую щ ее проис­
ходит из четырех начал, таких как
огонь, вода, воздух н земля, управляемь1х объединяющей и разъединяющей
силами — л ю б о вью
и ненавистью,
причем первыми из ж и вы х существ
появились растения, а затем ж и вотн ы е. Д ем о кри т ( V век до н. э.)
отмечал приспособление отдельных органов к вы п о лн яем ы м функциям.
А р и стотел ь (384— 322 годы до н. э .) указывал на черты сходства и
различия у отдельных ж и вы х сущ еств и составил классификацию расте­
ний и ж ивотн ы х . Ф . Эн гельс назвал м ы сли древних философов «гени­
альн ы м и догадками». Одновременно он подчеркивал их ум озритель­
ный, натурфилософский1 характер, поскольку зачастую они бы ли
плодом размыш лений и не опирались на данные систематических
наблюдений природы. Т аки м образом, представления мыслителей
древности отличались незавершенностью и вклю чали элем енты оче­
видной фантазии. П о этой причине б ы л о б ы неправильно рассматри­
в а т ь античных мыслителей как предшественников современных эволю
ционистов.
Аристотель ( I V
7.1.2.
век до
н э.)
Эволю ционная идея в биологии
конца X V I I — н а ча л а X V I I I веков
Период Средневековья характеризовался обш им упадком кул ьтур ­
ной жизни человечества. Расцвет искусства и наук в эпоху Возрождения
( X V — X V I век), послуживш ий началом рождения современных пред­
ставлений о природе, коснулся и биологии. Е стество и сп ы тател и этого
периода освоили огром ный фактический м атерил из области ботаники,
зоологии, анатомии. К X V I веку отн осятся первые м ноготом ны е
описания растительного и животного мира. В X V II веке У . Гарвей
создает учение о м алом и больш ом круге кровообращения, Г . Галилей
1 Натурф илософия
умозрительное истолкование природы взятой в целом
В натурфилософской оценке мири сочетаю тся фантастические представления с диа­
лектической идеей ра ш ития
230
конструирует микроскоп, а Р . Г у к и М . М альпиги о т к р ы в а ю т новое
направление в исследовании биологических стр уктур — микроскопиче­
скую ан атом и ю и цитологию. Вм есте с те м господствующ ее положение
в мировоззрении заняло учение об абсолю тной неизменности природы.
Считалось, ч т о современные виды растений и ж и вотн ы х установлены
раз и навсегда в м ом ент их возникновения. П ри это м подчеркивалась из­
начальная целесообразность природных процессов, строения и физио­
логических отправлений организма. Причину это го видели в сотворении
мира вы сш и м сущ еством — творцом , поэтом у само учение получило
название креационизма (о т лат. сгеайо— создаю, тво р ю ). Объяснение
приспособленности ж и вы х сущ еств к условиям жизни, гармонии в
природе высш ей м уд ростью творца, действующего с заранее наме­
ченной целью, составляет основу телеологического (о т греч. 1е1ео$ —
стремящ ийся к цели) взгляда на источник изначальных свойств
организма. Креационизм, о тр а ж а я идеалистические представления о
природе, неизменно поддерживался религией. Телеология ж е прямо
вела к теологическому (о т греч. 1ео5 — бог), божественному объяснению
органической целесообразности. Э т и концепции находили значительное
число приверженцев до середины X I X века.
Исследуя ж и вотн ы й и растительны й мир, те ла неживой природы и
сопоставляя их между собой, естествоиспы татели X V I I — X V I I I веков
отмечали различную сло ж но сть объектов изучения, в результате чего
б ы л сформулирован принцип градации и указано на наличие переходных
форм м ежду н еж и вы м и ж и в ы м , растениями и ж и во тн ы м и , ч т о нашло
отражение в «лестницах естественных тел». Т ак, Ш . Бонне (1720— 1793)
начинает та кую «лестницу» с «тонких материй», воздуха и воды через
м еталлы , камни, кораллы и далее грибы , растения, чувствую щ ие
растения, насекомых, червей, ры б, летаю щ их ры б, водных птиц и
просто птиц, л етучую м ы ш ь , четвероногих, обезьян доводит ее до
человека и продолжает через «ангельские чи ны » различных рангов до
бога. Д а ж е если отвлечься о т того, ч то теология находила прямое
отражение в «лестницах естественных тел», последовательность поло­
жения неодушевленных и одушевленных объектов, закономерное
расположение ж и вы х организмов в ней, не связы валось авторами с
м ы сл ью о развитии природы. Р е ч ь ш ла об общей конструкции
мира, созданного м уд ростью творца и с тех пор сущ ествую щ его в
неизменном виде. Такое понимание со ответствует метафизическим
представлениям, господствующ им в науке в X V I I I веке, которые
воплотились в создаваемых в это время искусственных классификациях
растений и животных.
7.1.3.
Эволю ционная идея у естествоиспытателей
и философов второй половины X V I I I века
П од влиянием научных ф актов и новых философских идей во второй
половине X V I I I века намечается ревизия креационизма в пользу эво­
люционной идеи. Э т о прослеживается в сочетаниях французских м ате­
риалистов. Ж . Л ам етр и (1709—-1751) в «Человеке-машине» отм ечает
сходство человека и ж и вотн ы х по происхождению и некоторы м чертам
строения. Д . Дидро (1713— 1784) в « М ы сл ях об объяснении природы»
■31
говорит о существовании в прош лом прототипа всех ж ивотны х,
трансформация признаков которого в естественных условиях и привела
к появлению современных форм. И л ея развития звучи т в э т о т период в
трудах ряда биологов. Ж . Бю ффои (1707— 1788) в «Естественной
истории» выдвигает положение об образовании ж и вы х сущ еств из
неорганических веществ и изменении их под влиянием среды, например
ландшафта. Сходство в строении ж и вотн ы х он объясняет общ но стью
происхождения. Эволю ционизм второй половины X V I I I века не
выходит однако за рамки трансформизма (о т лат. 1гап5 — пере,
Го гта — форма), которы й видит развитие органического мира в
простом превращении одних видов растений и ж и вотн ы х в другие.
Ограниченность трансформизма заклю чается в то м , ч то он лишен
понимания развития как исторического процесса, ко то ры й благодаря
закономерным движ ущ им силам обеспечивает определенную направ­
ленность органической эволюции, разнообразие ж и вы х форм, появле­
ние прогрессивных типов структурно-функциональной организации.
П оло ж и тельн о е значение трансформизма обусловлено его направлен­
н остью против креационизма, представлений о неизменяемости видов,
утверждением принципа развития природы под влиянием естественных
факторов.
7.1.4. Развитие эволюционной идеи в России в X V I I I веке
М атериалистические тенденции в объяснении мира и идея развития
нашли отражение в научной м ы сли России X V I I I века. Основоположник
русской науки М . В . Лом оносов (1711— 1765) утверждал первичность
материи, определение ею сущ ности природных тел и явлений.
В качестве основного свойства материи он назы вал движение, понимае­
мое как развитие. М атериалистичны идеи А . Н . Радищ ева (1749— 1802),
изложенные в тр акта те « О человеке, его смертности и бессмертии». Он
писал, ч го б ы ти е вещей независимо о т си лы познания о них сущ ествует
само по себе и что в это б ы ти е нельзя проникнуть иначе, как через опы т.
В построенной и м «лестнице вещ еств» А . Н . Радищ ев обходится без
божественных чинов. Ступени «лестн и ц ы » со о тв етс тв ую т значи­
тельн ы м этапам развития природы — превращению неорганических
веществ в органические, возникновению у ж ивых существ новых
качеств, таких как ощущение, мышление. Отражением идеи развития в
эмбриологии б ы л а концепция эпигенеза К . Ф . Вольф а (1733— 1794),
который в противовес метафизической теории префоризма утверждал,
ч то развитие зародыш а идет о т простого к сложному. Русски й ученый
К . Ф . Вольф обращ ал внимание на изменчивость ж и вотн ы х и растений,
причину которой он видел в действии климатических ф акторов и
особенностях питания.
7.1.5. Эволю ционная теория Ж . Б . Л ам арка
П редставления о развитии природы, изложенные вы ш е, зан им аю т
почетное место в истории естествознания. Д иалектико-материалисти­
ческие тенденции, прослеживающиеся в иих, иа протяжении веков
п ротивостояли метафизическому, идеалистическому и теологическому
восприятию мира. М ногие из них можно
г>ассматривать как гениальные предвидеПыя великих умов, однако они несли- в
'себ е ^ л н ш ь элем енты эволюционизма,
Никогда не поднимаясь до уровня эволю ­
ционной теории. П ричины эго го заклю ­
чались в малочисленности ф актов, которые с ост а вил й~~бьI фундамент такой Тбо1
рииГнёДОСтаточном оЬш ем уровне разви­
тия биоло гических и других есча,'шен^ных наук.____
Первая эволюционная теория б ы л а
создана в начале X 1X века Ж а н о м Б а ти ­
стом Д ам а ско м (1744— 1829). Н аряд у с
утверждением идеи развития и попыткой
обосновать ее фактическим материалом
из области зоологии и ботаники, Ж . Б . Л а ­
марк поставил вопрос о причинах и меха
низмах эволюционных преобразований.
ж 1 1 а м а р к (1744
1»29|
В своеЁГтёории он прежде всего о тм еча­
ет искусственный""характер таксономиче­
ских единиц. используемых биологами в классификациях органического
мира. П о его мнению в природе нет ни классов, ни родов, ни видов. О ни _
являю тся продуктами человеческого ума, созданными для упорядоче­
ния представлений о мире. Природа ж е создает то лько особей, т . е.
реально сущ ествует последовательный ряд сущ еств, для которого
характерны «постепенные переходы м ежду соседними видами и даже
родами». П ри чи ну таких переходов и непрерывности ряда в цело-уТ
Ж . Б . Л ам арк видит в изменчивости органи>мов. резул ьтаты которой
определяются двумя силами. ТЗо-первых, в природе действует принцип
градации, заклю чаю щ ийся в последовательном и непрерывном услож ­
нении и усовершенствовании ж ивых сущ еств в процессе развития
природы. Подобная градация не зависит о т влияния на организм
внешних условий и является следствием внутреннего стремления
оргаиизмслГ"к Совершенствованию. Во-вторых, на ж и в ы е существа
постоянно и случайны м образом д ействую т м ногообразные ф акторы.,
среды обитания, которые вм еш иваю тся в осуществление принципа гра­
дации и приводят к уклонениям о т непрерывного усовершенствования.
Ж . Б. Л ам арк пишет: «Теперешнее состояние ж и вотн ы х есть, с одной
стороны , следствие нарастаю щ ей сложности организации — сложно­
сти, стремящ ейся к правильной градации, а с другой — результат
влияния крайне многих и весьма различных внешних обстоятельств,
постоянно стремящ ихся наруш ить правильность градации в усложнении
организации». К приведенны м д вум м ом ентам, определяющим нанравления изменчивости организмов во времени, и сводится сущ ность
звБЛЮционной теории Ж. Ь. Ламарка, значительная рпдк н когопой —
отводится внешним условиям^ Влияние этих условий на ж ивотны х
осущ ествляется косвенным путём через первичное изменение п о требно­
стей и привычек, которое в ы з ы ва е т новые формы деятельности и как
следствие интенсивное упражнение одних органов и относительную
бездеятельность других. Упражнение органов стимулирует их развитие,
укрепление, увеличение, неупражнение — недоразвитие, уменьшение,
вп ло ть до исчезновения. Резул ь т а т о м этого с л у ж а т изменения ф ормы,
структуры _ организма, которые передаются по наследству и таки м
путем закрепляю тся в потом стве. Зависи м о сть состояния ор 1 днов о т их"
упражнения и сохранение изменений в п о т о м п в е пу тем ИХ Прямого
наследования известны как два^зайона Ж . Б . Л а м а р к а ~ Д л я низших
ж и вотн ы х и растений, где связь между потребностям и и привычкам и и
формами деятельности менее очевидна, Ж . Б. Л ам ар к допускал не­
посредственное влияние внешних условий.
Н епрерывны й ряд организмов в соответствии с принципом градации
напоминает «лестницу сущ еств» Ш . Бонне. Сходство здесь, одиако,
Видимое. О тличие заклю чается в то м , ч то по Ж . Б . Л а м а р ку — э т о ряд
последовательно развиваю щихся форм. Залож и в в основу это го ряда
принцип историзма, Ж . Ь . Л ам ар к внес в биологию совершенно новую
идею , в корне п од ры ваю щ ую метафизический '"взгляд на природу.
Им енно в это м заклю чается бол ьш ое значение теории Л а м а р ка.
Представления Ж . Б . Л ам арка о зависимости состояния и развития
органов о т внешних формирующих воздействий м о гут рассматри
ваться как стремление к научному, материалистическому объяснению
эволюции ж ивотн ы х . Н о э т о т материализм остается механистическим,
так как сводит сложное явление исторического развития к изменению
простых физиологических процессов. Вслед за многими предшественни
ками Л ам ар к подчеркивал целесообразность строения и проявлений
жизнедеятельности ж и вы х сущ еств, адекватность их* о твето в на
внешние ф акторы. Однако целесообразность в органическом м ирр
Лам арк пБт.Ягняп с т р е м л е н и е м Организмов к усоверш енствованию ,
которое рассматривал как изначальное, внутренне присущее свойство
ж ивых форм. В это м объяснении проявляется идеализм эволюционной
теории Ж . С Л ам арка.
7.1.6.
Эволю ционная теория Ч . Дарвина
Возникновению эволюционной теории Ч . Дарвина (1809— 1882)
сносов ствовали сон и ал ьно~экрномйчео<ие и естественнонаучны е пред­
посылки. К середине X I X века А н гл и я достигла значительных успехов в
развитии промыш ленности и средств сообщения, в интенсификации
сельскохозяйственного производства, селекции. В относительно ко­
роткий срок путем скрещиваний и браковки б ы л и выведены ценные
породы овец, свиней, крупного рогатого скота, кур, новы е сорта
культур н ы х растений. Росла уверенность в возм ож ности широкого
произвольного изменения ж и вы х форм. Го сп од ствую щ и е в т о время в
науке метафизические представления о неизменяемости ви дов вступали
в противоречие с новы м и ф актами. Д ля освоения колониальных
владении организовывались экспедиции, в состав которых вклю чались
биологн, привозившие богатейшие научные материалы о разнообразии
и особенностях ф лоры и ф ауны различных уголков земного шара.
В кругосветном плавании на экспедиционном корабле « Б и гл ь » , прод­
ливш емся 5 лет, принял участие Ч . Дарвин
В первые десятилетня X I X века нау­
ка, все еше находясь в идейном отнош е
ЙИИТюд~влиянием метафизических воз
даШТТТ; обогатилась рядом фундамен
И л ьн ы х " обобщении, способствовавших
утверждению эволюционной идеи. В а ж ­
ное место среди них принадлежит теории
Ч . Лайеля (1797— 1875) о медленном и
непрерывном развитии земной поверх
н бсти Т ю д ^ ёй дтви ем достоянных гг/июгйческихфакторов, таких как вода, ветер,
вулканы, остывание раскаленных недр.
Э т а теория исключала катастроф ы как
ведушйй<Ьакт о р эволю ции планеты , а
следовательно, и органического мира.
В области биологии значительных успе­
хов достигла сравнительная анатом ия
(Э . Сент-Илер, Ж . К ю в ь е , И . В . Гете),
бы ли заложены основы научной пале­
онтологии ( Ж . К ю в ь е ) и эмбриологии
( К . Бэр). Э л ем ен ты эволюционизма в э г о т период встречаются-Д-ТРУдах ряда крупных биологов. Т а к. по~Э7 С ент И л ср у выживание_вщ ю д
в прмропр ЭЯРИСИТ п т цх прцгппглбленности к среде. К . Б эр называет
вопрос происхождения органической жизни важ нейш им в биологии. Д л я
его решения он привлекает материал по изменчивости домашних
ж ивотных, данные по ископаемым формам, географическому распро­
странению животных. Встр еча ю тся отдельные вы сказы вания в пользу
значения естественного отб о ра в эволю ции организмов. Однако это
были разрозненные м ы сли, не прптнррж псиные ногтяточно~обосыо-.
ванными ф актами. Заслуга Ч . Д арвина состоит в то м , что он, с одной
стороны, дал систем атическое, полное~Для своего времени изложение
/Доказательств "эволюции, а с другой — предпринял успеш ную по'п ы тк у 1^ с к р ы т ь сущ ность эволюционного" процесса, показав его
^ /дви&ущйе силы Г
' ТГо Ч. Д арвину, в основе эволюции л еж и т взаимодействи е, та »щх
природных. ЯШ 1 СН11 .Й,
как:
1) изменчивость;
2) наследегвейность;
3) борьба за существование; 4) естественный отб ор; 5) расхождение
признаков.
Вслед "за некоторы ми предшественниками Ч . Дарвин обращ ает
внимание на вы раж енную изменчивость .ж и в ы х форм и о твод и т ей в
эволюции важное место. Новое, внесенное им в представления об
изменчивости, *заключается в разграничении ненаследственных и нас
ледственных изменений и утв<-ржлениепервостепённой~]рол~й в процессе”
Эволю ции последних. Т зс сМ Я тр и в а я вопрос о причинах изменений,
Ч . Д арвин вьщ еляет определенную и неопределенную изменчивость.
Неопределенными изменениями он называет те, которые п оявляю тся у
организмов внезапно, причём их направление у разных особей различно,
и в э т о м см ы сле они случайны. Им енно с таким и изменениями связана
эволюция, Определённая^ изменчивость в ы з ы ва ется воздействиями
известной природьП~Гакие изменения не наследуются.
‘
““
235
Ва ж н а я роль в эволюционном процессе отводится наследственно­
сти. Ч . Дарвин отм ечает распространенность этого явления, приводит
в д оказательство фактический материал, известный в его время. Вм есте
с тем ему не удалось вскры т ь законы наследственности. Опубликование
известного исследования^ Г Менделя в 1865 г. приш лось на период
активной раб оты Ч . Дарвина над теорией эволюции, однако оно
осталось неизвестным Ч . Дарвину, как и больш инству его современни
ков. Придавая больш ое значение неопределенным, случай н ы м измене­
ниям, Ч . Дарвин должен б ы л о бъясн и ть ф акт целесообразной приспо­
собленности организмов к среде обитания, закономерный характер
изменений строения ж и вы х суш еств в процессе эволюции., Возникнове
нне и закрепление новых разновидностей и видов при случайном
изменчивости Ч Г Д а рвин связал с явлением борьбы
за сущ ествование и естественным отбором^ Ъ о р ь б у за сущ ествование
Ч . Д арвин понимает в широком метафорическом см ы сле — это л ю б а я
зависимость одного организма о т другого или ж е о т условий
окруж аю щ ей среды. О н выделяет три формы борьбы за сущ ествование
Конституц иональная борьба зависит о т вариабельности ко н сти туц и и 1
организмов, ч то обусловливает разное соответствие их особенностям
действия абиотических факторов среды, например климата. М ежвидо­
вая борьба происходит м ежду организмами разных видов. Содержали ■
ем в н у т ривмппной борьб ы сл уж и т столкновение интересов особей
одного вида. Борьбу за сущ ествование " ^ . Д арвин Связывает со
стремлением организмов" разм нож ать с я в_геометр ической прогрессий.
Благодаря этой борьбе из огромного числа особёй~кажлш тт вида,
которое появляется на свет, до взрослого состояния дож ивает ли ш ь
незначительная часть, остальны е ж е погибаю т в результате жизненной
конкуренции, «бо р ьбы за ж и зн ь». Преим ущ ества в борьбе обусловлива­
ет не то лько сохранение ж изни осоОи. но и успех в оставлении потом
ства. Вследствие борьбы за сущ ествование в природе происходит
естественны й отбор. П од естественным отбором Ч . Д арвин понимал
« переживание наиболее приспособленных» или же избирательное
«сохранение полезных индивидуальных отличий или изменений и у нич­
т ожение вредных». Благодаря борьбе за существование и отоору в'
процессе эволюции накапли ваю тся полезны е,_биологически целесо­
образные особенности строения организма. Б о рьб а за существование и
происходят в природе постоянно и приводят к
появлению новых ж и вы х форм. Значительную роль в формировании
Ч . Д арвин ы м концепции естественного отбора сыграли его наблюдения
о значении искусственного отбора в происхождении пород домашних
животных.
П редставления об изменчивости, наследственности, борьбе за
существование и естественном отборе как ведущих движущ их силах
эволюции Ч . Дарвин дополнил указанием иа роль дивергенции,
1 К о н с т и т у ц и я ( о т л а т соп$1йи1ю
устройство)
совокупность морф ш оги
ческих и ф ункциональных характеристик организма
вы р а ж а ется в определенном
тине телосложения и других свойствах, например особенноетях реакций на внеш ­
ние воздействия
р
236
расхождения признаков. П о Ч . Дарвину виды с малой изменчиво стью в
Д щ у ограниченных "Возм ожностей приспособить ся к м еняю щ имся
условиям жизни обречены иа вы м и рание
Н апротив, наиболее
уклоняющиеся— от—среднего уровня ф орм ы - в пределах видов со
значительной изм енчивостью и м ею т высокие ш ансы на выживание
вследствие ослабления конкуренции и освоения разнообразных экологи­
ческих ниш . О собое значение принцип дивергенции имеет для
оК-кягнепия эволюции и надвидовом урбвнё~й~вбзнйкновения родов,
семейств, отрядов.
"
Эволю ционная теория Ч . Д арвина о ткр ы ла новую эру в биологии.
Она подорвала основы метафизических, телеологических и теологиче­
ских взглядов на природу и ее происхождение. Ф . Эн гельс отм ечал, что
Дарвин « ... нанес сильнейший удар метафизическому взгляду на
природу, доказав, ч то весь современный органический мир, растения и
ж и в о тные, а. елеловатёЛТ.но. такж е и человек, есть продукт процесса
развития»1 . П о опенке В . И . Ленина « Д арвин ... впервые поставил
биологЙ К П т вполне научную почву»2 . П ознание законов эволюции,
начавшееся с трудов Ч . Дарвина, расширяет горизонты науки и дает
возм ож ность о см ы сли ть ф акты специальных биологических дисциплин,
таких как морфология, зоология, физиология, биохимия, ботаника.
В области практической биологии теория Ч . Дарвина указала п у ть к
созданию естественной систем ы органического мира. Основанием к
группированию организмов в такой системе сл уж и т их генеалогия,
степень исторического, филогенетического родства.
7.1.7.
Развитие дарвинизма в X I X веке
Эволю ционная теория Ч . Д арвина б ы л а встречена современниками
с больш им интересом. Издание «Происхождения видов п утем есте
ственного отбора или сохранение избранных пород в борьбе за жизнь»
(1859) разош лось в первый ж е день его появления. П олож ения теории
Ч . Дарвина вызвали ожесточенную критику со стороны тех, к то видел в
ннх ниспровержение религиозных догм.
В лице передовых есте­
ствоиспы тателей то го времени эволюционная теория наш ла горячую
поддержку. Значительную роль в утверждении дарвиновских идей
сыграли публичные вы ступления и труд ы Т. Г ексли (А н глия), А за Г рея
(С Ш А ), Э . Геккеля (Герм ания), К . А . Тимирязева (Россия). Они
способствовали проникновению дарвинизма в различные области науки
о жизни, следствием чего явилось создание сравнительного метода в
зоологии и эмбриологии. Р езу л ьтаты , полученные с пом о щ ью теории
Ч . Д арвина, служили прежде всего укреплению самой эволюционной
идеи. Д арвинизм составил
теоретический
фундамент
развития
палеонтологии, экологии, биогеографии. Н е будет преувеличением
с к а з а т ь , —в— течение оолее Ч Ш полустолетия мировая наука
развивалась под непосредственным влиянием дарвинизма.
Н аряду с пропагандой эволюционной теории предпринимались
Мирке К .. Энгельс Ф С о ч 2-е изд., т 19. г 205
* Ленин В И Поли. собр. со ч . г I. < 139.
п опы тки пересмотра определенных положений дарвинизма. Так,
3 . Гекке ль1 , полн о стью принимая концепцию Ч . Дарвина о есте­
ственном отборе, одновременно признавал первостепенное значение
определенной изменчивости. О н некритически объединял взгляды
4 . Дарвина и Ж Б . Л ам арка. Согласно Л ам арку, эволю ция происходит
путем направленных и передающихся~По- наследству изменений
организмов в о тв е т на прямое или косвенное действие среды. Согласно
Д арвину, эволюция о сущ ествляется_а результате- мелких 1 неопреде­
ленных изменений организмов, кото ры е закреп ляю тся естественным
| ОТСОром.
П ризнание за естественным отбором исключительной роли в
эвол№ций~характерно для неодарвинизма. Ви д н ы й представитель этого
направления А . Всйсм ан (П П 4 — 1914) "отвергал наличие внутренних,
заложенных в грироде ж ивых сущ еств факторов эволюции в виде
«принципа градации» или «стремления к усовершенствованию»
Ж . Б . Лам арка. С т о л ь ж е негативно его отношение к наследованию
благоприобретаемых признаков, служащих результатом упражнения
или неупражнения органов. Гл а в н ы м фактором эволюции по Вейсману
является естественный отбор, который и производит все видовые
приспособления. Создав концепцию о неизменяемой зарод ы ш евой
плазме и связав ее с подовыми клеткам и , А Бейсман видел источник
новых наследственных изменений в смешении, дерекомбинации наслед­
ственных задатков гам ет родителей в процессе полового размножения.
Ограничивая изм енчивость амфимиксисом, А . Вейсман отходит о т
понимания изменчивости как всеобщего свойства ж и вы х сущ еств, чем
принижает ее роль в эволюции. Бо р ясь с «лам аркизм ом », А . Всйсман
очиш ал эволюционную теорию о г идеализма, но о тстаи вая неизменяем о с ть наследственных задатков, он способствовал возрождению
метафизических взглядов^
П редставители неоламаркизма считали, ч т о естественный отбор не
обеспечивает всех результатов эволю ции, та к как он не создает ничего
НОВОГО. П о К- Негелй Щ Г ? — 1891), В р р -гу п к тя -ге
тя рущрп-ягшание и естестБсттОГ5~отбора ф ормирую тся фнзнп логические особенно­
сти, тогд а как морфологические признаки зависят от способности
органиЗМОВ~С0 В1‘рпгсне"1"воваПься. К . Негели проводит аналогию между
процессом индивидуального развития и эволюцией. К а к в первом, так и
во втором случае закономерное развитие о т простого к слож ном у
обусловливается внутренними причинами, т. е. свойствами изменяю­
щегося ж ивого вещества. Благодаря эти м свойствам изменчивость
организмов изначально происходит по определенным направлениям.
К . Негели сводил закономерности исторического развития к физиоло­
гическим и механическим явлениям в организме, что типично для
механоламаркизма. Е ш е одно неоламаркистское течение конца X I X ве­
ка психоламаркизм (Э . К о п , 1840— 1897) связы вало историческое разви­
тие с особой силой роста, специфичной для каждого вида организмов и
И менно Э [ еккель вывел учение Ж
Б . Л а м а р к а нэ »лбиения. рассмотрев
его в книге «О б щ а я морфология», он первый поставил ими Ж
Б Л а м а р к а как
автора эволюционной теории рядом с именем Дариина
’18
направляющей наследственную изменчивость. Д л я механо и психола
маркизма в одинаковой мере характерно признание изначальной
целесообразности изменений ж и вы х сущ еств в эволюции, ч то не­
избежно приводит к идеализму в вопросах происхождения жизни и
природы движуш их сил исторического развития органического мира.
В X I X век уходит корнями мутационная теория эволюции русского
ботаника С . И . Ко рж инского (1861— 1900) и голландского ботаника
Г . де Ф ри за (1848— 1935). Согласно этой теории, образование новых
видов происходит не путем постепенных изменений, а внезапно,
скачкообразно, без пром ежуточны х переходных форм. В качестве
примера Г . де Ф р и з использовал наблюдения над энотерой — растени­
ем с выраженной способностью д авать новы е разновидности. Ж и в о й
организм представлялся Г . де Ф р и зу сум м ой независимых друг о т друга
наследственных единиц, а видообразование — чисто количественным
процессом изменения этих единиц. М утационная теория основывается
на механистических представлениях о развитии, в ней принижена роль
естественного отбора.
В истории эволюционной идеи, охватываю щ ей период о т первых
догадок древних мыслителей до начала текущего столетия, вы д ел яю т
следующие периоды: 1) непосредственно предшествовавший появлению
теории Ч . Дарвина; 2) распространения дарвинизма; 3) критики и даже
отрицания дарвиновских представлений о ходе эволюционного процесса.
Первая половина X X века ознаменовалась выд аю щ им ися о тк р ы ти ям и в
области генетики, цитологии, физико-химической биологии и экологии.
П оложения эволюционной теории Ч . Д арвина о см ы сливались в свете
механизмов наследственности и изменчивости, популяционной стр укту­
ры вида, закономерностей сущ ествования и развития биогеоценозов и
биосферы. В результате объединения достижений многих областей
биологии родилась современная синтетическая теория эволюции (С Т Э ).
Центральное место в ней отводится учению о популяииях^_а такж е
анализу природы элементарного эволюционного м а т е р и а ^ и эле­
ментарных эволюционных факторов.
7.2.
ПОНЯТИЕ О ВИДЕ
г
Видом н а зы в а ю т совокупность особей, сходных по основны м
морфологическим и функциональным признакам, кариотипу, поведенческим реакциям, имеющих общее пр(5йсхожденЯ^Г заселяющих
Определ е н н у ю
терр итор ию
Г я р е я п ),
гк рр щ и вяю н ги х ся
в
ппиполны х
условиях исключительно м ежду собой и при это м производящих
плодовитое п отом ство. Вид овая принадлежность особи- определяется
по со ответстви ю ее перечисленным критериям: морфологическому,
физиологическому, цитогенетическому, этологическому, экологическо­
му и др. Наиболее в аж н ы й признак вида — полная генетическая
(репродуктивная? ИЗОляция, заклю чаю щ аяся-в нескретпивяемпсти осо­
бей данного вида ^ п р ед ставителями других видов.
" С о времен- 1<ГЛиннея~вйд с л у ж и т 7УСНоинцй~сдиницей систематики.
Особое положение вида среди других систематических единиц (таксо­
нов) обусловливается тем , ч то в этой группировке отдельные особи
сущ ествую т реально. В составе вида в природных условиях особь
рождается, достигает половой зрелости и выполняет сво ю главную
биологическую функцию — участвуя в репродукции, обеспечивает про­
должение рода. В отличие о т вида таксон ы иадвидового ранга, такие
как род, отряд, семейство, класс, тип не яв л яю тся ареной реальной ж и з­
ни организмов. Выделение их в естественной системе органического
мира отраж ает р езультаты предшествующ их этапов исторического
развития ж ивой природы. Распределение организмов^ по над видовым
таксонам указы вает на степень их филогенетического родства.
Важ нейш им ф актором объединения организмов в виды сл уж и т
половой процесс. П редставители одного вида, скрещиваясь друг с
другом, обм ениваю тся наследственным материалом, что ведет кТтерекомбинации в_ каж д ом поколении генов_^аллелей), составляю щ их
генотипы отдельных особей. В результате достигается нивелировка
различий м ежду организмами внутри вида и длительное сохранение
основных морфологических, физиологических и прочих признаков,
отличаю щ их один вид о т другого. Благодаря половом у процессу
происходит т а к ж е объедннение генов (аллелей), распределенных по
генотипам разных особей, в общий генофонд-(аллелофоид)1 в и д а ^ Э то т
генофонд заклю чает ^ себе весь объем н'аСЛбДСтвенной информации,
ко то р ы м располагает вид на-определенном этапе его сущ ествования.
Форм ули ровка вида, приведенная вы ш е, ие м о ж е т б ы т ь применена к
агам н ы м (некоторые микроорганизмы, сине-зеленые водоросли),
самооплодотворяю щ имся и строго партеногенетическим организмам.
Группировки таких организмов, эквивалентные виду, в ы д ел яю т по
сходству фенотипов, общности ареала, близости генотипов по
происхождению. Практическое использование понятия «ви д » д аж е у
организмов с половы м размножением нередко затруднено. Э т о
обусловлено динам ичностью видов, проявляю щ ейся во внутривидовой
изменчивости, « р а зм ы то сти » границ ареала, образовании и распаде
внутривидовых группировок различного объема и состава (популяций,
рас, подвидов). Д и нам ичность видов является следствием действия
элементарных аволюПЖШных ф акторов.
7.3.
П О Н Я ТИ Е о П О П У Л ЯЦ И И
В природных условиях организмы одного вида заселяю т ареал
неравномерно, о тм ечается чередование участков повыш енной и пони­
женной концентрации особей (рис. 105). В результате вид распадается
на группировки или популяции, соответствую щ и е зонам более плотного
заселения. Д ействительно, радиусы индивидуальной активности о т ­
дельных особей ограничены. Т ак, виноградная улитка (НеНх рошасеа)
способна преодолеть расстояние в несколько десятков метров, оидатра
(Опйасга г н е й с а ) — в несколько со т метров, песец (А1орех 1а§орих) — в
несколько со т километров. Благодаря это м у размножение (репроО б ъем генетической информации, которой располагает вид иди популяция,
обусловливается совокупностью наследственных задатков во всех аллельных формах
!а к н м образом, более полно объем наследственной информации отраж ает термин
«адлелофонд», наиболее употребим
«генофонд».
240
Р яс.
105. Неравномерное распределение особен в пределах ареала вида
а - лситпчиый, в, к
(МТнттыА или островной типы
дуктивные ареалы ) в основном приурочено к территориям с повы
шенной п л отн о стью организмов. Вероятн ость случайных скрещиваний
(панмиксии), обусловливаю щ ая из поколения в поколение эффективную
пере комбинацию генов, внутри «сгущ ений» оказы вается вы ш е, чем в
зонах между ними и в среднем для вида. Т аки м образом, в репродуктивном процессе генофонд вида предоставлен генофондами популяций.
П опуляцией1 н а зы в а ю т группу особей одного вида, "населяющих
стюедёленну ю^территорию (ареал) д остаточно долго (в течение многих
поколений), __в _ которой фактически осущ ествляется сравнительно
высокий уровень панмиксии ^ которая в определенной степени отд елена
о т других таких групп то й или иной формой изоляции.
7.3.1.
Экологические характеристики популяции
Экологически популяция характеризуется величиной, оцениваемой
возрастного и п ^овото^остава^популяции. Р азм еры ареала зависят7от
«радиусов индивидуальной активности» организмов данного вида и
особенностей природных условий иа соответствую щ ей территории
Численность особей в популяциях организмов разных видов варьирует
Так, количество стрекоз (ЬеисоггЫша а1Ы1гсш) в популяции на одном из
подмосковных озер достигало 30 ООО, тогда как численность популяции
земляной улитки (Сераев петогаИз) оценивалась в 1000 экземпляров.
Сущ ес т в у ю т миним альн ы е значения численности, при которы х популяция способна поддержать себя во времени, а сокращение численности
ниже этого миним ум а п р и в о д и т к вы м иранию популяции. Величина
популяции постоянно колеблется, что зависит о т изменений'экологиче­
ской ситуации. Т ак, осенью благоприятного по ко рм овы м условиям
года популяция диких кроликов на одном из островов у юго-западного
побережья А нглии состояла из 10 000 особей. П осле холодной
малокормной зи м ы число особей упало до 100. Возрастная структура
популяций организмов разных видов варьирует в зависимости от
прбдблжитеЛБНосПГ ~ЖШТ№. интенсивности размножения,, вщ раста
достижения половой зрелости. 11олбвой состав популяций обусловлива
1 Определение справедливо для видов г половым размножением
ется эволюционно закрепленными механизмами формирования первич­
ного (на м ом ен т зачатия), вторичного (н а м ом ент рождения) и
третичного (во взрослом состоянии) соотношения полов.
7.3.2.
Генетические характеристики популяции
Генетически популяция характеризуется ее. генофондом (аллелофондом), ко то ры й представлен совокупно стью аллелей, ооразукмцнх
генотипы органЩ'моЁГ данной популяцщц Генофонды природных
популяций отличает наследственное разноооразие (г енеТиЧ?гка?Ггётерогенность или полиморфизм), генетическое единство, динамическое
равновесие лоли особей с разным и генотипами .
. ^ Наследс т венное разнообразие связано с наличием в генофонде
одновременно различных _аллелей отдельных генов. Первично оно
создается мутационны м процессом. М утации, будучи обычно- ре­
цессивными и не влияя 1 1 а фешзтипы гетерозиготных организмов,
сохраняю тся в генофондах популяций в скр ы то м о т естественного
отбора состоянии. Накапливаясь, они о бразую т «резерв наследственной
изменчивости». Благодаря комбинативной изменчивости э то т резерв
используется для создания в каждом поколении новых комбинаций
аллелей. О б ъем такого резерва огромен. Т ак, при скрешиааиии
организмов, различающихся по 1000 локусам ' , каж дый из которых
представлен десятью аллелями, количество вариантов генотипов
1 достигает Ю 1000,
что превосходит число электронов во Вселенной.
*)' Генетическое единство популяции обусловливается д о стато чн ы м уров
* нем панмиксии! В условиях случайного подбора скрещиввющихся
особей источником аллелей для генотипов организмов последова
угельных поколений является гсиофондГ популяций^
3
В пределах генофонда популяции ц о л и "теясггипов, содержащих
разные ялпепи одного^ гена, при собл юдении некоторы х условий из
поколения в поколение не изменяется. Э т и условия о писываю тся
основны м законом популяционной генетики, сф ормулированным в
1908 г. английским м атем ати ко м Д ж . Х ард и и немецким врачомгенетиком Г^ Ва й и б ер го м ., В популяции из бесконечно больш ого числа
свободно скрещивающ ихся особен ” в ^ о тсутствии ~мутац ий. и з бира­
тельной м и грации организмов с различны м и генотипами и давления
естественногоотбора пер ао Ш чальныё часто ты яппепей.сохраняются из
гюкблёцил в поколение" Д оп усти м , что в генофонде популяции,
удовлетворяющей описанным условиям, некий ген представлен аллеля­
ми А , и А 2, обнаруживаемыми с частотой р и д . С оответственно, особи
данной популяции о бр азую т р гам ет с аллелем А , и д гам ет с аллелем
А 2. Е с л и скрещивания происходят случай н ы м образом, т о доля
половых клеток, соединяющихся с гаметами А ]г равна р , а доля
половых клеток, соединяющихся с гам етам и у42 , — д. Возникаю щ ее в
результате описанного цикла размножения поколение Г , образовано
генотипами А , А ,, А , А 2, А 7 А 2 , количество которых описывается как
(Р + Ф ( Р + <7) = Р 2 +2р<? +д2 (рис. 106). П о достижении половой
1 Ч исл о локусов (ген о в> V человека превыш ает эту цифру на 2
242
3 порядка
зрелости о с о б и Д Л , и Д А * о бразую т по
одному типу гам ет — А х или А 2 — с ча­
стотой, пропорциональной числу орга­
низмов указанных генотипов ( р 2 и д 2).
Особи А хА 2 образую т оба типа гамет с
равной частотой (4 ^ )- Т аки м образом,
доля гамет А в поколении Г а составит:
<- Р (1 — Р ) - Р-
Р1 +
а доля гамет А 2 составит:
ч2 +
ч‘
иЧ (>—</)
ч
Рис. 106. Закономерное распре
депение генотипов в ряду ноко
Так как часто ты гам ет с разными . 1ений в зависимости от частоты
образования 1 амет разных ти
аллелями в поколении ^ в сравнении с поя Закон Харди Вейнберга
родительским поколением не изменены,
Объяснение в тексте
поколение Р2 будет представлено орга­
низмами с генотипами А , А „ А , А 2 и А 2А 2 в то м ж е соотношении: р 2 +
+ 2р ц - д . Благодаря это м у очередной цикл размножения произойдет
при наличии р гамет А х и д гам ет А 2. А налогичные расчеты можно
провести для локусов с л ю б ы м числом аллелей. В основе сохранения
частот аллелей л еж ат статистические закономерности случайных собы ­
тий в больших выборках.
Закон Х ар д и — Вайнберга описывает условия генетической
стабильности популяции. П оп ул яц ия, гено онд ко торой не изменяется
в ряду поколений, назы вается менделевской. Генетическая стабильность
менделевских популяций стави т их вне процесса эволюции, так как в
таких У с л о в и я х приостанавливается действие естественного отбора.
Выделение менделевских популяций имеет чисто теоретическое значе­
ние. В природе эти популяции не встречаю тся. В законе Харди —
Вайнберга перечислены условии, закономерно изменяющие генофонды
популяций. К указанному результату приводят, например, ф акторы,
ограничивающие свободное скрешивание (панмиксию ), такие, как
конечная численность организмов в популяции, изоляционные барьеры,
препятствую щ ие случайному подбору брачных пар. Генетическая
инертность преодолевается такж е благодаря м утациям , притоку в
популяцию или о тто ку из нее особей с определенными генотипами.
Отбору.
7.3.3.
М есто видов и популяций в эволюционном процессе
У ч и т ы в а я общ ую адаптивную (приспособительную) направлен­
ность эволюции, виды, возникающ ие в результате этого процесса,
представляю т собой совокупности организмов, так или иначе приспо­
собленных к определенной среде. Э т а приспособленность сохраняется
на протяжении длительного ряда поколений благодаря наличию в гено­
фондах и передаче п о то м ству при размножении соответствую щ ей
биологической информации. И з этого следует, что
при мало
243
меняющихся условиях обитания сохранность вида во времени зависит
от стабильности, консерватизма его генофонда. С другой стороны,
стабильны е генофонды не обеспечиваю т вы ж и ван и я в случае изменения
УСТОЧЩЙ жизни в историческом развитии планеты. Такие генофонды
д аю т меньше возможностей для расширения ареала вида и освоения
новых экологических ниш в текущ ий исторический период. П опуляци­
онная структура вида позволяет со вм естить долговременность приспо­
соблений, сформировавшихся на предшествующих этапах развития, с
эволюционными и экологическими перспективами. Генофонд вида
фактически распадается на генофонды популяций, каждый из которых
отличается собственны м направлением изменчивости. Популяции —
т о генетически о ткр ы ты е в рамках вида группировки организмов.
М ежпопуляционные миграции особей, сколь б ы незначительными они
не б ы л и , п р еп ятствую т углублению различий и объединяют популяции
б -линую систему вида. Однако в случае длительной изоляции,
например пространственной, некоторых популяций о т остальной части
вида первоначально минимальные различия нарастаю т. В конечном
итоге это приводит к генетической (репродуктивной) изоляции, что
соответствует появлению нового вида. В эволюционный ' Процесс
непосредственно вклю чены отдельны е популяции, а заверш ается он
образованием вида. Т аким образом, популяция является элементарной
эволюционирующей единицей, тогда как вид — каче ствен н ы ш этап ом
эволюции, ф иксирую щ им ее сущ ественный результат.
/З п
74
процесс
ВИ Д О О БРА ЗО ВА Н И Я.
Д Е Й С Т В И И Э Л Е М ЕН Т А РН Ы Х Э ВО Л Ю Ц И О Н Н Ы Х
Ф А К Т О Р О В В П РИ Р О Д Н Ы Х П О П У Л Я Ц И Я Х
IУ
Согласно синтетической теории эволюции элементарное эволюци­
онное явление, с которого начинается видообразование, заклю чается в
изменении генетического состава (генетической конституции или
генофонда) популяции. С о б ы ти я и процессы, способствующ ие преодо­
лению генетической инертности популяций, приводящие к изменению их
генофондов, н азы ваю тся элементарными эволюционными факторами.
К ним о тносятся м утационный процесс, популяционные волны,
изоляция, естественный отбор.
7.4.1.
М утационный процесс
Изменения наследственного м атериала половых клеток в виде
генных, хромосомных и геномных мутаций происходят постоянно.
Особое м есто в это м процессе принадлежит генны м ^м ухац иям ,
приводящим к возникновению серий аллелей и таки м образом к
разнообразию содержания биологической информации!
Влияние м утанипнного ппштесса на в идообразование носи т двоякий
характер. И зм ен яя ч асто ту одиогб'аллеля'по отнош ению к другому, он
оказывает на генофонд популяции прямое действие. Ещ е б ол ьш ее
значение имеет формирование за счет м утан тн ы х аллелей резерва
наследственной изменчивости. Э т о создает слови я для варьирования
44
ягшельного состава генотипов организмов в последовательных поколе­
ниях путем комбинативной изменчивости. Благодаря мутационному
'процессу поддерживается высокий уровень наследственного разиообра
зия природных популяций. С овокупность аллелей, возникающих в
результате м у т а ций, составляет исходный элемента рн ы й " эволкш и1 используется в процессе е
основа действия других элементарных эволюционных факторов.
Х о т я отдельная мутация — собы тие редкое, общее числСГмутаций
значительно. Д опустим , что некая мутация возникает с частотой 1 на
100 ООО гамет, количество локусов в геноме составляет 10 000,
численность особей вида в одном поколении равна 100 м лн ., а каждая
особь производит 1000 гамет. П ри таких условиях по всем локусам за
поколение в генофонде вида произойдет 10 10 мутаций. За среднее время
сущ ествования вида, равное нескольким десяткам ты сяч поколений,
количество мутаций составит 1014 . Б ольш и нство мутаций первона­
чально оказы вает иа фенотип особей неблагоприятное действие,
однако, будучи рецессивными, м утан тн ы е аллели обы чно при сутствую т
игенофпнрах популяций в гетерозиготных по со ответствую щ ем у локусу
генотипах. Благодаря это м у достигается тройственный положитель­
ный результат: ! ) исключается непосредственное б тр ицйтёльноё~влия
ние м утантного алл еля на фенотипическоёГ выражение признака,
контролируемого данны м геном; 2 ) п утем сохранения аллелей, не
имеющих приспособительной ценности в настоящих условиях суще­
ствования, но могущих приобрести та кую ценность в будущ ем или при
освоении новых экологических ннш, создается резерв наследственной
изменчивости; 3) благодаря явлению гетерозиса (гибридной мощ ности)
многие мутации, неблагоприятные по ю ГТГрШ ому-ф енотипическому
вы ражению , в гетерозиготном состоянии нередко п о вы ш аю т относи­
тельную ж и знеыхогпбнпсть организмов, х о тя доля полезных мутациимала, их абсолю тное количество в пересчете на поколение или период
существования вида м о ж е т б ы т ь больш им . Д опустим , что одна
полезная мутация приходится на миллион вредных. Тогда в рассмот­
ренном вы ш е примере среди Ю 10 мутаций за одно поколение 104 будут
полезными. За время сущ ествования вида его генофонд обогатится
Ю7 8
полезными мутациями.
М утационны й процесс, выполняя1 роль элементарного эволюнионного фактора, происходит постоянно на протяжении всего периода
существованиТ~жизни, а-отдельны е мутации возник а ю т Ш бгокр атн сГу
разных организмов^ Генофонды популяций и сп ы ты ва ю т непрерывное
давление м у т ационного процесса. Э т а компенсирует вы сокую вероятность потери в пялу поколений единичной мутации.
7.4.2.
П опуляционные волны
П оп уляц ионными волнами или «волнами ж изни» (С. С. Четвери­
ков) н а зы в а ю т периодические или апериодические колебания численности организмов в природных популяциях. Э т о явление распространя­
ется на все виды ж ивотны х и растений, а такж е на микроорганизмы.
П ричины колебаний часто и м ею т экологическую природу. Так, размеры
популяций «ж ертвы » (зайца)
растут при ослаблении дей
стви я на них популяций «хищ ­
ника» (ры си лисицы волка)
Отмечаемое в э то м случае уве­
личение кормовых ресурсов
способствует росту числен­
ности хищников, что в свою
очередь интенсифицирует ис­
требление ж ер твы (рис 107).
Отд ельны е «всп ы ш ки » чис
ленности организмов некото­
рых видов
наблюдавшиеся
в ряде регионов мира, бы ли
обусловлены деятельностью
человека. В X I X — X X веках
это наблюдалось в популя­
циях кроликов в А встралии,
домовых воробьев в Северной
Америке, канадской элодеи
в Евразии. В настоящее время
существенно впзпоггш . разме­
ры популяций домовой мухи,
находящей прекрасную кормо­
в у ю б а з у Т ви д е разлагающих
ся пищевых отбросов вблизи
поселений человека. НапроТИв; численность популяций
Рнс. 107. Колебания численности особей
домовых воробьев в городах
популяциях хищников и жертвы
падает вследствие прекраще­
Иунктнрчан линия а — рысь. Й
лисица, н — вол|
сплошная 1НННИ. заяц йетяи Численность особей дана
ния ш ирокого использования
процентах к минимальному из зарегистрированных зк;
ченнй. принятойV за 100%
лошадей. М а с ш та б ы колеба
ний численности организмов
разных видов варьирую т. Т ак, для одной из зауральских популяций май
ских ж уков отмечены изменения количества особей в 106 раз.
И зм енение генофондов популяций происходит кяу ня ппп ^ р м р так- и
на ш а д е Д ш ш п я щ ю ш ю й водщ л. При росте численности организмов
наблюдае!Х^Ш1ШшедзаЕее_42азобшеыных-попудящ4й-ч
генофондов. Т ак как популяции по своему генетическому составу
уникальны, в результате та ко го слияния возникаю т новые генофонды с
измененными по сравнению с исходными частотам и аллелей. ТГуслой н
ях возросшей численности интенсифицируются м еж популяционные
миграции особей, ч то т а кж е способст в у е т перераспределению яд
лелей.
Р о с т количества организмов сопровождается расширением занимае­
мой территории. Н а гребне популяционной волны некоторые группы
особей в ы сел яю тся за пределы ареала вида, оказы ваю тся в нетипичных
условиях существования и и сп ы ты ва ю т действие новых факторов
естественного отбора^ П о в ы ш ение коннентря 1цлц _а еп б ёй в__СВЯЗИ С
246
ростом их численности усиливает внутривидовую борьбу за сущ ество­
вание.
При спаде численности наблюдается распад крупных популяций
Возникаю щ ие малочисленные популяции характеризуются измеиенны
ми генофондами В условиях массовой гибели организмов редкие
м утан тн ы е аллели м огут о ы т ь генофондом вида потеряны
П ри
сохранении редкого аллеля его концентрация в генофонде м вло
численной популяции автоматически возрастает Н а спаде «волны
жизни» часть популяций, как правило, небольших по размерам, остается
за пределами обы чного ареала вида и, и сп ы ты вая действие необычных
условий жизни, вым ирает. Иногда, благодаря благоприятному генети­
ческому составу, такие популяции переживаю т период спада численно­
сти. Будучи изолированными о т основной м ассы вида, сущ ествуя в
необычной среде, они нередко сл уж а т родоначальниками новых видов
П опуляционные вол н ы — это эф ф ективней факхдр преодоления
генетической инептнпгтм ппирппных попупяттиП В м р г т р ■
их
действие на генофонды не является направленным. Д Р ЗЮ м у пни., так же
как и м утационный процесс, подготавливаю т эво л ю ционный м а тер и я к
действию других элементарных эволюционных факторов
7 .4 3 . Изоляция
Ограничение свободы скрещиваний (панмиксий) организмов называ
ю т 'изоляцией' С н и ж ая уровень паимиксии, изоляция приводит к
увеличению доли близкородственных скрещиваний.- С о п утствую щ ая
эТоЯУ гомозиготизацйя усиливает особенности генофондов популяций,
которые создаю тся вследствие мутаций, комЬннативной изменчивости,
популяционных волн. П р еп ятствуя нивелировке межпопуляционных
генотипических различий, изоляция сл уж и т необходимым условием
сохранения, закрепления и распространения в популяциях генотипов
повышенной ж и знеспособности.
В зависимости п т п р и р о д ы факторов ограничения панмиксии
р азли чаю т географическую и биологическую изоляцию . Географическая
изоляция заклю чается в пространственном разобщении популяций'
благодаря особенностям ландшафта в пределах ареала вида — наличию
водных преград для «сухопутны х» организмов, участков суши для
видов-гидробионтов, чередованию возвыш енных участков и равнии. Е й
способствует м алоподвижный или неподвижный (у растений) образ
жизни. Так, на Гавайских островах популяции наземных улиток
заним аю т долины, разделенные невысокими гребиями, преодоление
которых м оллю скам и затрудняет сухость почвы и редколесье
Вы раж енная, хотя и не полная изоляция в течение многих поколений
привела к о щ ути м ы м различиям фенотипов улиток из разных долин.
В горах острова Оаху, например, один из видов ули ток (АсЪайпеПа
ши5(еНпа) представлен более чем сотней рас, выделяемых по
морфологическим признакам. Пространственная изоляция м ож ет б ы т ь
и при о тсутстви и видимых физико-географических барьеров П ричины
ее в та ко м случае крою тся в ограниченных «радиусах индивидуальной
активности » Так, у береговой ры бы бельдюги (^оагсе5 уКзрагиз) в
247
зависимости о т м еста обитания (устье или конец фиорда) уменьшается
число позвонков и лучей некоторых плавников. Сохранение изменчиво
сти объясняется оседлым образом жизни бельдюги. Такая изменчивость
наблюдается и у подвижных видов ж и вотн ы х , например перелетных
птиц с гнездовым консерватизмом. Т ак, молодь ласточек возвращается
с зимовки на место своего рождения и гнездится в радиусе до 2 км от
матери некого гнезда, поэтом у скрещивания у ласточек ограничиваются
группой близко селящихся особей. В отличие о т разделения барьерами
э ту разновидность изоляции обозначаю т как разделение расстоянием
Биологическая изоляция возникает вследствие внутривидовых
различий организмов 'й имеет несколько форм.
Й &яогической
изоляции приводят особенности окраски покровов или"со става пищи"
размножение в разные сезоны, у паразитов — использование в качестве
хозяина организмов разных видов. Т ак, в М олдавии есть две
иесмвшивающихся популяции м ы ш ей — желтогорлая лесная м ы ш ь и
степная. Ф як т о р п м ря-чпепрнца их служ и т состав пиши. Разобщение
популяций способствовало вы явлен и ю и усилению особенностей
фенотипа степных м ыш ей — они мельче и и м ею т иную форму черепа
В описанном примере экологическая изоляция дополняется территори
альной. Сезонные расы, выделяемые по м есту и времени икрометания
описаны у лососевых, осетровых, карповых ры б. Д лительная экологи
ческая изоляция способствует дивергенции популяций вплоть до
образования новых видов Так, предполагают, что человеческая и
свиная аскариды, морфологически сходные, произошли о т общего
предка Их расхождению, согласно одной из гипотез, способствовал
запрет иа употребление человеком в пиш у свиного мяса, который по
религиозным соображениям распространялся длительное время иа
значительные м ассы людей
Экологическая изоляция сущ ествует
благодаря нюансам ритуала ухаживания, окраски, запахов, «пения»
самок и самцов из разных популяций
Так, подвиды щеглов —
седоголовый (Сап1иеК$ сагйиеНх сагйиеНк) и черноголовый (С . с.
ЬгеУ1го5(п$) и м ею т выраженны е о тм ети н ы иа голове Серые вороны
(С о с у щ согопе согшх) из крымской и североукраинской популяций
внешне неразличимые, о тл ичаю тся карканьем. П ри физиологической
изоляции препятствием к скрещиванию сл уж а т различия в структуре
органов размножения или просто разница в размерах тела. У растений к
этой форме изоляции ведет приспособление цветка к определенному
виду опылителей.
Описанные ф ормы изоляции, особенно в начальный период их
действия, сн и ж а ю т Гн о Т ГГ 'й ск л ю ча ю т полностью межпопуляциониые
скрещи в а н и я " I енетическая изоляция ^создает более жесткие, иногда
непреод олим ые барьеры скрещивания. О на заклю чается в несовместиШ с т и гам ет, гибели зигог непосредственно после оплппотворСТШГГ
стерильности или малой жизнеспособности гибридов.
Иногда разделение Популяций начинается именно с генетической
изоляции, к которой приводят полиплоидия или массивные хром о ­
сомные перестройки резко изменяющие хромосомные наборы гамет
музантов~по сравнению с исходными ф о р м а м и "П о т и лоидия распро­
странена среди расте нй (рис 108) Разны е виды плодовой мухи
нередко различаю тся хромосомными перестройками
Ги б ри д ы о т
скрещивания близкородственных форм со сниженной жизнеспособно­
с т ь ю известны для серой и черной ворон. У казан н ы й ф актор изолирует
популяции этих птиц в Евр опе (рис. 109). Ч а щ е генетическая изоляция
развивается вторично вследствие углубления морфологических разли­
чий организмов из популяций, длительно разобщенных другими
формами изоляции. В первом случае генетическая изоляция предше­
ствует дивергенции признаков и начинает процесс видообразования, во
втором — она его завершает.
И золяция в процессе видообразования взаимодействует с другими
элементарными эволюционными факторами. О н а усиливает генотипи­
ческие различия, создаваемые мутацион н ы м процессо^иТёнехическоЙ
комбинаторик о й Возникаю щ ие благодаря изоляции внутривидовые
группировки о тл и ча ю тся по генетическому составу и и сп ы ты ва ю т
неодинаковое давление отбора.
7.4.4.
Естествен н ы й отбор
В популяции организмов, размножаю щ ихся поло вы м способом,
сущ ествует больш ое разнообразие генотипов и, следовательно, феноти­
пов. Благодаря индивидуальной изменчивости в условиях конкретной
среды обитания приспособленность разных генотипов (фенотипов)
различна. В эволюционном контексте приспособленность определяют
как произведение жизнеспособности в данной среде, обусловливающей
б ольш ую или м еньш ую вероятность достижения репродуктивного
возраста, на репродуктивную способность особи. Р а зл и чи я м ежду
"организмами по приспособленности, оцениваемой "по передаче аЛ7ТбЛ?й
следующему поколению, в ы яв л яю тся в природе с п ом о щ ью есте­
ственного отб ора. Гл а в н ы й результат отбора заклю чается-не прост о в
вы ж ивании более жизнеспособных особей, а в относительном вкладе
таких особей в генофонд дочериед попудяпии_
Необходимой предпосылкой отбора сл уж и т борьба ^ с у щ е с т в о в а ­
ние — конкуренция за п и ш у , жизненное пространство, партнера д л я
"спаривания. Естественны й отбор происходит иа всех стадиях онтогенеза организмов и обеспечивает в конечном итоге дифференциальное
(избирательное) воспроизведение (размножение! генртиппв Благодаря
естественному о тбору аллели (признаки), повы ш аю щ ие вы ж и ваем ость
и репродуктивную способность, накапливаю тся в ряду поколений,
изменяя генетический состав популяций в биологически целесообразном
направлении В природных условиях естественны й отбор осущ ествля­
ется исклю чительно ЦП фрцт-мпу
генотипов происходит
вторично через о тбор фенотипов, которы е о тр а ж а ю т генетическую
конституцию организмов.
К а к элементарный эволюционный ф актор естественный отбор
действует в популяциях. П опуляция является полем действия,
отдельные особи — объектам и действия, а конкретные признаки —
точкам и приложения отбора.
Эф ф ективность отбора по качественном} и количественному
изменению генофонда популяции зависит о т величины давления и
350
1
направления его действия. Величина давления вы р аж ается коэффици­
ентом отбора (5}Г5СТбрый^характеризует интенсивность элиминации ИЗ
рггюодуутйвиого'пропессд. или сохранения. соответственно^ менее или
ббЛ€е” прйспособленных форм по сравнению с формой, принятой за
стандарт приспособленности. Т ак, если некий локус представлен
аллелями А , и А 7 , т о популяция по гепотипам делится иа 3 группы:
А , А ,1 А , А 2 ', А ^ А 2 . О бозначим приспособленность этих генотипов
А
Вы берем в качестве стандарта первый генотип,
относительная приспособленность которого максимальна и равна 1.
Тогда приспособленности других генотипов со ставят доли этого
стандарта:
А2А2
1РП
Ео
Vи
Ы I 5,
\х\
Величины 5, и 52 означаю т пропорциональное снижение в очередном
поколении воспроизводства генотипов А х А 7 и А 2 А 7 сравнительно с
генотипом А , А г
О тб о р особенно эффективен в отношении доминантны х аллелей при
условии их полного фенотипичного проявления^ О н идет медленно в
отношении рецессивных аллелей и при неполном доминировании. Н а
эффективность отбора вли яет исходная концентрация аллеля в гено­
фонде. П р и низких и в ы соких к о н ц е н т р а ц и я х п - т р р пргъцгугГпРт-медленно- Изменение доли доминантного аллеля в сравнении с рецессиШГБШ при коэффициенте отбора 0,01 приведено в табл. 22.
Т а б л и ц а 22
а ллеля в сравнении
отбора 0,01
В осстан ие.
Возрастание частоты доминантного
с рецессивным при коэффициенте
%
Число поколении
0.1
1
1 50
5 0 ‘>8
98
99.9
250
500
5 000
9 0 000
В теории, упрощая ситуацию, допускаю т, что отбор действует на
генотипы благодаря различиям в адаптивном значении отдельных
аллелей. В реальности приспособительная ценность генотипов зависит
о т действия и взаимодействия всей совокупности генов. Оценка
величины давления отбора по изменению концентрации конкретных
аллелей технически часто невыполнима. П оэтом у расчет проводят по
изменению концентрации организмов определенного фенотипа. П у с т ь в
популяции п р и сутствую т организмы двух фенотипических классов А и Б
в отнош ении --—
происходит
V . Вследствие различий по приспособленности
естественный
отбор,
которы й
изменяет
соотношение
Рис. 110. Ф о рм ы естественного отбора
I «табнмзирующан; б движущая, в —диэруптивнан, I .
особей с фенотипами А
_ Л . щ =. [/, ( I -г 5 ),
Г» 6', — 1.
5= ~
последовэтс |ьные п.
и Б . В следующем поколении оно станет:
где
5 — коэффициент
отбора.
О тсю д а
П р и селективном преимуществе фенотипов А Ь 7 > I/, , а
5 > 0 . При селективном преимуществе фенотипов Б С 2 < С,
и 5_<0.
Если приспособленность фенотипов А и Б сопоставима и С/7 = Ц , то
5 = 0. В рассмотренном примере при 5 >0 отбор сохраняет в популяции
фенотипы А и элиминирует фенотипы Б , при 5< 0 имеет место обратная
тенденция. О тбо р , сохраняющий определенные фенотипы, по своему
направлению является п оложительн ы м , тогда как о тбор, устраняющ ий
фенотипы и-Гполуляции, — отрицательн ы м .
В зависимости о т результата различаю т стабилизирую щ ую ,
д ви ж ущ ую и дизруптивную формы естественного отбора (рисН40>,
Стабилизирую щ ий отбор сохраняет в пштурянни сдедшш варцшгг фе­
нотипа или признака. О н уст р а н я е т'из репродуктивного процесса фе
нОтипьС уклоняю щ иёся"от сложившейся адаптивной «н о р м ы » , приво­
дит к пренмушественному р а зм Ш Ж е н и б -типичных организмов. Т ак,
сотрудник одного из университетов С Ш А подобрал после снегопада и
сильного ветр а 136 оглуш енных воробьев (Раххег йошезбсиз). 72 в ы ­
ж и в ш и х воробья имели кры лья средней длины, тогда как 64 погибших
птицы б ы л и либо длиннокры лы м и, либо ко роткокры лы м и. С табилизи­
рую щ ая форма со ответствует консервативной роли естественного
отбора. При относительном постоянстве условий среды благодаря этой
: этапов эволюции.
252
Д в и ж у щий
отбор— обусловливает
прогррггнднпр
изменение
фенотипа в определенном направлении, ч то проявляется " в сдвиге*
^сггедЩ ц значений о тб и в аем ы х признаков в сторону их усиления или
ослабления. Прим ером такого отбора с л у ж и т замещение в популяции
"крабов (Сагстих шаепах) ж и во тн ы х с широким головогрудным щ итком
на ж и во тн ы х с узким ш и тко м в связи с увеличением количества ила (га­
вань П л и м у т, А нглия). При смене условий обитания благодаря этой
форме отбора в популяции закрепляется Ф е н о т и п более соагведх х а у къ
щнй среде.
Д изруп ти вн ы й отбор сохраняет несколько разных фенотипов с
равной п риспособлен н п с т ь ю Г Он действует против особей со средним
или про м еж уто чн ы м зн ачением причч а к о в Т Так, в зависимости о т
преобладающего цвета почвы виноградные улитки (Сераеа петогаНз)
и м ею т раковины коричневой, ж елто й , розовой окраски. Д изруптивная
форма отбора «разры вает» популяцию по определенному признаку на
несколько групп. Она поддерживает в популяции состояние генетиче­
ского полиморфизма.
В зависимости о т формы отбор сокращает м а сш та б ы изменчиво­
сти, создает н овую или сохраняет п реж н ю ю картину разнообразия. К а к
и другие элементарные эволюционные ф акторы естественный отбор
вы зы ва ет изменения в соотношении аллелей в генофондах популяций,
и со Б сш ю с т ь его действия состоит _ в то м , ч то эти изменения
направленны. О тб о р приводит генофонды в соответствие с критерием
приспособленности, осущ ествляет ооратную связь м еж д у изменениями
генофонда и условиями обитания, накладывает на эти изменения печать
биологической целесообразности (полезности). Естествен н ы й отбор
действует совместно с другими эволюционными ф акторами. Поддер­
ж и вая генотипическое разнообразие особей в ряду поколений,
мутационный процесс, популяционные волны, комбииативиая изменчи­
вость создаю т для него необходимый материал.
Естествен н ы й отбор нельзя* рассм атривать как «си то », сортирую ­
щее генотипы по приспособленности. В эволюции ему принадлежит
творческая роль. И скл ю чая из репродукции генотипы с малой
приспособительной ценностью, сохраняя благоприятны е генные комби­
нации разного достоинства, он преобразует картину генотипической
изменчивости, склад ы ваю щ ую ся первоначально под действием случай­
ных факторов, в биологически целесообразном направлении. Р езульта ­
то м творческой роли отбора с л у ж и т процесс органической эволюции,
идущей в целом по линии прогрессивного усложнения морфофизиологи­
ческой организации, а в отдельных ветвях — по пути специализации.
7. 4.5. П у т и и способы видообразования
Процесс образования видов осущ ествляется в результате взаимо­
действия элементарных эволюционных факторов. Видообразование в
типичных случаях заклю чается в разделении первоначально единого
вида иа два или более новых в свищ с возникновением М5яотбпуляциоиных изоляционных барьеров и~~углубпением ^различий под действием
естественного о тб ора вп ло ть до генетическЫГйзоляцийГТ ако й процесс,
Энсчюцисннсе изменение
Рис. I I I . Основные способы видообразования,
л
фнлнтифское. б
диеср1ентнпе (истинное) и
путем гийрим
ведущий к увеличению количества видов, называется истинны м
видообразованием. Вы д еляется та к ж е филитическое видообразование,
которое заклю чается в погт р п рч м п м прёШЗашении во времени одного
вида в другой, ч то няйпюляется при изменении условий во всем ареале.
И звестн ы примеры видообразования путем гибридизации. Пере­
численные способы видообразования схематически изображены на
рис. 111.
С у щ е ств у ю т аллопатричсский и симпатрический пути образования
ыщовТ П р и аллопатрическом видообразовании, называем ом т а к ж е
географическим, "препятствия к скрещиванию первично обусловлена
'пространственной изоляцией популяций, а генетическая изоляция
(в зви вае тся вторично. Т ак, некогда в А встр ал и и сущ ествовал один вид
попугайчиков рода РасЪусерЬа1а. В засуш ливый период единый ареал
разделился на западную и восточную зоны , и со временем особи даух
популяций приобрели морфофизиологические различия,
которые
исключили скрещивания, когда ареал вновь стал общ им. П роизош ло
образование из одного предкового вида двух новых. П ри симпатрическом видообразовании новый вид образуется в н у т р и апеагтя идхппногб
ние создается в результате полиплоидии вследствие нарушений нормаль­
ного хода мейоза, крупных хромосомных перестроек, межвидовой
гибридизации. Аллопатрическое видообразование происходит медленно
и дает виды, как правило, отличаю щ иеся по морфофизиологическому
критерию о т вила-родоначальника. Симпатрический п уть видообразо­
вания относительно б ы стры й и в результате него о бразую тся виды,
близкие к исходному по морфофизиологическим показателям. Б ольш ин­
ство видов, особенно ж ивотны х, возникает аллопагрическим путем.
Симпатрическое видообразование на основе полиплоидии характерно
для растений. Т ак, разные виды пшениц со ставл яю т ряд с наборами 14,
28, 42 хромосомы. В клетках дикого хлопчатника 26 хромосом, а
культурного — 52. К у л ьт у р н а я слива возникла путем гибридизации
терна с алы чой. П римером гибридогенного вида сл уж и т рябиноки
зильник, распространенный в лесах ю ж но й Якути и . Симпатрический
п уть видообразования у паразитов часто связан с освоением популяций
гювых хозяев.
7.4.6. Генетико-автоматические процессы (дрейф генов)
М утаци и и ком бинативная изменчивость, периодические колебания
численности организмов, изоляция и зм еняю т генофонды популяций
случайны м образом. И х действие совместно с естественным отбором в
процессе видообразования придает биологической изменчивости в це­
лом приспособительный характер. Вы полнению отбором упорядочива
ю шей роли преп ятствую т изменения концентрации аллелей, зависящие
о т случайны х причин, которы е обусловли ваю т преимущественное
размножение генотипов вне связи с их адаптивной ценностью.
Д инам ика концентрации аллелей в генофондах последовательных покопенийТ ^ б с й т^ гг а ^ с г и ч е с к тш характер, поэтом у размах случайный
(Гбле&ш ий ч ас то т аллелей увеличивается по мере сокращения численности~попу ляци И ■ _____
Р а счеты показы ваю т, что при воспроизведении 5000 п о то м ­
ков родительской популяцией с частотой некоего аллеля р = 0,50 в
99,994^о вариантов дочерних популяций колебания концентрации этого
аллеля в силу случайных причин (в о тсутстви и отбора по это м у аллелю)
ие вы й д ут за пределы 0,48— 0,52. Есл и ж е родительская популяция мала
и воспроизводит 50 потомков, т о диапазон случайных колебаний
концентрации наблюдаемого аллеля в т о м ж е проценте вариантов
дочерних популяций составит 0,30— 0,70. Случайные, не обусловленные
действием естественного отбора колебания часто т аллелей назы ваю т
генетико-автоматическими процессами или дрейфом генов.
При
значительном
размахе колебаний в
последовательных
поколениях создаю тся условия для потери популяцией некоторых
аллелей и закрепления других. В результате происходит гомозиготизация особей и затухание изменчивости. Предположим, ч то популяция
со стоит из четырех особей и имеет аллель с частотой р =0,125. Э т о оз­
начает, ч т о указанный аллель присутствует в генофонде в единственном
экземпляре у одной из особей, гетерозиготной по соответствую щ ем у
локусу.
Л ю бо е случайное стечение о бстоятельств, исключающ ее такую
особь из размножения, приведет к утр ате аллеля и генофонд дочерней
популяции будет его лишен. Вероятность у тр а т ы со ставит 1/2 в случае
одного, 1/4 — двух, 1/8 — трех потом ков у данной особи. В популяции
из 4000 организмов при р = 0,125 м инимум 500 особей и м ею т соответ­
ствую щ ий аллель, причем в гом озиготном состоянии. Вероятность
исключения всех этих особей из размножения в силу случайных
о бстоятельств ни чтож н о мала, ч то гарантирует переход аллеля в
генофонд следующего поколения и его сохранение.
Дрейф генов обусловливает утрату (р = 0) или Фиксацию (р = П
аллёлеи и гим цзиГотнохГш стаянии У пгех чпенпв пппупяпии вне связи с
их адаптивной ценностью. О н играет в аж н ую роль в формировании
генофондов малочисленных гру п п ~опганизмов~— изолированНБ1Х~~РТ~ —
остальной час^и в и з?
5
7.4.7.
Наследственный полиморфизм природных популяций.
Генетический груз
Процесс видообразования путем естественного отбора создает
разнообразие ж и вы х форм, приспособленных к условиям обитания. К а к
удается со вм естить эти два результата? Среди разных генотипов,
возникающих в каждом поколении благодарЗТ'р^зёрву наследственной
йТМбТ-ГГивости и генетической комбинаторике, лишь ограниченное их
~чйсло обусловливает максимальнукт приспособленность к конкретной
среде. М о ж н о предположить, что дифференциальное воспроизведение
этих генотипов в конце концов приведет к то м у , чт о генофонды популяпий будут представлены ли ш ь «уд ачн ы м и » аллелями и ихкомвинация1П1Л с протнойДС!- затухание наследственной изм енчивости и
ГТОвышение степени гомозиготиости генотипов. В природных популяциях, однако^ наблюдается противоположное состояние — больш инст­
ва Организмов вы сокр гдтепошготно. Отд ельны е особи ге^'ЗрТГПгттны
ПО равным локусам , что п овы ш ает сум м арную гетерозиготиость
популяции. Так, методом электрофореза на 126 особях рачка ЕпрЪагша
щрегЬа, представляющего главную пищ у китов в антарктических водах,
изучали 36 локусов, кодирующих первичную стр уктур у ряда ферментов.
П о 15 локусам изменчивость о тсутствовал а, 21 локус имел по 3— 4
аллельные ф ормы. В целом в этой популяции рачков 5 8 % локусов были
гетерозиготными и имели по два и более аллелей. В среднем каждая,
особь имела 5 ,8 % гетерозиготных локусов. Средняя степень гетерозиготности у растений составляет 1 7 % , беспозвоночных — 13,4% ,
позво н очн ы х— 6 ,6 % . У человека э т о т показатель равен 6 ,7 % . С то л ь
в ы со кую степень гетерозиготиости нельзя о бъяснить то лько м утациями
в силу их относительной редкости.
Наличие в популяции нескольких геиртииргкид фррм (генотипов^ в
состоянии длительного равновесия в концентрации, превыш аю щ ей по
наиболее редкой форме 1 % ' , н а з ы в а ю т полиморфизмом. Н а след­
ственный полиморфизм создается м утациям и и комбинатианпЁГитмёнчиго стью . Он поддерживается естественным отбором и б ы ва ет
адаптационным и гетерозиготным (балансированным)! А д адтациоиный полиморфизм возникает, если в различных, но закономерно
изменяющихся условиях жизнн отбор благоприятствует разным
генотипам. Так, в популяциях двухточечных бож ьих коровок (АёаИа
Ы р и п с Ш а ) при уходе на зим овку преобладают черные ж уки , а весной
красные (рис. 112). Э т о происходит потом у, что красные ф ормы лучш е
переносят холод, а черные интенсивнее разм н ож аю тся в летний период.
7>.,лансированный полиморфизм возникает, если отбор благоприят^
<лаует гетерозйготам в сравнении с~рецессивиыми и д ом инантны м и
гомозиготами. Т ак, в экспериментальной численно равновесной популя­
ции мух (П)го5орЬНа теЫпоеаМ ег), содержащей сначала много м ута н то в
с более те м ны м и телами (рецессивная м утация еЬопу), концентрация
последних бы стр о падала, пока ие стабилизировалась иа уровне 10%
Уровень наследственного разнообразия, для поддержания которого достаточно
|гнойного процесса
Рис. И З . Балансированный поли мор
физм по л оку су окраски тела в экспоР"ментальной
популяц ии
п л о д о в ы х :'0'
мух. Справа
мутантная муха с чер- ю
ной окраской тела, слева
серая муха
I IIIКИ Й ТИП)
^ис. 112- Адаптивный полиморфизм > двух
точечных божьих коровок
>••■■тношепне чгриий 1.1аштриховано) и нраенс й ф чм
при тееннем ( В ) н «и«ннем (О! сборе
я ЧГрНОМ окрас
9 весенней и
(рис. 113). Анализ показал, ч то в созданных условиях гом озиготы по
мутации еЬопу и го м ози го ты по аллелю дикого типа менее
жизнеспособны, чем гетерозиготные мухи. Э т о и создает состояние
устойчивого полиморфизма по соответствую щ ем у локусу.
^Явление селективного преимущ ества гетерозигот называется
сверхдомйнантностью . М еханизм ы положительного "Отбора гетерозигот различны. О б щ и м правилом является зависимость интенсивности
отбора о т часто ты соответствую щ его фенотипа (генотипа). К примеру,
р ы б ы , птиц ы , м лекопитаю щ ие предпочитаю т о бы чн ы е фенотипические
ф ормы добычи, «не замечая» редкие, а сам цы относительно редких
генотипов м о гут п овы ш ать конкурентоспособность за самок. С е ­
лективное преимущество гетерозигот обусловливается та кж е явлением
гетерозиса. П овы ш ен н ая жизнеспособность межлинейных гибридов
о траж ает, по-виднмому, результат взаимодействия аллельных и не­
аллельных генов в системе генотипа в условиях гетерозиготности по
м ногим локусам. Гетерозис наблюдается в отсутстви е фенотипического
проявления рецессивных аллелей. Э т о сохраняет скр ы т ы м и о т
естественного отбора неблагоприятные и даже летальны е рецессивные
мутации.
В силу разнообразия факторов среды обитания естественный отбор
действует одновременно по многим направлениям, а его конечный
результат зависит о т соотношения интенсивности разных векторов
отбора. Т ак. высокая концентрация полулегального аллеля серпо­
видной ф ормы эритроцитов человека в определенных районах земного
шара поддерживается благодаря наложению на отриц втельны й отбор
по это м у аллелю интенсивного положительного контротбора по
9 56
257
суммарной приспособленности в условиях высокой заболеваемости
тропической малярией. Конечны й результат естественного отбора в
популяции зависит о т наложения многих векторов о тборов и кон трот­
боров, благодаря чем у достигается одновременно и стабилизация
генофонда, и поддержание неследствеииого разнообразия.
Балансированный полиморфизм придает популяции ряд ценных
свойств, ч т о определяет его биологическое значение. Генетически
разнородная популяция, осваивает более широкий^ спектр условий
ж и зни, использует среду обитания более эффективно. В ее генофонде
накапливается больш ий объем резервной'наследс гвенной изменчивости.
В результате она приобретает эволю ционную 'г и б к о сть и м ож ет,
изменяясь в то м или ином направлении, компенсировать колебания
среды в ходе исторического развития.
В генетически полиморфной популяции из поколения в поколение
рож даю тся организмы разных генотипов, приспособленность которых
к условиям среды неодинакова. В каждый м ом ент жизнеспособность
такой популяции ниже уровня, ко то ры й б ы л б ы достигнут при наличии
в ней ли ш ь наиболее «удачных» генотипов. Величина, на ко то рую
приспособленное!)? реальиой.цодудящш. отл*+ч«вт2й от присдособленносГтг-ияеальной популяции из «лучш и х» генотипов, возм о ж н ы х при
данном генофонде, назы вается генетическим г р у з о м . Он п п с л г т . ш п я ^ т
собой своеобразную плату за зкологическую~пластичиость и эволюци­
онные перспективы. Генетический груз сл у ж и т неизбежным следствием
генетического полиморфизма.
7.4.8.
Адаптации организмов к среде обитвния
Наряду с обш ей приспособленностью, зависящей о т генотипа в
целом и измеряемой успехом в размножении, в процессе эволюции
возникаю т приспособления или адаптации для решения организмом
экологических задач, предъявляемых средой обитания. Отд ельны е
адаптации — это постоянно возникающ ие в процессе развития жизни,
^13меняющиеся, совершгнетвутбщибся, иногда исчезающие эволюпионно
обусловленные приспособления к конкретны м факторам среды.
В результате вы работки адаптаций достигается состояние адаптированпости или соответствия мооФологни. Физиологии, попкпнш ч
организмов заним аем ы м ими экологическим ниш ам, которы е пред­
с тав л яю т совой всю совокупность^ условий среды и оораза жизни
данного организма.
~ Процесс выраоотки адаптаций происходит постоянно и в него
вовлечены многие признаки организма. Эво лю ц и я птиц о т рептилий
вклю чала, например, последовательные изменения костей, мыш ц,
покровов, конечностей. Увеличение грудииы, перестройка гистологиче­
ской стр уктур ы костей, придавшая им наряду с п рочностью легкость,
развитие оперения, обусловившего лучш ие аэродинамические свойства и
терморегуляцию, превращение пары конечностей в кры лья, обеспечило
решение проблемы полета. У некоторых представителей птиц впослед­
ствии развились приспособления к наземному или водному образу
жизни (страус, пингвин), при это м вторичны е адаптации захватили
258
также
ряд признаков. П ингвины, например, сменили кр ы л ь я на
а их покррвы стали водонепроницаемыми.
Одна экологическая задача м о ж е т реш аться путем выработки
разных адаптаций. Т ак, средством термоизоляции у медведей, песцов
сл уж и т густой мех, а у китообразных — жировой подкожный слой.
Адаптации возникаю т в ответ на конкретную экологическую задачу,
п т т б м у ' о н и всеТдаотиосительны. Относительность адаптаций заклю- ~
чяетсяТГо г р а н и ч ен ности их приспособительного значения определенны“ {^Тусловиями обитания. Т ак, приспособительная ценность пигментиро-рапности бабочек берёзовых пядениц по сравнению со светлы м и
формами очевидна ли ш ь на закопченных стволах деревьев. Приве­
денный пример показывает такж е, что о степени адаптивности признака
м ож но судить, ли ш ь сравнив два его разных состояния.
П риспособление образуется то л ько при наличии в генофонде вида
наследственной информации, способствующ ей изменению структур и
Д у и к ц й Гв требуемомиаправлении. Т ак, м лекопитаю щ ие и насекомые
использую т для дыхания, соответственно, легкие и трахеи, которые
развиваю тся из разных зачатков под контролем разных генов. Нередко
основу нового приспособления составляет предш ествующ ая структура,
которая выполняла другие функции, ио изменилась в тако м иаправле
нии и до такой степени, что смогла взять на себя новые Ф у н к ц и и . Н али ­
чие с'т'РУГ'П'р! способных расш ирить или изменить круг функций,
называется преадаптапией. Т ак, некогда у ры б, обитавших в мелководных водоемах со стоячей и бедной кислородом водой, появились
полые в ы р о сты в передней части пищевода и м ускулисты е плавники.
Первая структура способствовала решению задачи дыхания, а вторая —
перемещения по грунту, и позволила некоторым ры бам покидать на
время водоемы. Первоначально такие выходы совершались, видимо, в
дождливые дни или влаж н ы е иочи. Им енно так делает в настоящее вре­
мя американский сомик кошка ( 1 с Ы ш т пеЬи1ош5). Впоследствии эти
структуры развились в легкие и конечности наземных ж ивотных.
К адаптации иногда приводит новая мутация, которая, вклю чивш ись в
сис] с\1У Генотипа, изменяет фенотип в направлении более эффективного ~
решения экологических задан. Э т о т п уть возникновения адаптаций--называется комбинативным.
Е с т ь несколько классификаций адаптаций. П о механизму действия
выделяю т приспособлениЯ^пассивной заш и ты (вы сокая плодовитость.
покровительственная, о тпугиваю щ ая окраскаТ! активной за ш и т ы ,
перемещения и добывания пиш и (совершенствование аппарата движения^нервной системы, органов чувств, развитие средств нападения у
хищных), к общественному образу жизни (разделение «труд а» у пчел), к
сожительству с другими организмами (симбиоз, паразитизм). В зависи­
мости о т характера изменения р азли чаю т адаптации с успожением или
упрощением морфофизиологической организации. Последние типичны
для паразитов. П о м асш табу приспособления делят иа специализиро­
ванные и общие. С п ом ощ ью специализированных адаптаций организм
решает конкретные задачи в узколокальных условиях жизни вида.
К примеру, особенности строения язы ка муравьеда обеспечивают
питание муравьями. Общие адаптации позволяю т реш ать многие
плавн ики,
259
задачи в широком спектре условий среды. К ним отно сятся внутренний
скелет позвоночных и наружный членистоногих Г 1 е.шн либин ~как
переносчик кислорода и др. Такие адаптации способствую т освоению
разнообразных экологических ниш , обеспечивают значительную эколо­
гическую и эволюционную пластичность и о бнаруж иваю тся у предста­
вителей крупных таксонов организмов. Так, первичный роговой покров
предковых форм рептилий в процессе исторического развития дал
покровы современных рептилий, птиц, млекопитающих. М асш та б
приспособления вы является в ходе эволюции гой группы организмов, у
которой оно возникло впервые.
7.4.9.
Происхождение биологической целесообразности
Биологическая целесообразность организации ж ивых сущ еств
проявляется в гармонии между морфологией, физиологией, поведением
организмов разных видов и средой их обитания. Она заклю чается такж е
в удивительной согласованности строения и функций отдельных частей
н систем самого организма. Сторонники теологического объяснения
происхождения жизни видели в биологической целесообразности
проявление мудрости творца природы. Телеологическое объяснение
биологической целесообразности исходит из принципа «конечной цели»,
согласно которому жизиь развивается направленно в силу внутренне
присущего стремления к известной цели. С о времен Ж . Б. Л ам арка
сущ ествую т гипотезы, связы ваю щ ие биологическую целесообразность
с принципом адекватного о твета организмов иа изменения во внешних
условиях и наследованием таких «благоприобретенных признаков».
Убед ительны м аргументом в пользу целесообразности изменений под
влиянием среды долго признавался факт «привыкания» микроорга­
низмов к лекарственным препаратам — сульфаниламидам, антибиоти­
кам. О п ы т В. н Э . Ледербергов показал, ч то это не так. Схема о п ы та
приведена на рис. 114. В чаш ке П етри на поверхности твердой
питательной среды микроб образует колонии (1). Специальным
ш там пом (2) отпечаток всех колоний перенесли на среду со смертельной
дозой антибиотика (3). Е сл и в этих условиях вырастала хо ть одна
колония, то она происходила о т колонии микробов, такж е устойчивых к
данному препарату. В отличие от других колоний первой чаш ки П етри
(4) она давала рост в пробирке с антибиотиком (5). Если число исходных
колоний б ы л о велико, т о среди них, как правило, находилась и
устойчивая. Т аки м образом, речь идет не о направленном приспособле­
нии микроба, а о состоянии преадаптации, которое обусловлено
наличием в геноме микроорганизма аллеля, блокирующ его действие
антибиотика. В одних случаях «устойчивы е» микробы синтезирую т
фермент, разруш ающий лекарственное вещество, в других — стенка
клетки становится непроницаемой д зя препарата. П оявлению штаммов
микроорганизмов, устойчивых к лекарственным препаратам, способ­
ствует неправильная тактика врачей, которые, желая избежать
побочных эффектов, назначаю т низкие, сублетальные дозы лекарств.
Т акж е м ож н о объясн и ть появление форм, резистентных к ядам, среди
насекомых и млекопитающих — среди м ута н тн ы х организмов нахо-
260
днтся устойчивая форма, которая подверга­
ется положительному о тбору в условиях
действия отравляю щ его вещества. Напрн
мер, устойчивость кры с к варфарину,
используемому для их уничтожения, зави­
сит о т присутствия в генотипе определен­
ного доминантного аллеля.
Во зм ож н о сть «прямого приспособле­
ния» организмов к среде обитания, «пере­
делки природы путем ассимиляции условий»
утверждалась некоторы ми биологами еще
в 40— 50-е годы текущего столетия. Точки
зрения, приведенные выш е, со ответствую т
идеалистическим взглядам, и не м огут
объяснить биологическую целесообразность
без привлечения идеи если не бога, т о специа­
льной цели или программы развития жизни,
существовавших еще до ее возникновения.
Биологическая целесообразность строеНИЯ
И ФУНКЦИЙ
О р Ган И ЗМ О В
С К Л а Д Ы В а ет СЯ
I
V
р ис. 114. Схема ОЛЫГо. опро
в процессе развития жйзии. Она иредставляи^аю ш ио
факт прим ою
п сббои историческую категорию. О б это м
приспособлении
микроом о
свидетельствует смена типов организации,
■"■1357™
ш нимаюших господствующее положение в
органическом мире планеты. Так, господ­
ство амфибий на протяжении почти 75 млн. л е т сменилось господством
рептилий, затянувш им ся на 150 мли. лет. В периоды господства любой
труппы наблюдается несколько волн вымирания, которые изм еняю !
относительный видовой со став соответствую щ его крупного таксона.
Появление л ю бы х адаптаций и биологической целесообразности в
целом объясняется работой в природе иа протяжении более чем
3,5 млрд. лет естественного отбора. И з м нож ества случайных
отклонений он сохраняет и накапливает наследственные изменения,
имеющие приспособительную ценность. Э т о объяснение позволяет
понять, почему биологическая целесообразность, если ее рассм атривать
в пространстве и времени, является относительным свойством ж ивых
существ н почему в конкретных условиях обитания отдельные
приспособления достигаю т л и ш ь той степени развития, которая
достаточна для выживания в сравнении с приспособлениями конку­
рентов.
7.6. Д Е Й С Т В И Е
ЭЛЕМ ЬН ТАРН Ы Х э в о л ю ц и о н н ы х
Ф А К Ю Р О В И Г I: Н Е Т И К О - А В Т О М А Т И Ч Е С К И Х
ПРО Ц ЕССО В В ПО П УЛЯЦ И ЯХ Л Ю Д ЕЙ
7.5.1. П он яти е о популяции людей
Размножение человека осущ ествляется половы м способом, а репро­
дуктивные ареалы индивидуумов в то й или иной степени ограничены
определенной группой населения. Э т о позволяет вы д ели ть в человече-
стве с о о б щ е с т в а , а н а л о г и ч н ы е п о п у л я ц и я м в био­
логическом понимании этого термина. В антропогенетике популяцией
называю т группу людей, занимающих общую территорию и свободно
вступающих в брак. Изоляционные барьеры, препятствующие заключе­
нию брачных союзов, нередко носят выраженный социальный характер
(различия в вероисповедании). В формировании популяций людей
главную роль играет не общ ность территории, а родственные
связи.
Размер, уровень рождаемости и смертности, возрастной состав,
экономическое состояние, уклад жизни служат демографическими
показателями популяций людей. Генетически они характеризуются
генофондами. Демографические показатели оказываю т серьезное воз­
действие на состояние генофондов человеческих популяций, главным
образом через структуру браков. Большое, значение в определении
структуры браков имеет размер группы. "Популяции численностью /
1500— 4000 человек называю т д е м а м и, популяции численностью до |
1500 человек и з о л я т а м и . Для демов и изолятов типичен относи- Г
тельно низкий естественный прирост населения — для демов порядка |
2 0 % , для изолятов не более 25% за поколение. Частота внутригруппо­
вых браков в них составляет 80—90% для демов и свыш е 90% для
изолятов, а приток лиц из других групп сохраняется на уровне 1—2 %
для демов и менее 1% для изолятов. В силу высокой частоты
внутригрупповых браков члены изолятов, просуществовавших 4
поколения (примерно 100 Нет) и более, являю тся не менее чем
троюродными сибсами.
В больших по размерам популяциях распределение аллелей
отдельных генов в генотипах индивидуумов последовательных поколе­
ний подчиняется закону Харди— Вайнберга. Э то используется в медико­
генетической практике для расчета доли гетерозигот—носителей
определенного рецессивного аллеля. Так, в Швеции в 1965— 1974 г. по­
раженные фенилкетонурией лица встречались с частотой примерно
1 : 40 000.
Исходя из законов Харди — Вайнберга, по локусу, представ­
ленному двумя аллелями, три возможных генотипа (Л , Л , , Л, А 2
и Л 2 Л2) распределяются с частотой р 2
, 2р^, ц2 . Следовательно,
=1/40 000, а 9 = 1/200. Частота доминантного аллеля нормального
обмена фенилаланина равна: р = 1— 9 = 1— 1/200=199/200. Тог­
да,
частота гетерозигот
равна:
2рд=2х 1/200Х199/200=^
=2 х 199/20 000. При найденных частотах доминантного и рецессивно­
го аллелей популяция численностью 40 000 человек содержит одного
больного фенилкетонурией (Л 2Л 2), 400 носителей вредного аллеля в
гетерозиготном состоянии (Л, Л 2). Остальные члены популяции
гомозиготны по благоприятному доминантному аллелю (Л , Л , ).
В табл. 23 приведены данные о частоте гетерозиготного
носительства и соответствующ ей ей частоте гомозиготных индивидуу­
мов с фенотипическим проявлением определенного аллеля. Даже
по редким рецессивным аллелям количество гетерозигот оказывается
достаточно высоким, чтобы это учитывалось при медико-генетическом
консультировании вступающих в брак.
262
Таблица
23.
Соотношение представительства в
популяции гомозигот с соответствующим фенотипическим
(патологическим) признаком и гетерозигот— носителей
определенного аллеля (в пересчете на число членов
популяции)
В с тр е ч а е м о с т ь гомозигот (в пер е­
счете на чи сло чл е н о в п о п ул яц и и )
1:10
1:100
1:1000
1 :1 0 0 0 0
1:100 0 0 0
1:1 0 0 0 0 0 0
В с тр е ч а е м о с т ь гетерозигот
1 2,3
1 5,6
1 16
1 51
1 159 .
1 501
Клеточные и физико-химические механизмы наследственности и
изменчивости универсальны для всех живых суш еств, включая
человека. Очевидна зависимость жизнеспособности о т особенностей
генотипа индивидуума. Больш ую часть истории человечество было
совокупностью более или менее изолированных в репродуктивном
отношении групп. В отдельные исторические периоды происходили
миграции значительных людских контингентов, сопровождавшиеся
объединением ранее разобщенных групп, освоением новых территорий.
В наши дни миграции населения интенсифицировались в связи с ростом
численности людей, совершенствованием средств транспорта, неравно­
мерным развитием экономики. Благодаря отмеченным моментам
генофонды популяций людей испытывали ранее и продолжают
испытывать в настоящее время действие элементарных эволюционных
факторов. Социальность человека вносит в это действие определенную
специфику.
7.5.2. Мутационный процесс
Мутационный процесс у человека сходен с таковым у других живых
форм по всем основным показателям — средней частоте мутирования
на геном за поколение, генетико-физиологическим характеристикам
мутаций, наличию антимутационных барьеров. Такое совпадение
закономерно. Основные параметры спонтанного мутагенеза формиро­
вались на начальных этапах эволюции жизни под действием таких
постоянных факторов, как ультрафиолетовое и иные виды излучения,
температура, определенная химическая среда. В настоящее время
давление мутационного процесса на генофонд человечества, поввдимому, усиливается благодаря росту индуцированных мутаций,
причиной которых нередко служ ат факторы, возникающие в связи
с производственной деятельностью человека в условиях научно-техни­
ческой революции. М утагены антропогенной природы, например
щ щ зщ ую щ ие излучения, повыш аю т число заболеваний, к развитию
которых ведут соматические мутации. Так, при систематическом
йбйучЬййй больных анкилозирующим спондилитом с терапевтическими
263
'
\
д
\
|
.
; V
[ \
\
\
целями пропорциональ­
но дозе облучения росла
вероятность
заболева­
ния лЗД рзом .
7.5.3. Популяцион­
ные волны
Численность населе­
ния планеты за обозри­
мый исторический пери­
од в
целом возрос­
ла. В
эпоху неолита
она составляла пример­
но 5 м лн., в период появ­
ления городов — 20 —
40 м лн., во времена Рим ­
ской империи к на­
чалу
новой эры —
200 млн., к 1600 г. —
500 млн., в 1800 г. —
1 млрд., в настоящее
Рис. 115. Сокращение численности людей в средние
время — более 3 млрд.
века в связи с эпидемией чумы.
человек. Подтверждая
общую тенденцию увеличения народонаселения, эти данные гово­
рят о неравномерном изменении во времени темпов его прирос­
та. М оменты ускорения темпов прироста населения совпадают с
важнейшими достижениями человечества — развитием земледелия
(примерно 10 000 лет назад), началом индустриализации, эрой научнотехнической революции. Важ ны м следствием увеличения темпа приро­
ста служ ит изменение плотности населения. Так, в эпоху охотников и
собирателей она составляла менее 1 человека, в век бронзы и желе­
за — 10 человек, в настоящее время — от 4 до 900— 1200 человек на
3 км2. Крайне неравномерное распределение людей на земном шаре
наблюдалось на любом этапе развития человечества. Ускорение роста
численности при ограниченности заселяемой территории способствует
интенсификации миграций населения.
Н а фоне общей тенденции к увеличению численности населения
отмечались отдельные снижения этого показателя. Причиной сниже­
ния, показанного на рис. 115, явилась эпидемия чум ы , сопровождавшая­
ся большой смертностью и распространившаяся в средние века на
значительные территории. Предполагают, что еще одно временное
сокращение численности населения на территории Европы имело место
в начале мезолита. Причина его состояла в уничтожении племена­
ми, освоившими технику коллективной охоты , основного источника
п и щ и — крупных травоядных животных. Заметные колебания числен­
ности на ограниченных территориях происходили также вследствие
особо опасных инфекций и были закономерными явлениями в жизни
целых народов еще в начале текущего столетия (рис. 116).
264
1000000
500000
100000
0>0 =<МТО'9-Ю<01>СОО> о
г= СМсо
<0г- СО О) О
г: СМ
о > о ° о о о о о о о о
55 55
2 о 2 ® о ® ® ® ® 2 ® о>~ § 2 I Т 2I 2I 2| -I ®|®I 7
7 Т ' 7 7 7 7 7 7 7 7 0.2 = 2 та гг Ю ФгчСО ^ о ^
а>о>О = смС0-ч-иэ<0Ь:а> 0 0)®0)
2 §2 2 В й го
2 а . о ° о о о о о о о
® Ц ) 0 ) ® 0 ) 0 ! й © ® 0 ) в
\
Годы
Рис. 116. Смертность о т'чум ы в Индии по годам (1898— 1922 гг.).
Периодические колебания численности населения на обширных или
ограниченных территориях, изменяя плотность населения и вы зывая
миграции, влияли на состояние генофондов человеческих популяций.
7.5.4. Изоляция
Человеческое общество длительно развивалось как совокупность
изолированных производственных коллективов, внутри которых в ос­
новном и совершались браки. Природа изоляционных барьеров между
популяциями людей разнообразна. В ранней истории человечества
важное место принадлежало, по-видимому, географической изоляции.
Специфическими для человеческого общества являю тся формы изоля­
ции, зависящие от разнообразия культур, экономических укладов,
религиозных и морально-этических установок. Фактор изоляции
оказывал влияние на генофонды популяций людей. Длительным
проживанием в состоянии относительной культурной и географической
изоляции объясняю т, например, некоторые антропологические осо­
бенности представителей малых народностей: своеобразный рельеф
ушной раковины бушменов, больш ую ширину нижнечелюстного
диаметра коряков и ительменов, исключительное развитие бороды
айнов. Среди горных таджиков, проживающих в одном районе,
выделены группы с разным соотношением индивидуумов по антигенам
эритроцитов системы АВО. Указанные различия коррелируют со
степенью изолированности от главных перевальных путей сообщения.
Сохранению высокого уровня генетической изоляции двух популяций,
существующих на одной территории, способствуют отличия по
физическим признакам или образу жизни. Однако такие барьеры со
временем ослабевают, о чем свидетельствует судьба популяций белых и
265
V
Рис. 117. Неравномерное распределение аллеля
Европе.
В
системы групп крови
АВО
в
негров С Ш А и Бразилии — к настоящему времени доля генов от белых
составляет у американских негров 25% и у бразильских — 4 0 % . Между
двумя генетически различающимися популяциями, разделенными ге­
ографически, иногда вклиниваю тся другие популяции, через которые и
происходит обмен генами. В таких случаях наблюдается градиент
признака. Так, частота аллеля группы крови В в Европе постепенно
повыш ается с запада на восток (рис. 117). У коренного населения
Пиренейского полуострова этот ген практически отсутствует, тогда как
в районе Астрахани его частота достигает 30% .
В настоящее время круг возможных браков неуклонно расширяется.
Разрушение многовековых изоляционных барьеров — процесс, повидимому, необратимый.
7.5.5. Генетико-автоматические процессы
Предполагают, что человеческие сообщества в палеолите состояли
из нескольких сотен индивидуумов. Всего 1—2 столетия том у назад
люди жили преимущественно поселениями, состоящими из 25— 35 до­
мов. Вплоть до самого последнего времени количество индивидуумов
в отдельных популяциях, непосредственно участвующ их в размноже­
нии, редко превышало 400— 3500 человек. Причины географического,
экономического, расового, религиозного, культурного порядка ограни­
чивали брачные связи масштабами определенного района, племени,
поселения, секты. Высокая степень репродуктивной изоляции мало­
численных человеческих популяций на протяжении многих поколений
создавала благоприятные условия для дрейфа генов.
266
Генетико-автоматические процессы приводят к сглаживанию
изменчивости внутри группы и появлению случайных различий между
изолятами. Именно это выявили наблюдения за особенностями
фенотипов малочисленных групп населения, например в условиях
географической изоляции. Так, среди населения Памира резусотрицательные индивидуумы встречаются в целом в 2— 3 раза реже,
чем в Европе. В большинстве кишлаков такие люди составляю т 3— 5%
популяции, а в некоторых изолированных селениях их насчитывается
до 15%. В крови человека имеются гаптоглобины, которые связываю т
свободный гемоглобин после разрушения эритроцитов, чем предотвра­
щ ают его выведение из организма. Синтез гаптоглобина Нр 1— 1
контролируется геном Н Р 1, частота которого у представителей двух
соседних племен на севере Южной .Америки составляет 0,205 и 0,895,
отличаясь более чем в 4 раза. Примером действия дрейфа генов
в человеческих популяциях служит « э ф ф е к т
родоначаль­
н и к а». Он возникает, когда несколько семей порывают с роди­
тельской популяцией и создают новую на другой террито­
рии, причем такая популяция обычно поддерживает высокий уро­
вень брачной изоляции, способствуюший случайному закреплению
в ее генофонде одних аллелей и элиминации других. В результате
частота даже очень редкого аллеля может стать значительной. Так,
члены секты меннонитов (амишей) в округе Ланкастер ш тата Пенсиль­
вания, насчитывающей к середине текущего столетия примерно 8000
человек, почти все произошли от трех супружеских пар, иммигриро­
вавших в Америку в 1770 г. В этом изоляте обнаружено 55 случаев
особой формы карликовости с многопалостью, которая наследуется по
аутосомно-рецессивному типу и не зарегистрирована среди меннонитов
ш татов Огайо и Индиана. В мировой медицинской литературе описано
едва ли 50 таких случаев . В X V III веке из Германии в СШ А иммигриро­
вало 27 семей, основавших в ш тате Пенсильвания секту дункеров.
За 200-летний период существования в условиях сильной брачной
изоляции генофонд популяции дункеров изменился в сравнении с гено­
фондом населения Рейнской области Германии, причем степень
различий во времени увеличивалась. У лиц в возрасте 55 лет и выше
частоты аллелей системы групп крови
ближе к цифрам, типичным
для населения Рейнской области, чем у лиц в возрасте 28— 55 лет (табл.
24). В возрастной группе 3—27 лет этот сдвиг достигает еще больших
значений.
Т а б л и ц а 24. Прогрессивное изменение концентрации аллелей системы групп
кррви М N в популяции дункеров
Концентрация аллеля
Возраст членов изолята,
лет
1М
Более 55
28-5 5
3— 27
0,45
0,34
0,265
0,55
0,66
0,735
267
I
6
Рис. 118. Развитие нейтральных признаков в изоляте (секта дункеров штата Пен­
сильвания).
а — р о с т в о л о с н а ср е д н е й ф а л а н г е п а л ь ц е в к и с т и ; б — с п о с о б н о с т ь о т с т а в л я т ь б о л ь ш о й пален руки
Ро ст среди дункеров лиц с группой крови М и снижение — с группой
крови N нельзя объяснить действием отбора, так как направление
изменений не совпадает с таковым для населения ш тата Пенсильвания.
В пользу дрейфа генов говорит также то, что в генофонде американских
дункеров увеличилась концентрация аллелей, контролирующих разви­
тие нейтральных признаков, например оволосения средней фаланги
пальцев, способности отставлять большой палец руки (рис. 118).
Н а протяжении большей части истории человечества дрейф генов
оказывал влияние на формирование генофондов популяций людей. Так,
многие особенности узколокальных типов в пределах арктической,
байкальской, центральноазиатской, уральской групп населения Сибири
служ ат, по-видимому, результатом генетико-автоматических процессов
в условиях изоляции малочисленных коллективов. Э ти процессы,
однако, не имели решающего значения в дифференциации и длительной
эволюции человека.
Последствия дрейфа генов, представляющие интерес для медицины,
заклю чаю тся в неравномерном распределении некоторых наследствен­
ных заболеваний по группам населения земного шара. Так, изоляцией и
дрейфом генов объясняется, по-видимому, относительно высокая
частота церебромакулярной дегенерации в Квебеке и Ньюфаундленде,
детского цестиноза во Франции, алкаптонурии в Ч С С Р , одного из типов
порфирии среди белых колонистов в Ю жной Африке, адреногенитального синдрома у эскимосов. Э ти же факторы могли бы ть причиной
низкой частоты фенилкетонурии у финнов и евреев-ашкенази.
7.5.6. Естественный отбор
В процессе видообразования естественный отбор переводит
случайную индивидуальную изменчивость в биологически полезную
групповую — популяционную, видовую. Стабилизирующ ая его форма
сохраняет «удачные» комбинации аллелей о т предшествующих этапов
эволюции. Отбор поддерживает также состояние генетического поли­
морфизма. Смена биологических факторов исторического развития
социальными привела к том у, что в человеческих популяциях естественный отбор утратил функциювидообразования. За ним сбхрайИГ-'
268
лись функции стабилизации
генофондов и поддержания на'следственнего .дш нообхщ ия.
' В~пользу действия стаби­
лизирующей формы естест­
венного отбора говорит, на­
пример, большая перинаталь­
ная смертность среди недоно­
шенных и переношенных ново­
рожденных. Направление от­
бора среди таких детей зави­
сит, по-видимому, от сниже­
ния обшей жизнеспособности.
Отрицательный
отбор
по
одному локусу можно проил­
люстрировать на примере си­
стемы антигенов резус. Около
85% населения Европы имеет
в эритроцитах антиген КЬ и
образует группу резус-положительных
индивидуумов,
остальная часть европейской
популяции лишена этого анти­
гена и является резус-отрицаРис. 119. Генетическая основа резус-конф
тельной. Синтез антигена КЬ
контролируется
доминант­ ликта.
а - !-я бер ем ен н ость; б
2-я б ер е м ен н о е !ь. 1
р езус
ным аллелем (О ) и происходит антигены; 2 - ан ти тел а к резус-антигену
р организмах с генотипами
Б Б и М . Резус-отрицательные люди являю тся рецессивными
гомозиготами (йй). При беременности резус-отрицательной жен­
щины (йй) резус-положительным плодом (мужчина Б Б или Ш ,
плод Б й ) при нарушении целостности плаценты Б +-эритроциты плода проникают в организм матери и иммунизируют его
(рис. 119). При последующей беременности резус-положительным
плодом (Ш ) антирезус-антитела проникают через плаценту'в организм
плода и разруш ают его эритроциты. Развивается гемолитическая
болезнь новорожденного, ведущим симптомом которой служит
тяжелая анемия. В настоящее время медицина располагает способами
борьбы с этой формой патологии в виде быстрого переливания
младенцу резус-отрицательной крови или введения антирезус-антител
для предотвращения иммунизации матери. В отсутствие медицинской
помощи новорожденный с гемолитической болезнью нередко погибал.
Учиты вая резус-отрицательный фенотип матери, резус-положительный
плод генотипически является гетерозиготой (Ь й ). Э то означает, что со
смертью такого индивидуума из генофонда популяции, к которой он
принадлежит, удаляется равное количество доминантных и ре­
цессивных аллелей локуса «резус». Отбор в данном случае направлен
против
г е т е р о з и г о т . При неравенстве исходных частот
элиминируемых аллелей такой отбор приводит к постепенному
769
I
снижению доли более редкого из них, который в европейской популяции
представлен рецессивным аллелем ((I). Подсчитано, что снижение его
доли с 15 до 1% путем отбора против гетерозигот потребует
600 поколений или около 15 ООО лет.
Под действием естественного отбора находятся аллели, контроли­
рующие синтез антигенов системы групп крови АВО. Об этом
свидетельствует снижение против ожидаемого количества детей в АВО
несовместимых браках женщин 0 группы с мужчинами А , В и А В групп
(табл. 25).
Таблица
25.
Влияние АВО несовместимости на плодовитость и эмбриогенез
П о казател ь
Число обследованных семей
Бесплодные браки, %
Аборты на 1 беременность, %
Бездетные браки, %
Среднее число детей на 1 мать
А В О совм ести м ы е
А В О несовм естим ые
812
8,13
10,34
9,85
2,596
617
11,7
15,3
18,15
2,173
Особенность отбора по локусам группоспецифичных антигенов
эритроцитов, таких как резус или АВО заключается в том , что он
действует на уровне зиготы или ранних стадий эмбриогенеза. Замечено,
что в браках, где м ать и отец несовместимы по резус-фактору, число де­
тей с гемолитической болезнью меньше ожидаемого. Так, среди белого
населения С Ш А резус-отрицательные люди составляю т примерно 16%.
В такой популяции частота аллеля Нравна 4 0 % , а вероятность развития
гемолитической болезни при случайном подборе брачных пар — 9 % . На
самом деле частота этой болезни составляет 1 : 150— 1 : 200. Одной из
причин этого служит одновременная несовместимость родителей по
локусу АВО . В этом случае В +-эритроциты плода, попадающие в
организм матери, разруш аются и иммунизации не происходит.
Отрицательный отбор действует в большинстве популяций людей
по аллелям аномальных гемоглобинов. Его особая жесткость обуслов­
ливается тем , что он направлен п р о т и в г о м о з и г о т . Ребенок,
умирающий, например, от серповидноклеточной анемии, является
гомозиготным по аллелю 5. Каждая такая смерть элиминирует из
генофонда популяции аллели одного вида, что приводит к сравнительно
быстрому снижению изменчивости
по соответствующ ему локусу.
Во многих популяциях людей частота аллелей аномальных гемоглоби­
нов, в том числе и X, не превышает 1 % .
Высокая частота аллелей таких аномальных гемоглобинов, как 5, С,
Д
Е в некоторых районах планеты иллюстрирует действие
естественного отбора по поддержанию в человеческих популяциях
состояния балансированного генетического полиморфизма. Отрица­
тельный отбор в отношении аллеля 5 перекрывается мощным
положительным отбором гетерозигот НЪА/НЬ8 благодаря высокой
жизнеспособности последних в очагах тропической малярии (рис. 120).
Исследования в Уганде показали, что количество возбудителей маляио
б
Рис. 120. Совпадение районов распространения в Старом Свете:
а
а л л е л я сер п ови дн оклеточно сти , б
ал л ел я ( в п р оц ентах).
тр о пи ческо й м ал яр и и ; цифры о б о з н а ч а ю т концентрации»
рии в 1 мл крови зависит от генотипа ребенка и составляет до
10 ООО у Н Ь8/Н Ь8, до 160 ООО у НЬА/НЬ8 и до 800 ООО у НЬА/НЬА .
В Северной Греции обследовали 48 семей, в которых наблюдались и
серповидноклеточность, и малярия. Среди братьев и сестер больных
серповидноклеточной анемией болело малярией 16 из 25 с генотипами
НЬА/НЬА и 1 из 23 с генотипами Н ЬА/Н ЬБ. Таким образом нормальные
люди болели в 13 раз чаше, чем индивидуумы с аномальными
гемоглобинами. Устранение фактора контротбора приводит
к
снижению частоты аллеля серповиднокпеточности. Этой причиной,
действующей на протяжении уже нескольких столетий, наряду с
метисацией, объясняю т относительно низкую частоту гетерозигот
НЬА/НЬ8 среди североамериканских негров (8— 9 % ) в сравнении с
африканскими (около 2 0 % ).
В приведенных примерах действию отрицательного отбора,
снижающего в генофондах некоторых популяций людей концентрацию
определенных аллелей, противостоят к о и т р о т б о р ы , которые
поддерживают частоту этих аллелей на достаточно высоком уровне.
Результатом
наложения многочисленных и разнонаправленных
векторов отбора служит формирование и поддержание генофондов
популяций в состоянии, обеспечивающем возникновение в каждом
поколении генотипов достаточной приспособленности с учетом мес­
тных условий. Благодаря социально-экономическим преобразованиям,
успехам лечебной и особенно профилактической медицины влияние
отбора на генетический состав популяций людей снижается.
7.5.7. Генетическое разнообразие
и генетический груз в популяциях людей
Человечеству свойствен высокий уровень наследственного разно­
образия, что проявляется в многообразии фенотипов. Люди отлича­
ю тся друг от друга цветом кожных покровов, глаз, волос, формой носа
и ушной раковины, рисунком эпидермальных гребней на подушечках
пальцев и другими сложными признаками. Выявлены многочисленные
варианты отдельных белков, различающихся по одному или нескольким
аминокислотным остаткам , а следовательно, и функционально. Бел­
ки — это простые признаки и прямо отражаю т генетическую консти­
туцию организма. У людей не совпадают группы крови по системам
эритроцитарных антигенов резус, АВО и другим. Известно более 130
вариантов гемоглобина, более 70 вариантов фермента глюкоз о-6-фосидрогеназы (Г-6-Ф Д Г), который участвует в бескислородном
лении глюкозы в эритроцитах. В целом не менее 30% генов,
пирующих у человека синтез ферментных и других белков, имеет
*ко аллельных форм. Ч астота встречаемости разных аллелей
гена варьирует. Так, из многих вариантов гемоглобина лиш ь 4
обнаруживаются в некоторых популяциях в высокой концентр
ции — НЪ5 (тропическая Африка, Средиземноморье), Н ЬС (Западная
Африка), Н ЬИ (И ндия), Н Ь Е (Ю го-восточная А зия). Концентрация
других аллелей гемоглобина повсеместно не превышает, видимо,
0,01—0,0001%. Вариабельность распространенности аллелей в популя­
Ш
циях людей зависит от действия элементарных эволюционных
факторов, при этом важная роль принадлежит мутационному процессу,
естественному отбору, генетико-автоматическим процессам, мигра­
циям.
М утационный процесс создает новые аллели и действует
в человеческих популяциях ненаправленно, случайным образом, поэто­
му мутации не приводят к выраженному преобладанию концентраций
одних аллелей над другими. В достаточно большой популяции,
в которой каждая пара ролителей из поколения в поколение дает двух
потомков, вероятность сохранения новой нейтральной мутации через 15
поколений составляет всего 1/9. Все многообразие вариантов белков,
отражающее разнообразие аллелей в генофонде человечества, можно
разделить на две группы. К однОй из них относятся редкие варианты,
встречающиеся повсеместно с частотой менее 1 % . Появление их
объясняется исключительно мутационным процессом. Вторую группу
составляю т варианты, обнаруживаемые относительно часто в избран­
ных популяциях. Так, в примере с гемоглобинами к первой группе
относятся все варианты, кроме НЪ5, НЬС, Н Ь Б и Н Ь Е. Длительные
различия в концентрации отдельных аллелей между популяциями,
сохранение в достаточно высокой концентрации нескольких аллелей
в одной популяции зависят от действия естественного отбора или
дрейфа генов.
К межпопуляционным различиям в концентрации определенных
аллелей приводит стабилизирующая форма естественного отбора.
Неслучайное распределение по планете аллелей эритроцитарных анти­
генов АВО может бы ть обусловлено различной выживаемостью лиц,
отличающихся по группе крови, в условиях частых эпидемий особо
опасных инфекций. Области сравнительно низких частот аллеля 1° и
относительно высоких частот аллеля 1в в Азии примерно совпадают
с очагами чум ы , возбудитель которой имеет Н-подобный антиген. Э то
делает людей с группой крови 0 особенно восприимчивыми к чуме, так
как они, имея Н-антиген, не способны вы рабаты вать специфические
антитела в достаточном количестве. Указанному объяснению со­
ответствует факт, что относительно высокие концентрации аллеля 1°
обнаруживаются в популяциях аборигенов Австралии и Полинезии,
индейцев Америки, которые практически не поражались чумой. Вместе
с тем для популяций из одного географического района, но
изолированных в репродуктивном отношении, причиной различий в
концентрации аллелей АВО мог бы ть дрейф генов. Так, частота группы
крови А достигает у индейцев племени черноногих 8 0 % , а у индейцев из
ш тата Ю та 2 % .
В основе стойкого сохранения в популяции людей одновременно
нескольких аллелей одного гена лежит, как правило, отбор в пользу
Сетерозигот, который ведет к состоянию балансированного поли­
морфизма. Классическим примером такой ситуации служит распро­
странение аллелей гемоглобинов 8, С и Е в очагах тропической
малярии.
Выш е приведены примеры полиморфизма по конкретным локусам,
который объясняется действием известного фактора отбора. В есте27.?
-V
И
ственных условиях в силу воздействия на фенотипы организмов спектра
факторов отбор осущ ествляется по многим направлениям, в результате
чего формируются генофонды, сбалансированные по набору и частотам
аллелей, обеспечивающие в данных условиях обитания достаточную
выживаемость популяций. Э то справедливо и для популяций людей.
Так, люди с группой крови 0 более восприимчивы к чуме, чем люди с
групгЙУЙ крови В . Туберкулез легких у них лечится с большим трудом,
чем у лиц с группой крови А . Вместе с тем при одинаковой вероятности
заболевания при контакте с возбудителем сифилиса лечецце людей с
группой крови 0 быстрее вы зывает переход болезни в неактивную
стадию. Д ля лиц с группой крови 0 вероятность заболеть раком желуд­
ка, раком шейки матки, ревматизмом, ишемической болезнью сердца,
холециститом, желчнокаменной болезнью примерно на 20% ниже, чем
для лиц с группой крови А . Генетический полиморфизм по многим
локусам мог бы ть унаследован людьми от предков на досапиентной
стадии развития. Полиморфизм по таким системам групп крови, как
АВО и резус, обнаружен у человекообразных обезьян.
Факторы отбора, действие которых создавало современную
картину распределения аллелей в популяциях людей, для подавляющего
больш инства локусов точно не установлены, однако примеры,
рассмотренные выш е, указываю т на их экологическую природу.
Учиты вая слабую техническую оснащенность, плохие экономические и
гигиенические условия жизни основной массы населения планеты на
протяжении значительной части истории человечества, больш ую роль в
этом отношении играли, по-видимому, возбудители особо опасных
инфекций, паразитарных заболеваний, туберкулеза. В описанных усло­
виях наследственный полиморфизм способствовал расселению людей,
обусловливая удовлетворительную жизнеспособность в разных эколо­
гических ситуациях. Определенный вклад в наблюдаемое распределе­
ние аллелей внесли массовые миграции населения и сопутствую щ ая им
метисация. В период до великих географических открытий и начала
колониальной экспансии смешение больших контингентов людей
разной расовой принадлежности имело место в Восточной Африке,
Индии, Средней Азии, Западной Сибири, Индокитае, а впоследствии
в Ю жной и Центральной Америке.
Генетический полиморфизм служит основой межпопуляционной и
внутрипопуляционной изменчивости людей, которая проявляется в не­
равномерном распределении по планете некоторых заболеваний,
тяжести их течения в разных человеческих популяциях, разной степени
предрасположенности людей к определенным болезням, индивиду­
альных особенностях течения патологических процессов, различиях
в реакции на лечебное воздействие. Наследственное разнообразие
долгое время было препятствием для успешного переливания крови.
В настоящее время оно создает большие трудности в решении
проблемы пересадок тканей и органов.
Также как в популяциях других организмов наследственное
разнообразие снижает реальную приспособленность популяций людей.
Бремя генетического груза человечества можно оценить, введя понятие
л е т а л ь н ы х э к в и в а л е н т о в . С читаю т, что число их у от-
\у/, '>. г„.._
^
дельных людей колеблется от 3 до 8. Э то означает, что то значительное
количество неблагоприятных аллелей, которое есть в генотипе каждого
человека, оказывает суммарное вредное действие, эквивалентное
действию 3— 8 рецессивных аллелей, приводящих в гомозиготном
состоянии к смерти индивидуума до наступления репродуктивного
возраста. Благодаря наличию неблагоприятных аллелей и их сочетаний
примерно половина зигот, образующихся в каждом поколении людей,
в биологическом плане несостоятельна, т. е. не участвует в передаче
генов следующему поколению. Около 15% зачатых организмов гибнет
до рождения, 3% — при рождении, 2 % — непосредственно после
рождения. 3% людей умирает, не достигнув половой зрелости, 20% лиц
не вступает в брак и 10% браков бездетны. Исторические перспективы
человека в силу его социальной сущности не связаны с генетической
информацией, накопленной видом Н ото $ар1еп& в ходе эволюций. Тем не
менее человечество продолжает «оплачивать» эти перспективы, теряя в
каждом поколении часть своих членов в силу их генетической
несостоятельности.
1|
1
7.6. М И К Р О - И М А К Р О Э В О Л Ю Ц И Я
;I
Действий элементарных эволюционных факторов приводит к ди­
вергенции популяций организмов вплоть до образования новых видов.
Особи из разных популяций способны скрещиваться друг с другом и
таким образом обмениваться наследственным материалом, поэтому до
момента видообразования периодически происходит нивелировка
межпопуляционных различий. Рождение нового вида означает полную
репродуктивную изоляцию двух групп организмов. Э то устраняет
возможность сглаживания различий между ними. Представители
разных видов отличаю тся конкретными адаптациями, которые были
приобретены ими на предшествующих этапах эволюции. Историко­
географическая судьба и экологическая ситуация тех территорий,
которые они заселяю т, имеют свои особенности, поэтому развитие
совокупностей организмов видового ранга в условиях межвидовой
конкуренции обычно не совпадает как по направлению, так и по темпам
дальнейших изменений. Н а любом этапе своего существования органи­
ческий мир представляет сложную иерархическую систему групп
(таксонов) организмов, находящихся в состоянии репродуктивной
изоляции, но связанных друг с другом той или иной степенью родства.
Э то объясняется наличием в едином процессе исторического развития
природы двух сторон, различающихся по результатам ., Одна, из
щ д ,— видообразование или . м и к р о э в о л ю ц и я *
вторая —
формирование
системы
таксонов
надвидового
ранга
или
м а к р’ТТ’» '* о л * о и и я.
В силу растянутости процесса во времени закономерности
макроэволюции нельзя изучать путем прямых наблюдений или
экспериментально. Вскры ть их помогают данные палеонтологии,
сравнительной эмбриологии и сравнительной морфологии, а в настоя­
щее время также данные молекулярной биологии и генетики. Степень
275
.
филогенетического родства в этом случае определяют по доле обших
генов или различиям в первичной структуре белков.
Единого мнения о природе факторов макроэволюции нет. Согласно
одной из гипотез сущ ествуют общие факторы микро- и макроэволюционных изменений организмов и их групп. Несовпадение результатов
названных двух сторон процесса исторического развития объясняется
тем, что во втором случае эти факторы действуют в условиях сложных
межвидовых отношений в отсутствие нивелировки возникающих
различий. Э то приводит к изменению силы и направления давления
элементарных эволюционных факторов в разных группах, что замедля­
ет или ускоряет эволюционные преобразования, делает возможным
резкие уклонения направления развития одних групп по сравнению с
другими. В итоге органический мир образуют таксоны нескольких
уровней, каждый из которых представляет группу родственных видов,
родов, семейств и т. д. Степень отличий представителей разных
таксонов в целом соответствует времени дивергенции. Чем выше ранг
сравниваемых таксонов, тем раньше в филогенезе началось их
дивергентное развитие. Отдельные таксоны различаются и по
разнообразию входящих в них групп более низкого ранга.
Благодаря действию элементарных эволюционных факторов на
внутривидовом уровне формируются разнообразные адаптации. В про­
цессе исторического развития, растягивающегося на Миллионы лет,
выясняется универсальное значение некоторых из них. Такие адаптации
обеспечивают достаточную выживаемость в широком спектре условий
обитания и служат основой широкой дивергенции видов. Они
подхватываются и усиливаются естественным отбором, в чем и заклю­
чается его творческая функция. Эволю ция соответствующ ей группы
организмов ускоряется. В недрах этой группы образуется много видов,
которые, продолжая развитие, порождают целые «пучки» новых видов.
Отдельные адаптации и их сочетания становятся существенным приз­
наком определенного типа морфофизиологической организации,
характерной чертой всех представителей крупного таксона.
Особенность иерархической системы таксонов надвидового ранга,
которая складывалась в процессе макроэволюции, заключается в нали­
чии групп живых существ, отличающихся по сложности структурно­
функциональной- организации, уровню активности
по отношению
к среде. При этом более сложно устроенные и активные формы
появляю тся в эволюции позже менее сложных. Вместе с тем на планете
сосущ ествуют организмы, неизмеримо далекие по уровню и плану
строения, например микроорганизмы, простейшие, беспозвоночные,
млекопитающие, человек. Некоторые из просто организованных
существ имеют то же повсеместное распространение, что и ряд
высокоорганизованных форм. Х о тя поддержание жизни во времени
возможно по достижении сравнительно низких уровней структурно­
физиологической организации, процесс исторического развития в целом
имеет направленный, прогрессивный характер, что обеспечивается
ходом макроэволюции. К настоящему времени установлено много
фактов, освещающих разные стороны макроэволюционных преобразо­
ваний живой природы, сделаны важные эмпирические обобщения. Они
#
отражаю т закономерности эволюции групп1^эргани1к
Организмов на надвидовом
уровне, органов и систем органов, вопросы соотношения фило- и
онтогенеза, неограниченного, биологического и морфофизиологическо­
го прогресса в историческом развитии, антропогенеза.
7.7. Э В О Л Ю Ц И Я ГР У П П О РГ А Н И З М О В
7.7.1. Формы эволюции групп
Выделяю т несколько Ф о Р м . 3 вго л Ш Д л а . г,Р У п п. живых
существ. Одной из них я в л я е т а и М г я и т 'и д ^ ^ ^ ^ З А ^ Л ^ ^ и Ц и и ш
затрагивающая представителей одТОНГ*таксона, которы йвоврем ени
изменяется в определенном направлении как единое целое. Примером
может служить линия развития семейства лошадиных, в процессе
которого наблюдается постепенная смена древесноядных форм траво­
ядными и сокращение числа пальцев до одного (рис. 121). Н а микроэволюциолном уровне этой форме макроэволюции соответствуёт филити_ ческое. видообразование. Щде . одна форма ^эволюции представлена
‘ г г ^ 1 и т ц., 1 н _ з в . о д ю ц и
которая заключается в образотГёнии в процессе исторического развития нескольких новых групп от
одной предковой. Она создает разнообразие таксонов низшего ранга
в более крупных таксонах, например подклассов и отрядов в классе,
семейств в отряде. Н а микроэволюционном уровне ей соответствует
дивергентное видообразование. Рассмотренные формы эволюции отра­
ж аю т "ход"Исторического развития отдельных групп организмов.
Формами соотносительного развития цфжолыца ^а^рон^в^являются
М *3 $ а л л е л и зт^ и У б н в е с г е н Т н а я з в о. л ю .ц .у ; В перслучае два таксотаг дйвергпровавшие о т общего предка и,
следовательно, имеющие общую генетическую основу, в дальнейшем
претерпевают филитииажую эдодюцию в еходномнаправлении. В оли­
гоцене, например, саблезубость возникла у махайродонтов (Нор1орЬопеи$) и лжесаблезубых настоящих кошек (Й ш с т з ). Этап параллельного
развития в эволюции групп может смениться этапом конвергенции,
которая представляет собой Филитическую .эволюцию нррг>пг.тш>нныу
таксонов в сходном направлении. Конвергенция наблюдается, когда
представители разных групп встречаются со сходными экологическими
задачами. Так, конвергентное развитие формы тела происходило у
первичноводных (акуловые) и вторичноводных (китообразные)
животных. Генофонды конвергентно эволюционирующих неродствен­
ных групп различны, поэтому конвергентное сходство обычно
распространяется лиш ь на некоторые признаки. Так, акуловые и кито­
образные, сходные по форме тела, имеют резкие различия в строении
кровеносной системы, мускулатуры, покровов. Основные формы
эволюции групп представлены схемами на рис. 122.
7.7.2. Типы эволюции групп
В зависимости от масш таба адаптаций, на основе которых
происходит очередной этап эволюции, выделяю т два ее т и п а — алло­
генез и арогенез.
277
Рис. 121. Схема филитической эволюции в семействе Лошадиных
1 — ЕоЫрриз; 2
М ю Ы ррив; 3 — РагаЫрри&,в4 — Р1юЫрри$, 5 — Ецииз.
В2
I
А|
1
В2
\ / I
А ,
А д
\\ //
д
.
',
I
В(
\ /
д
1
В,
I / \
А)
А2
\\
А|
/
/
/
\
/
/
А
А
I
I
I
I
.
2
'
В2
Б
/
\\
\
Б
I
,
3
|
I
Рис.
групп
122. Формы
эволю ци и
1 "
ф и л н тн ч е с к а я ; 2 — ди вер ген тн а я; 3 — п а р а л л е л ь н а я ; 4
конверген тн ая. А , Б — исходное состо яни е
групп; А ^ , Б , — п р ом еж уто чн ы е
стадии
н а д п п эво
тоол
. нюиц
ц ии
г п ,; В
ж
-*,
,
— состояние Гр
упп в дан н ы й момент; Т
групп
время.
д
Рис. 123. Аллогенез в отряде насекомоядных млекопитающих.
Н а зе м н ы е ф орм ы: а — п р ы гу н чи к; б — зе м л е р ой ка; в — е ж ; зем н овод ны е ф орм ы: г —
в ы д р о в а я зе м л е р о й к а; е — вы х ух о л ь; подзем норою щ не ф орм ы: ж — к р от; з — зл ато кр о т.
к ут о р а; д —
Под а л л о г е н е з о н понимают развитие с сохранением у всех
представителей таксона высокого ранга главных черт структурнофизиолоГйческой организации. ^Н а этом фоне отдельные группы
организмов внутри таксона развиваю тся путем приобретения сопоста­
вимых по масш табу адаптаций. В результате эволюция осущ ествляется
в пределах одной а д а п т и в н о й з о н ы — совокупности экологи­
ческих ниш, различающихся в деталях, но сходных по степени давления
среды на организмы определенного морфофизиологического типа.
Примером аллогенеза служ ит разнообразие экологических форм в
отряде насекомоядных млекопитающих животных (рис. 123). Если
аллогенез происходит путем приобретения приспособлений к узколо­
кальным условиям существования, то говорят о развитии по типу
специализации.
Примером крайних форм специализации
279
Рис. 124. Типы эволюции групп.
а — арогенез; 6 — аллогенез; в — морфофнзиологический регресс.
служит удивительное соответствие строения цветка некоторых видов
растений определенному виду опылителей.
Под а р о г е н е з о м
понимают развитие с появлением у
представителей некоторых групп внутри крупного таксона таких черт
строения или физиологии, которые делают возможным их выход
н другую апяптивную зону. Так, своим возникновением и направления^
ми дальнейшего исторического развития класс птиц обязан появлению
крыла, четырехкамерного сердца, совершенствованию отделов го­
ловного мозга, участвующ их в координации двигательной активности,
теплокровности. Арогбнезами более крупного масш таба являю тся
возникновение эукариотического типа клеточной организации, мно­
гоклеточное™ .
Выход группы живых сущ еств в новую адаптивную зону
в результате арогенеза сопровождается ее
развитием по типу
аллогенеза и специализации с освоением всех подходящих экологических
ниш. Так, среди птиц выделяю т лесных, водоплавающих, горных,
равнинных.
Тип эволюции, при котором переход в новую адаптивную зону
сопровождается упрощением строения и физиологии организмов,
называется ^ й о р ф о ф и з и о л о г и ч е с к и м
регрессом.
Последний "Свойствен многим паразитам, которые составляю т, повидимому, не менее 4— 5 % всех современных видов животных.
Основные типы эволюции групп представлены на рис. 124.
Адаптации, составляющие основу разных типов эволюции групп,
в равной мере заклю чаю тся в изменении структурных или функцио­
нальных параметров органов, систем органов, механизмов реагирова­
ния на действие факторов среды обитания. Приспособительные из­
менения органов и систем организма, обусловливающие развитие
по типу аллогенеза, называю т и д и о а д а п т а ц и я м и ( а л л о ­
м о р ф о з а м и), по типу арогенеза — а р о м о р ф о з а м и ,
по тапу регресса — м о р ф о ф и з и о л о г и ч е с к о й д е г е ­
н е р а ц и е й . В процессе исторического развития крупного так­
сона наблодается смена типов развития. Группа живых существ,
вышедшая на данном этапе вследствие ароморфозов в новую
адаптивную зону, на следующем этапе характеризуется обычно в целом
280
развитием по типу аллогенеза, а в некоторых своих ветвях — по типу
специализации или морфофизиологического регресса. К определенному
моменту внутри такой группы может сложиться совокупность
организмов, накапливающих новые ароморфозы, что создает предпо­
сылки к смене типа эволюции на арогенез. (Специализация и регресс
соответствую т тупиковым ветвям эволюции, поэтому группы, развива­
ющиеся таким образом, при смене условий обитания вымираю т.
Сменой типов эволюции, различиями в стабильности адаптаций и
условий обитания объясняется значительная пестрота современного
органического мира по уровням структурно-физиологиЧеской организа­
ции и разнообразию специализаций. Так, сосуществуют--д ысшие и
низшие позвоночные, например млекопитающие и ам ф иб^. Вместе
с представителями процветающей группы костистых рыб в фауне
планеты сохраняется кистеперая рыба латимерия, отлйчающ аяся
строением, типичным для группы, вымершей т ч т и -целиком
200—300 млн. лет назад.
7.7.3. Правила эволюции групп
Эмпирическим путем установлен ряд п р а в и л э в о л ю ц и и
г р у п п . П равило^ е о б р а т и м о с т и э в о л ю ц и и [Л . Долло,
1893] утверждает, что в процессе исторического развития невозможен
возврат какой-либо группы организмов в состояние, уже пройденное ею
ранее. Генетическую основу его составляет бесконечно малая статисти­
ческая вероятность возникновения двух идентичных генофондов или
даже генотипов. Благодаря наличию обратных мутаций в филогенезе,
однако, возможно повторное возникновение отдельных простых
признаков.
р>
Согласно п р а в и л у ! п р о г р е с с и в н о й с п е ц и а л и з а ц и и
[Ш . Депере, 1876] группё, начавшая эволюционировать по этому типу, в
дальнейшем идет только по пути углубления специализации. Генетическую основу этого правила составляет ограниченность генофонда
соответствующ ей группы организмов и его формирование в филогенезе
под действием определенных факторов конкретной адаптивной зоны
или экологической ниши. Больш ое значение имеют также экологические
связи, которые накладывают свои ограничения. Так, в целом эволюция
позвоночных животных шла по пути увеличения размеров тела. Однако
освоение грызунами подземного образа жизни сопровождалось умень­
шением размеров тела. Аналогичным образом изменялся указанный
признак в ряду предков хищника ласки (Ми$(е1а шуаНх), питающегося
мышевидньдои грызунами.
Правило' п р о и с х о ж д е н и я
от н е с п е ц и а л и з и р о ­
в а н н ы х п ф е д к о в [Э . Коп , 1904] утверждает, что новые груп­
пы организмов происходят о т менее специализированных представи­
телей предковой формы. Основой его служит большее разнообразие
способов борьбы за существование в малоспециализированных груп­
пах. О тсутствие глубоких специализаций облегчает появление разно­
образных адаптаций, вклю чая и принципиально новые.
281
7.8. Э В О Л Ю Ц И О Н Н Ы Й П Р О Г Р Е С С
В зависимости от применяемых критериев и подходов выделяю т
несколько ф о р м э в о л ю ц и о н н о г о п р о г р е с с а .
7.8.1. Неограниченный прогресс
В условиях нашей планеты развитие жизни как особой формы дви­
жения материи в целом шло по пути н е о г р а н и ч е н н о г о
п р о г р е с с а . Он заключается в развитии живых сущ еств от прос­
тейших форм через ряд усложнений морфофизиологической организа­
ции особей, структуры популяций и сообществ более высокого ранга,
изменений отношений между средой и организмами вплоть до
возникновения новой, высшей формы движения материи — соци­
альной.
Процессу неограниченного прогресса свойствен многоступенчатый
характер. Сущ ественными его этапами служ ат образование эукариоти­
ческой клетки, многоклеточных организмов, позвоночных животных,
млекопитающих, приматов и, наконец, человека. С достижением
очередного этапа приобретались новые эволюционные и экологические
перспективы, которые обеспечивали выходы жизни в адаптивные ^оны
с более разнообразными, усложненными условиями. Н а любом этапе
эволюция группы организмов— носителей новых свойств шла по
мног". и направлениям. Э то нашло отражение в общей картине совре­
менного органического мира, которая представлена древом жизни.
Выход в социальную форму движения материи произошел только
в одной из ветвей развития жизни, остальные оказались тупиковыми.
Причина этого заклю чается в различии масш таба адаптаций, вы зы ваю ­
щих разные направления развития. Расширение экологических и сохра­
нение эволюционных перспектив наблюдались, если группа приобрета­
ла и накапливала адаптации общего значения.
7.8.2. Биологический прогресс
Современный органический мир представлен многообразием групп.
Несмотря на значительные различия в морфофизиологическом отноше­
нии, организмы из разных групп сосущ ествуют. Следовательно, им
всем сопутствует успех в борьбе за существование. В каждый отдельно
взятый период процесса исторического развития представители одних
групп отличаю тся особенно широким распространением, а представи­
тели других групп относительно малочисленны, но им свойственна
тенденция к экспансии. Третья категория групп находится в стадии
вымирания. Для сравнения состояния отдельных групп организмов
в конкретный исторический период, для оценки степени их
успеха
в
борьбе
за
существование
используется
критерий
б и о л о г и ч е с к о г о п р о г р е с с а , показателями которого слу­
ж ат: 1) увеличение численности представителей соответствующ ей
группы: 2) расширение ареала; 3) увеличение числа дочерних групп в
составе более крупного таксона — видов в роде, родов в семействе.
Среди выделенных выше групп отнесенные к первой категории
282
удовлетворяют перечисленным критериям и находятся в состоянии
биологического прогресса, группы второй категории — на пути к нему,
третьей — миновали период расцвета. Так, среди наземных позво­
ночных амфибии возникли в девонском периоде и господствовали на
суше в течение последующих 75 млн. лет. В каменноугольном периоде
появляю тся рептилии, занимавшие, однако, в то время среди наземных
животных достаточно скромное место. В триасе положение меня­
ется — амфибии уступаю т сушу рептилиям, при этом крупные амфибии
к концу триасового периода вымираю т. Н а протяжении юрского и
отчасти мелового периода доминируют динозавры, деля наземный мир
с примитивными млекопитающими. Волна вымирания динозавров
завершилась примерно 65 млн. лет том у назад, вслед за чем произошла
адаптивная радиация млекопитающих, которые господствуют на суше
последние 50 млн. лет.
Критерии биологического прогресса трудно использовать для
сравнительной оценки состояния представителей далеких ветвей
развития жизни, например простейших и млекопитающих. Поэтому
обычно их применяют при сравнении относительно близкородственных
групп, представители которых сходны в главных чертах морфофизио­
логической организации (классы в пределах типа, отряды в пределах
класса и т. д .). Биологический прогресс эквивалентен состоянию
б и о л о г и ч е с к о г о п р о ц в е т а н и я группы, зависящего от
успеха в борьбе за существование безотносительно к характеристикам
адаптаций, которые этот успех создают. Э то могут бы ть приспособле­
ния как общего (ароморфозы), так и частного (идиоадаптации)
значения. В группе паразитов процветание нередко сопровождается
морфофизиологической дегенерацией. Таким образом, состояние био­
логического прогресса достигается группой на любом из уровней
структурной и физиологической организации особей.
7.8.3. Морфофизиологический прогресс
Наиболее крупные ветви развития жизни, например царства,
представлены в современном мире ограниченным числом типов,
существенно различающихся по морфофизиологической организации
особей (членистоногие и хордовые). В процессе исторического развития
в типах последовательно возникают классы, которые, сохраняя свойс­
твенные типу принципиальные черты строения, отличаю тся прогрес­
сивным усложнением морфологии и функций. В типе Хордовых
подтипе Позвоночных, например, такой ряд образован классами Ры б,
Амфибий, Рептилий, Птиц и Млекопитающих. Возникновение и прог­
рессивное
совершенствование
в
эволюции
крупного
таксона
плана морфофизиологической организации соответствует понятию
м о р ф о ф и з и о л о г и ч е с к о г о п р о г р е с с а . Э то состоя­
ние достигается благодаря изменениям строения и физиологии, которые
ведут « ... к повышению интенсивности, разнообразия и сложности
функций органов, т. е. к подъему общей энергии жизнедеятельности
организма...»'
1 А. Н. Северцов. Главные направления эволюционного процесса (Морфобиоло­
гическая теория эваиоции). Изд. М Г У , 1967, с. 68.
283
Предложено несколько критериев морфофизиологического прогрес­
са. Так, он сопровождается нарастанием дифференциации и функцио­
нальной специализации частей организма на фоне более эффективной их
интеграции в единое целое (К . Бэр). Важным показателем служит
возрастание независимости организмов о т окружающей среды, все
большее избавление их от прямой власти условий обитания. О степени
независимости живых форм, различающихся по уровню морфофункци­
ональной организации, по отношению к среде обитания можно судить
по масш табам неизбирательного истребления гамет, зародышей и мо­
лоди. Так, от максимально возможного количества потомков в каждом
поколении до репродуктивного возраста доживает: у амфибий
менее
1%,
у
рептилий—
2— 3 % ,
у
птиц —
10%,
у
млекопитающих — 30— 50%. У лососевых на каждые 100 000 икринок
развивается 40 взрослых рыб, а у воблы — 15. Уходу от непосредствен­
ной власти условий обитания способствуют изменения, стабилизирую­
щие внутреннюю среду организма, совершенствование механизмов
гомеостаза. Достижение высших уровней независимости от внешней
среды сопряжено с приобретением живыми формами более сложного
строения. Такое усложнение требует увеличенных энергетических затра!т
на поддержание в организмах с прогрессивными чертами строения
более низких значений энтропии. Источником свободной энергии для
живых сущ еств служит среда обитания, поээтому в процессе
морфофизиологического прогресса возрастает сила действия (в изве­
стном смысле разрушающего) организмов на окружаюший мир.
Состояние морфофизиологического прогресса возникает всегда на
основе комплекса ароморфозов, который складывается на протяжении
длительных отрезков процесса исторического развития. Отдельные его
компоненты возникают в недрах предковой группы и объединяются у
переходной формы. Так, признаки, характеризующие в комплексе
млекопитающих — живорождение, теплокровность, развитие головно­
го мозга, совершенная терморегуляция, определенное строение зубной
системы, черепа, лицевых мышц, наличие диафрагмы — имеются
порознь у представителей разных групп их эволюционных предше­
ственников — рептилий. Эти признаки возникали в разное время и в
различных ветвях развития рептилий, но объединились хотя бы
в некоторой своей части у терапсид.
Приобретение черт прогрессивной структурно-функциональной
организации способствует биологическому прогрессу. Одновременно
группа сохраняет возможность дальнейшей эволюции, пределы которой
ограничены. Так, насекомых отличает повсеместное распространение и
чрезвычайное многообразие видов, число которых превышает количе­
ство видов всех остальных животных на Земле. Вместе с тем это тупи­
ковая ветвь развития жизни. Линия неограниченного прогресса прошла
через млекопитающих.
Причины утери группами эволюционных перспектив во многом не
ясны. Э то могут бы ть ограничения, накладываемые типом структурно­
функциональной организации в целом или выраженная специализация,
что препятствует освоению более сложной адаптивной зоны. Так,
некоторое приближение к социальному образу жизни встречается,
284
например, у насекомых и дельфинов. Предполагают, что общественные
насекомые не перешли рубеж между биологической и социальной
формой движения материи из-за выраженного преобладания у них
инстинктов над индивидуальным научением. М озг дельфинов по ряду
показателей превосходит мозг человека. Однако дельфины — это
млекопитающие, глубоко специализированные по многим признакам к
жизни в относительно однородной, стабильной водной среде.
Основу морфофизиологического прогресса составляю т адаптации
общего значения — ароморфозы. Так как масш таб последних варьиру­
ет, понятие морфофизиологического прогресса используется также при
сравнении особенностей строения и функций представителей разных
отрядов в пределах класса, семейств в пределах отряда.
Выдающийся вклад в развитие представлений о биологическом и
морфофизиологическом прогрессе внес А . Н . Северцов.
7.9. Э В О Л Ю Ц И Я И О Н Т О Г Е Н Е З
Особенности морфологии и физиологии современных организмов
представляют собой результат исторического развития (филогенеза)
соответствующ его таксона. Они отражаю т те преобразования, которые
происходили с предковыми формами на протяжении многих поколений.
Между тем у особей каждого поколения черты структурно-функцио­
нальной организации воспроизводятся в процессе индивидуального
развития (онтогенеза). Процесс исторического развития вида пред­
ставлен чредой сменяющих друг друга поколений особей. В биологиче­
ском смысле особь и ее онтогенез понятия тождественные (см. 2.2),
поэтому можно сказать, что историческое развитие является рядом
последовательно сменяющихся онтогенезов.
В связи с этим между историческим и индивидуальным развитием
сущ ествует тесная связь, познание сущности которой имеет большое
значение для понимания механизмов эволюционного процесса.
7.9.1. Закон зародышевого сходства
В начале прошлого века биологи обратили внимание на то, что
в индивидуальном развитии некоторые органы проходят стадии, на
которых они сходны с органами взрослых низших представителей того
же крупного таксона. Так, в эмбриогенезе человека последовательно
закладывается предпочка, первичная и, наконец, вторичная почка,
которая функционирует во взрослом состоянии.
Н а основании сравнительно-эмбриологических исследований К . Бэр
сформулировал з а к о н з а р о д ы ш е в о г о с х о д с т в а , соглас­
но которому черты, общие для большой группы родственных
животных, появляю тся у зародышей раньше частных признаков.
По К . Бэру, в зародышевом развитии животных наиболее рано форми­
руются признаки, отражающие принадлежность' к определенному
классу, затем — отряду, семейству, роду и, наконец, виду. Он обратил
внимание на то, что на ранних стадиях индивидуального развития
зародыши разных животных проявляют большое сходство между
собой.
285
Суждения о том, что организм в эмбриогенезе повторяет в той или
иной форме историю развития вида, высказывались до появления
дарвиновской теории эволюции. Однако лишь на основе положений
этой теории связь между фило- и онтогенезом получила правильное
естественнонаучное объяснение. Оно состоит в том, что зародышевое
сходство, наблюдаемое в большой группе родственных организмов,
например позвоночных животных, отражает факт их генетического
родства. На это обратил внимание сам Ч . Дарвин, который считал, что
зародыша можно рассматривать в какой-то мере как «свидетеля» тех
состояний, через которые в процессе исторического развития прошел
вид. В трудах Ч . Дарвина отсутствует указание на природу механизма,
при помоши которого историческое развитие находит свое отражение в
развитии индивидуальном.
Проблема связи между филогенезом и онтогенезом получила
дальнейшее развитие в трудах Ф . Мюллера, Э . Геккеля, А . Н . Северцова.
7.9.2. Биогенетический закон
Изучая филогенез ракообразных, Ф . Мюллер обратил внимание уа
сходство некоторых современных личиночных форм с формами их
вымерших предков. На основании этих наблюдений ой сделал заключе­
ние о том, что ныне живущие ракообразные в эмбриогенезе как бы
повторяют путь, пройденный в историческом развитии их предками.
Преобразования индивидуального развития в эволюции, по мнению
Ф . Мюллера, происходят путем добавления новых стадий к онтогене­
зу родителей. Повторение в онтогенезе потомков признаков нескольких
предков объясняется накоплением таких надставок.
Э.
Г еккель сформулировал
основной
биогенети­
ч е с к и й з а к о н , в соответствии с которым онтогенез представляет
собой краткое и быстрое повторение филогенеза.
В качестве доказательств справедливости биогенетического закона
используют примеры р е к а п и т у л я ц и й . Они заклю чаются в пов­
торении структуры органов взрослых предков на определенных стадиях
индивидуального развития потомков. Так, в эмбриогенезе птиц и
млекопитающих закладываются жаберные щели и соответствующие
им скелетные образования и кровеносные сосуды. Многие признаки
личинок бесхвостых амфибий соответствую т признакам взрослых
хвостатых амфибий. В эмбриогенезе человека эпидермис кожи сначала
представлен однослойным цилиндрическим, затем многослойным
неороговевающим, многослойным слабо ороговеваюшим и, наконец,
типичным ороговеваюшим эпителием. Соответствующ ие типы эпите­
лия встречаются у взрослых хордовых — ланцетника, костистых рыб,
хвостатых амфибий.
Согласно Э . Геккелю, новые признаки, имеющие эволюционное
значение, возникают во взрослом состоянии. По мере усложнения
организации взрослых форм зародышевое развитие удлиняется за счет
включения дополнительных стадий.
Признаки предковых форм, повторяющиеся в онтогенезе потомков,
Э . Геккелем названы п а л и н г е н е з а м и . Нарушение биогенетиче­
2X6
ского закона зависит от тех измене­
ний, не имеющих эволюционного
значения, которые возникают в ходе
индивидуального развития под дей­
ствием внешних условий. Они могут
заклю чаться в сдвигании процессов
зародышевого развития во времени
( г е т е р о х р о н и и ) и в простран­
стве (г е т е р о т о п и и). Наруше­
ния, обусловленные приспособления­
ми зародышей к условиям развития,
Э . Геккель назвал ц е н о г е н е з а м и. Примером гетерохроний служит
более ранняя закладка нервной си­
стемы и запаздывание в формирова­
нии половой системы у высших
позвоночных и человека по сравнению
с низшими, гетеротопий — закладка
легких, представляющих собой видо­
А. Н. Северцов (1866 1936)
изменение задней пары жаберных
мешков, расположенных по бокам
кишечника, на его брюшной стороне, ценогенезов — амнион, хорион,
аллантоис зародышей наземных позвоночных.
Основываясь на биогенетическом законе, Э . Геккель предложил
гипотезу филогенеза многоклеточных организмов (см. 7. 11.2). Стадии
морей, бластеи, гастреи исторического развития рекапитулируют, по
его мнению, в онтогенезе многоклеточных животных как стадии
морулы, бластулы, гаструлы.
7.9.3. Теория филэмбриогенезов
Решающее значение для раскрытия связи между онтогенезом и
филогенезом имеют труды А . Н . Северцова. Согласно А. Н . Северцову,
источником филогенетических преобразований служ ат изменения,
возникающие на ранних этапах онтогенеза, а не у взрослых форм. Если
они приводят к развитию признаков, имеющих полезное значение во
взрослом состоянии и наследуются, они передаются из поколения в
поколение и закрепляются. Такие признаки вклю чаю тся в филогенез
соответствующ ей группы организмов. Эмбриональные изменения,
отражающиеся в дальнейшем на строении взрослых форм и имеющие
эволюционное значение, называю тся ф и л э м б р и о г е н е з а м и ,
которые бы ваю т трех типов.
Эмбриогенез может изменяться путем включения дополнительной
стадии к уже имевшимся стадиям без искажения последних
( а н а б о л и я ) , или же ход эмбриогенеза нарушается в средней его
части ( д е в и а ц и я ) . Отклонение от обычного хода развития в начале
эмбриогенеза называется а р х а л л а к с и с о м .
Как видно, биогенетическому закону удовлетворяют изменения
онтогенеза по типу анаболий. В этом случае зародышевое развитие
2*7
представляет, по-существу, ряд последовательных рекапитуляций.
В случае девиации рекапитуляции наблюдаются, но в ограниченном
объеме, а при архаллаксисе они отсутствую т.
Согласно теории филэмбриогенезов изменения на ранних стадиях
индивидуального развития составляю т основу филогенетических пре­
образований органов. Таким образом, онтогенез не только отражает
ход эволюции организмов определенного вида, но, претерпевая измене­
ния, оказывает влияние на процесс исторического развития той или
иной группы животных. И з сказанного следует, что в известном смысле
филогенез можно рассматривать как причину онтогенеза (Э . Геккель).
Вместе с тем коль скоро эволюционно значимые изменения строения
органов во взрослом состоянии происходят путем изменения эмбриоге­
неза этих органов, филогенез представляет собой функцию онтогенеза
(А . Н . Северцов).
7.10. ЭВОЛЮЦИЯ и ПОВЕДЕНИЕ
7.10.1. Эволю ция поведения
Классическая эволюционная теория использовала в качестве
критериев исторического родства главным образом морфологические
признаки. Современная этология доказала, что не менее надежными
в установлении филогенетического родства являю тся о с о б е н ­
ности поведения.
Поведение складывается из более или менее сложных реакций
организмов на стим улы, поступающие из внешней среды. Поведенче­
ские реакции бы ваю т врожденными или приобретаются
путем
научения.
Способность к научению варьирует у представителей разных групп
организмов и внутри вида. Х о тя наследственные основы указанного
признака далеко не ясны, сам факт его генетической детерминации
сомнений не вы зывает. Э то означает, что способность к научению
подвержена действию естественного отбора, а эволюция такой
формы поведения не могла происходить согласно обычным меха:
низмам.
Показана наследуемость и некоторых врожденных форм поведения,
включая альтруизм и ритуальное поведение Животных, оспаривающих
какой-либо важный жизненный фактор — местообитание, партнер для
спаривания. В последнем случае нередко имеющееся в распоряжении
соперников оружие не используется максимально эффективно. Спор
ведется «по правилам», которые направлены на сокращение возможно­
сти серьезных повреждений. Соверш ая альтруистический акт, особь как
бы отказывается от собственных интересов. В связи с этим можно
ожидать, что эволюция таких поведенческих реакций должна идти
необычными путями. Действительно, теория утверждает, что в основе
эволюции лежит первичное изменение частот аллелей. При этом
частота определенного аллеля растет только в том случае, если он
повыш ает общую приспособленность его носителей. Напомним, что
общая или дарвиновская приспособленность пропорциональна произве­
288
дению жизнеспособности на репродуктивный потенциал особи и оцени­
вается числом оставляемых потомков.
Рассмотрим пример с родительским альтруизмом. Известно, что
родители нередко рискуют жизнью , защищая потомство. Закономерен
вопрос, мог ли естественный отбор в процессе эволюции видов с
альтруистическим поведением сохранить и размножить аллели,
повышающие вероятность смерти и, таким образом, объективно
снижающие вероятность их передачи дочернему поколению. Предста­
вим систему из двух особей, одна из которых (донор) благодаря нали­
чию в генотипе аллеля А способна к альтруистическому действию в
отношении второй особи (реципиента). У организмов, имеющих аллель
а, альтруистические наклонности снижены. Осуществление донором
акта альтруизма снижает его приспособленность на величину С, но
повыш ает приспособленность реципиента на величину В. Возможна ли
ситуация, когда альтруизм повышал бы суммарную приспособленность
популяции? Модельные расчеты показали, что направление изменений
частоты аллелей альтруизма (в данном случае А ) в ряду поколений
зависит о т коэффициента родства между донором и реципиентом. Если
его значение превышает ^еЯичину С / В , то альтруистические действия
приводят к подъему в генофонде популяции аллеля А. При таких усло­
виях этологический признак, вредный для отдельной особи, подверга­
ется положительному отбору и закрепляется в генофонде. Э ти расчеты
указы ваю т, что эволюция столь важных форм поведения как
сотрудничество и индивидуальный альтруизм могла происходить на
основе внутрипопуляционных разграничений на группы, закономерно
отличающ иеся степенью генетического родства.
Сделанное заключение подтверждается наблюдениями за пчелами,
для общественной организации которых характерны сотрудничество,
разделение на касты , индивидуальный, (односторонний) альтруизм
одной касты в отношении другой. Пчел, как и других перепончатокры­
лых, отличает первичная диплоидность самок и гаплоидность самцов,
поэтому самка имеет больше общих генов с сестрами, чем с собствен­
ными дочерьми (рис. 125). В самом деле самка (1) получает два набора
генов, один из которых — отцовский — у самок (1 и 2) одного
поколения идентичен. О тсю да коэффициент родства между сестрами
составляет 3/4. Отцовские наборы генов самки (1) и ее дочери (4) раз­
личны, что снижает коэффициент родства между ними до 1/2. У всех
полностью общественных насекомых, к которым принадлежит и пчела,
наблюдается период, когда м ать продолжает откладывать оплодотво­
ренные яйца, тогда как ее первая дочь (2) уже достигла половой зрело­
сти. Последняя, как правило, не осущ ествляет размножение, а остается
в семье и выполняет альтруистические действия в отношении сестер
(уход, кормление, защ ита), т. е. становится рабочей пчелой.
Казалось бы рассматриваемой гипотезе противоречит то т факт, что
для самки перепончатокрылых коэффициент родства с сыном (4) равен
1/2, а с братом (3) — 1/4. Анализ ситуации в семьях пчел показал, что у
них неоплодотворенные самки (2), осуществляющие уход за сестрами, в
то же время откладываю т яйца, развивающиеся партеногенетически в
самцов (5). Э то объясняет то т факт, что наряду с сестрами (1) они
10— 56
289
я о ш
а
р а з н ы е гены
Рис. 125. Степень генетического родства самки-матери с сестрами, дочерьми, сы­
новьими.
Сплош ные
в те ксте .
стр ел ки
—
поток
генов,
пун кти р ны е
н аправлени и
пр оявлен ии
ал ь тр у и зм а .
О б ъяс н е н и е
заботятся о сыновьях (5) больше, чем о братьях (3). Именно такая
стратегия, а не участие в репродукции гарантирует дочери макси­
мальные возможности сохранить в ряду поколений генотипы, близкие
по составу аллелей ее собственному. Такое поведение способствует
закреплению и аллелей альтруизма.
Известны проявления альтруизма между особями, не состоящ ими в
близком генетическом родстве, которые объясняю тся включением
поведенческих реакций, формирующихся на основе личного опыта. Так,
зафиксированы обращения самца павиана, пытающегося отбить самку
в период течки у другого самца, к третьему самцу. Нередко требуемая
помощь предоставляется, но в случае успеха самка неизменно достается
первому, а не третьему самцу. Наблюдения показали, что самцы,
которые часто отвечаю т на призыв помочь, сами получаю т помощь
особенно часто. Правилом служит также обращение к тем особям,
которые сами ранее получали поддержку. Таким образом, в отличие от
индивидуального альтруизма пчел здесь имеет место взаимный
альтруизм. Рассмотренные примеры показываю т, что эволюция
различных форм поведения происходит, по-видимому, согласно уни­
версальным эволюционным механизмам путем естественного отбора и
закрепления в генофондах видов поведенческих реакций, повышающих
приспособленность популяций.
290
7.10.2. Значение этологических признаков
в эволюции групп животных
Среди эволюционистов растет убеждение, что нередко именно
отклонения в поведении, приводящие к смене местообитания, источни­
ка пищи или образа жизни, вклю чаю т новые направления отбора и
стимулирую т формирование необычных адаптаций. Пример с пчелами
подсказывает, что положительный отбор по альтруизму мог бы ть
одним из факторов перехода к общественному укладу жизни. В пчели­
ном улье неоплодотворенные самки в силу особенностей генофонда
популяции взяли на себя уход, кормление развивающегося потомства,
защ иту гнезда. Э то привело к выделению касты рабочих пчел с четко
очерченными и генетически закрепленными функциями. В результате
оказалось возможным глубокое разделение труда, связанное с размно­
жением, выделение каст маток и трутней.
Сущ ествует гипотеза, согласно которой развитию орудийной
деятельности древнейших гоминид способствовало освоению такой
пищи, как костный мозг, содержащийся в трубчатых костях животных,
убитых хищниками. Так, большая часть костей, найденных в пещере
М акапансгатс (Африка), которая служила местом обитания австрало­
питеков, представлена фрагментами трубчатых костей крупных
млекопитающих со следами намеренных расколов.
7.11. С О В Р Е М Е Н Н А Я С И С Т Е М А
О РГА Н И ЧЕС КО ГО М ИРА
П о признаку клеточного строения мир живых существ представлен
и м п е р и я м и неклеточных (вирусы, фаги) и клеточных организмов.
Характер филогенетических отношений между ними не ясен. В соответ­
ствии
с
двумя
типами
клеточной
организации
выделяю т
п о д и м п е р и и прокариотических и эукариотических организмов,
имея в виду также то, что эукариоты произошли от прокариот. Эукари­
оты подразделяются на группы одноклеточных (простейших) и мно­
гоклеточных организмов. В процессе эволюции вторые произошли от
первых. В зависимости от общих особенностей образа жизни и, в
частности, способа добывания энергии многоклеточные объединены в
три ц а р с т в а : растений — Ме1арЫ1а, грибов — Рип§ 1 и живот­
ных — Мегагоа (рис. 126). Растения являю тся автотрофами и использу­
ю т энергию Солнца, грибы питаю тся органическими веществами, но по
многим другим признакам напоминают растения. Ж ивотны е пред­
ставляю т собой гетеротрофы, и их образ жизни существенно отлича­
ется от такового растений и грибов. Вопросы эволюционных предше­
ственников и путей происхождения названных царств живой природы
остаю тся предметом дискуссий.
Наиболее полно изучены история и филогенетические . связи
в царстве животных. Знакомство с этой ветвью развития жизни
помогает увидеть некоторые общие закономерности филогенеза — на­
личие ряда форм и типов эволюции групп, необратимость и прогрессив­
ный характер процесса исторического развития в целом. Предполагают,
Ю*
291
Рис. 126. Филогенетические отношении основных групп растений, грибов, животных
и прокариот (пунктиром обозначены предполагаемые положении групп).
что среди простейших имелось по крайней мере две разные предковые
формы многоклеточных животных. Потомки одной из них представле­
ны в современной фауне губками, а все другие животные произошли,
по-видимому, от иного предка. Наиболее крупные систематические
группы, на которые подразделяется царство животных, называю тся
типами. За период существования рассматриваемого царства их было
не менее 35. Имеется несколько классификаций типов современных
животных. Согласно одной из них в настоящее время на Земле обитает
26 типов. В зависимости от существенных особенностей строения эти
типы можно объединить в группы (рис. 127). Сравнительный анализ
292
П р о м е ж у тонн ая
1В т о р и ч н о группа
1 р оты е
П ервично\
ротие
\
'"Л /
Ж ивотны е без
втор и ’ш ой полости
тела
Ц В то р и ч н о
Ц полостные
Ц {л ело м и чесн и е)
П о явл е н и е вторичной полости те л а наи органа о б ш е го
зн аче н и я
ж и во тн ы е
Д в у с л о й н ы е или
осеволучевы е
животны е
II Iг-ар азо а
Тр е х сл о й н ы е или
двусторон
несим м етр ичны е
ж и во тн ы е
П р о сте й ш и е
м ногоклеточные
Н а ли чи е зар о д ы ш евы х л и с тк о в и стойких клеточных
специализаций. О б р а з у ю т с я ткани. П о я в л я е тс я нервная
си стем а. Д о с т и г а в ш и вы соки й у р овен ь интеграции
организма
| Эум етазоа
1
...................
Н а ли чи е трех зар о д ы ш е вы х л и стков. И м е ю тс я органы
и си ст ем ы органов. Д в у с г о р о н я я си м м етр и я тел а как
о с н о ва активизации д в и ж е н и я
■'
К л е то чн ы е механизмы повторены много раз.
В о з м о ж н о с т ь увеличить р азм е р ы и повысить
стаб и л ьн о сть организма. Фун кц и он альн ая
специализация к ле ток. Увел и че н и е чи сл а потомков
при р азм н о ж ен и и гаметами. У вел и че н и е
пр од о лж и тел ьн ости ж изн и
О дн онл ето ч н ы е
п редновые
формы
Рис. 127. Схема главных тупиковых и прогрессивных направлений эволюции групп
в царстве животных.
1 - ранг группы многоклеточных животных; 2 — название группы; 3 —
соотгететкующие прогрессивному направлению эволюции.
особенности организации,
последних дает представление об узловых пунктах процесса историче­
ского развития, в которых происходило разделение тупиковых и прог­
рессивных направлений филогенеза. В рисунке приведены ароморфозы
общего значения, способствовавшие неограниченному прогрессу жизни.
Представители отдельных типов современных животных изображены
на рис. 128 (см. на цвет. вкл.).
Клю чевыми событиями в истории развития жизни, которые
определили картину современного органического мира, считаю т
возникновение ее самой на планете, переход к эукариотическому типу
клеточной организации, появление многоклеточности, возникновение
человека.
7.11.1. Происхождение жизни
Есть две основные гипотезы, по разному объясняющие появление
жизни
на
Земле.
Согласно
разным
вариантам
гипотезы
п а н с п е р м и и жизнь была занесена из Космоса в виде спор
микроорганизмов или же разумные пришельцы из других миров
«засеяли» планету живыми существами намеренно. Вторая гипотеза
утверждает, что ж и з н ь в о з н и к л а н а З е м л е . В середине
прошлого столетия Л . Пастер окончательно доказал невозможность
29.'
самозарождения жизни в настоящих условиях. В 20-х годах текущего
столетия советский биохимик А . И . Опарин и английский биохимик
Д ж. Холдейн предположили, что в условиях, имевших место на планете
миллиарды лет назад, образование живого вещества было возможно.
Э ти условия включали отсутствие других живых существ и наличие
источников энергии, атмосферы восстановительного типа, воды и
приемлемого температурного режима. Основными этапами на пути
к возникновению жизни, по-видимому, были: 1) образование атмосфе­
ры из газов, которые могут служить «сырьем» для синтеза
органических веществ — метана, окиси и двуокиси углерода, аммиака,
сероводорода, паров воды; 2) абиогенный синтез биологических
мономеров — аминокислот, сахаров, азотистых оснований; 3) поли­
меризация мономеров в биологические полимеры — полипептиды и
полинуклеотиды; 4) образование предбиологических сложных сис­
тем — протобионтов, наделенных некоторыми свойствами живых
форм; 5) возникновение элементарной структурной, функциональной и
генетической единицы жизни — клетки.
Возможность а б и о г е н н о г о с и н т е з а б и о л о г и ч е ­
с к и х м о н о м е р о в при отсутствии живых организмов в условиях,
напоминающих существовавшие на Земле более 3,5 млрд. лет назад, до­
казана экспериментально. Если электрические разряды пропускать через
смеси газов, имитирующих примитивную атмосферу планеты, то в ка­
честве продуктов реакций получаю тся аминокислоты аланин, глицин,
глутаминовая и аспарагиновая, а также мочевина, молочная, янтарная,
уксусная кислоты. В абиогенных синтезах на заре развития планеты
использовалась энергия УФ-излучения, довольно значительная в то
время из-за отсутствия озонового экрана, а также энергия электриче­
ских разрядов, радиоактивного распада химических элементов, ударных
волн, порождаемых материалами и молниями, космических лучей,
вулканов. Биологические мономеры — предшественники биополиме­
ров — накапливались
в
водах
первичного
океана,
образуя
первичный бульон.
В водной среде термодинамические условия благоприятствую т
реакции диполимеризации биополимеров, что делает маловероятным
образование их непосредственно в первичном бульоне. Предполагают,
что р е а к ц и и п о л и м е р и з а ц и и м о н о м е р о в в поли­
пептиды и полинуклеотиды происходили в атмосфере или на склонах
вулканических конусов, покрытых остывающ ей лавой при отсутствии
воды. Возможность этого доказана в опытах С. Фокса, который
выдерживал сухую смесь аминокислот при 130°С в сосудах, приго­
товленных из кусков лавы, и получил в результате полипептиды.
Б и о п о л и м е р ы , образующиеся абиогенно в безводной среде,
возможно смывались ливнями в первичный бульон, что предотвращало
их распад под действием жесткого УФ-излучения.
П о достижении определенной концентрации органических веществ в
первичном бульоне спонтанно могли возникать к о а ц е р в а т ы или
м и к р о с ф е р ы — коллоидные капли, обогащенные нескольки­
ми или одним полимером, взвешенные в водной среде, имеющие
уплотненный наружный слой, имитирующий мембрану. При опреде­
294
ленных условиях коацерваты проявляю т некоторые свойства живых
систем — они способны до известной степени избирательно поглощать
химические соединения из окружающей среды и использовать их в
различных химических реакциях. Часть продуктов этих реакций
попадает обратно в среду. Накапливая вещества, коацерваты растут и
по достижении определенных размеров распадаются с сохранением
некоторых черт исходной химической организации. Стабильность
коацерватов, различающихся по химическому составу и ассортименту
каталитических реакций, различна. Э то создает условия для химическо­
го отбора систем, отличающ ихся благодаря особенностям метаболизма
и «обмена» со средой большей стабильностью . Дискретные, откры тые
фазовообособленные системы типа коаиерватных капель, возникавшие,
по-видимому, в первичном бульоне, которые были наделены некоторы­
ми чертами живого и способны к предбиологическому химическому
отбору,
получили
название
протобионтов
или
протоклеток.
Протобионты — это еще не жизнь в полном смысле слова. В них
отсутствует механизм, который обеспечивал бы в дочерних каплях
точное воспроизведение набора катализаторов и, следовательно,
метаболизма, гарантирующего выживание. Переход от протобионтов к
клеткам состоял в с о п р я ж е н и и ф у н к ц и й
биополи­
меров,
являющ ихся
высокоспецифичными
катализаторами
химических реакций (белки) и способных хранить и передавать в
ряду поколений полезную для выживания информацию (нуклеиновые
кислоты). В процессе становления клеточной организации возникли
также механизм м а т р и ч н о г о
синтеза,
обеспечивающий
точное копирование биологической информации, и
высоко­
эффективные
системы
энергообеспечения
жизнедеятельности, основанные на использовании А Т Ф . С этого момен­
та химический отбор уступил место биологическому и дальнейшее
развитие жизни пошло согласно законам биологической эволюции.
7.11.2. Происхождение многоклеточных животных
Сущ ествую т различные гипотезы происхождения многоклеточных
организмов, однако общепринятым является мнение о том , что предка­
ми современных многоклеточных животных были колониальные формы
гетеротрофных простейших. В обобщенном виде переход к многоклеточности мог иметь следующие этапы (рис. 129). Р о д о ­
н а ч а л ь н и к о м , по-видимому, служила ведущая свободный образ
жизни, шаровидная колония воротничковых жгутиконосцев типа
5рЪаегосеа с периферическим расположением особей (рис. 129, а).
Каждая из них имела ж гутак, окруженный у основания «во­
ротничком» — выростом цитоплазмы, образующим карман, в который
загонялись пищевые частицы, т. е. первичным способом питания был
фагоцитоз. Функцию движения обеспечивал жгутиковый аппарат.
Эволю ция описанной формы шла в направлении специализации клеток
на п о л о в ы е
исоматические
с выделением среди по­
следних ф а г о ц и т о в и к и н о ц и т о в . Первые обеспечивали
295
Рис. 129. Пути происхождения много­
клеточных животных.
а — родоначальник; б, в ~ две стадии эволюции
фагоцителлы; г — кишечнополостные; д
бес-'
кишечные турбеллярии; е — губкн; 1 — киноцит;
2 — фагоцит; 3 — яйцеклетки; 4 — спермато­
зоиды; 5 — другие клеточные типы; 6
рото­
вое отверстие.
пищеварительную, вторые — двигательную функцию. Н а ранних ста­
диях функции питания и движения, видимо, совмещались в одной
клетке, выступавш ей попеременно то в одной, то в другой роли.
Стойкому разделению функций соматических клеток способствовало
смещение фагоцитов внутрь колонии, тогда как киноциты сохранили
периферическое положение.
Образование вследствие этого ф а г о ц и т о б л а с т а и к ин о б л а с т а соответствует важному этапу перехода к многокле­
точное™ — возникновению двухслойной предковой формы, которую
И . И . Мечников назвал ф а г о ц и т е л л о й
или
паренхим е л л о й (рис. 129, б). Согласно рассматриваемой гипотезе, первич­
296
ным способом образования двухслойного организма была иммиграция.
Вначале выселение клеток из наружного слоя внутрь происходило, повидимому, равномерно, но затем локализовалось на одном из полюсов.
Поляризация организма способствовала образованию на заднем полюсе
первичного ротового отверстия. Благодаря упорядоченному биению
ресничек здесь создавалось «мертвое пространство», что обеспечивало
скопление пищевых частиц и лучшие условия для их поглощения
(рис. 129, в).
Н а следующем этапе дивергентная эволюция предка типа
фагоцителлы дала несколько вариантов многоклеточных, которые
напоминали по типу строения современных низших животных —
кишечнополостных, некоторых плоских червей и, возможно, губок.
Развитие в направлении к и ш е ч н о п о л о с т н ы х включало
образование стойкого ротового отверстия, полную эпителизацию
(превращение в пласт) фагоцитобласта и возникновение первичного
пищеварительного органа в виде энтодермы (рис. 129, г). Клетки
эктодермы переднего полюса такого организма приобрели повы­
шенную чувствительность, и здесь могли образоваться ловчие щупальца.
Организм в целом характеризовался осеволучевым типом симметрии
тела.
Развитие в направлении современных
бескишечных
т у р б е л л я р и_р (класс Ресничные черви, тип Плоские черви)
происходило без эпителизации фагоцитобласта в энтодерму, хотя
ротовое отверстие и возникло (рис. 129, д). Смещение у таких
животных ротового отверстия на брюшную сторону, будучи связано,
по-видимому, с освоением ползающего образа жизни, обусловило
переход к двусторонней симметрии тела. Э та ветвь развития жизни на
рассматриваемом этдпе филогенеза соответствовала направлению
неограниченного прогресса.
Ученые, которые допускают общую предковую форму для
кишечнополостных, червей и г у б о к , указы ваю т, что линия развития
последних должна была выделиться на^ стадии паренхимеллы до
образования ротового отверстия (рис. 129, е). Особенность эволюции в
этом направлении заклю чается в изменении локомоторной функции
кинобласта на гидродинамическую, что сопровождалось погружением
киноцитов внутрь тела.
Представления, изложенные выше, отражаю т современное состоя­
ние гипотезы происхождения многоклеточных через стадию фагоци­
теллы. Впервые эта гипотеза была сформулирована И . И . Мечнико­
вы м (1877— 1880). Он исходил из того, что: 1) в эволюции жизни
внутриклеточное пищеварение предшествует полостному; 2) эволюционно более ранним способом образования энтодермы служила не
инвагинация наружного слоя, а иммиграция клеток; 3) древнейшее
многоклеточное существо имело шаровидную форму и было одно­
слойным.
Альтернативная гипотеза была сформулирована Э . Геккелем в 70-е
годы прошлого века. Он сделал следующие предположения:
1) древнейшее многоклеточное (м о р е я) представляло собой шаро­
видную колонию плотно упакованных клеток с амебоидной активно­
297
V
стью ; 2) на следующем этапе оно путем накопления жидкости или
студнеобразного вещества эволюционировало в пузырькообразный
организм (б л а с т е ю ); 3) первичным способом образования двух­
слойного сущ ества (г а с т р е й) была инвагинация; 4) первичным в
эволюции многоклеточных животных было полостное пищеварение.
Данные современной сравнительной зоологии, зоологии беспозво­
ночных, сравнительной эмбриологии свидетельствую т в пользу гипоте­
зы И . И . Мечникова.
7.12. О Б Щ И Е З А К О Н О М Е Р Н О С Т И Ф И Л О Г Е Н Е З А
СИ СТЕМ О РГА Н О В М Н О ГО КЛ ЕТ О ЧН Ы Х
Ж И ВО ТН Ы Х И ЧЕЛ О ВЕКА
Строение и функции органов человека представляют результат
длительных эволюционных изменений структур предшествующих,
более примитивных форм. Задача данного раздела — проследить
основные направления и способы преобразования систем органов
человека в процессе исторического развития. Для решения этой задачи
используется исторический метод, значение которого в биологии и
медицине исключительно велико. Эволюционно-исторические исследо­
вания раскрываю т последовательные этапы процесса становления
систем органов человека, обусловившие их строение и функции. Знание
филогенеза систем органов позволяет врачу понять причины про­
исхождения аномалий развития, рудиментов и атавизмов, найти
оптимальные пути реконструкции органов, оценить возможности
восстановления исходной функции реконструированного органа, а так­
же дает доказательства животного происхождения человека.
Прежде чем перейти к разбору конкретного материала по
филогенезу систем органов человека, необходимо ознакомиться с общи­
ми закономерностями преобразований формы и функции в процессе
эволюции и методами их изучения.
7.12.1. Эволюционная морфология и ее задачи
Закономерности филогенетических преобразований изучает область
науки, получившая название э в о л ю ц и о н н о й м о р ф о л о г и и .
Она возникла в конце X IX века на базе трех самостоятельных наук:
сравнительной
анатомии,
эмбриологии
и
палеонтологии.
Сравнительная
анатомия
изучает строение сходных
органов взрослых форм животных, относящихся к различным система­
тическим группам. С р а в н и т е л ь н а я э м б р и о л о г и я сопо­
ставляет зародышевое развитие организмов, поскольку сходство
эмбрионов нередко проявляется более отчетливо, чем взрослых форм.
П а л е о н т о л о г и я изучает останки вымерших форм и последова­
тельную смену форм животного мира на Земле. Ее материалы
позволяю т сравнивать не только строение предковых форм, но и в
известной мере условия их существования на основании данных о
сопутствующ их организмах и характере осадочных пород.
Перед эволюционной морфологией сто ят три главные задачи:
установление фактов повторения предковых форм, т. е. преемственно­
298
сти строения или развития, позволяющего вы явить родственные связи
между организмами; установление способов и направлений филогенети­
ческих преобразований в процессе эволюции; выявление связи между
характером изменений и условиями существования. В настоящее время
эти задачи во многом решены. Установлена филогенетическая
преемственность структуры и функций между смежными группами и
родственные связи между основными группами животного мира.
Сформулированы принципы филогенетических преобразований формы
и функции, обусловивших смену форм в процессе исторического
развития и приспособительный характер эволюционных преобра­
зований.
Больш ой вклад в развитие эволюционной морфологии внес
А . Н . Северцов. Работы А . Н . Северцова „ в этой области были
продолжены И . И . Ш мальгаузеном (1884— 1963), В. Н . Беклемишевым
(1890— 1976), В . А . Догелем (1882— 1955).
7.12.2. М етоды эволюционной морфологии
В эволюционной морфологии основным является м е т о д
« т р о й н о г о п а р а л л е л и з м а » , сочетающий методы сравни­
тельной
анатомии,
эмбриологии
и
палеонтологии.
Палеон­
тологические данные даю т возможность представить события,
происходившие реально _в прошлом, и таким образом прове­
рить правильность выводов, вытекающ их из материалов сравнитель­
ного анализа строения современных организмов. Именно палеон­
тология представляет такие веские доказательства эволюции, как
п е р е х о д н ы е ф о р м ы о р г а н и з м о в , сочетающие признаки
более древних и более, молодых групп. Н а основании палеонтологи­
ческих находок удается построить ф и л о г е н е т и ч е с к и е
р я д ы , т. е. последовательности ископаемых форм, связанных друг с
другом
в процессе эволюции.
Сравнительно-эмбриологические
доказательства эволюции базирую тся на явлениях зародышевого
сходства и рекапитуляции (см . 7.9.1 и 7.9.2).
Основной вклад в исследования закономерностей филогенеза внесли
сравнительно-анатомические исследования.
С помощью метода тройного параллелизма эволюционная мор­
фология устанавливает и дифференцирует гомологичное и аналогичное
сходство структур, гомологичные и аналогичные органы, что составля­
ет первоначальную задачу филогенетических исследований.
Гомологичное
сходство
(о т греч. Ьото1о§ох —
одинаковый) проявляется в следующем: 1) единство плана строения
органов, одинаковое соотношение и расположение их частей; 2) одина­
ковое расположение органов в теле; 3) происхождение органов из одних
и тех же эмбриональных зачатков. При этом функция органов может
бы ть как одинаковой, так и различной. Примером гомологичных
органов служ ат передние конечности различных позвоночных, которые
имеют один и то т же план строения — сначала идет плечо, затем
предплечье, запястье, пястье и фаланги пальцев (рис. 130, см. на
цвет. вкл.).
299
Причина гомологичного сходства — историческое родство, т. е.
происхождение органов о т одного и того же органа общего предка.
Благодаря этому сходство организации сохраняется даже в различных
условиях сущ ествования, в то время как функция в новых условиях не­
редко видоизменяется. Так, у кровососущих насекомых слюнные
железы вместо секрета, смачивающего пищу, выделяю т вещество,
препятствующее свертыванию крови. Передняя конечность крота
служит для ры тья почвы, тогда как крыло птицы для полета. Благодаря
различной функции возникаю т некоторые второстепенные отличия и в
структуре, но общий план строения и соотношение частей гомоло­
гичных органов остаю тся неизменными. Изучение отличий в строении
гомологичных органов позволяет проследить способы и направления их
преобразования в процессе исторического развития.
А н а л о г и ч н о е с х о д с т в о (о т греч. апа1о§ох — сходный)
выражается в одинаковой функции. Оно может сопровождаться лишь
внешним морфологическим сходством. Аналогичными органами явля­
ю тся крыло бабочки и птицы, конечности членистоногих и позво­
ночных. Сходство аналогичных органов определяется одинаковыми
условиями существования и, следовательно, экологическими задачами и
не зависит о т филогенетических связей. Организмы, обитающие в
одинаковых условиях, нуждаю тся в сходных по функции приспособле­
ниях. Так, бабочка и птица, обитающие в воздушной среде, имеют
несущие поверхности в виде крыльев, источники развития которых у
названных животных совершенно различны.
Больш ое значение для понимания закономерностей филогенеза
имеет изучение эволюционных изменений как строения, так и функций
органов. С труктура и функции взаимно обусловлены. Нельзя предста­
вить себе изменения в строении органа без соответствующ их изменений
его функций и наоборот — изменение функции без изменения строения
(формы). Не сущ ествует также органов, не выполняющих какой-нибудь
функции, и нет функций, не связанных с каким-либо органом. В процессе
исторического развития с м е н а ф у н к ц и й всегда неразрывно
связана со с м е н о й
ф о р м . С другой стороны, эволюционно
обусловленные изменения морфологии всегда отражаю т преобразова­
ния определенных функций организма.
7.12.3. Главные принципы и предпосылки эволюционных
преобразований органов
В основе эволюционных преобразований структур лежат процессы
дифференциации и интеграции. Сущ ность процесса
диффе­
р е н ц и а ц и и заклю чается в том , что первоначально однород­
ная структура подразделяется на обособленные части, приобре­
тающие различное строение, и, одновременно различные, более
частные, функции по сравнению с исходной структурой. Благодаря
таким ф и л о г е н е т и ч е с к и м д и ф ф е р е н ц и р о в к а м ор­
ган из простого становится сложным, а функции органа более
разнообразными, что расширяет возможности приспособления орга­
низма к среде.
300
Примером дифференциации является изменение пищеварительной
трубки в подтипе позвоночных. В процессе эволюции пищеварительный
тракт подразделяется на ротовую полость, глотку, пишевод, желудок,
двенадцатиперстную кишку с печенью и поджелудочной железой,
тонкий и толсты й кишечник. В различных отделах пищеварительной
трубки пища подвергается определенным видам воздействия (механи­
ческое, химическое), что приводит в общей сложности к более полной
переработке и повышению усвояемости пищи. Более высокая степень
усвояемости пиши, в свою очередь, обеспечивает более высокий уровень
жизненных процессов и, следовательно, жизнеспособности.
Дифференциация неразрывно связана с процессом
инте­
г р а ц и и (усиления взаимозависимости частей организма).
Обособляющиеся части специализируются на выполнении одной,
более узкой функции исходного органа и в связи с этим в своем суще­
ствовании становятся более зависимыми от остальных компонентов
организма. Поэтому параллельно дифференциации происходит интег­
рация, нарастает степень соподчинения частей организма между собой и
подчинение их организму как целостной системе. Принцип интеграции
составляет основу функциональной кооперации отдельных структур и
органов. Важ ны м проявлением этого принципа служ ит объединение
органов и частей, различных по происхождению и положению в
организме, в функциональные системы (П . К . Анохин), которые
обеспечивают ту или иную существенную сторону жизнедеятельности.
Так, органы дыхания и кровообращения составляю т систему,^
поддерживающую в тканях требуемый уровень окислительных про­
цессов. Каждой группе организмов свойствен свой набор органов в той
или иной функциональной системе, что обусловлено филогенезом.
Необходимой предпосылкой филогенетических преобразований
органов служит и х м у л ь т и ф у н к ц и о н а л ь н о с т ь . Больш ин­
ство органов одновременно выполняю т несколько функций, одна из
которых является г л а в н о й ,
в то время как остальные —
в т о р о с т е п е н н ы м и . Так, передняя конечность собак выполняет
функции передвижения (главная), ры тья земли, удержания пищи,
плавания. У некоторых кольчатых червей органы выделения одновре­
менно служ ат и для выведения половых продуктов. Благодаря
мультифункциональности возможно разделение функций и их преобра­
зование.
Ещ е одной важной и обязательной предпосылкой эволюционных
преобразований
органов
является
способность
к
к о л и ч е с т в е н н ы м и з м е н е н и я м ф у н к ц и й (главных и
второстепенных), которые они выполняю т.
7.12.4. Способы морфофункциональных преобразований
органов
Способы морфофункциональных изменений органов в филогенезе
разнообразны. Более того, в процессе эволюции конкретных групп
организмов они сочетаю тся сложным образом, что затрудняет
выделение их «в чистом виде». Вместе с тем типичные способы таких
301
изменений могут б ы ть описаны. Наиболее часто эволюция органов
происходит способом с м е н ы ф у н к ц и й . В этом случае главная
функция может потерять свое значение, а одна из второстепенных стать
главной. Во всех случаях смены функции одновременно происходят и
изменения строения. Так, у предков позвоночных кожные чешуи при
переходе на челюсти приобрели функцию зубов. Плавательный пузырь
кистеперых рыб в процессе эволюции превратился в орган дыхания
атмосферным воздухом.
Среди других способов филогенетических преобразований наиболее
важными являю тся расширение, усиление и активация функций,
а такж е субституция органов и функций. Примером р а с ш и ­
р е н и я ф у н к ц и й служит эволюция грудных плавников рыб.
Первоначально они выполняли функции удержания тела на опреде­
ленном уровне и создания устойчивости, затем стали рулями глубины и
направления движения, а у донных рыб — одновременно органами
передвижения по дну и опоры.
Усиление
или
интенсификация
функций
сопровождается увеличением числа основных функциональных единиц,
из которых состоит орган. Нередко это связано с ростом размеров ор­
гана и перестройкой его гистолической структуры. Примером может
служить развитие переднего мозга позвоночных, эволюция молочных
желез путем увеличения числа долей. А к т и в а ц и ю ф у н к ц и й
или превращение пассивных органов в активные можно показать на
примере развития подвижных плавников рыб из боковых кожных
складок.
Способ
филогенетических
преобразований
известный как
субституция
или
замещение
органов
и
ф у н к ц и й заклю чается в том , что в процессе филогенеза один орган
замещается другим, принимающим на себя функцию первого.
Заменяемый орган или исчезает, или становится рудиментарным.
Например, хорда замещается позвоночником, головная почка позво­
ночных — туловищ ной.
Исследования филогенеза указы ваю т, что в ходе эволюции могут
наблюдаться г е т е р о т о п и и , которые заклю чаю тся в изменении
места закладки органа или смещении его относительно главных осей
тела. Так, сердце птиц и млекопитающих смешается в грудную полость,
передние конечности высших позвоночных смещены более кзади, чем
грудные плавники рыб. Филогенетические преобразования проявляются
также в виде г е т е р о х р о н и й — изменения времени закладки
органа. Так, у высших позвоночных закладка сердца происходит
раньше, чем у низших.
Д ля эволюции характерны явления гетеробатмии и компенсации
органов и функций. Под г е т е р о б а т м и е й понимают неодинако­
вый эволюционный уровень развития частей органа или организма.
Явление к о м п е н с а ц и и о р г а н о в и ф у н к ц и й состоит в
том, что в процессе филогенеза представителей разных групп
организмов сходные экологические задачи решаются путем прогрессив­
ного изменения лиш ь части органа или системы органов. Например, у
некоторых видов млекопитающих специализация к определенной пище
302
затрагивает желудок, а у других — зубную систему. Явления
гетеробатмии и компенсации органов и функций имеют большое
эволюционное значение, так как благодаря способности изменяться
частями орган и организм сохранят «резервные» неспециализированные
структуры , что может бы ть использовано в дальнейшем развитии в
Ш Ш Г ении'
7.12,5. Соотносительные преобразования органов
Онтогенетические корреляции. Организм представляет собой в
структурном и физиологическом отношении гармоничное целое и
соответствует определенному плану строения. Такой результат дости­
гается в эволюции благодаря согласованности филогенетических
преобразований органов и частей организмов. Коррелятивные измене­
ния структур организма в процессе исторического развития находят
отражение в особенностях соотносительного развития органов и частей
в онтогенезе современных животных. Предварительное знакомство с
онтогенетическими корреляциями дает возможность понять значение
филогенетических корреляций, для обозначения которых используется
термин «координации».
Различаю т геномные, морфогенетические и эргонтические онтогене­
тические корреляции.
В случае г е н о м н ы х к о р р е л я ц и й зависимость между
развитием различных признаков организма определяется действием
генов, а основным механизмом является плейотропия (см. 5.2.2).
Примерами геномных корреляций могут служить сочетание безрогости
и короткош ерстное™ у коз, серой окраски кожуры семян и пурпурных
цветов у гороха, мутация плодовой мухи, проявляющ аяся в редукции
крыльев и одновременном укорочении задней пары ног.
Морфогенетические
корреляции
характерны
прежде всего для ранних эмбриональных стадий развития, когда
функциональные связи между частями зародыша отсутствую т. При
этом зачатки, образующиеся раньше, определяют развитие и характер
изменений зачатков, возникающих позже. В основе корреляций такого
рода лежит явление эмбриональной индукции (см . 6.5.1).
Морфогенетические корреляции обусловливают у позвоночных
больш инство соотношений в величине, форме и расположении
отдельных органов и их частей в организме. При этом нарушение поло­
жения одного элемента корреляционной системы меняет положение или
ход развития другого элемента.
Морфогенетаческие корреляции могут встречаться и на более
поздних стадиях онтогенеза в том случае, если морфогенез не завершен.
Примером этому служит связь между развитием оперения ног у
мохноногих кур и укорочением 4-го пальца. Скелетогенная ткань пальца
в этом случае используется на формирование закладок перьев.
Э р г о н т и ч е с к и е к о р р е л я ц и и проявляю тся на более
поздних стадиях онтогенеза в растущем или взрослом организме. Они
обусловлены взаимозависимостью функций различных органов или
частей организма.
303
<
В качестве примера можно привести корреляции между развитием
периферических органов, нервов и нервных центров в Ц Н С . При
удалении органа чувств или конечностей нервы и соответствующ ие
нервные центры недоразвиваются. Такой же тип корреляции определяет
зависимость между степенью развития мыш цы и костного вы ступа, к
которому она прикрепляется, или степенью развития кровеносных
сосудов, ее снабжающих. Так, при экспериментальном параличе
жевательных мышц у молодых горилл наблюдается недоразвитие
скуловых дуг и гребней на черепной крышке. К эргонтическим
относятся корреляции между ходом развития половых желез и вто­
ричных половых признаков или некоторых эндокринных желез и
скелета.
Геномные корреляции составляю т основу для других форм
корреляций и прежде всего для морфогенетических. Морфогенетические
корреляции являю тся важным фактором формообразования в эмбрио­
генезе на нефункциональных стадиях развития органов. Ими в основ­
ном определяется тип организации, характеризующий ту или иную
систематическую группу. Эргонтические корреляции сменяют морфоге­
нетические на тех стадиях развития, когда органы уже осущ ествляю т
свои специфические функции. Благодаря функциональной согласованно­
сти частей дифференцированного организма, последний реагирует на
изменение среды как целое.
Система корреляций представляет собой главный фактор нормаль­
ного онтогенеза, обусловливающий согласованное течение формообра­
зовательных процессов. В ходе эволюции она претерпевает опреде­
ленные изменения. Причиной их служит непрекращающееся преобразо­
вание органов и структур, что вы зывает дифференцировку самой
корреляционной системы, появление в ней более частных, вторичных
звеньев корреляций.
Корреляции могут оказывать на онтогенез более частное
(распределение позвонков по отделам тела в зависимости от длины и
расчлененности его в разных классах позвоночных) или более общее
(зачаток хордомезодермы определяет развитие закладки нервной
трубки у всех позвоночных) влияние. В процессе естественного отбора
корреляции частного характера, имеющие ограниченное значение,
нередко исчезаю т. Корреляции общего характера, сохраняющие свое
полезное значение при широком круге условий, даже при глубоких
преобразованиях организма, сохраняются естественным отбором.
Примером может служить взаимозависимость между степенью разви­
тия мыш цы и нервного центра или степенью развития органа и
кровеносных сосудов его снабжающих. Такая корреляция полезна при
любых перестройках организма, поэтому она в процессе эволюции
сохраняется.
Подобные корреляции сущ ествую т длительное время и слу|
ж ат основой для дальнейшей прогрессивной эволюции. В результат^
в ходе исторического развития происходит медленное, но неуклонной
накопление корреляций общего значения, что обусловливает преем­
ственность форм, выражающ ую ся в повторении предковых черт\
организации (см . 7.9.2.).
304
Филогенетические корреляции (координации).
Координа­
ц и я м и называю т устойчивые взаимозависимости между органами
или частями организма, обусловленные филогенетически. В структур­
ной организации потомков координации проявляю тся в сосуществова­
нии и согласованном изменении определенных морфофизиологических
признаков, которые образуют постоянный комплекс у той или иной
группы организмов. Если два или более органа изменяются взаимозави­
симо, то можно говорить о координированной эволюции этих структур.
Указанная зависимость бывает прямой или косвенной, иногда обе части
взаимосвязаны определенными условиями среды, но важно, что даже в
ходе дальнейших эволюционных преобразований соотношение между
ними сохраняет постоянный характер.
Различаю т топографические, динамические и биологические
координации.
Топографические
координации
проявляю тся в
сопряженном изменении органов, связанных между собой простран­
ственно, но не связанных функционально. Примером может служить
общий план строения различных типов или более мелких систематиче­
ских групп животных. Так, у хордовых координированная система
признаков вклю чает хорду, нервную трубку, жаберные щели в глотке, у
млекопитающих — млечные железы, волосяной покров, диафрагму,
левую дугу аорты. Топографическими координациями определяется
также соотносительное расположение органов. У всех хордовых, как
известно, на дорсальной стороне тела расположена нервная трубка,
ниже идет хорда, под ней располагается кишечник, под которым
находится сердце или орган, его заменяющий. Основой топографиче­
ских координаций служат морфогенетические онтогенетические корре­
ляции.
Д и н а м и ч е с к и е к о о р д и н а ц и и выражаю тся в согласо­
ванном изменении частей, связанных функционально. Так, сущ ествует
зависимость между строением органа обоняния и обонятельными
долями переднего мозга. У млекопитающих органам обоняния при­
надлежит важная роль в ориентации во внешней среде. Высокоразвито­
му органу обоняния в этой группе животных соответствую т обоня­
тельные доли большого объема. У птиц орган обоняния играет
второстепенную роль в уравновешивании с внешней средой. В связи с
этим строение его достаточно примитивно, а обонятельные доли мозга
имеют небольшие размеры.
Б и о л о г и ч е с к и е к о о р д и н а ц и и проявляю тся в согла­
сованном развитии органов, которые не связаны ни функционально, ни
топографически. В качестве примера приведем зависимость между
формой зубов, длиной кишечника и специализацией конечностей у
плотоядных и растительноядных животных. У плотоядных в соответ­
ствии с особенностями экологии хорошо развиты клыки, кишечник
короткий, на ногах когти. У растительноядных развиты зубы с жева­
тельной поверхностью, кишечник длинный, конечности несут копыта.
Зависимость между координированными органами в этом случае
только косвенная. Связую щ им и определяющим звеном здесь служат
условия среды обитания организма (водная, воздушная, род пищи). Так,
405
способ добывания пиши определяет зависимость между длиной шеи и
длиной передних конечностей у копытных.
Акуловые рыбы не имеют плавательного пузыря, их вес ‘тяжелее
воды, поэтому им трудно удерживаться на одной глубине. В соответ­
ствии с этим они имеют уплошенное, широкое брюхо, а плавники рас­
положены в горизонтальной плоскости. У рыб, имеющих плава­
тельный пузырь, изменяются согласованно оба признака. Координация
определяется существованием рыб в водной среде.
Ещ е одним примером биологической координации является связь
между формой зубов и челюстного сустава у хищных и копытных мле­
копитающих. Редукция клыков и развитие складчатых зубов у траво­
ядных связана с изменением формы челюстного сустава — уплощение и
расширение суставных поверхностей, что определило возможность
скольжения нижней челюсти вперед, назад и в стороны. Полезность
таких движений первично определяется характером пищи.
Так же как и корреляции, многие координации характеризуются
устойчивостью . Таким свойством отличаю тся прежде всего динамиче­
ские координации, поскольку возможность изменения структуры
органов ограничивается необходимостью сохранения присущих им
функций. Устойчивостью характеризуются также многие топографиче­
ские координации, так как они основаны на стойких морфогенетических
корреляциях. Устойчивость координаций, так же как и корреляций,
составляет основу преемственности форм в процессе исторического
развития.
7.12.6. Возникновение и исчезновение органов в филогенезе.
Рудименты и атавизмы
В процессе эволюции одни органы появляю тся, тогда как другие
могут исчезать. Механизм этих явлений в настоящее время раскрыт.
Основу возникновения новых органов составляет принцип дифференци­
ации, а необходимую предпосылку — мультифункциональность предсу­
ществующих органов. В связи с развитием органов в новых направлени­
ях происходит смена функций. Примером этому служит возникновение
млечных желез млекопитающих, которые гомологичны потовым
железам. В классе Млекопитающих встречаю тся виды с групповым
расположением потовых желез в виде очаговых скоплений. Поскольку
предки млекопитающих высиживали яйца (ехидна, утконос) и согревали
детенышей телом, детеныши могли слизывать секрет потовых желез,
расположенных на брюхе. Очевидно, пот содержал некоторое
количество питательных веществ, поэтому мог играть определенную
роль в выживании потомства. В процессе естественного отбора
сохранялись формы с большим размером желез и увеличенным
количеством питательного материала в их секрете. Строение желез
постепенно усложнялось и в конечном итоге возник новый орган —
молочная железа, секрет которой — молоко — обеспечивает сущ ество­
вание детеныша на ранних стадиях постэмбрионального развития,
повышая тем самым выживаемость вида.
306
Р е д у к ц и я ( и с ч е з н о в е н и е ) о р г а н о в обычно связана
с превращением полезного органа в относительно бесполезный или.
даже, что встречается очень редко, вредный. Чащ е подобное
превращение вы зы ваю т изменения окружающей среды, иногда появле­
ние другого органа, функционально замещающего первый (субститу­
ция). Примером может служить редукция головной почки анамний
после появления туловищ ной. Редукция проявляется в уменьшении
размеров органа и упрощении его строения. Для редуцированных
Органов характерна повышенная изменчивость, что имеет значение в
медицинской практике. Так, длина червеобразного отростка (аппендик­
са) колеблется от 20 до 2 см. Степень редукции может б ы ть различной.
Иногда орган исчезает полностью, отсутствуя и у взрослых, и у
зародышей. В других случаях он может бы ть представлен только у
зародыша в виде эмбрионального зачатка (головная почка у высших
позвоночных). Наконец, орган нормально развивается до определенной
стадии, после чего его развитие останавливается, и он сохраняется у
взрослых особей в недоразвитом состоянии, не выполняя присущей ему
функции. Такие органы получили название р у д и м е н т а р н ы х
или р у д и м е н т о в . Примерами рудиментарных органов у человека
служат слепая кишка, аппендикс, мигательная перепонка глаза,
волосяной покров, мыш цы ушной раковины.
Редукция бргана, ставшего бесполезным, обусловлена тем , что он
перестает контролироваться естественным отбором. В результате
происходит накопление мутаций, вызывающ их нарушение его строения.
Возникающие изменения наруш ают коррелятивные связи органа с
другими частями организма, что приводит к отклонениям в нормаль­
ном развитии и, в конечном итоге, к остановке развития этого органа на
той или иной стадии. При тяжелых нарушениях связей не происходит
даже закладки органа в эмбриогенезе.
Процесс недоразвития органов имеет свои закономерности.
Последовательность выпадения этапов морфогенеза является обратной
по отношению к нормальной последовательности формирования
органа, т. е. прежде всего выпадаю т конечные стадии развития, затем
более ранние и т . д. Например, при недоразвитии конечностей прежде
всего выпадаю т закладки фаланг I и V пальцев, которые в норме
закладываются последними. Если редукция идет дальше, нарушается
закладка последующих дистальных отделов, в то время как прокси­
мальные (плечо, бедро) сохраняются. В отдельных случаях развитие
останавливается на стадии нерасчлененной конечности.
Некоторые органы, подвергающиеся редукции, присутствую т в виде
эмбриональных закладок на протяжении многих поколений. В этом
случае закладка органа сохраняется благодаря ее роли в эмбриогенезе в
качестве элемента коррелятивных систем, определяющих развитие
других жизненно важных органов. Поэтому процесс редукции не
распространяется на ранние этапы формирования органа. Примером
служит развитие у высших позвоночных хорды. В отсутствие редукции
органы предков переходят во взрослое состояние(атавизмы).
Выш е были рассмотрены общие принципы и способы филогенетиче­
ских преобразований органов. Далее следует материал, иллюстрирую307
щий основные направления и этапы филогенеза систем органов в ряду
позвоночных и человека.
Д ля удобства изучения филогенез каждой системы рассматрива­
ется отдельно. При этом не следует забы вать, что организм представ­
ляет собой целостную систему и изменение одного органа вы зывает
преобразования в других частях его. Прежде всего это касается
структур, которые образуют функциональные системы. Конечной
целью эволюционной морфологии является формирование представле­
ний о путях эволюции организма в целом.
Поскольку человек относится к типу Хордовых и подтипу
Позвоночных, наибольший интерес для будущих врачей представляют
преобразования органов в филогенезе этой группы животных.
7.13. Ф И Л О Г Е Н Е З С И С Т Е М О Р Г А Н О В П О ЗВ О Н О Ч Н Ы Х
7.13.1. Наружные покровы
Начиная с низших хордовых обнаруживается подразделение
наружных покровов или кожи на поверхностный эпителиальный слой
эктодермального происхождения (эпидермис) и подстилающий его
соединительнотканный, развивающийся из мезодермы (кориум или
собственно кожа). У л а н ц е т н и к а покровные ткани развиты
слабо, эпителий однослойный, цилиндрический, содержит отдельные
железистые клетки. Корнум представлен незначительным слоем
студенистой соединительной ткани.
В подтипе Позвоночных продолжается дифференцировка кожи на
отчетливо различимые эпидермис и кориум. Эпидермис становится
многослойным, его нижний слой состоит из цилиндрических клеток,
активно размножающихся и пополняющих поверхностные слои клеток.
Кориум представлен основным веществом, волокнами и клетками.
Кож а образует ряд придатков, главными из которых являю тся
защитные образования и железы.
Ры бы . У х р я щ е в ы х
р ы б эпидермис содержит большое
количество одноклеточных слизистых желез. Кориум плотный,
волокнистый. Все тело покрыто плакоидной чешуей, представляющей
собой пластинки, несущие шип или зубец. Ее основание лежит в
кориуме, а шип прободает эпидермис и выходит наружу. Чеш уя состоит
из дентина — соединения органического вещества с известью , более
твердого, чем кость, и не содержащего клеток.
ч, Закладка плакоидной чешуи образуется на границе эпидермиса и
кориума (рис. 131). Нижний слой эпидермиса приобретает форму
колпачка, в который внедряется в виде сосочка масса крезодермальных
клеток. Клетки, образующие стенки колпачка, становятся цилиндриче­
скими. Подлежащие клетки мезодермы (склеробласты) также распола­
гаю тся упорядоченно, сплошным слоем. Клетки этого слоя образуют
дентиновую пластинку — основание чешуи, охватывающ ее мезодермальный сосочек. Беспорядочно расположенные в его середине
клетки образую т пульпу. Дальнейшее утолщение дентина происходит за
счет слоя склеробластов, на поверхности которых возникаю т все новые
308
Рис. 131. Закладка плакоидной чешуи.
1
б -
сосочек кориума. 2
дентин
клетки эмалеобразователи; 3 — эмаль; 4 — эпидермис; 5 - склероблчсты;
слои дентина, благодаря чему шип растет и проходит через эпидермис.
Снаружи шип покрыт эмалью , еше более твердой, чем дентин.
У к о с т н ы х р ы б тело также покрыто чешуей, но в отличие от
хрящевых рыб — костной. Чеш уя имеет вид округлых тонких
пластинок, черепицеобразно налегающих друг на друга и снаружи
покрытых тонким слоем эпидермиса. Еаавитие костной чешуи идет
целиком за счет кориума, без участия эпидермиса. Филогенетически
"костная'чеш уя связана (Гболее примитивной плакоидной чешуей.
Амфибии. Кож а з е м н о в о д н ы х
голая, лишена чешуи.
Ороговение верхнего слоя выражено слабо. Кориум представлен
волокнами соединительной ткани, идущими строго параллельно, и
клеточными элементами. В коже много слизистых желез. Кожные
железы создают на поверхности жидкостную пленку, которая способ­
ствует газообмену (кожное дыхание) и защищает кожу от высыхания,
так как слабое ороговение не предохраняет земноводных от потери во­
ды. Кроме того, бактерицидные свойства секрета желез препятствую т
проникновению микробов. Ядовитые железы защищают животное от
врагов.
Рептилии. В связи с переходом к наземному образу жизни у
рептилий увеличивается степень ороговения эпидермиса (защ ита от
высыхания и от повреждений). Чеш уя становится роговой. Эпидермис
отчетливо подразделяется на д.ва слоя: нижний (мальпигиев), клетки
которого интенсивно размножаются, и верхний (роговой), содер­
жащий клетки, постепенно отмирающие в результате особого рода
перерождения. В клетках появляю тся капли кератогиалина — рого­
вого вещества, количество которого постепенно увеличивается, ядро
исчезает, клетка уплошается и превращается в твердую роговую че­
шуйку, которая затем слущивается. Благодаря размножению клеток
мальпигиевого слоя клетки рогового слоя постоянно восполняются.
Развитие роговой чешуи поначалу идет также как и костной. Отличия
развития наблюдаются на завершающей стадии и заклю чаю тся в
преобразовании эпидермиса. У рептилий отсутствую т кожные
железы.
Млекопитающие. Кож ны е покровы млекопитающих имеют осо­
бенно сложное строение. Оба слоя — эпидермис и кориум хорошо
развиты. Эпидермис дает начало многим производным кожи —
волосы, ногти, когти, копыта, рога, чешуя, различные железы.
Собственно кожа приобретает значительную толщ ину и состоит
преимущественно из волокнистой соединительной ткани. В нижней
части кориума формируется слой подкожной жировой клетчатки.
Характерной особенностью млекопитающих является волосяной
покров, основная функция которого — защ ита тела от потери тепла.
Волосы представляют собой роговые придатки сложного строения.
У взрослого .человека волосяной покров имеется на всем теле, кроме
ладоней и подошв, но сильно редуцированный.
В коже содержится большое количество многоклеточных желез —
потовые, сальные и млечные. Потовые железы млекопитающих
гомологичны кожным железам амфибий. Иногда потовые железы
образуют местные скопления. Секрет потовых желез, как правило,
имеет жидкую консистенцию и может бы ть слизистым или белковым
по составу, или содержит жир. П отовые железы играют важную роль в
процессах выделения и терморегуляции. Испарение пота связано с
большой потерей тепла.
Сальные железы выделяю т секрет, который смазывает волосы и
поверхность кожи, защищая ее от воздействия среды. Появление
сальных желез является отличительным признаком млекопита­
ющих.
"
—
М лечные железы гомологичны потовым железам. Наиболее
близкое сходство с потовыми железами имеют млечные железы
клоачных млекопитающих (ехидна, утконос), у которых они расположе­
ны группой на так называемом железистом поле, которое находится в
сумке для вынашивания яиц и детенышей. Секрет стекает на
поверхность и слизывается детенышами. У сумчатых животных име­
ется сосок, где каждая железа открывается собственным отверстием.
По краям развивающегося соска можно встретить все последова­
тельные переходы между обычными потовыми и типичными млечными
железами.
У живородящих по бокам брюха закладывается парная полоска
утолщенного эпителия — млечная линия, а на ней млечные железы и
соски. Расположение и число желез, а также сосков весьма различно.
У видов, отличающ ихся плодовитостью, число сосков доходит до
22—25. С уменьшением числа рождаемых детенышей уменьшается
число сосков, обычно встречается 1— 8 пар. У летучих мышей, обезьян
и человека только одна пара сосков на груди, однако в эмбриогенезе у
человека их закладывается 5 пар. Иногда рождаются люди с дополни­
тельными сосками, которые остаю тся во взрослом состоянии.
Основное направление эволюции наружных покровов — дифференци310
ровка слоев кожи и ее производных (железы, чешуя, перья, волосы),
-Обеспечивающих защ иту от разнообразных воздействий среды — вы•'сыхания, механических воздействий, потери тепла и перегревания.
7.13.2. Скелет
Среди беспозвоночных чаше встречается н а р у ж н ы й с к е л е т
в виде кутикулярных образований эктодермального эпителия. Наиболее
развит подобный скелет у членистоногих. Он состоит из хитина,
защищает тело от механических повреждений, высыхания и служит
местом прикрепления мышц.
У низших хордовых (бесчерепных) появляется в н у т р е н н и й
о с е в о й с к е л е т в виде хорды и плотных волокнистых тяжей, под­
держивающих плавники и жаберные щели. Хорда представляет собой
упругий тяж , состоящий из особых вакуолизированных клеток
(производных энтодермы). Она тянется по спинной стороне от
переднего конца тела до заднего. По поверхности хорду облегает
эластичная оболочка. Опорная функция хорды обеспечивается упруго­
стью оболочек и клеточных вакуолей, поддерживающих в клетках
значительное внутреннее давление (тургор).
У высших хордовых (позвоночных) скелет высокой степени
дифференцировки.
Осевой скелет. У низших позвоночных — к р у г л о р о т ы х
и
н и з ш и х р ы б — хорда сохраняется в течение всей жизни. Но
одновременно появляю тся верхние (у круглоротых) и нижние (у рыб)
дуги позвонков в виде парных хрящей, расположенных метамерно над
хордой и под хордой. Функционального значения дуги не имеют.
У в ы с ш и х р ы б , кроме дуг, развиваю тся тела позвонков — или за
счет разрастания оснований дуг, образующих вокруг хорды кольцо
хрящевой или костной ткани, или частично за счет дуг, а частично из
скелетогенной ткани, окружающей хорду. После образования тела
позвонка к нему прирастают дуги. Концы верхних дуг срастаю тся
между сосбой, образуя канал спинного мозга и остистый отросток,
нижние дуги дают боковые вы росты (поперечные отростки). Таким
образом, первоначально каждый позвонок складывается из нескольких
элементов. У р ы б хорда сдавливается позвонками н приобретает вид
четкообразного шнура. Позвоночник дифференцируется на туловищ ный
и хвостовой отделы. Все позвонки туловищного отдела несут ребра.
В хвостовом отделе ребра отсутствую т.
В позвоночнике
амфибий
дифференцируются два новых
отдела — шейный и крестцовый, представленные каждый одним
позвонком. Шейный отдел обеспечивает подвижность головы, необхо­
димую в более сложных условиях наземной среды. Позвонок несет
ребра. Крестцовый отдел возникает на границе хвостового и туло­
вищного, дает опору тазовым костям и задним конечностям. Туловищ­
ный отдел представлен пятью позвонками, которые несут ребра незна­
чительной длины. Они не доходят до грудины и кончаю тся свободно.
У
рептилий
число отделов позвоночника увеличивается;
появляется »| вый отдел — поясничный. Количество позвонков в отде­
лах увеличивается до 8— 12. В шейном отделе происходят прогрессив­
ные преобразования. Тело первого шейного позвонка не соединя­
ется дугами, а срастается с телом второго шейного позвонка, образуя
зубовидный отросток. Первый шейный позвонок приобретает форму
кольца и может свободно вращаться на втором позвонке, что резко
увеличивает подвижность головы. Ребра в шейном отделе редуциру­
ются. В грудном отделе все позвонки несут хорошо развитые ребра.
Большинство из них соединяется с грудиной, образуя грудную клетку.
Появление грудной клетки обеспечивает более совершенный механизм
дыхания. Для поясничного отдела характерны массивные поперечные
отростки, образованные за счет прнрастания рудиментарных ребер.
У
млекопитающих
во взрослом состоянии хорда
сохраняется только в виде пискш ри1ро$ш позвонков. Позвоночник
состоит из пяти отделов — шейный, грудной, поясничный, крестцовый,
хвостовой. Характерно постоянное число позвонков в шейном отделе,
равное 7. Ребра шейных позвонков полностью редуцированы. В груд­
ном отделе количество позвонков колеблется от 9 до 14, чаще 12— 13.
Позвонки несут ребра, большинство их соединяется с грудиной.
Поясничный отдел содержит от 2 до 9 позвонков, имеющих мощные
поперечные отростки. Крестец образован сросшимися позвонками, в
числе 10 и более. Количество позвонков в хвостовом отделе варьирует.
Как отражение некоторых этапов филогенеза позвоночных у
эмбрионов человека на ранних стадиях развития все позвонки имеют
ребра, которые в последующем развиваются в полной мере только в
грудном отделе. Однако иногда редукция ребер не происходит или она
задерживается, тогда возникают дополнительные ребра. Так, довольно
частой аномалией скелета у человека являются шейные ребра у нижнего
шейного позвонка. Аналогичным образом развивается добавочное
ребро, связанное с первым поясничным позвонком. Среди других
аномалий следует отметить развитие хвостового отдела позвоночника.
В возрасте 1,5— 3 мес хвостовой отдел позвоночника эмбрионов челове­
ка представлен 8— 11 зачаточными позвонками. К концу 3-го мес
наружный отдел хвоста редуцируется и погружается в складку между
быстро растущими ягодицами. Во внутренней части хвоста количество
позвонков уменьшается до 6— 2, чаше остается 4. Впоследствии они
срастаются между собой, образуя копчик. В отдельных случаях
хвостовой отдел позвоночника не останавливается в развитии, а
продолжает расти одновременно с другими частями тела. В результате
у человека развивается хвостовой придаток. В литературе описаны
случаи, когда длина хвоста превышала 20 см. Во взрослом состоянии
позвоночник человека приобретает 5-образный изгиб, связанный с
прямохождением.
Скелет свободной конечности. Впервые конечности возникают у
р ы б в виде парных плавников — грудных и брюшных, которые в
процессе эволюции преобразуются в пятипалые конечности — органы
передвижения наземных животных.
У большинства рыб в скелете грудного плавника различают
проксимальный отдел, состоящий из небольшого числа (1— 3)
сравнительно крупных хрящевых пластинок, и дистальный отдел,
312
б
в
Рис. 132. Скелет парного плавника кистеперой рыбы и стегоцефала.
а — п лечевой п ояс и п ла вн и к кистеперой р ы бы ; б — внутренний ск е л е т п л а вн и к а ; в — ске ле т передней
конечности стегоц еф ала; 1 — элем ент, гом ологичны й плечевой кости; 2 — элем ент, гомологичный л уч ев о й
кости; 3 — элем ент, гомологичный л октево й ко сти ; 4 - кости п р окси м ал ьно го р яд а з а п я с т ь я ; 5 — кости
мед иально го рода з а п я с т ь я ; 6 — кости д и ста льн о го р яд а з а п я с т ь я ; 7 — ф ал ан ги п ал ьц ев.
построенный из большого числа радиально расположенных тонких
лучей. Каж дый луч состоит их большого числа мелких элементов,
расположенных по его осй. Все части скелета плавника неподвижно
соединены между собой и образуют единую плоскость. С плечевым
поясом плавник соединен неподвижно, поскольку в сочленении
участвует несколько элементов проксимального отдела. У подавляю­
щего большинства рыб плавники не могут служить опорой для тела,
а используются как средство для изменения направления движе­
ния (повороты). Исключение составляют плавники ископаемых
к и с т е п е р ы х р ы б (Сго$5ор1еп§1а), широко распространенных в
девонском периоде (примерно 300 млн. лет назад) и затем вымерших.
Лиш ь одна из ветвей кистеперых сохранилась до наших дней в районе
юго-восточного побережья Африки.
Как показывают материалы палеонтологии, у кистеперых рыб в
скелете плавников возник ряд изменений, подготовивший их преобразо­
вание в пятипалую конечность. Произошло уменьшение числа костных
элементов скелета плавников и их укрупнение путем слияния.
Проксимальный отдел плавника состоит у этих рыб только из одного
крупного костного элемента. За ним следуют два массивных элемента,
гомологов костей предплечья наземных форм. К ним прикрепляются
радиально расположенные лучи, число которых уменьшается по
сравнению с остальными ры бам и— до 7— 12 (рис. 132). Вторая
особенность парных плавников кистеперых — высокая подвижность в
сочленении с плечевым поясом. Так как в соединении участвует толь­
ко один костный элемент, плавник может совершать вращательные
движения. Благодаря указанным преобразованиям плавники этой
группы рыб начинают служить опорой для тела и могут использоваться
для передвижения по твердому субстрату. Э т а новая функция плавников
была обусловлена особенностями существования кистеперых рыб. Они
обитали в мелких густо заросших, часто пересыхающих водоемах, где
нередко единственно возможным способом передвижения являлось
313
переползание с помощью плавников. Одновременно резкие колебания
уровня воды и содержания кислорода вызвали у этих рыб появление до­
бавочных органов дыхания, позволяющих использовать кислород
воздуха (см. 7.13.4). В период климатических изменений на Земле, в
частности при сокращении водной поверхности при крупных горнообра­
зовательных процессах, кистеперые рыбы, обладая органами атмос­
ферного дыхания и конечностями для передвижения по твердой
поверхности, смогли выжить и освоить новую среду обитания — сушу,
где они дали начало земноводным.
Первые
амфибии
(стегоцефалы) обладали конечностями
пятипалого типа. Их скелет по плану строения и соотношению костей
имел большое сходство с плавниками кистеперых (см. рис. 132, в). Так
же как и у кистеперых рыб, проксимальный отдел представлен одним
крупным элементом (плечо), за ним следует 2 костных элемента,
составляющих предплечье, затем идет 3—4 ряда мелких костей,
сохраняющих правильное радиальное расположение (запястье). После
запястья следует пястье (5 костей) и, наконец, фаланги пальцев, также
сохраняющие радиальный тип расположения костей. Такой план
строения скелета единый для всех наземных позвоночных.
Наряду с упрощением строения и уменьшением числа элементов
важным моментом в процессе преобразования плавников в конечности
наземного типа явилась замена прочного соединения элементов скелета
между собой подвижными сочленениями в виде суставов. В результате
конечность из простого рычага превратилась в сложный рычаг, части
которого подвижны относительно друг друга. Процесс упрощения
скелета конечности кистеперых продолжался и позднее. Основные
изменения затронули дистальный отдел. Так произошло дальнейшее
уменьшение количества лучей. Предки наземных форм имели 7 пальцев,
соединенных перепонкой. При выходе на сушу крайние пальцы
редуцировались и превратились в рудименты. Сократилось также
количество костных элементов в запястье. Амфибии имеют 3 ряда
костей запястья — проксимальный, средний и дистальный. У
в ы с ш и х п о з в о н о ч н ы х исчезает средний ряд, а количество
костей в каждом ряду последовательно уменьшается, так же как и
фаланг. Одновременно в процессе эволюции наземных форм происходит
значительное удлинение костей проксимальных отделов — плеча,
предплечья, а также дистального отдела (пальцы), в то время как кости
среднего отдела укорачиваются.
Все классы наземных позвоночных имеют общий план строения
передней конечности, но у различных форм мы встречаемся с большими
или меньшими отклонениями от единой схемы. Э ти изменения —
результат приспособления к различным условиям существования или
специализации. Например, роющая передняя конечность крота при­
обрела лопатообразную форму и несет шесть пальцев. В передней
конечности птицы, приспособленной для полета, сросшиеся кости
запястно-пястного отдела образуют опору для прикрепления мышц.
Рука
человека
сохраняет план строения конечности
предков — плечо, предплечье, запястье, пястье, фаланги пальцев.
Вместе с тем она имеет отличия, связанные с ее новой функцией — прев1
314
ращением в орган труда. Осо­
бенности строения и исключи­
тельное многообразие кон­
кретных функций руки челове­
ка возникли в процессе освое­
ния трудовой деятельности.
Рука, таким образом, как от­
мечал Ф . Энгельс, является не
только органом, но также и
продуктом труда.
Происхождение
парных
конечностей. Поскольку ко­
нечности ПОЗВОНОЧНЫХ ВОЗНИ- „
п
Рис. 13о. Происхождение парных плавникоп
кают в виде парных плавников ры6
у
у
рыб, ОСНОВНЫМ ЯВЛяеТСЯ ВО- I — боковы е п арны е с к л а д к и ; 2 — н а ча л ь н ы е ф азы
ППОС
О Пк
р ои схож д
обособления кон ц евы х о тд ело в; 3 — возникновение пар^
ехен и и в ПРОн
ных плавннков
цессе эволюции именно этих
структур.
Согласно существующим представлениям парные плавники возник­
ли из кожных складок, расположенных по бокам туловища. Такого рода
образования среди ныне живущих организмов встречаются у ланцетни­
ка в виде метаплевральных складок и у личинок рыб. Кожные складки
древнейших рыб располагались горизонтально и тянулись от переднего
конца тела к заднему, не имея ни скелета, ни собственной мускулатуры.
Считаю т, что они служили для увеличения брюшной поверхности
животного. Низшие рыбы не имеют плавательного пузыря, а удельный
вес их больше удельного веса воды. Поэтому горизонтальные складки,
увеличивая брюшную поверхность, замедляли погружение тела в воду.
Позднее в кожных складках появились собственный скелет в виде лучей
и мускулатуры, расположенные метамерно в каждом сегменте. С по­
явлением скелета, мускулатуры и увеличением размеров складки
приобрели новые функции. Они создавали поверхность, обеспечиваю­
щую подъемную силу (аналогично крыльям самолета) и служили
рулями глубины. Такие функции выполняют плавники современных
низших рыб.
Для изменения положения тела рыбы в воде наибольшее значение
имели крайние отделы складок, как наиболее удаленные от центра
тяжестн. Поэтому в процессе естественного отбора сохранились формы
с большим развитием крайних отделов складок. Средняя часть, не
имеющая функционального значения, постепенно редуцировалась.
В результате со временем передние и задние концы складок оказались
разобщенными, самостоятельными образованиями (рис. 133). За счет
слияния оснований лучей образовались пояса конечностей. У высших
рыб произошло перемещение плавников в вертикальную плоскость. На
последующих этапах эволюции богато расчлененные плавники дали
начало пятипалым конечностям наземных.
Теория происхождения парных плавников из боковых складок имеет
ряд доказательств. Так, парные плавники эмбрионов современных рыб
закладываются в виде горизонтальных складок, имеющих большую
протяженность. Позднее их длина значительно сокращается. Мускула315
тура парных плавников развивается в виде метамерно расположенных
мускульных почек, причем- у некоторых рыб мускульные почки
закладываются на всем пространстве между передней и задней парой
плавников. Скелет плавников вначале имеет также метамерное
расположение и лишь затем концентрируется.
Следует отметить, что у эмбрионов человека пятипалые конечности
закладываются в виде небольших горизонтальных складок, по форме
напоминающих плавники.
Скелет головы (череп). Череп позвоночных состоит из 2 основных
отделов — осевого и висцерального черепа. О с е в о й
отдел
( ч е р е п н а я к о р о б к а ) представляет собой продолжение осевого
скелета и служит для защиты головного мозга и органов чувств.
В и с ц е р а л ь н ы й о т д е л ( л и ц е в о й ч е р е п ) образует опору
для передней части пищеварительного тракта.
Оба отдела черепа развиваются независимо друг от друга и
различными путями. Наиболее значительные преобразования в про­
цессе эволюции происходят в висцеральном черепе, элементы которого
преобразуются в челюстной аппарат, а у высших, кроме того, дают
начало элементам органа слуха.
Особенно отчетливо происхождение основных элементов черепной
коробки у позвоночных прослеживается в эмбриональном периоде.
Черепная коробка у зародышей всех позвоночных закладывается под
головным мозгом в виде двух пар хряшей. Задняя пара хряшей пред­
ставляет собой удлиненные пластинки, расположенные по бокам
переднего конца хорды, и поэтому названные парахордалии. Передняя
пара хряшей или трабекула лежит впереди от хорды, под основанием
переднего мозга. Одновременно образуются парные хряшевые капсулы
вокруг зачатков органов слуха, обоняния, а также глазничные хряши.
Разрастаясь, трабекулы и парахордалии сливаются между собой,
образуя черепную коробку, прикрывающую мозг снизу и с боков.
Слуховые и обонятельные капсулы также прирастают к черепной
коробке. Глазничные хряши заполняют боковые стенки.
Положение парахордалий по отношению к хорде сходно с положе­
нием элементов позвоночника по- отношению к хорде в туловишной
части. Кроме того, в задней части парахордалий обнаруживаются
следы сегментации. Н а этом основании считают, что парахордальные
хряши возникли путем слияния определенного числа позвонков
(рис. 134). Трабекулу не имеют сходства со структурами осевого
скелета. Висцеральная часть черепа у зародышей позвоночных состоит
из метамерно расположенных хряшевых дуг, охватывающих подобно
обручам начальный отдел пищеварительного тракта. Дуги отделены
дгуг от друга висцеральными щелями. Дуги принято обозначать
порядковыми номерами в соответствии с их расположением по
от ношению к черепу. Ближайшая к черепной коробке пара — первая,
следующая кзади — вторая и т. д. Уж е у зародыша две передние дуги
О-я и 2-я) отличаются от остальных расчленением на меньшее
количество составных элементов и большей массивностью (рис. 135).
В процессе дальнейшего развития именно эти дуги претерпевают
сложное превращение, преобразуясь в челюсти, а у высших, кроме того.
б
II
Рис. 134. Распределение хрящевых закладок осевого черепа акулы (вид со спинной
стороны). Закладки обведены контурами черепа и головы взрослого животного.
1 — хорда; 2
парахордадии; 3 — трабекулы; 4 6 — глазничные хрящи; 7 — глаз;
8 — гипофиз
обонятельные капсулы; 5 — слуховая капсула;
Рис. 135. Хрящевой череп акулы.
а — зар о д ы ш : 1 — че л ю с т н а я д у га; 2 — п о д ъ я з ы ч н а я д у га; 3 -- 3 -Б-я ж а б е р н ы е д уги . 4
слуховая кап ­
сула; 5
хорда; 6 — к и ш к а ; 7 — спииной м озг; 8 — передний м озговой п узы р ь; 9 — средний
м озговой п у зы р ь;
10 — гл азн и чн ы е хрящ и; 1 1 — парахордалии; 12 — тр а б е к у л ы ; б — в » р о ?л т я ос*»бь;
1 — осевой чер еп; 2 — н еб н оквад р атн ы й х р ящ ; 3 — ниж н ий х р ящ 1-й д у ги ; 4 ~ 1 и ом андибудярны и х р я щ ;
5 — гиоид; 6
10 — ж аб ер н ы е дуги; 11 — коп ул а.
дают начало опорному аппарату языка и элементам органа слуха.
Поскольку 1-я дуга у большинства современных позвоночных приобре­
тает функцию челюстного аппарата, ее называют еше челюстной дугой,
а 2-ю дугу — тоже по функции — подъязычной или гиоидной.
Остальные дуги, начиная с 3-й, называют жаберными, поскольку у
низших они служат опорой для жаберного аппарата.
Н а ранних этапах развития висцеральный и осевой отделы черепа не
связаны между собой, но позднее такая связь возникает.
Общие для всех зародышей закладки осевого и висцерального черепа
в процессе постэмбрионального развития претерпевают изменения в
соответствии с особенностями исторического развития каждого класса.
У н и з ш и х р ы б (хрящевых) осевой череп во взрослом состоя­
нии более плотно охватывает мозг. Появляется затылочный отдел,
слуховые капсулы включаются в боковые стенки, обонятельные хряши
присоединяются к передней части черепа. Висцеральный череп состоит
из ряда хрящевых висцеральных дуг, охватывающих глотку наподобие
обруча (см. рис. 135), из которых 1-я (челюстная) дуга состоит всего из
двух крупных хрящей, вытянутых в передне-заднем направлении —
верхнего (небноквадратного) и нижнего (меккелева). Верхние и нижние
317
хряши каждой стороны срашены между собой и выполняют функции
челюстей (первичные челюсти). 2-я висцеральная дуга состоит из двух
парных и одного непарного хряша, соединяющего снизу парные хрящи
между собой. Верхний элемент пары, более крупный, — гиомандибулярный хрящ, нижний парный элемент — гиоид, непарный — копула.
Верхний край гиомандибулярного хряща соединен с черепной коробкой,
нижний с гиоидной, а передний — с лежащей впереди челюстной дугой.
Таким образом, гиомандибулярный хрящ выполняет роль подвеска для
челюстной дуги, она присоединяется к черепу с помощью подъязычной
дуги. Такой тип соединения челюстей с черепом называется гиостилией
(гиостильный
ч е р е п ) и свойствен низшим позвоночным,
Остальные дуги (3—7) образуют опору для дыхательного аппарата.
У в ы с ш и х р ы б (костные), наряду с первичным, хрящевым
черепом, гомологичным осевому черепу низших рыб, появляется
вторичный череп из накладных костей. Вторичный череп значительно
шире первичного. Он покрывает первичный череп сверху (парные
теменные, лобные, носовые кости), снизу (большая непарная кость —
парасфеноид) и с боков (надвисочная, чешуйчатая кости). Основные
изменения висцерального черепа касаются челюстной дуги. Верхняя
челюсть вместо одного крупного небноквадратного хряша состоит из
5 элементов — небный хрящ, квадратная кость и 3 крыловидных
кости. Впереди от первичной верхней челюсти образуются 2 крупные
накладные кости — предчелюстная и челюстная, снабженные крупными
зубами, которые становятся вторичными верхними челюстями.
Дистальный конец первичной нижней челюсти также покрывает
большая зубная кость, она выдается далеко вперед и образует
вторичную нижнюю челюсть. Таким образом, функция челюстей у
высших рыб переходит к вторичным челюстям, образованным
накладными костями. Подъязычная дуга сохраняет прежнюю функцию
подвеска челюстей к черепу. Следовательно, череп высших рыб также
гиостильный.
У земноводных
значительные изменения касаются в
основном висцерального отдела, поскольку с переходом к наземному
образу жизни происходит замена жаберного дыхания кожно-легочным.
Первичный череп амфибий почти не подвергается окостенению и не
отличается от первичного черепа рыб. Для вторичного черепа
характерно выраженное сокращение числа костных элементов.
В отношении висцерального черепа одно из главных отличий
заключается в новом способе соединения челюстной дуги с черепом.
Амфибии в отличие от гиостильного 4черепа рыб имеют череп
аутостильный,
т. е. у них челюстная дуга соединена с
черепом непосредственно, без помощи подъязычной дуги, благодаря
срастанию небного хряша челюстной дуги (первичная верхняя челюсть)
на всем протяжении с осевым черепом. Нижнечелюстной отдел
сочленяется с верхнечелюстным и, таким образом, тоже получает связь
с черепом без помощи подъязычной дуги. Благодаря этому гиомандибу­
лярный хрящ освобождается от функции подвеска челюстей (рис. 136).
Новый способ присоединения челюстей к черепу влечет ряд важных
преобразований в подъязычной дуге. Гиомандибулярный хряш сильно
318
Рис. 136. Типы сочленений нижней челюсти с черепом.
а
ги ости л ичсский череп а к у л ы ; б
а у то сти л и ч е с ки й череп ам ф ибии; I - гиом андибулирный хр ящ .
2
гиоид, 3 — ниж н ий х р ящ (м е к к е л е в ) че л ю стн о й дуги; 4
н еб н окнадратны й х р ящ верхней че л ю сти ;
5 — н и ж н я я че л ю с т ь
редуцируется и приобретает совершенно новую функцию — входит в
состав слухового аппарата в качестве слуховой косточки (столбик).
Смене функций способствовало расположение гиомандибулярного
хряща рядом со стенкой слуховой капсулы и превращение первой
висцеральной шели, в которой находился редуцированный гиомацдибулярный хрящ, в воздухоносную полость среднего уха. Оказавшись
внутри барабанной полости, гиомандибулярный хряш становится
слуховой косточкой, которая участвует в передаче звуковых колебаний
внутреннему уху. Э то имело важное значение при освоении амфибиями
наземной среды, поскольку обеспечило более совершенное восприятие
звуковых сигналов, которые в наземных условиях проводятся слабее,
чем в водной среде. Частично подъязычная дуга — гиоидный хрящ
вместе с жаберными дугами образует опору для языка и подъязычный
аппарат, частично — гортанные хрящи.
У зародышей р е п т и л и й также закладываются четыре пары
жаберных дуг и жаберные шели, из которых прорывается наружу
только одна, а именно первая, расположенная между челюстной и
подъязычной дугами, тогда как остальные быстро исчезают. Осевой
череп в отличие от земноводных состоит только из костной ткани.
Висцеральный череп рептилий, так же как и амфибий является
аутостильным. Однако есть и некоторые отличия. Передний элемент
первичной верхней челюсти — небный хряш — редуцируется. Поэтому
в причленении верхней челюсти к черепу участвует только задний
отдел — квадратная кость. Соответственно плошадь поверхности
причленения уменьшается. С квадратной костью верхней челюсти
соединяется нижняя челюсть и таким путем присоединяется к черепу.
Единственная жаберная щель, прорывающаяся наружу в эмбрио­
нальном периоде, преобразуется в полость среднего уха, а гиомандибу­
лярный хряш — в слуховую косточку. Остальная часть висцерального
скелета образует подъязычный аппарат, который состоит из тела
подъязычной кости и трех пар отростков. Тело подъязычной кости
образуется за счет срастания копул подъязычной дуги и всех жаберных
дуг (см. рис. 135). Передние рожки этой кости соответствуют нижнему
парному элементу подъязычной дуги — гиоиду, а задние — парным
элементам двух первых жаберных дуг.
319
В осевом черепе м л е к о п и т а ю щ и х происходит уменьшение
числа костей за счет их слияния. Резко меняется конфигурация черепа,
что связано с прогрессивным увеличением объема мозга. В частности,
передняя стенка черепной коробки приближается к обонятельным
капсулам, мозговая полость постепенно надвигается на носовую, а у
форм с наиболее развитым мозгом (человек) оказывается располо­
женной над носовой полостью, в то время как у низших мозговая по­
лость находится позади носовой. Главная особенность висцерального
черепа млекопитающих заключается в появлении принципиально нового
типа сочленения нижней челюсти с черепом, а именно нижняя челюсть
присоединяется к черепу непосредственно, образуя подвижный сустав с
чешуйчатой костью черепной коробки. В этом сочленении участвуют
только дистальный отдел покровной зубной кости (вторичная нижняя
челюсть). Ее задний конец у млекопитающих изогнут вверх и заканчива­
ется сочленовным отростком. Благодаря образованию указанного
сустава квадратная кость первичной верхней челюсти теряет функцию
подвеска нижней челюсти и превращается в слуховую косточку, которая
получила название наковальни (рис. 137). Первичная нижняя челюсть в
процессе эмбрионального развития полностью выходит из состава
нижней челюсти и также преобразуется в слуховую косточку, которая
получила название молоточек. И , наконец, верхний отдел подъязычной
дуги — гомолог гиомандибулярного хряша — преобразуется в третью
слуховую косточку — стремечко. Таким образом, у млекопитающих
вместо одной образуются три слуховые косточки, которые составляют
функционально единую цепь.
Нижний отдел подъязычной дуги у млекопитающих преобразуется в
передние рожки подъязычной кости. Первая жаберная дуга дает начало
задним рожкам, а ее копула — телу подъязычной кости; 2-я и 3-я
жаберные дуги образуют щитовидный хряш, впервые в процессе
эволюции появляющийся у млекопитающихся, а 4-я и 5-я жаберные дуги
дают материал для остальных гортанных хрящей, а также, возможно,
для трахеальных.
Как видно из приведенного сравнительно-анатомического обзора,
с к е л е т ч е л о в е к а полностью гомологичен скелету млекопитаю­
щих. У человека нет ни одной кости, которая отсутствовала бы у
представителей класса (рис. 138). Вместе с тем в процессе антропогенеза
в скелете человека появляется ряд особенностей. Большинство из них
прямо или косвенно связано с прямохождением. Согласно Ф . Энгельсу,
переход к прямохождению явился главным фактором, обусловившим
перестройку человеческого тела.
Прямым следствием перехода человека к передвижению на двух
ногах являются:
1) изменения стопы, которая потеряла хватательную функцию и
превратилась в орган с чисто опорной функцией, что сопровождалось
появлением продольного свода стопы (амортизирует сотрясение
внутренних органов при ходьбе);
2) мощное развитие большого (1) пальца по сравнению с другими,
поскольку он становится главной точкой опоры, и потеря им
значительной подвижности и способности к противопоставлению;
320
Рис. 137. Преобразование первых двух ж а ­
берных (висцеральных) дуг.
а — а к у л а ; б — к о ст и с та я р ы б а; в -- ам ф и б и я; г — реп­
ти л и я; д — ториодонт; е — м лекоп итаю щ ее. I - небно
квад р атн ы й х р ящ ; 2 — ниж ний хрящ (м е к к е л е в ) ч е л ю с т­
ной дуги; 3 — к ва д р а тн а я к о с т ь (н а к о в а л ь н я м ле ко п и таю ­
щ и х ); 4 — н еб н ая ко ст ь; 5 - кр ы ловидн ы е ко сти ; 6 - со
чл е н о вн ая к о ст ь (м о ло то че к м л е к о п и т а ю щ и х ); 7 — зу б
п ая к о ст ь; 8 - у гл о в а я ко сть; 9 - ги ом андибулярны й
хр ящ (стр е м е ч ко м л е к о п и т а ю щ и х ); 10 — че ш у й ч а т а я
к о ст ь; 11 — гиоид.
Рис. 138. Эмбрион человека в конце 5-й не­
дели после оплодотворения. Видны висце­
ральные дуги.
1
гиоидная д у га; 2
че л ю ст н а я д у га ; 3— 3-я вис
ц е ральн ая (ж а б е р н а я ) д у га; 4- 4-я ви сц е р ал ьн ая ( ж а ­
б е р н а я) д у га; 5
п очка р уки ; 6 почка ноги; 7
сердечный в ы с ту п ; 8 - хвост.
3) 5-образный изгиб позвоночника, смягчающий толчки внутренних
органов при ходьбе;
4) наклон таза под углом 60° к горизонтали в связи с перемещением
центра тяжести;
5) перемещение затылочного отверстия и изменение положения
головы относительно позвоночника;
6) появление сосцевидного отростка височной кости — места
прикрепления грудино-ключично-сосцевидной мышцы, удерживающей
голову в вертикальном положении.
Косвенно с прямохождением связаны: специализация верхних
конечностей как органа труда в связи с освобождением их от функции
передвижения; особенности мозгового черепа; характерные пропорции
тела — более короткие руки и более длинные ноги.
Независимо от изменений, связанных с прямохождением, шло
формирование подбородочного выступа нижней челюсти, возникшего в
связи с членораздельной речью.
Процесс приспособления человека к прямохождению еще не
закончился, о чем свидетельствуют относительно частые случаи
возникновения грыжи при поднятии больших тяжестей, выпадения
матки.
Аномалии скелета составляют более половины всех врожденных
заболеваний и пороков развития. Причинами их служат как эндогенные,
в том числе генетические, так и экзогенные факторы. Наиболее часто
встречаются аномалии нижних конечностей.
Поскольку при нарушениях формообразовательных процессов
прежде всего выпадают более поздние этапы развития структур нередко
отсутствует закладка пальцев. В то же время встречаются полидакти­
лия или многопалость, синдактилия или срастание пальцев, брахидактилия или укорочение пальцев. Чем раньше в эмбриогенезе наступа­
ю т нарушения развития, тем более выражены отклонения от нормаль­
ного состояния органа. При действии тератогенных факторов на
ранних стадиях внутриутробного развития конечностей последние
у новорожденного имеют вид коротких боковых складок.
(^7ЛЗ.З. Органы пищеварения
Органы пищеварения беспозвоночных. Впервые пищеварительная
система начинает формироваться у кишечнополостных. В процессе
гаструляции за счет впячивания энтодермы-эбразуётся первичная кишка
(гастральная полость). Она сообщается с внешней средой только одним
отверстием — ротовым, которое одновременно служит для выбрасыу вания непереваренных остатков пиши. Заднепроходного отверстия нет.
Ьльшинство типов животного мира, как и кишечнополостные,
(носятся к цервичнордхцм, так как рот, образовавшийся в эмбриоге­
незе, функционирует всю жизнь. Иглокожие, погонофоры и хордовые
рставляют группу вторичноротых (см. рис. 127). У них ротовое
гверстие сначала образуется на одном конце зародыша, а затем на
эотивоположном конце происходит впячивание эктодермы, и образугся второе ротовое отверстие (вторичный рот). Первичный рот
эрастает, а на его месте позднее формируется анальное отверстие.
У кишечнополостных внутриклеточное пищеварение начинает за­
вещаться вн у т р и п о л о с т н ы м . Пища первоначально подвергается воз­
действию ферментов и измельчается в полости, а затем захватывается
тетками энтодермы, где переваривается в пищеварительных вакуолях.
ГУ плоских червей (трематод! пищеварительная травка также заканчива­
й ся слепо и состоит из лвух отделов — 'переднего эктодермального,
(представленного хорошо развитой~тлоткойГ и среднего (кишечник),
развивающегося из энтодермы. Пищеварение внутпиполостное и внут­
риклеточное. У -КРУТЛЫУ ч р р в е . й ' п г Т д к п д р - г р о третий ОТДеЛ ПИЩеварИтельного тракта — задний. Он образуется путем впячивания эктодермы
на заднем конце тела, соединяется с полостью средней кишки и
заканчивается на заднем конце т ед&анайьцым отверстием.] С появлени­
ем задней кишки пища продвигается тол'Иой ЯЯнймшЙравлении, что
обеспечивает более полное ее усвоение. Пищеварение становится только
внутриполостным. Передний и задний отдел кишки, имеющие
эктодермальное происхождение, выстланы кутикулой, У к^дьчятых
322
Рис. 139. Ротовая полость позво­
ночны х.
а — р ы б а : 1 - з у б ы верхней че л ю сти ;
2 — к р ы ш а ротовой полости; 3
от­
вер стие п ищ евод а; 4 — ж аб ер н ы е щели;
5 — ж а б е р н ы е д уги : 6
язы к; 7 - з у ­
б ы н иж н ей че л ю сти ; б - ам ф ибии: 1 зу б ы верхней че л ю сти ; 2
хо ан ы ; 3
гл азн ы е яб л о к и ; 4
отвер стие слуховой
тр у б ы ; 5 — отве р сти е п ищ евод а; 6 - о тве р сти е гортани; 7 - о твер стие резо­
н ато р а; 8 -- н и ж н я я ч е л ю ст ь ; 9
я зы к ;
в — р е п ти л и я: I — зу б ы верхней ч е л ю ­
с т и ; 2 — со ш н и к ; 3 — хоаны, 4 - складкн вто р и чн о го неба; 5
отве р сти е с л у ­
ховой т р у б ы ; 6
отверстие пищ евод а;
7 — отве р сти е тр а х е и ; 8
язы к. 9
зу б ы н и ж н ей че л ю сти ; г
м л е к о п и таю ­
щее: 1 — ве р х н яя гу б а ; 2
верхние р е з­
цы; 3 — преддверие полости р та ; 4 верхние м ол яр ы ; 5
твердое небо,
6 — м ягкое небо; 7 — отвер стие п и щ е ­
вода; 8 — отве р сти е гортани, 9 - над­
гор тан н ик: 10 — и иж н ие м оляр ы . 11 —
я з ы к , 12
н и ж н ие резцы.
З у б ы оеп ти л ии
1
2
3
1 П левродонтны е
А нродонтны е
Те кто д он тн ы е
2
3
червей в стенке кишки появляются мышечные элементы, обеспечиваю­
щие перистальтику, и развивается се"ГБ~ кровеносных с о с у д о в . V ч пе­
ни('тоыпг-цу пппируопит дальнейшая дифференцировка кишечной труб­
ки и одновременно появляются приспособления дщцизмельчения пищи
(челюсти) и железы, декретирующие пищеварительные ферменты.
Органы пищеварения позвоночных. В связи с полупаразитическим
или паразитическим образом жизни у к р у г л о р о т ы х челюсти
Г
. 139). Вместо ротовой полости имеется присасыва-~
тельная воронка, на дне которой находится рот. ПищеварИТёЛЫШ
ттУЬка не г1 шКп,епсш1 ЩУ1кана. Желудок~7ПТуТСШ'е|, глотка, пронизанная жаберными щелями. непоспел(?твенно переходит П И И И Ч Н Ш ,'
котОрГОГ не разделен на отделы и имеет незначительную длину.
Кишечнуф трубка прямая, не образует изгибов. Печень развивается как
о т с у т с т в у ю
12‘
т
р и с
323
Рис. 140. Эволюции зубной системы позвоночных.
а — к о н и ч е ски е з у б ы перм ских р ептилий; б — т р е х зу б ч а т ы е зу б ы вы сш и х тер и одонтов; в
тр е х б у го р ч а ­
ты е зу б ы п р им итивны х м л екоп итаю щ и х; г — ч е ты р ех б у го р чаты е зу б ы и скопаемой п о л уо б е зь ян ы ; д -- т у п о ­
б уго р ча ты е з у б ы вы сш и х о б е зьян и че л о век а.
вырост начального отдела средней кишки и сохраняет примитивное
строение ветвистой трубчатой железы. Поджелудочная железа нахо­
дится в зачаточном с о с т о я н и й Пищеварительный тракт р ы б начитается ротовой полостью,
крыша
которой
образована
непосредственно'~^~---©снованием
черепа (первичное небо).
По краю челюстей, а у некоторых на всей поверхности ротовой по­
лости расположены зубы. Зубная система у рыб гомодонтная, гг. е.
зубы одинаковы по строению и’ДрунЩшГ (см. рис. 40)." обычно они
имеют коническую форму, обращены назад и служат лишь для
удержания пищи. По своему происхождению и развитию зубы
гомологичны плакоидной чешуе хрящевых рыб. Смена зубов происходиъ—а. течение—всей жизни. В ротовой полости рыб расположен
щиаедцхиввый яз ь ^ в виде двойной складки слизистой оболочки.
Жядезы~^»теутетвук)т-.
П о сравнению с низшими хордовыми пищеварительный тракт рыб
значительно дифференцирован, особенно у хрящевых. Ротовая полость
Переходит в глотку, стенки которой гшркмэаны жаберными щелями. За
ней следует короткий^ищёвдр, зате^окелудок) степень обособленности
которого различна. В кйШёчнике вьщ елякгГтонкий отдел и толетый,1
заканчивающийся,анусом. Длина кишечника увеличивается, он образует
петли. В петле тонкой* кишки лежит ^поджелудочная железа. Печень
324
...
рис.
\4\.) Секреторный
позвоночных.
аппарат
ротовой
секреторны й эпителий п ер вичны х черепны х ж и ­
рных;
м ногослойный эпителий круглороты х:
1 — се к р етор н ы й эпителий;-в э— м ногослойный энителий р ы б : I — сл и зи с ты е (м у к о з и ы е ) к л е т ки ; 2 —
■белковые (се р о зн ы е ) клетки;-'г V м ногослойны й эп и ­
телий ам ф иб ий : 1
слизистыЪ -клетки; 2 — м но го кле­
то чн ы е вн е эп и те л и ал ън ы е сл ю н н ы е ж е л е з ы / ^ / — м н о ­
гослойный п лоский эпителий рептилий: 1 — сл и зи сты е
клетки ; 2, 3 -— ви е эп и те л и ал ьн ы е альвеолярид-трубчаты е с л и зи с ты е и б ел ко вы е сл ю и и ы е ж е л е з ы Я е >— м но­
гослойный п лоский эпителий м ле ко п и таю щ и х : 2 —
б ел ко вы е сл ю н н ы е ж е л е зы ; 3 — сл и зи с ты е слю нные
ж е л е зы ; 4 — см е ш ан н ы е сл ю н н ы е ж е л е з ы ; 5
в н еэпи тел и ал ъны е м алы е сл ю н н ы е ж е л е зы .
развита хорошо, имеется желчный пузырь. У костистых рыб кишечник
м ру-р
п и Т К ^ р р Р Н ^11 ^ П В Я н .
я м (Ь и б и и1потовая полостьн^ «т. е т я д ^ ^ ф глотки. Зубная
^я. П оявляются Сс^ ^ньШ-^кедез^!) рис: 141). Их
секрет служит для смачивания пиши, нб оказывая нл *т;се химического
воздействия. В рото-глоточную полость открываю тся хаацы.^в,стахиевы трубы и гортанная щель. Полость" продолжается в, пищевод,
переходящий в желудок. Собственно кишечник имеет большую длину по
сравнению с рыбами и отчетливо подразделяется на .тонкий отдел и
толстый, открывающий в клоаку. Печень имеет больший~объём,
разветвленная поджелудочная железа леЖй'1 н ’петлк" '1ШКУ11!' "К!й:
щечника.
4— ---- -----------Ротовая полость р е п т и л и й более обособлена от глотки, у
325
большинства гомодонтная зубная система. Однако у некоторых, в
основном вымерших форм, обнаруживается начальная дифференци­
ровка зубов. Я зы к имеет иное происхождение, чем язык анамний. Он
развивается из зачатка, лежащего в области 2-й и 3-й жаберных дуг.
Форма и степень подвижности языка у разных видов рептилий
различна. Ротовые железы развиты лучше. Среди них выделяют
ябдъязычные. зубные и губньге^ /н д ц в и т х змрй задняя пара зубных
желёз лпёёппачуетёя в^ялбвиТУкТ желеа^чИз яда змей выделен ряд
биологически активных веществ, например, фактор роста нервов
(см. 6.5.2). Названный фактор, а также другие вещества, которые могут
бы ть отнесены по физиологическому действию к гормонам, обнаруже­
ны в гомологах яппнити»: ж г 1Р1 — 1- ч, чч!,|у ;&&]5 зах млекопитающих
У рептилий появляются (зачатки вторичного неба?) Оно образуется
боковыми складками верхнё'Н ЧРЛ1бСТ1|,‘ лЬтг»рыет1»*АЛ]шт до середины и
делят ротовую полость на верхний отдел — дыхательный и нижний —
вторичную ротовую полость.
Строение глотки, пищевода и желудка не имеет существенных отли­
чий по сравнению с амфибиями. Собственно кишечник подразделяется
на тонкую И толстую урчцги На ц ^яны.тр тонкого и толстого отдела
появляется небольшойодедаа^ццоост^Цлина кишечника по сравнению с
амфибиями увеличивается^&дняя кишка оканчивается клоакой..
Пищеварительный тракт
м л екопитающих
достигает
наибольшей степени дифференцировки. Он начинается препротово^'
, псшость в в д ^ зджддверьем рТ)а, расположенным межд^ г у б а ^ ^ щ к а м !?)
и 1КЯ1остями^М ясистые губы, свойственные только млекопитающим!
служат ДЛя захвата пищи. Ротовая полость ограничена стерх«Стгвердь1 м ■
небом. Кзади твердое небо продолжается ^ й я г ш ^ - В ^ Э — двойную
складку слизистой, отделяющую ротовую полость от глотки. На
твердом небе имеются поперечные валики, которые способствуют
перетиранию пищи. У человека при рождении также имеются такие
ы м.н.копитаюпщ Д неодинаковы по строению и функции —
гетородонтная зубная сист?МЯ. Различают резцы, клыки, малые
коренные (ложнокоренные) и большие коренные (истинные коренные).
Соотношение зубов различного типа составляет зубную формулу*
Резцы — передние зубы — имеют долотовидную форму и служат для
захвата и разрезания пищи. Следующие — клыки — сохранили
коническую форму, но имеют большие размеры и используются для
разрывания пищи. Задние зубы приобрели сложную бугристую или
складчатую поверхность и служат для перетирания пищи. Они
подразделяются на малые жевательные — (премоляры) и большие
жевательные (моляры). Следует отметить, что сложная структура
жевательных зубов млекопитающих является результатом преобразо­
вания простых конических зубов. Общее количество зубов у млекопита- ющих уменьшается. У низших млекопитающих число зубов составляет
44. У более высокоорганизованных животных происходит дальнейшее
сокращение числа зубов. Так, у многих приматов и человека количество
зубов равно 32
326
2 12 3
23
21
’
Располагаиэтся только на челюстях в
ячейках, основание зубов суживается, образуя корень. Смена зубов
только одна — молочные зубы сменяются постоянными. Эволюция
зубной системы человека пошла по пути редукции ее переднего отдела,
особенно клыков. Они сильно уменьшились в размере, не выступают из
зубного ряда и не перекрываются. Исчезли диастемы на верхних и
нижних челюстях, зубы стали в плотный ряд, зубная дуга получила ок­
руглую (параболическую) форму. Премоляры и моляры уменьшились
в размерах, но усложнилось их строение. В связи с перераспределением
основной нагрузки при жевании на первые моляры наблюдается
постепенное уменьшение размеров моляров от первого к третьему.
В связи с дифференцировкой и усложнением зубов у млекопитающих
увеличилась продолжительность их функционирования.
Во время жевания нижняя челюсть человека совершает сложные
вращательные движения по отношению к верхней, что стало
возможным только при неперекрываюшихся редуцированных клыках и
комплементарно расположенных буграх жевательных зубов на верхней
и нижней челюстях.
Ротовые железы у млекопитающих достигают наидысшего
развития. Имеются как мелкие ^елйУйст!
З Р ы ,.аак,
так ааю
иСвдмьге
ары
аь
слюнные
пцтЛТУычная, заднеяз
челюстная и околоушн;
в- ЦО'ГОВиИ Ш ЛТЭ Т ЕГТГОШ1ИЮ11Я Щ)У1
скопления лимфатической ткани Х*миндали ньй В глотку открываются
/^Носоглоточные ходы, евстахиевы трубы, гортанная~ТЦсжч Желудок
млекопитающих хорошо обособлен от других отдШВВ“Н“у'^Йзных видов
имеет свои специфические отличия. Общим служит разнообразие желез
слизистой оболочки, участвующих в образованиц^Желудочного сбЕр.
Собственно кишечник дифференцируется на отделы ^^енадцатиперррная? 'пуп гал1
, т л и ам.тЧТСТШяДГппямая кншкйЭ^лепаякИШКИ ИМРС1 нилнепарного сдецрга. аыроскц^-рт;110 Т1ОЖённого на границе толстрй
"*”1Гтонкой кишки, достигающего у некоторых животных (травоядные,
грызуны) больших размеров — от 10 до 27% всей длины кишечника.
У многих видов на слепой кишке имеется вырост
червеобразный
М'ростСТСтв стенке которого содержится большое количество"^лйм^о^ *
нищ 1Г1 Ппнм Длина кишечника по сравнению с рептилиями резко
увеличена.
ЗоХнИГХ Специальные органы дыхания отсутствую т, тазообмен происходит через покровы —Сдиффузное ды'ха'йиеч
(кишечнополостные, плоские, круглые черви). У кольчаз ы х' ЧериеЙ~гож^>
богато снабжена кровеносными капиллярами, в которые поступает
кислород. Диффузное дыхание встречается также у м ел ки ф Кёр ЯС ^ Н ^ 1
имеющих тонкий хитин и относительно большую "м тщ ж ю Ч\ в
тела. Энергетический обмен таких животных отличается малой интен­
сивностью. У многих беспозвоночных появляются приспособления,
Увеличивающие дыхательную поверхность в виде местных специализи­
рованных органов дыхания. У водных форм органы дыхания пред­
ставлены жабрами, у наземных — легкими и трахеями. Впервые жабры
327
III
Рис. 142. Эволюция легких у позвоночных.
1 — гл о тк а и п л а ват ел ь н ы й п у зы р ь (л е г к и е ) кистепер ы х рыб; I I — гло тка и легкие ам ф ибий ; 111 — легкие
а — х в о с т а т а я ам ф иб ия; б — б ес х во ста я ам ф и б и я; в — р ептил и я; г, д - м лекоп итаю щ ее; 1 — гл о тка;
2
н еп ар н ая кам ер а, со ед и н яю щ ая п ла вател ьн ы й п узы р ь (л е г к и е ) с глоткой; 3
меш ки п ла вател ьн о го
п у з ы р я ; 4 — гортаи и о-тр ахейи ая ка м е р а ; 5 — л его чн ы е м еш ки ; 6 — вн утр и ле гочн ы е перегородки; 7
т р а х е я; 8 — бронх; 9 — ве тви бронхов; 10
появляются у ) м ногощетинковых кольчецов!
ёцов| и представляют собой
вёнбсньп
разрастания эпителия, пронизанные кровеносными
сосудами. Многие
виды одновременно сохраняют диффузное дыхание. ^_У_наземных
&^паукообразныёУ появляются листовидные легкие, у (насекомых X
✓'Ърахеи.
* < р а л *4
Функцию органов дыхания у
(ланцетник)
,1 Г принимает на себя передняя частьГ’йппечцой трубки. В стенках глотки
имеется 100— 150 пар отверстий, или Жаберных щелей. Органами
дыхания служат межжаберные перегородки, вТ о то р ы х проходят
кровеносные сосуды — жаберные артерии. Вода, проходя через
жаберные щели, омывает названные перегородки и кислород диффун­
дирует через стенки артерий. Поскольку жаберные артерии ланцетника
не разветвляются на капилляры, общая поверхность, через которую
поступает кислород, невелика, окислительные процессы идут на низком
уровне. Соответственна этому ланцетник ведет малоподвижный,
пассивный образ жизни.
Позвоночные характеризуются переходом к активному образу"
жизни. Появление таких форм в процессе эволюции связано с возникно­
вением прогрессивных изменений в органах дыхания, обеспечивающих
более высокий уровень окислительных процессов в пределах каждого
Сукласса и всей группы в целом (рис. 142 ).)
Л/1
Прогрессивные изменения органов дыхания У ^ ^ б . заключаются в
л&явлении на^ е ж ж аберщлх пепегоролках многочисленных эпители­
альных выростов —7\жаберных лепестков. Жаберные лепестки, распо­
ложенные на одной перегородке, составляют жабру. Жаберные артерии
328
рыб в отличие от ланцетника образуют в жаберных лепестках густую
сеть капилляров. Дыхательная поверхность за счет лепестков резко
увеличивается, поэтому число жаберных перегородок у рыб сокрашается по четырех. Изменения дыхательной системы сочетаются у рыб с
прогрессивными изменениями органов кровообращения, о чем будет
сказано ниже.
ц\ Ж аберны^ ш**™ У рыб возникают путем выпячивания стенки
гло*ГкиГ~Сначала образуются парные слбЛые-"выросты — жаберные
мешки, растущие по направлению к периферии. Навстречу каждому из
них образуется впячивание кожных покровов. Выросты глотки и
выросты кожи растут друг другу навстречу. Н а месте их соединения
ткань прорывается и образуется шель, соединяющая полость глотки с
наружной средой, т. е. жаберная щель. Позднее на перегородках
образуются жаберные лепестки. У большинства рыб закладываются
пять пар жаберных мешков. У кистеперых рыб появляются наряду с
жабрами органы для использования атмосферного кислорода. Т а к т
дополнительным органом дыхания у них служит плавательный пузырь*
представляющий собой парный мешковиднвгй вырост брюшной
стороны глотки, стенки которого богаты кровеносными сосудами.
Пузырь соединен с глоткой короткой широкой камерой. Кровоснабже­
ние происходит за счет 4-й жаберной артерии, окисленная кровь
поступает прямо в сердце.
Земноводные
обладают способностью, хотя и о г р а н и ч ь
ченной" жить в наземных условиях, что обусловило дальнейшее^
развитие органов атмосферного дыхания в виде легких и кожи. Легкие
земноводных гомологичны плавательному пузырю кистеперых рыб.
Они представляют собой два мешка, соединенных с глоткой небольшой
гортанно-трахейной камерой. Так же, как плавательный пузырь
кистеперых рыб, они снабжаются кровью от 4-й жаберной артерии.
Легкие амфибий весьма примитивны. Как правило, стенки легочных
мешков гладкие, с небольшими перегородками, дыхательная площадь
мала. Поверхность легких относится к поверхности тела, как 2 к 3.
Количество кислорода, поступающего через легкие, составляет при­
мерно лишь 30—40% от его общего количества. Воздухоносные пути
слабо дифференцированы. В связи с недостаточным развитием легких
основным
■•ыхачия'щужцт р ж а в которой имеется большое
количество мелких кровеносных сосудов-капилляров.
Урептилий
с переходом к жизни на суше происходит'У'»
^дальнейшее развитие дыхательной системы. Кож а рептилий выключа- \ &
ет.ся из дыхания, поскольку толстая роговая чешуя, защищающая \
рептилий от высыхания, препятствует газообмену, ц_легкие становятся
основным о п г а н о м л ы х а н и я . Дыхательная поверхность легочных
мешков резко увеличивается благодаря появлению на их стенках
большого количества разветвленных перегородок, в которых проходят
кровеносные сосуды.
Одновременно у рептилий наблюдаются прогрессивные изменения в
воздухоносных путях. Е трахее Фо р м и р у ю т с я хрящевые
разделяясь, она дает два бронху Начинается формирование внутрилегочных бронхов. Отдельные крупные перегородки вдаются глубоко в
329
полость легкого, оставляя свободным лишь узкий центральный вход.
Дистальные края перегородок покрыты мерцательным эпителием, а в
наиболее крупных из них появляются хрящи. В результате образуются
стенки внутрилегочных бронхов.
М л е к о п и т а ю щ и е обладают легкими наиболее сложного
строения. Характерен древовидный тип разветвления бронхов. Основ­
ной бронх делится на довольно большое количество вторичных
бронхов, те в свою очередь распадаются на еще более мелкие бронхи
3-го порядка, а последние дают многочисленные мелкие бронхи 4-го
порядка и т. д., и, наконец, идут тонкостенные трубочки — бронхио­
лы. Н а концах бронхиол находятся мелкие пузырьки, выстланные
эпителием, или альвеолы. Стенки каждой альвеолы оплетены густой
сетью капилляров, где и происходит газообмен. Количество альвеол
достигает огромного числа, благодаря чему дыхательная поверхность
резко возрастает. У ряда млекопитающих поверхность легких в
50— 100 раз больше поверхности тела. У человека площадь легких
составляет 90 м 2 и превышает поверхность тела во много раз,
ветвления бронхов составляют 23 порядка.
Таким образом, основное направление эволюции дыхательной
системы заключается в увеличении дыхательной поверхности, обо­
соблении воздухоносных путей.
ГГГзТз. Органы кровообращ ения'"^
Необходимым условием жизнедеятельности организма является
непрерывное поступление в ткани кислорода и субстратов, а также
удалении продуктов распада.
Органы кровообращения беспозвоночных. У кишечнополостных
(гидроидные), тело которых состоит всего из двух слоев, пищевые
вещества, кислород и экскреты передаются путем диффузии от одного’
слоя к другому. ^м едуз в связи с мощным развитием мезоглеи фун­
кцию распределения, хотя и несовершенную, берут на себя каналы
гастроваскулярной системы. У плоских червей паренхима, заполняю­
щая промежуток между органами, не допускает перемещения веществ
на большие расстояния. Э то компенсируется появлением сильно
разветвленной пищеварительной и выделительной систем. Однако
такой механизм не может обеспечить единства внутренней среды
организма. Удкруглых червей с появлением первичной полости тела
функцию пере'ТСШЮТгтга—прщг^ктов обмена по организму начинает
выполнять полостная жидкость, которая, перемещаясь при движениях
червя, омывает все части тела и становится посредником между ними.
Однако правильной циркуляции и определенных путей передвижения
продуктов обмена здесь еще нет.
(-ДЯым ггтерптеинмм аппаратом распгслепницд /^пу^-ит
и.у~няп
система, появляющаяся впервые у аннелид. Очевидно, в процессе
эволюции в тканях организма постепенно определялись постоянные
пути циркуляции. Предполагают, что это произошло путем образова­
ния полостей в промежуточном веществе мезенхимы, принявших форму
правильных каналов. Попадавшие в полость каналов клетки мезенхимы
330
г
дали начало клеточным элементам крови. С появлением сократи­
тельных элементов в стенках сосудов возникла правильная циркуляция
крови, что обеспечило быстрое химическое взаимодействие разных
частей организма между собой и появление общей единой среды
организма. Кровеносная система аннелид замкнутая, т. е, кплвь течет—
I I ц у т р Т 7 7 и .'ц Ш - Ш - И м е е т с я С П И Н Н О Й И б р ю ш н о й г г ч - у п ч к
у п т п р т.»»
соединяются на концах тела и. кроме того, в каждом сегменте. Сеоппа
нет, его роль_дылолняют участки спинного и циркулярных сосудов,
сбЗё Р Ж ф л и е гп к п а ти те л ь н ы е элементы»
Кровеносная система л а н ц е т н и к а замкнутая, построена по
тому же принципу, что и система кольчатых червей (рис. 143, а, см. на
цвет. вкл.). Она представлена брюшным и спинным сосудом,
соединенным анастомозами в стенках кишки и теЛЦ, и ОДНИМ кругом
Грг.г.г»^Яр^л.^цца
Рпль
сепппя
выполняет
пульсирующий
совуД — брюшная аорта, п о брюшной аорте венозная кровь от органов
проходит в приносящие жаберные артерии (150 пар), где окисляется. По
выносящим жаберным артериям окисленная кровь поступает в парные
корни спинной аорты, которые на уровне заднего конца глотки
сливаются в непарный сосуд — спинную аорту. Последняя идет вдоль
тела к его заднему концу, образуя многочисленные артерии, направляю­
щиеся к органам, где кровь, отдавая кислород, превращается в
венозную. Венозная кровь от передней части тела поступает в парные
^ передние кардинадьные вены, а от задней части тела — в задние
кардинальные. Передняя и задняя кардинальная вены каждой стороны
да уровне заднего конца глотки соединяются цш роток (кювьеров),
впадающий^_бщошную аорту. О т внутренних органов, в основном от
^ишечника, венозная_кровыдостУпает в подкишечную вену, которая
входит в печеньпод названием воротной вены печени и там разветвля­
ется на густую сеть капилляров, образуя воротную систему печени.
Затем капилляры собираются вновь в венозный сосуд — деченочную
вену, по которой кровь поступает в брюшную аорту. Воротная система
печени имеет важное значение для организма. Кровь, поступающая из
кишечника, содержит наряду с питательными веществами токсические
продукты распада, которые нейтрализуются клетками печени, т. е. пе­
чень выполняет роль барьера и препятствует интоксикации организма.
Кровеносная система позвоночных. Имеется большое сходство в
строении кровеносной сйстемы ’ позвоночных и низших хордовых
(рис. 143, б— е, см. на цвет. вкл.). Вместе с тем в пределах этой группы
животных можно проследить ряд прогрессивных изменений.
Такие изменения в кровеносной системе р ы б направлены на
обеспечение более интенсивного метаболизма в связи с активным
образом жизни. Появляется сердце, обеспечивающее большую скорость
передвижения крови по сосудам. Сердце состоит из двух ка­
мер — предсердия и желудочка. О т желудочка у низших рыб начинается
мускулистая трубка — артериальный конус ь стенки которого содержат
поперечнополосатую мускулатуру и способны к пульсации. Внутри
конуса находится ряд клапанов. Конус переходит в брюшную аорту.
У высших рыб артериальный конус рудиментарен. Брюшная аорта в
■ачальном отделе образует эластичное вздутие — луковицу аорты.
А
331
В сердце рыбы содержится только, вендзная кровь. Она поступает от
(^ а н о в ШГвёнам в'предсердие, оттуда в желудочек и затем идет по
брюшной аорте в жаберные артерии, распадаюшиеся на капилляры,
где кровь окисляется.
А ^
■„ "
У з е м н о в о д н ы х в процессе приспособления к наземным
условиям"" Сущеед в 1 ШНИя-'исчезает жаберное дыхание и появляется
вторрй (дегочный) круг кровообращения. Одновременно в строении
сердца и сбСудов появляк)тся изменения, направленные на разделение
артериальной и венозной крови.
Сердце амфибий трехкамерное, состоит из двух предсердий и
желудочка. Оба предсердия открываются в желудочек общим отверсти­
ем. Левое предсердие содержит артериальную кровь, поступающую из
легких, правое — венозную, поступающую от органов большого круга
кровообращения. Изливающаяся из предсердий в желудочек артери­
альная и венозная кровь не успевают смешиваться полностью, поэтому
желудочек содержит 3 вида к^ови: срева — яртррияпьнуто в середи­
не — смешанную, справа — венозную. Из' желудочка выходит только
олин пощ ц —\ артериальный конус, от которого отходят--Л. ЦЯры
а^дот7^^жно-легочные"артерии (ближние'~к сердцу),^уги аорты и
-ХсонЕГЫе артерии. Каждая пара сосудов выносит из сердца определенный
вид крови: кожно-легочные — венозную ,, дуги аорты — смешанную,
сонные — артериальную. Механизм распределения крови по сосудам
следующий: артериальный конус отходит от правой стЬроны желу­
дочка, поэтому при сокращении последнего в ,него сначала поступает
венозная кровь, которая направляется в ближайщую пару сосу­
дов — кожно-легочные. Затем выходит порция смешанной крови. В это
время спиральный клапан, находящийся в артериальном конусе,
закрывает отверстия легочных сосудов и смешанная кровь идет в дуги
аорты. Последней из желудочка выходит порция артериальной крови,
которая не может пройти в Легочные артерии (спиральный клапан) и
дуги аорты (самая крупная по объему порция смешанной крови создает
большое давление в сосудах) и направляется в сонные артерии. Дуги
аорты огибают сердце: одна — слева, другая — справа, затем соединя­
ются за сердцем в непарный сосуд 2^ спинную аорту, несущую смешанную кровь. Она идет по спинной сто$онё~тела, отдавая артерии к
внутренним органам, затем делится на 2 подвздошные артерии, идущие
к задним конечностям.
Основные венозные стволы у а м ф и б и й также меняются по
сравнению с рыбами — вместо кардинальных вен появляю тся полые
* §ены. Венозная кровь от задней половины тела собирается1® две парЯые
подвздошные вены, которые соединяются в непарную1,заднюю полую
вену, впадающую в правоепред сердце. Кровь от кишечника по
воротной вене идет в печень, образует там воротную систему печени и
выходит по печеночной вене в заднюю полую вену. Венозная кровь от
передней чаети-ч=<ша. выносится по двум передним полым венам, в
которые впадает артериальная кровь из кожных вен. Передние полые
цены-вцадают в правое предсердие. —У р е п т и л и й происходит дальнейшее, более полное, разделениеНртериальной и венозной крови. Изменения касаются как строения
332
сердца, так и дифференцировки сосудов. Сердце рептилий трехкамерное, имеет два предсердия и желудочек, но отличается от сердца
амфибий. Предсердия обособлены полностью, каждое открывается в
желудёчек Собственным отверстием. В желудочке появляется неполная
перегородкаТвыпячивание дна), разделяющая его на левую и правую
половины. Поэтому у рептилий над перегородкой образуется не­
большая порция смешанной крови. В момент сокращения, желудочка
перегородка полностью разделяет желудочек на две половины — пра­
вую и левую. У некоторых рептилий, например у крокодилов,
перегородка полная. Артериальный ствол подразделен на три сосуда,
каждый из которых отходит от желудочка самостоятельно. Из.левой
половины желудочка отход^А иряпяя лугя яорты несущая_ артериалуцпо-кревь. Э то т сосуд огибает сердце с правой стороны и поэтому
получил название правой дуги аорты. О т него отходят сосуцы-я-голове и
передним конечностям. И з середины желудочка, над "перегородкой,
берёт начало левая дуга аорты. огибающая сердце слева и несущая сме­
шанную кровь. Ид ц и щ < й я е и и м *т*"1пу во н и в м е н ю ш че— я
артерия, несущая венозную—крепь. Правая и левая дуги аорты
соедйШтТТся'позади сердца и образуют спинную аорту. Кровь в
спинной аорте рептилий смешанная, но отличается более высоким
содержанием кислорода по сравнению с амфибиями, так как одна дуга
несет смешанную кровь, а другая — артериальную.
Большая насыщенность крови кислородом повышает уровень
обменных процессов и Обеспечивает более активный образ жизни,
который характеризует высших позвоночных. Прогрессивные измене­
ния кровеносной системы м л е к о п и т а ю ш и х заключаются в по­
явлении полной "перегородки в желудочке, благодаря чему сердце
становится четырехкамернцм. Э т о приводит к полному разделению
венозной и артериальной крови. Сосуд, гомологичный левой дуге аорты
рептилий, выносящий смешанную кровь, у млекопитающих редуциру­
ется. Теперь из желудочка выходят только два сосуда: из правого — ле­
гочная артерия, из левого — левая дуга аорты, которая огибает сердце с
левой стороны и переходит в спинную аорту, несущую артериальную
кровь. Все органы млекопитающих снабжаются артериальной кровью.
К ак видно, эволюция кповеногнойдшстемы идет по путиГувеличёния
количества камер” сердца, 9дифференцировки сосудов, отходящих от
сердца ,А(рвышения содержания кислорода в крови.
Ряд крупных артериальных сосудов человека развивается Эю
закладок жаберных, артерий. В эмбриональном периоде у всех классов
позвоночных в области глотки кровеносные сосуды закладываются по
одной и той же схеме. О т сердца вперед отходит непарный со­
суд — брюшная аорта, а от нее к жаберным перегородкам отходят
6 пар (у рыб 6—7 пар) крупных сосудов, охватывающих глотку и
соединяющихся на спинной стороне в два корня спинной аорты. Э ти
сосуды получили название а р т е р и а л ь н ы х , или ж а б е р н ы х
дуг.
У зародышей рыб первые две пары жаберных сосудов быстро исче­
зают, так как две первые жаберные перегородки (висцеральные дуги)
включаются в состав лицевого черепа. Оставшиеся 4— 5 пар артери­
I
1
333
альных сосудов функционируют в качестве жаберных артерий.
У наземных позвоночных, так же как и у рыб, первые две пары жабер­
ных дуг рано исчезают, а оставшиеся 4 пары в связи с переходом к ат­
мосферному дыханию подвергаются характерным преобразованиям.
Третья пара жаберных дуг у всех наземных теряет связь со следующей
парой артерий и вместе с участком корня спинной аорты превращается в
сонные артерии и несет кровь вперед в головной отдел (рис. 144, см. на
цвет. вкл.). Сосуды четвертой пары достигают наибольшего развития и
вместе с участком корня спинной аорты во взрослом состоянии
становятся основными сосудами большого круга кровообраще­
ния — дугами аорты. У амфибий и рептилий оба сосуда четвертой
пары развиты одинаково (симметрично). У млекопитающих происхо­
дит асимметричное развитие: развивается левая половина четвертой
пары сосудов (левая дуга), а правая исчезает. П ятая пара жаберных дуг
у наземных позвоночных (кроме хвостатых амфибий) редуцируется,
шестая — превращается в легочные артерии и теряет связь с дугами
аорты и корнями аорты. Таким образом, сонные артерии наземных
позвоночных гомологичны третьей паре жаберных артерий, дуги
аорты — четвертой, легочные — шестой паре жаберных артерий рыб.
Проток, соединяющий легочные артерии с дугами аорты (часть
четвертой жаберной артерии), называется артериальным протоком
(ФисШз айепозиз). Он сохраняется во взрослом с о с т о я н и и у хвостатых
амфибий и некоторых рептилий (черепахи).
Развитие сердца. В процессе развития сердца человека можно
проследить повторение ряда этапов филогенетических преобразований.
У низших позвоночных закладка сердца происходит непосредственно
под глоткой. Листки брюшной брыжейки на этом участке расходятся и
между ними и кишкой (энтодермой) появляется скопление мезодермальных клеток. Сначала они образуют пластинку, затем — труб­
чатую структуру, стенки которой состоят из одного слоя клеток. Э то
будущий эндокард. Стенки целома, прилежащие к трубке, утолщаются
за счет превращения мезенхимных элементов в мышечные и дают
начало миокарду. Часть стенки целомической полости обособляется и
образует околосердечную сумку, а ее стенки преобразуются в перикард.
У высших позвоночных и человека сердце развивается из парных
закладок, первоначально далеко отстоящих друг от друга. Сначала
обособляются правая и левая перикардиальные полости, в каждой из
них образуется эндокардиальная трубка. Энтодерма в это время
незамкнута, она распластана на поверхности желтка. По мере того как
энтодерма смыкается, обе закладки сердца сближаются, перемещаясь
под кишку, и сходятся, образуя единую трубку, расположенную
посередине. Продолжение трубки вперед и назад дает начало крупным
сосудам (рис. 145). Сначала сердце позвоночных имеет вид прямой
трубки. Н а переднем и заднем концах трубка фиксирована связанными с
ней сосудами, а ее средняя часть свободно лежит в перикардиальной по­
лости. Затем трубка начинает быстро расти, причем различные ее
участки растут с неодинаковой быстротой. В результате образуются
изгибы и сердце принимает 5-образную форму. После этого задняя
часть трубки смещается на спинную сторону и вперед, образуя
334
Рис. 145. Развитие сердца у эмбриона человека. Изгибание сердечной трубки и обра­
зование отделов в сердце (вид спереди).
Д л и н а эм бр и он а: а
р и и ).
2.08 м м; б
.4 м м; в -
5,2 мм; г
6 м м; д — 8,8 мм ( I — V I — ж аб ер н ы е а р т е ­
предсердие. Стенки этой части трубки остаются тонкими. Передняя
часть не подвергается смешению, стенки ее утолщаются и она дает
начало желудочку. К предсердию примыкает тонкостенный венозный
синус. У хрящ&вых рыб передняя часть желудочка преобразуется в
мускульную трубу — артериальный конус. У костных рыб он руди­
ментарен. Внутри конуса закладываются многочисленные клапаны,
играющие важную роль в процессе кровообращения.
У рыб одно предсердие. У амфибий оно разделяется перегородкой
на две половины, но оба предсердия открываются в желудочек общим
отверстием. У рептилий каждое предсердие имеет самостоятельное
отверстие. Полость желудочка у рыб остается общей, у амфибий на
внутренней поверхности появляются мышечные выросты (трабекулы),
образующие мелкие пристеночные камеры. У рептилий образуется
неполная перегородка за счет выроста, растущего снизу вверх. У птиц,
млекопитающих и человека желудочек разделяется полностью на две
половины. У человека, обособляющееся в процессе эмбриогенеза
предсердие, отделено от желудочка перехватом, внутри которого
находится атриовентрикулярный канал, сообщающий полости предсер­
335
дия и желудочка. Затем внутри предсердия от его краниальной части
вниз начинает расти перегородка, разделяющая его полость на две
половины.
Одновременно в стенках атриовентрикулярного канала с дорсальной
и вентральной сторон образуются утолщения (атриовентрикулярные
подушки), начинающие расти навстречу друг другу. Соединившись, они
разделяют общее атриовентрикулярное отверстие на два — правое и
левое. Каждое из них открывается в соответствующую камеру
предсердия. Позднее в каждом атриовентрикулярном отверстии
формируются клапаны. Меж желудочковая перегородка представляет
собой сложную структуру, формирующуюся за счет разных источников.
Верхняя часть перегородки возникает за счет клеток атриовентрику­
лярных подушек, дающих вырост вниз в полость желудочка, нижняя за
счет гребнеобразного выпячивания дна желудочка. В образовании
средней части принимает участие перегородка общего артериального
ствола. Общий артериальный ствол отходит от желудочка, затем в нем
формируется перегородка, подразделяющая его на два сосуда — аорту
и легочный ствол. Перегородка возникает в виде растущих вниз двух
продольных валиков на диаметрально противоположных сторонах
трубки. Посередине просвета валики соединяются и делят ствол на два
сосуда. Образовавшаяся перегородка спиралеобразно изгибается, ее
проксимальный отдел повернут по отношению к дистальному на 180°.
Часть этой перегородки принимает участие в образовании средней части
межжелудочковой перегородки. В месте соединения трех закладок
перегородки образуется перепончатая часть. Отклонения в развитии
межжелудочковой перегородки являются наиболее частой причиной
различных врожденных пороков сердца.
Наиболее часто встречающиеся аномалии развития сердца у
человека: дефекты межпредсердной и межжелудочковой перегородок,
транспозиция сосудов.
Дефекты межпредсердной перегородки чаще встречаются в области
овального отверстия или внизу, где не произошло ее срастания с
атриовентрикулярным кольцом. Дефекты межжелудочковой перего­
родки чаще отмечаются в области срастания ее трех зачатков — пере­
пончатой части. Обычно они сочетаются с нарушением процесса
разделения артериального ствола, хотя могут возникать и изолиро­
ванно.
Транспозиция сосудов заключается в неправильном отхождении
аорты (от правого желудочка) и легочного ствола (от левого
желудочка). Причиной транспозиции служит неправильное формирова­
ние артериального конуса и дефекты перегородки, разделяющей
артериальный ствол и участвующей в образовании межжелудочковой
перегородки. Если складка артериального ствола разделяет его
неравномерно, образуется сужение аортального или легочного сосуда.
Иногда ствол остается неразделенным.
Из сосудистых нарушений наибольшее значение имеют отклонения в
развитии аорты и крупных сосудов, являющихся производными
жаберных'дуг. В процессе эмбрионального развития у человека в норме
только некоторые фрагменты жаберных дуг и корней аорты идут на
336
образование магистральных сосудов и их ветвей, а остальные части
подвергаются редукции. Однако в ряде случаев редукции соответствую­
щих участков не происходит, и тогда формируется та или иная анома­
лия развития. С другой стороны, возможно исчезновение обычно
сохраняющихся отделов, что также будет служить причиной врожден­
ного дефекта.
Если у эмбрионов человека не происходит редукции правой артерии
четвертой жаберной дуги и корня аорты справа, то вместо одной
(левой) дуги аорты развиваются две дуги аорты. Одна из них, выходя из
сердца, охватывает пищевод и трахею справа, а другая — слева, после
чего они соединяются в непарную спинную аорту. В результате трахея и
пищевод оказываются в кольце (отсюда этот порок развития получил
название «аортальное кольцо»), которое с возрастом постепенно
сжимается. Э то приводит к нарушению глотания и требует хирургиче­
ского вмешательства.
Довольно часто (от 6 до 22% случаев по секционному материалу
различных авторов) у взрослых Встречается открытый артериальный
(боталлов) проток. Артериальный проток функционирует во время
внутриутробной жизни, направляя кровь из легких (спавшихся) в аорту.
После рождения в норме он зарастает в пределах 10 нед. Если проток
сохраняется во взрослом состоянии, у больного повышается давление в
малом кругу, развивается застой крови в легких, что приводит к
сердечной недостаточности.
Серьезной аномалией является развитие в качестве главного сосуда
правой артерии четвертой жаберной дуги и правого корня спинной аор­
ты вместо левых. В этом случае дуга аорты берет начало из левого
желудочка, но поворачивает вправо. При этом возможно нарушение
функции соседних органов.
/
(^ Х Ь .Ь . Нервная систем^
Нервная система беспозвоночных. Нервные клетки кишечнополо­
стных соединены ,между собой в единую сеть ^(«д~,и д><Ь,у з н аЭПГ
' н ё р в н Т я " с и с т е м а). 3 процессе последующей эволкщ Ж >гф б !исходит конДШТрЙция нервных клеток и образование нервных центров,
форма которых может бы ть разнообразной. У плоских червей, в связи с
дифференцировкой переднего и заднего конца тела, происходит
концентрация нервных клеток и образование нервного центра в виде
СйЩ н<5ш^
к^ ^ ^ ” О т них
оТХбджГвдоль тела нервные стволы, состоящие из нервных волокон и
клеток, у тп и ц атц ч червей нервная система состоит из (ЙЩыеиэ
1 П Л О М К Ч ^ Ы Ц Н Н б Й ^ р Е ^ Щ 'ц сщ чк гО тредставленной двумя
нервными стволам и,-идутшштГШПзрюшной стороне и образующими в
каждом сегменте по нервному узлу. Каждая пара узлов соединена
между собой поперечными связями <д'естнщШИй~ТйТ нервной системы)^
У многих аннелид продольные стволы брюш ной нервной цепочки
сближаются между собой, образуя непарную структуру. У,.чя««истоногн» р п м я я гцгтгчу[{1 в принципе не отличается от таковой кольчатых
червей. Характерно «влияние у зло».. вервной цепочки м^жду собой,’;)
337
В переднем отделе слияние ган­
глиев приводит к значительному
укрупнению и усложнению стро­
ения голового ганглия (голов­
ного мозга).
т прин­
ципиально новый 1
негпной
системы в виде
позвоночных() нец прогрессивно развивается и
, , « I превращается в головной мозг.
’ Г
■(
Головной мозг позвоночных.
, ^ !' Гомология нервной системы по' 1 звоночных нервной системы низших хордовых проявляется уже
при заклалке отделов головного
Рис. 146. Головной мозг позвоночных.
м ызгав эмбриональна
эх
иегиоде.
а — ст а д и я трех пузы р ей ( с глазным и п узы
У
всех
классов
позвоночных
за­
р я м и ); б — ст а д и я п яти отделов (с глазным и
б о к а л а м и ); 1 — передний м озг; 2 — п р о м е ж у то ч­
кладка нервной системы после­
ный м озг; 3 — средний м озг; 4
- м о зж е чо к .
довательно проходит стадии
5 — продолговаты й м озг; 6 — глазн ой б окал;
7 — гл азн ой п узы р ь
нервной пластинки, желоба и
нервной т п у Ь к и с полостью
невроцелем. Затем начинается период диф&еренцировки нервной трубкй НЙ головной и спинной мозг, когда прямая до сих пор трубка образу­
ет ряд вздутий, изгибов и дает утолщения стенки в определенных
местах. Формирование головного мозга у всех позвоночных начинается
с образования на переднем конце трубки трех вздутий, или мозговых
пузырей. Первый из них носит название переднег?Т,"фго5епсерЬа1оп),
второй — среднего (шезепсерЬаЬп) и третий — заднего мозгового
пузыря (гЬошЬепсерЬа1оп). После стадии трех мозговых пузырей
следует стадия пяти м о з г о в ы х п у з ы п е й Они дают начало дефинитив­
ным отделам мозга. Передний мозговой пузырь делится поперечной
перетяжкой на два отдела. Первый их них (передний) образует перед­
ний отдел головного мозга (1е1епсерЬа1оп), который у большинства
позвоночных образует так называемые(годушария~5одьщого мозга. г }о
И з задней части переднего мозгового пузыря развивается | промежу(<йепсерЬа1оп). Средний мозговой пузырь не делится и
целикомйреобразуется в (ахндниЙ,моз? (тезепсерЬаЬп). Задний моз­
говой пузырь также подразделяется на_дйа отдела: в передней части он
образует
или
(те1епсерЬа1оп), а из заднего
1 —
(шейииа оЫоп§а1а), который
отдела ^
без резкой границы переходит в спинной мозг (рис. 146).
Таким образом, у всех классов позвоночных головной мозг состоит
из пяти основных отделов: переднего, промежуточного, среднего,
заднего и продолговатого, расположенных в одной и той же
последовательности.
Степень развития отделов у различных классов не одинакова.
В процессе- образования пяти пузырей невроцель образует ряд расшире­
ний, которые носят названия желудочков мозга. Желудочки соответ338
*н
ствую т отделам мозга и сообщаются между собой и со спиннол
вым каналом.
В каждом отделе мозга различают крышу, или мантию (раШ ит), и
дно” или основание. Кры ш у составляют части мозга, лежащие над
желудочками, дно (основание) лежит под желудочками.
Вещество мозга неоднородно. Скопления нервных клеток имеют
более темную окраску — серое вещество, скопление отростков нервных
клеток более светлую — белое вещество. Слой серого вещества
(нервных клеток) на поверхности крыши любого отдела мозга носит
название коры (сопех).
Больш ую роль в эволюции нервной системы играет развитие
органов чувств. Раздражения, поступающие отних, служат фактором,
стимулирующим прогрессивные изменения нервной системы. Э т а связь
обусловлена филогенетически. Важнейшие органы чувств развиваются
непосредственно из стенок нервной трубки. Концентрация органов
чувств на переднем конце тела обусловила прогрессивное развитие в
процессе эволюции именно головного отдела нервной трубки. Основное
значение в развитии центральной нервной системы, в частности
головного мозга, отводят группе дистантных рецепторов (органы
зрения, слуха, обоняния). Полагаю т, что передний мозговой пузырь
сформировался под влиянием обонятельного, средний — зрительного и
задний — слухового рецепторов.
Рассмотрим строение головного мозга у представителей различных
классов позвоночных.
Головной мозг
р Ыб
имеет пиимит^упе гтппеииЕ- ч т о
•разлитии,.его
выражается в ксзначцтельцом -^&ьем»-мозга
Го отдел'
ПКрГОТОГЯЙг по сравнению с другими отделами мал и не разделен
нд долушагш^ Желудочек мозга представляет со(лл1"одп/
полость. Кры ш а переднего мозга тонкая, у костистых рыб она не
содержит нервнЪй ткани и состоит только из эпителия. Основную массу
переднего мозга составляет дно, где нервные клетки образуют два
скопления, получивших название полосатых тел (согрога хЗпаЗа). О т
переднего мозга вперед., отходят небольшие обонятельные доли. По
существуетереднйГ моз/^рыб связан только с органом обоняния и
служит
За пОрГТТОТГМ
^Л ом ежуточны^, который прикрыт
сверху передним и средним мозгом. О т 'к^ыщи^промежуточного мозга
отходит вырост — верхняя мозгоМая'жёлет^ ТэпифизУ от дна — во­
ронка С Г1пи гтежя1 иим к
, образующие
перекрест. Нижняя часть промежуточного мозга Ч^ л п ю гш ГШ у»^- со­
держит нейросекреторные клетки и тесно связанЗ С~~эндоиряпной
си с^ ш ш
представляеУ
отдел мозга
рыо. Он состоит ю двух полушарий (зрительных долей), покрытых
корой, и служит 6р'^^е'л"ыым ц ентров Орган зрения у рыб играет
ведущую роль при поисках шшЦтг поэтому величина среднего мозга
достигает значительных размеров. Кроме того, средний мозг пред­
ставляет собой ведущий интегрирующий отдел головного мбзга,
339
поскольку именно сюда поступает информация от всех органов чувств,
осуществляется ее анализ и вырабатываю тся ответные реакции. По
принятой классификации такой тип мозга, где ведущими являются
задние-шчшгы, носит название щдаюнеидрюш,
МёгЗастокДкоторый у всех позвоночн^ дщ щ д д щ ц д |'т п 1 " ш!1" ™
(согласование) движений, у рыб развит
поскольку
движения их отличаются..большой сложностью . ПродолтШааг.ыЛ.мозг
содержит ц е и г р ы ^ Г & ^ к ^
Прогрессивные изменения у а м_ф и б и й выражаются в увеличе­
нии общего объема головного ~ мозга и Ц
|^
; что связано с адаптацией к^словиямназемной среды.
мозг амфибий по сравнению с рыбами значительно
увеличивается."' Одновременно происходит разпегтание его ня пня
ПбТтушария, в связи с чем общая полость желудочка также подразделяетСЯ Н2Г"Я'ва боковых желудочка. Кры ш а переднего мозга состоит из
о нервной ткани. Поверхностное положение занимают нервные волокна
(белое вещество), в глубине, выстилая полость желудочка, лежит слой
нервных клеток. В основании переднего мозга лежат полосатые тела.
Как и у рыб, передний мозг амфибий выполняет функции обонятельного
центра. Промежуточный мозг хорошо виден сверху, так как меньше
прикрыт средним мозгом. Кры ш а его образует придаток — эпифиз, а
дно — часть гипофиза. Средний мозг еще сохраняет крупные размеры,
но несколько уменьшен в объеме по сравнению с тем же отделом рыб.
Как и у рыб, средний мозг может бы ть признан ведущим интегрирую­
щим отделом центральной нервной системы, сохраняя при этом
значение зрительного центра. Таким образом, амфибии имеют ихт!Ь-:
опсидный тип мозга. М озжечок развит слабо и имеет вид небольшого
пбйерёчного валика у переднего края продолговатого мозга. Слабое
развитие мозжечка соответствует простым движениям амфибий, не
требующим сложной координации (лягушка передвигается путем
однообразных движений — прыжков, большинство же амфибий ведет
малоподвижный образ жизни).
. р е п т и л и и являются первыми истинно наземными позво­
ночными, поэтому условия их обитания по сравнению с водной средой
характеризуются большим разнообразием и непостоянством действую­
щих факторов. Резкие и быстрые колебания температуры, влажности,
соотношения химических агентов, разнообразие звуковых и зрительных
раздражителей вызываю т у наземных форм прогрессивное развитие
органов чувств и головного мозга.
Передний мозг становится наиболее крупным отделом по
сравнению с остальными. Кпереди от него отходят хорошо развитые
обонятельные доли. Увеличение переднего мозга происходит в основ­
ном за счет развития полосатых тел, т. е. дна. Кры ш а остается тонкой.
На поверхности крыши впервые в процессе Эволюции появляется.кора,
которая еще не покрывает всей поверхности1 полушарий, а образует
тодькр два островка — на медиальной и латеральной сторонах каждого
полушария. Функциональное значение имеет только ее медиальный
•■^участок, представляющий собой высший обонятельный центр. В целом’
кора переднего мозга имеет примитивное строение и получила название
340
4
5 .
Рис. 148. Головной мозг позвоночных (продольный срез).
а — к о с т и с т а я р ы б а; б — эл ектр и че ски й с к а т ; в
л я г у ш к а ; г — р ептил и я; д — птица; е — м лекопитаю щ ее.
I — передний м озг; 2 - эп иф и з; 3
гипоф из; 4 — средний м озг; 5
м о зж е ч о к ; б — продолговаты й мозг;
7 — п р о м еж у то чн ы й м озг; 8 - м анти я ( к р ы ш а ) ; 9
п ол осаты е тел а.
древней, или агсЬког{ех. Полосатые тела переднего мозга сильно
развиты и принимают на себя функцию интегрирующего центра
нервной деятельности. Таким образом, передний мозг становится
ведущим отделом центральной нервной системы. Подобный тип
мозга получил название зауропсиднош. (стриарного).
Средний мрат тррд^т св'б? Значение ведущего отдела, одновременно
уменьшается его значение как зрительного центра, в связи с чем
размеры его сокращаются (рис. 147, см. на цвет, вкл., 148). Мозжечок
развит значительно лучше, чем у амфибий. Головной мозг млекопитаю­
щих, так же как и моз1_ п т и ц ,
отличается з начите льНы ^
непропорциональным развитиёмпоТфиарий переднего мозга. Но
у^лшевде^^аз^епрБ\^цлекопит^пшх шюи^зюдшне^а сяетдна^ш *
гиц, а
всей- ,
Ьвер^ноСТт^Чфьйпи пояш ТЯетЪГк&{$а/'кот5раяр^^
за счет С ?
клеток латерального островка древней коры рептилий. Последний,
341
разрастаясь по всей поверхности полушарий, оттесняет медиальный
участок к перемычке между полушариями, где он сохраняется под
названием гиппокампа и выполняет функцию высшего центра обоняния.
Возникшая кора, в отличие от древней коры рептилий, получила
название новой коры, или пеосоПех. По строению новая кора
представляет собой сложную структуру, состоящую из многих слоев
нервных клеток различного типа. У низших млекопитающих по­
верхность коры гладкая, у высших она образует многочисленные
извилины, резко увеличивающие ее поверхность. Количество нервных
клеток в коре человека исчисляется в миллиардах. Вместе с развитием
коры развивается сложная система волокон, соединяющих правое и
левое полушария (комиссура), а также ассоциативные волокна между
различными отделами мозга. Кора приобретает значение интегрирую­
щего отдела головного мозга, в ней находятся высшие чувствительные
(зрения, слуха, осязания) и двигательные центры, а также центры,
связанные с высшей нервной (психической) деятельностью.
Тип мозга^ где высшие центры нервной деятельности расположены
в крыше переднего мозга, получил название маммального. Э то наиболее
прогрессивная ступень эволюции нервной системы.
^
Промежуточный мозг прикрыт сверху передним и имеет*
характерные для всех нижеперечисленных классов придатки в виде
< эпифиза и гипофиза. Средний мозг резко уменьшен в размерах. Его
крыша, в отличие от Предыдущих классов, кроме продольной борозды,
имеет еще и поперечную, поэтому вместо двух полушарий (зрительные
доли) образуется четыре бугра (четверохолмие). Передние бугры
связаны со зрительными рецепторами, а задние — со слуховыми.
♦
^Мозжечок прогрессивно развивается, что выражается в резком
увеличении размеров органа и его сложной внешней и внутренней
структуре.
В продолговатом мозгу по бокам обособляются пучки нервных
волокон, идущих к мозжечку, а на нижней поверхности — продольные
валики, получившие название пирамид.
У человека происходит дальнейшее развитие коры, ччро привело к
появлению новой высшей формы нервной деятельности — абстрактно­
му мышлению. Таким образом, материальный субстрат сложной
психической деятельности возник в результате постепенных преобразо­
ваний в процессе эволюции нервных структур, обладавших первона­
чально лишь простой формой раздражимости. В последнее время
выявлена качественная особенность мозга млекопитающих и челове­
ка — функциональная асимметрия. Оказалось, что левое и правое
полушария неравнозначны по своим функциям. Правое отвечает за
образное мышление, левое — за абстрактное. В левом полушарии
находятся центры письменной и устной речи.
Органы чувств можно разделить на две группы — контактные и
дистантные. К контактным относятся органы вкуса и осязания,
сигнализирующие о раздражителе при непосредственном контакте с
342
ним. Дистантные рецепторы, в отличие от контактных, получают
сигнал от раздражителя, находящегося на расстоянии. Дистантным
рецепторам принадлежит главная роль во взаимодействии организма со
средой, поскольку они получают большее количество раздражений,
более разнообразных по своей природе. Поэтому мы ограничимся
рассмотрением филогенеза этой группы органов чувств.
Органы обоняния. По отношению к беспозвоночным правильнее
говорить об органах химического чувства, а не органах обоняния, так
как они воспринимают раздражения как от жидких, так и от
газообразных веществ.
У кишечнополостных, большинства червей и моллюсков химически
возбудимые клетки рассеяны по всему телу. У червей, кроме того,
нередко встречаются обонятельные ямки или специализированные
органы — антенны, пальпы и др. У членистоногих хеморецепторы
представлены обонятельными сенсиллами.
У р ы б органы обоняния имеют вид пары слепых мешков на спин­
ной стороне головы, выстланных мерцательным эпителием. Наружные
отверстия — ноздри — соединяют полость этих мешков с внешней
средой.
У а м ф и б и й обонятельные каналы соединены не только с
внешней средой, но и с ротоглоточной полостью (хоаны или внутренние
ноздри). Начинается дифференцировка носовой полости на обоня­
тельный и дыхательный отделы, появляется носослезный канал и
железы.
У рептилий
имеются обонятельные ходы с ноздрями и
хоанами. Средний отдел хода уже довольно четко подразделяется на
верхний — собственно обонятельный отдел и нижний — дыхательный.
Появляется обонятельная раковина. С появлением вторичного неба
хоаны отделяются вместе с верхней частью ротовой полости, образуя
носоглоточный ход. Отверстия хоан благодаря развитию вторичного
неба смещаются назад, ближе к глотке или гортани, что обеспечивает
возможность свободного дыхания во время нахождения во рту пищи.
Считаю т, что такое разделение дыхательных и пищевых путей явилосв
предпосылкой для процесса пережевывания пищи у млекопитающих.
Выстилка носовой полости приобретает характер слизистой, увлажня­
ется секретом слизистых бокаловидных клеток.
У м л е к о п и т а ю щ и х прогрессивные преобразования выра­
жаются в увеличении объема обонятельной полости и появлении
системы обонятельных раковин, число которых колеблется от 5— 8 до
2 (у человека 3). Развивается мясистый наружный нос, поддерживаемый
носовыми костями и хрящами.
Органы зрения. Восприятие светового раздражения свойственно
организмам различного уровня.
Низшие беспозвоночные не имеют истинного зрения, т. е. не могут
воспринимать внешний вид объектов, но способны различать свет и
темноту. Считаю т, что зрительной информации принадлежит главная
роль в эволюции животного мира. Очевидно, этим можно объяснить
появление уже у беспозвоночных, в частности низших кишечнополо­
стных, а также аннелид и моллюсков, наряду с примитивными
343
органами светоощущения (глазные пятна, ямки) сложно устроенных
глаз, содержащих роговицу, хрусталик, сетчатку.
Глазные пятна — группа чувствительных и пигментных клеток,
расположенных на наружных покровах. Если чувствительные клетки
образуют углубление, возникает глазная ямка. С превращением ямки в
замкнутый глазной пузырек его передняя часть и прилежащий участок
покровов приобретают прозрачность (роговица). Прозрачный участок
пузырька может утолщаться, приобретать выпуклую форму, превра­
щаясь в хрусталик. Внутренность глазного пузырька преобразуется в
бесструктурное стекловидное тело. Дно и боковые стенки выстланы
чувствительными и пигментными клетками (сетчатка). Считаю т,
однако, что, несмотря на сложное строение органа, кишечнополостные
обладают только светоощущением.
У низших хордовых (ланцетник) органом светоощущения являются
особые глазки (Гессе), расположенные в нервной трубке. Они
представляют собой светочувствительные клетки, сходные с таковыми
ресничных червей.
Парные глаза позвоночных имеют большое сходство со сложно
устроенными глазами беспозвоночных (сетчатка, хрусталик, стекло­
видное тело), но не гомологичны им. Органы зрения беспозвоночных
образуются за счет впячивания наружного покрова, а глаза позво­
ночных — за счет выпячивания стенок нервной трубки. Кроме того,
хрусталик позвоночных развивается за счет покровов, а не из передней
стенки глазного пузырька. Закладка в эмбриогенезе органов зрения у
всех позвоночных происходит однотипно.
У р ы б роговица плоская, а хрусталик имеет шаровидную форму.
Круговые мышцы зрачка и хрусталика отсутствую т. Аккомодация
(изменение фокусного расстояния) происходит за счет перемещения
хрусталика к передней камере или от нее с помощью отростка сосуди­
стой оболочки, приращенного к задней стенке хрусталика. Веки
отсутствую т.
У амфибий
роговица становится выпуклой, а хрусталик
приобретает форму двояковыпуклой линзы, что увеличивает дально­
зоркость. В воде роговица уплощается. Появляются веки (верхнее и
нижнее) и мигательная перепонка (третье веко) в переднем углу глаза,
предохраняющие глаз от высыхания и загрязнения. Появляется
круговая мышца зрачка и хрусталика, но аккомодация чаше осуще­
ствляется, как у рыб.
У р е п т и л и й изменяется механизм аккомодации, она осуще­
ствляется за счет сокращения ресничной мышцы, окружающей
хрусталик. Последняя располагается позади хрусталика, при ее
сокращении он перемещается вперед, кривизна меняется мало.
У млекопитающих
зрение играет относительно менее
важную роль. Аккомодация происходит за счет изменения кривизны
хрусталика, появляются слезные железы, ресницы.
Органы слуха. У большинства беспозвоночных и позвоночных
восприятия звуковых раздражений и восприятия положения тела в
пространстве совмещаются в одном органе.
У беспозвоночных органы этого типа обычно служат органами
344
равновесия. Они представляют собой ямки или пузырьки, выстланные
чувствующими клетками, в полости которых находятся «слуховые»
камешки, или статолиты. В зависимости от положения тела статолиты
давят на соответствующий участок стенки, что позволяет животному
ориентировать свое тело в пространстве.
У насекомых имеются и специальные органы, воспринимающие
звуковые колебания в виде так называемых хордотональных органов.
У позвоночных орган слуха также является одновременно и органом
равновесия, причем в процессе эволюции собственно орган слуха
развивается позднее.
У к р у г л о р о т ы х и р ы б орган слуха представлен только
внутренним ухом.
В процессе эмбриогенеза внутреннее ухо развивается за счет
утолщения эктодермы в задней части головы. Сначала образуется
парное впячивание, затем — замкнутый пузырек, который подразделя­
ется на два отдела — верхний и нижний. Из верхнего формируются
полукруглые каналы, лежащие во взаимно перпендикулярных плоско­
стях, и овальный мешок. И з нижнего отдела пузырька развивается
круглый мешочек с небольшим выростом — будущая улитка.
Совокупность каналов и мешочков образует перепончатый
лабиринт внутреннего уха, стенки которого состоят из эпителия.
Внутри его находится жидкость — эндолимфа. В ней взвешено множе­
ство мелких кристаллов извести (отолиты). В некоторых местах
лабиринта расположены скопления чувствительных, снабженных воло­
сками клеток — слуховые пятна. При давлении отолитов на те или
иные клетки раздражение передается в мозг и животное воспринимает
свое положение в пространстве. Перепончатый лабиринт свободно
взвешен в полости внутри слуховой капсулы (хрящевой или костной).
Стенки полости повторяют контуры перепончатого лабиринта. Внутри
полости также находится жидкость — перилимфа.
У круглоротых имеется только два или даже один полукружный ка­
нал. У рыб появляются три полукружных канала. Отдел гомологичный
улитке, у большинства рыб имеет вид небольшого слепого выроста
круглого мешочка, в котором обособляется слуховой сосочек от
слухового пятна. К нему подходит отдельная обособившаяся ветвь
слухового нерва. Ры бы воспринимают звук с частотой 30— 12 кГй.
У а м ф и б и й в связи с переходом к наземному образу жизни,
кроме внутреннего, появляется среднее ухо. Оно играет роль
добавочного аппарата для улавливания звуковых волн в воздушной
среде, где проводимость ниже, чем в водной, и передачи их внутреннему
уху. Среднее ухо развивается из остатка первой жаберной щели,
зарастающей снизу. Оставшаяся в верхней части полость располагается
позади глаз, вплотную примыкает к слуховой капсуле и сохраняет связь
с глоткой. С выходом амфибий на сушу она превращается в воздухо­
носную барабанную полость, которая граничит с внутренним ухом с
одной стороны и наружными покровами — с другой. Н а одном из
участков покров истончается и переходит в упругую барабанную
перепонку. В полости расположена одна слуховая косточка (гиомандибулярный хрящ), упирающаяся одним концом в барабанную перепонку,
345
,/
Рис. 149. Эволюция слухового аппарата у позвоночных. Левое внутреннее ухо,.
а
костистая рыба;
б
- ам ф иб ия; в — р е птил и я; г — п ти ц а; д — м лекоп итаю щ ее, I полу­
к р у ж н ы е к а н а л ы ; 2 — к р углы й м еш очек; 3 — овал ьны й м еш очек; 4 — вы сту п круглого м еш очка
(з а д а т о к у л и т к и ); 5 — кан ал ул и тк и ; 6 — слуховое пятн о кр угло го м еш очка.
1
а другим в слуховую капсулу, стенка которой в этом месте становится
перепончатой — овальное окно. Колебания барабанной перепонки
усиливают звуковые колебания и передают их на внутреннее ухо. Во
внутреннем ухе у амфибий прогрессивно развивается зачаток улитки.
Вырост удлиняется.
У р е п т и л и й во внутреннем ухе слепой вырост круглого
мешочка еще больше увеличивается в длину и начинает изгибаться в
канал улитки. Внутри канала начинают фо'-мироваться части восприни­
мающего аппарата.
Строение среднего уха в принципе не меняется. Н а границе с внут­
ренним ухом в задней стенке появляется, кроме овального, второе
перепончатое окно — круглое, что уменьшает несжимаемость перилимфы и совершенствует передачу звуковых колебаний. Барабанная
перепонка расположена несколько ниже общего уровня кожных
покровов, образуя зачаток наружного слухового прохода. У млекопита­
ющих орган слуха состоит из трех отделов: внутреннего, среднего и
наружного. Внутреннее ухо достигает наивысшего развития. Канал
улитки сильно удлиняется и изгибается спирально. Основной слуховой
сосочек преобразуется в спиральный орган. Таким образом, у наземных
позвоночных прогрессивно развивается слуховой аппарат, в то время
как строение органа равновесия остается на том же уровне.
346
В среднем ухе усложняется передаточный аппарат — появляются
3 косточки — молоточек, стремечко и наковальня. В результате звуко­
вые колебания в процессе передачи усиливаются примерно в 16 раз
(рис. 149).
Кроме того, у млекопитающих формируется наружное ухо:
наружный слуховой проход и наружная слуховая раковина. Последняя,
перемещаясь с помощью специальной мускулатуры, концентрирует
звуковые волны и направляет их в слуховой проход.
Органы выделения беспозвоночных. \^_.1щшечнодолостных, тело
которых состоит всего из 2 слоев, продукты диссимиляции удаляются
во внешнюю спепуптискЬузно.
,
У ддооких -червей появляется выделительная система в виде
Чрротонефрндиев (см. ГГ,П . У к р у г л ы х червей наряду с измененными
'Я}5от5НефрйДиями существуют гигантские кожные клетки.
\
К о л ьчатые черви имеют сёгмёнтарнуЮ-выделительную~Ейстему в
^]видё метанефридаев. В каждом сегменте расположена пара метанефридаемГЙаждый" состойт из воронки,^открывающейся в целом и отходя­
щего от нее вьщелительного канальца и яыделитедьной поры.
Органы выделения членистоногих^пелставлены видоизмененными
метяне^пи лиями. особыми (мальпигиевыми! сосудами (см. главу 18) и
--’""~
5кш ю в ы м телом0
^
■
-^^ОрганьГБьщ еления позвоночных. Органом выделения позвоночных
служат почки. По своему внешнему виду почки (парный компактный
орган) отличаются от нефридиев беспозвоночных и низших хордовых,
но по строению во многом сходны , с ними. О сновной—структурной
единицей почек, так же как и нефридиев, является воронка,
открывающаяся в полость тела, с отходящ им.от нёё~вьЩёли тельным
■ЧШГР-'У" В р »--т л д ^ н т едьшлс—-каывтгьцы впадают в "одия—общий
выводной проток —^мочеточник, в то время как каждый каналец
нефридия имеет самостоятельное отверстие. Первоначально воронки с
канальцами закладываются метамерно, однако позднее в филогенезе
метамерность утрачивается.
У низших позвоночных в эмбриогенезе последовательно закладыва­
ются органы выделительной системы сначала в головном отделе
(преддочка), затем в туловищном (первичная почка). У высших
позвоночных, кроме этих двух, образуется еще третья закладка в
тазовом—отделе (вторичная понка). Более ранние закладки после
появления псюледукшйхТэедуцируются (субституция).
Подобная смена органов выделения в онтогенезе отражает этапы
филогенеза и позволяет проследить пути эволюции выделительной
системы.
П р е д п о ч к а или головнг
меет наиболее
примитивное строение. Она закладывается у всех позвоночных на
ранних стадиях эмбрионального развития в головном конце тела и
состоит всего из 6— 12 нефронов, представляющих собой структурно­
функциональные единицы органа выделения. Нефрон предпочки
347
Рис. 150. Эволюция нефрона.
а — п р едп о чка; 6 — п ер ви чн а я п о чк а; в
в т о р и ч­
н ая п о ч к а ; г ~ п очка че л о в е к а ; 1 -— неф ростом ;
2 — вы д е ли тел ьн ы й к ан ал ец ; 3 — м очеточн и к; 4 —
к л у б о че к к а п и л л яр о в ; 5 — целом; 6 - к а п с у л а п о че ч­
ного к л у б о ч к а ; 7 — п р о кси м ал ьны й и зви то й кан алец ;
8 — д и ста л ь н ы й и зви то й к ан ал ец ; 9 — п ет л я нефроиа.
начинается воронкой(нефростом) с ресничками, открывающейся в
целом, а короткий и прямой выделительный каналец, отходящий от
воронки, открывается в общий для "них всех мочеточник, который
растет вдоль позвоночника и открывается в клоаку. Рядом с воронкой,
за брюшиной, развивается несколько сосудистых клубочков. Продукты
диссимиляции из клубочков поступают в целомическую жидкость, а
затем, смешиваясь с ней, попадают в нефростомы, канальцы и
мочеточник. Несовершенство предпочки заключается в отсутствии
прямой^.;фвязи между кповеносной и-выделительной системами.
Продукты распада постоянно присутствуют в целомической жилкости.
У современных позвоночных предпочка существует только в
эмбриональном периоде. Во взрослом состоянии пронефрос функциони­
рует лишь у некоторых круглоротых. У эмбриона человека канальцы
пронефроса функционального значения не имеют.
, /
Первичная,
или
туловищная,
почка
(тё§оперйгох) — второй этап эволюции выделительной системы позвоночных.
Она закладывается в туловищных сегментах тела. Строение нефрона
усложняется — на спинной стенке выделительного канальца появляется
слепой вырост в виде двустенной чаши (капсулы почечного клубочка).
348
эту капсулу врастает сосудистый клубочек, образуя вместе с
капсулой (почечное тельце/ Благодаря этому возникает прямая связь
между кровеносной и выделительной системами. Теперь продукты
! диссимиляции из крови сразу поступают в почки, не попадая в целом.
Удаление продуктов распада из организма происходит более полно и
быстро. Воронки первцчной..почки утпачивают свое чначениег и в тече­
ние жизшГТГТюкоторых нефронах происходит их редукция, связь с
целомом частично утрачивается (рис. 150).
Прогрессивным изменением нефрона следует считатц (удлинение
; выделительного канальца, благодаря чему он становится извитым, а
~также7|ДиФФеренпиповку его отделов. В итоге в канальцах первичной
| почки'тначинаюТ осуществляться процессы обратного всасывания таких
продуктов, как вода, глюкоза и др., т. е. происходит концентрация
*- мочи.
~I
Количество нефронов в первичной почке по сравнению с предпочкой
Р увеличивается, так как на каждом первичном канальце позднее
| возникает один или несколько добавочных нефронов путем своеI
образного почкования, ^тдл^рно^ть.строеыилндддшсх.утращщаться,1
поверхность выделения значительно увеличивается. Первичная почка
I' функционирует у низших позвоночных (рыбы, амфибиТ|)Гв'т^ченйе всей
Г жизни, у высших (рептилии, птицы, млекопитаюпше) сохраняется
только в эмбриональном состоянии.
Вторичная,
или
тазовая,
почка
(те*аперЬго$)
закладывается у высших позвоночных в сегментах тела, лежащих кзади
от туловищной почки.
Отличительный признак нефронов — ш хутсхаие в о р о н к и , благода­
ря чему'связь с целомом полностью утрачивается Нефрон начинается
прямо с точечного тельца. ВьшелительныйТаналёй джшЬсрен шшлет ся
на ряд отделов — проксимал1Гнь® ''’изШТб1Г”” каналец, дистальный
извитой каналец, петля нефрона и т. д. Клубочковый аппарат
упрощается, в частности количество капиллярных петель в клубочке
I
уменьшается, фильтрационная способность отдельного клубочка сни■ жается, зато усложняется строение канальцев, увеличивается их длина.
( Усиливается секреция клеток стенок канальцев, выделяющих продукты
распада в просвет канальцев. Таким образом, в тазовой почке продукты
[ диссимиляхщи-поступают в нефрон двумя путямйГТгуТём фильтрации
| плазм ьм^подость.|щдсудь1 ^ у БЩ ка и за счет выделения~ЛСТПсств в прос[ вет "канальцев. Одновременно в канальцах интенсивно происходят
^тЬрцёссы обратного всасывания. Моча в полости капсулы клубочка
содержит некоторое количество полезных для организма низкомолеку­
лярных соединений: сахара, витамины, аминокислоты, хлориды и т. д.
При прохождении мочи по отделам канальцев происходит реабсорбция
этих веществ и большей части воды обратно в кровь.
У рептилий канальцы имеют проксимальный извитой отдел,
промежуточный, дистальный извитой отдел, петля нефрона отсут­
ствует.
—
*
“
У птиц и млекопитающих появляется петля—нефрона — главное
звено в механизме реабсорбции воды.
О т каждого первоначального нефрона путем почкования образуется
349
несколько вторичных, в связи с чем количество нефронов возрастает во
много раз, и обшая выделительная поверхность резко увеличивается.
Мочеточник тазовой почки образуется независимо от мочеточников
пре^ествую лш х~5о1Гекг-Основное направление эволюции вьщелителнйбй системы —^Увеличение выделительной поверхности, что обес­
печивает более б ы с т р О Л Г полное удаление пр'бй^тов” обмена, и
диффершщфовка.вьшеди-телы40Ед.зсанала, предупреждающего потерю
организмом важных компонентов (вода, углеводы).
Связь выделительной и половой систем. Выделительная система
позвоночных связана с органами половой системы. У некоторых
позвоночных, например осетровых рыб, закладка половых желез
происходит непосредственно за счет эпителия нефростомов первичной
почки. У большинства позвоночных эта связь выражается в том, что
некоторые части предпочки и первичной почки принимают на себя
функцию выведения половых продуктов.
Половые железы позвоночных, как прамыю, закладываются в виде
парных складок на вентральных краях мезшгефро&та Зачаток гонад
состоит из утолщенного эпителия с большим колитеством соедини­
тельной ткани. Сначала мужские и женские половые железы имеют
одинаковое строение, позднее происходит специализация их и возникает
связь с различными для каждого пола частями выделительной системы,
которые становятся половыми протоками.
У самок анамний после появления первичной почки предпочка
освобождается от функции выведения мочи и редуцируется. Обычно
остается лишь одна воронка, которая сильно увеличивается в размерах
(иногда она образуется за счет соединения нескольких воронок) и вместе
с мочеточником предпочки (парамезонефральный канал) преобразу­
ется в яйцевод. Подобная смена функций объясняет отсутствие
непосредственной связи между яичником и яйцеводом у позвоночных.
При созревании яйцеклеток в яичнике происходит разрыв его стенок.
Яйцеклетки выходят в полость тела, попадают в воронку, а затем в
яйцевод.
Продукты диссимиляции у самок анамний выводятся через
первичную почку и ее мочеточник, или мезонефральный (вольфов)
проток.
У самцов анамний в эмбриональном периоде происходит полная
редукция предпочкй — исчезают не только выделительные канальцы,
но и мочеточник (рис. 151).
Одновременно возникает связь между семенником и первичной
почкой. И з эпителия, выстилающего стенку полости тела, образуются
тяжи, соединяющие выделительные канальцы первичной почки и
семенные канальцы. Затем в этих тяжах образуется просвет, и они
превращаются в семявыносящие канальцы. Созревшие сперматозоиды
по семявыносящим канальцам попадают в почку и по мочеточнику
выделяются наружу. Поскольку первичная почка функционирует как
орган выделения, в мочеточник одновременно поступает моча и он
служит как для выведения мочи, так и половых клеток (мочеполовой
проток).
У высших позвоночных (амниот) с появлением вторичной почки не
350
29
Д
е
Рис. 151. Взаимоотношения органов мочеполовой системы у позвоночных.
Р ы б ы и ам ф ибии : а
сам ец ; б — с а м к а , рептилии и птицы; в
сам е ц и г
с а м к а , м л е к о п и таю ­
щие: д - сам ец и е — са м к а . I - предпочка; 2
сем енник; 3 — ссм яв ы во д ящ и й проток; 4
п ер вичная п о чк а; 5
за д н я я к и ш ка ; 6 — м очевой п у зы р ь; 7 — к л о а к а ; 8
воронка яйцевода;
9 — яи чн и к ; 10 — яйцевод; I I
придаток сем енника (о ст а то к передней ч а сти первичной п о чк и );
12 — сем яп р о во д (м езон еф р альн ы й п р о то к ); 13 - вто р и чн ая п очка (м е т а н е ф р о с ); 14
м очеточник
м етан еф р оса; 15
рудимент м езонеф р оса; 16
яй ц евод; 17 — яй ц е к л е т к а ; 18 — б елок, вы деляем ый
яйц еводом; 19
м атк а, 20 - зар о д ы ш в м атке ; 21
в д ш а л и ш с ; 22
ан ал ьн о е о твер стие; 23
рудим ент п ир ам езон еф р альн ого п ротока; 24 — м езо неф р альн ы й проток (м о чепо ло во й к а н а л ).
Рис. 152. Дифференцировка яйцеводов у млекопитающих.
а — клоачные; б — молодая двуутробка; в
плацентарное млекопитающее с двойной маткой; г — с двуро­
гой маткой; Д — с простой маткой; I - клоака; 2
прямая кишка; 3
яйцевод, 4
мочеполовой синус;
5 — мочеточники; 6 — матка; 7 — влагалище; 8
мочевой пузырь.
только предпочка, но и первичная почка освобождается от функции
выведения мочи.
У самок амниот, так же как и у анамний, из остатков предпочки и ее
мочеточника развивается яйцевод. Первичная почка и ее мочеточник у
взрослых самок редуцируются, лишь часть канальцев первичной почки
сохраняется в виде незначительных рудиментов ероорЬогоп и
рагаорНогоп. Существует представление, что эти рудименты склонны к
злокачественному перерождению.
Яйцеводы у амниот дифференцируются на отделы. У млекопитаю­
щих в связи с появлением функции живорождения дифференцировка
яйцеводов становится наиболее сложной (рис. 152). Яйцеводы подраз­
деляются на 3 отдела — маточные трубы, матку и влагалище.
У высших млекопитающих — плацентарных — происходит срастание
дистальных отделов яйцеводов на разных уровнях. У некоторых видов
срастается только влагалищная часть, а матки остаются парны­
ми — двойная матка (грызуны); у некоторых матки слиты в дисталь­
ном отделе, образуя общую полость, в то время как их проксимальные
отделы сохраняют самостоятельность — двурогая матка (хищники,
парнокопытные). Наконец, матки могут бы ть полностью слиты в одну
352
на всем протяжении — простая матка (полуобезьяны, обезьяны,
человек и некоторые летучие мыши).
У человека встречаются различные аномалии матки и влагалища,
соответствующие филогенетическим этапам изменения этого органа в
процессе эволюции. Как правило, аномалии связаны с ненормальным
срастанием парамезонефральных протоков. Наиболее часто встреча­
ется двурогая матка, иногда Щ ЯФШ Р&ххС
У самцов амниот п ро неф лРси еЬ мочеточник полностью
редуцируются. У млекопитающих. и человека за счет остатков
дистального отрезка парамезонефрального канала формируется слепое
выпячивание, являющееся гомологом матки и влагалища (и*еги$
шахсиНпих).
Канальцы передней части первичной почки у самцов сохраняются и
преобразуются в придаток семенника — эпидидимис, а мочеточник
первичной почки (мезонефральный канал) превращается в семяпровод.
.13.9. Эндокринные железы
Железами внутренней секреции (эндокринными) называют органы,
секретирующие биоЯОТйПеС№~ШШшнь1 е вещества, оказывающие регу­
лирующее влияние на жизнедеятельность организма. В отличие от
желез внешней секреции, эндокринные железы не имеют протоков и
выделяю т секреты , получившие название гормонов, непосредственно
в кровь.
Наибольшего развития эндокринные железы достигают у позво­
ночных. Эндокринные железы развиваются в процессе филогенеза из
разных источников и имеют различное местоположение.
Щитовидная
железа
развивается за счет эпителия
жаберного отдела глотки. У рыб закладка щитовидной железы имеет
вид продольного желобка на брюшной стороне глотки — между первой
и второй жаберными щелями. Желобок выстлан мерцательным
эпителием и содержит железистые клетки. В последующем этот
участок обособляется от глотки и превращается в замкнутый пузырек.
Стенки его утолщаются и в них появляются отдельные фоллику­
лы — структуры, характерные для щитовидной железы. Клетки стенок
фолликулов выделяют секрет, накапливающийся в их полостях.
У последующих классов меняется положение железы, появляются доли
и перешеек.
Тимус
у рыб развивается в виде множественных закла­
док — плотных эпителиальных выступов спинной части всех жаберных
мешков. Выросты отшнуровываются и на каждой стороне соединяются
между собой, образуя полоску с узкой полостью внутри. О т полости
наружу растут многочисленные выпячивания, состоящие из лимфо­
идной ткани, которая делится на дольки врастающей соединительной
тканью. У амфибий и рептилий количество зачатков, из которых
формируется тимус, значительно уменьшается, иногда развиваются
только закладки второй и третьей пары жаберных мешков. У млекопи­
тающих в образовании тимуса принимают участие выросты трех пар
мешков (в основном второй пары). Э ти выросты лежат на брюшной
стороне впереди сердца непосредственно над грудиной (рис. 153).
353
13— 56
и, б
аил мини, и
ам иш пы ;
пара щ итовидной ж е л е зы ; I
IV
I
щ и ю н и д н аи ж е л е з а ; 2
ж аб ер н ы е м еш ки.
Тимусу присуща эндокринная функция. Вместе с тем он является
важнейшим органом иммунной системы, филогенез которой рассмат­
ривается ниже (см .7.13.10).
Г и п о ф и з, или нижний придаток мозга, образует с нейросекре­
торной зоной промежуточного мозга — гипоталамусом — единую гипоталамо-гипофизарную систему. Гипофиз состоит из трех долей:
передней (аденогипофиз), средней (промежуточной) и задней (нейроги­
пофиз). Он соединен с нижней поверхностью промежуточного мозга
выростом дна этого отдела мозга, или воронкой. Доли гипофиза в
процессе фило- и онтогенеза развиваются из разных источников.
В процессе эволюции гипофиз впервые появляется у круглорохых. но
имеет весьма примитивное строение и состоит только из передней доли.
У рыб гипофиз закладывается как широкое мешковидное выпячивание
крыши ротовой полости. По мере роста его дистальный конец
оказывается под дном промежуточного мозга, образующего вы­
рост — воронку. Последняя имеет широкую полость и плоское тонкое
дно. Воронка выпячивает дно кармана так, что образуется двустенная
структура.
Из передней стенки гипофизарного кармана образуется передняя
доля. В ней развивается множество ветвящихся трубочек, которые
распадаются на клеточные тяжи и группы клеток, образуя железистую
ткань. За счет задней стенки кармана образуется средняя доля. Учрыб
она развита очень сильно и имеет больший объем, чем у всех последую­
щих классов позвоночных. Функции пепелней— и средней доли
сходны — ^бе вырабатываю т гонадотропные горкюдьД
Воронка состоит из мозговой ткагт, нмсот тОйкое дно, но из него
начинают образовываться пальцеобразные выросты, глубоко вдающи­
еся в ткань средней доли. Вместе с выростами в среднюю долю начина­
ю т проникать отростки нейросекретных клеток гипоталамуса. Гипофиз
354
хрящевых рыб во взрослом состоянии сохраняет связь с ротовой
полостью.
У .амфибий появляется обособленная задняя доля гипофиза. Ее
появление в процессе филогенеза связываю т с переходом к наземному
образу жизни, поскольку нейрогипофиз принимает участие в регуляции
Задняя доля развивается из дистального отдела
воронки промежуточного мозга. Дно и задний отдел последней
утолщаются в основном за счет разрастания нейроглии. Благодаря
сходству с тканью мозга задняя доля получила название нейрогипофиза.
В толще задней доли проходят отростки, связывающие нейрогипофиз с
определенными ядрами гипоталамуса. По объему задняя доля сямяя
крупная из долей гипофиза. В ней происходит накопление антидиуретиЧРГКИ У
гпр м пнод
Филогенетическая связь между развитием нейрогипофиза и пере­
ходом к наземному существованию находит подтверждение в том, что
задняя доля у амфибий развивается только после окончания метаморфо­
за. Г репняя лппя гипосЬиза. начиная с амфибий, теряет способность
вырабатывать гонадотропный гормон.
У амниот продолжается обособление и дифференцировка гипоталамо-гипофизарной системы в связи с возрастающей у наземных
животных потребностью в сохранении воды организмов. Развитие
задней доли сочетается у амниот с дифференцировкой ядер гипоталаму­
са, часть из которых сохраняет связь с аденогипофизом, а другая уста­
навливает контакт с нейрогипофизом. В результате формируются две
системы — гипоталамо-аденогипофизарная и гипоталамо-нейрогипофизарная. У человека железистая зона аденогипофиза составляет
70— 80% от общей массы железы, задняя — 18—2 3% , а сред­
н я я — всего 1—2 % . У некоторых людей гипофиз сохраняет связь с
ротовой полостью. В этом случае остается узкая полоска клеток длиной
5— 6 мм и толщиной 0,5— 1 мм в подслизистой оболочке крыши глотки
и основания клиновидной кости. Клеточные элементы уплощенной
формы сходны с тканью мозга и имеют тенденцию к злокачественному '
перерождений^
• У
ВысшйО^Рпозвоночных и человека
надпочечники
представляют парный компактный орган, расположенный на верхних
полюсах почек. Железа состоит из двух слоев — коркового (наружного)
и мозгового (внутреннего). Слои возникают в процессе филогенеза из
разных зачатков. К о р к о во е вещ ество об р а з уется за счет утолщения
эпителия—б р ю ш и н ы , расположенного на участке между корнем
брыжейки и мочеполовым бугорком, мозговое — т е ч е т обособления
комплексов клеток о т^ачаткодхимпатинеских ганглиев (рис. 154).
х- У низших хордовых зачатки коркового и мозгового слоев
появляются как^ И Т О Л з?1связанные между собой, самостоятельные
образования в виде так называемых межпочечных и надпочечных тел.
У рыб закладки мозгового вещества (надпочечные тела) образуют
ряд парных зачатков, расположенных метамерно, т. е. вдоль спинной
поверхности первичных почек, в тесной связи с их сосудами.
Межпочечные тела (корковое вещество) развиваются также в виде
метамерно расположенных утолщений эпителия брюшины поблизости
355
13*
от половых валиков по обе стороны
корня брыжейки. Затем зачатки
межпочечных тел каждой стороны
сливаются и образуют тяж , лежа­
щий между почками. Иногда за­
кладки обеих сторон соединяются
и образуют непарный тяж .
У амфибий между надпочечны­
ми телами возникает пространст­
венная связь, причем строгая парность'не соблюдается.
У амниот все закладки надпо­
чечников сливаются между собой,
образуя парный околопочечный
орган, имеющий сложное строение.
.13.10. Иммунная систем:
4'
Иммунная система служит для
защиты от вирусов, микроорганиз­
мов и других клеток, генетически
чужеродных организму. Она пред­
ставлена совокупностью лимфоид­
ных органов и скоплений лимфоид­
ных клеток. Среди таких клеток
Рис.
154.
Развитие
надпочечника
выделяют Т-лимфоциты (тимусI — хор д а; 2 - ган глий си м п а ти че ско й » с т о . и
зависимые) и В-лимфоциты, разви­
Л - з а ч а т о к м о зго вого в е щ е с т в а ; 4
зачат»
тие которых не зависит от вилочкоко р ко во го с л о я ; 5 — половой буго р о к; 6
ка
н а л ь ц ы п е р в и ч н о й п о ч к и ; 7 — п а р а м е з о н е ф р а .и ,
вой железы. Их взаимодействие при
ные п р отоки ; 8 — п р оток п е р ви чн о й по чки
участии клеток-макрофагов обеспе­
чивает весь спектр иммунологи­
ческих реакций.
В эволюции сформировалось три главных формы иммунологи­
ческих реакций: 1) фагоцитоз1 или неспецифическое уничтожение генетически
------ чужеродного материала особыми клетками;
кЛетками; 21 -------клеточный
иммунитет, основанный на специфическом распознавании и уничтоже­
нии такого материала Т-лимфоцитами; 3) гуморальный иммунитет,
осуществляемый путем образования потомками В-лимфоцитов (плаз­
матическими клетками) иммуноглобулинов (антител) и связывания
ими антигенов. Различаю т три главных этапа эволюции реакций
клеточного и гуморального иммунитета. К в а з и и м м ^ н д о е
р а с п о з н а в а н и е клеток организмов свз5ег&ЛрЯллотенные)или
щругогб'^ксёногенныеУ' вида обнаружено у животных разного плана
структурной организации — от кишечнополостных до млекопитаю­
щих. Его механизм не_связан с синтезом антител и лишен и м м у н ол о г и ч е с к о й п а м я т и. олагодаря которой при друпВГрёакциях
1 Некоторые иммунологи не включают в понятие иммунитета фагоцитоз на том
основании, что этот защитный механизм лишен специфичности.
356
иммунитета ответ организма на повторное проникновение чужерод­
ного материала усиливается и ускоряется.
П
обнару­
живается у целомических беспозвоночных (кольчаты ечёрви, игло­
кожие). Он обеспечивается _целомоцитами — клетками вторичной
полости тела, уничтожающими чужеродный материал. Н а этом этапе
иммунные реакции приобретают свойство специфичности, появляется
иммунолргичегкаа память
—
------ '
н а следующем этапе сложилась ^ и ^ т ^ м ^ ^ 1^ ^ т^ ^ _^ и д ^ в а нг ^ 1 3 -ДЛ,Л-^н^_Г-0___и дщи у н и т е т а,
для'кбторой'типичны специфические гум.г ^ т н м ^ и к ге т^ тТ ^ гйакпии на чужепопное. нал^ДГД-МЮ.И'.УИ ЙЫХ .-щмфон :ш.1Л- органов и м м у нйтета. образование антител. Ни один из “перечисленных показателей
в достаточно развитом состоянии не встречается у беспозвоночных —
этот этап эволюции иммунитета характеризует лишь позвоночных
животных.
•По мнению одних авторов иммунитет формировался в филогенезе
какГриеханизм зашиты от микроорганизмов. Согласно другой точке
зрения (концепция и м м у н о л о г и ч е с к о г о н а д з о р а ) , раз­
витие системы иммунитета стимуяЬшвда со и ^ бходимостью защиты
организма от внутренней опасноГ/тт У^ т* < ственных соматических
клеток, ставших генетически чужими вследствие мутаций. Нарушение
процессов жизнедеятельности из-за соматических мутаций (мутацион­
ный риск) возрастает с увеличением количества клеток в организме.
Так, тело человека образовано примерно 1015 клетками, причем не
менее 1013 из них являются пролиферирующими. В таких условиях
при существующей частоте мутирования генов в каждой смене деля­
щихся клеток ежесуточно должно накапливаться 106 соматических
мутаций, т. е. мутационный риск достигает значительной величины.
В ряду позвоночных животных наблюдается усиление иммунного
ответа по мере усложнения структурно-функциональной организации.
Круглопотые способны отторгать аллотрансплантат и синтезировать антитела, но вопрос о наличии у них органа иммунитета —
вилочковой железы — остается открытым. Р а эвить<й--тйцу|[ГЪбнару►*йиваетсяГлишь у акул и других низших рыб. Акулы — первые в ряду
позвоночных, имеющие типичные плазматические клетки, вырабаты­
вающие антитела.
Эволюционные предшественники лимфоидных оргпнов млекопита­
ющих и человека (тимус, селезенка, скопления лимгЬоилной ткани)
обнаруживаются в полном объеме у бесхвостых амфибий, ^.низш их
позвоночных (рыбы, амфибии) в отличие от птиц и млекопитающих
вилочковая железа активно выделяет антитела.
Особенность системы иммунитета _птиц заключается в наличии
особого лимфоидного органа — фабрициевой сумки. В эмбриогенезе
этому органу принадлежит ведущая роль в развитии клеточной линии
В-лимфоцитов, т. е. способности организма к образованию антител.
Млекопитающие отличаются сложностью и разнообразием строе­
ния и расположения в организме лимфоидных органов. Предположи­
тельно гомологом фабрициевой сумки у млекопитающих служит
)
357
\
Л
.
'МЛЛЛЛДАА^
"
I 1 т а а и ш и д Д В Д ЮДОК а ш Я и м и Согласно данным сравнитель* / но-морфологических исследований, лимфоидные структуры хордовых
у эволюционировали из элементов выделительной системы.
#
Глава
8
.А Н Т Р О П О Г Е Н Е З
^
8.1. С О Ц И А Л Ь Н А Я С У Щ Н О С Т Ь И Б И О Л О Г И Ч Е С К О Е
Н АС Л ЕД С ТВО Ч Е Л О В Е К А
Соответственно г^орфофизиологической организации люди образу­
ю т единственный вид рода Ношо, относящийся к надсемейству Вдшших
узконосыхдюдотряда Человекоподобных^отряда Приматов (табл. 26).
•Последний принадлежит к^подклассу ТГлацентардых ^ласса Млекопита­
ющих
а Хордовых. Вместе с тем в органическом мире планеты люди
занимают уникальное место, что обусловлено приобретением ими в
процессе антропогенеза социальной сущности, которая «... в своей
действительности есть совокупность общественных отношений»1 . Э то
означает, что именно общество й производство, а не биологические
механизмы обеспечивают выживание, всесветное и даже космическое
распространение, процветание людей. Из социальной сущности людей
вытекаю т также закономерности и главные направления исторического
развития человечества. Социальность не противопоставляет людей
остальной природе: «... мы отнюдь не властвуем над природой, так
как... кто-либо, находящийся вне природы, — ... а мы, наоборот, нашей
плотью, кровью и мозгом принадлежим ей и находимся внутри ее...»2.
Человек включен в систему органического мира, которая складывалась
на протяжении большей части истории планеты независимо от
социального фактора и породила этот фактор в ходе своего развития.
Человек и человечество составляют своеобразный, но обязательный и
неотъемлемый компонент биосферы: «Человек должен понять, ...что
он не есть случайное, независимое от окружающего (биосферы или
ноосферы) свободно действующее природное явление. Он составляет
неизбежное проявление большого природного процесса, закономерно
длящегося в течение по крайней мере двух миллиардов лет»3.
Благодаря животному происхождению жизнедеятельность человеческо­
го организма основывается на фундаментальных биологических меха­
низмах, которые составляют биологическое наследство людей.
Особенности развития жизни в одной из ее ветвей привели к соеди­
нению в' человеке социального и биологического. Такое соединение
отражает объективный результат биологической предыстории и насто­
ящую историю вида Человек разумный. Характер взаимодействия
социального и биологического в человеке нельзя представлять как их
простое сочетание в некоторой пропорции или прямое подчинение
1 М аркс К .. Энгельс Ф . Соч. Изд. 2-е, т. 3, с. 3.
2 М аркс К-. Энгельс Ф . Соч. Изд. 2-е, т. 20, с. 496.
Вернадский В. И. Размышления натуралиста. М.: Н аука, 1977, с. 19.
358
-
Таблица
26.
Комплекс признаков, определяющих
О тр яд П р и м аты
Конечности хватательного ти­
па - 1 палец противопостав­
лен остальным; на пальцах
имеются ногти; сосков млеч­
ных желез одна пара; ключи­
цы развиты хорошо: кисть
способна к пронации и супи­
нации (локтевая кость сво­
бодно.вращается вокруг луче­
вой); зубная система неспе­
циализированная с присут­
ствием всех типов зубов; мо­
лочные зубы в процессе онто­
генеза сменяются постоян­
ными; глазница отделена от
височной ямы и глаза направ­
лены вперед; сезонность по
ловон жизни
отсутствует;
обычно рождается один дете­
ныш
По до тр яд
Чело векоп од о бн ы е
Ногти плоские, значитель­
ное развитие
головного
мозга, особенно больших
полушарий; заметная ре­
дукция
обонятельного
мозга;
присутствуют
2
нижних и 2 верхних верти­
кально
расположенных
резца; коренные зубы не­
сут 3 и более бугров на же
вательной
поверхности;
хрящевые кольца трахеи
не замкнутые; матка про­
стая; плацента гемо.хориальная, отпадающая
положение человека в
и мин т|1>И1 ШТИИМии
Мидсе м р йот но В ы с ш и е \ ш л ю е ы е
Присутствие дуг, петель и замкну­
тых фигур в пальцевых узорах кисти;
редукция хвостового отдела позво­
ночника; укрепление таза посред­
ством уменьшения числа грудо-поясничных позвонков до 16— 18 и увели­
чения числа крестцовых
(4— 5);
грудная клетка бочкообразная; по­
звоночный столб е изгибами; гру­
дина образована единой костью,
короткая и широкая, У-образная бо­
роздка между 5 бугорками на ж е ва ­
тельной поверхности коренных зубов
(«узор дриопитека»), последний (тре­
тий) коренной самый малый по раз­
мерам; присутствует червеобразный
отросток слепой кишки (аппендикс);
большое число извилин на полуша­
р и я х головного мозга; полное пере­
крытие мозжечка полушариями го­
ловного мозга, заметное развитие
лобных долей полушарий головного
мозга; присутствие четырех основ
ных групп крови (А, В , 0, А В ) ; моче­
вая кислота не превращается в аллантоин, а выделяется с мочой;
сравнительно крупные общие разме­
ры тела; присутствие хорошо разви­
той подвижной верхней губы; разви­
тие мимической мускулатуры; значи­
тельное пореденне волосяного по­
крова
В и д Ч ей и во к
Объем мозга выше 900 см3 при
очень сложном строении, прямо­
хождение и связанные с ним
признаки строения скелета (см.
7.1.3.2); резкие изгибы позвоноч­
ника как рессоры; развитие седа­
лищных и икроножных мыши;
положение черепа; относительно
длинный и очень толстый I палец
на руках; некоторые изменения в
топографии и строении гортани
(связанные со способностью про
износить членораздельные зву­
к и ); резкое сокращение лицевого
отдела черепа и увеличение моз­
гового; малый размер клыков;
отсутствие гребней на черепе;
развитие подбородочного выстхна; специфическое распределе­
ние волос на теле; сильное разви­
тие эротических зон на теле (гу­
бы, грудь, ягодицы), на верхних
предкоренных зубах часто два
корня
одного другому. Особенность человеческого биологического заклю ча­
ется в т о м , что оно проЯВлме 1 си в условиях действии законов высшей,
социальной ф ормы движения м атерин Г В философии это называется
действием в сн ятом виде!
Биологические процессы с необходимостью происходят в организме
человека, и и м принадлежит фундаментальная роль в определении
важнейших сторон жизнеобеспечения и развития. Однако в популяциях
людей эти процессы не приводят к результатам , о б ы чн ы м для
остального мира ж и вы х сущ еств. В качестве примера рассмотрим
процесс эволюции, котором у в конечном итоге подчинены механизмы,
функционирующие на всех основных уровнях организации жизни —
молекулярно-генетическом, клеточном, онтогенетическом и др. Генофонды популяций людей вп ло ть до настоящего времени исп ы т и н я т т
давление мутаций, комбинативной изменчивости, избирательного
отбора. _ О б этом ш ла речь в предыдущих разделах. Вм есте с те м
благодаря действию социальных факторов естественный о гбор утр ати л
функци ю видообразования^ Э т о делает невозм ож ным достижение
закономерного биологического результата — появление новы х видов
рода Н о т о . Одно из необычных последствий действия элементарных
эволюционных факторов в таких условиях заклю чается в выраженном
наследственном разнообразии людей, которое в т аких масш т абах среди
животны х не наблюдается.
Приобретение социальной сущности и сохранение биологических
механизмов жизнеобеспечения изменило процесс инливилуягшнпгп
развития людец. В онтогенете человека используется информация двух
вндов^ П ерв ы й вид представляет собой биологически целесообразную
информацию, которая отбиралась, и сохранялась в процессе эволюции
предковых форм и зафиксирована в Д Н К клеток в виде генетической
праграммБГ. Благодаря ё!Г1Г индивидуальном развитии складывается
уникальный комплекс структурны х и функциональных признаков,
отличаю щ их человека о т других животных. Возникновение этого
комплекса с л у ж и т необходимой предпосылкой становления человека
как социального сущ ества. Второй внд информации представлен
суммой знаний* ко то ры е создаю тся, сохраняются" и использую тся
поколениями людей в ходе развития общ ества и произво дственнд й
деятельности. _ З т о программа социального наследования,” освоение
которой человеком происходит в процессе его воспитания н обучения.
8.2. ЗА К О Н О М Е РН О С Т И А Н Т Р О П О ГЕН ЕЗА
В процессе антропогенеза на Земле возникла в ы сш а я форма
сущ ествования материи — социальная. О траж ением социальной сущ ио
СТИ л ю пей ГДУЖИТ в ы д а ю т а п г .я .р п п ь Г Г Т И т ц чгш ш рц ^ Тна оЬш ественных отношений и труда. Труд, по замечанию Ф . Энгельса, —
« ... первое, основное условие всей человеческой жизни, и п р и т о м в такой
степени, ч то м ы в известном
пппжны сказать: труд создал
"Самого человека»1 . Предпосылки к возникновении? социальной формы
1 М аркс К . Энгельс Ф
160
Сич
И «л 2-е, г 20. <
•
.
. ».
- т е р т .т и к <
м
•• М М М 1Н11 1*1
Ьмрег Шмлн нч
I
млн
Л» Г
Прямохождение
•>где пенис встни
ИсПОЛЫОИПНМГ рдзщичных
Прсгоминидн •»*
Г и ш и и 1т т
«V • «И *
•
Рнмпнтск
М^нсе 1 9
Индия, Африки
До» грали" • -
1>'-*о 500
Африка
СОМИНИд
Асия
{*)
НреДМеЮ15
. М1Н
И
ЛСI
Ранние
фм>мы
2 ,6м лн мет, рас­
цвет
6Г*‘—
100 ты с **«•
Ранние формы
1 5 млн
ч , р<с
цвет
250 <10
гыс ■
“* *
1 Т О В .1 1 Н И С м | 1 * 1ИЙ
Поддсрж • ние 01 ня
Речи {примитивная, состоя­
т с я из отдельных выкрн
ков) Простые формы кол
лсктивпой
деятельности
Добывание О! ня
Г ***жныС формы коллектив­
ной ч чтелыюсти {сгонная
Архантроиы
Па ••чанпропы
Человек ме)лый
Ч си'чек прямо
ходящий (пи­
текантропы)
750
КЮ -И0О
Африке
Африки, * "-|дн •»
и
Центрильн-<н
Европа, Поемочи **п
Асия Иидоиет'1’
Н«.андерч-1ец
До 1500
Европа,
Асия
Афрнь
Кроманьонец
Око-'О 1400
Европа, Азия, Аф
рнка, Авст])д.шя
Америка
Всесветное
0X01*1
Работа о ближних
Речь (продвинутая
форма
типа лалий ( печча)
М№ № 40 тыс
сет
'Аенее 10 тыс лет
| Неоанчрены
I Настоящая речь
| Мыш пение4
I Искчсство
Расвнтие Сельского чозяйI сгва, промышленного про|. н^водства, техники, науки
Око со 1400
движения м атерии склады вались в ходе биологической эволюции в связи
сопределенны м иусловиям и обитания. М ногие сущ ественные м ом енты
процесса становления человека о ста ю тся предметом дискуссий. Общ ая
схема антропогенеза представляется следующим образом (табл . 27).
Предком гоминнд б ы л наземный примат, ведущий наземный образ
жнзни и перемещавшийся по земле и ветвям деревьев на четырех конеч­
ностях. П ри хождении он опирался на фаланги согнутых пальцев, что
позволило переносить в передних лапах различные предметы.
Н ачальн ы й этап гоминизаиии заклю чается в освоении прямохождения,
чему способствовали использование внешних предметов для добывания
пиши, всеядность, необходимость расш ирить крутозор на основе
зрительной ориентировки. П рямохождение н простейшая орудийная
деятельность с п ом ощ ью случайных предметов были освоены, повидимому, на п р е г о м н н " и д н о й
стадии"
а н т р о п ог е н е з аТ Согласно арчеи'Ю гическим данным, изготовление простей^
т и х каменных орудий стало возм о ж н ы м по достижении определенного
уровня развития головного мозга. Австралопитеки и Человек умелый
(Н о т о НаЫНк) сосуществовали на протяжении многих сотен ты сячеле­
тий, однако то лько последний б ы л способен и зго то вл ять орудия.
Проводя аналогию с процессом онтогенеза современного человека,
антропологи предполагают, что освоение речи не м огло произойти
прежде, чем объем мозга достигнет 750— 800 с м 3 . Укорочение челюсти
и расширение альвеолярного отростка у пекинского питекантропа
допускает относительно б о л ьш у ю подвижность язы ка, облегчает
опускание его корня в гортань. О бъем мозга около 1000 с м 3 в сочетании
с указанны м и морфологическими особенностями дает возм о ж н ость
датировать
появление
примитивной
речи
стадией
арх а н т р о п о в . _
Р ечь и м ыш ление развиваю тся единым комплексом, поэтому
единицы речевой деятельности (слово) н м ыш ления (понятие) не­
разрывно связаны. Н а с т а д и и п а л е о а н т р о п о в возникаю т
сло ж ны е Ф о р м ы коллективной деятельности, соверш енствую тся ору­
дия. К этой стадии, по-видимому, гоминиды приобретаю т способность
воспроизводить форму орудия в уме, д о б ы ва ть огонь. О б ъем мозга
палеоантропов сравним с объемом мозга современного человека,
однако их речь примитивна, интеллектуальны е функции ограниченны.
Примерно 200— 50 ты сяч л ет назад палеоантропы у т ратили
положение преобладающей группы гоминид н процесс антропогенеза
вступил в сталию н е о а н т р о п е в: Т акие качества последних, как
социаОильность, развитая речь, интеллект, коллективная трудовая
деятельность, привели в процессе дальнейшего развития к возникнове­
нию человеческого общества. Важнейш ими этапами развития этого
общ ества яв л я ю т ся неолитическая н научно-техническая революции.
В результате первой из них человек благодаря окультуриванию
растений и одомашниванию ж и во тн ы х вы ш ел из полной зависимости
от сил природы, в результате второй получил мощнейшие источники
энергии и технические возм ож ности активного изменения природы в
глобальном масш табе.
В последние годы б ы стр ы м и тем пам и ш ло накопление археологиче-
Рнс. 155. < кем я эволюции юминнп
йы
грал<и иН'ьивье. 6
^рхаптропы, в
ского материала, которы й свидетельствовал о разнообразии вариантов
основных предковых форм гоминид, различающ ихся по морфологиче­
ским показателям . Т ак, известны массивный н грацильный типы
австралопитеков. Полиморфна группа питекантропов. Среди палео­
антропов вы д ел яю т более грацильных, так называем ых ранних, и
физически более м ощ ных поздних неандертальцев.
Е с т ь основания предполагать, что в отдельные периоды на планете
сосуществовали австралопитеки, Человек ум елый, архантропы. М о ж н о
заклю чи ть, ч то на протяжении больш ей части антропогенеза развитие
гоминид ш ло в каждой линии по нескольким направлениям (рис. 155).
В пределах каждого направления возникали разновидности, которые
изменялись под влиянием разных факторов естественного отбора.
363
В результате отдельные формы вымирали, другие ассимилировались.
В целом отбор доминировал в пользу образа жизни, основанного на
применении орудий, о чем свидетельствуют некоторые морфологиче­
ские признаки современного человека, стабилизировавшиеся на уровне
неоантропов. Так, пропорции кисти руки, сильно развитый и мускули­
стый большой палец, особенности анатомии головного мозга формиро­
вались, по-видимому, под действием отбора, направленного на умение
применять орудия, способность к ручному труду, социабильность.
Морфофизиологические характеристики человека не изменились со
времени стадии неоантропов. Предполагают, что на этом рубеже
благодаря включению мощных факторов социального развития есте­
ственный отбор утратил доминирующую роль и биологическая
эволюция уступила место социальной.
8.3. В И Д О В О Е Е Д И Н С Т В О Ч Е Л О В Е К А .
П О Н Я Т И Е О РА С А Х
Современное человечество п р и н а д л е ж и т
к
одному
виду
Н о то 5ар1еп5. Э то доказывается рождением плодовитого и
полноценного потомства в скрещиваниях между представителями резко
различающихся по некоторым признакам этногеографических групп
населения. Видовое единство людей основывается на воспроизведении
трех главных признаков рода Н ото — выпрямленного положения тела.
хватательного типа верхних конечностей, развитой речевой функции и
мышления. Названным признакам принадлежит ведущая роль в обеспе­
чении выживания и развития всех гоминид. Значительным консерва­
тизмом обладают особенности строения опорно-двигательного аппара­
та и головного мозга, от которых зависит прямохождение, орудийная
деятельность, социабильность. Представители разных этногеографиче­
ских групп характеризуются одинаковым интеллектуальным потенциа­
лом.
Вместе
с
тем
Человек
разумный — это
резко
политипический
вид, что проявляется в наличии трех
«больших» рас людей и некоторого числа более мелких групп,
различающихся главным образом комплексом внешних признаков.
Выделяют е в р о п е о и д н у ю (евразийскую), а в с т р а л о ­
негроидную
(экваториальную)
и
монголоидную
(азиатско-американскую) «большие» расы (рис. 156, 157).
Европеоиды имеют светлую или смуглую кожу, прямые или
волнистые волосы, развитый волосяной покров на лице, узкий
выступающий нос, тонкие губы. Монголоиды отличаются светлой или
смуглой дожей, прямыми, нередко жесткими волосами, уплощенным
лицом с выступающими скулами, косым разрезом глаз, выраженным
«третьим веком» (эпикантом), средними показателями ширины носа и
губ. У австрало-негроидов кожные покровы темные, волосы курчавые
шерстистые или волнистые, губы толстые, нос широкий, маловыст^паюший, с поперечным расположением ноздрей. Представители различ­
ных рас отличаются по некоторым физиологическим и биохимическим
признакам. Так, основной обмен у негров и у большей части других на­
родов экваториальной зоны ниже, чем у европейцев. У последних
364
Рис. 156. Схема расовой классификации
современного человека. По окружности
приведены названия больших рас, в сек­
торах — названия малы)( рас; между
большими расами по две переходных.
Рис.
а
г
157. Представители
- запад н оаф ри кански й
ш вед
больших
н е гр , 6, в
бурят
рас.
и ки таец ;
содержание холестерина в плазме крови достигают 4,64 ммоль/л, тогда
как у первых —‘ 3,48 ммоль/л.
Общность основных человеческих признаков и главной линии
исторического развития, полноценность потомства в межрасовых
браках указываю т на то, что разделение на расы относится к
достаточно продвинутым стадиям эволюции гоминид. На основании
сравнительно-биохимических и антропологических данных предполага­
ю т, что первоначально в человечестве выделились монголоидная и
европеоидно-негроидная расы. Позже из последней выделились евра­
зийская и австрало-негроидная. Указанные события имели место, повидимому, на стадии перехода от палеоантропов к неоантропам.
До эпохи великих географических открытий «большие» расы
характеризовались определенным расселением по планете. Представи­
тели монголоидной расы размещались на территории Северной,
Центральной, Восточной и Юго-Восточной Азии, Северной и Южной
Америки, австрало-негроидной — в Старом Свете к югу от тропика
Рака, европеоидной — на территории Европы, Северной Африки,
Передней Азии, Северной Индии. Многие расовые признаки адаптивно
целесообразны в той части Ойкумены, в которой складывались и
365
обитали расы на протяжении многих тысячелетий. К таковым
относятся пигментация кожных покровов и шерстистые волосы негрои­
дов (повышенный уровень солнечной радиации), крупные размеры
носовой полости европеоидов (действие охлажденного в зимний период
воздуха), эпикант, узкая глазная щель, своеобразное отложение
жировой ткани на лице монголоидов (предохранение глаза от ветра,
пыли, отраженного от снега солнечного света и от переохлаждения
тканей лица). Можно предположить, что формирование расовых
комплексов признаков происходило под действием естественного
отбора. Вместе с тем следует избегать упрошенного понимания
адаптивной природы таких комплексов в целом. Некоторые признаки,
входящие в расовый комплекс, могли появиться в силу коррелятивной
изменчивости. Так, главную роль в развитии уплошенности лица
монголоидов играли, по-видимому, первичные изменения жевательного
аппарата и общей конструкции лицевого скелета. В выделении внутри
«больших» рас различных морфологических типов и групп могли играть
роль метисация, длительное размножение в популяции с высокой
степенью генетического родства, а в отношении народностей,
размещавшихся по окраинам Ойкумены, — дрейф генов.
Р а з д е л
II
ЧЕЛОВЕК И БИОСФЕРА
Глава
9
ВВЕД ЕН И Е В У Ч ЕН И Е О БИ О С Ф ЕРЕ
9.1. С О В Р Е М Е Н Н Ы Й К О Н Ц Е П Ц И И Б И О С Ф Е Р Ы
Термин «биосфера» введен австралийским геологом Э . Зюссом в
1875 г. для обозначения особой оболочки Земли, образованной со­
вокупностью живых организмов, что соответствует
биологи­
ческой
концепции биосферы. В указанном смысле названный
термин используется рядом исследователей и в настоящее время.
Представление о широком влиянии жизни на природные процессы
было сформулировано В. В. Докучаевым, который показал зависи­
мость процесса почвообразования не только от климата, но и от
совокупного влияния растительных и животных организмов.
В. И . Вернадский развил это направление и разработал учение о
биосфере как глобальной системе нашей планеты, в которой основной
ход геохимических и энергетических превращений определяется живым
веществом. Он распространил понятие биосферы не только на сами
организмы, ной на среду их обитания, чем придал концепции биосферы
б и о г е о х и ' м и ч е с к и й смысл. Большинство явлений, меняющих
в масштабе геологического времени облик Земли, рассматривались
ранее как чисто физические, химические или физико-химические
(размыв, растворение, осаждение, выветривание пород и т. д.).
В. И . Вернадский создал учение о геологической роли живых
организмов и показал, что деятельность последних представляет собой
важнейший фактор преобразования минеральных оболочек планеты.
С именем В. И . Вернадского связано также
формирование
с о ц и а л ь н о-э к о н о м и ч е с к о й концепции биосферы, отража­
ющей ее превращение на определенном этапе эволюции в ноосферу
(см. главу 10) вследствие деятельности человека, которая приобретает
роль самостоятельной геологической силы. Учитывая системный
принцип организации биосферы, а также то, что в основе ее функциони­
рования лежат круговороты веществ и энергии, современной наукой
сформулированы б и о х и м и ч е с к а я , т е р м о д и н а м и ч е с ­
кая,
биогеоценотическая,
кибернетическая
концепции биосферы.
Б и о с ф е р о й называется оболочка Земли, которая населена и
активно преобразуется живыми существами. Согласно В. И . Вернад­
скому, биосфера — это такая оболочка, в которой существует или
существовала в прошлом жизнь и которая подвергалась или подверга­
ется воздействию живых организмов. Она включает: 1) живое
вещество, образованное совокупностью организмов; 2) биогенное
вещество, которое создается и перерабатывается в процессе жизнедея­
тельности организмов (газы атмосферы, каменный уголь, нефть,
367
сланцы, известняки и др.); 3) косное
вешество, которое образуется без уча­
стия живых организмов (продукты
тектонической деятельности, метеори­
ты ); 4) биокосное вещество, пред­
ставляющее собой совместный резуль­
тат жизнедеятельности организмов и
абиогенных процессов (почвы).
9.2. С Т Р У К Т У Р А И Ф У Н К Ц И И
БИ О СФРРЫ
Биосфера представляет собой мно­
гоуровневую систему, включающую
подсистемы различной степени слож­
ности. Границы биосферы определя­
ю тся областью распространения орга­
низмов в атмосфере, гидросфере и
литосфере (рис. 158). Верхняя граница
биосферы проходит примерно на выв. и. Вернадский (1кбЗ 194Г>)
соте 20 км. Таким образом, живые
организмы расселены в тропосфере и в нижних слоях стратосферы.
Лимитирующим фактором расселения в этой среде является
нарастающая с высотой интенсивность ультрафиолетовой ради­
ации. Практически все живое, проникающее выше озонового
слоя атмосферы, погибает. В гидросферу биосфера проникает
на всю глубину мирового океана, что подтверждается обнаруже­
нием живых организмов, и органических отложений до глуби­
ны 10— 11 км. В литосфере область распространения жизни во многом
определяется уровнем проникновения воды в жидком состоянии —
живые организмы обнаружены до глубины примерно 7,5 км.
Атмосфера. Э т а оболочка состоит в основном из &зо га и кислорода.
В меньших концентрациях она содержит углекисл 1й газ и озон.
Состояние атмосферы оказывает большое влияние на физические,
химические и, особенно, биологические процессы на земной поверхности
и в водной среде. Наибольшее значение для биологических процессов
имеют: кислород атмосферы, используемый для дыхания организмов и
минерализации омертвевшего органического вещества, углекислый газ,
расходуемый при фотосинтезе, а также озон, экранирующий земную
поверхность от жесткого ультрафиолетового излучения. Вне атмосфе­
ры существование живых организмов невозможно. Э то видно на
примере лишенной жизни Луны, у которой нет атмосферы. Историче­
ски развитие атмосферы связано с геохимическими процессами, а также
жизнедеятельностью организмов. Так, азот, углекислый газ, пары воды
образовались в процессе эволюции планеты благодаря в значительной
мере вулканической активности, а кислород — в результате фото­
синтеза.
Гидросфера. Вода является важной составной частью всех
компонентов биосферы и одним из необходимых факторов существова368
10ОО00 м
оО О О О
20000
10000
2000
1000
100-
1000
10000 м
Рис. 158. Область распространения организмов в биосфере.
1
3
у р о в е н ь о з о н о в о г о 1 , ю я , л а д е р ж н в а ю щ е !о ж е с т к о е у л ь т р а ф и о л е т о в о е и з л у ч е н и е , 2
почва; 4
ж и во тн ы е, о б итаю щ и е в пещ ерах, 5
бактер ии в неф тяны х с к в а ж и н а х
гр ани ц а снегов,
ния живых организмов. Основная ее часть (95 % ) заключена в мировом
океане, который занимает примерно 70% поверхности земного шара.
Общая масса океанических вод составляет свыше 1300 млн. км 3. Около
24 млн. км3 воды содержится в ледниках, причем 90% этого объема
приходится на ледяной покров Антарктиды. Столько же воды
содержится под землей. Поверхностные воды озер составляют
369
приблизительно 0,18 млн. км3 (из них половина соленые), а рек —
0,002 млн. км3 . Количество воды в телах живых организмов достигает
примерно 0,001 млн. км3 . Из газов, растворенных в воде, наибольшее
значение имеют кислород и углекислый газ. Количество кислорода в
океанических водах изменяется в широких пределах в зависимости от
температуры и присутствия живых организмов. Концентрация угле­
кислого газа также варьирует, а общее количество его в океане в 60 раз
превышает его количество в атмосфере. Гидросфера формировалась в
связи с развитием литосферы, выделившей за геологическую историю
Земли значительный объем водяного пара и так называемых
ювенильных (подземных магматических) вод.
Литосфера. Основная масса организмов, обитающих в пределах
литосферы, сосредоточена в почвенном слое, глубина которого обычно
не превышает нескольких метров. Почвы, будучи, по терминологии
В. И . Вернадского, биокосным веществом, представлены минеральны­
ми веществами, образующимися при разрушении горных пород, и
органическими веществами — продуктами жизнедеятельности орга­
низмов.
Ж ивые организмы (живое вещество). В настоящее время описано
около 300 тыс. видов растений и более 1,5 млн. видов животных. Из
этого количества 93% представлено сухопутными, а 7 % водными
видами животных. Суммарная биомасса организмов сухопутных видов
образована на 99,2% зелеными растениями (2,4 х 1012 т ) и на 0,8% —
животными и микроорганизмами (0,2 х 10" т). В океане, напротив, на
долю растений приходится 6,3% (0,2 х 10 е т), а на долю животных и
микроорганизмов 93,7% (0,3 х 10,0 т ) совокупной биомассы. Несмотря
на то что океан покрывает немногим более 70% поверхности планеты, в
нем содержится лишь 0,13% биомассы всех живых существ, обитающих
на Земле. Расчеты показывают, что растения составляют около 21%
всех учтенных видов. Однако на их долю приходится более 99%
биомассы, тогда как вклад в биомассу планеты животных, которыми
представлено79% видов, составляет менее 1% . Среди животных 96%
видов приходится на долю беспозвоночных и только 4 % на долю
позвоночных, среди" которых млекопитающие составляют примерно
10%. Приведенные соотношения иллюстрируют фундаментальную
закономерность организации биосферы: в количественном отношении
преобладают формы, достигшие в процессе эволюции относительно
низких степеней морфофизиологического прогресса.
Живое вещество по массе составляет 0,01—0,02% от косного
вещества биосферы, однако играет ведущую роль в биогеохимических
процессах благодаря совершающемуся в живых организмах обмену
веществ. Так как субстраты и энергию, используемые в обмене веществ,
организмы черпают из окружающей среды, они преобразуют ее уже
тем, что живут. Ежегодная продукция живого вещества в биосфере
равняется 232,5 млрд. т сухого органического вещества. За это же время
в масштабе планеты в процессе фотосинтеза синтезируется 46 млрд. т
органического углерода. Для этого требуется, чтобы 170х Ю 9 т угле­
кислого газа прореагировало с 68x10 т воды. Таким образом, в
результате фотосинтеза ежегодно образуется 115x10е т сухого органи­
370
ческого вешества и 123x10® т кислорода. В процесс фотосинтеза
вовлекается также в течение года 6х 10® т азота, 2 х 10® т фосфора и
другие элементы, например калий, кальций, магний, сера, железо.
Приведенные цифры показывают, что живое вешество является
наиболее активным компонентом биосферы. Оно производит гигантс­
кую геохимическую работу, способствуя преобразованию других оболо­
чек Земли в геологическом масштабе времени.
Биотический кру го в о ро т . Главная функция биосферы заключается в.
обеспечении круговоротов химических элементов. Глобальный^ биоти"ческий—круГОВПрот осуществляется при участии всех населяющих
планету организмов. Он заключается в циркуляции веществ между
почвой, атмосферой, гидросферой и живыми организмами. Благодаря
биотическому круговороту возможно длительное существование и раз­
витие жизни при ограниченном запасе доступных
химических
элементов. Используя неорганические вещества, зеленые растения за
счет энергии Солнца создают органическое вещество, которое другими
живыми существами (гетеротрофами-потребителями и деструкторами)
разрушается, с тем чтобы продукты этого разрушения могли бы ть
использованы растениями для новых органических синтезов.
Важная роль в глобальном круговороте веществ принадлежит
циркуляции воды между океаном, атмосферой и верхними слоями
литосферы. Вода испаряется и воздушными течениями переносится на
многие километры. Выпадая на поверхность суши в виде осадков, она
способствует разрушению горных пород, делая их доступными для
растений и микроорганизмов, размывает верхний почвенный слой и
уходит вместе с растворенными в ней химическими соединениями и
взвешенными органическими частицами в океаны и моря. Подсчитано,
что с поверхности Земли за 1 мин испаряется около 1 млрд. т воды (на
образование 1 г водяного пара затрачивается 2,248 кД ж ). Энергия,
затрачиваемая на испарение воды, возвращается в атмосферу (рис. 159).
Циркуляция воды между мировым океаном и сушей представляет собой
важнейшее звено в поддержании жизни на Земле и основное условие
взаимодействия растений и животных с неживой природой. Благодаря
этому процессу происходит постепенное разрушение литосферы,
перенос ее компонентов в глубины морей и океанов.
Н а создание органического вещества расходуется всего 0,1—0,2%
'солнечной энергии, достигающей поверхности планеты. Энергия
биотического круговорота мала по сравнению с энергией, расходуемой в
абиотических геохимических процессах. Благодаря ей осуществляется
значительный объем работы по перемещению химических элементов.
В качестве примеров биотического круговорота рассмотрим
круговороты углерода и азота в биосфере (рис. 160). К р у г о в о р о т
у г л е р о д а начинается с фиксации атмосферной двуокиси углерода в
процессе фотосинтеза. Часть образовавшихся при фотосинтезе углево­
дов используется самими растениями для получения энергии, а часть
потребляется животными. Углекислый газ выделяется в процессе
дыхания растений и животных. Мертвые растения и животные
разлагаю хся, углерод их тканей окисляется и возвращается в атмосфе­
ру. Аналогичный процесс происходит и в океане.
371
Д ис'.ипацир а ю м о в
водорода в ноомо(
1
Рис. 159. Круговорот воды в биосфере.
Рис. 160. Круговорот углерода в биосфере.
1НЫИ
а зо т
Атм осф ер н ая
, ф и кса ц и я
'/Ми."//
О венилн-/^
Р о м ^|Ш П е н
„и
ю ван ны й* 1 о
IV а з о т
«Гъ
Белон
* и кса
*^ии
Д К
’ВДкДО
Изверженные"
породы
Биологичесная
Ч фиксация
Расти тельн ы е
и ж ивотны е
отходы м ер твы е
о р ган и зм ы
Д енитри
Восстановление
I ф инация
н и т р а то в
Занись
азо та у
А м миак
Н итриты
Н и т р а ты .
В п о д з е м н ы е воды
Рис. 161. Круговорот азота в био­
сфере.
Рис. 162. Темпы циркуляции
ществ в биосфере.
ве­
Круговорот
азота
также охватывает все области
биосферы (рис. 161). Х о тя его запасы в атмосфере практически
неисчерпаемы, высшие растения могут использовать азот только после
соединения его с водородом или кислородом. Исключительно важную
роль е этом процессе играют азотфиксирующие бактерии. При распаде
белков этих микроорганизмов азот снова возвращается в атмосферу.
Показателем масштабов биотического круговорота служат темпы
оборота углекислого газа, кислорода и воды. И з рис. 162 видно, что
весь кислород атмосферы проходит через организмы примерно за
2 тыс. лет, углекислый газ — за 300 лет, а вода полностью разлагается
и восстанавливается в биотическом круговороте за 2000 лет.
Благодаря биотическому круговороту биосфере присущи опреде­
ленные геохимические функции: газовая — биогенная миграция газов в
результате фотосинтеза и азотфиксации; концентрационная — аккуму­
ляция живыми организмами химических элементов, рассеянных во
внешней среде; окислительно-восстановительная — превращение ве­
373
ществ, содержащих атомы с переменной валентностью (например,
железо, марганец); биохимическая — процессы, протекающие в живых
организмах.
Стабильность биосферы. Биосфера представляет собой сложную
экологическую систему, работающую в стационарном режиме.
Стабильность биосферы обусловлена тем, что результаты активности
трех групп организмов, выполняющих разные функции в биотическом
круговороте — продуценты (автотрофы), потребители (гетеротрофы) и
деструкторы (минерализующие органические остатки) — взаимоуравновешиваются. Гомеостатическое состояние биосферы не исключа­
ет способности ее к эволюции.
9.3. Э В О Л Ю Ц И Я Б И О С Ф Е Р Ы
Эволюция биосферы на протяжении большей части ее истории
осуществлялась под влиянием двух главных факторов: естественных
геологических и климатических изменений на планете и изменений
видового состава и количества живых существ в процессе биологической
эволюции. На современном этапе в третичном периоде к ним
присоединился третий фактор — развивающееся человеческое обще­
ство.
Этапы возникновения жизни, пути и механизмы эволюционного
процесса рассмотрены выше (см. главу 7). Ж изнь зародилась на Земле
свыше 3 млрд. лет назад. Первыми живыми существами были
анаэробы, которые получали энергию путем брожения. Так как
брожение представляет собой относительно малопродуктивный способ
энергообеспечения, примитивная жизнь не могла эволюционировать
далее одноклеточной формы организации. Питание таких примитивных
организмов зависело от опускавшихся на дно водоемов органических
веществ, синтезируемых в поверхностных слоях воды абиогенным
способом.
Недостаток органических веществ создал давление отбора,
приведшее к возникновению фотосинтеза. Прогрессивное увеличение
кислорода в воде за счет жизнедеятельности фотосинтезирующих
организмов и его диффузия в атмосферу вызвали изменения в химиче­
ском составе оболочек Земли, и прежде всего атмосферы, что, в свою
очередь, сделало возможным быстрое распространение жизни по
планете и развитие более сложно организованных живых форм. П о мере
увеличения содержания кислорода в атмосфере формируется доста­
точно мощный слой озона, который защищает поверхность Земли от
проникновения жесткого ультрафиолетового излечения. В таких усло­
виях жизнь смогла продвинуться к поверхности моря. Развитие
механизма аэробного дыхания сделало возможным появление мно­
гоклеточных организмов. Примечательно, что первые такие организмы
появились после того, как концентрация кислорода в атмосфере
планеты достигла примерно 3 % , что произошло около 600 млн. лет
назад (начало кембрия).
Благодаря способности фотосинтезирующих морских организмов
продуцировать такое количество кислорода, которое превышало
374
потребности в нем обитателей планеты того времени, с 1 ало
возможным на основе возникновения в процессе эволюции организмов
более высокого уровня структурно-физиологической организации ши
рокое расселение и ^проникновение жизни в различные среды обитания,
причем за сравнительно короткое время. В течение палеозойской эры
жизнь не только заполнила все моря, но и вышла на сушу. Развитие зе
леных растений обеспечило образование больших количеств кислорода
и органических веществ, что создавало благоприятные условия для
последующей прогрессивной эволюции.
В середине палеозоя темпы потребления кислорода живыми
организмами и расход его в абиотических процессах и темпы ею
■образования сравнялись. Содержание кислорода в атмосфере, начиная с
этого периода истории Земли, стабилизировалось на уровне примерно
20 %.
С появлением человеческого общества в развитии биосферы
намечается переход от биогенеза, обусловливаемого факторами биоло­
гической эволюции, к ноогенезу — развитию под влиянием разумной
созидательной деятельности человечества.
Глава
10
У Ч Е Н И Е О Н О О С Ф ЕРЕ
10.1. Б И О Г Е Н Е З И Н О О Г Е Н Е З
Эволюция органического мира прошла несколько этапов. Первый из
них — возникновение первичной биосферы с биотическим круговоро­
том, второй — усложнение структуры биотического компонента
биосферы в результате появления многоклеточных организмов. Эти два
этапа-, осуществлявшиеся в связи с чисто биологическими закономер­
ностями жизнедеятельности и развития, могут бы ть объединены
в п е р и 'о д б и о г е н е з а . Третий этап связан с возникновением
человеческого общества. Разумная по своим намерениям деятельность
людей в масштабе биосферы способствует превращению последней в
ноосферу. На рассматриваемом этапе эволюция происходит под
определяющим воздействием человеческого сознания в процессе
производственной (трудовой) деятельности людей, что свойственно
периоду ноогенеза.
Понятие н о о с ф е р ы было введено в науку французским фило­
софом Э . Леруа (1927). Ноосферой Э . Леруа назвал оболочку Земли,
включающую человеческое общество с его индустрией, языком и
прочими атрибутами разумной деятельности.
Высказав правильную идею о сознательной человеческой деятельно­
сти как факторе, преобразующем биосферу, Э . Леруа и некоторые его
последователи дали ей идеалистическую трактовку. Ноосфера, по их
мнению, представляет собой «...мыслящий пласт», который, заро­
дившись в конце третичного периода, разворачивается с тех пор над
миром растений и животных вне биосферы и над ней.
В противоположность приведенной трактовке В. И. Вернадский
развивал материалистическое представление о ноосфере, понимая под
ноосферой не нечто внешнее по отношению к биосфере, а новый этап в
ее развитии, заключающийся в разумном регулировании отношейий
человека и природы.
В
непосредственной связи с революционными социальными
преобразованиями, со сменой капиталистических отношений социали­
стическими возникает реальная возможность
научно
обос­
нованного природопользования.
Науку управления взаимоотношениями между человеческим обще­
ством и природой можно назвать н о о г е н и к о й . Основная цель
ноогеники — планирование настоящего во имя лучшего будущего, а ее
главная задача — исправление нарушений в отношениях человека и
природы, вызванных прогрессом техники.
Помимо охранных функций, ноогеника должна способствовать
увеличению многообразия форм жизни путем создания новых видов
растений, животных и микроорганизмов. Э ти новые виды призваны не
только служить источником пищи, кислорода, сырья для про­
мышленности, но и помогать человеку, осуществляя буферные
функции, бороться с вредными побочными результатами технического
прогресса, способствовать еще более активному освоению неживой
природы, сопровождать человека в космических полетах.
Таким образом, ноогеника не ставит целью достижение какого-то
постоянного равновесия между человеком и природой, которое в
принципе неосуществимо. Ее задача — сознательное управление
эволюцией биосферы, включающей человеческое общество в качестве
ведущего компонента.
Для рационализации отношений человека и биосферы особенно
много делается в С С С Р. Только за последние годы Верховным Советом
С С С Р приняты такие важные законы, как «Основы земельного
законодательства Союза С С Р и союзных республик», «Основы
законодательства Союза С С Р и союзных республик о здравоохране­
нии», «Основы водного законодательства Союза С С Р и союзных
республик». В союзных республиках приняты специальные законы об
охране природы. Энергичные меры предпринимаются для предотвра­
щения загрязнения Каспийского моря, Байкала, бассейнов Волги и
Урала. Н а IV сессии Верховного Совета С С С Р восьмого созыва (1972)
было принято историческое Постановление «О мерах по дальнейшему
улучшению охраны природы и рациональному использованию при­
родных ресурсов». В этом Постановлении подчеркивается, что дости­
жения научно-технической революции и мощная база нашей индустрии
позволяют в условиях социалистического хозяйствования разумно
пользоваться всеми природными богатствами, успешно решать истори­
чески важную задачу — нейтрализовать вредные для природы и челове­
ка побочные явления хозяйственной деятельности.
В дополнение к этим документам в 1978 г. было принято Постанов­
ление Ц К К П С С и Совета Министров С С С Р «О дополнительных мерах
по усилению охраны природы и улучшению использования природных
ресурсов».
376
10.2. П У Т И В О З Д Е Й С Т В И Я Ч Е Л О В Е Ч Е С Т В А
ЭКО Л О ГИ ЧЕС КИ Й КРИ ЗИ С
НА П Р И Р О Д У .
Качественно новый этап в развитии биосферы наступил с возникно­
вением человеческого общества. Сначала интенсивность воздействия
людей на среду обитания не отличалась от воздействия других
организмов. Получая из окружающей среды средства к существованию
в таком количестве, которое полностью восстанавливалось за счет
естественных процессов биотического круговорота, люди возвращали в
биосферу то, что использовалось другими организмами для их
жизнедеятельности. Универсальная способность микроорганизмов
разрушать органическое вещество, а растений превращать минеральные
вещества в органические обеспечивала включение продуктов хозяй­
ственной деятельности людей в биотический круговорот.
В настоящее время человек извлекает из биосферы сырье в
значительном и все возрастающем количестве, а современные
промышленность и сельское хозяйство производят или применяют
вещества, не используемые другими видами организмов, а нередко к
тому же и ядовитые. В результате этого биотический круговорот
становится незамкнутым. Вода, атмосфера, почвы загрязняются
отходами производства, вырубаются леса, истребляются дикие
животные, разрушаются природные биогеоценозы.
Возможность нежелательных последствий неконтролируемой чело­
веческой деятельности осознавалась естествоиспытателями уже в конце
X V III
(Ж . Л . Л . Бюффон) — первой половине
X IX (Ж . Б. Л а­
марк) века. В настоящее время человечество стоит перед перспективой
э к о л о г и ч е с к о г о к р и з и с а , т. е. такого состояния среды
обитания, которое вследствие произошедших в ней изменений
оказывается непригодной для жизни людей. Ожидаемый кризис по
своему происхождению является антропогенным, так как к нему ведут
изменения в природе Земли, развивающиеся в связи с воздействием на
нее человечества.
По своим последствиям воздействия человеческого общества на
среду обитания могут бы ть положительными и отрицательными.
Именно последние привлекают к себе особое внимание. Основные пути
воздействия людей на природу заключаются в расходовании есте­
ственных богатств в виде минерального сырья, почв, водных ресурсов,
загрязнения среды, истреблении видов и разрушении биогеоце­
нозов.
Естественные богатства планеты делятся на невоспблняемые и
восполняемые. К первым, например, относятся полезные ископаемые,
запасы которых ограниченны.
Тенденцию в изменениях восполняемых природных ресурсов можно
проследить на примере леса. В настоящее время лесом покрыта
примерно треть суши (без Антарктиды), тогда как в доисторические
времена им было занято не менее 70% суши. Особенно пострадал лес в
районах древних цивилизаций. Оголение горных склонов Ливана
началось 5000 лет назад, когда по приказу царя Соломона 80 000
египетских дровосеков вырубили для строительства дворца и храмов
377
роши ливанских кедров на значительной территории. Густы е леса
Далмации начали интенсивно уничтожать при создании римского
флота, а затем при строительстве Венеции. На большей части
территории Китая и Индии леса были почти полностью вырублены уже
в прошлом тысячелетии.
Уничтожение лесов прежде всего резко нарушает водный режим
планеты. Мелеют реки, их дно покрывается илом; что приводит, в свою
очередь, к уничтожению нерестилищ и сокращению численности рыб.
Уменьшаются запасы грунтовых вод, создается недостаток влаги в
почве. Талая вода и дождевые потоки смывают, а ветры, не сдерживае­
мые лесной преградой, выветривают верхний почвенный слой.
В результате возникает эрозия почвы. Древесина, ветви, кора,
подстилка аккумулируют минеральные элементы питания растений.
Уничтожение лесов ведет к вымыванию этих элементов из почв и,
следовательно, падению ее плодородия. С вырубкой лесов гибнут
населяющие их птицы, звери, насекомые-энтомофаги. В результате
беспрепятственно размножаются вредители сельскохозяйственных
культур. Лес очищает воздух от ядовитых загрязнений, в частности, он
задерживает радиоактивные осадки и препятствует их дальнейшему
распространению, т. е. вырубка лесов устраняет важный компонент
самоочищения воздуха. Наконец, уничтожение лесов на склонах гор
является существенной причиной образований оврагов и селевых
потоков.
Благодаря нерациональному землепользованию человечество поте­
ряло вследствие эрозии почв обширные территории, ставшие практиче­
ски не пригодными для земледелия. Так, за период, равный примерно
150 годам, в С Ш А эрозия привела к резкому снижению плодородия почв
на территории, равной 120 млн. га.
Промышленные отходы, пестициды, применяемые для борьбы с
вредителями сельскохозяйственных культур, радиоактивные вещества,
образуемые при испытании оружия массового уничтожения, загрязня­
ю т природную среду. Так, только автомобили в Париже за год
выбрасывают в атмосферу около 50 млн. м3 угарного газа, кроме того,
каждый автомобиль ежегодно выделяет около 1 кг свинца.. Обнаруже­
но, что в организме людей, проживающих вблизи крупных магистра­
лей, содержание свинца повышено. Из-за бесконтрольного использова­
ния количество известного инсектицида Д Д Т, находящегося в распы­
ленном состоянии на обширных территориях планеты и оказывающего
неблагоприятное действие на живые организмы, достигло 1 млн. т.
Троекратная обработка инсектицидом, сходным с ДДТ, района
живописного озера Клири в С Ш А привела к почти полному
уничтожению популяции гагар, достигавшей 1000 пар. В 1961 г. в орга­
низме жителя С Ш А обнаруживалось примерно 925 мг хлорорганических
соединений.
Считая с 1600 г. человеком было истреблено более 160 видов и
подвидов птиц и не менее 100 видов млекопитающих. В настояшее
время около 600 видов позвоночных животных находятся на грани
полного истребления. К ним относятся киты, австралийские сумчатые
(кенгуру), крокодилы, бегемоты, носороги, ряд крупных хищников.
178
Отдельные виды животных исчезают не только в результате их
непосредственного истребления человеком. Между естественными и
искусственными биоценозами все время идет борьба за территорию. Но
человеческий труд оказывается фактором настолько мощным, что
искусственные биоценозы, сами по себе малоустойчивые, тем не менее
теснят биоценозы естественные.
Деятельность человека изменяет структуру земной поверхности,
отчуждая под сельскохозяйственные угодья, строительство населенных
пунктов, коммуникаций, водохранилищ территорию, занимаемую
природными биогеоценозами. К настоящему времени указанным обра­
зом преобразовано около 20% суши.
К числу отрицательных влияний относится нерегулируемый
промысел рыбы, млекопитающих, беспозвоночных, водорослей, изме­
нение химического состава вод, воздуха, почвы в результате сброса
отходов промышленности, транспорта и сельскохозяйственного про­
изводства.
Положительное влияние выражается в выведении новых пород
домашних животных и сортов сельскохозяйственных растений, созда­
нии культурных биогеоценозов, а также в выведении штаммов
полезных микроорганизмов как основы микробиологической промыш­
ленности, развитие прудового рыбного хозяйства, интродукции по­
лезных видов в новых условиях обитания.
Прогнозы будущего человечества с учетом экологических проблем,
стоящих перед ним, представляют непосредственный интерес для всего
населения Планеты. По мнению экспертов, экологическая ситуация,
складывающаяся на Земле, действительно таит в себе опасность
серьезных и, возможно, необратимых нарушений биосферы в ток*
случае, если деятельность человечества не приобретет планомерный,
согласующийся с законами существования и развития биосферы
характер. Вместе с тем расчеты показывают, что человеческое
общество не использует значительные резервы биосферы.
Одной из наиболее острых проблем современности является
проблема быстрого роста населения Земли. Ежегодный прирост
населения в абсолютном исчислении достигает 60—70 млн. человек или
примерно 2 % , что составит к 2000 году численность в пределах от 4,9 до
6,9 млрд. человек. Площадь поверхности суши на планете равна
1,5 х 1014 м2 , что достаточно для размещения 15— 20 млрд. человек со
средней плотностью 300—400 человек на 1 км 2, имеющей место в
настоящее время в Бельгии, Нидерландах, Японии.
Растущее население Земли должно бы ть обеспечено пишей.
Известно, что производство продовольствия на душу населения растет
медленнее, чем производство энергии, одежды, различных материалов.
Многие миллионы людей в слаборазвитых странах испытывают
острую нехватку продовольствия. Вместе с тем из всей территории
суши, пригодной для земледелия, в среднем по земному шару под
селькохозяйственные угодья занято лишь 41 % . При этом на используе­
мой территории, по мнению разных экспертов, получают от 3—4 до
30% от возможного при современном уровне развития агротехники
количества продуктов. Причины этого отчасти заключаются в недоста­
379
точной энерговооруженности сельского хозяйства. Так, в Японии,
получая урожай в 5 раз больший, чем в Индии (с 1 га сельскохозяйствен­
ных угодий), затрачивают на 1 га в 20 раз больше электроэнергии и в
20— 30.раз больше удобрений и пестицидов.
Уж е сейчас 30% металлоизделий изготовляется из вторичного
сырья. При существующей технологии из месторождений нефти
излекается лишь 30— 50% запасов. Выход полезных ископаемых, таким
образом, может бы ть увеличен путем разработки прогрессивных
способов добычи. Около 95% энергии получают в настоящее время за
счет сжигания ископаемого топлива, 3—4 % за счет энергии речного
стока и только 1— 2 % за счет атомного горючего. Использование
атомной энергии в мирных целях решает проблему энергетического
кризиса.
Р а з д е л
III
БИОЛОГИЧЕСКИЕ АСПЕКТЫ
ЭКОЛОГИИ Ч Е Л О В Е К А
Глава
11
Э Л Е М Е Н Т Ы О Б Щ ЕЙ Э К О Л О Г И И
11.1. О С Н О В Н Ы Е П О Н Я Т И Я Э К О Л О Г И И
Ж ивы е существа, населяющие территории с разнообразными
условиями обитания, испытываю т на себе влияние последних и сами
оказывают действие на окружающую среду.| Закономерности взаимо­
отношений организмов и среды их обитания, законы развития и суще­
ствования биогеоценозов, представляющих собой комплексы взаи­
модействующих живых " и неживых'"1ГоШонен^6В' ’Б
ле пенны х
участках биосферы, изучаются специальной биологической няукой
у
к о'тго г и е ятТ
"
Экологические закономерности проявляются на уровне осо­
би, популяции особей, биоценоза (сообщества), биогеоценоза.
Б иоценозом
(сообществом организмов) называется ппостпанственно ограниченная ассоциация взаимодействующих растений и
ЖИВОТНЫХ. В КОТОРОЙ Пам1ПГшт^ТТГТГ]Т|^^
" " " ф»зичщщй
фактор. Предметом экологии, таким образом, являются физиология и
поведение отдельных организмов в естественных условиях обитания
(а уТ"0 э к о л о г и я), рождаемость, смертность, миграции, внутри
видовые отношения (дГИ н а М ТПГа п огГ у л я ц и й), межвидовые
отношения, потоки энергии и круговороты веществ
(с и н-
экология).
К основным методам экологии относятся полевые наблюдения,
эксперименты в природных условиях, моделирование процессов и
ситуаций, встречающихся в популяциях и биоценозах, с помощью
вычислительной техники.
С р е д а — это вся совокупность элементов, которые действуют на
осооь в месте ее обитания.\ Элемент среды, способный оказывать
прямое влияние на живой организм хотя бы на одной из стадий инди­
видуального развития, называется э к о л о г и ч е с к и м
фак­
тором.
В соответствии с распространенной и удобной клас­
сификацией экологические факторы делят на б и о т и ч е с к и е и
а б и о т и ч е с к и е , хотя указанное деление до некоторой степени
условно. Абиотический фактор температура может, например, регули­
роваться изменением состояния популяции организмов. Так, при
снижении температуры воздуха ниже 13°С интенсифицируется двига­
тельная’ активность пчел, что повышает температуру в улье до
25— 30°С. Учиты вая социальную сущность человека, проявляющуюся в
его активном отношении к природе, целесообразно выделение также
антропогенных
экологических
факторов.
По
381
мере роста народонаселения и технической вооруженности человече­
ства удельный вес антропогенных экологических факторов неуклонно
возрастает.
Согласно другой классификации различают п е р в и ч н ы е
и
вторичные периодические и непериодические
экологические факторы. С действием первичных факторов жизнь
столкнулась на ранних стадиях эволюции. К ним относятся температу­
ра, изменение положения Земли по отношению к Солнцу. Благодаря им
в эволюции возникла суточная, сезонная, годичная периодичность
многих биологических процессов. Вторичные периодические факторы
являются производными первичных факторов. Например, уровень
влажности зависит от температуры, поэтому в холодных областях
планеты атмосфера содержит меньше водяных паров. Непериодические
факторы действуют на организм или популяцию эпизодически,
внезапно. К ним относят стихийные силы природы — извержение
вулкана, ураган, удар молнии, наводнение, а также хищник,
настигающий жертву, и охотник, поражающий цель. Благодаря
многообразию экологических факторов наблюдается закономерное
расселение видов по планете. Колебания интенсивности их действия
проявляются в исчезновении некоторых видов с определенные
территорий, изменении плотности популяций, показателей рождаемо­
сти, смертности. Под влиянием экологических факторов в эволюции
сложились такие адаптивные модификации, как зимняя или летняя
спячка, диапауза.
Лю бая особь, популяция, сообщество испытывают одновременное
воздействие многих факторов, но лишь некоторые из них являются
жизненно
важными.
Такие
факторы
называются
лим и­
тирующими,
и их отсутствие или наличие концентрации
ниже и выше критических уровней делает невозможным освоение среды
организмами определенного вида. Н а рис. 163 представлены типы
растительности в зависимости от структуры почвы и особенностей
климата, выполняющих функцию лимитирующих факторов. Благодаря
наличию лимитирующих экологических факторов для каждого биоло­
гического вида существуют о п т и м у м
и пределы
вы­
н о с л и в о с т и . Так, устрицы наилучшим образом развиваются
в воде с содержанием солей 1,5— 1,8%. При снижении концентрации
солей до 1,0% более 90% личинок погибает в течение двух недель, а при
концентрации 0,25% все поголовье их гибнет за одну неделю.
Повышение концентрации соли по сравнению с оптимальной величиной
также оказывает неблагоприятное действие на устриц. В общем виде
зависимость выживаемости организмов определенного вида от интен­
сивности лимитирующего экологического фактора представлена графи­
чески на рис. 164. Взаимодействие нескольких экологических факторов
усложняет картину. Так, некоторые виды тропических орхидей в
природе при относительно высокой температуре воздуха растут только
в тени. При искусственном понижении температуры окружающего
воздуха они прекрасно развиваются в условиях прямой инсоляции.
Способность вида осваивать разные среды обитания выражается
величиной э к о л о г и ч е с к о й в а л е н т н о с т и . Виды с малой
382
400
Годовые
осадки
см
П ределы
вы н о сл и в о сти
Эн ологичесн ии
оптимум
300
200
-
С р е д н я я тем пература
С
М инимум
ф антора
М ак си м у м
Рис. 163. Типы растительности в зависимости от климата.
Рис. 164. Выживаемость вида в зависимости от интенсивности действия экологиче­
ского фактора.
экологической валентностью называются с т е н о т о п н ы м и ,
с
большой — э в р и т о п н ы м и .
Эвритропные виды могут быть
представлены несколькими
э к о т и п а м и — разновидностями,
приспособленными к выживанию в средах, различающихся по некото­
рым факторам. Так, сложноцветное растение тысячелистник (АсЫНеа
тШ е& И и т) образует равнинные и горные экотипы. При выращивании
горного экотипа в равнинных условиях растения сохраняют присущие
им особенности на протяжении ряда поколений.
11.2. К О Н Ц Е П Ц И Я Б И О Г Е О Ц Е Н О З А
Всю полноту взаимодействий и взаимозависимости живых существ
и элементов неживой природы в области распространения жизни
отражает концепция биогеоценоза (В . Н . Сукачев).
К понятию биогеоценоза близко по смыслу понятие экосистемы,
которое введено в науку английским ботаником А . Тенсли в 1935 г.
В отличие от биогеоценозов, границы которых задаются рамками
растительных сообществ (фитоценозов), экосистемы не имеют опреде­
ленного объема и могут охватывать пространства разной протяженно­
сти — от капли воды или аквариума до океана или всей поверхности
планеты.
Б и о г е о ц е н о з — это динамическое и устойчивое сообщество
растений,-животных и микроорганизмов, находящееся в постоянном
взаимодействии и непосредственном контакте с компонентами атмос­
феры, гидросферы и литосферы. Биогеоценоз состоит из биотической
(биоценоз) и абиотической (экотоп) части, которые связаны непре­
рывным обменом веществом и представляет собой энергетически и
183
вещественно открытую си­
стему (рис. 165). В него
поступают энергия солнца,
минеральные вещества поч­
вы и газы атмосферы,
Оолнечная
А
вода. Из него выделяются
энергия
Кр у гоТепло
минеральныетепло, кислород, углекис­
ворот
О 2 С 0 2
вещества
—
энергии
лый
газ, биогенные веще­
и
вещб
почвы, газы “
биогенные
ства I
атмосферы.
ства, переносимые водой,
вещ ества
Биотоп
вода
перегной. Основными функ­
абиотическая среда
неорганические
циями биогеоценоза явля­
соединения
ю тся поток энергии и круго­
бедные энергией
вороты веществ.
Биогеоценоз содержит
следующие обязательные
компоненты: 1) абиотиче­
Рис. 165. Схема круговорота энергии и веществ в
биогеоценозе (экосистеме).
ские неорганические и орга­
нические вещества среды;
2) автотрофные организмы — продуценты биотических органических
веществ; 3) гетеротрофные организмы — потребители (консументы)Готовых органических веществ первого (растительноядные животные)
и следующих (плотоядные животные) порядков; 4) детритоядные
организмы — разрушители (деструкторы), разлагающие органическое
вещество.
Особая роль в экономике биогеоценоза принадлежит цепям или
сетям питания (рис. 166). Они составляют трофическую структуру,
по которой происходит перенос энергии и круговороты ве­
ществ. П и щ е в а я ц е п ь состоит из ряда т р о ф и ч е с к и х
у р о в н е й , последовательность которых соответствует направлению
потока энергии. Первичным источником энергии служит солнечное
излучение,
энергия
которого
составляет
4,6 ж 1026 Дж/с
(1,1 ж 1026 кал/ с). 1/2 ООО ООО часть этого количества энергии достигает
поверхности Земли, при этом 1,0—2,0% ассимилируется растениями,
30—70% поглощенной энергии используется ими для обеспечения
собственной жизнедеятельности и синтеза органических веществ.
Эффективность, с которой растения ассимилируют солнечную
энергию, оценивается величиной первичной продуктивности. В разных
биогеоценозах эта величина различна.
Энергия, накопленная в растительной биомассе, составляет
ч и с т у ю п е р в и ч н у ю п р о д у к ц и ю биогеоценоза. Фитоби­
омасса используется в качестве источника энергии и материала для
создания биомассы потребителей первого порядка — растительно­
ядных животных и далее по пищевой цепи. Количество энергии,
расходуемой на поддержание собственной жизнедеятельности, в цепи
трофических уровней растет, а продуктивность падает. Обычно
продуктивность последующего трофического уровня составляет не
более 5—20% предыдущего. Э то находит отражение в соотношении
на
планете
биомасс
растительного
и
животного
проБиогеоценоз
Биоценоз
биотическая среда
богатые энергией
органические
соединения
384
исхождения. Так, суммар­
ная биомасса организмов,
обитающих на суше, со­
ставляет примерно 3 х
х 1012 т. Лиш ь 1— 3 % это­
го количества приходится
на зообиомассу. Масса жи­
вого вещества, приходяще­
гося на людей, составляет
около 0,0002% от суммар­
ной массы живого вещества
планеты. Объем энергии,
необходимой для обеспече­
ния жизнедеятельности ор­
ганизма, растет с повыше­
нием уровня морфофункци­
ональной организации. Со­
ответственно
количество
биомассы, создаваемой на
более высоких трофических
уровнях, снижается. Так, в
в разных биогеоценозах
95— 99,5%
зообиомассы
приходится на беспозвоноч­
ных животных. Прогрес­
сивное снижение ассимили­
рованной энергии в ряду
трофических уровней нахо­
дит отражение .в структу­
ре э к о л о г и ч е с к и х
п и р а м и д (рис. 167).
Продукция живого вещест­
ва растительноядными жи­
вотными составляет в дан­
ном случае 12,5%, а челове­
ком — 0,6%
продукции
растений. Снижение коли­
чества доступной энергии
на каждом последующем
трофическом уровне сопро­
вождается
уменьшением
биомассы и численности
особей. Таким образом,
пирамиды биомассы и чис­
ленности. организмов для
данного биогеоценоза по­
вторяю т в общих чертах
конфигурацию пирамиды
продуктивности.
14— 56
Органические
остатки и
минерализован
ные вещ ества
Вы сш ие
растения
Водоросли
сапрофит­
ные банте-
рии
Моллюсни
Ноловратни
П р о стей ш и е
Насекомые
и их
личинни
Свободноживущие
черви
Низшие ра­
кообразные |
Хищные
насекомые
Мальки
рыб
Паразити­
ческие
черви
Хищные
рыбы
Рис. 166. Схема простейшей пищевой
Стрелки направлены к источнику пищи.
1
Мальчи!
1А
Телята
| Л ю ц ерна
1
10
цепи.
4,5
2 * |0 7
|
100
масш таб
[Мальчин
4 8 кг
|Телята
8211 кг |
| Л ю ц ерна
1
___
1035 иг |
10
100
масш таб
[Ткани человека
| Телята
| Л ю ц ерна
3 4 .7 5 * Ю 3 Д ж
4,98*106 Д ш |
6.24*107 Дж ^
И спользованная солнечная энергия
26,4*109 Д Ж
10 100
—3
— __ I
Рис.
167.
Экологические
пирамиды.
а — чи сл а ; б — б и о м а сса; в
энергия. Р а с ч е т ы вы полнены.
исходя из д о пущ ен ия, что м а л ь ч и к п отр еб ляет 1 пищ у то л ьк о
те л я т и н у , а т е л я т а — лю ц ерну.
385
Размеры биогеоценозов, выделяемых экологами, различны.
Совокупности определенных биогеоценозов образуют г л а в н ы е
п р и р о д н ы е э к о с и с т е м ы , имеющие глобальное значение в
обмене энергии и вещества на планете, к которым относятся:
1) тропические леса; 2) леса умеренной климатической зоны; 3) пас­
тбищные земли (степь, саванна, тундра, травянистые ландшафты);
4) пустыни и полупустыни; 5) озера, болота, реки, дельты; 6) горы;
7) острова; 8) океан.
Понятие биогеоценоза применимо к хозяйственным угодьям,
создаваемым человеком, — пашням, лесопосадкам, паркам, прудам,
водохранилищам. Их называют а г р о б и о г е о ц е н о з а м и или
к у л ь т у р б и о г е о ц е н о з а м и . Благодаря целенаправленному
вмешательству человека, создающему благоприятные условия для
произрастания культурных растений путем вспашки, рыхления почвы,
прополки сорняков, внесения ядохимикатов, удобрений, агробиогеоце­
нозы существенно отличаются от природных биогеоценозов по
видовому составу, показателям вещественно-энергетического обмена,
устойчивости к внешним воздействиям.
11.3. Х А Р А К Т Е Р И С Т И К А Б И О Ц Е Н О З А
(С О О Б Щ Е С Т В А )
-
Главной частью биогеоценоза, от состояния которой зависят его
существование и изменения во времени, служит биоценоз. Биоценозы
отличаются по видовому составу, и важнейшей их характеристикой
является постоянное прямое /щи опосредованное взаимодействие
популяций организмов друг с другом. Влияние любой популяции
распространяется до экологически отдаленных элементов биоценоза
через взаимодействие с конкурентами, хищниками, жертвами. Так,
насекомоядные птицы не оказывают прямого действия на растения, но,
снижая численность насекомых, питающихся листьями или опыляющих
растения, они тем самым воздействуют на воспроизведение фитобио­
массы. Последнее существенно для состояния популяций и про­
дуктивности растительноядных животных, хищников, паразитов.
Экологические влияния отдельной популяции распространяются в био­
ценозе во всех направлениях, но по мере прохождения последователь­
ных звеньев в цепи взаимодействия интенсивность влияния ослабевает.
Показателями структуры и функционирования биоценозов служат
их видовой состав, число трофических уровней, первичная продуктив­
ность, интенсивность потока энергии и круговоротов веществ.
Структура биоценозов складывается в процессе эволюции, причем
каждый вид организмов эволюционирует таким образом, чтобы занять
в биоценозе определенное место. Совместное историческое развитие
многих видов на одной территории способствует их специализации к
использованию лишь части наличных пищевых ресурсов и ограниченно­
му местообитанию. В результате достигается состояние взаимоприспособленности видов друг к другу или коадаптации, которая служит
обязательным условием стабильности биоценоза.
В качестве примера рассмотрим ситуацию, возникшую в искус386
сгвенном озере Гатун, которое образовалось в начале текущего
столетия в зоне Панамского канала. В течение нескольких десятилетий
биоценоз озера отличался стабильностью благодаря коадаптации
организмов основной пищевой цепи: фитопланктон — зоопланктон —
планктоноядные рыбы. Последние, поедая зоопланктон, снижали его
численность, что способствовало поддержанию количества фитоплан­
ктона на достаточно высоком уровне. В 1967 г. случайно в озеро была
интродуцирована хищная, прожорливая рыба туканаре. Она быстро
сократила численность планктоноядных рыб, что привело к размноже­
нию зоопланктона и сокращению количества фитопланктона. Одновре­
менно снизилась численность обитающих на озере крачек и зимородков,
питающихся рыбой, и повысилась численность комаров, личинки
которых прежде поедались рыбой. Таким образом, появление нового
вида вызвало серьезные нарушения в экономике биоценоза озера и
дестабилизировало его структуру. В дальнейшем по мере развития
коадаптации при измененном видовом составе стабильность биоценоза
может восстановиться. Состояние коадаптации достигается даже
между видами-антагонистами: хищником и жертвой, хозяином и пара­
зитом. В биоценозах обнаруживаются все возможные виды взаимодей­
ствия тех организмов, которые их образуют. К ним относятся
мутуализм, внутри- и межвидовая конкуренция, хищничество, па­
разитизм.
11.4.
И З М Е Н Е Н И Е Б И О Ц Е Н О З О В ВО В Р Е М Е Н И
Лю бая теприто^&йд пригодная к жизни по набору абиотических
факторов, зааеляСтсй. Э т о т процесс называется
сукцессией.
В соответстви11 с трофической структурой биоценоза первостепенная
роль в освоений—йовых местообитаний принадлежит растительным
организмамТ~Р»4 §итие растительности в местообитаниях, в которых
прежде растении4 не \было> оббтца/ают как
первичную
с у к ц д с с идр, а в местах с предсутрствовавшим, но разрушенным
растительном Эюкрово^-?— как в туо р и ч н у кь^с у к ц е[с с и ю.
1$ процесса сукцессийЛдзменя ку'Ься | видовой соткав бцрценоза и
харш?геотсти1ш месторби-^^йй^/ Сукцессия завершается \
к лдрм а к с о м^Ообразован^ем кю б тъ ствк, видовой составЧю'Ьорого в
дальнейшем изменяется \незнаЧ)ггельноч Скорость сукцессшКро мере
приближения к состояний клилАкса.снижается. Процесс прекращается,
когда добавление или исюпЬч^иЙгйдов не приводи-* к изменению среды
развивающегося биогеоценоза, т .V . по достижений климакса; между
элементами биоценоза и фйзическоц средой устанавливается равнове­
сие.
Ч
4
/
Из наблюдений за заселением пеАаных дюн или ^нов^/образованных потоков лавы в результате первичной сукуебсйц, аТЙкже вырубок
или заброшенных пашен в результате вторичной сукцессии следует, что
для достижения состояния климакеауфебуетсЯ'Ссотвед ственно 1000 или
200 лет. В качестве примера приводим схему перБичной и вторичной
сукцессий после пожаров в районехззерг* Верхнее, ш тат Мичиган
(рис. 168). Климаксные сообщества, воз)вЙсающие в результате сукцес-
Вторичная сукцессия
Выжженный лес
{береза осина)
Пирогенные
сорняни
Цинл начался с небольшого пожара,
перегной не выгорел
== Климаксный л е с .
(канадская ель,
пихта,
у
береза)
Сухой лес
(сосна Банкса,
черная ель)
/
Высонорослые
кустарники
(ольха кизил, ива)
Л ес по торфянику Заболоченный лес
(туя. американская
(американская
лиственница,
лиственница,
черная ель)
черный ясень)
Вересковая дернина
Низкорослые
кустарники
(малина,
I
Лишайники (кладоиия)
♦
Ж И М ОЛОСТЬ)
Листовые лишайнини
Нустарники по
Нустарники
торфянику
(восковница, ясень)
со сфагнумом
(хамедафия
болотная, багульник
Злановые тоавы
♦
*
Норковые лишайники
Отдельные
травянистые
растения
Густы е заросли
осоки
1
I
Водные
растения
Водные
растения
Сукцессия на
поверхности
к породы________
Сукцессия на
морском берегу
Густые
заросли осоки
С укцессия
на болоте
С укц ессия на
торфянине
Ч_________
Первичная сукцессия
Рис. 168. Первичная и вторичная сукцессия после пожаров разной силы в районе
озера Верхнее.
Площадь, %
Тип экосистемы
10
5
Чистая первичная продунция. %
0 0
10
20
30
Г
Наземные
Тропический лес
Л ес умеренной зоны
Северный лес
Нустарнини
Саванна
/гЛУ//////л
У/////А
Ш
772/7ЯА
Степи умеренной зоны
Тундра и альпийская зона
Пустыня
Возделываемые земли
Болота и марши
Зодные
________
Открытый о кеан.
1
^
Нонтинентальный шельф
Заросли водорослей и кораллы
Эстуарии
О зера и реки
Рис. 169. Площадь поверхности и годовая продукция главных биогеоценозов (эко­
систем). Площ адь и первичная продукция даны в процентах от соответствующих
величин для всего земного шара.
сии разных местообитаний, различаются по производимой биомассе:
тропический лес, леса умеренной зоны, болота. Максимальный
объем
биомассы
ограничивается
климатом
соответствующего
района.
Сукцессия проявляется также в саморазвитии биогеоценозов и
постепенной смене видового состава растений, животных и микроорга­
низмов, изменении показателей продуктивности, численности орга­
низмов. Примером такой сукцессии служит зарастание водоемов.
Последовательное отмирание и придонное отложение мелких планк­
тонных организмов, донных водорослей, водоплавающих растений,
сопровождаемые сменой преобладающих видов животных и микро­
организмов обусловливает смену водных биогеоценозов на биогеоце­
нозы болотного типа.
Климаксные сообщества характеризуются состоянием устойчивого
равновесия, что проявляется в их способности возвращаться в исходное
состояние после кратковременных внешних воздействий, изменяющих
условия существования, а также противостоять этим воздействиям.
Так, в одном из климаксных биогеоценозов при временном понижении
осадков на 50%^го сравнению с их обычным количеством продукция фи­
тобиомассы снижалась на 2 5 % , а численность популяций растительно­
ядных — всего на 10%. Устойчивость таких сообществ зависит как от
гомеостатических реакций организмов и популяций, так и от условий
физической среды. В приведенном примере она могла бы ть обусловлена
запасом влаги в почве и реакцией растений на засуху. Устойчивость со­
общества растет с повышением видового разнообразия и увеличением
биомассы.
Главные природные климаксные сообщества, в частности сухо­
путные (рис. 169), отличаются значительной биомассой, но малой
первичной продуктивностью. В наиболее продуктивных из них в ре­
акциях фотосинтеза используется- всего 1—2 % солнечной энергии.
В связи с этим они не дают достаточно продукции, чтобы прокормить
растущее человечество.
Обратные соотношения — относительно малая биомасса и высокая
первичная продуктивность — свойственны агробиогеоценозам, кото­
рые в силу этого экономически выгодны. Однако без постоянного ухода
и защиты со стороны человека они обречены на быструю смену
природными биогеоценозами.
11.5. Г О М Е О С Т А Т И Ч Е С К И Е Р Е А К Ц И И
ЭКО Л О ГИ ЧЕС КО Й
НА И З М Е Н Е Н И Е
СИТУАЦИИ
Для большинства видов экологический оптимум биологической
активности ограничен жесткими рамками. Сохранение должного уровня
биологической активности, несмотря на колебания интенсивности
экологических факторов, обеспечивается гомеостатическими меха­
низмами на уровне особи
или популяции.
У
видов-к о нформистов
в ответ на смену экологической ситуации про­
исходят изменения определенных структурных и физиологических
параметров. Виды-р е г у л я т о р ы противостоят колебаниям среды,
389
поддерживая постоянство своей собственной среды. Типичными кон­
формистами являются пойкилотермные, а регуляторами — гомойотермные животные, хотя, строго говоря, в природе нет абсолютных
конформистов и регуляторов.
Примером реакции конформистского типа на изменение условий
окружающей среды, наблюдаемой у человека, служит увеличение
частоты дыхания и сердечных сокращений при восхождении на высокую
гору. Пребывание в горах в течение нескольких дней включает другой,
долгосрочный, гомеостатический механизм — увеличение количества
эритроцитов. Э то увеличивает концентрацию кислорода в крови и
позволяет человеку вернуться к обычному уровню активности.
В высокогорных популяциях структурно-функциональные приспособ­
ления к сниженному парциальному давлению кислорода, например
увеличенное содержание гемоглобина и эритроцитов, обнаруживаются
не только у взрослых, но и у детей, что указывает на стойкое воспроиз­
ведение гомеостатических механизмов в ряду поколений. Сопоставление
популяций кечуа и аймара из высокогорных районов Перу с их
равнинными сородичами, переселившимися в горы относитель­
но недавно, показало, что содержание гемоглобина у мигрантов
ниже.
Биологические механизмы упомянутых гомеостатических реакций
различны. Предполагают, что характер реакции организма или
популяции в значительной мере зависит от скорости нарастания и
продолжительности действия изменения среды. При внезапных откло­
нениях в среде организм использует быстрые, но кратковременные
физиологические или поведенческие реакции, основанные на принципе
обратной связи. Сохранение ситуации в течение длительного времени
приводит к преходящим изменениям определенных физиологических
процессов в пределах нормы реакции организма. Действие необычных
условий на популяцию в течение многих поколений приводит к
соответствующим изменениям ее генофонда. В таком случае приспосо­
бительный комплекс признаков складывается в процессе индивиду­
ального развития и характеризует определенный экотип организмов
данного вида. Последнее находит отражение в экологических правилах
К . Бергмана (1847) и Д. Аллена (1906). Согласно
правилу
Б е р г м а н а размер тела животных теплокровного политипического
вида с понижением температуры среды увеличивается. В соответствии с
правилом
А л л е н а в условиях жаркого климата у теплокров­
ных животных наблюдается тенденция к увеличению размеров
выступающих частей тела по сравнению с особями того же вида,
проживающими в районах с холодным климатом.
Приспособления, компенсирующие периодические и апериодические
колебания условий среды, многообразны. Кроме поведенческих,
физиологических, морфогенетических реакций, отмеченных выше, к ним
относятся особенности морфологии организмов, способствующие
экономии или рассеиванию энергии, диапауза, спячка, опреде­
ленный ритм суточной активности, сезонные миграции, запасание
пищи.
390
Глава
12
В В Е Д Е Н И Е В ЭКО Л О ГИ Ю Ч Е Л О В ЕК А
В настоящее время термином «экология человека» обозначают
комплекс полностью еще не очерченных вопросов, касающихся
взаимодействия человека с окружающей средой. Главной особенностью
экологии человека как самостоятельной области науки служит ее
междисциплинарный характер, так как в ней сходятся социологические,
философские, географические, естественнонаучные, медико-биологиче­
ские проблемы. Э к о л о г и я ч е л о в е к а изучает закономерности
возникновения, существования и развития антропоэкологических сис­
тем, которые представляют собой сообщество людей, находящееся в
динамической взаимосвязи со средой и удовлетворяющее благодаря
этому свои потребности.
Размеры таких систем различны в зависимости от численности и
характера организации человеческих популяций. Э то могут быть
изоляты, демы, нации, наднациональные ассоциации, различающиеся
по способу производства, укладу жизни, наконец, человечество в целом.
Большое значение в определении размера антропоэкологической
системы имеют природные условия. Наиболее многочисленные совре­
менные популяции, объединяющие более 80% человечества, обитаю т на
44% суши в области тропических лесов и саванн, а также в зоне умерен­
ного пояса с кустарниковой растительностью или смешанными лесами.
Напротив, на засушливых землях и в зоне пустынь, на которые прихо­
дится 18% суши, размещено 4 % населения.
Главной отличительной чертой антропоэкологических систем по
сравнению с природными экосистемами служит наличие в их составе
человеческих сообществ, которым в развитии всей системы принадле­
ж ит доминирующая роль. Сообщества людей различаются по способу
производства материальных ценностей и структуре социально-эконо­
мических отношений, от которых зависят способ организации труда,
объем и способ распределения производимой продукции между членами
сообщества. Активностью сообществ людей на занимаемой территории
определяется уровень воздействия их на окружающую среду. Развиваю­
щиеся сообщества (например, в период индустриализации) характеризу­
ются наряду с ростом численности населения, увеличением потребно­
стей его в продуктах питания, сырье, водных ресурсах, размещении
отходов. Э то повышает нагрузку на природную среду, интенсифициру­
ет использование биотических и абиотических факторов.
В процессе существования антропоэкологических систем взаимодей­
ствие людей и природной среды осуществляется по двум главным
направлениям. Во-первых, происходят изменения биологических и соци­
альных показателей отдельных индивидуумов и сообщества в целом,
направленные на удовлетворение требований, предъявляемых человеку
средой. Во-вторых, осуществляется перестройка самой среды для
удовлетворения требований человека. Н а протяжении истории челове­
чества соотношение названных изменений менялось в сторону
преобладающей роли второго направления. Естественная среда, в
которой зарождалось человечество, в результате перехода к культурно­
391
му земледелию и скотоводству уступила место
частично
о ч е л о в е ч е н н о й с р е д е сельских жителей. С возникновением
городов современного типа произошел переход к существованию
сообществ
людей
в
полностью
очеловеченной
с р е д е , границы распространения которой неуклонно расширяются.
Общим результатом биологических и социальных процессов в
антропоэкологических системах служит индивидуальная и групповая
приспособленность человеческих сообществ к жизни в средах обитания,
различающихся по природным условиям, формам хозяйствования и
культуры. Особенность такой приспособленности в отличие от
приспособленности к среде популяций любых других живых организмов
состоит в. том, что человек адаптируется к условиям жизни не только
физиологически, но прежде всего экономически, технически, эмоцио­
нально. Различные стороны и направления индивидуальной и групповой
адаптации человека, вся совокупность условий жизни и экологических
связей людей являю тся предметом изучения экологии человека. Именно
это делает ее междисциплинарной наукой. В курсе биологии допустимо
ограничиться рассмотрением отдельных вопросов, имеющих непосред­
ственное отношение к задачам охраны здоровья людей. Среди них
важное значение принадлежит биологической изменчивости популяций
людей в связи с биогеографическими особенностями среды, а также
медико-биологической характеристике антропогенных экологических
систем. Экологические вопросы паразитологии включены в соответ­
ствующий раздел учебника и излагаются ниже.
12.1. О Б Щ А Я Х А Р А К Т Е Р И С Т И К А
С РЕД Ы О БИ ТАН И Я Л Ю Д ЕЙ
Человек в среде обитания, с одной стороны, является объектом
действия экологических факторов, с другой — сам оказывает воздей­
ствие на среду. И как объект приложения экологических факторов, и как
самостоятельный экологический фактор он характеризуется важными
особенностями.
Отличительная черта человека как экологического фактора
заключается в осознанности, целенаправленности и массированное™
действия на природу. Ф . Энгельс писал: «животное только пользуется
внешней природой и производит в ней изменения просто в силу своего
присутствия; человек же вносимыми им изменениями заставляет ее
служить своим целям, господ ствует над ней» . Любой биологический
вид имеет ограниченные энергетические ресурсы, что лимитирует его
воздействия на среду. Зеленые растения используют энергию Солнца.
Все другие существа — часть энергии органических веществ, образуе­
мых организмами предшествующего трофического уровня. Человече­
ство в процессе трудовой и интеллектуальной деятельности расширяет
круг доступных источников энергии вплоть до использования ядерных и
термоядерных реакций. Э то позволило людям преодолеть естественные
ограничения роста их численности. Прирост народонаселения, энерго1 М аркс К ., Энгельс' Ф . Соч. Изд. 2-е, т. 20, с. 495.
392
Природа
О бщ ество
ооеспеченности, технической
вооруженности людей создает
предпосылки для заселения
любых экологических ниш.
Человечество
представляет
собой единственный на Земле
вид с всесветным распростра­
нением, что превращает че­
ловека в экологический фак­
тор с глобальным распрост­
ранением влияния. Благодаря
воздействию на все главные
компоненты биосферы влия­
ние человечества достигает
самых отдаленных экологи­
ческих зон планеты. Печаль­
ным примером этому слу­
жит, в частности, обнаруже­
ние Д Д Т в ^печени пингвинов
и тюленей, отловленных в
Рис. 170. Среда жизни человека.
Антарктиде, где никогда ни
один из инсектицидов не
применялся. Еще одна особенность человека как экологического
фактора заключается в активном, творческом характере его деятельно­
сти. Энергия, которой манипулируют люди, обращается ими на
изменение среды обитания. Экологический оптимум существования
человека на основе его биологических механизмов ограничен, и возмож­
ность широкого расселения достигается не путем изменения людьми их
собственной биологии, а путем создания очеловеченной среды.
Созданием вокруг себя искусственной среды обусловливается также
и специфика человека как объекта действия экологических факторов.
Это действие всегда опосредовано результатами производственной
деятельности людей: «Труд есть прежде всего процесс, совершающийся
между человеком и природой, в котором человек своей собственной
деятельностью опосредует, регулирует и контролирует обмен веществ
между собой и природой»1
. Естественные экосистемы вытесняются
антропогенными экосистемами, абсолютно доминирующим экологиче­
ским фактором которых является человек.
Обобщенное представление о среде обитания людей дает рис. 170.
Она включает биоприродный и социально-культурный компоненты или
естественную и искусственную среды. В естественной и искусственной
средах человек представлен как социальное существо.
Факторы естественной и искусственной среды оказывают на чело­
века постоянное действие. Результаты действия природных факто­
ров, различающихся в разных районах обитаемой части Планеты, на
протяжении истории человечества проявляются в настоящее время в
1 М аркс К.. Энгельс Ф . Соч. Изд. 2-е, т. 23, с. 188.
393
Л
экологической дифференциации населения земного шара, подразделении
его на адаптивные типы. Результатом действия социальных факторов
служит образование и закономерная смена в историческом развитии
х о з я й с т в е н н о - к у л ь т у р н ы х т и п о в сообществ людей,
которые представляют собой комплекс хозяйства и культуры,
характеризующий народы, различающиеся по происхождению, но
обитающие в сходных природно-ресурсных условиях и находящиеся на
одинаковом социально-экономическом уровне.
В настоящее время на планете сосуществуют различные по времени
возникновения, производительности труда, благосостоянию и демогра­
фическим показателям населения хозяйственно-культурные типы со­
обществ людей. В ограниченном числе сохраняется «присваивающий»
тип с преобладанием экономической роли охоты, рыболовства,
собирательства (пигмеи-охотники на территории Заира, племена аэта,
кубу, обитающие в лесах Юго-Восточной Азии, отдельные группы
индейцев в бассейне реки Амазонки). Достаточно широко представлены
хозяйственно-культурные типы, экономическую основу которых сос­
тавляю т ручное (мотыжное) или плужное (пашенное) земледелие и
скотоводство. В связи с научно-технической революцией в про­
мышленно развитых странах сложились хозяйственно-культурные ти­
пы с высокоразвитым товарным земледелием и животноводством.
Формирование хозяйственно-культурных типов зависит от есте­
ственной среды обитания людей. Э та зависимость была наиболее
сильна на ранних стадиях развития человеческого общества. Однако уже
тогда и особенно в более поздние периоды развития человечества
зависимость формирования хозяйственно-культурных типов от при­
родных условий опосредовалась уровнем социально-экономического
развития народа. Н а всех этапах истории общество активно приспо­
сабливает природу к собственным нуждам. Инструментом такого
приспособления, связующим звеном между естественной и очелове­
ченной средой служит трудовая деятельность людей, в процессе
которой человек создает хозяйственную и культурную среду, от
которой зависят образ жизни, показатели здоровья, структура
заболеваемости.
Среда обитания человека представляет собой сложное переплетение
взаимодействующих естественных и антропогенных экологических
факторов, набор которых различается в разных природно-географиче­
ских и экономических регионах планеты. В таких условиях необходим
единый и н т е г р а л ь н ы й к р и т е р и й к а ч е с т в а с р е д ы
с точки зрения ее пригодности для обитания человека. Согласно Уставу
Всемирной Организации Здравоохранения, принятому в 1968 г., этим
критерием служит состояние здоровья населения. В исследованиях по
экологии человека термин «здоровье» используется в широком смысле
как показатель полного физического и душевного благополучия.
Главная линия развития экологии человека в настоящее время
нацелена на решение проблем управления средой, выработку путей
рационального природопользования, оптимизации условий жизни
людей в различных антропоэкологических системах.
394
12.2. Э К О Л О Г И Ч Е С К А Я Д И Ф Ф Е Р Е Н Ц И А Ц И Я
ЧЕЛ О ВЕЧЕС ТВА
Искусственная среда способствует нивелировке прямого воздей­
ствия природных экологических факторов на человека. Степень этой
нивелировки, будучи в настоящее время в экономически развитых
странах значительной, различалась в разные периоды развития
человечества. Эра технической революции по времени занимает в
летописи человеческого общества менее 1 % . Эра научно-технического
прогресса началась в текущем столетии. Таким образом, на протяжении
большей части истории вида Человек разумный климатический,
алиментарный, геохимический, биологический, в частности, микробный
и паразитарный, факторы естественной среды продолжали оказывать
серьезное воздействие на популяции людей, не совпадающее по
направлению в разных частях Ойкумены. Последствия этого давления
сохраняются в современном человечестве в виде наличия адаптивных
типов людей.
А д а п т и в н ы й т и п представляет собой норму биологической
реакции на преобладающие условия обитания и проявляется в развитии
комплекса морфофункциональных, биохимических, иммунологических
признаков, обусловливающих лучшую биологическую приспособлен­
ность человека к определенной физической среде. Комплекс признаков,
свойственный конкретному адаптивному типу, не зависит от расовой и
этнической принадлежности популяции. Так, приспособления к жизни в
условиях хронической гипоксии обнаруживаются у аборигенов Арктики
и жителей высокогорья. В комплексы признаков адаптивных типов из
разных географических зон входят общие и специфические элементы.
К первым относят, например, показатели костно-мускульной массы
тела, количество иммунных белков сыворотки крови человека. Такие
элементы повыш ают общую сопротивляемость организма к небла­
гоприятным условиям среды. Специфические элементы отличаются
разнообразием и тесно связаны с преобладающими условиями в данном
месте обитания — гипоксией, жарким и холодным климатом. Именно
их сочетание служит основанием к выделению адаптивных типов:
арктического, тропического, зоны умеренного климата, высокогорного,
пустынь и полупустынь и др.
Особенности природной среды Арктики и Антарктики заключаются
в постоянно низких температурах воздуха, отсутствии лесов, скудной
растительности тундр и лесотундр. Ж ивотный мир довольно богат и
представлен рыбой, морскими млекопитающими, арктическими видами
животных — лис, медведей, зайцев, поэтому пиша богата белками и
жирами, но бедна углеводами. Кроме того, существует очевидная
зависимость количества добываемой пищи от сезона. Летом в ежеднев­
ный рацион жителей входят ягоды, молодые побеги карликовых
деревьев. Организм человека в достаточном количестве получает
железо, витамины А , В , С , Э . Воздух и почва в Антарктиде и Заполярье
содержит мало микроорганизмов.
И з перечисленных факторов наибольшее влияние на формирование
комплекса признаков
арктического
адаптивного
395
т и п а оказали, по-видимому, холодный климат и преимущественно
животная пища. Арктическому комплексу признаков свойственны
относительно сильное развитие костно-мускульного компонента тела,
большие размеры грудной клетки, высокий уровень гемоглобина,
относительно большое пространство, занимаемое костным мозгом,
повышенное содержание минеральных веществ в костях, высокое
содержание в крови белков, холестерина, повышенная способность
окислять жиры. Среди аборигенов Арктики почти не встречаются лица
с астеническим телосложением. В целом арктический тип характеризу­
ется усиленным энергетическим обменом, который отличается ста­
бильностью показателей в условиях переохлаждения. Им ею т свои
особенности и механизмы терморегуляции. Так, при одинаковой
степени охлаждения у канадских индейцев резко падает температура
кожи, но уровень обмена веществ меняется незначительно, а у пришлого "
белого населения наблюдается меньшая степень снижения кожной
температуры, но появляется сильная дрожь, т. е. интенсифицируется
обмен.
В тропиках и субтропиках располагается чуть ли не большая часть
Ойкумены. Э то т регион отличается в целом экстремальным количе­
ством тепла и влаги, большей или меньшей сглаженностью сезонных
колебаний условий обитания. Вместе с тем благодаря особенностям
рельефа наблюдается значительная контрастность распределения тепла
и влаги — массивы влажных лесов нередко соседствуют с засушливыми
плато, обширными равнинами и редколесьем. В экваториальной и
субэкваториальной областях сосредоточены огромные количества
растительной биомассы. Влажные тропические леса относительно
бедны животными, тогда как в субтропиках животный мир разнообра­
зен и включает крупных стадных животных, издавна используемых
человеком как объект охоты. К преобладающим экологическим факто­
рам, под влиянием которых формировался комплекс признаков
тропического
а д а п т и в н о г о т и п а , относятся жаркий
и влажный климат, рацион с относительно низким содержанием
животного белка. Свое влияние оказало также большое разнообразие
экологических условий от района к району.
В субтропической и тропической областях наблюдается исключи­
тельно широкая вариабельность групп населения в расовом, этническом
и экономическом отношениях. Э т о проявляется в поразительном
размахе изменчивости, например по соматическим признакам. Так,
именно здесь проживают самые низкорослые и самые высокорослые
племена. Пределы этой изменчивости таковы, что в них укладываются
все известные ныне человеческие популяции. Тем не менее преобладаю­
щие экологические факторы, особенно климатический, способствовали
образованию определенного комплекса морфофизиологических призна­
ков обитателей тропиков и субтропиков Африки. К характерным
признакам негроидов относятся удлиненная форма тела, сниженная
мышечная масса. У них наблюдается относительное уменьшение массы
тела при увеличении длины конечностей, уменьшение окружности
грудной клетки, более интенсивное потоотделение за счет повышенного
количества потовых желез на 1 см2 кожи, низкие показатели основного
396
обмена и синтеза жиров, понижена в крови концентрация холестерина.
Антропологическое изучение современных обитателей зоны уме­
ренного климата под углом зрения формирования биологических
механизмов адаптации к природным условиям затруднено, так как
большая часть населения проживает в промышленно развитых странах
с большой долей городского населения. Результаты наблюдения
позволяют, однако, говорить о том, что и в этом случае в процессе ис­
торического развития человеческих популяций сформировался комплекс
признаков, соответствующий особому а д а п т и в н о м у т и п у
у м е р е н н о г о п о я с а . По соматическим показателям, уровню
основного обмена население умеренного пояса занимает промежуточное
положение между коренными жителями арктического и тропического
регионов. Э то соответствует условиям биогеографической среды в зоне
умеренного климата. Для нее характерны неравномерное распределение
районов, отличающихся по количеству тепла и влаги, типу расти­
тельности (от сухих степей и полупустынь до тайги), богатству
животного мира. Вместе с тем температура и влажность воздуха здесь
не достигают экстремальных величин, хорошо выражен сезонный ритм
биоклиматических условий.
Условия высокогорья для человека во многих отношениях
экстремальны. Их характеризует низкое атмосферное давление,
сниженное парциальное давление кислорода, холод, относительное
однообразие пищи. Основным экологическим фактором формирования
г о р н о г о а д а п т и в н о г о т и п а явилась, по-видимому, ги­
поксия. У жителей высокогорья независимо от климатической зоны, ра­
совой и этнической принадлежности наблюдается повышенный уровень
основного обмена, относительное удлинение длинных трубчатых
костей скелета, расширение грудной клетки, повышение кислородной
емкости крови за счет увеличения количества эритроцитов, содержания
гемоглобина и относительной легкости его перехода в оксигемоглобин.
В процессе исторического развития, осваивая новые местообитания,
различающиеся преобладающими условиями географической среды,
человечество подвергалось экологической дифференциации. Адаптив­
ные типы формировались независимо от расы в связи с конкретными
экологическими задачами на основе резерва приспособительных
механизмов вида, соответствующих его генофонду. Выделение этих ти­
пов не носило черт экологической специализации, о чем свидетельствует
возможность проживания людей одного адаптивного типа на террито­
рии, которой в большей степени отвечает комплекс признаков другого
адаптивного типа. Скорее процесс заключался в образовании
популяций, генофонды которых соответствовали местным условиям
лучше, чем генофонд вида в целом. В пользу такого объяснения говорят
результаты метисации, например, между русскими и бурятами.
Предполагается, что в период заселения русскими Забай­
калья процесс метисации с местным населением был интенсивным.
Русские из Европейской части С С С Р характеризуются по сравнению с
бурятами несколько иными пропорциями тела, а также сниженным
процентом лиц, ощущающих вкус фенилтиокарбамида. В настоящее
время вследствие метисации произошло сближение потомков русских
397
старожилов Сибири, мигрировавших в X V II в., с бурятами по
пропорциям тела, повышение числа русских и уменьшение числа
бурятов, ощущающих вкус фенилтиокарбамида, среди жителей Баргузинской котловины в Сибири.
Особенности определенного адаптивного типа формируются в
эмбриогенезе. Так, различия в пропорциях тела между негроидами и
европеоидами вполне заметны к концу внутриутробного периода
развития.
Наличие различных адаптивных типов свидетельствует о значи­
тельной экологической изменчивости человека, которая послужила, повидимому, одной из предпосылок всесветного расселения людей. Н а
современном этапе эта изменчивость отражает преобладающие
направления действия отбора на генофонды человеческих популяций в
разных местообитаниях на заре развития человечества.
12.3. А Н Т Р О П О Г Е Н Н Ы Е Э К О С И С Т Е М Ы .
О СО БЕН НО СТИ ЭК О Л О ГИ И ГО РО Д О В
Отличительная черта антропогенных экосистем состоит в том, что
доминирующий экологический фактор в них представлен сообществом
людей и продуктами его производственной и общественной деятельно­
сти. Благодаря целенаправленной преобразующей активности человека
в отношении природы антропогенным экосистемам свойственно пре­
обладание искусственной среды над естественной. Последняя входит в
среду обитания людей в очеловеченном виде, приспособленной к
нуждам людей, включенной в круг их трудовых, культурных, бытовых
интересов. Преобразующая деятельность людей в современных услови­
ях строится, как правило, на основе предварительного планирования,
однако предполагаемый ход развития антропогенных экосистем не­
редко искажается благодаря действию стихийных, неучтенных челове­
ком заранее сил. В результате возникают состояния «экологических
стрессов», которые могут в перспективе привести к экологическому
кризису.
Важнейшие современные антропогенные экосистемы — города,
сельские поселения, транспортные коммуникации — характеризуются
определенным сочетанием биоприродных и хозяйственно-культурных
условий. Медико-санитарная оценка антропогенных экосистем приво­
дится в курсе гигиены. Рассмотрим в качестве примера такой системы
города, в которых наиболее заметны положительные и отрицательные
стороны процесса превращения естественных экосистем в антропо­
генные. Города в истории человечества представляют особую среду
обитания с неповторимым, качественно новым сочетанием производ­
ственных и социально-бытовых условий. Они возникли всего около
7000 лет назад. К 1950 г. в них проживало 28% , а к 1970 г. 40% населе­
ния планеты. В 2000 г., согласно расчетам разных исследователей,
ожидается дальнейшее возрастание доли городского населения от
56—62 до 70— 90%. В настоящее время более трети горожан проживают
в городах с числом жителей не менее 1 млн. В странах с высокой плот­
398
ностью населения происходит слияние соседних городов, что приводит
к образованию мегаполисов — обширных территорий с высоким уров­
нем урбанизации.
Урбанизация
в целом явление прогрессивное. В связи с
концентрацией производства, научных и культурных учреждений,
учебных заведений она создает благоприятные условия для производ­
ственной деятельности и организации бы та людей — легче решаются
вопросы трудоустройства, образования, снабжения продовольствием,
медицинского обслуживания, быта.
Вместе с тем в городах наиболее выражены изменения природной
среды. Так, климатические факторы действуют в черте города с иной
интенсивностью, чем на территории, его окружающей. Обилие
промышленных и бытовых отходов приводит к необычному распреде­
лению в почве, водах, растительности городов многих микроэлементов,
т. е. к геохимическим аномалиям. Города отличаются высокой
плотностью населения, что создает благоприятную обстановку для
распространения инфекционных заболеваний.
Благодаря загрязнению воздуха аэрозолями, препятствующему
ночному излучению тепла, накоплению тепла каменными зданиями,
тепловым выделениям промышленных предприятий и транспорта,
средняя годовая, месячная, суточная температура в городах на
несколько градусов выше, чем на окружающей территории. В СанФранциско, например, температура воздуха на окраинах на 6—7 °С
ниже, чем в центре. Э то типично и для других крупных городов. Неред­
ко в таких городах в летнее время образуются «острова жары», что
оказывает резкое отрицательное воздействие на здоровье людей. Так, в
июне 1966 г. в С Ш А прошла волна высоких температур и в центральных
районах многих крупных городов благодаря возникновению «островов
жары» при среднесуточной температуре воздуха 35°С фактическая
смертность превысила ожидаемую на 2 7 % , при температуре
3 7 ,8 °С — н а7 5 % , при40,6 °С — на200%, при 4 Т ,3 °С — на|546%.
Задымленность воздуха снижает0в городах интенсивность ультра( фиолетовой радиации зимой на 30%, а летом — на 5 % . Длительность
солнечного освещения сокращается на 5— 15%. Развивается «световой
голод», который вызывает авитоминоз Э , способствует повышенной
утомляемости людей, ухудшению их самочувствия и настроения,
снижению сопротивляемости к инфекционным и простудным заболева­
ниям. При сокращенном световом дне ослабевают антимикробные
свойства среды. Так, по сравнению с больными из сельской местности у
горожан, больных хроническими формами воспаления легких, в мокро­
те чаще обнаруживаются стафилококки и стрептококки. К техногенным
факторам, достигающим в городах значительных величин, кроме
загрязнения среды, относятся шум и вибрация.
Среди социальных факторов наибольшее влияние на здоровье людей
имеет высокая плотность (контактность) населения в условиях города.
С одной стороны, эт^т фактор имеет положительное значение для
организации производства, а с другой — нередко приводит к перенапря­
жению нервной системы. Города отличаются в целом все более
возрастающим темпом жизни, но наряду с этим образ жизни горожан
399
становится все менее подвижным. Падение физической активности
вызывает детренированность сердечно-сосудистой системы.
Для питания современного городского населения типично повыше­
ние калорийности пищи, увеличение в рационе жиров и углеводов,
уменьшение потребления растительной пищи и молока. Одновременно
сокращается доля продуктов, нерафинированных или не имеющих
химических добавок.
Таким образом, город как особая антропогенная экосистема имеет и
положительные, и отрицательные стороны. Урбанизированная среда
характеризуется повышением уровня жизни, снижением общей заболе­
ваемости, что проявляется в росте такого показателя, как средняя
продолжительность жизни. В конце X IX века мужчины в России жили в
сельской местности дольше, чем в городе, на 3,4 года, в Пруссии — на
3,9 года, в Великобритании — на 8,8 лет, в С Ш А — на 10 лет._
В 1926— 1927 гг. средняя продолжительность жизни в городах С С С Р
была выше, чем в селах, почти на 1 год. Н а рубеже 50—60-х годов этот
показатель для города и села сравнялся. Вместе с тем на фоне снижения
общей заболеваемости в городах повышается частота заболеваний,
ранее не имевших широкого распространения. Уровень заболеваемости
болезнями органов кровообращения, дыхания, нервной системы среди
городского населения выше в 1 ‘/г —2 раза. Города отличаются более
низкими показателями рождаемости. В 1973 г. при сопоставимых
показателях смертности прирост составил в городах 5,9, а в сельской
местности 8,9 человек на 1000 населения.
12.4. А Д А П Т А Ц И И Ч Е Л О В Е К А К С Р Е Д Е О Б И Т А Н И Я
Биологические адаптации рассмотрены выше (см. 7.4.8). Благодаря
биосоциальной природе адаптации людей к условиям обитания имеют
отчасти биологическую, но главным образом социальную природу.
В настоящее время преобладающее значение для освоения человеком
новых сред обитания и создания лучших условий жизни в уже освоенных
средах имеют социально-гигиенические мероприятия, результатом
которых служит совершенствование средств и систем жизнеобеспече­
ния, достижение состояния комфорта в местообитаниях людей.
Адаптации создаются по отношению к факторам как природной, так и
искусственной среды, поэтому они носят не только экологический (в
узком смысле этого термина), но и социально-экономический характер.
Так как каждый человек представляет собой индивидуальность,
экологические и социально-экономические адаптации дополняются
психологическими адаптациями. Индивидуальные и групповые адапта­
ции человека в отличие от биологических адаптаций растений и
животных обеспечивают наряду с выживанием и воспроизведением
потомства выполнение им социальных функций, важнейшей из которых
является общественно полезный высокопроизводительный труд.
Социально-гигиенические мероприятия, направленные на оптимиза­
цию условий жизни и производственной деятельности, включаю т
устройство жилищ и других помещений, конструкцию одежды,
организацию питания и водоснабжения, рациональный режим труда и
400
отдыха, сознательную направленную тренировку организма и многое
другое. При этом исходят из принципа «разумного максимума»
удобств, при котором поддерживается высокая работоспособность и
сохраняется здоровье населения, но в то же время не допускаются изне­
живание организма и чрезмерные экономические затраты.
Х о тя в основе адаптации человека лежат социально-экономические
механизмы, важная роль принадлежит также состоянию естественных
приспособительных и защитных механизмов, составляющих биологи­
ческое наследство людей. Достаточно демонстративно эта роль
выявляется при переходе в местообитания с экстремальными условия­
ми, которые создаются благодаря наличию на заселяемой территории
экологического фактора или комбинации факторов, оказывающих на
здоровье человека выраженное неблагоприятное действие. Они могут
складываться не только в естественных (Арктика, высокогорье), но и в
антропогенных (крупные города) местообитаниях. Так, выходцев из
зоны умеренного климата, прибывающих на работу в Арктику или
Антарктиду, встречают суровый климат, необычные для средних
широт атмосферные явления, резко пониженное количество микроорга­
низмов в почвах и воздухе, жизнь в относительно малочисленных,
скученных коллективах. Как правило, такие люди по прибытии в
Заполярье длительное время испытываю т болезненные состояния и
ощущения, усиливающиеся, например, при смене полярных дня и ночи.
Они проявляются в повышении артериального давления и учащении
пульса, которые сменяются затем понижением давления, иногда до
уровня 70/30 мм рт. ст., и урежением пульса. Э ти явления, обозначае­
мые некоторыми исследователями как «метеоневроз», сопровождаются
падением работоспособности. Описанная ситуация находит отражение
в рекомендациях гигиенистов, ограничивающих продолжительность
работы для вновь прибывших в Заполярье. Так, при температуре до
—30°С и скорости ветра 4— 8 м/с основной состав полярной станции
может работать на открытом воздухе полный рабочий день, тогда как
вновь прибывшие — не более 1 ч. У полярников количество лейкоцитов
в крови обычно снижено до уровня 3000—3500 в 1 мм3. В период смены
состава зимовщиков при контакте с вновь прибывшими, как правило,
наблюдается почти поголовная заболеваемость простудными и ки­
шечными заболеваниями. Выявляю тся признаки утомления и даже
истощения нервной системы — ухудшается оперативная память, сни­
жается надежность работы человека, увеличивается продолжитель­
ность скрытого периода двигательных реакций.
По прошествии времени у части людей функциональные показатели
возвращаются к нормальному уровню. У других они остаются
измененными по сравнению с исходными значениями, наблюдавшимися
до приезда в Арктику, однако восстанавливается работоспособность и
самочувствие. В таких случаях говорят об а к к л и м а т и з а ц и и
людей к новым условиям обитания. Для растений и животных,
перенесенных в необычную среду обитания, критерием акклиматизации
служит их выживание, для людей — восстановление высокого уровня
трудоспособности. Изменение физиологических механизмов при аккли­
матизации нередко имеет сложный характер. Так, у адаптированных к
401
жизни в холодном климате людей при охлаждении наблюдается
изменение кровоснабжения кожных покровов. П ри э то м степень
изменения различается в разных частях тела. У акклиматизированных в
Заполярье людей, например, на холоде тепловой поток с рук возрастает
на 4 0 % , тогда как с груди — на 1 9 % . Т аки м образом, благодаря под­
держанию достаточно высокой тем пературы сохраняется работоспо­
собность рук.
Д лительное существование групп людей на территориях, различаю ­
щихся преобладающ ими климатическими, алим ентарным и и другими
факторами, привело к образованию воспроизводящихся в ряду
поколений комплексов признаков. Э т и комплексы со о тветствую т эко­
логическим ти п ам людей и обусловливаю-! более высокий уровень
приспособленности к проживанию в определенной биогеографической
среде за счет биологических механизмов. Такое положение свидетель­
ствует об относительности понятия экстремальности природных
условий в тех или иных регионах планеты. Вм есте с тем перед человече­
ством сто и т задача глубокого освоения и более плотного заселения
м алокомф ортных территорий — п у сты н ь и полупусты нь, вы соко ­
горных, полярных, тропических районов, дна океанов и морей, на долю
которых приходится 7 1 % поверхности планеты, космоса. П ризнаки
экстремаль ноет и приобретают и некоторые антропогенные ф акторы
среды обитания людей. Решение этих задач ст о и т в связи с социальноэкономическим прогрессом человечества.
Раздел
IV
БИ О Л О ГИ Ч ЕС К И Е О СНОВЫ
П АРАЗИ ТИ ЗМ А
Г л а в и 13
П АРАЗИ ТИ ЗМ К А К Ф О РМ А
БИО ТИ ЧЕСКИ Х
С ВЯЗЕЙ
Качествен ная особенность ж и вы х организмов со стои т в непре­
рывной связи с окружаю щ ей средой — живой и неживой природой.
Биотические связи (м еж д у ж и в ы м и организмами) характеризуются
больш ой сл о ж но стью и разнообразием, но в основе их л еж ат прежде
всего пространственные и пиш евые отнош ения. Такие типы связей
объединяю т между собой различные компоненты биогеоценозов и
антропобиогеоценозов.
13.1.
Ф О РМ Ы Б И О ТИ Ч Е С К И Х С В Я З Е Й В П Р И Р О Д Е
Р а зл и ч аю т следующие формы взаимоотношений м еж д у особями
различных видов: симбиоз, квартирантство, комменсализм, хищниче­
ство и паразитизм.
С и м б и о з (о т греч. $уп — вместе, Ьюз — жизнь) — обоюдополезное сожительство, при котором оба партнера приносят пользу друг
другу. Т ак, в кишечнике человека ж и в у т киш ечные бактерии зшерихии,
которые п и таю тся его содержимым и в саою очередь способствую т
вы р аботке витам инов группы В, а та кж е о бладаю т способностью
подавлять активность возбудителей кишечных заболеваний, например
брюш ного тифа и бактериальной дизентерии.
Следует им еть в вицу, что иногда употребляю т термин «симбиоз»
в ш ироком см ы сле слова, объединяя этим понятием все формы
отношений между организмами. В это м случае взаимовыгодное
сож ительство или собственно симбиоз о бозначаю т термином « м ут у а ­
лизм».
К в а р т и р а н т с т в о — пространственная форма связи, пос­
кольку оба партнера в это м случае м огут б ы т ь индифферентны друг к
другу или извлекает пользу то л ько один партнер, ко то ры й использует
организм или убежище другого в качестве м еста обитания Т ак, норы
грызунов использую тся другими ж и в о тн ы м и (пауки м оски ты , блохи
и др.); молодь некоторых морских ры б держится около щупалец медуз
и в случае опасности прячется под их зонтом.
К о м м е н с а л и з м
(о т фран. соштеп5а1 — сотрапезник) —
вы раж ается не то л ько в пространственных, ио и в пиш евых связях.
Один из партнеров использует для питания или излишки, или отходы
пиши другого, не причиняя ему вреда. В качестве примера м ож но
привести р о то вую амебу, о би таю щ ую в ротовой полости человека.
Х и щ н и ч е с т в о — однократное использование добычи хищни­
ком, так как используемый организм погибает.
П а р а з и г и з м (о т греч. рагазло* — тунеядец) — представляет
собой ф орму взаимоотношений между организмами различных видов,
при которой один организм (паразит) использует другого (хозяина) как
источник питания и место обитания, причиняя ему вред, но, как
правило, не ун и что ж а я его.
П аразитический образ жизни обы чно сл у ж и т специфическим
признаком вида, он
свойствен всем особям без исключения и
закреплен филогенетически.
Ф о р м ы проявления паразитизма чрезвычайно многообразны.
П арази ты м о гут о би тать в различных тканях и органах хозяина,
питаться его тканям и или переваренной пищей, проводить на теле или в
теле хозяина всю свою жизнь илн то л ько часть ее, а такж е б ы т ь посто­
янными паразитами или временными. Д а ж е такое свойство паразитов,
как вредоносность, проявляется не всегда, а зависит о т вида и состояния
паразита и хозяина и окружающ ей среды. Т ак, например, мелкая
вегетативная форма дизентерийной ам ебы при определенных условиях
не обладает вредоносным действием, а под влиянием провоцирующих
факторов приобретает патогенные свойства.
Наиболее близки к паразитизму хищ ничество и комменсализм, и в
ряде случаев б ы ва ет трудно провести границу между этим и формами
биотических связей.
П аразитизм широко распространен в природе. П о данным
В . А . Догеля, сущ ествует 60—65 ты с. видов ж ивотны х, ведущих
паразитический образ жизни, что со ставляет 6— 7 % о т общего числа
всех видов на Земле. Ко ли чество паразитических форм в разных типах
животного мира неодинаково. Н аибольш ее количество паразитов
установлено в типе простейших, плоских и круглых червей, а такж е
членистоногих.
13.2.
О БШ А Я И М Р Д и ЦИ Н С КА Я
П Р Ь Д М Ь Т И ЗА Д А Ч И
П А РА ЗИ Т О Л О ГИ Я .
П а р а з и т о л о г и я
(о т греч. рагазЛоБ — тунеядец, 1о^об —
учение) — комплексная
биологическая
наука,
изучаю щ ая
явле­
ние паразитизма, биологию и экологию паразитов, а та кж е вы зы ва е­
м ы е ими заболевания и меры борьбы с паразитами.
М е д и ц и н с к а я
п а р а з и т о л о г и я
изучает паразитов
человека, разрабаты вает научные основы борьбы с ними, а такж е
методы диагностики, лечения н предупреждения заболеваний, вы зы ва е­
м ы х или распространяемых паразитами.
Медицинская паразитология подразделяется на 3 раздела: медицин­
скую п р о т о з о о л о г и ю , изучаю щ ую паразитов человека из типа
простейших; медицинскую
ге л ь м и н т о л о г и ю ,
изучаю щ ую
гельминтов, паразитирующ их у человека, из типа плоских и круглых
червей; медицинскую а р а х н о э н т о м о л о г и ю ,
изучаю щ ую
представителей типа членистоногих (клешей и насекомых), являющ ихся
404
переносчиками, природными резервуарами и возбудителями заболева­
ний человека.
О сн овн ы е задачи медицинской паразитологии следующие:
— изучение особенностей строения паразитов на всех стадиях
развития, что необходимо для точного определения вида. Правильное
определение видовой принадлежности паразита имеет для врача
первостепенное значение, так как помогает устан овить правильный
диагноз, ч то в сво ю очередь определяет методы лечения. Так,
ленточны е черви (свиной н бы чи й цепень), паразитирующ ие в кишечнике
человека, сходны по строению и циклу развития, но присутствие
свиного цепня м о ж е т в ы зв а ть тяжелейш ие осложнения, иногда закан­
чивающиеся см ер тью больного, в т о время, как заболевание, в ы з ы ­
ваемое б ы чьи м цепнем таких осложнений никогда не дает. П ра­
вильное определение вида паразита позволяет своевременно принять
соответствую щ ие меры, предотвращающие тяж елы е последствия;
—: изучение систематики объектов паразитологии, нх принадлеж­
ность к то й или иной систематической группе для последующего
определения вида;
— изучение особенностей циклов развития (экологии) паразитов и
переносчиков позволяет устан овить пути циркуляции паразитов в
природе и способы попадания в организм человека, а та кж е определить
наиболее «слабое» звено цикла для последующего воздействия на него
в целях профилактики заболеваний и уничтожения паразита;
— изучение взаимоотнош ений в системе паразит — хозяин;
— разработка научных основ диагностики и лечения паразитарных
заболеваний на основании знания вредоносного действия паразитов, а
такж е методов профилактики и борьбы с паразитами и переносчиками;
— решение перечисленных задач определяет основную задачу
медицинской паразитологии — создание систем ы , обеспечивающей
профилактику и ликвидацию паразитарных заболеваний.
Медицинская паразитология неразрывно связана как с дисциплина­
ми клинического, так и санитарно-гигиенического профиля, особенно
эпидемиологией.
П а р а зи ты , как у ж е отм ечалось, м о гут п ораж ать лю бой орган
человека, поэтом у врачу лю б ой специальности в той или иной мере
приходится им еть дело с паразитарными заболеваниями (рис. 171).
К врачам-терапев там обращ аю тся б ольны е с поражением кишечни­
ка и ж елчны х путей (жгутиконосец лям блия), печени (трем атоды ,
эхинококк и альвеококк), легких (легочный сосальш нк), а такж е
страдаю ш ие висцеральным лейш маниозом, малярией, киш ечными
гельминтозами и их последствиями в виде интоксикации, злокаче­
ственным малокровием (анкилостом атоз, дифнллоботриоз н др.).
Врачи-хирурги о сущ ествл яю т оперативное лечение целого ряда
паразитарных заболеваний: эхинококкоза и альвеококкоза печени и
легких, кишечной непроходимости, вызванной аскаридами, закупорки
желчного протока аскаридами или сосальщ иками, дракункулеза и т. д.
Наиболее часто с паразитарными заболеваниями приходится иметь
дело педиатрам, поскольку дети в связи с недостаточно сформиро­
ванной иммунологической системой чаш е страд аю т этими заболевания405
17
Рис. 171. Л о кал и зац и я паразитов человека.
I
малярийный плазмодий и комар анофелес; 2 — лейшманни, 3 — трипаносома; 4 — токсоплазма;
5
лям б ли я; 6
дизентерийная амеба, 7 - балантидий, 8 — некатор; 9 — острица; 10
аскарида;
I I — трихомонача. 12
криноголовка: 13
риш та; 14
нласо 1ла в, 15 — чесоточный зудень, 16 — иксо>внй клеш ; 17 — клеш дермацентор; 18 - поселковый клещ ; 19 — финна вооруженного цепня.
^0
трихина; 21 — ланцетовидный сосальщ ик; 22 — печеночный сосальщ ик; 23 — кош ачий сосаль
■дик. 24
вооруженный цепень; 25 — эхинококк (ф и н н а ); 26 - невооруженный цепень; 27
лентец
широкий; 28 — карликовый цепень; 29 — москит; 30 — вош ь; 31 - клоп постельный; 32 — блоха,
муха
1ка
ми. Присутствие паразитов в ряде случаев вы зы вает тяжелые
нарушения. Так, паразитирование анкилостомид служит причиной не
только физического, но и умственного недоразвития детей.
Нейрохирурги оперативным путем удаляют финны свиного цепня
или эхинококка, поражающие мозг.
Недооценка лечащим
врачом
сопутствующих
паразитарных
болезней снижает эффективность лечения и м ож ет затягивать его на
длительные сроки (см. 13.4.5.). Так, при операциях на кишечнике
послеоперационные осложнения в определенном проценте случаев
вызваны нарушениями целостности кишечного шва аскаридами,
поскольку в результате хирургического вмеш ательства нарушается
химизм кишечника, что вы зы вает активные перемещения паразитов.
Следовательно, каждый врач обязан помнить о возможности
паразитарных заболеваний, сопутствующ их основному заболеванию
человека.
Многие паразитарные заболевания распространены повсеместно
(аскаридоз), зараженность в очагах может составлять 70— 90%
(описторхоз), в связи с чем некоторые инвазионные заболевания
становятся серьезным социальным фактором. К сожалению, стойкий
иммунитет ко многим паразитарным заболеваниям отсутствует и
возможны многократные повторные заражения.
Знание морфологии и цикла развития паразита составляет основу,
необходимую для изучения будущими врачами на последующих курсах
симптомов заболевания, вызванного паразитом, установления клиниче­
ского диагноза и способов лечения данной патологии.
Врачам санитарно-гигиенического профиля материал медицинской
паразитологии необходим для обоснования и осуществления профи­
лактических мероприятий.
Очерк по истории паразитологии. В самостоятельную дисциплину
паразитология оформилась во второй половине X I X века. В этот
период была экспериментально установлена роль комаров в переносе
возбудителей малярии, обнаружены возбудители балантидиаза, лейшманиозов, лямблиоза, амебиаза (у человека), пироплазмоза и кокцидио­
за (у млекопитающих и птиц). К том у времени были расшифрованы
жизненные циклы некоторых патогенных гельминтов.
Весомый вклад в развитие паразитологии внесли русские ученые.
А . Д. Федченко впервые описал жизненный цикл риш ты (Е)гасипси1и5
тесИпеп515); И . А . Порчинский опубликовал подробные данные о слеп­
нях, комарах, оводах, мухах, Н . А . Холодковский — о вшах. Изучению
комаров были посвящены работы В. В. Фавра, Н . М . Кулагина,
Г . А . Кожевникова.
При советской власти паразитология вступила в новый этап своего
развития. Мероприятия по профилактике и борьбе с паразитарными
заболеваниями стали проводиться на строго научной основе.
Руководитель ш колы советских гельминтологов Герой Социалисти­
ческого Труда лауреат Ленинской и Государственной премий академик
К . И . Скрябин (1879— 1972) организовал первый в мире институт
гельминтологии, которому еще при жизни выдающегося ученого было
присвоено его имя (Всесоюзный институт гельминтологии им. акад.
407
К И. Скрябин {1878
1972)
Е . Н. Павловский (1884
1965)
К . И . Скрябина), а также Лабораторию гельминтологии А Н С С С Р,
являющуюся основным теоретическим центром общей гельминтоло­
гии. Значительное место в исследованиях школы К . И . Скрябина
занимали практические мероприятия по борьбе с гельминтозами. Им и
его многочисленными учениками были разработаны методы диагно­
стики гельминтозов, изучены их клиника и терапия. По предложению
К. И. Скрябина, комплекс мероприятий, направленных на лечение
больного тем или другим гельминтозом, а также очищение внешней
среды от инвазивного материала получил название дегельминтизации.
К И . Скрябиным было сформулировано учение о девастации — ком
плексе мероприятий, направленных на полное уничтожение некото­
рых видов гельминтов, а также создание условий, при которых эти ви­
ды не смогли бы возникнуть вновь. Девастация подразумевает
уничтожение гельминта как зоологического вида.
Большое значение для развития паразитологии имеют труды акад.
Е . Н. Павловского (1884— 1965), под руководством которого успешно
разрабатывалось арахноэнтомологическое направление. И м была
обоснована теория об организме хозяина как среде обитания паразита и
теория паразитоценозов. Последняя успешно разрабатывается и углуб­
ляется в настоящее время акад. А . П . Маркевичем. Итогом изучения
переносчиков, их хозяев и путей циркуляции возбудителей в природе
стало учение Е. Н. Павловского о природной очаговости трансмиссив­
ных болезней.
Экологическая паразитология, изучающая зависимость фауны па­
разитов от биологического цикла и специфики физиологии их хозяев, а
также от факторов окружающей среды в нашей стране развивалась под
руководством лауреата Ленинской премии члена-корреспондента
А Н С С С Р проф. В . А . Догеля (1882— 1955). 408
За сравнительно короткий срок обширная территория нашей страны
стала одной из наиболее полно изученных в паразитологическом
отношении. Достижения в области паразитологии нашли широкое
применение в медицинской практике.
13.3 О С Н О В Н Ы Е П О Н Я Т И Я
П А РА ЗИ ТО Л О ГИ И
13.3.1. Экологические основы классификации паразитизма
Формы паразитизма в ппиполе чрезвычайно разнообразны.
Паразиты подразделяются на истинных и ложных, временных и посто­
янных.
И с т и н н ы е паразиты связаны с хозяином на значительном
протяжении (большем или меньшем) жизни Паразитический оора
жизни в данном случае обусловлен филогенетически и служит
специфическим признаком вида.
Ложный
паразитизм состоит в том, что отдельные особи
свободно живущего вида случайно попадают в организм особи другого
вида, где сохраняют некоторое время жизнеспособность и вызываю т
нарушения нормальной жизнедеятельности хозяина. Примером может
служить попадание личинок комнатной мухи или падальных мух в
кишечник человека.
Не всегда удается четко разграничить истинный и ложный
паразитизм. Например, у таких паразитов, как комары и москиты,
кровью питаются только самки и только в определенные периоды своей
жизни, в то время как самцы ведут свободный образ жизни.
В р е м е н н ы е паразиты (в основном кровососущие членистоно­
гие) только часть своей жизни находятся в непосредственном контакте с
хозяином, как правило, в процессе питания Длительность сроков
питания и степень связи с хозяином варьирует.
П о с т о я н н ы е паразиты подразделяются на относительно
постоянных и безусловно постоянных. Первые проводят на хозяине
только одну из фаз своего жизненного цикла (личиночную или
имагинальную), а в течение остальной части цикла ведут свободный
образ жизни.
Безусловно постоянные паразиты проводят всю жизнь на теле
хозяина или внутри его и не могут существовать во внешней среде. Пос­
тоянные паразиты могут бы ть связаны только с одним хозяином (вши)
или переходить от хозяина одного вида к хозяину другого вида В пос­
леднем случае передача паразита происходит непосредственно от
одного хозяина к другому без выхода во внешнюю среду. Так,
малярийный плазмодий передается от человека к комару при сосании
последним крови больного. Личинки трихинеллы, живущие в мышцах,
передаются непосредственно от свиньи к человеку.
Резкую границу между постоянным и временным паразитом также
трудно провести.
В зависимости от локализации паразитов в организме хозяина
различают паразитов наружных и внутренних.
Э к т о п а р а з и т ы (наружные) обитают на внешних покровах
хозяина (кожа, волосы). К ним относятся кровососушие насекомые и
клещи.
Э н д о п а р а з и т ы (внутренние) паразитируют во внутренних
органах, тканях, клетках, целомической полости хозяина и практически
могут поражать любой орган. Известны паразиты кишечника
(ленточные и круглые черви, простейшие), паразиты печени (сосальщи­
ки), паразиты легких (легочный сосальщик), мочеполовой системы
(шистозомы, простейшие) и т. д. В большинстве случаев локализация
тех или иных видов паразитов строго определена.
В то же время многие паразиты могут паразитировать в разных
органах хозяина. Например, возбудитель висцерального лейшманиоза
живет в печени, селезенке, костном мозгу и лимфатических узлах.
Личинки свиного цепня могут развиваться в мышцах, глазах, мозгу.
Один и тот же паразит может на разных стадиях развития поражать
различные органы. Так, малярийный плазмодий сначала развивается в
печени, а затем в эритроцитах человека.
13.3.2. Понятие о хозяине
\ Хозяева паразитов подразделяются на промежуточных, оконча­
тельных, резервуарных.
Хозяином паразита называют живой организм, используемый
паразитом как источник питания и место обитания. Ряд паразитических
форм эазные периоды жизненного цикла переходят от хозяина одного
вида к другому. Э то явление получило название смены хозяина.
Например, личиночные формы бычьего цепня развиваются в организме
крупного рогатого скота, а половозрелые формы только у человека.
В соответствии с э^им различают промежуточных и основных или
окончательных хозяев.
О к о н ч а т е л ь н ы м (основным или дефинитивным) хозяином
называют организм, в котором паразит находится в половозрелой
форме или размножается половым путем.
П р о м е ж у т о ч н ы м хозяином называют организм, в кото­
ром паразит находится в личиночной стадии или размножается
бесполым путем. Выделяют также категорию р е з е р в у а р н ы х
хозяев. В них паразит сохраняет жизнеспособность, происходит его'
накопление, что повышает выживаемость вида, но дальнейшее развитие
его не происходит.
13.3.3. Происхождение паразитизма
«Явление паразитизма имеет многообразное происхождение. Даже
один и то т же вид паразитизма, например, эктопаразитизм, у разных
животных мог возникнуть разными путями» (В. А . Догель).
Наиболее изучен переход свободноживуших форм к эктопарази­
тизму. Большинство эктопаразитов перешло к паразитизму от
хищничества путем постепенного удлинения сроков питания и контакта
с хозяином. В процессе эволюции можно проследить увеличение
продолжительности сроков питания. Самки москита питаются меньше
410
минуты, личинка иксодовых клещей — 5— 8 дней, а половозрелые
формы — от 5 до 14 дней, блохи проводят на хозяине большую часть
своей жизни, вши — всю жизнь.
Второй путь перехода к паразитизму можно проследить на примере
птичьих клешей-пухоедов. В этом случае паразитизм возникает на
основе комменсализма. Вероятно, предки клещей-пухоедов были ком­
менсалами птиц, обитавшими в гнезде, где они питались органическими
остатками, скапливающимися на дне гнезда. Постепенно клещи стали
переходить с подстилки на хозяев гнезда и питаться ороговевшими
придатками кожи и в конечном счете превратились в эктопаразитов.
Объяснить превращение свободноживущих форм в эндопаразитов
значительно труднее.
В отдельных случаях эндопаразитизм происходит из эктопарази­
тизма. Например, пухоед пеликана с перьев перешел в подклювный
мешок, где стал питаться кровью. Филогенетическая связь с эктопара­
зитизмом прослеживается в том, что яйца пухоеды откладывают
на перья.
В ряде случаев возможно происхождение эндопаразитизма,
например, кишечного паразитизма, из комменсализма. Кишечная среда
выгодна для обитания многих форм, поскольку она богата пищевыми
веществами. Среди комменсалов есть группа так называемых пищерасхитителей. Они используют не отходы пиши, а известную долю
рациона хозяина, потребляя пищу, подготовленную им для усвоения, и,
таким образом, причиняют ему косвенный ущерб. С увеличением
количества потребляемой пиши, например за счет увеличения поверхно­
сти тела у ленточных, комменсальная форма начинает приносить
прямой вред и переходит в паразитическую.
Однако основная масса случаев внутреннего паразитизма, особенно
кишечного, согласно В. А . Догелю, представляет собой первичное
явление, возникшее благодаря случайному заносу в пищеварительную
систему яиц или покоящихся стадий различных свободно живущих
организмов.
13.3.4. Взаимоотношения в системе паразит— хозяин
на уровне особей
Паразит и хозяин составляют взаимосвязанные элементы единой
биологической системы «паразит— хозяин», обитающие в конкрет­
ных условиях среды.
Для конкретизации их взаимодействия рассмотрим раздельно
действия паразита на хозяина, хозяина на паразита и воздействие среды
на обоих партнеров.
Паразит обычно оказывает вредоносное действие на хозяина,
вызывая заболевание. Такое свойство паразита обозначают как
патогенност ь
(от греч рагНох — страдание, ^епемх — разви­
тие). В противоположность этому формы, не обладающие способно­
стью вы зы вать заболевание, -называются н е п а т о г е н н ы м и .
Действие паразита на хозяина. Воздействие паразита на хозяина
разнообразно. Механическое воздействие проявляется в виде поврежде­
411
ний тканей, наносимых органами прикрепления (крючья, присасыва­
тельные щели, присоски) или частями ротового аппарата паразита.
Ряд крупных кишечных паразитов (широкий лентец, аскарида и др.)
могут вы звать закупорку просвета и даже разрыв кишечника. Желчные
протоки печени нередко закупориваются печеночными сосальщиками.
Наряду с половозрелыми особями механические повреждения могут
причинять личиночные формы, проникая через неповрежденную кожу
(личинки кровяных сосальщиков).
Яйца кровяных сосальщиков, имеющие своеобразный шип, проходя
через стенки сосудов и слизистую оболочку мочевого пузыря или стенку
кишечника, сильно их травмируют. Скопление яиц паразитов в печени,
занесенных током крови, вызывает цирроз печени.
Т о к с и ч е с к о е. действие оказывают продукты жизнедея­
тельности паразита. Например, выход продуктов диссимиляции
малярийных плазмодиев из эритроцитов в плазму вызывает лихора­
дочные приступы у больного. С помощью многочисленных экспери­
ментов доказано, что полостная жидкость и экстракты тканей аскариды
и других червей обладают высокой токсичностью. При введении в вену
собаки полостной жидкости человеческой или лошадиной аскариды,
наблюдается снижение кровяного давления, одышка, торможение
свертывания крови, что в конечном итоге вызывает смерть жи­
вотного.
Токсическое действие гельминтов проявляется у больного в потере
аппетита, уменьшении массы тела, малокровии, повышенной утомляе­
мости, бессоннице, тошноте, рвоте, снижении работоспособности,
поносе и т. д. Токсические продукты жизнедеятельности анкилостом
вызываю т у детей задержку не только физического, но и умственного
развития.
П и т а н и е п а р а з и т о в п р о и с х о д и т за счет хозяина. Паразиты
поглощают тканевую жидкость, ткани, кровь, а также переваренную
пищу. Особенно много пищи поглощают кишечные гельминты, в
частности ленточные черви, имеющие в длину несколько метров.
Развивающийся бычий цепень увеличивается за месяц в длину на
несколько метров, а в день на 7— 10 см. Для осуществления таких
темпов роста гельминту необходимо большое количество пищи, что
может вызвать резкое истощение больного.
При паразитировании анкилостом, питающихся кровью, развива­
ется тяжелое малокровие. Широкий лентец поглощает боль­
шое количество витамина В 12 , что приводит к злокачественному
малокровию.
Патогенное
действие
некоторых
гельминтов
связано
с
м и г р а ц и е й ^ передвижением) личиночных форм по организму
хозяина. В процессе миграции личинки разрушают ткани, вызываю т
воспалительные процессы, способствуют проникновению инфекции,
оказывают токсическое действие.
Личинка аскариды пробуравливает стенку кишечника, попадает в
ток крови и последовательно проходит в печень, правое сердце, лег­
кие, бронхи, трахею, глотку, откуда заглатывается в кишечник. При
интенсивном заражении в печени возникают множественные кровоизли­
412
яния и некрозы, развиваются соединительнотканные прослойки, в
легких наблюдается очаговая пневмония.
Р а с п р о с т р а н е н и е возбудителей инфекционных и инвази­
онных заболеваний осуществляется кровососущими эктопаразитами
Твши, блохи, клещи). Исключительно важное значение для медицины
имеет распространение возбудителей заболеваний человека, имеющих
массовый характер, или эпидемических.
Как правило, паразит одновременно действует на хозяина
различными путями, но обычно не вызывает смерти хозяина, поскольку
гибель хозяина означает и гибель паразита. В процессе эволюции
системы паразит — хозяин вырабатываются взаимные приспособле­
ния, обеспечивающие относительную устойчивость этой системы.
Действие хозяина на паразита. Организм хозяина также оказывает
влияние на паразита, направленное на подавление его жизнедеятельно­
сти или уничтожение.
Ответные реакции хозяина можно разделить на клеточные,
тканевые (местные), гуморальные (общие).
К л е т о ч н а я р е а к ц и я проявляется, например, в увеличении
размеров клетки. Например, эритроциты, пораженные малярийным
плазмодием, заметно крупнее «здоровых».
Т к а н е в а я р е а к ц и я проявляется в образовании вокруг
покоящейся стадии паразита соединительнотканной капсулы, которая в
той или иной мере изолирует паразита от окружающих тканей.
Г у м о р а л ь н ы е р е а к ц и и являются иммунологическими и
заключаются в выработке защитных специфических антител в ответ на
поступление антигенов, вырабатываемых паразитом.
Механизмы и формы иммунитета при паразитарных заболеваниях в
принципе не отличаются от иммунитета при бактериальных и вирусных
инфекциях. При заражении многими простейшими (лейшмании,
трипаносомы) развивается стойкая невосприимчивость, другие протозойные заболевания (амебиаз, трихомонадоз) не дают длительного
приобретенного иммунитета.
Иммунитет при гельминтозах изучен недостаточно. Установлено,
что наряду со сходством защитных реакций имеется ряд особенностей,
связанных со спецификой строения и биологией гельминтов.
Основное отличие состоит в кратковременности иммунологических
реакций и слабой степени напряженности иммунитета, хотя в случаях
трихинеллеза и шистозоматоза наблюдается стойкий иммунитет.
Относительный характер иммунитета объясняют отсутствием размно­
жения гельминтов в организме хозяина, крупными размерами и слож­
ным строением их, что препятствует тесному контакту паразита с
клетками тканей хозяина. Затрудняет выработку иммунитета и смена
стадий паразита (яйцо, личинка, половозрелая форма), поскольку
антигены каждой стадии специфичны. Наиболее интенсивный иммуни­
тет развивается во время присутствия личиночных стадий, поскольку
личинка имеет более тесный контакт с тканями хозяина и выделяет ан­
тигены большей активности.
За счет относительного иммунитета в очаге снижается как
интенсивность (число паразитов у местного населения в среднем ниже,
413
чем у приезжих), так и экстенсивность (процент зараженности местного
населения ниже, чем среди приезжих) заражений. Имеются данные, что
в очагах аскаридоза до 25% заражений прерывается в ранней фазе, сразу
после окончания миграции паразитов, т. е. половозрелые формы не
образуются.
Таким образом, иммунитет при гельминтозах вырабатывается, но
не обеспечивает полной невосприимчивости к повторным заражениям, а
лишь снижает продолжительность жизни паразитов и их плодовитость.
Отсутствие стойкого иммунитета означает возможность многократных
повторных заражений, что обусловливает особенности профилактиче­
ских мероприятий.
Вопрос искусственной иммунизации людей против гельминтов пока
не разработан, хотя в эксперименте получены данные о возможности
создания такого иммунитета.
Влияние внешней среды на систему паразит — хозяин на уровне
особей. Взаимодействие паразита и хозяина происходит в конкретных
условиях среды. Организм хозяина служит внешней средой для
паразита. На хозяина, в свою очередь, влияет окружающая его среда,
которая опосредованно оказывает влияние и на паразита. Для человека
внешняя среда включает в себя как природные, так и социальные факто­
ры. Известно, что неблагоприятные изменения физиологического
состояния — охлаждение, перегревание, перенесенные заболевания,
голодание, переутомление, психические травмы, — возникающие под
влиянием окружающей среды, способствуют активизации паразитов.
Гак, непатогенная комменсальная форма дизентерийной амебы при
перечисленных условиях превращается в патогенную, вызывающую
тяжелое заболевание — амебиаз. Как показали наблюдения, переход
амеб в патогенную форму часто наблюдается у раненых, поэтому в
военное время численность больных амебиазом резко увеличивается.
Переход непатогенных форм амебы в патогенные идет также гораздо
интенсивнее в районах с жарким климатом, чем в районах с умеренным
климатом.
Таким образом, конечный результат взаимодействия паразита и
хозяина зависит от следующих факторов: видовых особенностей
паразита и хозяина; индивидуальных качеств данной пары партнеров;
конкретных особенностей среды обитания паразита и хозяина на фоне
их филогенетического развития.
13.3.5. Паразитоценоз
В организме хозяина одновременно могут присутствовать паразиты
разных видов. Они вступают во взаимодействие друг с другом,
вследствие чего различные комбинации видового состава паразитов
могут как усиливать, так и ослаблять совместное отрицательное
воздействие.
Совокупность паразитов, населяющих организм хозяина или его
отдельные органы, получила название п а р а з и т о ц е н о з .
М ысль о том, что паразиты разных видов находятся во взаимодей­
ствии, была впервые высказана Е . Н . Павловским и затем подтвержде­
414
на многочисленными исследованиями его сотрудников. Им же предло­
жен термин «паразитоценоз». Наиболее изучены связи в паразитоценозах кишечника, в частности между гельминтами и простейшими. По
имеющимся данным, у лиц, зараженных аскаридозом и анкилостомидозами, процент сопутствуюшего заражения лямблиями в 2—4 раза ниже,
чем у людей, не зараженных этими видами гельминтов. В отношении
ленточных червей получены противоположные результаты — при
наличии карликового цепня и широкого лентеца зараженность
лямбиозом в 2— 3 раза выше, чем у людей, не страдающих гельминтозами.
Установлена взаимосвязь между простейшими, гельминтами и
бактериями кишечника.
Известно, что бактериальная дизентерия у больных, страдающих
аскаридозом и другими гельминтозами, протекает более тяжело и
плохо поддается лечению. Поэтому у детей, больных дизентерией,
положено проводить обследование на гельминтозы с последующей
дегельминтизацией.
У инвазированных кишечными гельминтами больных тяжелее
протекает туберкулез, болезнь Боткина, брюшной тиф и другие
инфекционные болезни.
Знание взаимосвязей внутри паразитоценозов и их характера очень
важно для врача, так как позволяет ему повысить эффективность
лечения.
Дальнейшие исследования внутривидовых и межвидовых соотноше­
ний между компонентами различных паразитоценозов следует считать
актуальной задачей современной эволюционной и экологической па­
разитологии. В настоящее время паразитоценология в нашей стране
выделилась в самостоятельную науку, которая успешно развивается.
13,3.6. Морфофизиологические адаптации
к паразитическому образу жизни
Переход к паразитическому образу жизни сопровождается
появлением ряда приспособлений паразита к новым условиям существо­
вания.
Как правило, у паразитов развиваются органы прикрепления или
фиксации, так как большинство паразитов, выйдя из организма
хозяина, не может попасть к другому хозяину и погибает. Примером
органов фиксации могут служить коготки вшей, присоски гельминтов,
ротовой аппарат клешей и т. д.
У кровососущих характерно увеличение вместимости пищевари­
тельной системы за счет появления слепых выростов кишечной трубки
(пиявка, клеши). Кишечные паразиты обладают антиферментными
свойствами, защищающими их от переваривания в кишечнике. Наконец,
для паразитов характерна высокая плодовитость, поскольку в процессе
цикла развития огромное количество личиночных форм погибает, не
достигая половой зрелости. К ак компенсация гибели личиночных форм
у паразитов наблюдается мощное развитие половой системы и образо­
вание огромного количества половых продуктов.
415
У сосальщиков, кроме того, гибель зародышей компенсируется
размножением лцчиночных стадий путем партеногенеза. В результате
из одного яйца может образоваться множество инвазионных стадий.
Развитие одних органов сочетается у паразитов с исчезновением других,
не играющих существенной роли в новых условиях существования. Так,
постоянство внутренней среды обитания приводит к редукции органов
чувств (глаза и др.), исчезновению органов передвижения, у ленточных
червей отсутствует пищеварительная система.
Г л а в а 14
П У ТИ Ц И РК У Л Я Ц И И ВО ЗБУ Д И ТЕЛ ЕЙ
ЗА БО Л ЕВА Н И Й В П РИ РО Д Е
Для сохранения вида экто- и эндопаразиты в процессе размножения
покидают организм хозяина и переходят во внешнюю среду или в
организм другого хозяина.
Круг возможных хозяев и механизм передачи паразита от одного
хозяина к другому или от хозяина во внешнюю среду и снова к хозяину
определяют пути циркуляции паразитов и вызываемые ими забо­
левания.
14.1. РЕЗЕРВУАРЫ И ПЕРЕНОСЧИКИ
И с т о ч н и к о м возбудителей болезней может бы ть человек или
животное, в организме которого паразит обитает, размножается, а
затем передается новым хозяевам.
Организм, в котором возбудитель сохраняется длительное время,
называется р е з е р в у а р о м . Для возбудителей одних заболеваний
резервуаром служит человек (малярия, эпидемический сыпной и воз­
вратный вшивый тиф и др.), для других — животные. Диких животных
называют п_р и р о д н ы м р е з е р в у а р о м . Н апример, грызуны
служат природным резервуаром лейшманиоза, чумы, туляремии.
Некоторые виды клешей могут длительное время (до 20 лет) хранить
возбудителей возвратного клещевого тифа, клещевого энцефалита,
туляремии.
Кроме резервуаров, для циркуляции многих возбудителей необходи­
мы . п е р е н о с ч и к и , роль которых выполняют кровососущие
членистоногие (насекомые и клещи). В результате активных перемеще­
ний переносчиков возбудители заболеваний могут распространяться на
значительные расстояния.
Различают
специфических
и
механических
переносчиков.
С п е ц_И-ф^и ч е с к и м и п е р е н о с ч и к а м и называют тех чле­
нистоногих, в организме которых возбудитель проходит цикл развития.
Поскольку между возбудителями и переносчиками существует биоло­
гическая связь, то обычно выполнять роль последнего могут только
организмы одного вида или рода (малярийный плазмодий —
малярийный комар.
М е х а н и ч е с к и м и п е р е н о с ч и к а м и называю т члени­
416
стоногих, в организме которых возбудитель не проходит цикла
развития, а лишь перемещается с помощью их в пространстве. Так, на
наружных покровах, лапках, а также в кишечнике комнатной мухи
могут находиться возбудители различных заболеваний. Механическими
переносчиками одного и того же возбудителя могут бы ть членистоно­
гие различных видов (комнатная муха, таракан, домовая муха).
Некоторые переносчики могут бы ть и природным резервуаром.
Например, аргазовые клеши могут сохранять возбудителей возвратно­
го тифа в своем организме до 20 лет. Блоха может при температуре
37 °С сохранять возбудителей чумы до 27 дней, а при температуре
0— 5 °С — 358 дней.
В процесс циркуляции возбудителей заболеваний могут вовлекаться
животные многих видов, связанные между собой биоценотическими
связями.
14.2. Т Р А Н С О В А Р И А Л Ь Н А Я
П ЕРЕД АЧА
ВО ЗБУД И ТЕЛЕЙ
Исключительно важную роль в удлинении сроков хранения воз­
будителей заболеваний переносчиками играет механизм трансовари­
альной передачи, т. е. передача возбудителей через яйцевые клетки от
одного поколения к другому. Циркулируя в организме переносчика,
возбудитель может попадать в различные ткани и органы, в том числе в
его яичник и в яйцеклетки. При оплодотворении такой яйцеклетки
происходит нормальное развитие, но все стадии его сохраняют в себе
возбудителя, а развившаяся самка в свою очередь будет передавать его
последующим поколениям. В лаборатории Е . Н. Павловского была в
свое время прослежена трансовариальная передача аргазовыми клеща­
ми возбудителей возвратного тифа трем последующим поколениям,
теперь обнаружена передача риккетсий иксодовыми клещами двенадца­
ти поколениям. В настоящее время обнаружена трансовариальная
передача возбудителей двум поколениям у москитов.
14.3. Э К О Л О Г И Ч Е С К И Е О С Н О В Ы
КЛАССИ ФИ КАЦ И И БО ЛЕЗН ЕЙ
14.3.1. Инвазионные и инфекционные болезни
Болезни человека и животных классифицируют прежде всего по
причинам, их вызывающим. Различают инфекционные и инвазионные
(паразитарные) заболевания.
Инфекционные
заболевания вызываю тся различными
микроорганизмами — бактериями, спирохетами, вирусами, грибками.
И н в а з и о н н ы е или паразитарные болезни вызываю тся
патогенными простейшими (протозоозы), гельминтами (гельминтозы),
насекомыми (энтомозы).
Название паразитарных болезней образуется из корней родового
названия паразита и суффикса «аз», «оз», «ез». Например, заболевание,
вызываемое лейшманиями, — лейшманиоз, трематодами — трематодоз, дизентерийной амебой — амебиаз и т. д.
417
15— 56
Кроме того, болезни классифицируют по способу передачи бо­
лезнетворного начала. Так, различают п и ш е в ы е — паразитарные
болезни, возбудители которых попадают в организм хозяина через рот
вместе с пищей, водой или через грязные руки (дизентерия,
гельминтозы, брюшной тиф), и к о н т а к т н ы е — возбудители
проникают через неповрежденную кожу или слизистые при соприкосно­
вении с зараженными предметами, почвой или водой. Проникать таким
путем из почвы могут личйнки нематод (анкилостомы). Чесоточный
зудень переходит к здоровому человеку через рукопожатие с больным
и т. д.
14.3.2. Трансмиссивные болезни
Возбудители трансмиссивных заболеваний передаются посредством
переносчиков. К ним относятся как паразитарные, так и инфекционные
болезни. Различают облигатно-трансмиссивные и факультативно­
трансмиссивные заболевания.
О б л и г а т н о-т р а н с м и с с и в н ы е болезни передаются от
одного хозяина к другому только через переносчика. Например,
малярией или сыпным тифом человек может заразиться только через
укус насекомого, так как возбудитель должен попасть в кровь.
Ф а к у л ь т а т и в н о-т р а н с м и с с и в н ы е болезни могут
передаваться какчерез переносчика, так и без него другими путями, т. е.
участие переносчика не обязательно. Примером таких заболеваний
могут служить туляремия и чума. Возбудители туляремии могут
передаваться от зараженных животных к человеку как с помощью
кровососущих членистоногих, так и через загрязненные фекалиями
пищевые продукты и воду. Чум а может передаваться человеку от
грызунов через укус блох, а также контактным путем при снятии шку­
рок с больных животных, при соприкосновении с больным человеком.
Специфические связи между возбудителем и хозяином позволяют
выделить следующие группы трансмиссивных заболеваний:
— з о о н о з ы — болезни, свойственные только животным (маля­
рия птиц);
- а н т р о п о з о о н о з ы — болезни, возбудители которых мо­
гут поражать как животных, так и человека. В этом случае переносчик
может передавать возбудителя от животных к человеку и обратно
(таежный энцефалит, лейшманиоз, чума);
— а н т р о п о н о з ы — болезни, которые свойственны только
человеку (трихомонадоз, амебиаз).
Таким образом, переносчикам принадлежит исключительно важная
роль в распространении многих, иногда принимающих массовый
характер, эпидемических заболеваний.
14.3.3. Природноочаговые заболевания
Основоположником учения о природной очаговости заболеваний
является Е . Н . Павловский. На основании экспедиционных, полевых,
лабораторных и экспериментальных работ в 1939 г. им была выделена
новая категория заболеваний — заболевания с природной очаговостью.
418
Заболевания этой группы имеют ряд характерных особенностей
1) циркулируют в природе независимо от человека; 2) резервуаром слу­
жат дикие животные, составляющие с возбудителями и переносчиками
биоценотический комплекс; 3) распространены не повсеместно, а на
ограниченной территории большей или меньшей протяженности, с
определенным географическим ландшафтом, что связано с ареалом
распространения компонентов биоценоза.
Примером заболевания с природной очаговостью служит таежный
весенне-летний энцефалит. Дикие животные (бурундук, белка, заяц, еж,
птицы), зараженные вирусом энцефалита, обитают в тайге и составля
ю т природный резервуар. Иксодовые клеши, паразитирующие на
животных, передают вирус энцефалита от одного животного к другому,
благодаря чему в природе возбудители циркулируют в течение многих
лет независимо от человека. Животные-резервуары, переносчики и
возбудители заболевания имеют свой видовой ареал распространения,
поэтому энцефалит встречается только на определенной территории и
не встречается за ее пределами. Территорию, на которой обитают дикие
животные, служащие резервуаром возбудителя и существование
которых не связано с деятельностью человека, а также переносчики,
называют п р и р о д н ы м о ч а г о м з а б о л е в а н и я .
Таким образом, условиями возникновения и существования очага
служит присутствие возбудителя, восприимчивого к возбудителю
животного-резервуара, переносчика и соответствующих природных
(климатических, ландшафтных) условий, обеспечивающих существова­
ние компонентов данного биоценоза. Так, природный очаг таежного
энцефалита представляет собой участок неосвоенной тайги с соответ­
ствующими обитателями. Природный очаг лейшманиоза — это часть
территории полупустыни или пустыни, населенная песчанками,
зараженными лейшманиями, и москитами, обитающими в их
норах.
Существующий природный очаг потенциально опасен для человека.
Если человек оказывается на его территории (геологическая разведка,
охота, экспедиция, лесоразработки и т. д.), то ему может бы ть передан
возбудитель антропозоонозов и человек заболевает.
Зараженность животных-резервуаров и переносчиков на территории
очага может бы ть весьма значительной. Оказалось, что зараженность
песчанок в очагах лейшманиозов достигает 6 0 % , а москитов в норах
3—6 % , крысы заражены спирохетами возвратного тифа более чем в
20% случаев.
Важную роль в длительности существования очагов играет
упомянутая выше способность переносчиков (клещей, насекомых) к
трансовариальной передаче возбудителей заболеваний. Наряду с биоти­
ческими факторами в существовании очага определяющую роль, как
уже отмечалось, играют климато-географические природные условия.
Так, развитие и размножение вируса японского энцефалита в комарах
возможно при температуре не ниже 21—22°С . Поэтому природные
очаги энцефалита встречаются только в районах с теплым климатом,
хотя ареалы комаров гораздо шире.
Иногда один и тот же ландшафт может включать очаги различных
заболеваний. Например, норы грызунов полупустынь и пустынь могут
быть очагами лейшманиоза,клещевого возвратного тифа и чумы.
Первоначально природная очаговость была установлена по
отношению только к трансмиссивным заболеваниям, таким, как
таежный весенне-летний энцефалит, клещевой возвратный тиф,
лейшманиоз и др. Позднее выяснилось, что природно-очаговый
характер имеют заболевания, распространяемые без участия переносчи­
ка, — нетрансмиссивные, в том числе гельминтозы, такие, как
огшеторхоз, парагонимоз, дифиллоботриоз и др. В природных очагах
не трансмиссивных заболеваний (токсоплазмоз, бешенство, гельминто­
зы) передача возбудителя осуществляется в основном пищевым и
контактным путем. Перечень заболеваний с природной очаговостью в
настоящее время расширился. Сюда относятся чума, туляремия,
клещевой возвратный тиф, лейшманиоз, таежный энцефалит, геморра­
гические нефрозо-нефриты, многие гельминтозы и т. д.
Учение Е. Н. Павловского о природной очаговости заболеваний
сыграло огромную роль в дальнейшем развитии медицинской паразито­
логии. Оно определило совершенно новый подход к профилактике этой
группы заболеваний. Если раньше основной мерой предупреждения
заболеваний считалось лечение больных и уничтожение переносчиков,
теперь основной целью стало уничтожение животных-резервуаров.
Эффективность такого подхода можно показать на примере снижения
частоты заболеваемости лейшманиозом. После уничтожения грызунов
и обитающих в их норах москито&число больных в очагах снизилось с
90 до 0,04%.
■ Учение о природной очаговости заболеваний нашло быстрое
признание во всем мире и принято Всемирной Организацией Здраво­
охранения. Во многих странах выявлены специфические для них
заболевания с природной очаговостью (болезнь Чагаса в Южной
Америке, трипаносомозы в Африке, шистозоматозы, желтая лихорадка
и т. д.).
Глава
15
ЭКО Л О ГИ Ч ЕС КИ Е А С П ЕКТЫ
П А Р А ЗИ Т О Л О ГИ И
П а р а з и т о л о г и я — наука экологического профиля, посколь­
ку паразитизм является одной из форм межвидовых отношений. Вместе
с тем экологическим аспектам паразитологии (расселение паразитов,
поиски хозяина, взаимоотношения паразита и хозяина) уделяется
меньше внимания, чем морфологии, систематике паразитов, изучению
их циклов развития и т. д.
Среди советских паразитологов вопросами экологии паразитов
занимались Е. Н Павловский, К . И. Скрябин, В. Н . Беклемишев и ос­
нователь советской экологической школы паразитологов В. А. Догель,
труды которых внесли существенный вклад в развитие этого
направления.
420
15.1. в з а и м о о т н о ш е н и я В С И С Т Е М Е
.
П А Р А З И Т — Х О З Я И Н НА У Р О В Н Е П О П У Л Я Ц И Й
Д ля современной экологической паразитологии характерен переход
от изучения отношений в системе паразит — хозяин н а у р о в н е
о т д е л ь н ы х о с о б е й к изучению этого вопроса н а у р о в н е
популяций.
Развивающаяся популяционная экология паразитов уже накопила
интересный фактический материал, позволяющий дать ответ на ряд
вопросов, остававшихся нерешенными.
15.2. З Н А Ч Е Н И Е П А Р А З И Т И З М А К А К Ф А К Т О Р А
У С Т О Й Ч И ВО С Т И С И С Т Е М Ы П А Р А З И Т — ХО ЗЯИ Н
Подход к изучению проблемы паразитизма на популяционном
уровне позволил выявить ряд новых закономерностей, среди которых
важно отмстить неодинаковую биологическую значимость паразитизма
для отдельных особей хозяина и для популяции хозяина.
На уровне отдельных особей паразит, как правило, приносит вред
организму хозяина, ослабляет его жизнедеятельность иногда даже
убивая его. В популяции хозяина гибель части особей под воздействием
паразитов оказывается не вредной, а «полезной» для этой группы
организмов. Например, при возрастании скорости размножения
количество особей в популяции хозяина может превысить пищевые
ресурсы, имеющиеся в ее ареале, и вызвать самоуничтожение
популяции. Благодаря гибели части животных под воздействием
паразитов вновь восстанавливается оптимальное соотношение между
числом особей популяции и конкретными возможностями среды
обитания. В результате популяция хозяина сохраняется, т. е. ее
устойчивость повышается. Одновременно выживает и популяция
паразита. Следовательно, паразитизм, регулируя численность популя­
ции хозяина, повышает устойчивость системы паразит— хозяин.
Э т о т регулирующий механизм вступает в действие при опреде­
ленных условиях — бескормица, авитаминоз, повышение плотности
популяции и др. Главную роль при этом играют наиболее патогенные
паразиты, вызывающие массовые острые заболевания животных или
эпизоотии.
Популяции хозяина являются составным компонентом различных
биогеоценозов. При резком повышении их плотности происходит
разрушение нижележащих трофических уровней биоценоза, что наруша­
ет равновесие между элементами экосистемы. Так, при резком
увеличении численности насекомых, гусеницы которых питаются
листьями, последние могут уничтожить листву деревьев полностью.
Вызванная дефолиация означает разрушение первичного трофического
уровня, что имеет тяжелые последствия для всех элементов экоси­
стемы
Таким образом, согласно новым данным, паразитизм следует
рассматривать как экологический фактор, который регулирует числен­
ность популяции хозяина и повышает устойчивость не только простых
421
систем паразит— хозяин на уровне популяции, но и экосистем более
сложного уровня — биоценозов.
С этой точки зрения представление о паразитарном заболевании как
отрицательном явлении по отношению к природным биоценозам
должно бы ть пересмотрено как не соответствующее сущности явления
(В. Н. Беклемишев). Однако следует сразу отметить, что подобное
значение паразитизма ни в коей мере не распространяется на популяции
современного человечества.
Новые представления о «полезной» роли паразитизма как
возможного универсального фактора, регулирующего устойчивость
экосистем, позволили объяснить широкое распространение парази­
тизма и относительную устойчивость систем паразит—хозяин.
Получил объяснение и другой вопрос, а именно почему у хозяев
паразитов,
принадлежащих
к
высокоорганизованным
группам
животного мира и обладающих хорошо сформированными защитными
реакциями, в процессе эволюции не выработалась зашита от
паразитизма. Стало очевидно, что подобные приспособления не могли
закрепляться в процессе естественного отбора. В освободившихся от
паразитов популяциях хозяев численность особей стала бы возрастать,
что привело бы к быстрому разрушению элементов среды, необходи­
мой для популяции, в частности к истощению пищевых ресурсов и
жизненного пространства. По данным некоторых авторов у млекопи­
тающих, наоборот, обнаружены приспособления, облегчающие парази­
тизм.
Современный регулирующий численность популяций хозяев меха­
низм паразитизма является результатом длительной эволюции экоси­
стем. В соответствии с разнообразием паразитов, их циклов и парази­
тических связей, паразитизм представляет собой гибкий и быстро
реагирующий фактор экосистем.
Как уже указывалось выше, по отношению к популяциям человека в
настоящее время паразитизм не играет положительной роли, хотя,
безусловно, биологическая эволюция человека также контролировалась
паразитами, поддерживавшими численность популяций людей в со­
ответствии с экологическими возможностями среды. Считаю т, что на
ранних этапах цивилизации рост популяции человечества еще контроли­
ровался паразитарным регулирующим механизмом и возникавшие
эпидемии вызывали гибель больших масс людей. Однако с развитием
хозяйственной деятельности человека и увеличением производства
продуктов питания численность популяций человека стала расти, что
привело к нарушениям устойчивости системы. На более поздних
этапах цивилизации обществу пришлось организовывать защиту от
действия паразитарного механизма и разработать для этого систему
лечебных и профилактических мероприятий.
Однако в развивающихся странах, особенно в тропических, до
настоящего времени паразитарные заболевания служат причиной
высокой смертности среди населения. По данным ВОЗ, около 400 млн.
человек поражены гельминтами, в связи с чем ежегодно погибают
свыше 1 млн. детей.
Человек научился использовать паразитический механизм в своих
422
целях. Искусственно разводя паразитических насекомых, человек затем
использует их для подавления резких вспышек численности насекомыхвредителей.
15.3
Р А С П Р Е Д Е Л Е Н И Е П АРАЗИ ТО В
В П О П УЛ ЯЦ И И ХО ЗЯИ НА
В пределах одной популяции степень зараженности разных особей
неодинакова. Как показали исследования, на уровне популяций
распределение паразитов между особями хозяина соответствует не
случайному или нормальному, а так называемому персрассеянному
типу распределения, т. е. паразиты сосредоточены лишь в немногих
особях хозяина, но в большом количестве.
Наиболее частая причина неоднородности заражения особей хозяина
заключается в различной вероятности инвазии для различных особей
хозяина и различных особей паразита. Для того чтобы распределение
паразита в популяции хозяина было случайным, вероятность инвазии
должна бы ть одинаковой для всех особей хозяина и всех особей парази­
та, т. е. среда должна бы ть однородной. Поскольку в природе такие
условия отсутствую т, возникают отклонения в концентрации паразитов
в отдельных особях.
Второй причиной перерассеянного распределения паразита является
возможность бесполого или партеногенетического размножения пара­
зита в организме хозяина (малярийный плазмодий в человеке,
трематоды в промежуточных хозяевах и др.).
И, наконец, суперинвазия становится возможной вследствие
ослабления защитной реакции хозяина, что повышает вероятность его
последующих повторных заражений.
Псрерассеянное распределение паразита в популяции необходимо
учитывать при разработке мер борьбы и профилактики паразитарных
заболеваний.
15.4. Р А С С Е Л Е Н И Е И П О И С К Х О З Я И Н А
Расселение животных необходимо для избежания перенаселенности,
повышающей смертность, и поэтому служит важным условием
сохранения вида. Особенно важно расселение для паразитов, поскольку
супсринвазия приводит к смерти хозяина и гибели паразита. Поэтому
паразиты обладают способностью покидать на определенной стадии
развития хозяина и находить новых хозяев.
Переход от одного хозяина к другому, обеспечивая расселение
паразита, одновременно влечет за собой высокую его смертность,
поскольку вероятность нахождения нового хозяина, как правило, крайне
мала. Гибель большого числа особей на разных стадиях цикла развития
паразита компенсируется высокой плодовитостью паразитических
форм.
Расселение паразитов происходит различными способами и на
различных стадиях жизненного цикла.
Наиболее часто переход от одного хозяина к другому осуществляет­
423
ся с помощью свободноживущей (находится вне организма хозяина),
активной подвижной (личинка) и неактивной покоящейся (яйцо, циста,
спора) стадий. Последняя имеет защитные приспособления (оболочка),
обеспечивающие переживание неблагоприятных условий, и обычно
неподвижна. Благодаря этому она не может обеспечить расселение в
пространстве, но позволяет продлить срок существования паразита,
т. е. обеспечивает его расселение во времени, что повышает шансы на
встречу с новым хозяином.
Активная подвижная стадия (церкарий, рабдитная, филяриевидная
личинка) способствует расселению паразита в пространстве, обычно не
питается и живет недолго. Кроме функции расселения, она выполняет и
функции поиска хозяина.
Включение в цикл развития промежуточного хозяина дает
возможность паразиту избежать неблагоприятных воздействий среды и
продлить срок своего существования.
Расселению паразита способствует также перемещение хозяев.
Особенно эффективно перемещение окончательного хозяина, содержа­
щего половозрелые формы и рассеивающего яйца по территории
обитания.
Если цикл развития паразита включает переносчика, то функцию
поиска нового хозяина выполняет переносчик. При наличии в цикле
промежуточного хозяина паразит, как правило, не участвует в поиске
окончательного хозяина, а попадает к нему благодаря существующим
пищевым связям.
Паразиты могут изменять поведение промежуточных хозяев таким
образом, что шансы попасть к окончательному хозяину будут
повышаться. Например, муравьи, зараженные метацсркариями трема­
тоды, выползают на кончик травинки и здесь впадают в состояние свое­
образного оцепенения, что обеспечивает заглатывание их травоядными
животными. Ры бы , зараженные некоторыми видами цестод, значи­
тельно медленнее плавают, отчего становятся добычей хищников в
первую очередь.
У многих паразитов вероятность встречи с хозяином облегчается за
счет синхронизации их цикла развития с циклом развития хозяина или
переносчика. Так, у трематоды, паразитирующей в мочевом пузыре
лягушки, яйца созревают и выделяются в то т период, когда лягушки
переходят в воду для размножения (продолжительность периода почти
неделя). Из отложенных яиц паразита инвазионные личинки выходят в
тот момент, когда у головастика появляются жабры, на которых
личинки паразитируют. Таким образом, и инвазионные стадии, и
хозяева находятся одновременно в одном месте, синхронность циклов
развития паразита и хозяина вызывается одновременным воздействием
гипофизарного гормона лягушки и на организм паразита.
Яйца БсЬкЮкоша й аетаю Ьш т выделяются из организма челове­
ка с мочой, причем наибольшее их количество поступает в полость мо­
чевого пузыря в полдень, когда вероятность купания особенно
велика (промежуточный хозяин — водный моллюск).
Примером синхронизации циклов паразита и переносчика может
служить филярия \УисЬегепа ЪапкгоШ, личинки которой поступают в
424
периферические сосуды человека в ночные часы, т. е. в то время, когда
переносчик комар рода Мапзоша нападает на человека для сосания
крови. В дневные часы личинки концентрируются в сосудах внутренних
органов.
15.5. П Р И Н Ц И П Ы С И С Т Е М А Т И К И
Объектом медицинской паразитологии, как уже отмечалось, служат
паразиты и переносчики, принадлежащие к различным группам
животного мира. Необходимой предпосылкой их изучения является
знание основ систематики или классификации животного мира.
Систематика — наука, подразделяющая животный и растительный
мир на соподчиненные группы в зависимости от их сходства или
различия, и строяшая систему животного мира. Возникновение
систематики было вызвано необходимостью ориентироваться в много­
образии животных форм. Первоначально ее задача была чисто
практической и состояла в выделении полезных и вредных для человека
организмов (съедобные, несъедобные, ядовитые, неядовитые). С мо­
мента появления теории Ч . Дарвина и до настоящего времени основной
задачей систематики является отражение исторического родства
существующих форм.
Принципы систематики были разработаны шведским естествоиспы­
тателем К . Линнеем (1707— 1778) и изложены в работе «Система
природы» (1735). К . Линней выделил четыре систематические катего­
рии: вид, род, отряд и класс. Вид, по К . Линнею, наименьшая система­
тическая единица, объединяющая организмы, наиболее сходные между
собой. Сходные виды объединяются в род, роды — в отряд, отряды —
в класс, который представлял высшую систематическую единицу. Весь
животный мир, по К . Линнею, включал всего 6 классов: млекопитаю­
щие, птицы, гады, рыбы, насекомые и черви.
Система К . Линнея была искусственной, поскольку не отражала
реального исторического родства между организмами, так как в основу
деления на группы было положено формальное сходство отдельных
признаков.
Многие классификации, создававшиеся в последующие годы, также
были искусственными или в значительной мере формальными.
Современная система животного мира является результатом дли­
тельного развития биологических наук. Она естественна и отражает
реальные филогенетические связи между группами организмов, пос­
кольку в основу классификации положена совокупность признаков
организмов.
Система соподчиненных групп современной классификации включа­
ет вид (хрес1С5), род (§епш), семейство (ГатШ а), отряд (оп1о), класс
(с1а5515) и тип (1урш). Для некоторых групп применяют промежуточные
группы, подтипы, подотряды, подсемейства. Количество видов в
животном мире составляет более 2 млн., типов — свыше 20.
В систематике принята двойная, или бинарная, номенклатура,
впервые введенная К . Линнеем. Сущность бинарной номенклатуры
состоит в том, что каждый вид имеет двойное наименование. Первое
425
слово означает название рода, к которому относится вид, второе —
собственно видовое название. Например, клеш таежный — 1хос1с5
рег5и!саш5, клеш собачий — 1хойе5 папих (слово 1хойе5 означает
название рода, а регаЛсашх и пеших — название видов). Бинарная
номенклатура представляет общепринятую международную систему, в
которой используется только латинская терминология. Благодаря
этому возможно взаимопонимание специалистов любых стран.
Врачу необходимо знать систематическое положение важнейших
паразитов человека и уметь определять их видовую принадлежность,
поскольку каждый вид обладает своими особенностями строения и
экологии. Без этого невозможно поставить точный клинический
диагноз и обосновать соответствующие меры профилактики.
Г л а в а 16
П РО С ТЕЙ Ш И Е
(М Е Д И Ц И Н С К А Я П РО Т О ЗО О Л О ГИ Я )
К типу простейших (РгоЮ/оа) относится ряд патогенных для
человека форм, поражающих отдельные ткани и органы и вызывающих
заболевания различной тяжести, в том числе и с летальным (смер­
тельным) исходом.
Наука о паразитических простейших — возбудителях болезней
человека — носит название протозоология и является важным разделом
в подготовке врачей разных специальностей.
Морфофизиологическая
характеристика.
По строению соответствую т отдельной клетке многоклеточных
(отсюда название «одноклеточные»), по функциям — целому са
мостоятельному организму. Принципиальным отличием простейших
от клетки многоклеточных служит специализация последних, т. е.
каждый вид клеток выполняет в организме какую-нибудь одну функ­
цию и потому зависит от деятельности других клеток и не может су­
ществовать изолированно. В противоположность этому единственная
клетка, которая представляет собой организм простейших, пере­
двигается, захватывает пищу, размножается, защищается от врагов,
т. е. обладает всеми свойствами целого организма и физиологически
ему соответствует. Поэтому в настоящее время простейших назы­
вают организмами на клеточном уровне. В системе животного мира
простейшие представлены всего одним типом.
Тело простейших имеет микроскопические размеры и состоит из тех
же компонентов, что и клетка многоклеточных — наружной мембраны,
цитоплазмы, ядра и органоидов
Наружная мембрана имеет типичное трехслойное строение, но за
счет дополнительных образований у некоторых видов может достигать
большой толщины и плотности. Цитоплазма подразделяется на два
слоя: наружный и внутренний. Наружный слой (эктоплазма) более
плотный, однородный и прозрачный, внутренний (эндоплазма) —
зернистый, имеет более жидкую консистенцию. В эндоплазме нахо­
дятся органоиды общего назначения — митохондрии, эндоплазматиче426
ская сеть, сетчатый аппарат и др. Кроме того, в соответствии с функци­
ями, присущими целому организму, простейшие имеют органоиды
специального назначения, осуществляющие функции передвижения,
питания, выделения, защиты и др.
Органоидами движения простейших служат: 1) псевдоподии или
, прспгтяипяюшие собой временные выросты цитоплазмы;
2)_жгутики— постоянные-органоиды. имеющие вид длинной тонкой
нити, начинающейся обычно на переднем конце; 3) реснички —
Тюстоянные органоиды, представляющие собой многочисленные ко_
~роткйе нитттг~
"I
Строение органоидов питания не одинаково и зависит от способа
питания разных простейших. Большая часть простейших питается
частицами твердой пищи. У т^ких организмов для переваривания пищи
существует пищеварительная вакуоль — капля жидкости, содержащая
пищеварительные ферменты, которая образуется при поступлении
пиши в эндоплазму. Пищеварительная вакуоль окружает пишевую
частицу и перемещается по телу простейшего. Пиша переваривается и
всасывается в цитоплазму. Остатки непереваренной пиши вместе с
пищеварительной вакуолью выбрасываются наружу.
Многие простейшие, ведущие паразитический образ жизни,
усваивают пищу всей поверхностью, тела, используя в основном
механизм пиноцитоэа. Наконец, небольшая группа простейших пи­
тается подобно растениям и имеет хлоропласты.
Органоиды выделения представлены сократительной, шли лудьси=_
рующей вакуолью, имеющей вид небольшого пузырька, наполненного
жидкостью, который периодически'увеличивается, достигая опреде­
ленного объема, а затем сокращается, выбрасывая жидкость наружу.
Накопление и сокращение ритмически чередуются. Основная функция
сократительной вакуоли — поддержание осмотического давления на
постоянном уровне. Концентрация минеральных и органических^
веществ, определяющая осмотическое давление, в теле простейших
выше, чем в окружающей среде, поэтому вода по закону осмоса1 посто­
янно поступает в цитоплазму. Если воду не удалять, то произойдет на­
бухание и гибель простейшего.
У паразитических и морских форм, обитающих в среде с более
высоким осмотическим давлением, сократительная вакуоль может
отсутствовать.
Одновременно с излишками воды через сократительную вакуоль
удаляются жидкие продукты диссимиляции и осуществляется снабже­
ние клетки кислородом, поступающим с водой.
Органоиды защиты см. 16. 4.
Большинство простейших имеет одно ядро, но существуют и
многоядерные формы. Ядро имеет строение, характерное для
эукариот. Морфология и формы деления ядер разнообразны, но, как
л о ж
н о н п ж
у и
1 Явление осмоса состоит в следующем: если два раствора с разной концентра­
цией разделены полупроницаемой мембраной, то растворитель (вода) переходит из
раствора с меньшей концентрацией в раствор с большей концентрацией
427
доказано в настоящее время, деление ядра в любом случае представляет
собой митоз.
Р а з м н о ж е н и е . Простейшие м о г у т размножаться бесполым и
половым путем. Бесполое ра шцожшие. встречается как в виде деления
на две част и, так и в пиле множественного деления^ Половой процесс у
большинства простейших представлен копуляцией, у инфузорий —
конъюгацией.
Жизненный
цикл.
В отличие от соматических клеток
многоклеточных, жизненный цикл простейших складывается из
последовательных стадий развития, которые повторяются с опреде­
ленной закономерностью. Чаще всего исходной стадией является
зигота, затем следует .беенеяое размножение путем деления, затем
образование половых особей и вновь образование зиготы.
И н ц и с т'ТГрЪ в а н и е. При неблагоприятных^ условиях внеш­
ней среды (повышение или понижение температуры, высыхание и т. д.)
вегетативные формы простейших превращаются в цисты. Они
перестают питаться, двигаться и покрываются толстой оболочкой;
процессы обмена веществ при этом резко замедляются. При
восстановлении благоприятных условий из цисты вновь выходит
активная вегетативная форма. Инцистированис — защитная реакция,
возникшая в процессе эволюции, обеспечивающая выживание в небла­
гоприятных условиях. Цисты могут сохраняться в течение длительного
времени, месяцы и даже годы.
Распространение.
Простейшие обитают в пресной и
морской воде, жидких средах организмов.
К л а с с и ф и к а ц и я . В настоящее время классификация типа
простейших пересматривается. Появились новые классификации. Неко­
торые авторы (В. Л. Догель) выделяют простейших как царство,
которое делится на 5 самостоятельных типов: саркомастигофора,
споровики, книдоспоридии, микроспоридии и инфузории. Общеприня­
той является классификация, согласно которой все простейшие делятся
иа 4 класса: 1. Класс саркодовые (5агсос1та). 2. Класс жгутиковые
(Р1а§е11а(а 8. Махб^орЬога). 3. Класс споровики (5рогогоа). 4. Класс ин­
фузории (Ь й ф о п а ). Паразиты человека встречаются в каждом классе.
16.1. К Л А С С С А Р К О Д О В Ы Е ( 5 А К С О О Ш А )
Морфофизиологическая
характеристика.
Наиболее примитивные простейшие, что проявляются прежде всего в
слабой степени дифференцировки. Цитоплазма ограничена только
наружной мембраной, форма тела непостоянная. Органоидами движе­
ния и захвата пиши_ служат псевдоподии^ Специальное ротовое
отверстие отсутствует. Поступление пиши и выделение непереваренных
остатков может происходить в любом участке тела. Сократительная
вакуоль одна. Выделение продуктов диссимиляции и излишков воды
также может происходить в любом месте. Обычно имеется только одно
ядро, хотя встречаются и многоядерные формы.
Р а з м н о ж е н и е . Размножение в основном бесполым путем —
делением на две части.
428
\л.
И н ц и с т и р о в а п и е . В неблагоприятных условиях образую!
цисту.
Медицинское значение имеет только один отряд подкласса
Корненожек, а именно отряд Лмеб (АтосЫ па).
16.1.1. Отряд Амебы (АгпоеЫпа)
В организме человека обитает несколько видов амеб. Наиболее
важное значение имеет дизентерийная амеба.
1^/' Дизентерийная амеба (Е т а т о е Ь а Ы$(о1уйса). Возбудитель тяжелого
заболевания — амебной дизентерии или амебиаза.
Л о к а л и з а ц и я . Толстый кишечник.
Г е о г р а ф и ч е с к о е р а с п р о с т р а н е н и е . Повсеместно,
но в районах с жарким климатом встречается чаше.
Морфофизиологическая
характеристика и
Рис. 172. Дизентерийная и кишечная амебы
Д изен тер и й н ая ам еб а: а
к р у п н а я в е г е т а т и в н а я ф о р м а , б, в
м е л к а я в е г е т а т и в н а я ф орма- I
эктоплаз
ма; 2
эн д оп лазм а: 3
псевд о под ии ; 4
ядро; 5
кариосом а; 6
ф аго ц и ти р о в ан н ы е эри тр оц и ты
в п и щ е вар и тельн ы х ва к у о л ях , г — ц и ста; к и ш ечн а я ам еб а: д
вегетати вн ая ф орма, е
- ц и ста
429
основное значение; общественная — борьба с загрязнением среды
фекалиями свиней, а также людей, соответствующая организация
условий труда на свиноводческих фермах, своевременное выявление и
лечение больных.
Глава
17
ГЕЛ ЬМ И Н ТЫ
(М Е Д И Ц И Н С К А Я Г Е Л Ь М И Н Т О Л О Г И Я )
\
17.1. Т И П
ПЛО СКИЕ ЧЕРВИ
(Р 1 .А Т Н Г Ц И 1 М Т Н Е 5 )
М о р ф о ф и з и о л о г и ч е с к а я х а р а к т е р и с т и к а . Те­
ло сплющено в дорсовентральном направлении (наиболее характерД ный внешний признак). Ф о рма его, как правило, лентовидная или
листовидная. Стенка представлена кожно-мускульным мешком, состо­
ящим из слоя кожного эпителия и трех слоев гладкой мускулатуры — •
кольцевого, лродолъного~и диагонального.
У паразитических червей поверхностный покровный слой приобре­
тает особое строение. Как показали электронно-микроскопические
исследования, он преобразован в утративший клеточное строение
__ тегумент, в наружной части которого находится цитоплазма с большим
количеством вакуолей, митохондрий и гранул. С помощью ци­
топлазматических тонких тяжей этот слой соединяется с глубже
расположенным внутренним слоем, который находится под про­
дольными и кольцевыми мышцами. Погруженная часть тегумента
представлена грушевидными участками цитоплазмы, содержащими
ядра. Внутри кожно-мускульного мешка располагаются внутренние
органы, пространство между которыми заполнено особой рыхлой
соединительной тканью — паренхимой, в которой накапливаются
запасные питательные вещества. "Полость тела отсутствует.
У плоских червей -'развиты "пйщеварительная, выделительная,
нервная и половая системы. Дыхательная и кровеносная- системь
отсутствую т. Пищета^ительная^с^стема^представлена ртом, глоткой,
пищеводом и дбумя кишечными каналами — прямыми или разветвлен­
ными. Рот, глотка и пищевод, выстланный кутикулой, составляют
передний, эктодермальный отдел пищеварительной трубки, кишеч­
ник — средний энтодермальный отдел. В нем происходит переварива
ние и всасывание пищи. Заканчивается кишечник слепо. Анального
отверстия нет. Непереваренные остатки пиши выбрасываются через
рот. Vнекоторы х плоских червей (ленточные) пищеварительная
система отсутствует.
Выделительная^ си^з^ма протонефридиального типа, состоящая из
спещшШ йрованных терминальных, или звездчатых, клеток, разбро­
санных по всему телуТ^Гсйстемы отходящих от них ветвистых каналь­
цев. В середине каждой клетки находится полость с пучком ресничек.
Электронно-микроскопические исследования показали, что стенки
начального участка канальцев звездчатых клеток состоят из продольно
расположенных плотных цитоплазматических тяжей, разделенных
448
3
Рис. 183. Выделительная система трематод.
л
р асп ол о ж ен ие к а н а л о в 1
со би р ате л ьн ы е к ан ал ьц ы протонеф ридиев; 2
центр альн ы й вы д е л и ­
те л ьн ы й к а н а л ; 3
вы д е ли тел ьн ая пора, б
эл ектр о нн о -м и кр о скоп ическая схем а стр оени я протонефридиальной к ле тки : 1
зв е з д ч а т а я ч а с т ь м ер ц ательно й к ле тки ; 2 — ц и то п л азм ати че ск и е т я ж и ; 3
тр у б ка
( вн утр и м е р ц ате л ьн о е п л а м я ).
тончайшими щелевидными просветами, затянутыми слизистой плен­
кой. Через эти щелевидные отверстия поступают жидкие продукты
диссимиляции и избыток жидкости, подлежащие удалению. Канальцы
соединяются между собой, образуют два латеральных канала, которые
сливаются и открываются непарной выделительной порой на заднем
конце тела. Движение ресничек в клетках и канальцах создает ток
жидкости по направлению к выделительному отверстию (рис. 183).
Протонефридиальная система удаляет продукты диссимиляции и регу­
лирует осмотическое давление.
Нервная система представлена двумя нервными узлами, которые
находятся в переднем конце тела и соединены комиссурами. О т них
отходят вдоль тела нервные стволы, из которых наиболее развиты
боковые. Нервные стволы соединяются между собой комиссурами,
образуя систему, именуемую ортогоном. К органам чувств относятся
нервные чувствительные окончания (сенсиллы).
Подавляющее большинство плоских червей
каждая особь имеет мужскую и женскую половую
сложно.
К л а с с и ф и к а ц и я . Тип плоские черви
1. Класс Ресничные черви (ТигЪеИапа). 2. Класс
шёа). 3. Класс Ленточные черви (Се$Нн1а).
относятся к двум последним классам.
■
17.1.1. Класс Сосальщики (Тгсша1о<1а)
гермафродиты, т. е.
систему, устроенную
включает 3 класса:
Сосальщики (ТгешаПаразиты человека
' ■
' К -
Класс включает только паразитические формы.
М о р ф о ф и з и о л о г и ч е с к а я х а р а к т е р и с т и к а . Те­
ло в большинстве случаев имеет листовидную форму и размеры от 2 до
80 мм. Кожно-мускульный мешок представлен тегументом и тремя
мышечными слоями — кольцевым, диагональным, продольным^
Им ею т специальные органы прикрепления (фиксации) к телу
хозяина или присоски. Каждая присоска представляет собой цирку­
лярную мышцу с полостью внутри. Обычно имеется 2 присоски —
ротовая и брюшная. Ротовая расположена на переднем конце тела,
терминально, и связана с ротовым отверстием, брюшная — на
брюшной стороне и служит только для фиксации.
Пищеварительная система имеет типичное_строение. Пищей для
сосальщиков служат содержимое кишечника, клеточные элементы,
слизь, жидкая часть крови и т. д.
Выделительная система протонефридиального типа. Диссимиляция
осуществляется анаэробным путем, энергия освобождается за счет
гликогена клеток паренхимы.
Нервная система представлена<бртогоном,^> органы чувств — сен­
сорным аппаратом (сенсиллы, чувсУвительные щетинки и др.).
Половая система достигает исключительного развития и очень
сложно устроена. Все трематоды, за исключением шистозом,
гермафродиты.
'
М ужская половая система состоит обычно из двух компактных
семенников, от каждого из которых отходят семяпроводы. Н а пе­
реднем конце тела они соединяются и образуют непарное соединение —
семенной пузырь, переходящий в семяизвергательный канал. Дистальный.отдел этого канала проходит внутри совокупительного органа —
(шрруап Мужские половые клетки образуются в семенниках, затем по
се&гяТфоводам поступают в циррус.
Женская половая система начинается яичнйком (непарный), в
котором образуются женские половые клетки. Он имеет форму
разветвленной трубки или округлого тельца. О т яичника отходит
яйцевод, который открывается в небольшую полость — оотип.
По яйцеводу женские половые клетки проходят из яичника в камеру
оотипа, где происходит оплодотворение (рис. 184, 185).
К женской половой системе относятся также желточники,
состоящие из большого числа округлых пузырьков, расположенных в
боковых частях тела. О т желточников отходят каналы, впадающие в
оотип. В желточниках образуются клетки, богатые желтком, который
используется развивающимся
зародышем. Оотип или цент­
ральная камера — небольшая
полость, куда открываются
протоки всех органов женской
половой системы и где проис­
ходит процесс оплодотворе­
ния и формирования яиц.
Маткапредставлена узкой
трубкой, "образующей много­
численные петли. Один ее ко­
нец открывается в оотип, дру­
гой заканчивается на переднем
конце тела женским половым
отверстием рядом с отверсти­
ем цирруса. В матке происхо­
дит развитиев яйце зародыша.
Кроме того, у сосальщиков
матка одновременно выполня­
ет и функцию влагалища —
через нее в оотип поступают
мужские половые клетки.
Тельце Мелиса — мелкие
Рис. 184. Схема строении половой системы
одноклеточные железы, рас­ трематод.
положенные около оотипа.
1
сем ен н ик; 2
сем япр о вод , 3
цирруснаи с у м к а с
р р усом ; 4
яи чн и к , 5
сем нп р и»м н и к; 6
ж елтом
Жидкий секрет, выделяемый аи
н ики, 7
те л ьц е М е л и с а , 8
Л аур ер о в канал; 9
ими, заполняет оотип и, воз­ «« т и п 1, 10 м а тк а (в л а г а л и щ е )
можно, участвует в образова­
нии оболочки (скорлупы) яйца. Отходящий от оотипа на спинную
сторону короткий канал называется лауреровым каналом. Предпо­
лагают, что он служит для удаления избытка половых продуктов.
Все перечисленные органы связаны между собой в единую систему.
О п л о д о т в о р е н и е-И ф о р м и р о в а н и е я й ц а . Опло­
дотворение, как правило, перекрестное, т. е. при размножении черви
соединяются попарно и обмениваются мужскими половыми клетками.
Сперматозоиды из цирруса поступают в матку партнера и проходят в
оотип, где соединяются с яйцеклетками. Оплодотворенное яйцо
окружается желточными клетками, которые прилипают к его поверхно­
сти, после чего снаружи образуется оболочка. Сформированное яйцо
поступает из оотипа в матку и продвигается к наружному половому от­
верстию, в то время как в яйце развивается зародыш. Созревшее яйцо
выходит через отверст
ки наружу.
Яйцо имеет, характ^, ’ь^е особенности: форма овальная, на одном
полюсе находится крышечка, через которую выходит личинка.
Ж и з н е н н ы й ц и к л . Сложный, со сменой хозяев и нескольки­
ми поколениями личиночных стадий.
Окончательным хозяином служат позвоночные и человек,
промежуточным, обязательным — моллюски. Некоторые трематоды,
кроме того, имеют второго промежуточного хозяина, которым могут
*
16*
*
451
Рис. 185. Трематоды
- паразиты человека.
2
к о ш а чи й с о с а л ь щ и к : I
р о то в ая п р исо ска; 2
гл о тк а ; 3
б р ю ш н а я п р и со ск а; 4
ветви ки ш еч н и к а; 5
сл епы е о к о н ч ан и я
ветвей к и ш е ч н и к а ; 6
ж ен ск о е половое о тве р сти е ; 7
м уж ­
ское половое о тве р сти е ; 8
м а тк а (в л а г а л и щ е ), 9
ж елточники, 10
яи чн и к I I
се м япр и ем н ик; 12
сем енники, 13
к ан ал вы д е ли тел ьн о й си стем ы , 14
вы д е л и т е л ь н а я пора; б
печен о чн ы й со с а л ь ш и к ; в
л ан ц етовид н ы й с о са л ьщ и к ; г
ле
точный со са л ьш и к .
Рис. 18Н. Строение личиночных стадий трематод.
<1
мнрдцмдии; б
спороцистег, в
редия, г - ц еркарий; д
метацср
ресн ички ; 2
гл аза ; 3 — зар о д ы ш е вы е к ле тки , 4
зароды ш и
церкарии, 8
присоска; 7
хвост; 8 • о б о ю ч к а цисты
к«1р и й , 1
редий, 5
быть низшие позвоночные и представители различных групп беспозво­
ночных.
Характерной особенностью жизненного цикла служит размножение
личиночных стадий путем партеногенеза.
Половозрелая форма — марита - г откладывает яйца, которые
выносятся наружу. Для дальнейшего развития яйцо, как правило,
должно попасть в воду. Из яйца выходит первая личиночная стадия —
мипяпилий. имеющий овальную форму, ресничный покров, 2
пиГй' нтных глазка на переднем конце тела и протонефридии. В задней
453
части тела мираиидия находятся так называемые зародышевые клетки,
которые дают начало следующему поколению личиночных форм.
Мирацидцй^плавает в воде и активно проникает в тело моллюска —
промежуточного хозяина. Здесь мирацидий превращается в мешковиднукщ ю пгаш сту. внутри которой сохраняются зародышевые клетки.
Через некоторое время из каждой зародышевой клетки внутри
спороцисты развиваются без оплодотвоврения, т. е. путем партеноге­
неза, следующая лечиночная стадия — редия. Она имеет удлиненное
тело, глотку, зачатки кишечника, нервй'бй Тг^Тзыделительной систем и
также содержит зародышевые клетки. Количество редий, развиваю­
щихся в спороцисте, зависит от вида трематод (от 8 до 100). Редии вы ­
ходят из спороцисты в окружающие ткани моллюска. В теле редии из
зародышевых клеток также партеногенетически образуется следующее
1—' поколение личинок—■церкарии (рис. 186). Последние имеют тело с
хвостовым придатком, / присоски, кишечник, сформированную выде­
лительную систему, а иногда и Зачаток половой системы. Н а переднем
конце тела у некоторых форм находится острый стилет или пучок
шипов, выполняющих перфорирующую функцию, и группа желез
проникновения. Церкарии выходят из редии и затем из тела модлюсков
и свободно плавают в воде.
^
{/0 У Н №
У большинства трематод в дальнейшем церкарий проникает во
второго промежуточного хозяина (позвоночные, беспозвоночные) и там
превращаются в инцистированную форму — мехацеркария. У тех
трематод, которые имеют одного промежуточного хозяина, церкарий
инцистируется непосредственно во внешней среде. Э та стадия называетс&ддолееКврртУ'Метацеркарий и адолескарий являю тс^Зш вазионнй-^
4
о ш ш ч а т 5^ п ь н о г о хозяина^ в организме которого они преврашаются в м а р и т у . ---------В связи со сложностью цикла развития значительная часть
личиночных стадий трематод погибает, не достигая половозрелой
формы. Действительно, развитие будет прервано, если яйцо не попадет
в воду; если мирацидий не встретит моллюска соответствующего вида
или моллюск будет уничтожен другими животными; если церкарий не
встретит окончательного хозяина и т. д. В результате лишь ничтожный
процент личинок достигает стадии мариты. Вышеуказанная огромная
гибель компенсируется двумя путями: 1) мошным развитием половой
системы у мариты и продукцией огромного числа половых продуктов
(тысячи и десятки ты сяч); 2) размножением в стадии личиночных форм;
каждая спороциста дает от 8 до 100 редий, каждая редия, в свою оче­
редь, дает от 20 и более цсркарисв. В результате потомство одного
мирацидия может составлять от 600 до 200 000 церкариев.
Трематоды распространены по всему земному шару. Класс
включает большое количество форм, патогенных для человека. Заболе­
вания, вызываемые сосальщиками, называются трематодозами
Кошачий, или сибирский, сосальщик (ОрЫЬогсЫк ГеПпеи*). Вызывает
заболевание описторхоз, впервые описанное К . Н. Виноградовым в
1891 г. в Сибири.
Л о к а л и з а ц и я . Половозрелые особи паразитируют в желчных
протоках печени, желчном пузыре, поджелудочной железе.
Г е о г р а ф и ч е с к о е р а с п р о с т р а н е н и е . На террито­
рии С С С Р встречается в районе Оби и Иртыш а, реже в бассейне Кам ы ,
Днепра.
Морфофизиологическая
характеристика.
Передний конец тела уже заднего. Каналы кишечника не разветвленные,
заканчиваются не доходя до заднего конца тела. М атка занимает
среднюю часть тела. Кзади от нее расположен округлый яичник и
бобовидный семяприемник. В задней четверти тела находятся два
лопастных семенника, между которыми виден 5-образно изогнутый
канал выделительной системы. Желточники находятся между каналами
кишечника и краем тела. Яйца бледно-желтой окраски, с крышечкой на
одном полюсе.
Жизненный
ц и к л . {-Окончательные х о зя е в а ч е л о в е к ,
кошка, собака^ лисица, песец и другие плотоядные животные.
ПТц(Тч1ёжУточные' хозяеваГ^первый — дресноводный моллюск ВнИуша
(ёасЫ, второй — рыбьГсемейства карповых (язь, плотва, сазан, вобла и
др.). Через желчные протоки окончательного хозяина яйца попадают в
кишечник и с фекалиями выносятся наружу. Для развития яйцо должно
попасть в воду и бы ть заглоченным моллюском (рис. 187), в организме
которого из яйпа выходит мирацидий, затем последовательно
образуются 9 пог)оиисты, редии, церкарии. П оследние выходят из
моллюска и активно или пассивно проникают в тело рыбы, где в
подкожной клетчатке или мышцах превращаются в ^ метацеркарии.
При поедании окончательным хозяином зараженной рыбы метацеркарии попадают в его пищеварительный тракт, затем паразит проникает в
печень и желчный пузырь, где превращается в половозрелую форму
(мариту).
Человек заражается при употреблении в пишу плохо прожаренной и
проваренной рыбы. Особенно широко распространен описторхоз в тех
районах, где существует обычай употреблять в пишу сырую свежеморо­
женую рыбу. Зараженность в очагах описторхоза может дости­
гать 90%.
Описторхоз является природно-очаговым заболеванием. Его
резервуаром в природе служат дикие животные, участие человека для
распространения болезни необязательно.
П а т о г е н н о е д е й с т в и е . Паразиты оказываю т токсиче­
ское действие на организм больного. Скопление паразитов от 20 тыс.
экземпляров (известно обнаружение до 75 тыс. паразитов) вызывает
задержку тока желчи и сока поджелудочной-железы, - приводит к
развитию цирроза в пораженных органах. При большом количестве
гельминтов возможен смертельный исход.
Лабораторная
д и а г н о с т и к а . Диагноз ставится на
основании обнаружения яиц паразита в фекалиях больного или в
содержимом двенадцатиперстной кишки. Яйца начинают выделяться
через месяц после заражения.
П р о ф и л а к т и к а : личная — не упот