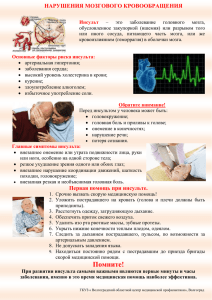
ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ МИНИСТЕРСТВА ЗДРАВООХРАНЕНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
advertisement

1 ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ «КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ МЕДИЦИНСКИЙ УНИВЕРСИТЕТ» МИНИСТЕРСТВА ЗДРАВООХРАНЕНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ На правах рукописи ТРОФИМЕНКО Артем Иванович ПАТОГЕНЕТИЧЕСКОЕ ОБОСНОВАНИЕ ПРИМЕНЕНИЯ ТЭС-ТЕРАПИИ ПРИ ИШЕМИЧЕСКОМ ИНСУЛЬТЕ (экспериментальное исследование) 14.03.03 — патологическая физиология Диссертация на соискание ученой степени кандидата медицинских наук Научный руководитель: доктор медицинских наук, профессор Каде Азамат Халидович Краснодар — 2014 2 ОГЛАВЛЕНИЕ СПИСОК СОКРАЩЕНИЙ ............................................................................. 7 ВВЕДЕНИЕ ....................................................................................................... 9 ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ ................................................................ 17 1.1. Этиология ишемического инсульта ......................................................... 17 1.2. Классификация ишемического инсульта ................................................. 17 1.3. Общие аспекты патогенеза ишемического инсульта .............................. 19 1.4. Роль воспаления в патогенезе ишемического инсульта ......................... 22 1.5. Роль системного воспаления в патогенезе ишемического инсульта ...... 29 1.6. Понятие об опиоидергической нейротрансмиттерной системе ............. 31 1.7. Основные принципы профилактики и лечения ишемического инсульта ............................................................................ 35 1.8. Понятие о транскраниальной электростимуляции ................................. 37 1.8.1. Центральные эффекты ТЭС-терапии .............................................. 38 1.8.2. Периферические эффекты ТЭС-терапии ......................................... 40 ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ ..................... 45 2.1. Характеристика групп животных ............................................................ 45 2.2. Методы исследования .............................................................................. 46 2.2.1. Методика наркоза ............................................................................. 46 2.2.2. Методика моделирования ишемического инсульта у крыс ............ 46 2.2.3. Методика проведения ТЭС-терапии у крыс .................................... 48 2.2.4. Методика эвтаназии у крыс .............................................................. 49 2.2.5. Методика забора крови ..................................................................... 49 2.2.6. Методика иммуноферментного анализа β-эндорфина в плазме крови крыс ................................................... 50 3 2.2.7. Методика иммуноферментного анализа интерлейкина-1β в плазме крови крыс ............................................ 51 2.2.8. Методика иммуноферментного анализа интерлейкина-6 в плазме крови крыс .............................................. 52 2.2.9. Методика иммуноферментного анализа фактора некроза опухоли-α в плазме крови крыс ......................................... 53 2.2.10. Методика иммуноферментного анализа интерлейкина-10 в плазме крови крыс ............................................ 53 2.2.11. Методика иммуноферментного анализа трансформирующего фактора роста-1β в плазме крови крыс ....... 54 2.2.12. Методики гистологических исследований ...................................... 55 2.2.13. Методика определения размеров очага ишемии в головном мозге у крыс ................................................................... 59 2.3. Статистические методы обработки полученных данных ....................... 60 ГЛАВА 3. ГИСТОЛОГИЧЕСКАЯ КАРТИНА ЗОНЫ ИШЕМИИ ПРИ МОДЕЛИРОВАНИИ ИШЕМИЧЕСКОГО ИНСУЛЬТА БЕЗ ПРОВЕДЕНИЯ И С ПРИМЕНЕНИЕМ ТЭС-ТЕРАПИИ ........................................................................... 61 3.1. Гистологическая картина зоны ишемии при моделировании ишемического инсульта ............................................................................ 61 3.2. Гистологическая картина зоны ишемии при моделировании ишемического инсульта при применении ТЭС-терапии ........................ 70 3.3. Изменение объема очага инфаркта мозга при моделировании ишемического инсульта без проведения и с применением ТЭС-терапии ................................................................... 77 4 ГЛАВА 4. ДИНАМИКА СОДЕРЖАНИЯ ЦИТОКИНОВ И β-ЭНДОРФИНА В ПЛАЗМЕ КРОВИ КРЫС ПРИ МОДЕЛИРОВАНИИ ИШЕМИЧЕСКОГО ИНСУЛЬТА .................................................................................. 80 4.1. Динамика содержания цитокинов в плазме крови крыс при моделировании ишемического инсульта ................................................. 80 4.1.1. Динамика содержания фактора некроза опухоли-α у крыс с экспериментальным ишемическим инсультом ............................. 80 4.1.2. Динамика содержания интерлейкина-1β у крыс с экспериментальным ишемическим инсультом ............................. 83 4.1.3. Динамика содержания интерлейкина-6 у крыс с экспериментальным ишемическим инсультом ............................. 85 4.1.4. Динамика содержания интерлейкина-10 у крыс с экспериментальным ишемическим инсультом ............................. 87 4.1.5. Динамика содержания трансформирующего фактора роста-1β у крыс с экспериментальным ишемическим инсультом ................ 89 4.2. Динамика содержания β-эндорфина у крыс с экспериментальным ишемическим инсультом .......................................................................... 91 4.3. Динамика содержания цитокинов в плазме крови крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии ................................................................. 94 4.3.1. Динамика содержания фактора некроза опухоли-α у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии ........................ 94 4.3.2. Динамика содержания интерлейкина-1β у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии ........................ 96 4.3.3. Динамика содержания интерлейкина-6 у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии ........................ 98 5 4.3.4. Динамика содержания интерлейкина-10 у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии ...................... 100 4.3.5. Динамика содержания трансформирующего фактора роста-1β у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии ...................... 102 4.4. Динамика содержания β-эндорфина у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии ........................................................................................... 104 4.5. Динамика содержания цитокинов в плазме крови крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии ............................................................... 106 4.5.1. Динамика содержания фактора некроза опухоли-α у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии ....................................................... 106 4.5.2. Динамика содержания интерлейкина-1β у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии ....................................................... 108 4.5.3. Динамика содержания интерлейкина-6 у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии ....................................................... 110 4.5.4. Динамика содержания интерлейкина-10 у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии ....................................................... 112 4.5.5. Динамика содержания трансформирующего фактора роста-1β у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии ............................ 114 4.6. Динамика содержания β-эндорфина у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии ........................................................................................... 117 6 ГЛАВА 5. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ .............................................. 120 ВЫВОДЫ ....................................................................................................... 136 ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ ..................................................... 138 СПИСОК ЛИТЕРАТУРЫ ........................................................................... 140 ПРИЛОЖЕНИЯ ........................................................................................... 169 7 СПИСОК СОКРАЩЕНИЙ АНС — антиноцицептивная система АЭ — анальгетический эффект АФК — активные формы кислорода БАВ — биологически активные вещества БОФ — белки острой фазы ГГНС — гипоталамо-гипофизарно-надпочечниковая система ГМ — головной мозг ГЭБ — гемато-энцефалический барьер ИИ — ишемический инсульт ИЛ-1 — интерлейкин-1 ИЛ-1α — интерлейкин-1α ИЛ-1β — интерлейкин-1β ИЛ-2 — интерлейкин-2 ИЛ-4 — интерлейкин-4 ИЛ-6 — интерлейкин-6 ИЛ-8 — интерлейкин-8 ИЛ-10 — интерлейкин-10 ИФА — иммуноферментный анализ Лтр — лейкотриены Лф — лимфоциты МФ — макрофаги МЦ — моноциты Нф — нейтрофил НК — натуральный киллер ОНМК — острое нарушение мозгового кровообращения ОП — опиоидные пептиды ОР — опиатные рецепторы 8 ПОЛ — перекисное окисление липидов ПГ — простагландины РАИЛ-1 — рецепторный антагонист интерлейкина-1 СКАО — синдром компенсаторного антивоспалительного ответа ССВО — синдром системного воспалительного ответа ТАП — тканевой активатор плазминогена ТФР-1β — трансформирующий фактор роста-1β ТЭС — транскраниальная электростимуляция ФНО-α — фактор некроза опухоли-α ЦНС — центральная нервная система ЭЦ — эндотелиоцит Bcl-2 — белковый фактор регулятор апоптоза CD11 — кластер дифференцировки-11 CD11α — кластер дифференцировки-11α CD18 — кластер дифференцировки-18 CD11/18 — кластер дифференцировки-11/18 ICAM-1 — молекула клеточной адгезии NO — оксид азота rtPA — тканевой активатор плазминогена Th1 — Т-хелпер 1 9 ВВЕДЕНИЕ (общая характеристика работы) Актуальность исследования. Среди современных общемировых медикосоциальных проблем ишемический инсульт (ИИ) занимает одно из ведущих мест ввиду высоких, имеющих тенденцию к росту показателей заболеваемости и смертности [A.D. Lopez, C.D. Mathers, 2006; A. Di Carlo, 2009; D. Lloyd-Jones et al., 2009; P.U. Heuschmann et al., 2009; M.A. Moskowitz, E.H. Lo, C. Iadecola, 2010]. Рост числа инсультов головного мозга (ГМ) среди трудоспособного населения нашей страны является на данный момент одним из самых важных вопросов отечественного здравоохранения [А.Н. Редько, Р.К. Карипиди, В.В. Завьялова, 2013; В.И. Скворцова, Г.С. Алексеева, Н.Ю. Трифонова, 2013]. В Российской Федерации заболеваемость инсультом среди лиц старше 25 лет составила 3,48 ± 0,21, смертность от инсульта 1,17 ± 0,06 на 1000 населения в год. Для сравнения в США частота инсульта среди жителей европеоидной расы составляет 1,38–1,67 на 1000 населения. Доля острых нарушений мозгового кровообращения (ОНМК) в структуре общей смертности в России составляет 21,4 %. Смертность от инсульта среди лиц трудоспособного возраста увеличилась за последние 10 лет более чем на 30 % (41 на 100000 населения). Ранняя 30дневная летальность после инсульта составляет 34,6 %, а в течение года умирают приблизительно 50 % больных, то есть каждый второй заболевший. Инсульт — преобладающая причина инвалидизации населения (3,2 на 1000 населения). По данным Национального регистра инсульта, 31 % пациентов, перенесших инсульт, нуждаются в посторонней помощи для ухода за собой, 20 % не могут самостоятельно ходить. Лишь 8 % выживших больных могут вернуться к прежней работе [Е.И. Гусев и соавт., 2009]. ИИ — острое нарушение кровообращения в ГМ с развитием стойких симптомов поражения, вызванных инфарктом в мозговое вещество [Е.И. Гусев, А.Н. Коновалов, А.Б. Гехт, 2007]. 10 Независимо от причины и патогенетического варианта развития ИИ, его развитие характеризуется формированием области острой локальной ишемии в ткани мозга [Е.И. Гусев, В.И. Скворцова, 2001; А.А. Фирсов и соавт., 2013; N.W. Manning et al., 2014]. Церебральная ишемия является процессом, обусловливающим метаболический этап патогенеза ИИ, в основе которого лежат универсальные реакции ткани мозга на нарушения деятельности сосудистых систем, обеспечивающих жизнедеятельность мозга [Л.П. Соколова; 2011; К.П. Иванов, Ю.И. Зильбертер, Т.М. Зильбертер, 2012; K.A. Hossmann, 2012; Е.И. Гусев, М.Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014]. Очаг ишемии мозга неоднороден, в нем выделяют: зону необратимого повреждения которую называют ядром инфаркта и зону ишемического поражения обратимого характера которую обозначают термином «пенумбра» (ишемическая полутень) [Е.И. Гусев, В.И. Скворцова, 2001; A. Durukan, T. Tatlisumak, 2007; Е.И. Гусев, М.Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014]. За счет сокращения зоны пенумбры происходит последующее увеличение размеров зоны инфаркта мозга [Е.И. Гусев, В.И. Скворцова, 2001; Е.И. Гусев, М.Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014]. В настоящее время считается, что единый организованный ответ организма на острую церебральную ишемию осуществляется путем взаимодействия нервной, иммунной и эндокринной систем, использующих для этого нейротрансмиттеры, цитокины, факторы роста, гормоны и воздействующих на сходные мишени (рецепторы) [Е.И. Гусев, В.И. Скворцова, 2001; А.Х. Каде, А.П. Парахонский, 2004; А.А. Мольберг, Т.В. Гришина, 2006; А.Ф. Повещенко, В.В. Абрамов, В.А. Козлов, 2007; А.А. Олейник, Р.С. Вастьянов, 2008; М.А. Самоструева, Д.Л. Теплый, И.Н. Тюренков, 2009; Г.Н. Крыжановский, С.В. Магаева, 2010; Ж.В. Титова, Г.М. Бодиенкова, 2013]. Патологическим процессом, лежащим в основе расширения зоны инфаркта мозга в течение первой недели после возникновения инсульта, а также ее организации в более поздние сроки является острое асептическое воспаление [K.J. Becker, 2001; G.J. del Zoppo, K.J. Becker, J.M. Hallenbeck, 2001; M.D. Gins- 11 berg, 2003; M. Fisher, R. Ratan, 2003; P.L. Wood, 2003; R. Markus, D.C. Reutens, S. Kazui, 2004; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; Q. Wang, X.N. Tang, M.A. Yenari, 2007; А.Х. Хана-Мурад, Л.И. Павлинова, А.А. Мокрушин, 2007; A. Tuttolomondo, D. Di Raimondo, R. Di Sciacca, 2008; J. Jordan et al., 2008; D. Amantea et al., 2008; C. Benakis, L. Hirt, R.A. Du Pasquier, 2009; Ю.Д. Губарев, А.О. Шеремет, 2009; А.С. Кость, Б.Д. Луцик, Л.Е. Лаповець, 2010; A.G. Ceulemans et al., 2010; Y. Lee et al., 2014; М.В. Онуфриев и соавт., 2014]. Значительное влияние на жизнеспособность клеток в зоне пенумбры оказывает изменение концентрации β-эндорфина в крови, происходящее при нарушении мозгового кровообращения [М.А. Барабанова, 2003; Н.Н. Волосникова, 2004; A.I. Faden, B. Stoica, 2007; V.C. Borlongan et al., 2009]. В патогенетической терапии ИИ выделяют два основных направления: улучшение перфузии ткани мозга и нейропротективную терапию [Е.И. Гусев, В.И. Скворцова, 2001; М. Бэр, 2011]. Ключевой концепцией терапии ИИ является воздействие на зону пенумбры для предотвращения гибели клеток [Е.И. Гусев, В.И. Скворцова, 2001; W.S. Smith, 2004; A. Bose et al., 2008; М. Бэр, 2011]. Это в перспективе приведет к максимальному сохранению неповрежденной ткани мозга, сокращению объема инфаркта мозга и, следовательно, менее выраженному неврологическому дефициту в последующем [Е.И. Гусев, В.И. Скворцова, 2001; М. Бэр, 2011; M. Fisher, R. Ratan, 2003; R. Markus, D.C. Reutens, S. Kazui, 2004; Г.В. Цаканова и соавт., 2011]. Основная задача лечения состоит в разработке таких воздействий и лекарственных препаратов, которые этому способствуют. В наше время почти все используемые нейропротекторные препараты недостаточно эффективны [J. Rother, 2008]. Многие препараты, показавшие высокую эффективность в эксперименте, зачастую обладают выраженными побочными эффектами, которые делают их применение в клинической практике невозможным [Е.И. Гусев, В.И. Скворцова, 2001; М. Бэр, 2011; J. Rother, 2008]. Поэтому важнейшей задачей является разработка новых подходов к профилактике и терапии ИИ. 12 Одним из перспективных в этом плане методов является транскраниальная электростимуляция (ТЭС-терапия) [В.П. Лебедев, А.Б. Савченко, Н.В. Петряевская, 1988; С.В. Рычкова, 2006]. Метод ТЭС-терапии направлен на избирательную активацию защитных (антиноцицептивных) механизмов мозга, ключевую роль, в деятельности которых играет эндогенная опиоидергическая система [Я.С. Кацнельсон, 1985; В.П. Лебедев и соавт., 1986; Ю.Д. Зильбер, М.В. Степанова, Ю.В. Киселев, 1987; А.Б. Савченко, 1994]. ТЭС-терапия модулирует активность стресс-лимитирующих систем организма [Л.Н. Айрапетов и соавт., 1987]. Ее воздействие носит комплексный, системный, гомеостатический характер, а все эффекты обусловлены центральным или периферическим влиянием опиоидергической системы [Л.Н. Айрапетов и соавт., 1987; А.Б. Савченко, 1994]. Показан иммуномодулирующий эффект ТЭС-терапии [Ю.А. Богданова, А.Х. Каде, Р.А. Ханферян, 2000; Ю.А. Богданова, А.Х. Каде, Р.А. Ханферян, 2002; Ю.А. Богданова, 2003; А.В. Рубцовенко, 2006]. Большой интерес представляет способность ТЭС-терапии повышать толерантность органов к ишемии [В.Г. Борисенко, 2009]. В настоящее время показан ряд эффектов эндогенных опиоидных пептидов (ОП): антигипоксический, влияние на синтез гипофизарных гормонов, иммуномодулирующий и др. [D.J.J. Carr, 1991; M.L. Simmons, C. Chavkin, 1996; Л.Н. Маслов, Р.С. Карпов, Ю.Б. Лишманов, 2002; О.Б. Сеин и соавт., 2009; С.В. Гейн, 2010; В.А. Дубынин, А.А. Каменский, 2010; T.V. Lasukova, S.V. Nizkodubova, A.S. Tayurskaya, 2011; S. Sharma, R. Singh, S. Rana, 2011; А.Г. Полунина, Е.А. Брюн, 2013; Т.В. Ласукова, С.В. Низкодубова, Е.Ю. Мухтобарова, 2013]. Таким образом, может быть перспективно применение ТЭС-терапии при лечении ИИ [Н.Л. Грицкевич и соавт., 1993; М.Г. Пшенникова, 2003]. Однако для подтверждения данной гипотезы требуется углубленное исследование влияния ТЭС-терапии на баланс про- и противовоспалительных цитокинов совместно с динамикой морфологических изменений в очаге острой локальной церебральной ишемии. 13 Исходя из этого целью работы являлось изучение влияния ТЭС-терапии на динамику баланса про- и противовоспалительных цитокинов при экспериментальном ишемическом инсульте. Задачи исследования 1. Исследовать уровень про- (интерлейкина-1β, интерлейкина-6, фактора некроза опухоли-α) и противовоспалительных (интерлейкина-10, трансформирующего фактора роста-1β) цитокинов, а также β-эндорфина в крови при экспериментальном ишемическом инсульте на 1, 3, 7 и 14 сутки. 2. Изучить динамику морфологических изменений в очаге острой фокальной ишемии на 1, 3, 7 и 14 сутки после моделирования ишемического инсульта. 3. Изучить влияние ТЭС-терапии на уровень про- (интерлейкина-1β, интерлейкина-6, фактора некроза опухоли-α) и противовоспалительных (интерлейкина-10, трансформирующего фактора роста-1β) цитокинов, β-эндорфина в крови при экспериментальном ишемическом инсульте на 1, 3, 7 и 14 сутки. 4. Изучить влияние ТЭС-терапии на динамику морфологических изменений в очаге острой фокальной ишемии на 1, 3, 7 и 14 сутки после моделирования ишемического инсульта. Научная новизна исследования Впервые установлены закономерности изменений цитокинового профиля у крыс в разные сроки после моделирования ИИ, и их ассоциация с динамикой морфологических изменений в очаге острой фокальной ишемии и уровнем β-эндорфина в крови. Впервые установлены закономерности изменений цитокинового профиля у крыс в разные сроки после моделирования ИИ при применении ТЭС-терапии. 14 Впервые представлено патогенетическое обоснование эффективности применения ТЭС-терапии в острейшем периоде ИИ для ограничения поражения мозга. Впервые обоснована возможность применения ТЭС-терапии с целью профилактики последствий ИИ. Полученные данные позволили получить целостную картину изменения баланса про- и противовоспалительных цитокинов, роли эндогенной опиоидергической стресс-лимитирующей системы в динамике морфологических изменений в очаге острой локальной церебральной ишемии при экспериментальном ИИ. Проведенные исследования дополняют представления о возможности нейропротекторного воздействия ТЭС-терапии в острейшем периоде ИИ и ее влиянии на размеры формирующегося очага инфаркта мозга. Результаты исследования вносят вклад в изучение патогенеза ИИ, расширяют представление о нарушениях цитокинового гомеостаза и деятельности опиоидергической стресс-лимитирующей системы при данной форме патологии, а также возможности их коррекции с помощью ТЭС-терапии. Практическая значимость Экспериментальные исследования показали принципиальную возможность использования ТЭС-терапии в коррекции цитокинового статуса в острейшем периоде ИИ. Результатом работы стало патогенетическое обоснование использования ТЭС-терапии как немедикаментозного метода терапии в острейшем периоде ИИ. Применение ТЭС-терапии позволяет путем коррекции цитокинового статуса и нарушений активности эндогенной опиоидергической системы ограничивать расширение очага инфаркта мозга при развитии экспериментального ИИ. Достоинством ТЭС-терапии является ее доступность, простота в выполнении, отсутствие больших экономических затрат, хорошая переносимость. 15 Основные положения, выносимые на защиту: – динамика показателей цитокинового статуса при развитии экспериментального ишемического инсульта характеризуется преобладанием содержания провоспалительных над уровнем противовоспалительных цитокинов; – при развитии экспериментального ишемического инсульта наблюдается падение концентрации β-эндорфина; – применение ТЭС-терапии как до, так и после моделирования ишемического инсульта позволяет повысить активность опиоидергической системы и нормализовать цитокиновый статус; – ТЭС-терапия рекомендуется для использования в качестве нейропротекторного средства в комплексном лечении больных в острейшем периоде ишемического инсульта. Апробация работы. Основные положения работы представлялись и обсуждались на Международной научной конференции «Фундаментальные исследования», Доминиканская республика, 2013; Международной научной конференции «Приоритетные направления развития науки, технологий и техники», Италия, 2014; Международной научной конференции Фундаментальные исследования, Доминиканская республика, 2014; Международной научной конференции «Инновационные медицинские технологии», Франция, 2014; Научно-практической конференции молодых ученых и студентов юга России «Медицинская наука и здравоохранение», Россия (Краснодар), 2014. Внедрение результатов исследования. Результаты диссертационной работы внедрены на кафедре общей и клинической патофизиологии ГБОУ ВПО КубГМУ Минздрава России (приложение 2) и кафедре клинической иммунологии, аллергологии и лабораторной диагностики ФПК и ППС ГБОУ ВПО КубГМУ Минздрава России (приложение 3). Публикации. По материалам диссертации опубликовано 13 работ, из них 7 в журналах, рекомендованных ВАК (приложение 1). 16 Личный вклад автора в исследование. Автором разработаны основные идеи и алгоритмы получения и обработки результатов экспериментального исследования. Автор самостоятельно провел сравнительный анализ современной литературы по проблеме эффективности диагностики, профилактики и лечения острого нарушения мозгового кровообращения с использованием немедикаментозных средств. Разработано и осуществлено многократное моделирование ИИ путем создания у крыс острой локальной церебральной ишемии за счет коагуляции правой средней мозговой артерии на экспериментальных животных. Автор самостоятельно контролировал условия содержания животных в виварии, производил забор биологического материала и проводил морфологические и участвовал в проведении иммуноферментных исследований. Автором разработаны протоколы исследования, позволяющие зафиксировать и подвергнуть статистической обработке данные по теме диссертационного исследования. На основании проведенных исследований сделаны достоверные, обоснованные выводы и разработаны практические рекомендации. Авторский вклад в выполнении работы 80 % и в написание научных работ по теме диссертации 70 %. Объем и структура диссертации Диссертация изложена на 174 страницах машинописного текста, который включает 21 таблицу и 63 рисунка. Работа состоит из введения, обзора литературы, описания материалов и методов исследования, 2-х глав, отражающих результаты собственных исследований, обсуждения полученных результатов, выводов, списка литературы, содержащего 122 отечественных и 144 иностранных источников и 3-х приложений. 17 ГЛАВА I. ОБЗОР ЛИТЕРАТУРЫ 1.1. Этиология ишемического инсульта ИИ — острое нарушение кровообращения в ГМ с развитием стойких симптомов поражения, вызванных инфарктом в мозговое вещество [Е.И. Гусев, А.Н. Коновалов, А.Б. Гехт, 2007]. Факторы риска развития ИИ, ассоциируются с повышенной частотой развития сердечно-сосудистых заболеваний [Рекомендации по ведению больных с ишемическим инсультом и транзиторными ишемическими атаками (ESO), 2008]. Их можно разделить на не модифицируемые (возраст, пол, наследственная предрасположенность) и модифицируемые (артериальная гипертензия любого происхождения, заболевания сердца, фибрилляция предсердий, инфаркт миокарда в анамнезе, дислипопротеинемия, сахарный диабет, бессимптомное поражение сонных артерий). Выделяют также факторы риска связанные с образом жизни: табакокурение, избыточная масса тела, низкий уровень физической активности, неправильной питание, острый стресс или длительное психоэмоциональное напряжение [Рекомендации по ведению больных с ишемическим инсультом и транзиторными ишемическими атаками (ESO), 2008; Е.И. Гусев, А.Н. Коновалов, В.И. Скворцова и соавт., 2009; E.C. Jauch, J.L. Saver, H.P. Adams et al., 2013]. 1.2. Классификация ишемического инсульта Наиболее употребляемой классификацией ишемического инсульта является TOAST (Trial of Org 10172 in Acute Stroke Treatment) [H.P. Adams, B.H. Bendixen, L.J. Kappelle et al., 1993; E.C. Jauch, J.L. Saver, H.P. Adams et al., 2013], согласно ей в зависимости от основного механизма возникновения острой локальной церебральной ишемии, выделяют пять патогенетических вариантов инсульта: 18 – атеротромботический возникает в результате стеноза или окклюзии крупных артерий, являющихся следствием атеротромбоза или фрагментации атеросклеротической бляшки; – кардиоэмболический — источником эмболов, приводящих к окклюзии сосудов мозга, является сердце (аритмии, фибрилляция предсердий, патология клапанного аппарата сердца, инфаркт миокарда); – лакунарный — развивается вследствие окклюзии артерий малого калибра (артериальная гипертензия и сахарный диабет); – гемодинамический — связан с нарушениями гемодинамики и развитием острой локальной церебральной ишемии, не атеросклеротическими васкулопатиями, расслоением стенки артерий, состояниями, сопровождающимися гиперкоагуляцией крови. – неизвестного происхождения (инсульты с неустановленной причиной или с наличием двух и более возможных причин). По степени тяжести выделяют [В.Н. Шток, О.С. Левин, 2006]: – малый инсульт- возникающая неврологическая симптоматика регрессирует в течение первых 21 дней заболевания; – инсульт легкой и средней тяжести — преобладает очаговая неврологическая симптоматика; – тяжелый инсульт — присутствует выраженная общемозговая симптоматика с угнетением сознания, признаками отека мозга, грубым очаговым неврологическим дефицитом, часто с дислокационными синдромами, вегетативными и трофическими нарушениями; В зависимости от локализации инфаркта мозга в определенном сосудистом бассейне выделяют [МКБ-10; В.Н. Шток, О.С. Левин, 2006]: синдром средней мозговой артерии; синдром передней мозговой артерии; синдром задней мозговой артерии; синдромы верхней, передней и задней нижних мозжечковых артерий и другие сосудистые синдромы головного мозга. 19 1.3. Общие аспекты патогенеза ишемического инсульта Ткань ГМ характеризуется чрезвычайно высоким уровнем аэробного энергетического метаболизма. Отсюда следует, что именно уровень оксигенации, напрямую связанный с кровоснабжением, является лимитирующим фактором определяющим жизнеспособность ткани мозга в краткосрочной перспективе [Е.И. Гусев, В.И. Скворцова, 2001; S.H. Graham, J. Chen, 2001; J. Graulich et al., 2002; A.N. Clarkson, B.A. Sutherland, I. Appleton, 2005; A. Durukan, T. Tatlisumak, 2007; К.П. Иванов, 2010; М. Бэр, 2011; Ю.И. Зильбертер, Т.М. Зильбертер, 2012; N.W. Manning et al., 2014]. Поэтому основу патогенеза ИИ, вне зависимости от патогенетического варианта его развития, составляет «острая локальная церебральная ишемия» и реакция организма на нее, определяемая, прежде всего, реактивностью единой нейроиммуноэндокринной системы [Е.И. Гусев, В.И. Скворцова, 2001; А.Х. Каде, А.П. Парахонский, 2004; А.А. Мольберг, Т.В. Гришина, 2006; А.Ф. Повещенко, В.В. Абрамов, В.А. Козлов, 2007; А.А. Олейник, Р.С. Вастьянов, 2008; М.А. Самоструева, Д.Л. Теплый, И.Н. Тюренков, 2009; Г.Н. Крыжановский, С.В. Магаева, 2010; Ж.В. Титова, Г.М. Бодиенкова, 2013]. «Острая локальная церебральная ишемия» означает острое ограничение кровоснабжения ГМ в отдельном артериальном бассейне. Она является динамическим и потенциально обратимым процессом [Е.И. Гусев, В.И. Скворцова, 2001; W.S. Smith, 2004; К.П. Иванов, 2010; М. Бэр, 2011; N.W. Manning et al., 2014]. Инфаркт мозга — область нежизнеспособной ткани мозга, возникшая, в том числе, в результате острой локальной церебральной ишемии. Инфаркт является зоной необратимого морфологического дефекта [Е.И. Гусев, В.И. Скворцова, 2001; Е.И. Гусев, М Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014;]. Последовательность метаболических реакций ткани ГМ, возникающих в ответ на снижение мозгового кровотока [T.R. Anderson, R.D. Andrew, 2002; W.S. Smith, 2004; Л.П. Соколова, 2011; K.A. Hossmann, 2012; Е.И. Гусев, 20 М.Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014;] подразделяют на несколько критических этапов: ● первый критический этап — развивается при снижении кровотока до 70 % от нормального уровня (менее 50–55 мл на 100 гр вещества мозга в минуту) — возникает торможение белкового синтеза с изменением профиля образующихся белков; ● второй критический этап — развивается при дальнейшем снижении кровотока до 45–50 % от нормальной величины (до 35 мл на 100 гр вещества мозга в минуту) — происходит активация анаэробного гликолиза, завершающаяся развитием тканевого лактат-ацидоза и цитотоксического отека; ● третий критический этап — наступает при дальнейшем снижении кровотока до 20 мл на 100 гр вещества мозга в минуту — характеризуется прогрессирующим энергодефицитом, приводящим к нарушению активного ионного транспорта и мембранных электрических процессов, сопровождается выбросом возбуждающих аминокислот и затруднением их обратного поглощения (ранняя фаза глутаматной эксайтотоксичности); ● четвертый критический этап — наступает при снижении мозгового кровотока до 20 % от нормальной величины (10–15 мл на 100 гр вещества мозга в минуту) — возникает необратимое повреждение клеток, характеризующееся аноксической деполяризацией мембран; Современные представления о патогенезе церебральной ишемии, позволили создать принципиальную схему молекулярных реакций ткани мозга в ответ на ее развитие, так называемый «ишемический каскад» [Е.И. Гусев, В.И. Скворцова, 2001; A. Aggarwal, P. Aggarwal, M. Khatak et al., 2010]: 1 этап — снижение мозгового кровотока; 2 этап — глутаматная «эксайтотоксичность»; 3 этап — внутриклеточное накопление кальция; 4 этап — активация внутриклеточных ферментов; 5 этап — повышение синтеза NO и развитие оксидативного стресса; 6 этап — экспрессия генов; 21 7 этап — «отдаленные» последствия ишемии (местная воспалительная реакция, микроваскулярные нарушения, повреждения гемато-энцефалического барьера); 2–8 этапы — активация апоптоза. Область острой локальной церебральной ишемии характеризуется наличием градиента кровоснабжения, которое возрастает от центра к периферии формирующегося очага инфаркта [L. Rohl et al., 2001; W.S. Smith, 2004; N.W. Manning et al., 2014]. Соответственно этому, в области локальной церебральной ишемии можно выделить «ядро» — зону с крайне низким уровнем кровотока, в которой находится ткань, погибшая преимущественно путем некроза, и «пенумбру» — зону со сниженным уровнем кровотока (не менее 20 мл на 100 грамм вещества мозга за минуту), ввиду чего, не смотря на нарушения функционирования нейронов, они остаются жизнеспособными [L. Rohl et al., 2001; W.S. Smith, 2004; Е.И. Гусев, М.Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014]. Зона «ядра» в ткани мозга, с уровнем кровоснабжения ниже 10 мл на 100 гр вещества мозга за 1 минуту, формируется в течение 6–8 минут после развития острой локальной церебральной ишемии [L. Rohl et al., 2001; W.S. Smith, 2004; Е.И. Гусев, М.Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014]. Окончательное формирование очага инфаркта мозга, развивающееся за счет уменьшения объема зоны «пенумбры» в пользу зоны «инфаркта», завершается через 3–6 часов с момента возникновения нарушений мозгового кровообращения [L. Rohl et al., 2001; W.S. Smith, 2004; A. Bose et al., 2008; Е.И. Гусев, М.Ю. Мартынов, П.Р. Камчатнов, 2013; N.W. Manning et al., 2014]. Во многих исследованиях показано, что прирост значительной части области инфаркта мозга происходит вследствие асептического воспаления, за счет развития процессов некроза и апоптоза в зоне пенумбры в течение 72 часов с момента развития острой локальной церебральной ишемии [S.H. Graham, J. Chen, 2001; M.P. Mattson et al., 2001; N.W. Manning et al., 2014]. 22 В основе расширения зоны инфаркта мозга, в течение первой недели после возникновения инсульта, лежит острое асептическое воспаление. В более поздние сроки идет организация зоны инфаркта мозга, за счет процессов резорбции тканевого детрита и формирования глиомезодермального рубца, что сопровождается некоторым уменьшением объема области инфаркта [K.J. Becker, 2001; G.J. del Zoppo, K.J. Becker, J.M. Hallenbeck, 2001; M.D. Ginsberg, 2003; M. Fisher, R. Ratan, 2003; P.L. Wood, 2003; R. Markus, D.C. Reutens, S. Kazui, 2004; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; Q. Wang, X.N. Tang, M.A. Yenari, 2007; А.Х. Хана-Мурад, Л.И. Павлинова, А.А. Мокрушин, 2007; A. Tuttolomondo, D. Di Raimondo, R. Di Sciacca, 2008; J. Jordan et al., 2008; D. Amantea et al., 2008; C. Benakis, L. Hirt, R.A. Du Pasquier, 2009; Ю.Д. Губарев, А.О. Шеремет, 2009; А.С. Кость, Б.Д. Луцик, Л.Е. Лаповець, 2010; A.G. Ceulemans et al., 2010; Y. Lee et al., 2014; М.В. Онуфриев и соавт., 2014]. Прирост объема инфаркта ткани мозга может быть вызван ростом потребления энергии клетками зоны пенумбры в условиях недостаточного кровоснабжения, связанным с периинфарктной деполяризацией (волны деполяризации) [E. Pinard et al., 2001; T.R. Anderson, R.D. Andrew, 2002; M. Umegaki et al., 2005; M. Fabricius et al., 2006]. На основании этого выделяются следующие временные периоды в развитии ИИ [Е.И. Гусев и соавт., 2009]: – острейший период — первые 3 суток; – острый период — до 28 суток; – ранний восстановительный период — до 6 месяцев; – поздний восстановительный период — до 2-х лет; – период остаточных явлений — после 2-х лет. 1.4. Роль воспаления в патогенезе ишемического инсульта Острое асептическое воспаление является тем патологическим процессом, который лежит в основе расширения зоны инфаркта мозга в течение первой недели после возникновения инсульта [K.J. Becker, 2001; G.J. del Zop- 23 po, K.J. Becker, J.M. Hallenbeck, 2001; M.D. Ginsberg, 2003; M. Fisher, R. Ratan, 2003; P.L. Wood, 2003; R. Markus, D.C. Reutens, S. Kazui, 2004; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; Q. Wang, X.N. Tang, M.A. Yenari, 2007; А.Х. ХанаМурад, Л.И. Павлинова, А.А. Мокрушин, 2007; A. Tuttolomondo, D. Di Raimondo, R. Di Sciacca, 2008; J. Jordan et al., 2008; D. Amantea et al., 2008; C. Benakis, L. Hirt, R.A. Du Pasquier, 2009; Ю.Д. Губарев, А.О. Шеремет, 2009; А.С. Кость, Б.Д. Луцик, Л.Е. Лаповець, 2010; A.G. Ceulemans et al., 2010; Y. Lee et al., 2014; М.В. Онуфриев и соавт., 2014]. Развитие воспалительной реакции в очаге ишемии связано с изменением содержания цитокинов: провоспалительных — интерлейкин-1 (ИЛ-1), фактор некроза опухоли-α (ФНО-α), интерлейкин-6 (ИЛ-6), интерлейкин-8 (ИЛ-8), а также противовоспалительных интерлейкин-4 (ИЛ-4), интерлейкин-10 (ИЛ-10), трансформирующий фактор роста-1β (ТФР-1β) [Е.И. Гусев, В.И. Скворцова, 2001; N. Quan, M. Herkenham, 2002; А.А. Аракелян и соавт., 2005; Л.Н. Кашаева, Л.М. Карзакова, В.Н. Саперов, 2005; Q. Wang, X.N. Tang, M.A. Yenari, 2007; D. Amantea et al., 2008; M.Y. Zuidema, C. Zhang, 2010; М.В. Онуфриев, 2014]. Такие цитокины как интерлейкин-1α (ИЛ-1α) и интерлейкин-1β (ИЛ-1β), ФНО-α являются ключевыми медиаторами микроглиальных нейроиммунных функций и вырабатываются локально в ответ церебральную ишемию [A.C.H. Yu, L.T. Lau, 2000; H. Boutin et al., 2001; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; A. Tuttolomondo et al., 2008; S. Shenhar-Tsarfaty et al., 2008; S. Choi, W.J. Friedman, 2009; A. Maddahi, L. Edvinsson, 2010; M. Zhang et al., 2011]. Их продуцируют активированные лейкоциты, поврежденные нейроны, микроглиоциты, астроциты [С.М. Белоцкий, Р. Р. Авталион, 2008]. Известно, что к запуску воспалительной реакции приводит именно поражение нейронов, в свою очередь неповрежденные нейроны наоборот предотвращают способность к индуцированию экспрессии генов участвующих в процессе воспаления [R.M. Adibhatla, J.F. Hatcher, 2007; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; H.K. Jeong et al., 2010; Y. Huang et al., 2011]. В условиях церебральной ишемии синтез ИЛ-1 микроглией является главным активирующим сигналом для запуска образования других провоспалитель- 24 ных цитокинов, а также стимуляции астроцитов к продукции потенциальных нейротоксичных веществ, таких как оксид азота (NO) и метаболиты арахидоновой кислоты [H. Boutin et al., 2001; L.D. Church, P.G. Cook, M.F. Mc Dermott, 2008; S. Choi, W.J. Friedman, 2009; C.A. Dinarello, 2009; C.A. Dinarello, 2011; H.F. Green et al., 2012; J. Galea, D. Brough, 2013; C. Garlanda, C. A. Dinarello, A. Mantovani, 2013]. Эндогенным протективным агентом защищающим мозг от негативных эффектов ИЛ-1 является рецепторный антагонист ИЛ-1 (РАИЛ-1), при снижении биологической активности которого отмечается усиление гибели нейронов в ишемизированной области мозга [С.А. Кетлинский, А.С. Симбирцев, 2008; Е.В. Супрун и соавт., 2009; J. Galea, D. Brough, 2013]. Действие ФНО-α — плейотропного полипептида, вырабатываемого нейтрофилами (Нф), активированными макрофагами (МФ), астроцитами и эндотелиальными клетками, во многом сходно с действием ИЛ-1 [J. Zaremba, 2000; С.А. Кетлинский, А.С. Симбирцев, 2008]. ФНО-α оказывает токсическое действие на нейроны путем активации системы генерации активных форм кислорода (АФК), что резко индуцирует процессы перекисного окисления липидов (ПОЛ), в том числе, за счет взаимодействия с сигнальной системой сфингомиелинового цикла [J.J. Viel, D.Q. Mc Manus, S.S. Smith, 2001; A. Quintana et al., 2005; А.Е. Бугрова, 2007; R.M. Adibhatla, J.F. Hatcher, 2007; R.M. Adibhatla, R. Dempsey, J.F. Hatcher, 2008; N. Liu et al., 2009; А.В. Алексеенко, 2013]. Церамид, в большом количестве синтезирующийся в очаге церебральной ишемии, является одним из основных продуктов сфингомиелинового цикла. Он способствует увеличению содержания ФНО-α в мозге, непосредственно вызывает нарушения в цепи переноса электронов, повышает проницаемость митохондриальной мембраны, что приводит резкому увеличению продукции активных форм кислорода [А.Е. Бугрова, 2007; А.В. Алексеенко, 2013]. Снижение клеточной концентрации глутатиона наблюдающееся в условиях окислительного стресса при церебральной ишемии, приводит к активации нейтральной сфингомиелиназы, что увеличивает синтез церамида [А.Е. Бугрова, 2007]. 25 Таким образом порочный круг замыкается, увеличение концентрации эндогенного ФНО-α, вследствие асептического воспаления в области инфаркта мозга, за счет активации сфингомиелиназы, приводит к росту продукции церамида, а ФНО-α индуцирует образование активных форм кислорода в клетках, в результате чего развивается активация реакций свободнорадикального окисления, в том числе перекисного окисления липидов, дефицит антиоксидантов (в том числе глутатиона), что в свою очередь дополнительно повышает уровень активности сфингомиелиназы приводя к образованию новых порций церамида и ФНО-α [А.Е. Бугрова, 2007; А.В. Алексеенко, 2013]. ФНО-α помимо токсического влияния, в низкой (физиологической) концентрации, оказывает защитный эффект на нейроны, путем увеличения экспрессии факторов антиоксидантной системы [J.J. Viel, D.Q. Mc Manus, S.S. Smith, 2001; A. Quintana et al., 2005]. ИЛ-6 — является типичным ранним индуцибельным цитокином и синтезируется многими типами клеток, участвующими в инициации и регуляции воспаления и иммунного ответа [D.A. Papanicolaou et al., 1998; С.А. Кетлинский, А.С. Симбирцев, 2008]. ИЛ-6 оказывает выраженное провоспалительное действие за счет активации экспрессии молекул адгезии и активации хемотаксиса лейкоцитов, а также является важнейшим индуктором острофазного ответа [K. Paul-Pletzer, 2006; I.M. Cojocaru et al., 2009; С.А. Кетлинский, А.С. Симбирцев, 2008; T. Tanaka et al., 2013]. Установлено, что ИЛ-1 и ФНО-α являются мощными индукторами ИЛ-6-синтетазы в астроцитах: быстрое увеличение содержания микроглиального ИЛ-1β в первые 1-2 часа церебральной ишемии вызывает усиленный синтез ИЛ-6 [H. Boutin et al., 2001]. Важно отметить, что ИЛ-1, ИЛ-6, ФНО-α регулируют функцию гипоталамо-гипофизарно-надпочечниковой системы (ГГНС), непосредственно воздействуя на переднюю долю гипофиза, что задействует стресс-реализующие системы в ответ на ишемию мозга, в рамках общей реакции нейроиммуноэндокринной системы [A. Johansson et al., 2000; N. Quan, M. Herkenham, 2002; F. Eskandari, J.I. Webster, M.S. Esther, 2003; И.Г. Акмаев, В.В. Гриневич, 2003; С.А. Кетлинский, А.С. Симбирцев, 2008]. 26 Существует прямо пропорциональная зависимость между изменением концентрации провоспалительных цитокинов (ИЛ-1, ФНО-α, ИЛ-6, ИЛ-8) при развитии острой локальной церебральной ишемии и конечными размерами инфаркта мозга (зоной некроза) [Е.И. Гусев, В.И. Скворцова, 2001; H. Boutin et al., 2001; K.Y. Kwan, B.H. Jeon, 2001; N. Quan, M. Herkenham, 2002; Q. Wang, X.N. Tang, M.A. Yenari, 2007; D. Amantea et al., 2008; I.M. Cojocaru et al., 2009; M. Zhang et al., 2011; М.В. Онуфриев, 2014]. Противовоспалительные цитокины, такие как ИЛ-10 и ТФР-1β, подавляя развитие воспаления, оказывают защитный эффект в условиях ишемии ГМ [K. Strle et al., 2001; Y. Zhu et al., 2002; N. Quan, M. Herkenham, 2002; N. Vila et al., 2003; Г.Н. Жданов, 2006; M. Kaste, H.J. Castillo, S. de Compostela, 2006; N. Liu et al., 2009]. Хорошо известен нейропротективный эффект ТФР-1β, который обеспечивается за счет угнетения иммунных реакций в ткани мозга и понижения чувствительности клеток ГМ к ишемии [J.J. Letterio, 2000; Y. Zhu et al., 2002]. Дополнительное повреждение мозговой ткани при ИИ обусловлено активацией сосудистого эндотелия провоспалительными цитокинами (ИЛ-1β, ФНО-α и ИЛ-6) [D. Stanimirovic, K. Satoh, 2000; J.P. Konsman, B. Drukarch, A.M. van Dam, 2007; R. Tanasescu et al., 2008]. Процесс эндотелиальной активации сопровождается повышением экспрессии факторов адгезии (ICAM-1, CD11α и CD18), благодаря которым происходит прикрепление лейкоцитов к эндотелию [M. Fisher, R. Ratan, 2003; Q. Wang, X.N. Tang, M.A. Yenari, 2007; R. Tanasescu et al., 2008; M. Zhang et al., 2011]. ИЛ-8, выработка которого резко возрастает под действием ИЛ-1β и ФНО-α, активирует миграцию лейкоцитов с повреждением сосудистой стенки, что приводит к повышению проницаемости гематоэнцефалического барьера (ГЭБ) и сопровождается воспалительной инфильтрацией зоны церебральной ишемии [J.P. Konsman, B. Drukarch, A.M. van Dam, 2007]. Особое значение имеет миграция моноцитов в ткань ГМ. Кроме прямого повреждающего действия, треть из них дифференцируется в клетки по функции и морфологии подобные микроглиоцитам [J. Priller, A. Flugel, T. Wehner, 2001; M. Schilling et al., 2005]. 27 Прогрессирование повреждения в зоне пенумбры может происходить не только под влиянием абсолютного увеличения концентрации провоспалительных цитокинов, но и вследствие недостатка противовоспалительных и нейротрофических факторов [K. Strle et al., 2001; Alzheimer, C., 2002; N. Vila et al., 2003]. Защитное противоишемическое действие ТФР-1β проявляется угнетением адгезии лейкоцитов к эндотелию, подавлением выработки МФ микроглией повреждающих кислородных и NO метаболитов [J.J. Letterio, 2000; N. Vila et al., 2003]. Нейропротективная способность ТФР-1β связана со стабилизацией нейронального кальциевого гомеостаза и повышением экспрессии гена — белкового фактора регулятора апоптоза (bcl-2), что приводит к значительному торможению свободнорадикального повреждения, замедлению процессов некроза и апоптоза [J.J. Letterio, 2000; Y. Zhu et al., 2002]. Установлена важная роль ТФР-1β, в увеличении васкуляризации ткани мозга после острой церебральной ишемии, особенно в зоне пенумбры [J.J. Letterio, 2000]. ИЛ-10 является мощным регулятором клеточно-опосредованного иммунного ответа моноцитов/макрофагов: снижает продукцию простогландина Е2 (ПГЕ2), угнетает экспрессию ICAM-1 активно участвующих в развитии местного воспаления, подавляет выработку многочисленных провоспалительных цитокинов, включая ИЛ-1, ФНО-α, ИЛ-6, ИЛ-8 [С.А. Кетлинский, А.С. Симбирцев, 2008; N. Liu et al., 2009; T. Nabe et al., 2012]. Установлено также, что ИЛ-10 подавляет синтез супероксидного аниона и реактивных промежуточных продуктов обмена кислорода. Он оказывает модулирующее влияние на синтез NO в зависимости от продолжительности воздействия на активированные МФ. Содействие синтезу NO является одним из многочисленных активирующих эффектов ИЛ-10 на моноциты/МФ наряду с индукцией им выработки РАИЛ-1. Таким образом, ИЛ-10 является фактором, модулирующим функцию МФ и обладающим мощной противовоспалительной активностью, что объясняет его нейропротекторные свойства при ИИ, а его низкая концентрация в динамике заболевания наблюдается в случаях тяжелого, неблагоприятного течения инсульта [K. Strle et al., 2001; N. Vila et al., 2003; Г.Н. Жданов, 2006; N. Liu et al., 2009]. 28 Значительную роль в последующем расширении очага инфаркта мозга, которое происходит за счет гибели клеток из зоны пенумбры, играет активация клеток микроглии происходящая под воздействием провоспалительных цитокинов [K.J. Becker, 2001; M.D. Ginsberg, 2003; Y. Lee et al., 2014]. Клетки микроглии являются единственными иммунокомпетентными клетками центральной нервной системы (ЦНС) [Р.И. Сепиашвили, 2013]. Они активно участвуют в формировании асептического воспаления, развивающегося при острой локальной церебральной ишемии [Y. Lee et al., 2014] и тем самым вносят большой вклад как в процесс формирования окончательного объема инфаркта мозга, так и в процесс последующей организации зоны деструкции в ткани мозга [K.J. Becker, 2001; M.D. Ginsberg, 2003; M. Schilling et al., 2005; Q. Wang, X.N. Tang, M.A. Yenari, 2007; A.G. Ceulemans et al., 2010; Y. Lee et al., 2014]. Активированные микроглиоциты синтезируют большой спектр нейротоксичных веществ, приводя тем самым к поражению клеток в зоне пенумбры и расширению очага инфаркта мозга [A. Ceulemans et al., 2010; Y. Lee et al., 2014]. Микроглиоциты, являясь основными поставщиками провоспалительных цитокинов и свободных радикалов, поддерживают воспаление в очаге ишемии, обусловливая тем самым дополнительные нейрональные потери. Активно участвуя в процессах репарации ткани мозга, клетки микроглии вносят большой вклад в формирование глиального рубца [A.Y. Lay, K.G. Todd, 2006; Q. Wang, X.N. Tang, M.A. Yenari, 2007; Y. Lee et al., 2014]. Большое значение в процессе асептического воспаления, возникающего в ответ на повреждение ткани мозга, играют астроциты, являясь одними из основных источников продукции провоспалительных цитокинов, свободных радикалов и других нейротоксичных веществ, но наряду с этим они синтезируют и некоторые нейропротективные агенты — ТФР-1β [J.J. Letterio, 2000]. Систематизация знаний по микроциркуляторным и клеточный реакциям в очаге острой локальной церебральной ишемии позволила нам представить их в виде хронологической последовательности — V стадий микроциркуляторно-клеточного каскада [Y. Tomita et al., 2002]. 29 1.5. Роль системного воспаления в патогенезе ишемического инсульта Синдром системного воспалительного ответа (ССВО) — воспалительный ответ на уровне целостного организма, в частности при состояниях без четко определяемой бактериемии [F. Hietbrink et al., 2006; С.М. Белоцкий, Р.Р. Авталион, 2008; J.M. Sinning et al., 2012; В.В. Иващенко и соавт., 2012]. Вследствие нарушения гематоэнцефалического барьера (ГЭБ) с последующей генерализацией воспалительного процесса при ИИ наблюдается развитие ССВО, который еще больше отягощает течение заболевания и играет значительную роль в развитии и прогрессировании экстрацеребральной патологии [D. Stanimirovic, K. Saton, 2000; M. Di Napoli, F. Papa, 2006; B.W. Mc Coll, N.J. Rothwell, S.M. Allan, 2007]. Переход постишемического воспалительного ответа с локального уровня на системный обусловлен нарушением регуляторных механизмов контролирующих развитие воспаления и в основе его развития лежит значительный рост концентрации провоспалительных цитокинов (ИЛ-1β, ИЛ-6, ФНО-α) в системной циркуляции [P.E. Marik, G.P. Zaloga, 2001; А.А. Аракелян и соавт., 2005; M. Di Napoli, F. Papa, 2006; B.W. Mc Coll, N.J. Rothwell, S.M. Allan, 2007; С.М. Белоцкий, Р.Р. Авталион, 2008]. Одной из основных причин развития ССВО организма при ИИ является нарушение целостности ГЭБ, которое способствует увеличению притока иммуномодуляторов в ЦНС, что в свою очередь еще больше усиливает воспалительный ответ [D. Stanimirovic, K. Saton, 2000; K.J. Becker, 2001; M. Di Napoli, F. Papa, 2006]. При этом ИЛ-1β попавший в системный кровоток индуцирует образование ФНО-α, они вместе запускают провоспалительный каскад в результате чего возрастает продукция ИЛ-6, что в целом приводит к росту содержания и активации Нф в кровотоке, в дальнейшем эти процессы приводят развитию эндотелиальной дисфункции [H. Hirasawa et al., 2004; M.F. Osuchowski et al., 30 2006; F. Hietbrink et al., 2006; С.М. Белоцкий, Р.Р. Авталион, 2008; D.H. Yu, B. Kim, J. Park, 2011]. ИЛ-1 при поступлении в системный кровоток является важнейшим медиатором острофазного ответа на уровне организма обеспечивая: запуск единого нейроимунноэндокринного ответа организма на воспаление; активацию циркулирующих иммунных клеток (в первую очередь НФ) и запуск иммунопоэза; запуск синтеза белков острой фазы (БОФ) в печени [С.А. Кетлинский, А.С. Симбирцев, 2008]. ИЛ-6 является важнейшим регулятором иммунитета, главной его функцией является переключение реакций первоначально развивающегося воспаления по механизмам врожденного иммунитета на реакции приобретенного иммунитета [S. Jones, 2005]. Гиперпродукция ИЛ-6 приводит к запуску воспалительных и аутоиммунных заболеваний человека [K. Ishihara, T. Hirano, 2002]. ФНО-α играет главную роль в прогрессировании полиорганной недостаточности при ССВО, его уровень коррелирует с летальностью пациентов с синдромом системного воспалительного ответа, а использование антицитокиновых препаратов зачастую предотвращает летальный исход [С.А. Кетлинский, А.С. Симбирцев, 2008]. Известно, что системное действие провоспалительных цитокинов ИЛ-1β, ИЛ-6, ФНО-α приводит к прогрессированию имеющейся кардиальной патологии, так ФНО-α является важнейшим агентом вызывающим развитие ремоделирования миокарда, потерю сократимости кардиомиоцитов, его содержание является важнейшим прогностическим фактором развития неблагоприятного исхода хронической сердечной недостаточности [А.Я. Кравченко, Ю.М. Черняева, 2013]. Известные факты гиперпродукции противовоспалительных цитокинов (ИЛ-4, ИЛ-10, РАИЛ-1, ТФР-1β) у больных сепсисом привели к появлению термина «синдром компенсированного антивоспалительного ответа» (СКАО) [M.M. Faas et al., 2002; С.М. Белоцкий, Р.Р. Авталион, 2008]. В настоящее время считается, что СКАО развивается при системной воспалительной реакции, выполняя функции ограничения гиперпродукции 31 провоспалительных цитокинов и системного воспаления, что может приводить к развитию иммунодефицитного состояния у некоторых пациентов [L. Perry, C.P. Love, 2001; K. Tschaikowsky et al., 2002; F. Venet et al., 2008; A. Vogelgesang et al., 2010]. Тесное взаимодействие цитокинов и гормонов на центральном и периферическом уровнях объясняет включение стресс-реализующих и стресслимитирующих систем в ответ на ишемию мозга в рамках общей реакции нейроиммуноэндокринной системы [A. Johansson et al., 2000; N. Quan, M. Herkenham, 2002; F. Eskandari, J.I. Webster, M.S. Esther, 2003; И.Г. Акмаев, В.В. Гриневич, 2003; С.А. Кетлинский, А.С. Симбирцев, 2008; Л.К. Хныченко, Н.С. Сапронов, 2003; М.А. Самоструева, Д.Л. Теплый, И.Н. Тюренков, 2009; В.В. Иващенко и соавт., 2012]. Результаты изучения динамики изменений уровней цитокинов показывают, что для ССВО характерно двух стадийное развитие. В ранней стадии наблюдается резкое увеличение синтеза провоспалительных цитокинов, затем в поздней — происходит постепенное истощение их выработки [K. Tschaikowsky et al., 2002; С.М. Белоцкий, Р.Р. Авталион, 2008; A. Vogelgesang et al., 2010; J.M. Sinning et al., 2012]. Однако у части больных переживших фазу гиперцитокинемии, в последующие дни развивается иммуносупрессивное состояние, которое проявляется присоединением вторичной госпитальной инфекции [K. Tschaikowsky et al., 2002; С.М. Белоцкий, Р.Р. Авталион, 2008; A. Vogelgesang et al., 2010; J.M. Sinning et al., 2012]. 1.6. Понятие об опиоидергической нейротрансмиттерной системе Опиоидные рецепторы (ОР), с которыми реагирует широкий набор эндогенных лигандов, представлены 5 типами: µ, δ, κ, σ, ε [А.А. Зозуля, С.Ф. Пшеничкин, 1990; Л. Н. Маслов, Ю.Б. Лишманов, Г.Н. Смагин, 2002]. Существуют классические типы опиоидных рецепторов (µ, δ и κ) все эффекты которых блокируются полным рецепторным антагонистом опиоидных пептидов налоксо- 32 ном, а также так называемые неопиоидные — стимуляция которых налоксоном не блокируется [Л.Н. Маслов, Ю.Б. Лишманов, Г.Н. Смагин, 2002; В.А. Дубынин, А.А. Каменский, 2010; Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011]. Различные лиганды ОР по-разному взаимодействуют с указанными типами рецепторов, создавая сложную и многообразную систему опиоидной регуляции, которая обеспечивает контроль над различными функциями организма от нервно-психической деятельности до функционирования висцеральных органов [В.В. Брагин, 1991; В.В. Брагин, Е.О. Яснецов, 1991; R.J. Bodnar, 2011; А.Г. Полунина, Е.А. Брюн, 2013]. В первую очередь для опиоидергической нейротрансмиттерной системы характерен стресс-лимитирующий эффект, в частности ее активация может уменьшать стрессовое повреждение органа [Н.В. Соленкова, 2002; М.Г. Пшенникова, 2003; А.Г. Полунина, Е.А. Брюн, 2013], что является актуальным и для острого поражения ЦНС. Доказаны такие эффекты эндогенных ОП: антигипоксический, антиоксидантный, антиангинальный, антиишемический, антиаритмический, влияние на синтез гипофизарных гормонов, иммуномодулирующий и др. [Л.Н. Маслов и соавт., 2002; В.А. Черешнев, С.В. Гейн, 2009; В.А. Дубынин, А.А. Каменский, 2010; Т.В. Ласукова, С.В. Низкодубова, Е.Ю. Мухтобарова, 2013]. Важнейшим представителем среди семейства ОП является β-эндорфин, который является агонистом µ и δ классических опиоидных рецепторов, а также обладает действием не связанным с активацией классических ОР [В.А. Дубынин, А.А. Каменский, 2010; Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011; И.Л. Шаравьева, 2011; О.В. Нечайкина и соавт., 2013]. ОП в настоящее время рассматриваются как важнейшие посредники между нервной и иммунной системами организма [О.Б. Сеин и соавт., 2009; В.А. Дубынин, А.А. Каменский, 2010; Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011; И.Л. Шаравьева, 2011; К.Г. Горшкова, 2012]. ОР широко представлены на разных типах клеток иммунной системы, что подчеркивает их важную роль в процессах иммунорегуляции [I.H. Jonsdottir, 2000; J.M. Bidlack, 2000; H. Machelska, 33 K. Stein, 2003; Т.А. Баева, 2006; H. L. Rittner, A. Brack, C. Stein, 2008; И.Л. Шаравьева, 2011; Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011; Т.А. Баева, В.О. Небогатиков, 2012; К.Г. Горшкова, 2012; J.B.S. Garsia, M.G. de Melo Cardoso, M.C. Dos-Santos, 2012]. Показано, что стимуляция µ и δ-опиоидных рецепторов при развитии гуморального и клеточного иммунного ответа, за счет стимуляции синтеза ИЛ-4 оказывает прямое иммуносупрессивное действие и опосредованно через мононуклеары иммуностимулирующий эффект [Т.А. Баева, 2006; С.В. Гейн, Т.А. Баева, О.А Кичанова, 2006; С.В. Гейн, К.Г. Горшкова, 2008; М.А. Чейдо, Г.В. Идова, 2009; В.А. Черешнев, С.В. Гейн, 2009; И.Л. Шаравьева, 2011; Т.А. Баева, В.О. Небогатиков, 2012; K.S. Iwaszkiewicz, J.J. Schneider, S. Hua, 2013]. Активация δ- опиоидных рецепторов приводит к ингибированию эффектов ФНО-α [M.R. Hutchinson et al., 2011; N. Akhter et al., 2013]. На модели ишемии/реперфузии сетчатки получены данные свидетельствующие о том, что агонисты δ- опиоидных рецепторов уменьшают выработку ФНО-α и оказывают протективное действие [S. Husain, D.E. Potter, C.E. Crosson, 2009; S. Husain, G.I. Liou, C.E. Crosson, 2011]. Использование агонистов δ-опиоидных рецепторов приводит к уменьшению свободнорадикального повреждения ишемизированных нейронов [Y. Yang et al., 2009]. β-эндорфин действуя через опиоидные рецепторы (µ и δ) на клетки иммунной системы в целом оказывает ингибирующее действие [M.J. Finley. et al., 2008; C. Borner et al., 2009; S. Boehncke et al., 2010]. Огромную важность представляют данные показывающие нейропротекторный эффект активации δ-опиоидных рецепторов [M. Iwata et al., 2007; D.S. Su et al., 2007; M. Govindaswami et al., 2008; M. Staples et al., 2013]. По данным литературы некоторые агонисты δ-опиоидных рецепторов способны индуцировать выработку ТФР-1β, который при ишемии мозга оказывает нейропротекторное действие [C. Happel et al., 2008]. В условиях гипоксии показано активирующее действие δ-опиоидных рецепторов на экспрессию рецептора ГАМК, что потенциально способно также оказывать нейропротективное действие [Y. Feng et al., 2011]. 34 Обращает на себя внимание тот факт, что β-эндорфин посредством активации µ и δ опиоидных рецепторов значительно повышает экспрессию мРНК BDNF, что также может лежать в основе его нейропротекторного действия [H. Zhang et al., 2006]. Феномен прекондиционирования при использовании β-эндорфина, при связывании с µ и δ опиоидными рецепторами реализуется за счет деятельности АТФ-зависимых калиевых каналов [G.J. Gross, 2000; Н.В. Соленкова, 2002; Е.И. Барзах, 2006; J.M. Gidday, 2006; A.K. Rehni, N. Singh, S.A. Jaggi, 2007; К.П. Иванов, 2013]. Основная функция опиоидных рецепторов связана с пресинаптическим торможением передачи сигнала в различных нейромедиаторных системах. Соответственно, опиоиды, как правило, оказывают ингибирующее действие на высвобождение нейротрансмиттеров, в частности снижается выброс возбуждающих медиаторов-аминокислот глутамата и аспартата [M. Kawalec, J.E. Kowalczuk, 2011]. Большинство типов ОР сопряжены с Gi/o-белками, чувствительными к пертуссин-токсину [O.A. Dravolina, E.E. Zvartau, A.Y. Bespalov, 2000]. В свою очередь Gi/o-белки подавляют активность аденилатциклазы, что приводит к уменьшению количества цАМФ и изменению состояния протеинкиназ. В результате снижается кальциевая проводимость пресинаптической мембраны и уменьшается количество высвобождаемого медиатора. Именно этот механизм был описан первым в качестве способа, с помощью которого опиоиды влияют на работу синапсов в нейронах, передающих боль, и ингибируют выброс субстанции Р [H. Machelska, K. Stein, 2003]. В дальнейшем была показана способность Gi/o-белков влиять на калиевую проводимость (как пре-, так и постсинаптическую), именно этому механизму в настоящее время отводится ведущая роль в механизме ишемического прекондиционирования (открытие АТФ-зависимых калиевых каналов) [G.J. Gross, 2000; R.M. Fryer et al., 2000; Н.В. Соленкова, 2002; Е.И. Барзах, 2006; К.Н. Мельников и соавт., 2009]. В пресинаптических окончаниях дополнительное открывание калиевых каналов повышает скорость 35 реполяризации, ограничивая, таким образом, кальциевый ток и высвобождение нейротрансмиттеров [J.D. Wood, J.J. Galligan, 2004; К.Н. Мельников и соавт., 2009]. 1.7. Основные принципы профилактики и лечения ишемического инсульта Основными видами профилактики и лечения ИИ являются: первичная и вторичная профилактика, базисная и специфическая терапия [Рекомендации по ведению больных с ишемическим инсультом и транзиторными ишемическими атаками (ESO), 2008]. Первичная профилактика Целью первичной профилактики инсульта является снижение риска его развития среди здорового населения. Первичная профилактика инсульта направлена на контроль сосудистых факторов риска. К мероприятиям по первичной профилактике в первую очередь относят — рекомендации по изменению образа жизни (диета, отказ от курения и злоупотребления алкоголем, мероприятия направленные на снижение веса), контролю уровня артериального давления, глюкозы и холестерина в крови. Вторичная профилактика Целью вторичной профилактики является снижение риска развития повторного инсульта. Она также направлена на контроль сосудистых факторов риска. К мероприятиям вторичной профилактики относят — рекомендации по изменению образа жизни, контроль уровня артериального давления, глюкозы и холестерина в крови, проведению антитромботической (антитромбоцитарная и антикоагулянтная) терапии, использование хирургических методов (стентирование и ангиопластика). 36 Базисная терапия Базисная терапия направлена на стабилизацию состояния тяжело больных пациентов и коррекцию тех нарушений, которые могут осложнить и замедлить восстановление неврологических функций [C.J. Klijn, G.J. Hankey et al., 2003; D. Leys, E.B. Ringelstein, M. Kaste, 2007]. Она включает поддержание функций дыхания и кровообращения, коррекцию метаболических и волемических нарушений, контроль уровня артериального давления, коррекцию повышенного внутричерепного давления, профилактику и лечение таких состояний, как эпиприступы, венозные тромбозы, дисфагия, аспирационные пневмонии и другие инфекционные осложнения. Общепринятой практикой является активный мониторинг неврологического статуса и таких жизненно важных показателей, как артериальное давление, пульс, уровень глюкозы в крови и температура тела. Специфическая терапия Специфическая терапия включает в себя мероприятия направленные на реперфузию: тромболитическая терапия с использованием тканевого активатора плазминогена (ТАП), антитромбоцитарная терапия, хирургическая внутриартериальная реканализация, сюда также входят мероприятия, направленные на нейропротекцию. Главной целью специфической терапии инсульта является патогенетически обоснованный комплекс воздействий направленный на предотвращение роста зоны инфаркта мозга за счет предотвращения гибели зоны пенумбры. При этом мероприятия направленные на реперфузию восстанавливают кровоток в области очага ишемии, а целью нейропротекции является повышение устойчивости клеток к ишемии. Выделяют первичную нейропротекцию (мероприятия направленные на блокирование реакций глутаматной эксайтотоксичности и свободнорадикального окисления) и вторичную нейропротекцию (мероприятия направленные преимущественно на уменьшение отдаленных последствий ишемии, в основном за счет воздействия на воспаление и апоптоз) [Е.И. Гусев, В.И. Скворцова, 2001; М. Бэр, 2011]. 37 Наибольшую доказательную базу среди методов специфической терапии ИИ имеет тромболитическая терапия с использованием ТАП. Известно, что тромболитическая терапия с помощью ТАП вводимого в течение 3 часов от начала инсульта, значительно улучшает исход у пациентов с острым ИИ [The National Institute of Neurological Disorders and Stroke rt-PA Stroke Study Group, 1995; W.M. Clark et al., 2000; M.G. Lansberg et al., 2007]. Несмотря на замечательные результаты тромболитическая терапия доступна незначительному числу пациентов, в связи с тем фактом, что отступление от достаточно строгих критериев отбора пациентов сопровождается высоким риском осложнений в виде внутричерепных кровоизлияний после использования ТАП [M.G. Lansberg et al., 2007]. Несмотря на большую перспективность различных методов нейропротективной терапии в эксперименте, крупных рандомизированных многоцентровых клинических исследований продемонстрировавших достоверное улучшение исхода заболевания при ее использовании не существует [J. Rother, 2008]. Многие препараты, показавшие высокую эффективность в эксперименте, зачастую обладают выраженными побочными эффектами, которые делают их неприменимыми в клинической практике. Поэтому разработка новых подходов к терапии и профилактике инсульта является важнейшей задачей медицины. 1.8. Понятие о транскраниальной электростимуляции Транскраниальная электростимуляция (ТЭС-терапия) представляет собой воздействие электрическим током на мозг через покровы черепа, приводящее к селективной активации структур, относящихся к антиноцицептивной системе (АНС) ствола мозга [В.П. Лебедев, А.Б. Савченко и соавт., 1988]. На данный момент проведена и продолжает проводиться огромная работа по изучению эффектов ТЭС-терапии в различных областях медицины. Основные эффекты ТЭС-терапии, обусловлены усиленной продукцией и выделением β-эндорфинов (рис. 1.1). 38 Рисунок 1.1 — Основные эффекты ТЭС-терапи Все эффекты ТЭС-терапии по механизму условно делят на центральные, связанные с действием непосредственно на структуры мозга, и периферические — с воздействием веществ, поступивших в системный кровоток. 1.8.1. Центральные эффекты ТЭС-терапии В Институте физиологии им. И.П. Павлова АН СССР и ВНИИ пульмонологии МЗ СССР В.П. Лебедевым и соавт. (1983) был разработан и экспериментально обоснован оптимальный режим ТЭС: частота непрерывных импульсов или пачек высокочастотных импульсов 77 ± 0,5 Гц, длительность импульса 3,75 ± 0,25 мс при соотношении постоянного и среднеимпульсного тока 2 : 1 – 5 : 1 [Я.С. Кацнельсон, 1985]. Указанные параметры тока являются строго критичными. При изменении частоты или длительности импульса, а также соотношения между величинами постоянного и импульсного тока анальгетический эффект (АЭ) резко ослабевает. Расположение электродов при ТЭС — фронто-мастоидальное. Проводится ТЭС при помощи электростимуляторов «ТРАНСАИР» и «ЭТРАНС» различных модификаций. В ауторадиографических экспериментах с помощью [14С] деоксиглюкозы, которая накапливается в нервных клетках в соответствии с уровнем их 39 возбуждения, было установлено, что ТЭС приводит к селективной активации структур, относящихся к антиноцицептивной системе (АНС) ствола мозга (дорсомедиальные ядро гипоталамуса, серое околоводопроводное вещество, ядра шва) [В.П. Лебедев и соавт., 1986], при этом происходит блокада проведения восходящих болевых импульсов в кору головного мозга. При исследовании конкурентного взаимодействия эндогенных опиоидов, выделяющихся на фоне ТЭС, наиболее выраженное вытеснение меченого лиганда-µопиоидных рецепторов (2-125-I-морфина) было отмечено именно в структурах АНС [Л.Н. Айрапетов и соавт., 1987]. При исследовании внутричерепных путей распространения тока с помощью ядерно-магнитнорезонансной томографии было установлено, что только при расположении электродов, обеспечивающим сагитальное направление тока, последний имеет доступ к основным элементам АНС [M.L.G. Joy et al., 1993; M.L.G. Joy et al., 1994]. При проведении ТЭС — терапии в активации структур АНС [В.П. Лебедев и соавт., 1983] ведущая роль принадлежит эндогенным опиатным механизмам [А.Б. Савченко, 1994]. В более поздних публикациях выявлено участие серотонинергического, дофаминергического и холинергического механизмов в реализации стимуляционной анальгезии [В.П. Лебедев и соавт., 1991; А.Б. Савченко, 1994]. Опиоидная природа медиаторов, выделяемых при ТЭС подтверждена многочисленными экспериментальными и клиническими наблюдениями [В.П. Лебедев и соавт., 1986]. Показано, что во время и после сеанса ТЭС-терапии в мозге и спинномозговой жидкости увеличивается уровень ключевого опиоидного пептида — β-эндорфина [Л.И. Айрапетов и соавт., 1985; В.П. Лебедев и соавт., 1986]. Подобные эффекты наблюдаются также при непосредственной электростимуляции структур ствола мозга, относящихся к АНС [Е.О. Брагин, 1991]. Наряду с увеличением содержания опиоидных пептидов в структурах мозга и ликворе животных, на фоне ТЭС-терапии в режиме анальгезии отмечается, определяемое радиоиммунным методом, резкое увеличение 40 β-эндорфина в плазме крови экспериментальных животных (кролики, мыши, крысы, кошки) и человека [А.Б. Савченко, 1994]. Устранение обезболивания после введения налоксона — блокатора µ-опиоидных рецепторов, свидетельствует об участии опиоидного нейрохимического механизма в формировании АЭ при ТЭС [В.П. Лебедев и соавт., 1988]. У животных с толерантностью к морфину АЭ отсутствует и появляется по мере ее исчезновения [В.П. Лебедев и соавт., 1988]. У лиц с пониженной чувствительностью к опиатам, обусловленной их периодическим применением или врожденными особенностями организма, АЭ также не наблюдается [В.П. Лебедев и соавт., 1986]. Д-лейцин и Д-фенилаланин — ингибиторы энкефалиназ (карбоксипептидаза А и др.) — ферментов, осуществляющих деградацию энкефалинов, потенцируют АЭ [В.П. Лебедев и соавт., 1988], что также свидетельствует об участии опиоидных пептидов в эффектах стимуляции АНС. К центральным эффектам ТЭС-терапии помимо АЭ, относят центральный гемодинамический эффект. Экспериментальные исследования показали, что ТЭС в том же узком диапазоне характеристик, который необходим для развития АЭ, эффективно стабилизирует гемодинамику [В.П. Лебедев и соавт., 1986; Я.С. Кацнельсон, В.А. Леоско, 1987]. ТЭС-терапия широко используется для купирования болевых синдромов различной этиологии (висцеральных, спондилогенных, неврогенных, мигренозных). 1.8.2. Периферические эффекты ТЭС-терапии Основой исследований периферических эффектов ТЭС-терапии, которые в значительной степени опосредованы иммунными механизмами, послужило открытие ОР на разных типах клеток иммунной системы, что подчеркивает важность их роли в процессах иммунорегуляции [I.H. Jonsdottir, 2000; J.M. Bidlack, 2000; H. Machelska, K. Stein, 2003; H.L. Rittner, A. Brack, C. Stein, 2008; Т.А. Баева, 2006; И.Л. Шаравьева, 2011; Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011; Т.А. Баева, В.О. Небогатиков, 2012; К.Г. Горшкова, 2012; J.B. S. Garsia, M.G. de Melo Cardoso, M.C. Dos-Santos, 2012]. 41 Модуляция ТЭС динамики острофазного ответа в условиях асептического воспалительного процесса в значительной степени обусловлено указанной способностью опиоидных пептидов [Н.Л. Грицкевич и соавт., 1987; Н.Л. Грицкевич и соавт.,1991; Н.Л. Грицкевич и соавт.,1993]. Способность ТЭС-терапии в режиме анальгезии вызывать противовоспалительный эффект используется при использовании ТЭС-терапии в комплексной терапии воспалительных заболеваний в различных системах организма [А.П. Голиков и соавт., 1989; В.А. Александрова и соавт., 1994; В.С. Тиликин и соавт., 2012]. Изучение влияния ОП, высвобождающихся при ТЭС-терапии, на процесс репарации тканей и регенераторные процессы нервных стволов, в работах с экспериментальной перерезкой седалищного нерва у крыс, показано, что ТЭС-терапия ускоряет процесс репарации тканевых дефектов в среднем на 17–20 % [В.П. Лебедев и соавт., 1986; А.Б. Савченко 1994], реиннервацию двигательных нервных волокон на 30 %, чувствительных — на 20 % [Г.Н. Акоев и соавт., 1990; Л.И. Колосова и соавт., 1990]. Благодаря этому ТЭС-терапия используется в комплексной терапии нейросенсорной тугоухости [А.С. Роземблюм и соавт., 1998; Н.И. Краева и соавт., 1998], язвенной болезни желудка и двенадцатиперстной кишки [Ю.Д. Зильбер и соавт., 1987; Ю.Д. Зильбер и соавт., 1988; С.В. Рычкова, В.А. Александрова, 1991; С.В. Рычкова 1992; В.А. Александрова и соавт., 1994; Р.В. Шапоренко, 2008]; инфаркта миокарда [А.П. Голиков и соавт., 1989; Е.А. Губарева, 2009; В.Г. Борисенко, 2009], гестоза [С.П. Вчерашнюк, 2011]. ТЭС-терапия применяется для лечения ожогов и операционных травм. По данным кафедры общей и клинической патофизиологии Кубанского государственного медицинского университета благодаря использованию ТЭС-терапии сокращалось время заживления ожогов («легкие» на 25,8 %, «тяжелые» на 16,8 %) и операционной травмы на 18,7 %, что позволяло сокращать сроки пребывания больных в стационаре [Ю.А. Богданова, 2003]. ТЭС-терапия оказывает гомеостатическое действие на уровень артериального давления [В.П. Лебедев и соавт., 1992; Н.В. Зюзина, 2006]. 42 Показано что, под влиянием ТЭС-терапии отмечается тенденция к нормализации артериального давления [В.А. Заболотных, И.И. Заболотных, 1998]. Этот эффект ТЭС-терапии особенно выражен при вегетососудистых дистониях, как эссенциальных, так и климактерических [Г.А. Акимов и соавт., 1991]. Приоритетность и новизна такого способа лечения подтверждена авторским свидетельством СССР [В.А. Заболотных и соавт., 1987]. ТЭС-терапия оказывает положительные иммуномодулирующие эффекты [А.В. Рубцовенко, 1996], в том числе при вторичной иммунодепрессии, вызванной термической травмой [Ю.А. Богданова и соавт., 2000; Ю.А. Богданова и соавт., 2002; Ю.А. Богданова и соавт.,2003]. ТЭС-терапия оказывает положительный эффект при лечении заболеваний в патогенезе которых существенное значение имеют аллергические реакции [И.И. Заболотных и соавт., 1998]. Основными отличительными особенностями эффектов ТЭС-терапии являются. 1. Проявляются комплексно и одновременно. 2. Имеют гомеостатическую направленность. 3. Носят многокомпонентный системный характер. Нам известно, что в ходе течения ИИ содержание β-эндорфина претерпевает значительные изменения [М.А. Барабанова, 2003; F. Nyberg, 2004; Н.Н. Волосникова, 2004]. Важность данного факта подчеркивается тем, что в настоящее время ОП рассматриваются как важнейшие посредники между нервной и иммунной системами организма [О.Б. Сеин и соавт., 2009; Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011; В.А. Дубынин, А.А. Каменский, 2010; И.Л. Шаравьева, 2011; К.Г. Горшкова, 2012]. β-эндорфин важнейший представитель среди семейства ОП является агонистом µ и δ классических опиоидных рецепторов [В.А. Дубынин, А.А. Каменский, 2010; Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011; И.Л. Шаравьева, 2011; О.В. Нечайкина и соавт., 2013]. 43 β-эндорфин действует через ОР (µ, δ) на клетки иммунной системы, в целом оказывая ингибирующее влияние [M.J. Finley. et al., 2008; C. Borner et al., 2009; S. Boehncke et al., 2010]. По данным литературы широко известно, что формирование очага инфаркта мозга в ходе ИИ продолжается в течение 72 часов с момента развития острой локальной церебральной ишемии [S.H. Graham, J. Chen, 2001; M.P. Mattson et al., 2001; N.W. Manning et al., 2014]. В основе данного процесса лежит развитие острого асептического воспаления [K.J. Becker, 2001; G.J. del Zoppo, K.J. Becker, J.M. Hallenbeck, 2001; M.D. Ginsberg, 2003; M. Fisher, R. Ratan, 2003; P.L. Wood, 2003; R. Markus, D.C. Reutens, S. Kazui, 2004; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; Q. Wang, X.N. Tang, M.A. Yenari, 2007; А.Х. Хана-Мурад, Л.И. Павлинова, А.А. Мокрушин, 2007; A. Tuttolomondo, D. Di Raimondo, R. Di Sciacca, 2008; J. Jordan et al., 2008; D. Amantea et al., 2008; C. Benakis, L. Hirt, R.A. Du Pasquier, 2009; Ю.Д. Губарев, А.О. Шеремет, 2009; А.С. Кость, Б.Д. Луцик, Л.Е. Лаповець, 2010; A.G. Ceulemans et al., 2010; Y. Lee et al., 2014; М.В. Онуфриев и соавт., 2014]. Огромную важность представляют данные показывающие нейропротекторный, противовоспалительный, антиоксидантный, нейротрофический, антигипоксический эффекты активации δ-опиоидных рецепторов [G.J. Gross, 2000; Н.В. Соленкова, 2002; Е.И. Барзах, 2006; H. Zhang et al., 2006; M. Iwata et al., 2007; D.S. Su et al., 2007; A.K. Rehni, N. Singh, S.A. Jaggi, 2007; C. Happel et al., 2008; M. Govindaswami et al., 2008; Y. Yang et al., 2009; S. Husain, D.E. Potter, C.E. Crosson, 2009; M.R. Hutchinson et al., 2011; Y. Feng et al., 2011; S. Husain, G.I. Liou, C.E. Crosson, 2011; N. Akhter et al., 2013; M. Staples et al., 2013; J.M. Gidday, 2006; К.П. Иванов, 2013]. Таким образом, мы предполагаем значительное участие опиоидэргической системы в формировании очага инфаркта мозга при ИИ. Предполагается позитивный эффект воздействия β-эндорфина на течение экспериментальной модели ИИ. Однако, учитывая сложность синтеза в искусственных условиях, крайнюю нестойкость пептидов в организме (слишком быстрый распад), трудности по- 44 ступления через ГЭБ [K.A. Witt, T.P. Davis, 2006; R.K. Upadhyay, 2014], становится рациональным использование ТЭС-терапии как стимулятора образования эндогенного β-эндорфина [В.П. Лебедев и соавт. 1983; Л.И. Айрапетов и соавт., 1985; В.П. Лебедев и соавт., 1986; А.Б. Савченко, 1994] при ИИ. 45 ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ 2.1. Характеристика групп животных Эксперименты проведены на 140 взрослых белых нелинейных крысахсамцах средней массой 200 ± 25 гр. Возраст животных колебался от 8 месяцев до 1 года. У крыс моделировали ИИ. У них производили забор крови, тканей ГМ. Содержание животных и постановка экспериментов проводилась в соответствии с требованиями приказов № 1179 МЗ СССР от 11.10.1983 года и № 267 МЗ РФ от 19.06.2003 года, а также международными правилами «Guide for the Care and Use of Laboratory Animals» [Р. А. Когаладзе, 2004; Y. Wang-Fischer, 2009]. Животные были разделены на 4 группы (табл. 2.1). Таблица 2.1 Количественная характеристика исследования № группы 1 группа 2 группа Название группы Контрольная группа Крысы с моделированием ишемического инсульта Крысы с проведением ТЭС-терапии и последующим 3 группа моделированием ИИ Крысы с моделированием ишемического инсульта и по4 группа следующим проведением ТЭС-терапии Всего животных Количество животных (n) 20 40 40 40 140 1 группа (n = 20) — контрольная — интактные животные подвергались эвтаназии без оперативного вмешательства. 2 группа (n = 40) — животные с моделированием ИИ. Выведение животных из эксперимента производили по 10 животных на 1, 3, 7 и 14 сутки после моделирования ИИ. 3 группа (n = 40) — животные с моделированием ИИ, с проведением ТЭСтерапии: 7 сеансов до моделирования ИИ (n = 40). Выведение животных из эксперимента производили по 10 животных на 1, 3, 7 и 14 сутки после операции. 46 4 группа (n = 40) — животные с моделированием ИИ, с проведением ТЭС-терапии в течение 7 дней после моделирования ИИ. Выведение животных из эксперимента производили по 10 животных на 1, 3, 7 и 14 сутки после операции. 2.2. Методы исследования 2.2.1. Методика наркоза Все потенциально болезненные вмешательства в проводимых экспериментах (процедуры по созданию модели ИИ, проведение ТЭС-терапии), а также эвтаназия осуществлялись под общим наркозом. Эксперименты над крысами производились под комбинированным инъекционным наркозом [А.И. Трофименко и соавт., 2013]: золетил 0,3 мг в/м («Virbac» Франция), ксиланит 0,8 мг в/м (ЗАО «НИТА-ФАРМ, Россия, г. Саратов), атропина сульфат 0,1 % раствор — 0.01 мл п/к из расчета на 100 гр массы тела животного. Наркоз верифицировали по угнетению роговичного рефлекса и исчезновению реакции на болевые раздражители (укол лапы). 2.2.2. Методика моделирования ишемического инсульта у крыс Моделирование ИИ осуществляли путем создания у крыс острой локальной церебральной ишемии за счет коагуляции правой средней мозговой артерии (ПСМА) [Y. Wang-Fischer, 2009; А.И. Трофименко и соавт., 2013]. Моделирование острой локальной ишемии мозга у крыс Для проведения операции использовали налобную лупу с увеличением 4,8 крат. В области операционного поля кожу выбривали и обрабатывали спиртом и спиртовым раствором йода. Разрез кожи производили по ходу скуловой кости (около 1,5 см) (рис. 2.1, а). Обнажали слюнную железу в задненижнем квадрате операционного поля (рис. 2.1, б). Затем ее вместе с сосудистым сплетением отделяли от окружающих тканей и перемещали в задневерхний квадрат операционного поля (рис. 2.1, в). 47 Рисунок 2.1 — Этапы операции: а) обработка операционного поля и разрез кожи; б) доступ к слюнной железе; в) препаровка слюнной железы После удаления части скуловой кости обнажали края прикрепления височной мышцы к нижней челюсти и разрезали ее вдоль волокон (рис. 2.2, а). Разводили края височной мышцы и обнажали височную ямку, дно ямки образует крыловидная мышца с проходящим рядом нижнечелюстным нервом. Крыловидную мышцу раздвигали и обнажали поверхность черепа между овальным отверстием и отверстием зрительного нерва (рис. 2.2, б). Здесь сверлили отверстие диаметром около 3 мм и открывали место расположения ПСМА. ПСМА коагулировали электрокоагулятором собственного изготовления до прекращения тока крови. Прекращение кровотока по ПСМА контролировали визуально: побледнение артерии дистальнее места коагуляции и прекращение кровотечения при повреждении артерии дистальнее места коагуляции. Рисунок 2.2 — Моделирование локальной ишемии: а) удаление скуловой кости; б) доступ к поверхности черепа 48 После моделирования церебральной ишемии, восстанавливали топографию мышц и мягких тканей, пришивали сухожилия височной мышцы к нижней челюсти для возможности процесса жевания в последующем (рис. 2.3). \ Рисунок 2.3 — Завершающий этап операции В постоперационный период производили антибиотикотерапию (бициллин-5 в дозе 30 тысяч Ед на 100 гр. веса животных). 2.2.3. Методика проведения ТЭС-терапии у крыс ТЭС-терапию экспериментальным животным из групп № 3 и № 4 осуществляли модифицированным двухпрограммным электростимулятором «ТРАНСАИР-03» (ООО «Центр транскраниальной электростимуляции», г. Санкт-Петербург) в анальгетическом режиме (рис. 2.4). Рисунок 2.4 — Аппарат транскраниальной электростимуляции «Трансаир-03» 49 Параметры ТЭС-терапии для крыс представлены в таблице 2 [В.П. Лебедев, А.Б. Савченко, Н.В. Петряевская, 1988; В.П. Лебедев, А.Б. Савченко, А.В. Красюков, 1991; V.P. Lebedev et al., 2001]. Таблица 2.2 Параметры электрического тока, использующегося при проведении ТЭС-терапии в режиме анальгезии у крыс Частота, Гц Длительность импульса, мс Величина суммарного тока,мА 70 ± 2 3,75 ± 0,25 0,6–2,5 Подкожные игольчатые электроды располагали фронто-мастоидально (катод — в области лба над глазницами, анод — позади ушных раковин). Продолжительность первого сеанса составляла 15 минут, всех последующих по 30 минут. В группе № 3 7 сеансов ТЭС-терапии были проведены до моделирования ИИ. В группе № 4 ТЭС-терапия производилась после моделирования ИИ, в течение 7 дней. 2.2.4. Методика эвтаназии у крыс У крыс изымались кормушки с кормом за 12 часов до проведения эвтаназии. Потребление воды оставалось свободным. Эвтаназию крыс выполняли с соблюдением правил проведения работ при использовании экспериментальных животных, путем декапитации под стандартным наркозом [Y. WangFischer, 2009; Р.А. Когаладзе, 2004]. Во время эвтаназии проводили забор крови у крыс. Смерть животного констатировали по отсутствию дыхания и сердечной деятельности. После наступления летального исхода для последующего гистологического исследования проводили изъятие ГМ. 2.2.5. Методика забора крови Под наркозом проводили выделение общей яремной вены. При сборе крови, с целью предотвращения ее свертывания, на 1 мл крови добавляли 50 1000 ЕД гепарина. Кровь, в объеме 3 мл, помещали в пластиковые центрифужные пробирки с пробкой. Затем ее центрифугировали на центрифуге ОП-12 (Россия) в течение 15 минут со скоростью 1500 об/мин. Супернатант собирали пастеровской пипеткой и замораживали в криопробирках. Хранили в холодильнике при –70 оC. 2.2.6. Методика иммуноферментного анализа β-эндорфина в плазме крови крыс Иммуноферментный анализ (ИФА) проводили на базе ЦНИЛ отдел клинической экспериментальной иммунологии и молекулярной биологии ГБОУ ВПО КубГМУ Минздрава России. Определение уровня β-эндорфина в плазме крови количественно проводили иммуноферментным методом с помощью набора «Elabscience Biotechnology Co., Ltd», (Китай). Набор представлял собой комплект, содержащий реагент — моноклональные антитела к β-эндорфину. Они были сорбированы на поверхности лунок полистирольного планшета и содержали конъюгаты поликлональных антител к β-эндорфину с биотином и калибровочные образцы с β-эндорфином. На первой стадии анализа, исследуемые и контрольные образцы, инкубировали в лунках с иммобилизованными антителами. β-эндорфин, имеющийся в образцах, связывался с иммобилизованными антителами. Материал, который не связался, удаляли буферным раствором из набора. Основными компонентами раствора являлся фосфатно-солевой буфер с детергентом. β-эндорфин, который связался, взаимодействовал с конъюгатом № 1 (антитела к β-эндорфину с биотином). Конъюгат № 1, который не связался, удаляли отмывкой. Связавшийся конъюгат № 1 на третьей стадии взаимодействовал с конъюгатом № 2 (стрептавидин с пероксидазой хрена). Количество связавшегося конъюгата № 2, после третьей отмывки, определяли цветной реакцией с использованием субстрата пероксидазы хрена — перекиси водорода и хромогена — тетраметилбензидина. Протекающую реакцию останавливали добавлением раствора стоп-реагента — 0,1 N HCl и измеряли оптическую плотность растворов в лунках при длине вол- 51 ны 450 нм. Интенсивность окрашивания раствора в лунке была пропорциональна количеству содержащегося в образце β-эндорфина. Все этапы реакции проходили в термостатируемых условиях на шейкерах-инкубаторах «ST-3» (Латвия). Учет реакции, построение калибровочных графиков и определение концентрации β-эндорфина проводили на фотометре вертикального сканирования «ANTHOS 2010» (Австрия) с помощью программного обеспечения «Auswerte-Softwere anthos labtec», версия 2.3.0.7. 2.2.7. Методика иммуноферментного анализа интерлейкина-1β в плазме крови крыс Определение уровня ИЛ-1β в плазме крови проводили иммуноферментным методом с помощью набора «Ray Biotech, Inc.» (Германия). Основным реагентом набора являются моноклональные антитела к ИЛ-1β, сорбированные на поверхности лунок полистирольного планшета, конъюгаты поликлональных антител к ИЛ-1β с биотином и калибровочные образцы, содержащие ИЛ-1β. Исследуемые и контрольные образцы на первой стадии анализа, инкубировали в лунках с иммобилизованными антителами. ИЛ-1β, который имелся в образцах связывался с иммобилизованными антителами. Материал, который не связался отмывали буферным раствором из набора, основным компонентом которого является фосфатно-солевой буфер с детергентом. ИЛ-1β, который связался, взаимодействовал с конъюгатом № 1 (антитела к ИЛ-1β с биотином). Конъюгат № 1, который не связался удаляли отмывкой. Конъюгат № 1, который связался, на третьей стадии взаимодействовал при инкубации с конъюгатом № 2 (стрептавидин с пероксидазой хрена). Количество связавшегося конъюгата № 2, после третьей отмывки, определяли цветной реакцией с субстратом пероксидазы хрена — перекиси водорода и хромогена — тетраметилбензидина. Добавлением раствора стоп-реагента — 0,1 N HCl реакцию останавливали и оптическую плотность растворов в лунках измеряли при длине волны 450 нм. Количество содержащегося в образце ИЛ-1β было пропорционально интенсивности окрашивания раствора в лунке. Все этапы реакции проходили в термостатируемых условиях на шейкерах-инкубаторах «ST-3» (Латвия), учет реакции, построение 52 калибровочных графиков и определение концентрации анализов проводили на фотометре вертикального сканирования «ANTHOS 2010» (Австрия) с помощью программного обеспечения «Auswerte-Softwere anthos labtec», версия 2.3.0.7. 2.2.8. Методика иммуноферментного анализа интерлейкина-6 в плазме крови крыс Определение уровня ИЛ-6 в плазме крови проводили иммуноферментным методом с помощью набора «Cusabio Biotech Co., Ltd» (Китай). Основным реагентом комплекта являются моноклональные антитела к ИЛ-6, сорбированные на поверхности лунок полистирольного планшета, конъюгаты поликлональных антител к ИЛ-6 с биотином и калибровочные образцы, содержащие ИЛ-6. Исследуемые и контрольные образцы, на первой стадии анализа, инкубировали в лунках с иммобилизованными антителами. ИЛ-6, который имелся в образцах, связывался с иммобилизованными антителами. Отмывкой буферным раствором из набора удаляли не связавшийся материал. Основным компонентом раствора являлся фосфатно-солевой буфер с детергентом. С конъюгатом № 1 (антитела к ИЛ-6 с биотином) взаимодействовал связавшийся ИЛ-6. Конъюгат № 1, который не связался удаляли отмывкой. Связавшийся конъюгат № 1 на третьей стадии взаимодействовал при инкубации с конъюгатом № 2 (стрептавидин с пероксидазой хрена). Количество связавшегося конъюгата № 2, после третьей отмывки, определяли цветной реакцией с субстратом пероксидазы хрена — перекиси водорода и хромогена — тетраметилбензидина. Для остановки реакции добавляли раствора стоп-реагента — 0,1 N HCl. Оптическую плотность растворов в лунках измеряли при длине волны 450 нм. Количеству ИЛ-6, содержащегося в образце было пропорционально интенсивности окрашивания раствора в лунке. Все этапы реакции проходили на шейкерах-инкубаторах «ST-3» (Латвия) в термостатируемых условиях. Учет реакции, построение калибровочных графиков и определение концентрации анализов проводили на фотометре вертикального сканирования «ANTHOS 2010» (Австрия) с помощью программного обеспечения «Auswerte-Softwere anthos labtec», версия 2.3.0.7. 53 2.2.9. Методика иммуноферментного анализа фактора некроза опухоли-α в плазме крови крыс Определение уровня ФНО-α в плазме крови проводили иммуноферментным методом с помощью набора «Ray Biotech, Inc.», (Германия). Основным реагентом комплекта являются моноклональные антитела к ФНО-α, сорбированные на поверхности лунок полистирольного планшета, конъюгаты поликлональных антител к ФНО-α с биотином и калибровочные образцы, содержащие ФНО-α. Исследуемые и контрольные образцы на первой стадии анализа инкубировали в лунках с иммобилизованными антителами. ФНО-α, имеющийся в образцах, связывался с иммобилизованными антителами. Материал, который не связался удаляли отмывкой буферным раствором, основным компонентом которого является фосфатно-солевой буфер с детергентом. Конъюгат № 1 (антитела к ФНО-α с биотином) взаимодействовал со связавшимся ФНО-α. Не связавшийся конъюгат № 1 удаляли отмывкой. Связавшийся конъюгат № 1 на третьей стадии при инкубации взаимодействовал с конъюгатом № 2 (стрептавидин с пероксидазой хрена). Количество связавшегося конъюгата № 2 после третьей отмывки определяли цветной реакцией с субстратом пероксидазы хрена — перекиси водорода и хромогена — тетраметилбензидина. Добавлением раствора стопреагента — 0,1 N HCl реакцию останавливали и измеряли оптическую плотность растворов в лунках при длине волны 450 нм. Количество, содержащегося в образце ФНО-α, было пропорционально интенсивности окрашивания раствора в лунке. Все этапы реакции проходили на шейкерах-инкубаторах «ST-3» (Латвия), в термостатируемых условиях. Учет реакции, построение калибровочных графиков и определение концентрации анализов проводили на фотометре вертикального сканирования «ANTHOS 2010» (Австрия) с помощью программного обеспечения «Auswerte-Softwere anthos labtec», версия 2.3.0.7. 2.2.10. Методика иммуноферментного анализа интерлейкина-10 в плазме крови крыс Определение уровня ИЛ-10 в плазме крови проводили иммуноферментным методом с помощью набора «Ray Biotech, Inc.» (Германия). Основным реаген- 54 том комплекта являются моноклональные антитела к ИЛ-10, сорбированные на поверхности лунок полистирольного планшета, конъюгаты поликлональных антител к ИЛ-10 с биотином и калибровочные образцы, содержащие ИЛ-10. Исследуемые и контрольные образцы, на первой стадии анализа, инкубировали в лунках с иммобилизованными антителами. ИЛ-10, имеющийся в образцах, связывался с иммобилизованными антителами. Материал, который не связался удаляли отмывкой буферным раствором, содержащим фосфатно-солевой буфер с детергентом. С конъюгатом № 1 (антитела к ИЛ-10 с биотином) взаимодействовал связавшийся ИЛ-10. Конъюгат № 1, который не связался удаляли отмывкой. Связавшийся конъюгат № 1 на третьей стадии взаимодействовал при инкубации с конъюгатом № 2 (стрептавидин с пероксидазой хрена). Количество связавшегося конъюгата № 2 после третьей отмывки определяли цветной реакцией с использованием субстрата пероксидазы хрена — перекиси водорода и хромогена — тетраметилбензидина. Добавлением раствора стоп-реагента — 0,1 N HCl реакцию останавливали и измеряли оптическую плотность растворов в лунках при длине волны 450 нм. Количество содержащегося в образце ИЛ-10 было пропорционально интенсивности окрашивания раствора в лунке. Все этапы реакции проходили на шейкерах-инкубаторах «ST-3» (Латвия) в термостатируемых условиях. Учет реакции, построение калибровочных графиков и определение концентрации анализов проводили на фотометре вертикального сканирования «ANTHOS 2010» (Австрия) с помощью программного обеспечения «Auswerte-Softwere anthos labtec», версия 2.3.0.7. 2.2.11. Методика иммуноферментного анализа трансформирующего фактора роста-1β в плазме крови крыс Определение уровня ТФР-1β в плазме крови проводили иммуноферментным методом с помощью набора «Boster Biological Technology Co., Inc.» (США). Основным реагентом комплекта являются моноклональные антитела к ТФР-1β, сорбированные на поверхности лунок полистирольного планшета, конъюгаты поликлональных антител к ТФР-1β с биотином и калибровочные образцы, содержащие ТФР-1β. Исследуемые и контрольные образцы на первой стадии ана- 55 лиза инкубировали в лунках с иммобилизованными антителами. ТФР-1β, имеющийся в образцах, связывался с иммобилизованными антителами. Материал, который не связался удаляли отмывкой буферным раствором, содержащим фосфатно-солевой буфер с детергентом. С конъюгатом № 1 (антитела к ТФР-1β с биотином) взаимодействовал связавшийся ТФР-1β. Конъюгат № 1, который не связался, удаляли отмывкой. Связавшийся конъюгат № 1 на третьей стадии взаимодействовал при инкубации с конъюгатом № 2 (стрептавидин с пероксидазой хрена). Количество связавшегося конъюгата № 2 после третьей отмывки определяли цветной реакцией с использованием субстрата пероксидазы хрена — перекиси водорода и хромогена — тетраметилбензидина. Добавлением раствора стоп-реагента — 0,1 N HCl реакцию останавливали и измеряли оптическую плотность растворов в лунках при длине волны 450 нм. Количество содержащегося в образце ТФР-1β было пропорционально интенсивности окрашивания раствора в лунке. Все этапы реакции проходили на шейкерах-инкубаторах «ST-3» (Латвия) в термостатируемых условиях. Учет реакции, построение калибровочных графиков и определение концентрации анализов проводили на фотометре вертикального сканирования «ANTHOS 2010» (Австрия) с помощью программного обеспечения «Auswerte-Softwere anthos labtec», версия 2.3.0.7. 2.2.12. Методики гистологических исследований Фиксация, проводка и заливка гистологического материала Из ГМ, взятого после эвтаназии крыс, (половина органа из каждой группы) на границе очага ишемии вырезались кусочки размером 3 мм. Их фиксировали в 10 % нейтральном забуференном фосфатами формалине в течение 2 суток. Затем в течение 2 часов фиксированную ткань ГМ выдерживали в проточной водопроводной воде. Ручную проводку забранного материала (ткани ГМ) выполняли с использованием изопропилового спирта с минеральным маслом до парафина [М.В. Пешков, И.И. Дыгало, 2009; Д.Э. Коржевский, А.В. Гиляров, 2010]. Заливку осуществляли парафином. После вырезки парафиновых блоков их устанавливали на деревянные блоки. 56 Протокол ручной гистологической проводки тканей с использованием изопропанола и минерального масла (время протокола 30 часов). 1. Изопропанол 60 минут — вылить после использования. 2. Изопропанол 60 минут — перелить в № 1 3. Изопропанол 60 минут — перелить в № 2 4. Изопропанол 60 минут — перелить в № 3 5. Изопропанол 60 минут — перелить в № 4 каждая партия (40 кассет) — свежий реагент 6. Изопропанол: минеральное масло 5 : 1, 50 оС — 90 минут, каждая 3-я партия свежий реагент. 7. Изопропанол: минеральное масло 2 : 1, 50 оС — 90 минут, каждая 3-я партия свежий реагент. 8. Минеральное масло, 50 оС — 120 минут (или до утра и на выходные) — каждая 3-я партия свежий реагент. 9. Парафин 60 оС — 60 минут вылить после 5 партий. 10. Парафин 60 оС — 60 минут перелить в № 9. 11. Парафин 60 оС — 60 минут перелить в № 10. 12. Парафин 60 оС — 60 минут каждая 5-я партия свежий реагент, использованный реагент перелить в № 11. Изготовление срезов и их наклейка на предметные стекла На микротоме МПС-2 (CCCP) парафиновые блоки нарезались на срезы толщиной 10 мкм. Срезы помещали влажным способом на предметные стекла с адгезивным покрытием. Стекла со срезами выдерживались в термостате при температуре 37 оС в течение 12 часов для расправления. Депарафинирование и регидратация срезов перед окраской ● Помещение стекол в о-ксилол на 5 минут. ● Помещение стекол в о-ксилол на 3 минуты. ● Помещение стекол в этиловый спирт 96о на 3 минуты. 57 ● Помещение стекол в этиловый спирт 96о на 2 минуты. ● Помещение стекол в этиловый спирт 70о на 2 минуты. ● Помещение стекол в этиловый спирт 50о на 2 минуты. ● Помещение стекол в дистиллированную воду на 5 минут. Протокол окраски срезов гематоксиллином и эозином Методика окраски гематоксилином и эозином по Д.Э. Коржевскому, А.В. Гилярову (2010). ● Депарафинирование и регидратация срезов. ● Окраска срезов в гематоксилине Эрлиха 2 минуты. ● Промывка стекол в дистиллированной воде 2 минуты. ● Промывка стекол в аммиачной воде 2 минуты. ● Промывка стекол в дистиллированной воде 5 минут. ● Окраска стекол водным раствором эозина 2 минуты. ● Промывка стекол в дистиллированной воде 30 секунд. ● Дегидратация, просветление и заключение срезов по протоколу. Методика окраски ткани головного мозга толуидиновым синим по Нисслю [Д.Э. Коржевский, А.В. Гиляров, 2010]. ● Депарафинирование и регидратация срезов. ● Окрашивание срезов на стеклах в 0,5 % водном растворе толуидинового синего в течение 15 минут. ● Промывка стекол в дистиллированной воде 2 минуты. ● Помещение стекол в этиловый спирт 96о на 2 минуты. ● Помещение стекол в этиловый спирт 96о на 2 минуты. ● Дифференцировка окрашивания под контролем микроскопа 96о уксуснокислым спиртом. ● Ополаскивание стекол в 96о этиловом спирте. ● Далее продолжить дегидратацию начиная с пункта 4. 58 Дегидратация, просветление и заключение срезов после окраски. ● Помещение стекол в этиловый спирт 50о на 2 минуты. ● Помещение стекол в этиловый спирт 70о на 2 минуты. ● Помещение стекол в этиловый спирт 96о на 2 минуты. ● Помещение стекол в этиловый спирт 96о на 3 минуты. ● Помещение стекол в о-ксилол на 3 минуты. ● Помещение стекол в о-ксилол на 5 минут. ● Подсушивание стекол и заключение срезов в кедровый бальзам. Методика окрашивания ткани мозга по Чиаччио [Р. Лилли, 1969]. ● Депарафинирование и регидратация срезов. ● Ополаскивание стекол в водном растворе 1 Н соляной кислоты. ● Выдержка стекол в водном растворе 1 Н соляной кислоты в термостате при 60 °С в течение 6 минут. ● Ополаскивание стекол в водном растворе 1 Н соляной кислоты. ● Окрашивание стекол в реактиве Шиффа в течение 10 минут (в темноте). ● Ополаскивание срезов в 3-х порциях 0,5 % водного раствора метабисульфита натрия в каждой по 2 минуты. ● Ополаскивание нескольких порциях дистиллированной воды в течение 5 минут. ● Помещение стекол в красящую смесь на 10 минут. Красящая смесь получена путем фильтрации 0,5 % раствора судана черного на изопропаноле и 0,5 % водного раствора буры. ● Ополаскивание стекол в дистиллированной воде 2 минуты. ● Заключение срезов в глицерин — желатину с последующей окантовкой покровных стекол даммар — лаком. Полученные стекла микроскопировали на микроскопе Микромед-6 (Россия). Фотографирование осуществляли с помощью окулярной камеры «Levenhuk 230» (США). 59 2.2.13. Методика определения размеров очага ишемии в головном мозге у крыс Размер очага церебральной ишемии определяли тетразолиевым методом [Y. Wang-Fischer, 2009; Д.Н. Силачев, 2009; А.И. Трофименко и соавт., 2013], с применением 2,3,5-трифенилтетразолия хлорида (2,3,5-ТТХ) (Россия). Тетразолиевый метод использовали, потому что он дает сравнимые данные объема очага повреждения по сравнению с МРТ в режиме Т2-ВИ. Результаты, полученные при этом коррелируют со степенью неврологического дефицита у животных с ишемией мозга в поведенческих тестах [Д.Н. Силачев, 2009]. 2,3,5-ТТХ является акцептором ионов водорода. Он присоединяет водород. Водород, проходя через всю систему переносчиков, соединяется с молекулярным кислородом. Ферментативное окисление субстрата приводит к образованию НАДН или НАДФН. 2,3,5-ТТХ на втором этапе реакции в сопряженной «индикаторной реакции» восстанавливается при участии НАДФН-дегидрогеназы до водонерастворимого формазана красного цвета. Затем снова НАДН или НАДФН окисляются. В месте локализации НАДФН-дегидрогеназы, как правило, в митохондриях происходит образование формазана. От количества образующегося формазана зависит интенсивность окраски окрашиваемой ткани, которая свидетельствует об активности фермента. У половины животных каждой группы проводили визуализацию с последующим определение размеров очага ишемии головного мозга тетразолиевым методом [Y. Wang-Fischer, 2009; Д.Н. Силачев, 2009; А.И. Трофименко и соавт., 2013]. Протокол визуализации очага ишемии ГМ тетразолиевым методом Быструю промывку ГМ от крови проводили сразу после его выделения двукратно в холодном 0,9 % растворе хлорида натрия с последующей его заморозкой и нарезкой во фронтальной плоскости на срезы по 2 мм. Срезы ткани мозга помещали в размеченную чашку Петри с фосфатным буфером с pH 7,2 объемом 10 мл, затем прибавляли 2 мл 2 % раствора 2,3,5-ТТХ и с 2 мл 2 % раствор сукцината натрия. Срезы выдерживали в указанной смеси в термостате при температуре 38 °С в течение 1 часа. 60 Срезы мозга подвергались фиксации в растворе 10 % нейтрального формалина в течение 1 часа. Затем срезы мозга сканировали на фотосканере и изображение сохраняли в натуральном масштабе в формате PDF. Площадь очага церебральной ишемии определяли на полученных сканах срезов в (мм2) с использованием программы AreaS (в основе метода лежит принцип сравнения площадей двух фигур, площадь одной из которых известна) [А.В. Пермяков и соавт., 2003]. Объем очага инфаркта мозга (в мм3) и объем мозга (в мм3) определяли на основе имеющихся данных: известная площадь срезов, известная площадь очага инфаркта мозга на срезе, число и толщина срезов [Г.Г. Автандилов, 1990; К.А. Дроздов и соавт., 2011]. Показатель «удельный объем инфаркта мозга» определяли как соотношение объема очага инфаркта мозга к объему всего мозга. Расчет указанного показателя позволял нивелировать разброс по объему мозга и соответственно объему очагов инфаркта у животных разной массы [Г.Г. Автандилов, 1990]. 2.3. Статистические методы обработки полученных данных Статистическую обработку полученных данных осуществляли методами непараметрической статистики с использованием программного обеспечения «Statistika 6.0 for Windows» фирмы «Stat Soft Inc.» [О. Ю. Реброва, 2006]. Полученные результаты исследуемых групп после статистической обработки выражали в виде средних значений (M) и ошибки среднего (m). Сравнение выборок проводилось по непараметрическому критерию Манна-Уитни, с установлением уровня значимости *p ≤ 0,05. Величины средних значений в таблицах указаны в границах M ± m. 61 ГЛАВА 3. ГИСТОЛОГИЧЕСКАЯ КАРТИНА ЗОНЫ ИШЕМИИ ПРИ МОДЕЛИРОВАНИИ ИШЕМИЧЕСКОГО ИНСУЛЬТА БЕЗ ПРОВЕДЕНИЯ И С ПРИМЕНЕНИЕМ ТЭС-ТЕРАПИИ 3.1. Гистологическая картина зоны ишемии при моделировании ишемического инсульта На микропрепаратах мозга интактных крыс группы № 1 (контрольная), окрашенных гематоксилин-ээозином, выявлено, что в коре теменной доли ГМ крыс в зоне кровоснабжения ПСМА можно выделить шесть слоев, представленных разными видами клеток (рис. 3.1). Рисунок 3.1 — Кора головного мозга интактной крысы. Ув. об. 40. Окраска гематоксилин-эозин I слой коры представлен мелкими клетками веретеновидной формы (рис. 3.1, поз. 1). II, III и IV слои коры имели общие черты морфологической организации, были представлены в основном зернистыми клетками и сливались между собой (рис. 3.1, поз. 2). V слой состоял из многочисленных крупных пирамидных клеток (рис. 3.1, поз. 3). VI слой был слабо различим, состоял из веретеновидных клеток и сливался с V слоем (рис. 3.1, поз. 3). 62 В ткани мозга обнаружено, что капилляры (рис. 3.2, поз. 1) окружены оптически пустыми пространствами умеренных размеров, в связи с наличием плотно прилегающих к стенке сосудов отростков астроцитов (рис. 3.2, поз. 2). На микропрепаратах ткани мозга интактных крыс группы № 1 (контрольная), окрашенных по Нисслю, выявлено, что в коре теменной доли головного мозга крыс в области кровоснабжения правой средней мозговой артерии отмечается преобладание нормохромных клеток (рис. 3.3, поз. 1). Они симметричной формы с равномерно распределенной хроматофильной субстанцией цитоплазмы и ядрами округлой формы (рис. 3.3, поз. 2). Рисунок 3.2 — Взаимоотношение астроцитов и капилляра. Ув. об. 100. Окраска гематоксилин-эозин Рисунок 3.3 — Нормохромные пирамидные нейроны. Ув. об. 40. Окраска по Нисслю Для выявления ДНК ядер клеток нейроглии в коре теменной доли головного мозга, в зоне кровоснабжения правой средней мозговой артерии применяли реакцию Фельгена (специфическая реакция фуксинсернистой кислоты с альдегидными группами дезоксирибозы, полученной путем мягкого кислотного гидролиза ДНК. Ее результатом является образование стойкого красителя красно-фиолетового цвета). При окраске по Чиаччио на микропрепаратах ткани мозга, имеет место четкое окрашивание ядер в красный цвет, полученное за счет протекания реакции Фельгена. Фон, из-за окрашивания липидов суданом черным, был равномерного серо-зеленоватого цвета (рис. 3.4). 63 Рисунок 3.4 — Ядра клеток нейроглии в ткани мозга. Ув. об. 10. Окраска по Чиаччио Рисунок 3.5 — Очаг инфаркта в ткани головного мозга. Крысы группы № 2. Ув. об. 40. Окраска гематоксилин-эозин При моделировании ИИ через 1 сутки, на микропрепаратах ткани мозга крыс группы № 2, окрашенных гематоксилин-эозином, начинает прослеживаться формирование области инфаркта мозга (рис. 3.5, поз. 1). Появляются клетки-тени (рис. 3.5, поз. 3), единичные нейтрофилы (рис. 3.5, поз. 4), признаки перицеллюлярного отека ткани мозга (рис. 3.5). На периферии очага инфаркта мозга отмечено развитие клеточного ответа на формирование очага некроза ткани мозга (рис. 3.5, поз. 2). В капиллярах в центре области ишемии выявляется запустевание, они становятся незаметными на общем фоне ткани (рис. 3.5, поз. 5). По периферии очага ишемии капилляры хорошо прокрашены (рис. 3.6), в них выявляются признаки отека эндотелиоцитов (рис. 3.6, поз. 1), стаза крови. Развивается адгезия нейтрофилов к эндотелию капилляров (рис. 3.6, поз. 2), в некоторых признаки сдавления сосудов отечными ножками астроцитов. Через 1 сутки после моделирования ИИ на микропрепаратах ткани мозга крыс группы № 2, окрашенных по Нисслю, отмечается появление гиперхромных дистрофически измененных нейронов (рис. 3.7, поз. 1), прослеживаются признаки инфильтрации ткани мозга нейтрофилами (рис. 3.7, поз. 2), единичные явления нейронофагии (рис. 3.7, поз. 3), практически полный тигролиз (признаки распада хроматофильной субстанции Ниссля в нейронах) (рис. 3.7, поз. 4). 64 Рисунок 3.6 — Периферия очага ишемии в мозге крысы группы № 2. Ув. об. 90. Окраска гематоксиллин-эозин Рисунок 3.7 — Ткань мозга крысы группы № 2. Ув. об.40. Окраска по Нисслю На микропрепаратах ткани мозга, окрашенных по Чиаччио, в те же сроки после моделирования ИИ, отмечается гипохромия ядер клеток в очаге ишемии (рис. 3.8, поз. 1), что возможно отражает текущие процессы деполимеризации ДНК в некротизированных клетках. Кроме того происходит повышение интенсивности восприимчивости окраски ткани по периферии очага ишемии к судану черному, что отражает процесс демиелинизации (рис. 3.8, поз. 2), также в очаге ишемии видна значительная разреженность нейропиля, что показывает выраженность перицеллюлярного отека (рис. 3.8). На 3 сутки после моделирования ИИ на микропрепаратах ткани мозга, окрашенных гематоксилин-эозином, четко определяется область инфаркта мозга (рис. 3.9). Имеет место значительная воспалительная клеточная Рисунок 3.8 — Срез мозга крысы группы № 2. Ув.10. об. Окраска по Чиаччио Рисунок 3.9 — Срез мозга крысы группы № 2. Ув. об. 40. Окраска гематоксилин-эозин 65 инфильтрация зоны инфаркта ткани мозга (рис. 3.9, поз. 1). Воспалительный клеточный инфильтрат в последующем послужит основой для формирования глиомезодермального рубца. Отмечено значительное увеличение численности погибших нейронов (множественные клетки-тени) (рис. 3.9, поз. 2), разреженность нейропиля, выраженный перицеллюлярный отек. На границе между зоной инфаркта в ткани мозга и интактной тканью отмечается скопление множества мелких круглых клеток, предположительно лейкоцитарного и лимфоцитарного происхождения. По периферии очага ишемии отчетливо выявляются сосуды, в них имеются признаки стаза крови (рис 3.10, поз. 1). Наблюдаются явления адгезии к эндотелию капилляров полиморфоядерных нейтрофилов и их миграция в ткань мозга (рис. 3.10, поз. 2). На 3 сутки после моделирования ИИ исследование микропрепаратов ткани мозга животных группы № 2, окрашенных по Нисслю, показало наличие небольшого числа дистрофически измененных нейронов (рис. 3.11, поз. 1), признаки нейронофагии (рис. 3.11, поз. 2) и выраженную воспалительную клеточную инфильтрацию (рис. 3.11, поз. 3). При исследовании на 3 сутки, после моделирования ИИ, микропрепаратов ткани мозга крыс группы № 2, окрашенных по Чиаччио, отчетливо выявляется зона деструкции ткани мозга (рис. 3.12, поз. 1). Отсутствие в ней ядер, окрашенных в красный цвет, указывает на деполимеризацию ДНК. На периферии зоны деструкции видна область воспалительной клеточной инфиль- Рисунок 3.10 — Срез мозга крысы группы № 2. Ув. об. 40. Окраска гематоксилин-эозин Рисунок 3.11 — Срез мозга крысы группы № 2. Ув. об. 40. Окраска по Нисслю 66 трации (рис. 3.12, поз. 2), которая служит основой формирования в последующем глиомезодермального рубца. На 7 сутки, после моделирования ИИ, на микропрепаратах ткани мозга животных группы № 2, окрашенных гематоксилин-эозином, отмечается скопление «зернистых шаров» в зоне деструкции и по ее границам (рис. 3.13, поз. 1). В этот период в зоне деструкции нейронов не обнаружено (рис. 3.13). Рисунок 3.12 — Срез мозга крысы группы № 2. Ув. об. 20. Окраска по Чиаччио Рисунок 3.13 — Срез мозга крысы группы № 2. Ув. об. 40. Окраска гематоксилин-эозин На микропрепаратах ткани мозга крыс группы № 2 на 7 сутки после моделирования ИИ, при окраске по Нисслю, видим, что нейроны в области инфаркта ткани мозга отсутствуют (рис. 3.14). Отмечено присутствие значительного количества воспалительного клеточного инфильтрата (рис. 3.14, поз. 1), в частности «зернистых шаров» (рис. 3.14, поз. 2). На микропрепаратах ткани мозга, окрашенных по Чиаччио, виден формирующийся незрелый глиомезодермальный рубец (рис. 3.15, поз. 1). Он расположен перпендикулярно направлению нахождения ядер олигодедроглиоцитов и нервных волокон в мозге (рис. 3.15, поз. 2). Отмечается выраженная клеточная реакция и разреженность нейропиля в области самого инфаркта мозга (рис. 3.16). На 14 сутки, после моделирования ИИ, на микропрепаратах ткани мозга крыс группы № 2, окрашенных гематоксилин-эозином, по периферии очага видны формирующиеся капилляры, прорастающие в зону деструкции 67 (рис. 3.17, поз. 1). Основу глиомезодермального рубца в очаге инфаркта мозга и по его границам составляет скопление клеток глии (рис. 3.17, поз. 2). Рисунок 3.14 — Срез мозга крысы группы № 2. Ув. об. 40. Окраска по Нисслю Рисунок 3.15 — Срез мозга крысы группы № 2. Ув. об. 20. Окраска по Чиаччио Рисунок 3.16 — Срез мозга крысы группы № 2. Ув. об. 20. Окраска по Чиаччио Рисунок 3.17 — Срез мозга крысы группы № 2. Ув. об. 40. Окраска гематоксилин-эозин На всех микропрепаратах, по сравнению с 7 сутками, несколько увеличено содержание «зернистых шаров». На микропрепаратах ткани мозга крыс группы № 2, окрашенных по Нисслю, на 14 сутки в области зоны деструкции виден формирующийся глиальный рубец (рис. 3.18). На микропрепаратах мозга, окрашенных по Чиаччио, видна происходящая организация зоны деструкции и формирующийся избыточный глиомезодермальный рубец (рис. 3.19). 68 Рисунок 3.18 — Срез мозга крысы группы № 2. Ув. об. 40. Окраска по Нисслю Рисунок 3.19 — Срез мозга крысы группы № 2. Ув. об. 10. Окраска по Чиаччио Таким образом, через 1 сутки после моделирования ИИ, начинает выявляться область формирующегося инфаркта мозга. Это сопровождается появлением клеток-теней, видны единичные Нф, появляются признаки перицеллюлярного отека ткани мозга. На периферии очага инфаркта мозга формируется клеточный ответ в связи с появлением очага некроза ткани мозга. В центре области ишемии капилляры запустевают. По периферии очага ишемии капилляры хорошо видны. В них выявляются признаки отека эндотелиоцитов и стаза крови. Развивается адгезия нейтрофилов к эндотелию капилляров. Видна инфильтрация ткани мозга Нф. При окраске по Нисслю появляются гиперхромные дистрофически измененные нейроны. В них произошел практически полный тигролиз. При окраске по Чиаччио, в те же сроки после моделирования ИИ, в очаге ишемии отмечается гипохромия ядер клеток, отражающая процессы деполимеризации ДНК в некротизированных клетках. Одновременно идет процесс демиелинизации. Наблюдаемая значительная разреженность нейропиля в очаге ишемии указывает на выраженность перицеллюлярного отека. На 3 сутки, после моделирования ИИ, на микропрепаратах ткани мозга, окрашенных по Чиаччио, отчетливо контурируется зона деструкции ткани мозга. Выявляется значительная воспалительная клеточная инфильтрация зо- 69 ны инфаркта ткани мозга. На границе между зоной инфаркта в ткани мозга и интактной тканью скапливается много мелких круглых клеток, предположительно лейкоцитарного и лимфоцитарного происхождения. В последующем воспалительный клеточный инфильтрат послужит основой формирования глиомезодермального рубца. По периферии очага ишемии отчетливо видны сосуды. В них проявляются признаки стаза крови, а также явления адгезии полиморфоядерных нейтрофилов к эндотелию капилляров и миграции их в ткань мозга. Усиливается разреженность нейропиля, а значит и выраженность перицеллюлярного отека. Появляются множественные клетки-тени (погибшие нейроны). На микропрепаратах ткани мозга, окрашенных по Нисслю, выявляются дистрофически измененные нейроны. Отсутствие ядер (окраска по Чиаччио) окрашенных в красный цвет, свидетельствует о деполимеризации в них ДНК. На 7 сутки, после моделирования ИИ, в зоне деструкции (тканевый детрит) и по ее границам отмечается скопление «зернистых шаров». Нейроны в области инфаркта ткани мозга отсутствуют. Отмечается присутствие значительного количества воспалительного клеточного инфильтрата. Отчетливо виден формирующийся избыточный глиомезодермальный рубец. Он располагается перпендикулярно направлению расположения ядер олигодедроглиоцитов, нервных волокон. При этом в области самого инфаркта мозга отмечается выраженная клеточная реакция и разреженность нейропиля. На 14 сутки, после моделирования ИИ, появляются новые капилляры по периферии очага, с прорастанием в зону деструкции. Значительное скопление клеток глии, отмечающееся в очаге инфаркта мозга и по его границам, составляет основу глиомезодермального рубца. При окраске по Чиаччио и по Нисслю, в области зоны деструкции видна происходящая ее организация. Виден формирующийся избыточный глиомезодермальный рубец. Следовательно, формирование зоны деструкции ткани мозга происходит к 3 суткам после моделирования ИИ. В ней выявляется много погибших нейронов. В зоне инфаркта ткани мозга возникает значительная воспалительная клеточная инфильтрация. Формирующийся незрелый глиомезодермаль- 70 ный рубец появляется на 7 сутки. При этом отмечается значительное количество воспалительного клеточного инфильтрата. Новые капилляры по периферии очага, с прорастанием в зону деструкции появляются на 14 сутки. Формируется избыточный глиомезодермальный рубец. 3.2. Гистологическая картина зоны ишемии при моделировании ишемического инсульта при применении ТЭС-терапии Экспериментальным животным группы № 3 проводили 7 сеансов ТЭС-терапии до моделирования ИИ и крысам в группе № 4 проводилось 7 сеансов ТЭС-терапии после моделирования ИИ. Значительных различий между микропрепаратами ткани мозга у животных групп № 3 и № 4 обнаружено не было, поэтому для сравнения с микропрепаратами ткани мозга животных группы № 2 будут представлены только микропрепараты ткани мозга животных группы № 4. Через 1 сутки после моделирования ИИ на микропрепаратах ткани мозга крыс этих групп, окрашенных гематоксилин-эозином, начинает выявляться область формирующегося инфаркта мозга. Зона инфаркта ткани мозга менее выражена, чем у крыс группы № 2. Развитие клеточного ответа на появление очага некроза ткани мозга и признаки перицеллюлярного отека ткани мозга были менее выражены по сравнению с животными группы № 2 (рис. 3.20). На микропрепаратах ткани мозга крыс появлялись единичные клетки-тени (рис. 3.20, поз. 2) и единичные нейтрофилы (рис. 3.20, поз. 3). По сравнению с животными группы № 2 отмечается преобладание гиперхромных дистрофически измененных нейроцитов (рис. 3.20, поз. 1), над содержанием «клеток-теней» (некротизированных нейронов). В центре области ишемии в капиллярах выявлялось запустевание, они становились незаметными на общем фоне ткани (рис. 3.20, поз. 4). По периферии очага ишемии капилляры хорошо прокрашены (рис. 3.20, поз. 5), в них выявляются признаки отека эндотелиоцитов, стаза крови, адгезия Нф к эндотелию капилляров (рис. 3.20, поз. 6), в некоторых признаки сдавления сосудов отечными ножками астроцитов (рис. 3.20, поз. 7). 71 Рисунок 3.20 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска гематоксилин-эозин Рисунок 3.21 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска по Нисслю На срезах мозга, окрашенных по Нисслю, обращает на себя внимание значительно меньшее количество гиперхромных дистрофически измененных нейронов по сравнению с крысами группы № 2 (рис. 3.21, поз. 1), преобладают нейроны с признаками частичного тигролиза (рис. 3.21, поз. 2). Значительно меньше встречается Нф и явлений нейронофагии. Это отражает большую сохранность нейроцитов и меньшую выраженность воспаления. На микропрепаратах ткани мозга, окрашенных по Чиаччио, отмечается некоторая гипохромия ядер клеток (рис. 3.22, поз. 1) в очаге ишемии, что возможно отражает текущие процессы деполимеризации ДНК в некротизированных клетках. Кроме того отмечается повышение интенсивности восприимчивости окраски ткани по периферии очага ишемии к судану черному, что отражает текущий процесс демиелинизации (рис. 3.22, поз. 2). В очаге ишемии также видна разреженность нейропиля, что показывает выраженность перицеллюлярного отека (рис. 3.22, поз. 1). Следует отметить, что проявления вышеуказанных изменений на микропрепаратах крыс группы № 4 (рис. 3.22) выражены меньше, чем на микропрепаратах крыс группы № 2. На 3 сутки после моделирования ИИ четко определяется область инфаркта мозга (рис. 3.23, поз. 1). Обращает на себя внимание воспалительная клеточная инфильтрация зоны инфаркта ткани мозга (рис. 3.23, поз. 2), которая более выражена на микропрепаратах крыс группы № 2. 72 Рисунок 3.22 — Срез мозга крысы группы № 4. Ув. об. 10. Окраска по Чиаччио Рисунок 3.23 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска гематоксилин-эозин Воспалительный клеточный инфильтрат служит в последующем основой формирования глиомезодермального рубца. Выявляются погибшие нейроны (клетки-тени), количество которых значительно меньше, чем на микропрепаратах крыс группы № 2 в тот же период (рис. 3.23, поз. 3). На микропрепаратах ткани мозга животных группы № 4, окрашенных по Нисслю, по сравнению с животными группы № 2, определяется значительно большее количество нейронов, некоторые из которых имеют признаки частичного тигролиза (рис. 3.24, поз. 1). Выявляется мало гиперхромных дистрофически измененных нейронов (рис. 3.24, поз. 2), единичные признаки нейронофагии (рис. 3.24, поз. 3), воспалительная клеточная инфильтрация (рис. 3.24, поз. 4) выражена значительно меньше, чем у животных группы № 2. На микропрепаратах ткани мозга животных группы № 4, окрашенных по Чиаччио, в отличие от животных группы № 2, зона деструкции значительно менее выражена (рис. 3.25, поз. 1). В ней выявляются редкие рассеянные гипохромные ядра клеток красного цвета. По периферии зоны деструкции располагается область плотного компактно сгруппированного расположения ядер клеток (область воспалительного клеточного ответа) интенсивно краснофиолетового цвета (рис. 3.25, поз. 2). На 7 сутки после воссоздания модели ИИ в зоне деструкции наличия нейронов на микропрепаратах ткани мозга животных групп № 2, № 3 и № 4 73 Рисунок 3.24 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска по Нисслю Рисунок 3.25 — Срез мозга крысы группы № 4. Ув. об. 20. Окраска по Чиаччио не обнаружено. На микропрепаратах ткани мозга крыс группы № 4, по сравнению с животными группы № 2, обнаружено больше «зернистых шаров» (рис. 3.26, поз. 1). На микропрепаратах ткани мозга крыс группы № 4, в отличие от животных группы № 2, обращает на себя внимание значительное количество новообразованных капилляров по периферии области инфаркта мозга с прорастанием их вглубь зоны деструкции (рис. 3.26, поз. 2). На 7 сутки после моделирования ишемического инсульта исследование микропрепаратов ткани мозга крыс группы № 4, окрашенных по Нисслю, показало, что нейроны в области инфаркта ткани мозга отсутствуют (рис. 3.27, поз. 1). Рисунок 3.26 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска гематоксилин-эозин Рисунок 3.27 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска по Нисслю 74 Наблюдается выраженный воспалительный клеточный инфильтрат (рис. 3.27, поз. 1). По сравнению с животными группы № 2 «зернистых шаров» (рис. 3.27, поз. 2) больше. Глиомезодермальный рубец на микропрепаратах ткани мозга животных группы № 4 (рис. 3.28, поз. 1), по сравнению с животными группы № 2, выглядит более сформированным. В области формирования глиомезодермального рубца (рис. 3.28, поз. 1) отмечены признаки активного ангиогенеза в виде появления новообразованных капилляров (рис. 3.28, поз. 2) по периферии с прорастанием вглубь очага инфаркта мозга. На микропрепаратах ткани мозга крыс группы № 4, окрашенных по Чиаччио, в области самого инфаркта мозга, по сравнению с животными группы № 2, отмечается значительно менее выраженная разреженность нейропиля и клеточная реакция (рис. 3.29). При анализе обзорных микропрепаратов ткани мозга крыс группы № 4 (рис. 3.29) отмечено, что формирующийся глиомезодермальный рубец не блокирует ход нервных волокон, о чем можно судить по направлению расположения окрашенным красным ядер олигодендроглиоцитов (рис. 3.29, поз. 2). О большей степени зрелости глиомезодермального рубца у животных группы № 4 свидетельствует более равномерное распределение ядер клеток по зоне деструкции (рис. 3.29, поз. 1). Это отражает процесс ее организации. Рисунок 3.28 — Срез мозга крысы группы № 4. Ув. об. 10. Окраска по Нисслю Рисунок 3.29 — Срез мозга крысы группы № 4. Ув. об. 10. Окраска по Чиаччио 75 На микропрепаратах ткани мозга, окрашенных гематоксилин-эозином, крыс группы № 4 на 14 сутки после моделирования ИИ видны активно текущие процессы организации в зоне деструкции (рис. 3.30, поз. 1). По ее периферии образуется глиомезодермальный рубец. Он выглядит более зрелым и гораздо менее плотным (рис. 3.31) по сравнению с животными группы № 2. По сравнению с 7 сутками на всех микропрепаратах видно некоторое увеличение содержания зернистых шаров. Рисунок 3.30 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска гематоксилин-эозин Рисунок 3.31 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска гематоксилин-эозин На микропрепаратах крыс группы № 4 на 14 сутки после моделирования ИИ, окрашенных по Нисслю, в области зоны деструкции виден формирующийся глиальный рубец, который выглядит более зрелым по сравнению с животными группы № 2 (рис. 3.32). На 14 сутки на микропрепаратах ткани мозга крыс группы № 4, окрашенных по Чиаччио, формирующийся глиомезодермальный рубец выглядит более зрелым по сравнению с животными группы № 2 (рис. 3.33). Таким образом, проведение ТЭС-терапии до моделирования ИИ или сразу после уменьшает степень повреждения мозга в зоне ишемии. В 1 сутки менее выражена зона инфаркта ткани мозга. В связи с этим менее активны: развитие клеточного ответа на появление очага некроза ткани мозга и признаки перицеллюлярного отека ткани мозга. 76 Рисунок 3.32 — Срез мозга крысы группы № 4. Ув. об. 40. Окраска по Нисслю Рисунок 3.33 — Срез мозга крысы группы № 4. Ув. об. 10. Окраска по Чиаччио В зоне ишемии преобладают гиперхромные дистрофически измененные нейроциты, над содержанием некротизированных нейронов. Больше нейронов с признаками частичного тигролиза. Значительно меньше встречается нейтрофилов и явлений нейронофагии. Это отражает большую сохранность нейроцитов и меньшую выраженность воспаления. Гипохромия ядер клеток (отражает текущие процессы деполимеризации ДНК в некротизированных клетках) в очаге ишемии, повышение интенсивности восприимчивости окраски ткани по периферии очага ишемии к судану черному (отражает текущие процесс демиелинизации) также менее выражены. На 3 сутки после моделирования ИИ четко определяется область инфаркта мозга. Обращает на себя внимание выраженность воспалительной клеточной инфильтрации зоны инфаркта ткани мозга. Значительно меньше выявляется некротизированных нейронов (клетки-тени). Мало определяется гиперхромных дистрофически измененных нейронов, но значительно большее количество интактных нейронов, некоторые из которых имеют признаки частичного тигролиза. Значительно меньше выражена воспалительная клеточная инфильтрация. На 7 сутки после воссоздания модели ИИ в зоне деструкции обнаружено больше «зернистых шаров». Обращает на себя внимание более значительное количество новообразованных капилляров по периферии области инфаркта 77 мозга с прорастанием их вглубь зоны деструкции. Глиомезодермальный рубец более сформирован. В области формирования глиомезодермального рубца отмечены признаки активного ангиогенеза в виде появления новообразованных капилляров по периферии с прорастанием вглубь очага инфаркта мозга. Ход нервных волокон не блокируется формирующимся глиомезодермальным рубцом, на что указывает направление расположения окрашенных красных ядер олигодендроглиоцитов. О большей степени зрелости глиомезодермального рубца на 14 сутки свидетельствует более равномерное распределение ядер клеток по зоне деструкции. Это отражает процесс ее организации. 3.3. Изменение объема очага инфаркта мозга при моделировании ишемического инсульта без проведения и с применением ТЭС-терапии Для оценки динамики объема очага инфаркта произведен анализ его размеров на срезах окрашенных тетразолиевым методом у крыс групп № 2, № 3 и № 4 на 1 и 3 сутки после моделирования ИИ. Определяли показатель «удельный объем инфаркта мозга» путем подсчета соотношения объема очага инфаркта мозга к объему всего мозга. Указанный показатель позволяет значительно уменьшить разброс, обусловленный разными размерами мозга и соответственно объемами очагов инфаркта у животных разной массы [Г.Г. Автандилов, 1990]. На срезах мозга группах № 2, № 3 и № 4 сделанных на 1 сутки четко выявлялась область инфаркта мозга, локализованная в зоне кровоснабжения правой средней мозговой артерии с захватом неокортекса и небольшого участка каудопутамена. Показатель удельного объема очага инфаркта мозга у крыс группы № 2 на 1 сутки составил 0,108 ± 0,003 у.е. На 3 сутки он достоверно (р ≤ 0,05) возрастал на 18,3 % и составил 0,132 ± 0,010 у.е. (табл. 3.1). 78 Таблица 3.1 Удельный объем очага инфаркта мозга у крыс групп № 2, № 3 и № 4 на 1 и 3 сутки после моделирования ишемического инсульта 2 группа 2 группа 3 группа 3 группа 4 группа 4 группа 1 сутки 3 сутки 1 сутки 3 сутки 1 сутки 3 сутки N=5 N=5 N=5 N=5 N=5 N=5 М 0,108 0,132 0,104 0,104 0,099 0,095 m 0,003 0,010 0,001 0,001 0,004 0,004 р < 0,05 р* > 0,05 < 0,05 р** > 0,05 < 0,05 < 0,05 р*** < 0,05 > 0,05 < 0,05 Примечание: р — достоверность различия между 1 и 3 сутками в группе; р* — достоверность различия в 1 сутки между группами; р** — достоверность различия в 3 сутки со 2 группой; р*** — достоверность различия между 3 и 4 группой. У крыс группы № 3 показатель удельного объема очага инфаркта мозга на 1 сутки составил 0,104 ± 0,001 у.е. На 3 сутки размер объема очага инфаркта мозга достоверно (р ≥ 0,05) не изменялся и составлял 0,104 ± 0,001 у.е. (табл. 3.1). Показатель удельного объема очага инфаркта мозга у крыс группы № 4 на 1 сутки составил 0,099 ± 0,004 у.е. На 3 сутки его размер достоверно (р ≥ 0,05) не изменялся и составлял 0,095 ± 0,004 у.е. (табл. 3.1). Таким образом на 1 сутки после моделирования ИИ у крыс группы № 2 удельный объем очага инфаркта мозга был достоверно (р ≤ 0,05) больше на 4,2 %, чем у животных группы № 3, которым проводили 3 сеанса ТЭС-терапии до моделирования ИИ. Удельный объем очага инфаркта мозга у крыс группы № 2 на 1 сутки после моделирования ИИ был также достоверно (р ≤ 0,05) больше на 8,9 %, чем у крыс группы № 4, которым к этому времени проводили 1 сеанс ТЭС-терапии после моделирования ИИ. 79 На 1 сутки после моделирования ИИ у крыс групп № 3 и № 4 объем очага инфаркта мозга достоверно (р ≥ 0,05) не отличался. Удельный объем очага инфаркта мозга на 3 сутки после моделирования ИИ у крыс группы № 2 составил 0,132 ± 0,010 у.е., что достоверно (р ≤ 0,05) больше на 21,6 % по сравнению с удельным объемом очага инфаркта мозга у крыс группы № 3. Сравнение удельного объема очага инфаркта мозга крыс группы № 2 и № 4 на 3 сутки после моделирования ИИ показало, что у крыс группы № 2 он достоверно (р ≤ 0,05) на 28,0 % больше, чем у животных группы № 4. У крыс группы № 3 удельный объем очага инфаркта мозга на 3 сутки после моделирования ИИ был достоверно (р ≤ 0,05) больше на 8,3 % по сравнению с животными группы № 4. Итак, использование ТЭС-терапии, как до, так и после моделирования ИИ, снижало степень повреждения ткани мозга. Удельный объем очага инфаркта мозга был меньше у крыс группы № 3 (на 4,2 %) и группы № 4 (на 8,9 %) по сравнению с животными группы № 2, которым ТЭС-терапию не проводили. При этом, как показал анализ полученных результатов, удельный объем очага инфаркта мозга у крыс групп № 3 и № 4 не возрастал к моменту его полного формирования (3 сутки), что имело место у крыс группы № 2. Таким образом, к 3 суткам после моделирования ИИ удельный объем очага инфаркта мозга у крыс группы № 3 был на 21,6 % и у крыс группы № 4 — на 28,0 % меньше, чем у крыс группы № 2. Кроме того, у крыс группы № 3 он к 3 суткам был достоверно (р ≤ 0,05) больше на 8,3 % по сравнению с животными группы № 4. Следовательно, применение ТЭС-терапии наиболее эффективно после моделирования ИИ. 80 ГЛАВА 4. ДИНАМИКА СОДЕРЖАНИЯ ЦИТОКИНОВ И β-ЭНДОРФИНА В ПЛАЗМЕ КРОВИ КРЫС ПРИ МОДЕЛИРОВАНИИ ИШЕМИЧЕСКОГО ИНСУЛЬТА 4.1. Динамика содержания цитокинов в плазме крови крыс при моделировании ишемического инсульта 4.1.1. Динамика содержания фактора некроза опухоли-α у крыс с экспериментальным ишемическим инсультом Острое асептическое воспаление лежит в основе расширения зоны инфаркта мозга в течение первой недели после возникновения инсульта [K.J. Becker, 2001; G.J. del Zoppo, K.J. Becker, J.M. Hallenbeck, 2001; M.D. Ginsberg, 2003; M. Fisher, R. Ratan, 2003; P.L. Wood, 2003; R. Markus, D.C. Reutens, S. Kazui, 2004; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; Q. Wang, X.N. Tang, M.A. Yenari, 2007; А.Х. Хана-Мурад, Л.И. Павлинова, А.А. Мокрушин, 2007; A. Tuttolomondo, D. Di Raimondo, R. Di Sciacca, 2008; J. Jordan et al., 2008; D. Amantea et al., 2008; C. Benakis, L. Hirt, R.A. Du Pasquier, 2009; Ю.Д. Губарев, А.О. Шеремет, 2009; А.С. Кость, Б.Д. Луцик, Л.Е. Лаповець, 2010; A.G. Ceulemans et al., 2010; Y. Lee et al., 2014; М.В. Онуфриев и соавт., 2014]. Активность воспалительной реакции в очаге ишемии обусловлена возрастанием уровня провоспалительных цитокинов (ИЛ-1, ФНО-α, ИЛ-6, ИЛ-8) [Е.И. Гусев, В.И. Скворцова, 2001; N. Quan, M. Herkenham, 2002; А.А. Аракелян и соавт., 2005; Л.Н. Кашаева, Л.М. Карзакова, В.Н. Саперов, 2005; Q. Wang, X.N. Tang, M.A. Yenari, 2007; D. Amantea et al., 2008; M.Y. Zuidema, C. Zhang, 2010; М.В. Онуфриев, 2014]. ФНО-α, ИЛ-1β и ИЛ-6, являются ключевыми медиаторами микроглиальных нейроиммунных процессов и вырабатываются локально в ответ церебральную ишемию [A.C.H. Yu, L.T. Lau, 2000; H. Boutin et al., 2001; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; A. Tuttolomondo et al., 2008; S. ShenharTsarfaty et al., 2008; S. Choi, W. J. Friedman, 2009; A. Maddahi, L. Edvinsson, 2010; 81 M. Zhang et al., 2011]. ФНО-α — плейотропный полипептид, вырабатываемый Нф, активированными МФ, астроцитами и эндотелиальными клетками [J. Zaremba, 2000; С.А. Кетлинский, А.С. Симбирцев, 2008]. Он оказывает токсической действие на нейроны путем активации системы генерации АФК, что резко индуцирует процессы ПОЛ [J.J. Viel, D.Q. Mc Manus, S.S. Smith, 2001; A. Quintana et al., 2005; А.Е. Бугрова, 2007; R.M. Adibhatla, J.F. Hatcher, 2007; R.M. Adibhatla, R. Dempsey, J.F. Hatcher, 2008; N. Liu et al., 2009; А.В. Алексеенко, 2013]. Таким образом, изменение его содержания влияет на степень повреждения ткани мозга. При исследовании плазмы крови у крыс группы № 1 (интактные животные) уровень ФНО-α составил 8,41 ± 2,53 пг/мл (табл. 4.1). Таблица 4.1 Динамика уровня ФНО-α при экспериментальном ишемическом инсульте у животных Показатель ФНО-α, М ±m р р* р** р*** Контроль 8,41 2,53 1 сутки 17,82 6,31 < 0,05 3 сутки 18,36 7,20 < 0,05 > 0,05 7 сутки 17,82 3,79 < 0,05 > 0,05 > 0,05 14 сутки 21,01 8,06 < 0,05 > 0,05 > 0,05 > 0,05 Примечание: ФНО-α — фактор некроза опухоли-α; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. В 1 сутки после моделирования ИИ содержание ФНО-α в плазме крови животных группы № 2 составило 17,82 ± 11,31 пг/мл, что было достоверно (р ≤ 0,05) больше на 52,80 %, чем в плазме крови интактных животных (рис. 4.1). 82 Рисунок 4.1 — Изменения содержания ФНО-α при экспериментальном ишемическом инсульте К 3 суткам после моделирования ИИ уровень ФНО-α в плазме крови животных, по сравнению с 1 сутками той же группы, достоверно не изменился (р ≥ 0,05) и составил 18,36 ± 7,20 пг/мл. Это на 54,19 % достоверно (р ≤ 0,05) больше, чем в плазме крови животных группы № 1 (табл. 4.1). На 7 сутки после моделирования ИИ содержание ФНО-α в плазме крови животных группы № 2, по сравнению с 1 сутками, и животными группы № 2 и 3 сутками крыс группы № 2 достоверно не изменилось (р ≥ 0,05) и составило 17,82 ± 3,79 пг/мл, что достоверно (р ≤ 0,05) больше на 52,80 %, чем в плазме крови интактных животных (рис. 4.1). К 14 суткам после моделирования ИИ уровень ФНО-α в плазме крови животных группы № 2 составил 21,01 ± 8,06 пг/мл, что, по сравнению с животными группы № 1, было достоверно (р ≤ 0,05) больше на 59,97 %, а по сравнению с 3 и 7 сутками у животных группы № 2 достоверно не изменился (р ≥ 0,05). Таким образом, содержание ФНО-α в плазме крови животных группы № 2 повышалось с момента моделирования ИИ и оставалось повышенным в течение всего периода наблюдения. 83 4.1.2. Динамика содержания интерлейкина-1β у крыс с экспериментальным ишемическим инсультом Синтез ИЛ-1 микроглией в условиях церебральной ишемии является активирующим сигналом для запуска образования других провоспалительных цитокинов, а также стимуляции астроцитов к продукции потенциальных нейротоксичных веществ, таких как NO и метаболиты арахидоновой кислоты [H. Boutin et al., 2001; L.D. Church, P.G. Cook, M.F. Mc Dermott, 2008; S. Choi, W.J. Friedman, 2009; C.A. Dinarello, 2009; C.A. Dinarello, 2011; H.F. Green et al., 2012; J. Galea, D. Brough, 2013; C. Garlanda, C.A. Dinarello, A. Mantovani, 2013]. Содержание ИЛ-1β в плазме крови у крыс группы № 1 (интактные животные) составило 30,93 ± 7,90 пг/мл (табл. 4.2). Таблица 4.2 Динамика уровня ИЛ-1β у животных с экспериментальным ишемическим инсультом Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-1β, М 30,93 464,60 1798,99 623,08 407,73 ±m 7,90 127,43 586,25 56,97 125,34 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 < 0,05 < 0,05 р р* р** р*** < 0,05 Примечание: ИЛ-1β — интерлейкин-1β; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Через 1 сутки в плазме крови животных группы № 2 после моделирования ишемического инсульта концентрация ИЛ-1β составила 464,60 ± 127,43 пг/мл, что достоверно (р ≤ 0,05) больше на 93,34 %, чем в плазме крови животных группы № 1 (рис. 4.2). 84 Рисунок 4.2 — Изменения содержания ИЛ-1β при экспериментальном ишемическом инсульте К 3 суткам в плазме крови животных группы № 2 уровень ИЛ-1β составил 1798,99 ± 586,25 пг/мл, по сравнению с 1 сутками он стал достоверно (р ≤ 0,05) больше на 74,17 % и на 98,28 % достоверно больше (р ≤ 0,05), чем в плазме крови у животных группы № 1. К 7 суткам после моделирования ИИ в плазме крови животных группы № 2 содержание ИЛ-1β по сравнению с 1 сутками было достоверно (р ≤ 0,05) больше на 25,43 %. По сравнению с 3 сутками уровень стал достоверно (р ≤ 0,05) меньше на 65,36 % и составило 623,08 ± 56,97 пг/мл (табл. 4.2). Это достоверно (р ≤ 0,05) больше на 95,03 %, чем в плазме крови животных группы № 1 (рис. 4.2). На 14 сутки после моделирования ИИ в плазме крови у животных группы № 2 содержание ИЛ-1β вернулся к уровню 1 суток. По сравнению с 3 сутками уровень ИЛ-1β стал на 77,33 % достоверно меньше (р ≤ 0,05). По отношению к 7 суткам был достоверно (р ≤ 0,05) меньше на 34,56 % и составил 407,73 ± 125,34 пг/мл (табл. 4.2 и рис. 4.2). Итак, уровень ИЛ-1β с началом ишемии значительно возрастал, достигая максимума к 7 суткам. К 14 суткам он снижался до уровня, наблюдаемого в 1 сутки после моделирования ИИ. Однако содержание ИЛ-1β оставалось выше контроля на 75,8 %. 85 4.1.3. Динамика содержания интерлейкина-6 у крыс с экспериментальным ишемическим инсультом ИЛ-6 синтезируется многими типами клеток, обеспечивающими запуск и регуляцию воспаления и иммунного ответа [D.A. Papanicolaou et al., 1998; С.А. Кетлинский, А.С. Симбирцев, 2008]. Он активирует экспрессию молекул адгезии и хемотаксис лейкоцитов, а также является важнейшим индуктором острофазного ответа [K. Paul-Pletzer, 2006; С.А. Кетлинский, А.С. Симбирцев, 2008; I.M. Cojocaru et al., 2009; T. Tanaka et al., 2013]. Показано, что ИЛ-1 и ФНО-α являются мощными индукторами ИЛ-6-синтетазы в астроцитах: быстрое увеличение содержания микроглиального ИЛ-1β в первые 1–2 ч церебральной ишемии вызывает усиленный синтез ИЛ-6 [H. Boutin et al., 2001]. Увеличение концентрации провоспалительных цитокинов (ФНО-α, ИЛ-1, ИЛ-6) при развитии острой локальной церебральной ишемии расширяет конечные размеры инфаркта мозга (зона некроза) [Е.И. Гусев, В.И. Скворцова, 2001; H. Boutin et al., 2001; K.Y. Kwan, B.H. Jeon, 2001; N. Quan, M. Herkenham, 2002; Q. Wang, X.N. Tang, M.A. Yenari, 2007; D. Amantea et al., 2008; I.M. Cojocaru et al., 2009; M. Zhang et al., 2011; М.В. Онуфриев, 2014]. При исследовании плазмы крови у крыс группы № 1 на содержание ИЛ-6 его концентрация составила 0,43 ± 0,16 пг/мл (табл. 4.3). У животных из группы № 2 в 1 сутки после моделирования ИИ в плазме крови уровень ИЛ-6 составил 6,87 ± 1,93 пг/мл, что было достоверно (р ≤ 0,05) больше на 93,74 %, чем у животных группы № 1 (рис. 4.3). На 3 сутки после моделирования ИИ в плазме крови животных группы № 2 содержание ИЛ-6, по сравнению с 1 сутками, стало достоверно (р ≤ 0,05) больше на 33,75 % и составило 10,37 ± 1,91 пг/мл. Это достоверно (р ≤ 0.05) больше на 95,85 %, чем в плазме крови животных группы № 1 (рис. 4.3). Уровень ИЛ-6 в плазме крови животных группы № 2 на 7 сутки после моделирования ИИ, по сравнению с 1 сутками, был достоверно (р ≥ 0,05) больше на 40,36 %. По отношению к 3 суткам он достоверно не изменился и составил 11,52 ± 2,73 пг/мл. Это достоверно (р ≤ 0,05) больше на 96,26 %, чем в плазме крови животных группы № 1 (табл. 4.3. и рис. 4.3). 86 Таблица 4.3 Динамика уровня ИЛ-6 у животных с экспериментальным ишемическим инсультом Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-6, М 0,43 6,87 10,37 11,52 10,20 ±m 0,16 1,93 1,91 2,73 2,83 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 р р* р** р*** > 0,05 Примечание: ИЛ-6 — интерлейкин-6; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.3 — Изменения содержания ИЛ-6 при экспериментальном ишемическом инсульте К 14 суткам после моделирования ИИ содержание ИЛ-6 в плазме крови животных группы № 2, по сравнению с 1 сутками, стало достоверным (р ≤ 0,05) больше на 32,64 %. По отношению к 3 и 7 суткам достоверных (р ≥ 0,05) изменений не отмечено, а по сравнению с животными группы № 1 оно стало достоверно (р ≤ 0,05) больше на 95,88 %, составив 10,20 ± 2,83 пг/мл. 87 Таким образом, после моделирования ИИ содержание ИЛ-6, достигало максимума на 3 сутки. На 7 и 14 сутки уровень ИЛ-6 не претерпевал изменений, оставаясь на высоком уровне. 4.1.4. Динамика содержания интерлейкина-10 у крыс с экспериментальным ишемическим инсультом Противовоспалительные цитокины, такие как ИЛ-10 и ТФР-1β, подавляя развитие воспаления, оказывают защитный эффект в условиях ишемии ГМ [K. Strle et al., 2001; Y. Zhu et al., 2002; N. Quan, M. Herkenham, 2002; N. Vila et al., 2003; Г.Н. Жданов, 2006; M. Kaste, H.J. Castillo, S. de Compostela, 2006; N. Liu et al., 2009]. ИЛ-10, являясь мощным регулятором клеточно-опосредованного иммунного ответа моноцитов/макрофагов, снижает продукцию ПГЕ2, угнетает экспрессию ICAM-1 активно участвующих в развитии местного воспаления, подавляет выработку многочисленных провоспалительных цитокинов, включая ИЛ-1, ФНО-α, ИЛ-6, ИЛ-8 [С.А. Кетлинский, А.С. Симбирцев, 2008; N. Liu et al., 2009; T. Nabe et al., 2012]. ИЛ-10 также подавляет синтез супероксидного аниона и реактивных промежуточных продуктов обмена кислорода. Содействие синтезу NO является одним из многочисленных активирующих эффектов ИЛ-10 на моноциты/макрофаги наряду с индукцией им выработки антагониста рецептора для ИЛ-1. Таким образом, ИЛ-10 является фактором, модулирующим функцию макрофагов и обладающим мощной противовоспалительной активностью, что объясняет его нейропротекторные свойства при ишемическом инсульте, а его низкая концентрация в динамике заболевания наблюдается в случаях тяжелого, неблагоприятного течения инсульта [K. Strle et al., 2001; N. Vila et al., 2003; Г.Н. Жданов, 2006; N. Liu et al., 2009]. У крыс группы № 1 (интактные животные) содержание ИЛ-10 в плазме крови составило 1,21 ± 0.66 пг/мл (табл. 4.4). 88 В плазме крови животных из группы № 2 в 1 сутки после моделирования ИИ уровень ИЛ-10 составил 39,79 ± 10,76 пг/мл, что достоверно (р ≤ 0,05) больше на 96,95 %, чем в плазме крови животных группы № 1 (рис. 4.4) Таблица 4.4 Динамика уровня ИЛ-10 у животных с экспериментальным ишемическим инсультом Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-10, М 1,21 39,79 49,85 78,69 46,48 ±m 0,66 10,76 11,67 15,96 10,67 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 < 0,05 < 0,05 < 0,05 > 0,05 р р* р** р*** < 0,05 Примечание: ИЛ-10 — интерлейкин-10; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.4 — Изменения содержания ИЛ-10 при экспериментальном ишемическом инсульте 89 На 3 сутки после моделирования ИИ содержание ИЛ-10 в плазме крови животных группы № 2 по сравнению с 1 сутками достоверно (р ≥ 0,05) не изменилось и составило 49,85 ± 11,67 пг/мл, что достоверно (р ≤ 0,05) больше на 97,57 %, чем в плазме крови животных группы № 1 (рис. 4.4). Уровень ИЛ-10 в плазме крови животных группы № 2 на 7 сутки после моделирования ИИ стал достоверно (р ≤ 0,05) больше на 49,43 % по сравнению с 1 сутками. По сравнению с 3 сутками он стал достоверно (р ≤ 0,05) больше на 36,65 % и составил 78,69 ± 15,96 пг/мл. Это достоверно (р ≤ 0,05) больше на 98,46 %, чем в плазме крови животных группы № 1 (табл. 4.4, рис. 4.4). На 14 сутки после моделирования ИИ содержание ИЛ-10 в плазме крови животных группы № 2 вернулось (р ≤ 0,05) к уровню 1 суток. Оно стало достоверно (р ≤ 0,05) меньше на 48,55 %, в сравнении с 7 сутками. По сравнению с животными группы № 1 содержание ИЛ-10 в плазме крови оставалось достоверно (р ≤ 0,05) больше на 97,01 % и составляло 40,48 ± 10,67 пг/мл. Итак, уровень ИЛ-10 резко увеличивался на 1 сутки ишемии мозга. Затем значительно возрастал к 7 суткам. К 14 суткам он снижался до значений, наблюдаемых в 1 сутки, оставаясь, таким образом, намного выше исходного уровня, отмеченного у интактных животных. 4.1.5. Динамика содержания трансформирующего фактора роста-β у крыс с экспериментальным ишемическим инсультом ТФР-1β угнетает иммунные реакции в ткани мозга, а также понижает чувствительность клеток ГМ к гипоксии [J.J. Letterio, 2000; Y. Zhu et al., 2002]. Защитное противоишемическое действие ТФР-1β обусловлено угнетением адгезии лейкоцитов к эндотелию, подавлением выработки МФ повреждающих кислородных и NO метаболитов [J.J. Letterio, 2000, N. Vila et al., 2003]. Нейропротективная способность ТФР-1β связана со стабилизацией нейронального кальциевого гомеостаза и повышением экспрессии гена bcl-2, что приводит к значительному торможению свободнорадикального повреждения, замедлению процессов некроза и апоптоза [J.J. Letterio, 2000; Y. Zhu 90 et al., 2002]. Установлена важная роль ТФР-1β, в увеличении васкуляризации ткани мозга после острой церебральной ишемии, особенно в зоне пенумбры [J.J. Letterio, 2000]. Содержание ТФР-1β в плазме крови у крыс группы № 1 (интактные животные) составило 22,22 ± 7,67 пг/мл (табл. 4.5). На 1 сутки после моделирования ИИ уровень ТФР-1β в плазме крови животных группы № 2 составил 43,35 ± 10,93 пг/мл, что достоверно (р ≤ 0,05) больше на 48,74 %, чем в плазме крови из группы № 1 (рис. 4.5). Таблица 4.5 Динамика уровня ТФР-1β у животных с экспериментальным ишемическим инсультом Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ТФР-1β, М 22,22 43,35 62,97 66,30 45,72 ±m 7,67 10,93 11,41 14,93 12,53 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 < 0,05 р р* р** р*** < 0,05 Примечание: ТФР-1β — трансформирующий фактор роста β; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Содержание ТФР-1β в плазме крови животных группы № 2 к 3 суткам после моделирования ИИ достоверно (р ≤ 0,05) возросло на 31,15 % по сравнению с 1 сутками и составило 62,97 ± 11,41 пг/мл, что достоверно (р ≤ 0,05) больше на 64,71 %, чем в плазме крови животных группы № 1 (табл. 4.5, рис. 4.5). На 7 сутки после моделирования ИИ в плазме крови животных группы № 2 уровень ТФР-1β по сравнению с 1 сутками стал достоверно (р ≤ 0,05) 91 больше на 34,61 %. По сравнению с 3 сутками он достоверно (р ≥ 0,05) не изменился и составил 66,30 ± 14,03 пг/мл. Это достоверно (р ≤ 0,05) больше на 66,48 %, чем в плазме крови животных группы № 1. Рисунок 4.5 — Изменения содержания ТФР-1β при экспериментальном ишемическом инсульте К 14 суткам после моделирования ИИ содержание ТФР-1β в плазме крови животных группы № 2 по сравнению с 1 сутками достоверно (р ≥ 0,05) не изменилось. Однако оно стало достоверно (р ≤ 0,05) на 27,39 % меньше по сравнению с 3 сутками. К 7 суткам уровень ТФР-1β стал достоверно (р ≤ 0,05) меньше на 31,04 %. Однако по сравнению с группой № 1 был достоверно (р ≤ 0,05) больше на 51,39 % и составлял 45,72 ± 12,53 пг/мл. Таким образом, уровень ТФР-1β значительно возрастал в 1 сутки после моделирования ИИ. К 3 суткам он достигал максимума, не меняясь на 7 сутки. На 14 сутки уровень ТФР-1β снижался до величины, отмеченной в 1 сутки, оставаясь, следовательно, достаточно высоким. 4.2. Динамика содержания β-эндорфина у крыс с экспериментальным ишемическим инсультом β-эндорфин важнейший представитель среди семейства ОП является агонистом µ и δ классических ОР [В.А. Дубынин, А.А. Каменский, 2010; 92 Ю.А. Ковалицкая, Е.В. Наволоцкая, 2011; И.Л. Шаравьева, 2011; О.В. Нечайкина и соавт., 2013]. Имеются данные свидетельствующие о нейропротекторном, противовоспалительном, антиоксидантном, нейротрофическом, антигипоксическом эффектах активации δ-опиоидных рецепторов [G.J. Gross, 2000; Н.В. Соленкова, 2002; H. Zhang et al., 2006; J.M. Gidday, 2006; Е.И. Барзах, 2006; M. Iwata et al., 2007; D.S. Su et al., 2007; A.K. Rehni, N. Singh, S.A. Jaggi, 2007; C. Happel et al., 2008; M. Govindaswami et al., 2008; Y. Yang et al., 2009; S. Husain, D.E. Potter, C.E. Crosson, 2009; M.R. Hutchinson et al., 2011; S. Husain, G.I. Liou, C.E. Crosson, 2011; Y. Feng et al., 2011; N. Akhter et al., 2013; M. Staples et al., 2013; К.П. Иванов, 2013]. Содержание β-эндорфина в плазме крови интактных животных составило 27,52 ± 6,14 пг/мл (табл. 4.6). Таблица 4.6 Содержание β-эндорфина при моделировании ишемического инсульта у животных группы № 2 Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки β-эндорфин, М 27,51 13,95 11,10 9,63 9,62 ±m 6,13 1,88 1,33 2,65 3,02 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 р р* р** р*** > 0,05 Примечание: р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. В плазме крови животных из группы № 2 на 1 сутки после моделирования ИИ уровень β-эндорфина составил 13,95 ± 1,88 пг/мл, что достоверно (р ≤ 0,05) меньше на 49,31 %, чем в плазме крови животных группы № 1 (рис. 4.6). 93 Рисунок 4.6 — Изменения содержания β-эндорфина при экспериментальном ишемическом инсульте К 3 суткам после моделирования ИИ в плазме крови животных группы № 2 уровень β-эндорфина по сравнению с 1 сутками стал достоверно (р ≤ 0,05) меньше на 20,35 % и составил 11,11 ± 1,33 пг/мл. Это достоверно (р ≤ 0,05) меньше на 59,62 %, чем в плазме крови животных группы № 1 (табл. 4.6, рис. 4.6). На 7 сутки после моделирования ИИ в плазме крови животных группы № 2 содержание β-эндорфина по сравнению с 1 сутками стало достоверно (р ≤ 0,05) меньше на 30,89 %. По сравнению с 3 сутками содержание β-эндорфина достоверно (р ≥ 0,05) не изменилось и составило 9,64 ± 2,66 пг/мл, что достоверно (р ≤ 0,05) меньше на 64,97 %, чем в плазме крови животных группы № 1 (рис. 4.6). К 14 суткам после моделирования ИИ в плазме крови животных группы № 2 уровень β-эндорфина по сравнению с 3 и 7 сутками достоверно (р ≥ 0,05) не изменился. По сравнению с 1 сутками он остался достоверно (р ≤ 0,05) меньше на 30,96 % и составил 9,63 ± 3,02 пг/мл (рис. 4.6). Итак, моделирование ИИ вызывало падение уровня β-эндорфина на 1 сутки. К 3 суткам снижение содержания β-эндорфина достигало максимума и оставалось на этом уровне в 7 и 14 сутки. 94 4.3. Динамика содержания цитокинов в плазме крови крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии 4.3.1. Динамика содержания фактора некроза опухоли-α у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии У крыс группы № 1 (интактные животные) в плазме крови содержание ФНО-α составило 8,41 ± 2,53 пг/мл (табл. 4.7). К 1 суткам после моделирования ИИ в плазме крови животных группы № 3 уровень ФНО-α составил 8,73 ± 5,03 пг/мл, что достоверно (р ≥ 0,05) не отличалось по сравнению с животными группы № 1. У животных группы № 2 содержание ФНО-α было достоверно (р ≤ 0,05) меньше на 51,01 % (рис. 4.7). У животных группы № 3 на 3 сутки после моделирования ИИ в плазме крови уровень ФНО-α по сравнению с 1 сутками той же группы и группы Таблица 4.7 Динамика уровня ФНО-α при экспериментальном ишемическом инсульте у животных с предварительным использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ФНО-α, М 8,41 8,73 8,84 8,79 8,76 ±m 2,53 2,53 3,37 1,85 2,04 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 р р* р** р*** > 0,05 Примечание: ФНО-α — фактор некроза опухоли-α; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. 95 Рисунок 4.7 — Изменения содержания ФНО-α при экспериментальном ишемическом инсульте с предварительным использованием ТЭС-терапии № 1 достоверно (р ≥ 0,05) не изменился и составил 8,39 ± 1,85 пг/мл. По сравнению с содержанием ФНО-α в плазме крови животных группы № 2 на 3 сутки это было достоверно (р ≤ 0,05) меньше на 54,30 % (табл. 4.7, рис. 4.7). На 7 сутки в плазме крови животных группы № 3 после моделирования ИИ содержание ФНО-α составило 8,79 ± 1,85 пг/мл. По сравнению с 1 и 3 сутками той же группы и группы № 1 достоверных (р ≥ 0,05) отличий не было. Однако по сравнению с 7 сутками группы № 2 уровень ФНО-α стал достоверно (р ≤ 0,05) меньше на 52,91 %. К 14 суткам после моделирования ИИ в плазме крови животных группы № 3 уровень ФНО-α по сравнению с 1, 3 и 7 сутками той же группы и группы № 1 достоверно (р ≥ 0,05) не отличался и составил 8,76 ± 2,04 пг/мл. Это по сравнению с содержанием ФНО-α в плазме крови группы № 2 было достоверно (р ≤ 0,05) меньше на 58,30 %. Таким образом, предварительное использование ТЭС-терапии до моделирования ИИ предупреждало повышение уровня ФНО-α в плазме крови группы № 3. 96 4.3.2. Динамика содержания интерлейкина-1β у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии В плазме крови у интактных крыс группы № 1 содержание ИЛ-1β составило 30,93 ± 7,90 пг/мл (табл. 4.8). На 1 сутки после моделирования ИИ в плазме крови животных группы № 3 уровень ИЛ-1β составил 235,27 ± 69,95 пг/мл, что достоверно (р ≤ 0,05) больше на 86,85 %, чем в плазме крови животных группы № 1. Однако по сравнению с животными группы № 2 содержание ИЛ-1β в плазме крови на 1 сутки достоверно (р ≤ 0,05) меньше на 49,36 % (рис. 4.8). В плазме крови животных группы № 3 на 3 сутки после моделирования ИИ содержание ИЛ-1β составило 190,92 ± 46,71 пг/мл, что достоверно (р ≥ 0,05) не отличалось от уровня ИЛ-1β в плазме крови на 1 сутки у животных той же группы. Однако было достоверно (р ≤ 0,05) больше на 83,79 %, чем у животных Таблица 4.8 Динамика уровня ИЛ-1β при экспериментальном ишемическом инсульте у животных с предварительным использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-1β, М 30,93 235,27 190,92 171,65 120,28 ±m 7,90 69,95 46,71 51,93 56,24 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 < 0,05 < 0,05 > 0,05 < 0,05 р р* р** р*** < 0,05 Примечание: ИЛ-1β — интерлейкин-1β; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. 97 Рисунок 4.8 — Изменения содержания ИЛ-1β при экспериментальном ишемическом инсульте с предварительным использованием ТЭС-терапии группы № 1. По сравнению с содержанием ИЛ-1β в плазме крови животных группы № 2 на 3 сутки это было достоверно (р ≤ 0,05) меньше и на 89,38 % (табл. 4.8, рис. 4.8). На 7 сутки после моделирования ИИ в плазме крови животных группы № 3 уровень ИЛ-1β составил 171,65 ± 51,93 пг/мл, что на 27,04 % достоверно (р ≤ 0,05) меньше, чем на 1 сутки у животных той же группы. Однако по сравнению с содержанием в плазме крови группы № 1 этот уровень оставался достоверно (р≤0,05) на 81,98 % больше. По сравнению с содержанием ИЛ-1β на 7 сутки в плазме крови группы № 2 был достоверно (р ≤ 0,05) меньше на 72,45 % (табл. 4.8, рис. 4.8). К 14 суткам после моделирования ИИ содержание ИЛ-1β в плазме крови животных группы № 3 стало достоверно (р ≤ 0,05) меньше на 48,87 % по сравнению с 1 сутками той же группы. По сравнению с 3 сутками той же группы оно стало достоверно (р ≤ 0,05) на 36,99 % меньше. Также стало достоверно (р ≤ 0,05) меньше на 29,92 % по сравнению с 7 сутками и составило 120,28 ± 56,24 пг/мл. Это на 74,28 % достоверно (р ≤ 0,05) больше, чем в плазме крови животных группы № 1 и достоверно (р ≤ 0,05) меньше на 70,50 %, чем у животных группы № 2 в тот же период. 98 Таким образом, уровень ИЛ-1β, хотя и возрастал в 1 сутки после моделирования ИИ, но значительно меньше, чем у животных группы № 2. На 3 сутки содержание ИЛ-1β не изменялось по сравнению с 1 сутками. К 7 суткам уровень ИЛ-1β снижался по отношению к 3 суткам. Затем содержание ИЛ-1β продолжало снижаться (14 сутки), хотя и не достигало исходных значений. При этом оно все время оставалось значительно ниже, чем у животных группы № 2. Следовательно, использование ТЭС-терапии ограничивало рост уровня ИЛ-1β. 4.3.3. Динамика содержания интерлейкина-6 у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии Содержание ИЛ-6 в плазме крови у крыс группы № 1 (интактные животные) составило 0,43 ± 0,16 пг/мл (табл. 3.9). Таблица 4.9 Динамика уровня ИЛ-6 при экспериментальном ишемическом инсульте у животных с предварительным использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-6, М 0,43 3,53 5,49 0,64 0,68 ±m 0,16 0,77 1,29 0,18 0,17 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 р р* р** р*** > 0,05 Примечание: ИЛ-6 — интерлейкин-6; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. 99 Рисунок 4.9 — Изменения содержания ИЛ-6 при экспериментальном ишемическом инсульте с предварительным использованием ТЭС-терапии В плазме крови животных из группы № 3 на 1 сутки после моделирования ИИ уровень ИЛ-6 составил 3,53 ± 0,77 пг/мл, что достоверно (р ≤ 0,05) больше на 87,81 %, чем у животных группы № 1. По сравнению с содержанием ИЛ-6 в плазме крови на 1 сутки из группы № 2 это было достоверно (р ≤ 0,05) меньше на 48,61 % (рис. 4.9). На 3 сутки после моделирования ИИ содержание ИЛ-6 в плазме крови животных группы № 3 по сравнению с 1 сутками стало достоверно (р ≤ 0,05) больше на 63,29 % и составило 5,49 ± 1,29 пг/мл. Это достоверно (р ≤ 0,05) больше на 92,16 %, чем у животных группы № 1 и достоверно (р ≤ 0,05) меньше на 47,05 % по сравнению с уровнем ИЛ-6 в плазме крови у животных группы № 2 (табл. 4.9, рис. 4.9). К 7 суткам в плазме крови животных группы № 3 уровень ИЛ-6 по сравнению с 1 сутками той же группы стал достоверно (р ≤ 0,05) меньше на 81,86 %. В сравнении с 3 сутками уровень стал достоверно (р ≤ 0,05) меньше на 88,34 % и составил 0,64 ± 0,18 пг/мл, что по сравнению с содержанием ИЛ-6 в плазме крови животных группы № 1 достоверно (р < 0,05) больше на 32,8 %. Содержание ИЛ-6 на 7 сутки в плазме крови у животных группы № 2 было достоверно (р ≤ 0,05) больше на 94,44 % (табл. 4.9, рис. 4.9). 100 На 14 сутки после моделирования ИИ содержание ИЛ-6 в плазме крови составило 0,68 ± 0,17 пг/мл. По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) меньше на 80,73 %. По сравнению с 3 сутками содержание ИЛ-6 уменьшение на 87,61 % было достоверным (р ≤ 0,05). Однако на 7 сутки достоверных (р ≤ 0,05) отличий не отмечено. Уровень ИЛ-6 достоверно (р ≤ 0,05) превышал на 36,76 % таковой у животных группы № 1. При этом он был достоверно (р ≤ 0,05) меньше на 93,33 % уровня ИЛ-6 в плазме крови животных группы № 2. Итак, уровень ИЛ-6, после предварительной ТЭС-терапии, возрастал, однако менее значительно, чем у животных группы № 2. К 3 суткам он достигал максимума, а затем быстро падал и к 14 суткам приближался к исходной величине. Таким образом, проведение ТЭС-терапии до моделирования ИИ значительно ограничивает рост содержания ИЛ-6 после моделирования ИИ. 4.3.4. Динамика содержания интерлейкина-10 у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии Содержание ИЛ-10 в плазме крови у крыс группы № 1 (интактные животные) составило 1,21 ± 0,66 пг/мл (табл. 4.10). В 1 сутки после моделирования ИИ в плазме крови животных группы № 3 содержание ИЛ-10 составило 49,56 ± 8,97 пг/мл, что достоверно (р ≤ 0,05) больше на 97,55 %, чем в плазме крови животных группы № 1. Уровень ИЛ-10 в плазме крови на 1 сутки достоверно (р ≥ 0,05) не отличался по сравнению с животными группы № 2 (рис. 4.10). На 3 сутки после моделирования ИИ в плазме крови животных группы № 3 уровень ИЛ-10 по сравнению с 1 сутками стал достоверно (р ≤ 0,05) больше на 41,22 % и составил 84,32 ± 10,49 пг/мл. Это достоверно (р ≤ 0,05) больше на 98,56 %, с таковым у животных группы № 1. По сравнению с содержанием ИЛ-10 в плазме крови у животных группы № 2 он достоверно (р ≤ 0,05) больше на 40,87 % (табл. 4.10, рис. 4.10). 101 Таблица 4.10 Динамика уровня ИЛ-10 с экспериментальным ишемическим инсультом у животных с предварительным использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-10, М 1,21 49,56 84,32 85,04 76,52 ±m 0,66 8,97 10,49 5,76 11,73 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 р р* р** р*** > 0,05 Примечание: ИЛ-10 — интерлейкин-10; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.10 — Изменения содержания ИЛ-10 при экспериментальном ишемическом инсульте с предварительным использованием ТЭС-терапии Уровень ИЛ-10 в плазме крови животных группы № 3 на 7 сутки после моделирования ИИ составил 85,04 ± 5,76 пг/мл и стал достоверно (р ≤ 0,05) больше на 41,72 % по сравнению с 1 сутками. По сравнению с 3 сутками он достоверно (р ≥ 0,05) не отличался. Уровень ИЛ-10 достоверно (р ≤ 0,05) стал 102 больше на 98,57 % по сравнению с содержанием ИЛ-10 в плазме крови животных группы № 1. Однако в сравнении с животными группы № 2 достоверной разницы (р ≥ 0,05) не отмечено (табл. 4.10, рис. 4.10). К 14 суткам после моделирования ИИ в плазме крови животных группы № 3 содержание ИЛ-10 составило 76,52 ± 11,73 пг/мл. По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) больше на 35,23 %. По отношению к 3 и 7 сутками достоверных (р ≥ 0,05) отличий не выявлено. У животных группы № 1 уровень ИЛ-10 был достоверно (р ≤ 0,05) меньше на 98,41 %. По сравнению с уровнем ИЛ-10 в плазме крови у животных группы № 2 у животных группы № 3 он был достоверно (р ≤ 0,05) больше на 39,25 %. Таким образом, содержание ИЛ-10 после моделирования ИИ возрастает достигая максимума на 3 сутки и остается на этом уровне до 14 суток. При этом в сравнении с группой № 2 уровень ИЛ-10 был значительно выше. 4.3.5. Динамика содержания трансформирующего фактора роста-1β у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии Содержание ТФР-1β у крыс группы № 1 (интактные животные) составило 22,22 ± 7,67 пг/мл (табл. 4.11). У животных группы № 3 в плазме крови на 1 сутки после моделирования ИИ уровень ТФР-1β составил 53,20 ± 13,06 пг/мл, что достоверно (р ≤ 0,05) больше на 58,23 %, чем у животных группы № 1 и достоверно (р ≥ 0,05) не отличалось от содержания у животных группы № 2 (рис. 4.11). На 3 сутки в плазме крови животных из группы № 3 после моделирования ИИ содержание ТФР-1β составило 21,72 ± 8,26 пг/мл. По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) меньше на 59,17 %, что достоверно (р ≥ 0,05) не отличается от животных группы № 1. По сравнению с уровнем ТФР-1β в плазме крови у животных группы № 2 он достоверно (р ≤ 0,05) меньше на 65,50 % (табл. 4.11, рис. 4.11). 103 Таблица 4.11 Динамика уровня ТФР-1β с экспериментальным ишемическим инсультом у животных с предварительным использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ТФР-1β, М 22,22 53,20 21,72 19,25 30,17 ±m 7,67 13,06 8,26 5,90 9,05 < 0,05 > 0,05 > 0,05 > 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 р р* р** р*** < 0,05 Примечание: 1ТФР-1β — трансформирующий фактор роста β; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.11 — Изменения содержания ТФР-1β при экспериментальном ишемическом инсульте с предварительным использованием ТЭС-терапии В плазме крови животных группы № 3 на 7 сутки после моделирования ИИ уровень ТФР-1β составил 19,25 ± 5,90 пг/мл. По сравнению с 1 сутками он стал достоверно (р ≤ 0,05) меньше на 63,81 %. Содержание ТФР-1β в сравнении с 3 сутками и группой № 1 достоверно (р ≥ 0,05) не отличалось. По 104 сравнению с животными группы № 2 уровень ТФР-1β в плазме крови был достоверно (р ≤ 0,05) меньше на 70,96 % (табл. 4.11, рис. 4.11). На 14 сутки после моделирования ИИ в плазме крови животных группы № 3 уровень ТФР-1β составил 30,17 ± 9,05 пг/мл. В сравнении с 1 сутками он стал достоверно (р ≤ 0,05) меньше на 43,28 %. По отношению к 3 суткам достоверных (р ≥ 0,05) не было. По сравнению с 7 сутками он стал достоверно (р ≤ 0,05) больше на 36,19 %. Содержание ТФР-1β плазме крови животных группы № 1 стало достоверно (р ≤ 0,05) меньше на 26,35 %. У животных группы № 2 оно было достоверно (р ≤ 0,05) больше на 34,01 %. Таким образом, содержание ТФР-1β значительно возрастал через 1 сутки после моделирования ИИ. К 3 суткам оно падало до исходного уровня, оставаясь таковым до 7 суток. Однако к 14 суткам опять возрастало, превышая исходный уровень. 4.4. Динамика содержания β-эндорфина у крыс с экспериментальным ишемическим инсультом с предварительным использованием ТЭС-терапии Уровень β-эндорфина в плазме крови у крыс группы № 1 (интактные животные) составил 27,52 ± 6,14 пг/мл (табл. 4.12). В 1 сутки после моделирования ИИ в плазме крови животных группы № 3 содержание β-эндорфина составило 50,31 ± 9,67 пг/мл, что достоверно (р ≤ 0,05) больше на 45,3 %, чем в плазме крови животных группы № 1 и достоверно (р ≤ 0,05) больше на 72,98 % по сравнению с животными группы № 2 (рис. 4.12). В плазме крови животных группы № 3 на 3 сутки после моделирования ИИ уровень β-эндорфина составил 44,89 ± 5,47 пг/мл. По сравнению с 1 сутками достоверных (р ≥ 0,05) изменений не отмечено. По отношению к животным группы № 1 он достоверно (р ≤ 0,05) больше на 38,69 %. На 75,25 % он достоверно (р ≤ 0,05) больше у животных группы № 2 (табл. 4.12, рис. 4.12). 105 Таблица 4.12 Содержание β-эндорфина при моделировании ишемического инсульта у животных группы № 3 Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки β-эндорфина, М 27,51 50,31 44,89 44,10 43,69 ±m 6,13 9,66 5,47 6,99 6,03 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 р р* р** р*** > 0,05 Примечание: р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.12 — Изменения β-эндорфина при экспериментальном ишемическом инсульте с предварительным использованием ТЭС-терапии На 7 сутки после моделирования ИИ в плазме крови животных группы № 3 содержание β-эндорфина составило 44,11 ± 7,00 пг/мл. По сравнению с 1 и 3 сутками достоверных (р ≥ 0,05) отличий не было. У животных группы № 3 уровень β-эндорфина достоверно (р ≤ 0,05) больше на 37,61 %, чем в плазме крови из группы № 1. По сравнению с животными группы № 2 β-эндорфина достоверно (р ≤ 0,05) больше на 78,14 %. 106 В плазме крови животных группы № 3 на 14 сутки после моделирования ИИ содержание β-эндорфина составило 43,69 ± 6,04 пг/мл, достоверных (р ≤ 0,05) отличий по сравнению с 1, 3 и 7 сутками не было. По сравнению с животными группы № 1 уровень β-эндорфина был достоверно (р ≤ 0,05) больше на 37,01 %, и достоверно (р ≤ 0,05) больше на 77,95 % по сравнению с животными группы № 2. Таким образом, использование ТЭС-терапии до моделирования ИИ позволяет поддерживать содержание β-эндорфина в течение периода наблюдения (14 суток) на достаточном уровне, обеспечивающим цитокиновый гомеостаз. 4.5. Динамика содержания цитокинов в плазме крови крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии 4.5.1. Динамика содержания фактора некроза опухоли-α у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии У крыс группы № 1 (интактные животные) содержание ФНО-α составило 8,41 ± 2,53 пг/мл (табл. 4.13). На 1 сутки после моделирования ИИ в плазме крови животных группы № 4 уровень ФНО-α составил 8,33 ± 6,39 пг/мл, что достоверно (р ≥ 0,05) не отличалось от животных групп № 1 и № 3. Однако было достоверно (р ≤ 0,05) меньше на 53,25 %, чем в плазме крови животных группы № 2 (рис. 4.13). В плазме крови животных из группы № 4 на 3 сутки после моделирования ИИ уровень ФНО-α составил 8,96 ± 4,67 пг/мл. По сравнению с 1 и 3 сутками группы № 3 и № 1 достоверных изменений (р ≥ 0,05) не происходило. Однако по сравнению с животными группы № 2 он был достоверно (р ≤ 0,05) меньше на 51,19 %. На 7 сутки после моделирования ИИ в плазме крови животных из группы № 4 содержание ФНО-α составило 8,01 ± 3,21 пг/мл. По сравнению с 1 и 3 сутками группы № 4, 7 сутками группы № 3 и группой № 1 достоверных 107 (р ≥ 0,05) изменений не отмечено. У животных группы № 4 на 7 сутки по сравнению с группой № 2 уровень ФНО-α был достоверно (р ≤ 0,05) меньше на 55,05 % (табл. 4.13, рис. 4.13). Таблица 4.13 Динамика уровня ФНО-α при экспериментальном ишемическом инсульте у животных с последующим использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ФНО-α, М 8,41 8,33 8,96 8,01 8,87 ±m 2,53 6,39 4,67 3,21 7,61 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 р р* р** р*** > 0,05 Примечание: ФНО-α — фактор некроза опухоли-α; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.13 — Изменения содержания ФНО-α при экспериментальном ишемическом инсульте с последующим использованием ТЭС-терапии 108 В плазме крови животных из группы № 4 на 14 сутки после моделирования ИИ уровень ФНО-α составил 8,87 ± 7,61 пг/мл. По сравнению с 1, 3 и 7 сутками группы № 4, 14 сутками группы № 3 и группы № 1 он достоверно (р ≥ 0,05) не изменился. У животных группы № 4 по сравнению с группой № 2 уровень ФНО-α на 14 сутки был достоверно (р ≤ 0,05) меньше на 57,78 %. Итак, содержание ФНО-α после моделирования ИИ, благодаря использованию ТЭС-терапии, не отличалось от контрольной величины и не изменялось в течение всего периода наблюдения. 4.5.2. Динамика содержания интерлейкина-1β у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии В плазмы крови крыс группы № 1 (интактные животные) содержание ИЛ-1β составило 30,93 ± 7,90 пг/мл (табл. 4.14). На 1 сутки после моделирования ИИ содержание ИЛ-1β в плазме крови животных группы № 4 составило 220,85 ± 73,59 пг/мл. Это достоверно (р ≤ 0,05) выше на 85,99 %, чем в плазме крови животных группы № 1. У животных группы № 2 содержание ИЛ-1β достоверно (р ≤ 0,05) меньше на 52,46 %, а группы № 3 достоверно (р≥0,05) не отличается от группы № 4 (рис. 4.14). В плазме крови животных группы № 4 на 3 сутки после моделирования ИИ уровень ИЛ-1β составил 211,58 ± 73,71 пг/мл и достоверно (р ≥ 0,05) не отличался от уровня, наблюдаемого в 1 сутки. Это достоверно (р ≤ 0,05) больше на 85.38 %, чем в плазме крови животных группы № 1. Содержание ИЛ-1β у животных группы № 2 достоверно (р ≤ 0,05) больше на 88,23 %. Уровень ИЛ-1β у животных группы № 4 достоверно (р ≥ 0,05) не отличается от животных группы № 3. К 7 суткам после моделирования ИИ содержание ИЛ-1β составило 70,56 ± 10,09 пг/мл. По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) меньше на 68,05 %. К 3 суткам уровень ИЛ-1β стал достоверно 109 (р ≤ 0,05) меньше на 66,65 %. У животных группы № 1 он был достоверно (р ≤ 0,05) меньше на 56.16 %. По сравнению с животными группы № 2 содержание ИЛ-1β было достоверно (р ≤ 0,05) меньше на 88,67 % и на 58,89 % достоверно (р ≤ 0,05) меньше, чем у животных группы № 3 (табл. 4.14, рис 4.14). На 14 сутки после моделирования ИИ уровень ИЛ-1β в плазме крови животных из группы № 4 составил 46,66 ± 15,56 пг/мл, что по сравнению с 1 сутками достоверно (р ≤ 0,05) меньше на 78,87 %. По сравнению с 3 сутками он стал достоверно (р ≤ 0,05) меньше на 77,94 %. По сравнению с 7 сутками уровень ИЛ-1β стал достоверно (р ≤ 0,05) меньше на 33,87 %. Содержание ИЛ-1β у животных группы № 1 было достоверно (р ≤ 0,05) меньше на 33,71 %. У животных группы № 2 — достоверно (р ≤ 0,05) больше на 88,55 %, а группы № 3 достоверно (р ≤ 0,05) больше на 61,20 % (табл. 4.14, рис 4.14). Таблица 4.14 Динамика уровня ИЛ-1β при экспериментальном ишемическом инсульте у животных с последующим использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-1β, М 30,93 220,85 211,58 70,56 46,66 ±m 7,90 73,59 73,71 10,09 15,56 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 < 0,05 < 0,05 < 0,05 < 0,05 р р* р** р*** < 0,05 Примечание: ИЛ-1β — интерлейкин-1β; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. 110 Рисунок 4.14. Изменения содержания ИЛ-1β при экспериментальном ишемическом инсульте с последующим использованием ТЭС-терапии Таким образом, содержание ИЛ-1β после моделирования ИИ на фоне проводимой ТЭС-терапии в 1 сутки повышался менее значительно. Затем к 14 суткам падал почти до исходной величины. 4.5.3. Динамика содержания интерлейкина-6 у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии У крыс группы № 1 (интактные животные) уровень ИЛ-6 составил 0,43 ± 0,16 пг/мл (табл. 4.15). В 1 сутки после моделирования ишемического инсульта уровень ИЛ-6 составил 1,08 ± 0,31 пг/мл, что достоверно (р ≤ 0,05) больше на 60,18 %, чем в плазме крови животных группы № 1. Он был достоверно (р ≤ 0,05) меньше на 84,27 % по сравнению с животными группы № 2. Содержание ИЛ-6 у животных группы № 3 достоверно (р ≤ 0,05) меньше на 69,40 % (рис. 4.15). В плазме крови животных группы № 4 на 3 сутки после моделирования ИИ содержание ИЛ-6 составило 1,62 ± 0,38 пг/мл. 111 Таблица 4.15 Динамика уровня ИЛ-6 при экспериментальном ишемическом инсульте у животных с последующим использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-6, М 0,43 1,08 1,62 0,77 0,41 ±m 0,16 0,31 0,38 0,25 0,17 < 0,05 < 0,05 < 0,05 > 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 р р* р** р*** < 0,05 Примечание: ИЛ-6 — интерлейкин-6; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.15 — Изменения содержания ИЛ-6 при экспериментальном ишемическом инсульте с последующим использованием ТЭС-терапии По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) больше на 33,3 %. Это достоверно (р ≤ 0,05) больше на 73,45 %, чем в плазме крови из группы № 1. Уровень ИЛ-6 у животных группы № 4 был достоверно (р ≤ 0,05) меньше на 84,37 % по сравнению с животными группы № 2 и до- 112 стоверно (р ≤ 0,05) меньше на 70,49 %, чем в группе № 3 (табл. 4. 15, рис. 4.15). На 7 сутки после моделирования ИИ в плазме крови животных группы № 4 уровень ИЛ-6 составил 0,77 ± 0,25 пг/мл, что достоверно (р ≤ 0,05) меньше на 28,70 % по сравнению с 1 сутками. По сравнению с 3 сутками он стал достоверно (р ≤ 0,05) меньше на 52,46 %. У животных группы № 1 уровень ИЛ-6 был достоверно (р ≤ 0,05) меньше на 44,15 % и достоверно (р ≤ 0,05) меньше на 93,31 %, чем в группе № 2. Достоверных (р ≥ 0,05) отличий по сравнению с группой № 3 не выявлено (табл. 4.15, рис. 4.15). Содержание ИЛ-6 в плазме крови животных группы № 4 на 14 сутки после моделирования ИИ составило 0,41 ± 0,17 пг/мл. По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) меньше на 62,03 %, — с 3 сутками достоверно (р ≤ 0,05) меньше на 74,69 %. На 7 сутки оно стало достоверно (р ≤ 0,05) меньше на 46,75 %. По сравнению с животными группы № 1 достоверных (р ≥ 0,05) отличий нет. У животных группы №4 по сравнению с группой №3 уровень ИЛ-6 стал (р ≤ 0,05) достоверно меньше на 39,70 %. Применение ТЭС-терапии после моделирования ИИ ограничивало рост уровня ИЛ-6 на 1 и 3 сутки. К 14 суткам содержание ИЛ-6 достоверно (р ≥ 0,05) по сравнению с группой № 1 не отличалось. 4.5.4. Динамика содержания интерлейкина-10 у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии У крыс группы № 1 (интактные животные) содержание ИЛ-10 составило 1,21 ± 0,66 пг/мл (табл. 4.16). На 1 сутки после моделирования ИИ уровень ИЛ-10 в плазме крови животных группы № 4 составил 68,22 ± 9,43 пг/мл. Это достоверно (р ≤ 0,05) больше на 98,22 %, чем у животных группы № 1. По сравнению с животными группы № 2 содержание ИЛ-10 достоверно (р ≤ 0,05) больше на 41,67 %. У животных группы № 3 уровень ИЛ-10 достоверно (р ≤ 0,05) меньше на 27,35 % (рис. 4.16). 113 Таблица 4.16 Динамика уровня ИЛ-10 при экспериментальном ишемическом инсульте у животных с последующим использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ИЛ-10, М 1,21 68,22 101,45 92,40 72,48 ±m 0,66 9,43 17,40 9,47 9,47 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 < 0,05 р р* р** р*** < 0,05 Примечание: ИЛ-10 — интерлейкин-10; р — достоверность различия по отношению исходному показателю; р*- — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.16 — Изменения содержания ИЛ-10 при экспериментальном ишемическом инсульте с последующим использованием ТЭС-терапии В плазме крови животных группы № 4 на 3 сутки после моделирования ИИ содержание ИЛ-10 составило 101,45 ± 17,40 пг/мл. По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) больше на 32,75 %. Это достоверно (р ≤ 0,05) превысило на 98,80 % уровень ИЛ-10 в плазме крови животных 114 группы № 1. По сравнению с животными группы № 2 он достоверно (р ≤ 0,05) меньше на 50,86 %. У животных группы № 3 уровень ИЛ-10 на 16,88 % достоверно (р ≤ 0,05) меньше (табл. 4.16, рис. 4.16). На 7 сутки после моделирования ИИ в плазме крови животных группы № 4 уровень ИЛ-10 составил 92,40 ± 9,47 пг/мл. По сравнению с 1 сутками он стал на 26,16 % достоверно (р ≤ 0,05) больше, по сравнению с 3 сутками — достоверно (р ≥ 0,05) не изменился. У животных группы № 1 уровень ИЛ-10 достоверно (р ≤ 0,05) меньше на 98,69 %. В плазме крови животных группы № 2 содержание ИЛ-10 (р ≥ 0,05) достоверно не отличалось, также как (р ≥ 0,05) по сравнению с животными группы № 3. К 14 суткам после моделирования ИИ в плазме крови животных из группы № 4 содержание ИЛ-10 составило 72,48 ± 9,47 пг/мл. Достоверных (р ≥ 0,05) отличий по сравнению с 1 сутками не отмечено. По сравнению с 3 сутками уровень ИЛ-10 стал достоверно (р ≤ 0,05) меньше на 28,55 %, с 7 сутками — на 21,55 %. По сравнению с содержанием ИЛ-10 в плазме крови животных группы № 1 оно стало достоверно (р ≤ 0,05) больше на 98,33 %. Достоверных (р ≥ 0,05) изменений по сравнению с животными группы № 3 не выявлено. На фоне проведения ТЭС-терапии отмечен более высокий уровень ИЛ10 в 1 сутки. Содержание ИЛ-10 достигало максимума к 3 суткам. Затем происходило снижение уровня ИЛ-10, и к 14 суткам он возвращался таковому, наблюдаемому в 1 сутки. 4.5.5. Динамика содержания трансформирующего фактора роста-1β у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии У группы № 1 (интактные животные) содержание ТФР-1β составило 22,22 ± 7,67 пг/мл (табл. 3.17). В плазме крови животных из группы № 4 на 1 сутки после моделирования ИИ уровень ТФР-1β составил 46,41 ± 8,16 пг/мл, что достоверно 115 (р ≤ 0,05) больше на 52,12 %, чем в плазме крови животных группы № 1 и достоверно (р ≥ 0,05) не отличается от животных группы № 2 и группы № 3 (рис. 4.17). Таблица 4.17 Динамика уровня ТФР-1β с экспериментальным ишемическим инсультом у животных с последующим использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки ТФР-1β, М 22,22 46,41 33,19 36,31 56,36 ±m 7,67 8,16 6,28 7,76 9,33 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 < 0,05 р р* р** р*** < 0,05 Примечание: ТФР-1β — трансформирующий фактор роста β; р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. Рисунок 4.17 — Изменения содержания ТФР-1β при экспериментальном ишемическом инсульте с последующим использованием ТЭС-терапии 116 На 3 сутки после моделирования ИИ в плазме крови животных из группы № 4 содержание ТФР-1β составило 33,19 ± 6,28 пг/мл. По сравнению с 1 сутками оно стало достоверно (р ≤ 0,05) меньше на 28,48 %. В плазме крови животных группы № 1 оно было достоверно (р ≤ 0,05) меньше на 48,55 %. По сравнению содержанием ТФР-1β в плазме крови животных группы № 2 достоверно (р ≤ 0,05) меньше на 47,29 %. У животных группы № 3 уровень ТФР-1β достоверно (р ≤ 0,05) больше на 34,55 % (табл. 4.17, рис. 4.17). В плазме крови животных из группы № 4 на 7 сутки после моделирования ИИ уровень ТФР-1β составил 36,31 ± 7,76 пг/мл. По сравнению с 1 сутками он стал достоверно (р ≤ 0,05) больше на 21,76 %, а по сравнению с 3 сутками он достоверно (р ≥ 0,05) не изменился. В плазме крови из группы № 1 содержание ТФР-1β было достоверно (р ≤ 0,05) меньше на 38,80 %. У животных группы № 2 оно было достоверно (р≤0,05) больше на 45,23 %, а в группе № 3 достоверно (р ≤ 0,05) меньше на 46,98 %. К 14 суткам после моделирования ИИ уровень ТФР-1β в плазме крови животных из группы № 4 составил 56,36 ± 9,33 пг/мл. По сравнению с 1 сутками он стал достоверно (р ≤ 0,05) больше на 17,65 %, а по сравнению с 3 сутками — достоверно (р ≤ 0,05) больше на 41,11 %. К 7 сутками уровень ТФР-1β достоверно (р ≤ 0,05) вырос на 35,57 %. В плазме крови из группы № 1 его было достоверно больше (р ≤ 0,05) на 60,57 %. По сравнению с животными группы № 3 уровень ТФР-1β был достоверно (р ≤ 0,05) больше на 46,46 %. Следовательно, ТЭС-терапия, проводимая сразу после моделирования ИИ, ограничивала рост содержания ТФР-1β на 3 и 7 сутки. Однако к 14 суткам он превышал уровень, наблюдаемый у животных группы № 2. 117 4.6. Динамика содержания β-эндорфина у крыс с экспериментальным ишемическим инсультом с последующим использованием ТЭС-терапии В плазме крови крыс группы № 1 (интактные животные) уровень β-эндорфина составил 27,52 ± 6,14 пг/мл (табл. 4.18). Таблица 4.18 Содержание β-эндорфина при моделировании ишемического инсульта у животных группы с последующим использованием ТЭС-терапии Показатель Контроль 1 сутки 3 сутки 7 сутки 14 сутки β-эндорфина, М 27,51 48,67 42,42 42,63 42,20 ±m 6,13 15,28 11,59 9,29 9,93 < 0,05 < 0,05 < 0,05 < 0,05 > 0,05 > 0,05 > 0,05 > 0,05 > 0,05 р р* р** р*** > 0,05 Примечание: р — достоверность различия по отношению исходному показателю; р* — достоверность различия по отношению показателя 1 суток после моделирования ишемического инсульта; р** — достоверность различия по отношению показателя 3 суток после моделирования ишемического инсульта; р*** — достоверность различия по отношению показателя 7 суток после моделирования ишемического инсульта. В плазме крови животных группы № 4 на 1 сутки после моделирования ИИ содержание β-эндорфина составило 48,68 ± 15,28 пг/мл, что достоверно (р ≤ 0,05) больше на 43,46 %, чем в плазме крови животных группы № 1 и достоверно (р ≤ 0,05) больше на 71,34 % по сравнению с животными группы № 2. Однако достоверно (р ≥ 0,05) не отличается от уровня β-эндорфина у животных группы № 3 (рис. 4.18). На 3 сутки после моделирования ИИ в плазме крови животных группы № 4 уровень β-эндорфина составил 42,42 ± 11,59 пг/мл. По сравнению с 1 сутками он достоверно (р ≥ 0,05) не изменился. Однако был достоверно 118 (р ≤ 0,05) больше на 35,12 %, чем в плазме крови животных группы № 1 достоверно (р ≤ 0,05) больше на 73,81 %, чем в группе № 2. По сравнению с животными группы № 3 содержание β-эндорфина и достоверно (р ≥ 0,05) не отличалось. Рисунок 4.18 — Изменения β-эндорфина при экспериментальном ишемическом инсульте с последующим использованием ТЭС-терапии К 7 суткам после моделирования ИИ в плазме крови животных группы № 4 содержание β-эндорфина составило 42,64 ± 9,29 пг/мл. По сравнению с 1 и 3 сутками достоверных (р ≥ 0,05) изменений не было. Оно оставалось достоверно (р ≤ 0,05) больше на 35,46 %, чем в плазме крови животных группы № 1 и достоверно (р ≤ 0,05) больше на 77,39 % по сравнению с группой № 2. От содержания у животных группы № 3 достоверно (р ≥ 0,05) не отличалось. На 14 сутки после моделирования ИИ в плазме крови животных из группы № 4 уровень β-эндорфина составил 42,21 ± 9,93 пг/мл. по сравнению с 1, 3 и 7 сутками достоверных (р ≥ 0,05) изменений не отмечено. Оно также оставалось достоверно (р ≤ 0,05) больше на 34,80 %, чем в плазме крови животных группы № 1 и достоверно (р ≤ 0,05) больше на 77,18 % животных группы № 2. Достоверных (р ≥ 0,05) отличий с животными группы № 3 не было. 119 Таким образом, использование ТЭС-терапии после моделирования ИИ позволяет поддерживать содержание β-эндорфина в течение периода наблюдения (14 суток) на достаточном уровне, ограничивающем активность воспалительного процесса. Итак, применение ТЭС-терапии до или после моделирования ИИ уменьшает нарушения цитокинового гомеостаза, приводящие к выраженной активации системного воспалительного ответа, что ограничивает повреждение ткани мозга в очаге ишемии. 120 ГЛАВА 5. ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ Воспаление играет ключевую роль в патогенезе ИИ [K.J. Becker, 2001; G.J. del Zoppo, K.J. Becker, J.M. Hallenbeck, 2001; M.D. Ginsberg, 2003; M. Fisher, R. Ratan, 2003; P.L. Wood, 2003; R. Markus, D.C. Reutens, S. Kazui, 2004; Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; Q. Wang, X.N. Tang, M.A. Yenari, 2007; А.Х. Хана-Мурад, Л.И. Павлинова, А.А. Мокрушин, 2007; A. Tuttolomondo, D. Di Raimondo, R. Di Sciacca, 2008; J. Jordan et al., 2008; D. Amantea et al., 2008; C. Benakis, L. Hirt, R.A. Du Pasquier, 2009; Ю.Д. Губарев, А.О. Шеремет, 2009; А.С. Кость, Б.Д. Луцик, Л.Е. Лаповець, 2010; A.G. Ceulemans et al., 2010; Y. Lee et al., 2014; М.В. Онуфриев и соавт., 2014]. Известно, что развитие воспалительной реакции в очаге ишемии связано с изменением концентрации провоспалительных (ИЛ-1, ФНО-α, ИЛ-6) и противовоспалительных цитокинов (ИЛ-4, ИЛ-10, ТФР-1β) [Е.И. Гусев, В.И. Скворцова, 2001; N. Quan, M. Herkenham, 2002; А.А. Аракелян и соавт., 2005; Л.Н. Кашаева, Л.М. Карзакова, В.Н. Саперов, 2005; Q. Wang, X.N. Tang, M.A. Yenari, 2007; D. Amantea et al., 2008; M.Y. Zuidema, C. Zhang, 2010; М.В. Онуфриев, 2014]. Запуск продукции цитокинов, регуляторов развития воспалительной реакции, происходит в результате гиперактивации поврежденных нейронов, микроглиоцитов и астроцитов. Кроме того, важным источником становятся активированные лейкоциты, которые проникают через поврежденный ГЭБ [Y.D. Wen, H.L. Zhang, Z.H. Qin, 2006; R.M. Adibhatla, J.F. Hatcher, 2007; С.М. Белоцкий, Р.Р. Авталион, 2008; H.K. Jeong et al., 2010; Y. Huang et al., 2011]. При моделировании ИИ уровень цитокинов повышается (группа животных № 2), по отдельным показателям более чем сто раз, что является основой развития ССВО. ССВО воспалительный ответ на уровне целостного организма, в том числе при состояниях без определяемой бактериемии [F. Hietbrink et al., 2006; С.М. Белоцкий, Р.Р. Авталион, 2008; J.M. Sinning et al., 2012; 121 В.В. Иващенко и соавт., 2012]. При ИИ наблюдается переход воспалительного ответа на системный уровень, что проявляется в виде развития ССВО, который значительно отягощает течение заболевания и играет важную роль в развитии и прогрессировании экстрацеребральной патологии [D. Stanimirovic, K. Saton, 2000; M. Di Napoli, F. Papa, 2006; B.W. Mc Coll, N.J. Rothwell, S.M. Allan, 2007]. Нарушение регуляторных механизмов, контролирующих развитие воспалительного процесса, ответственно за негативные постишемические эффекты и переход воспалительного ответа с локального уровня на системный [P.E. Marik, G.P. Zaloga, 2001; А.А. Аракелян и соавт., 2005; M. Di Napoli, F. Papa, 2006; B.W. Mc Coll, N.J. Rothwell, S.M. Allan, 2007; С.М. Белоцкий, Р.Р. Авталион, 2008;]. Индуцируемый ишемией воспалительный ответ организма обусловливает нарушение целостности ГЭБ при инсульте. Возникающие нарушения ГЭБ могут, в свою очередь, способствовать увеличению притока иммуномодуляторов в ЦНС и усиливать воспалительный ответ [D. Stanimirovic, K. Saton, 2000; K.J. Becker, 2001; M. Di Napoli, F. Papa, 2006]. Главная роль в развитии патологических изменений при развитии ССВО отводится провоспалительным цитокинам [H. Hirasawa et al., 2004; M.F. Osuchowski et al., 2006; F. Hietbrink et al., 2006; С.М. Белоцкий, Р.Р. Авталион, 2008; D.H. Yu, B. Kim, J. Park, 2011]. Важнейшим провоспалительным цитокином является ФНО-α, который в больших концентрациях оказывает токсической действие на нейроны путем активации системы генерации АФК, что резко индуцирует процессы ПОЛ, в том числе, за счет взаимодействия с сигнальной системой сфингомиелинового цикла [J.J. Viel, D.Q. Mc Manus, S.S. Smith, 2001; A. Quintana et al., 2005; А.Е. Бугрова, 2007; R.M. Adibhatla, J.F. Hatcher, 2007; R.M. Adibhatla, R. Dempsey, J.F. Hatcher, 2008; N. Liu et al., 2009; А.В. Алексеенко, 2013]. Однако ФНО-α, помимо токсического влияния, в низкой (физиологической) концентрации оказывает защитный эффект на нейроны, путем увеличения экспрессии факторов антиоксидантной системы [J.J. Viel, D.Q. Mc Manus, S.S. Smith, 2001; A. Quintana et al., 2005]. 122 После моделирования ИИ уровень ФНО-α у животных группы № 2 возрастал и оставался достоверно (р ≤ 0,05) стойко повышенным (более чем на 50 %) без существенной динамики с 1 до 14 суток, что объясняет большую активность процессов свободно-радикального окисления в данной группе крыс [В.Д. Левичкин и соавт., 2013]. Увеличение концентрации эндогенного ФНО-α вследствие асептического воспаления в области инфаркта мозга, за счет активации сфингомиелиназы приводит к росту продукции церамида, также ФНО-α индуцирует образование АФК в клетках, в результате чего развивается активация реакций СРО, в том числе ПОЛ, дефицит антиоксидантов (в том числе глутатиона), что в свою очередь дополнительно повышает уровень активности сфингомиелиназы приводя к образованию новых порций церамида и ФНО-α [А.Е. Бугрова, 2007; А.В. Алексеенко, 2013]. ИЛ-1β является наряду с ФНО-α ключевыми медиатором микроглиальных нейроиммунных функций. В ответ на развитие церебральной ишемии его уровень возрастает наиболее сильно начиная с 1 суток с пиком на 3 сутки после моделирования ИИ. Это объясняется тем, что синтез ИЛ-1β микроглией является главным активирующим сигналом для запуска образования других провоспалительных цитокинов, а также стимуляции астроцитов к продукции потенциальных нейротоксичных веществ, таких как NO и метаболиты арахидоновой кислоты [H. Boutin et al., 2001; L.D. Church, P.G. Cook, M.F. Mc Dermott, 2008; S. Choi, W.J. Friedman, 2009; C.A. Dinarello, 2009; C.A. Dinarello, 2011; H.F. Green et al., 2012; J. Galea, D. Brough, 2013; C. Garlanda, C.A. Dinarello, A. Mantovani, 2013]. У крыс группы № 2 на 1 сутки после моделирования ИИ содержание ИЛ-1β в плазме крови достоверно (р ≤ 0,05) выросло на 93,34 % по сравнению с интактными животными. К 3 суткам уровень ИЛ-1β достоверно (р ≤ 0,05) увеличился еще на 74,17 %. Только на 7 сутки после моделирования ИИ содержание ИЛ-1β достоверно (р ≤ 0,05) снизилось на 65,36 % по сравнению с 3 сутками почти до уровня 1 суток (достоверно (р ≤ 0,05) больше на 25,43 %). 123 На 14 сутки после моделирования ИИ содержание ИЛ-1β в плазме крови животных группы № 2 вернулось к уровню 1 суток. Однако оставалось достоверно (р ≤ 0,05) выше на 93,34 % по сравнению с интактными животными. ИЛ-6, содержание которого исследовали в плазме крови крыс, является типичным ранним индуцибельным цитокином и синтезируется многими типами клеток, участвующими в инициации и регуляции воспаления и иммунного ответа [D.A. Papanicolaou et al., 1998; С.А. Кетлинский, А.С. Симбирцев, 2008]. ИЛ-6 оказывает выраженное провоспалительное действие за счет активации экспрессии молекул адгезии и активации хемотаксиса лейкоцитов, а также является важнейшим индуктором острофазного ответа [K. Paul-Pletzer, 2006; А.С. Симбирцев, 2008; I.M. Cojocaru et al., 2009; С.А. Кетлинский, T. Tanaka et al., 2013]. Особенностью для животных группы № 2 являлось стойкое значительное повышение уровня ИЛ-6, что отражает активное течение асептического воспаления в ответ на развитие церебральной ишемии. Известно, что ИЛ-1 и ФНО-α являются мощными индукторами ИЛ-6-синтетазы [С.А. Кетлинский, А.С. Симбирцев, 2008; H. Boutin et al., 2001]. При этом важно отметить, что ФНО-α, ИЛ-1, ИЛ-6 и некоторые другие цитокины регулируют функцию гипоталамо-гипофизарно-надпочечниковой системы (ГГНС), непосредственно воздействуя на переднюю долю гипофиза, что объясняет включение стресс-реализующих систем в ответ на ишемию мозга в рамках общей реакции нейроиммуноэндокринной системы [A. Johansson et al., 2000; N. Quan, M. Herkenham, 2002; F. Eskandari, J.I. Webster, M.S. Esther, 2003; И.Г. Акмаев, В.В. Гриневич, 2003; С.А. Кетлинский, А.С. Симбирцев, 2008]. Отмечена прямо пропорциональная зависимость между уровнем провоспалительных цитокинов (ИЛ-1, ФНО-α, ИЛ-6, ИЛ-8) при развитии острой локальной церебральной ишемии и конечными размерами инфаркта мозга (зона некроза) [Е.И. Гусев, В.И. Скворцова, 2001; H. Boutin et al., 2001; K.Y. Kwan, B.H. Jeon, 2001; N. Quan, M. Herkenham, 2002; Q. Wang, X.N. Tang, 124 M.A. Yenari, 2007; D. Amantea et al., 2008; I.M. Cojocaru et al., 2009; M. Zhang et al., 2011; М.В. Онуфриев, 2014]. Эту зависимость легко проследить сравнив между собой уровни провоспалительных цитокинов (ИЛ-1, ФНО-α, ИЛ-6) на 1 и 3 сутки после воссоздания модели ИИ с размерами очага инфаркта мозга, что и наблюдалось в группах № 2, № 3 и № 4 [А.И. Трофименко и соавт., 2014]. Важное значение при развитии ИИ имеет изменение содержания противовоспалительных цитокинов ИЛ-10 и ТФР-1β, что определяется их биологическим действием, которое направлено на подавление воспалительной реакции в очаге ишемии ГМ [K. Strle et al., 2001; Y. Zhu et al., 2002; N. Quan, M. Herkenham, 2002; N. Vila et al., 2003; Г.Н. Жданов, 2006; M. Kaste, H.J. Castillo, S. de Compostela, 2006; N. Liu et al., 2009]. Прогрессирование повреждения зоны пенумбры в области острой фокальной ишемии мозга может происходить не только под влиянием абсолютного увеличения концентрации провоспалительных агентов, но и вследствие недостатка противовоспалительных и нейротрофических факторов [K. Strle et al., 2001; C. Alzheimer, 2002; N. Vila et al., 2003]. Трофические факторы наряду с аутокринной стимуляцией собственных рецепторов индуцируют дальнейшие сигнальные каскады, приводящие к синтезу регуляторных протеинов, которые предотвращают индукцию апоптоза и способствуют клеточному жизнеобеспечению. Защитное противоишемическое действие ТФР-1β проявляется угнетением адгезии лейкоцитов к эндотелию, подавлением выработки МФ повреждающих кислородных и NO метаболитов [J.J. Letterio, 2000, N. Vila et al., 2003]. Нейропротективная способность ТФР-1β связана со стабилизацией нейронального кальциевого гомеостаза и повышением экспрессии гена bcl-2, что приводит к значительному торможению свободнорадикального повреждения, замедлению процессов некроза и апоптоза [J.J. Letterio, 2000; Y. Zhu et al., 2002]. Установлена важная роль ТФР-1β, в увеличении васкуляризации ткани мозга после острой церебральной ишемии, особенно зоны пенумбры [J.J. Letterio, 2000]. 125 На 1 сутки после моделирования ИИ уровень ТФР-1β в плазме крови животных группы № 2 был достоверно (р ≤ 0,05) больше на 48,74 %, чем у интактных животных. К 3 суткам достигал максимума (больше на 64,7 %). Оставался на этом уровне и на 7 сутки. Затем к 14 суткам снижался до уровня 1 суток. Однако он превышал содержание у интактных животных на 51,39 %. ИЛ-10 является мощным регулятором клеточно-опосредованного иммунного ответа моноцитов/МФ: снижает продукцию ПГЕ2, угнетает экспрессию ICAM-1 активно участвующих в развитии местного воспаления, подавляет выработку многочисленных провоспалительных цитокинов, включая ИЛ-1, ФНО-α, ИЛ-6, ИЛ-8 [С.А. Кетлинский, А.С. Симбирцев, 2008; N. Liu et al., 2009; T. Nabe et al., 2012]. Он также подавляет синтез супероксидного аниона и реактивных промежуточных продуктов обмена О2, модулирует синтез NO в зависимости от продолжительности воздействия на активированные макрофаги. Содействие синтезу NO является одним из многочисленных активирующих эффектов ИЛ-10 на моноциты/МФ, наряду с индукцией им выработки антагониста рецептора для ИЛ-1. Таким образом, ИЛ-10 является фактором, модулирующим функцию МФ и обладающим мощной противовоспалительной активностью. В плазме крови животных группы № 2 на 1 сутки после моделирования ИИ уровень ИЛ-10 был достоверно больше (р ≤ 0,05) на 96,95 %, чем у интактных животных. На 3 сутки рост содержания цитокина продолжался. Он достигал максимума на 7 сутки, когда уровень ИЛ-10 стал достоверно (р ≤ 0,05) больше на 98,46 %, чем в плазме крови из группы № 1. К 14 суткам содержание ИЛ-10 вернулось к уровню 1 суток. Таким образом рост уровня важнейшего противовоспалительного цитокина, который происходил на фоне высокого содержания провоспалительных цитокинов (ФНО-α, ИЛ-1β, ИЛ-6), с одной стороны отражал высокую активность воспалительного процесса в ткани мозга и бóльшие размеры инфаркта мозга, с другой стороны — явную дисрегуляцию при развитии воспалительной реакции. 126 При изучении срезов ткани мозга крыс группы № 2, сделанных на 1 сутки после моделирования ИИ обнаружено формирование выраженной зоны инфаркта мозга, локализованной преимущественно в неокортексе с захватом небольшого участка каудопутамена, расположенного в зоне кровоснабжения правой средней мозговой артерии. Это хорошо согласуется с данными Y. Wang-Fischer, 2009. При моделировании ИИ при изучении срезов нативной ткани мозга окрашенных тетразолиевым методом, можно было оценить размеры и расположение очага инфаркта мозга [А.И. Трофименко и соавт., 2012; А.И. Трофименко и соавт., 2013]. Использовали показатель «удельный объем инфаркта мозга», который определяли оценки соотношения объема очага инфаркта мозга к объему всего мозга [Г.Г. Автандилов, 1990]. Определение показателя «удельный объем инфаркта мозга» позволило наблюдать динамику изменения размеров очага инфаркта мозга на 1 и 3 сутки после воссоздания модели. Показатель «удельного объема очага инфаркта мозга» у крыс группы № 2 на 1 сутки составил 0,108 ± 0,003 у.е., на 3 сутки он достоверно (р ≤ 0,05) возрос на 18,3 % и составил 0,132 ± 0,010 у.е. Согласно данным литературы именно к этому сроку завершается изменение размеров очага инфаркта мозга [S.H. Graham, J. Chen, 2001; M.P. Mattson et al., 2001; N.W. Manning et al., 2014]. Морфологическая картина к 3 суткам после моделирования ИИ характеризовалась формированием очага деструкции. В нем выявлялось большое количество погибших нейронов. В сосудах по периферии очага ишемии наблюдались признаки стаза крови. Возникала значительная воспалительная клеточная инфильтрация. К 14 суткам по периферии очага ишемии появлялись новые капилляры, которые прорастали в зону деструкции, формировался избыточный глиомезодермальный рубец. Указанные процессы происходили на фоне значительного роста содержания провоспалительных цитокинов (ФНО-α, ИЛ-1β, ИЛ-6), которое оставалось высоким даже на 14 сутки. Одновременно имело место повышение 127 уровня противовоспалительных цитокинов (ТФР-1β и ИЛ-10), которое достигало максимума к 7 суткам и затем падало к 14 суткам, хотя и оставалось достаточно высоким. Все изменения происходили на фоне постоянно снижающегося уровня βэндорфина, который к 14 суткам стал достоверно (р ≤ 0,05) меньше на 65,0 %, чем у интактных животных (рис. 5.1). Таким образом, высокий уровень провоспалительных цитокинов и снижение содержания β-эндорфина в сочетании с гиперпродукцией противовоспалительных цитокинов обусловливает значительное повреждение ткани мозга в зоне ишемии. Опиоидные пептиды, которые выделяются в достаточном количестве при ТЭС-терапии, потенциально способны ограничивать глутаматную эксайтотоксичность, а также благодаря открытию АТФ-зависимых калиевых каналов, запускать реакции ишемического пре- и посткондиционирования, что в свою очередь способно вызвать нейропротективный эффект в 1 сутки после развития острой локальной церебральной ишемии [J.M. Cidday, 2006]. Рисунок 5.1 — Изменение содержания β-эндорфина при моделировании ишемического инсульта без и с применением ТЭС-терапии Применение ТЭС-терапии влияло на динамику содержания ФНО-α у животных групп № 3 и № 4, оно имело выраженные отличия от животных 128 группы № 2. У животных группы № 2 содержание ФНО-α было достоверно (р ≤ 0,05) больше на 51,01 %. Уровень ФНО-α у животных групп № 3 и № 4 не имел достоверных отличий между собой и животными группы № 1 (рис. 5.2). Рисунок 5.2 — Изменение содержания ФНО-α при моделировании ишемического инсульта без и с применением ТЭС-терапии В плазме крови животных группы № 3 на 1 сутки после моделирования ИИ содержание ИЛ-1β было достоверно (р ≤ 0,05) меньше на 49,36 %, чем у животных группы № 2. Одним из ключевых отличий динамики уровня ИЛ-1β у животных группы № 3 от животных группы № 2, наряду со значительно более низким уровнем, является сдвиг пика содержания ИЛ-1β в плазме крови с 3 на 1 сутки, постепенным снижением концентрации к 14 суткам. Однако несмотря на это оно оставалось достоверно (р ≤ 0,05) больше на 74,28 %, чем у интактных животных (рис. 5.3). Схожие изменения уровня ИЛ-1β отмечены и у животных группы № 4. Отличием было только то, что к 14 суткам содержание ИЛ-1β приближалось к нормальным значениям, достоверно (р ≤ 0,05) превышая их на 33,71 % (рис. 5.2). 129 Рисунок 5.3 — Изменение содержания ИЛ-1β при моделировании ишемического инсульта без и с применением ТЭС-терапии Таким образом, проведение ТЭС-терапии после моделирования ИИ обладает более выраженным гомеостатическим действием на уровень ИЛ-1β по сравнению с предшествующим ишемии применением ТЭС-терапии. В остальном динамика изменения содержания ИЛ-1β в группах № 3 и № 4 имела общие черты, отличаясь от динамики уровня ИЛ-1β у животных группы № 2. Сдвиг максимума содержания цитокина с 3 на 1 сутки, является косвенным признаком уменьшения выраженности реакций острого асептического воспаления в ответ на развитие церебральной ишемии. Особую важность этому факту придает то, что по данным литературы существует прямо пропорциональная зависимость между уровнем ИЛ-1β и размерами очага инфаркта мозга [Е.В. Супрун и соавт., 2009; J. Galea, D. Brough, 2013]. ИЛ-6 является типичным ранним индуцибельным цитокином и синтезируется многими типами клеток, участвующими в инициации и регуляции воспаления и иммунного ответа [D.A. Papanicolaou et al., 1998; С.А. Кетлинский, А.С. Симбирцев, 2008]. Он оказывает выраженное провоспалительное действие за счет активации экспрессии молекул адгезии и активации хемотаксиса лейкоцитов, а также является важнейшим индуктором острофазного ответа [K. Paul-Pletzer, 2006; С.А. Кетлинский, А.С. Симбирцев, 2008; I.M. Cojocaru et al., 2009; T. Tanaka et al., 2013]. 130 Динамика содержания ИЛ-6 в плазме крови крыс групп № 2, № 3 и № 4 на 1, 3, 7 и 14 сутки с момента воспроизведения модели ИИ значительно отличалась. Особенностью у животных группы № 2 явилось значительное стойкое повышение уровня ИЛ-6, достигавшее максимума на 3 сутки и остававшееся на этих значениях до 14 суток, что отражало активное течение асептического воспаления в ответ на развитие церебральной ишемии (рис. 5.4). Рисунок 5.4 — Изменение содержания ИЛ-6 при моделировании ишемического инсульта без и с применением ТЭС-терапии У животных группы № 3 содержание ИЛ-6 также достигало максимума на 3 сутки, однако на значительно более низком уровне (на 47,1 %). К 7 суткам уровень ИЛ-6 резко снижался почти до значений как у животных группы № 1 (превышал на 48,8 %). У животных группы № 4 динамика уровня ИЛ-6 имела тот же характер, однако содержание было еще более низким (на 84,3 %). К 7 суткам оно уменьшилось, а к 14 суткам возвращалось к уровню животных группы № 1. Указанные отличия объясняются более высоким уровнем β-эндорфина в группах № 3 и № 4 в отличие от группы № 2. Кроме того, известно, что ИЛ-1 и ФНО-α являются мощными индукторами ИЛ-6-синтетазы [H. Boutin et al., 2001]. 131 Как уже было отмечено при развитии острой локальной церебральной ишемии существует прямо пропорциональная зависимость между содержанием провоспалительных цитокинов (ИЛ-1, ФНО-α, ИЛ-6, ИЛ-8) и конечными размерами инфаркта мозга (зона некроза) [Е.И. Гусев, В.И. Скворцова, 2001; H. Boutin et al., 2001; K.Y. Kwan, B.H. Jeon, 2001; N. Quan, M. Herkenham, 2002; Q. Wang, X.N. Tang, M.A. Yenari, 2007; D. Amantea et al., 2008; I.M. Cojocaru et al., 2009; M. Zhang et al., 2011; М.В. Онуфриев, 2014]. В 1 сутки после моделирования ИИ у крыс группы № 2 удельный объем очага инфаркта мозга был достоверно (р ≤ 0,05) больше на 4,2 %, чем у животных группы № 3, которым до моделирования ИИ проводилась ТЭС-терапия и на 8,9 % достоверно (р ≤ 0,05), чем у крыс группы № 4, которым проводили ТЭС-терапию после моделирования ИИ (рис. 5.5). Рисунок 5.5 — Динамика удельного объема очага инфаркта мозга после моделирования ишемического инсульта без и с проведением ТЭС-терапии На 1 сутки после моделирования ИИ у крыс групп № 3 и № 4 объем очага инфаркта мозга достоверно (р ≥ 0,05) не отличался. Следовательно, проведение сеансов ТЭС-терапии как до, так и после формирования очага церебральной ишемии, даже в таких жестких условиях как перманентное нарушение мозгового кровотока, вследствие коагуляции ПСМА, приводит достоверному сокращению размеров очага инфаркта мозга, по сравнению с животными, которым ТЭС-терапия не выполнялась. 132 Удельный объем очага инфаркта мозга на 3 сутки после моделирования ИИ у крыс группы № 2 стал достоверно (р ≤ 0,05) больше на 21,6 %, в сравнении с удельным объемом очага инфаркта мозга у крыс группы № 3. Сравнение удельного объема очага инфаркта мозга крыс группы № 2 и № 4 на 3 сутки после моделирования ИИ показало, что у крыс группы № 2 он достоверно (р ≤ 0,05) больше на 28,0 %, чем у животных группы № 4. У крыс группы № 3 удельный объем очага инфаркта мозга на 3 сутки после моделирования ИИ был достоверно (р ≤ 0,05) больше на 8,3 % по сравнению с животными группы № 4. Согласно данным литературы именно к этому сроку завершается увеличение размеров инфаркта мозга [S.H. Graham, J. Chen, 2001; M.P. Mattson et al., 2001; N.W. Manning et al., 2014]. Вышеуказанные различия в размерах очага инфаркта мозга у крыс групп № 2, № 3 и № 4, ассоциируются со значительными отличиями в содержании β-эндорфина в плазме крови. Уровень β-эндорфина в плазме крови у крыс группы № 3 на 72,98 %, а у крыс группы № 4 на 71,34 % достоверно больше (р ≤ 0,05), по сравнению с содержанием у крыс группы № 2 (рис. 5.1). После проведения ТЭС-терапии в 1 сутки морфологически зона инфаркта ткани мозга менее выражена. Значительно меньше наблюдается некротизированных и гиперхромных дистрофически измененных нейронов. В связи с этим менее активно развивается воспалительный клеточной ответ и меньше перицеллюлярный отек ткани мозга. На 14 сутки формирующийся глиомезодермальный рубец выглядит более зрелым. Таким образом, ОП, в большом количестве выделяющиеся при ТЭС-терапии способны ограничивать глутаматную эксайтотоксичность, а также благодаря открытию АТФ-зависимых калиевых каналов запускать реакции ишемического пре- и посткондиционирования, что в свою очередь способно объяснить нейропротективный эффект на 1 сутки после развития острой локальной церебральной ишемии [G.J. Gross, 2000; Н.В. Соленкова, 2002; H. Zhang et al., 2006; Е.И. Барзах, 2006; J.M. Gidday, 2006; M. Iwata 133 et al., 2007; D.S. Su et al., 2007; A.K. Rehni, N. Singh, S.A. Jaggi, 2007; C. Happel et al., 2008; M. Govindaswami et al., 2008; Y. Yang et al., 2009; S. Husain, D.E. Potter, C.E. Crosson, 2009; M.R. Hutchinson et al., 2011; S. Husain, G.I. Liou, C.E. Crosson, 2011; Y. Feng et al., 2011; N. Akhter et al., 2013; M. Staples et al., 2013; К.П. Иванов, 2013]. Противовоспалительные цитокины ИЛ-10 и ТФР-1β, подавляя развитие воспаления, оказывают защитный эффект в условиях ишемии ГМ [K. Strle et al., 2001; Y. Zhu et al., 2002; N. Quan, M. Herkenham, 2002; N. Vila et al., 2003; Г.Н. Жданов, 2006; M. Kaste, H.J. Castillo, S. de Compostela, 2006; N. Liu et al., 2009]. В плазме крови животных из группы № 3 на 1 сутки после моделирования ИИ содержание ИЛ-10 достоверно (р ≤ 0,05) повышалось, по сравнению с животными группы № 1 и достоверно (р ≥ 0,05) не отличалось от животных группы № 2. Содержание ИЛ-10 достигло максимума на 3 сутки (достоверно (р ≤ 0,05) выросло на 41,2 %) и оставалось на том же уровне до 14 суток. У животных группы № 2 максимум достигался только на 7 сутки и к 14 суткам достоверно (р ≤ 0,05) падал на 40,9 % (рис. 5.6). Рисунок 5.6 — Изменение содержания ИЛ-10 при моделировании ишемического инсульта без и с применением ТЭС-терапии 134 В плазме крови животных группы № 4 на 1 сутки после моделирования ИИ уровень ИЛ-10 был наиболее высоким достоверно (р ≤ 0,05) больше на 98,22 %, чем у интактных животных и достоверно (р ≤ 0,05) выше на 41,67 % по сравнению с животными группы № 2. Он достоверно (р ≤ 0,05) превышал на 27,35 % уровень в группе № 3. На 3 сутки содержание ИЛ-10 достигло максимума как и в группе № 3. Однако достоверно (р ≤ 0,05) превышало последний на 14,9 %. Затем как и у животных группы № 3 снижался к 14 суткам (рис. 5.6). Исследование уровня ТФР-1β у животных групп № 3 и № 4 показало, что в группе № 3 наблюдался достоверно (р ≤ 0,05) меньший рост концентрации цитокина на 1 сутки по сравнению с группой № 2, а уже на 3, 7 и 14 сутки концентрация цитокина достоверно не отличалась от интактных животных. У животных группы № 4 на 1 сутки, как и в группе № 3 также наблюдалось возрастание уровня цитокина. На 3 и 7 сутки как и в группе № 3 содержание ТФР-1β также как в группе № 3 снижалось, однако было достоверно (р ≤ 0,05) более высоким (на 47,0 %). Имело место небольшое плато от 3 до 7 суток. Затем у животных групп № 3 и 4 на 14 сутки отмечено достоверное (р ≤ 0,05) повышение уровня ТФР-1β на 36,2 % и 35,6 % соответственно (рис. 5.7). Рисунок 5.7 — Изменение содержания ТФР-1β при моделировании ишемического инсульта без и с применением ТЭС-терапии 135 Нейропротективный эффект ТФР-1β обеспечивается за счет угнетения иммунных реакций в ткани мозга, а также понижения чувствительности клеток ГМ к ишемии [J.J. Letterio, 2000; Y. Zhu et al., 2002]. Но имеет место и другая сторона биологической активности ТФР-1β, а именно усиление образования внеклеточного матрикса. Однако в условиях значительного долговременного повышения концентрации цитокина запускается и поддерживается хроническая воспалительная реакция [С.А. Кетлинский, А.С. Симбирцев, 2008]. Выраженным повышением ТФР-1β у животных группы № 2 можно объяснить большую выраженность глиомезодермального рубца на 14 сутки, так как ТФР-1β является, наряду со своим нейропротективным действием, одним из главных индукторов фиброза в тканях [S. Fichtner-Feigi et al., 2006; A. Popa-Wagner et al., 2006]. У животных из групп № 3 и № 4 уровень ТФР-1β, низкий на 3 и 7 сутки, к 14 суткам его содержание повышается, данный паттерн выработки цитокина видимо способствует более качественному формированию рубца, без излишнего его разрастания [A. Popa-Wagner et al., 2006]. Применение ТЭС-терапии в условиях моделирования ИИ приводило к формированию очага инфаркта мозга меньших размеров, а также нормализовало процессы репарации в очаге инфаркта мозга (более зрелый и значительно менее выраженный глиомезодермальный рубец который не блокировал ход нервных волокон). Это обусловлено гомеостатическим ТЭС-терапии на баланс про- и противовоспалительных цитокинов. влиянием 136 ВЫВОДЫ 1. Нарушения цитокинового профиля в острейший период экспериментального ишемического инсульта характеризуются резким повышением уровня про- и противовоспалительных, со значительным преобладанием провоспалительных цитокинов. Даже к 14 суткам уровень ФНО-α, ИЛ-1β и ИЛ-6, значительно превышал контрольные показатели (на 59,9 %, 75,8 % и 95,8 % соответственно). Это сочеталось с повышением содержания ИЛ-10 и ТФР-1β на 97,1 % и 51,4 % соответственно. Содержание β-эндорфина было на 57,27 % меньше контроля. 2. При экспериментальном ишемическом инсульте морфологическая картина в зоне ишемии характеризовалась завершением формирования очага деструкции к 3 суткам после моделирования ИИ, с возрастанием удельного объема очага инфаркта мозга на 18,3 %. К 14 суткам формировался выраженный избыточный глиомезодермальный рубец. 3. Использование ТЭС-терапии до моделирования ишемического инсульта снижало уровни ФНО-α (не отличался от контрольного), ИЛ-1β на 70,5 %, ИЛ-6 на 93,3 %, соответственно, к 14 суткам по сравнению с животными группы № 2. При этом по сравнению с животными группы № 2 возрастало содержание ИЛ-10 на 39,3 % и снижался уровень ТФР-1β на 34 %. Уровень β-эндорфина в плазме крови был выше (чем у животных группы № 2) на 77,95 %. 4. При проведении ТЭС-терапии после моделирования ишемического инсульта динамика ФНО-α не отличалась от предыдущей группы. Содержание к 14 суткам ИЛ-1β было ниже на 88,5 % по сравнению с животными группы № 2. Содержание ИЛ-6 на 14 сутки не отличалось от контрольного. Эти данные свидетельствуют о более низкой активности воспалительного процесса под влиянием ТЭС-терапии. На этом фоне также не происходило падения содержания ТФР-1β и возрастал уровень ИЛ-10 (у животных группы 137 № 2 он падал). Уровень β-эндорфина в плазме крови был выше (чем у животных группы № 2) на 77,2 %. 5. Проведение ТЭС-терапии в обеих группах уменьшало размеры зоны инфаркта ткани мозга уже на 1 сутки после воссоздания модели инсульта. Значительно меньше наблюдалось некротизированных и гиперхромных дистрофически измененных нейронов. В связи с этим менее активно развивался воспалительный клеточной ответ и был меньше перицеллюлярный отек ткани мозга. На 14 сутки сформированный глиомезодермальный рубец выглядит более зрелым, не блокировал ход нервных волокон. Удельный объем очага инфаркта мозга на 1 сутки у крыс из групп № 3 и № 4 по сравнению с группой № 2 был меньше на 8,9 %. Удельный объем очага инфаркта мозга на 3 сутки у крыс из группы № 3 был на 22 % и в группе № 4 на 28 % меньше по сравнению с группой № 2. 6. Полученные факты свидетельствуют о наличии выраженного нейропротекторного и противовоспалительного эффекта ТЭС-терапии при применении в острейшем периоде экспериментального ишемического инсульта. 138 ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ 1. Экспериментальные исследования показали принципиальную возможность использования ТЭС-терапии с целью коррекции гипоэндорфинемии, дисбаланса цитокинового статуса в острейшем периоде ишемического инсульта. Применение ТЭС-терапии позволяет путем коррекции цитокинового статуса и нарушений в активности эндогенной опиоидергической системы ограничивать расширение очага инфаркта мозга при развитии ишемического инсульта. 2. Применение ТЭС-терапии в острейшем периоде ишемического инсульта показало ее нейропротекторные свойства в следующем режиме: использование биполярного импульсного тока ежедневно, сила тока подбирается индивидуально (от 1 до 2 мА, в среднем 1,7 мА), положение электродов фронто-мастоидальное. Длительность 1-го сеанса составляет 15 минут, всех последующих — 30 минут. Сеансы проводят в течение 7–10 дней после возникновения ишемического инсульта. 3. Применение ТЭС-терапии для профилактики возникновения ишемического инсульта при действии на организм факторов риска: использование биполярного импульсного тока ежедневно, сила тока подбирается индивидуально (от 1 до 2 мА, в среднем 1,7 мА), положение электродов фронтомастоидальное. Длительность 1-го сеанса составляет 15 минут, всех последующих — 30 минут. Сеансы проводят в течение 7–10 дней перед вмешательством с высоким риском возникновения ишемического инсульта. 139 В заключении выражаю сердечную благодарность своему научному руководителю, учителю и высокоуважаемому мною человеку — доктору медицинских наук, профессору Азамату Халидовичу Каде за предоставленную тему исследования, руководство и создание всех условий для выполнения диссертационной работы. Искренне признателен кандидату медицинских наук, доценту Сергею Александровичу Занину за критическое обсуждение и консультативную помощь по ходу выполнения всех этапов исследования. Хочу отметить помощь и поблагодарить кандидата медицинских наук, доцента Евглевского Андрея Александровича — его советы в области гистотехники, микрофотографии и описания микропрепаратов были воистину бесценны. Отдельно за повседневную помощь, поддержку в работе и доброжелательное отношение благодарю коллектив кафедры общей и клинической патофизиологии ГБОУ ВПО КубГМУ Минздрава России. 140 СПИСОК ЛИТЕРАТУРЫ 1. Автандилов, Г.Г. Медицинская морфометрия. Руководство. – М. : Медицина, 1990. – 384 с. 2. Акимов, Г.А. Транскраниальное воздействие в лечении вегетососудистой дистонии / Г. А. Акимов [и соавт.] // Журн. невропатологии и психиатрии. – 1991. – № 7. – С. 75–78. 3. Акмаев, И.Г., Гриневич, В.В. Нейроиммуноэндокринология гипоталамуса. – М. : Медицина, 2003. – 168 с. 4. Акоев, Г.Н. Влияние транскраниальной электростимуляции опиоидных структур мозга на регенерацию периферических нервов крысы / Г.Н. Акоев [и соавт.] // Нейрофизиология. – 1990. – Т. 22. – № 1. – С. 76–79. 5. Александрова, В.А. Транскраниальная электростимуляция в терапии хронических гастродуоденитов у детей / В.А. Александрова, С.В. Рычкова, В.П. Лебедев // Междунар. мед. обзоры. – 1994. – Т. 2. – № 3. – С. 204–209. 6. Алексеенко, А.В. Потенциальная роль сфинголидипидов в нейропатогенезе болезни Альцгеймера / А.В. Алексеенко // Биомедицинская химия. – 2013. – Т. 59. – № 1. – С. 25–50. 7. Айрапетов, Л.Н. Топография зон связывания эндогенных опиоидных пептидов, высвобождаемых на фоне транскраниальной электроанальгезии / Л.Н. Айрапетов [и соавт.] // Новый метод транскраниального обезболивания: тез. докл. науч.-практ. конф. – Л., 1987. – С. 14–15. 8. Аракелян, А.А. Роль цитокинов при ишемическом инсульте / А.А. Аракелян [и соавт.] // Клиническая медицина. – 2005. – Т. 83. – № 10. – С. 22–24. 9. Баева, Т.А. Иммунорегуляторные эффекты естественных и синтетических лигандов сигма- и мю-опиоидных рецепторов : автореф. дис. … канд. мед. наук. – Пермь, 2003. – 22 с. 141 10. Баева, Т.А. Роль блокады µ и δ-опиатных рецепторов в динамике развития гуморального иммунного ответа / Т.А. Баева, В.О. Небогатиков / Вестник Уральской медицинской академической науки. – 2012. – № 14. – С. 13–14. 11. Барабанова, М.А. Изменение содержания Р-эндорфина в сыворотке крови больных с ишемическим инсультом в остром периоде / М.А. Барабанова // Неврологический журнал. – 2003. – Т. 8. – № 5. – С. 18. 12. Барзах, Е.И. Использование лигандов дельта-опиоидных рецепторов для патологически обоснованной защиты сердца при ишемии и реперфузии : автореф. дис. … канд. мед. наук. – Томск, 2006. – 22 с. 13. Белоцкий, С.М., Авталион, Р.Р. Воспаление. Мобилизация клеток и клинические эффекты. – М. : Издательство БИНОМ, 2008. – 240 с. 14. Богданова, Ю.А. Иммуномодулирующие эффекты транскраниальной электростимуляции у больных с вторичной иммунной недостаточ- ностью : автореф. дис. … канд. мед. наук. – Краснодар, 2003. – 23 с. 15. Богданова, Ю.А. Динамика иммунного статуса в послеоперационном периоде при использовании транскраниальной электростимуляции / Ю.А. Богданова, А.Х. Каде, Р.А. Ханферян // Int. J. Immunorehab. – 2002. – Т. 4. – № 1. – С. 137. 16. Богданова, Ю.А. Применение транскраниальной электростимуляции у больных с вторичной иммунной недостаточностью / Ю.А. Богданова, А.Х. Каде, Р.А. Ханферян // Аллергология и иммунология. – 2000. – № 1(2). – С. 33–34. 17. Борисенко, В.Г. Повышение устойчивости миокарда к ишемии методом ТЭС-терапии : автореф. дис. … канд. мед. наук. – Краснодар, 2009. – 22 с. 18. Бояджян, А.С. Циркулирующие иммунные комплексы в семьях с наследственной отягощенностью ишемическим инсультом // А.С. Бояджян [и соавт.] // Журнал неврологии и психиатрии им. С.С. Корсакова. – 2007. – Т. 107. – № S20. – С. 43–46. 142 19. Брагин, Е.О. Нейрохимические механизмы регуляции болевой чувствительности. – М. : Медицина, 1991. – 193 с. 20. Брагин, Е.О., Яснецов, В.В. Опиоидные и моноаминовые механизмы регуляции функций организма в экстремальных условиях. – М. : Медицина, 1991. – 228 с. 21. Бугрова, А.Е. Роль сигнальной системы сфингомиелинового цикла в развитии болезни Альцгеймера : автореф. дис. … канд. биол. Наук. – М., 2007. – 23 с. 22. Бэр, М. Нейропротекция: модели, механизмы, терапия / М. Бэр. – М. : БИНОМ. Лаборатория знаний, 2011. – 429 с. 23. Волосникова, Н.Н. Состояние активности эндорфинергической системы головного мозга при острых нарушениях мозгового кровообращения : автореф. дис. … канд. мед. наук. – Саратов, 2004. – 24 с. 24. Вчерашнюк, С.П. Влияние ТЭС-терапии на гормональный гомеостаз и микроциркуляцию у беременных с гестозом : автореф. дис. … канд. мед. наук. – Краснодар, 2011. – 23 с. 25. Гейн, С.В. Опиоидергическая модуляция иммунных реакций // Молекулярная медицина. – 2010. – № 4. – С. 17–21. 26. Гейн, С.В. Влияние b-эндорфина на антителогенез и продукцию ИЛ-4 в условиях блокады опиатных рецепторов / С.В. Гейн, Т.А. Баева, О.А Кичанова // Бюлл. экспериментальной биол. и мед. – 2006. – Т. 142. – № 8. – С. 192–195. 27. Гейн, С.В. Оценка влияния бета-эндорфина на продукцию ИЛ-4 и гамма-ИФН CD4+-лимфоцитами / С.В. Гейн, К.Г. Горшкова // Бюлл. экспериментальной биол. и мед. – 2008. – Т. 145. – № 10. – С. 427–430. 28. Голиков, А.П. Способ лечения больных с острым инфарктом миокарда / А.П. Голиков, В.А. Рябинин, В.П. Лебедев [и соавт.]. – А. с. СССР, 1989. – № 1507404. 29. Голиков, А.П. Влияние транскраниальной электростимуляции на репаративные процессы у больных инфарктом миокарда / А.П. Голиков [и соавт.] // Кардиология. – 1989. – Т. 29. – № 12. – С. 45–48. 143 30. Горшкова, К.Г. Опиоидные пептиды в регуляции функций эффекторов врожденного иммунитета : автореф. дис. … канд. биол. Наук. – Екатеринбург, 2012. – 25 с. 31. Грицкевич, Н.Л. Неспецифическая резистентность организма при транскраниальном электрическом воздействии в режиме электроанальгезии / Н.Л. Грицкевич [и соавт.] // Новый метод безлекарственного лечения : тез. докл. науч. конф. – СПб., 1993. – С. 61–67. 32. Грицкевич, Н.Л. Влияние транскраниальной стимуляции в анальгетическом режиме на рост различных штаммов опухолей у крыс / Н.Л. Грицкевич [и соавт.] // Синтез, фармакологические и клинические аспекты новых обезболивающих средств : тез. докл. науч.-практ. конф. – Н. Новгород, 1991. – С. 56. 33. Грицкевич, Н.Л. Изменение интенсивности неспецифических защитных реакций у животных при транскраниальном электрическом воздействии / Н.Л. Грицкевич [и соавт.] // Новый метод транскраниального обезболивания. Теоретические основы и практическая оценка : тез. докл. науч.практ. конф. – Л., 1987. – С. 53–54. 34. Губарева, Е.А. Влияние ТЭС-терапии на показатели прооксидантноантиоксидантной системы при остром инфаркте миокарда : автореф. дис. … канд. мед. наук. – Краснодар, 2009. – 22 с. 35. Губарев, Ю.Д. Роль иммунной системы в патогенезе острых и хронических повреждений головного мозга / Ю.Д. Губарев, А.О. Шеремет // Научные ведомости Белгородского государственного университета. Серия : Медицина. Фармация. – 2009. – Т. 59. – № 7. – С. 47–52. 36. Гусев, Е.И. Ишемический инсульт. Современное состояние проблемы / Е.И. Гусев, М.Ю. Мартынов, П.Р. Камчатнов // Доктор.Ру. – 2013. – Т. 83. – № 5. – С. 7–12. 37. Гусев, Е.И. Неврология: национальное руководство / Е.И. Гусев, А.Н. Коновалов, В.И. Скворцова [и соавт.]. – М. : ГЭОТАР-Медиа, 2009. – 1035 с. 144 38. Гусев, Е.И. Клинические рекомендации. Неврология и нейрохирургия / Е.И. Гусев, А.Н. Коновалов, А.Б. Гехт. – М. : ГЭОТАР-Медиа, 2007. – 368 с. 39. Гусев, Е.И., Скворцова, В.И. Ишемия головного мозга / Е.И. Гусев, В.И. Скворцова. – М : Медицина, 2001. – 328 с. 40. Дроздов, К.А. Исследование динамики повреждений мозга крыс при черепно-мозговой травме методом магнитно-резонансной томографии / К.А. Дроздов [и соавт.] // Тихоокеанский медицинский журнал. – 2011. – № 1. – С. 93–95. 41. Дубынин, В.А., Каменский, А.А. Бета-казоморфины и их роль в регуляции поведения. – М. : Товарищество научных изданий КМК, 2010. – 306 с. 42. Жданов, Г.Н. О связи течения ишемического инсульта головного мозга с содержанием интерлейкина-10 в сыворотке крови больных / Г.Н. Жданов // Иммунология. – 2006. – Т. 27. – № 1. – С. 26–27. 43. Заболотных, И.И. Эффективность лечения больных с психорганическим синдромом различной этиологии с помощью транскраниального электровоздействия / И.И. Заболотных [и соавт.] // Транскраниальная электростимуляция : эксперим.-клинич. исслед. / под ред. Д.П. Дворецкого. – СПб., 1998. – С. 380–389. 44. Заболотных, В.А. Применение транскраниального электровоздействия для лечения сосудистых заболеваний с нарушением центральной регуляции / В.А. Заболотных [и соавт.] // Новый метод безлекарственного лечения : тез. докл. науч.-практ. конф. – СПб., 1993. – С. 96. 45. Заболотных, В.А. Применение транскраниальной электроанальгезии при цефалгиях различного происхождения / В.А. Заболотных [и соавт.] // Вопр. курортологии, физиотерапии и лечебн. физ. культуры. – 1986. – № 2. – С. 26–28. 46. Зильбер, Ю.Д. Транскраниальная электростимуляция опиоидных структур мозга при лечении язвенной болезни желудка и двенадцатиперстной 145 кишки: репарационный эффект и изменение содержания β-эндорфинов и гастрина плазмы / Ю.Д. Зильбер [и соавт.] // Физиология пептидов : тез. докл. симп. – Л., 1988. – С. 75–76. 47. Зильбер, Ю.Д. Использование транскраниальной электроанальгезии в комплексной терапии язвенной болезни желудка и двенадцатиперстной кишки / Ю.Д. Зильбер, М.В. Степанова, Ю.В. Киселев // Новый метод транскраниального обезболивания : тез. докл. науч.-практ. конф. – Л., 1987. – С. 52–53. 48. Зильбертер, Ю.И. Энергетический метаболизм: от нейронов и глии к целому мозгу; норма, патология и коррекция / Ю.И. Зильбертер, Т.М. Зильбертер // Успехи физиологических наук. – 2012. – Т. 43. – № 2. – С. 37–53. 49. Зозуля, А.А. Нейроиммунология: молекулярные и клинические аспекты / А.А. Зозуля, С.Ф. Пшеничкин // Итоги науки и техники. – Серия: Иммунология. – М., 1990. – Т. 25. – С. 50–111. 50. Зюзина, Н.А. Коррекция центральной и периферической гемодинамики у больных гипертонической болезнью с помощью транскраниальной электростимуляции : автореф. дис. … канд. мед. наук. – Ростов-на-Дону, 2006. – 22 с. 51. Иванов, К.П. Гипоксия мозга и гибель нейронов вследствие нарушения микроциркуляции в мозге и регионарного мозгового кровообращения / К.П. Иванов // Регионарное кровообращение и микроциркуляция. – 2010. – Т. 9. – № 2. – С. 5–17. 52. Иванов, К.П. Критический обзор механизмов прекондиционирования / К.П. Иванов // Вестник Российской академии медицинских наук. – 2013. – № 4. – С. 58–62. 53. Иващенко, В.В. Стресс и синдром системного воспалительного ответа / В.В. Иващенко [и соавт.] // Экспериментальная и клиническая урология. – 2012. – № 4. – С. 20–22. 54. Илюхина, В.А. Особенности динамики функционального состояния центральной нервной системы при болевых синдромах под влиянием 146 транскраниальной электроанальгезии / В.А. Илюхина, М.П. Ломарев // Новый метод транскраниального обезболивания : тез. докл. науч.-практ. конф. – Л., 1987. – С. 37–38. 55. Каде, А.Х. Клинико-эксперименальные аспекты взаимодействия нервной и иммунной систем / А.Х. Каде, А.П. Парахонский // Фундаментальные исследования. – 2004. – № 4. – С. 34–35. 56. Канцельсон, Я.С. Оценка эффективности нового метода транскраниальной электроаналгезии в клинической анестезиологии / Я.С. Канцельсон, В.А. Леоско // Новый метод транскраниального обезболивания: тез. докл. науч. конф. – Л., 1987. – С. 20–22. 57. Кацнельсон, Я.С. Транскраниальная электроанальгезия как компонент анестезиологического пособия и средство купирования болевых синдромов : автореф. дис. … канд. мед. наук. – Л., 1985. – 18 с. 58. Кашаева, Л.Н. Иммунологические нарушения при церебральных инсультах и их коррекция / Л.Н. Кашаева, Л.М. Карзакова, В.Н. Саперов // Медицинская иммунология. – 2005. – Т. 7. – № 1. – С. 57–62. 59. Кетлинский, С.А., Симбирцев, А.С. Цитокины. – СПб. : ООО «Издательство Фолиант», 2008. – 552 с. 60. Ковалицкая, Ю. А. Неопиоидное действие β-эндорфина / Ю.А. Ковалицкая, Е.В. Наволоцкая // Биохимия. – 2011. – Т. 76. – № 4. – С. 469–486. 61. Когаладзе, Р.А. Работа с лабораторными животными в контексте биоэтики – история, современность, перспективы / Р.А. Когаладзе // Успехи физиологических наук. – 2004. – Т. 35. – № 2. – С. 92–109. 62. Колосова, Л.И. Стимуляция регенерации нерва опиоидными пептидами при экзогенном введении и усилении их эндогенной продукции / Л.И. Колосова [и соавт.] // Физиологическое и клиническое значение регуляторных пептидов : тез. докл. науч. конф. – Пущино, 1990. – С. 90. 63. Коржевский, Д.Э., Гиляров, А.В. Основы гистологической техники. – СПб. : СпецЛит, 2010. – 95 с. 147 64. Кость, А.С. Воспалительный ответ при разном характере мозгового инсульта в остром периоде заболевания / А.С. Кость, Б.Д. Луцик, Л.Е. Лаповець // Загальна патологiя та патологiчна фiзiологiя. – 2010. – Т. 5. – № 3. – С. 215–219. 65. Кравченко, А.Я. Роль цитокинов в развитии и течении сердечной недостаточности / А.Я. Кравченко, Ю.М. Черняева // Клиническая медицина. – 2013. – Т. 91. – № 10. – С. 11–16. 66. Краева, Н.И. Транскраниальная электростимуляция при лечении сенсоневральной тугоухости. Прогноз эффективности с помощью фокусированного ультразвука / Н.И. Краева [и соавт.] // Транскраниальная электростимуляция: эксперим.-клинич. исслед. / под ред. Д.П. Дворецкого. – СПб., 1998. – С. 462–467. 67. Крыжановский, Г.Н. Патофизиология нейроиммунных взаимодействий / Г.Н. Крыжановский, С.В. Магаева // Патогенез. – 2010. – Т. 8. – № 1. – С. 4–9. 68. Кузин, М.И. Влияние чрезкожной трансцеребральной электростимуляции в режиме электроанестезии на содержание β-эндорфина в спинномозговой жидкости и плазме крови / М.И. Кузин [и соавт.] // Бюл. эксперим. биологии и медицины. – 1984. – Т. 97. – № 5. – С. 515–516. 69. Ласукова, Т.В. Опиоидная система, ее возможная роль в адаптации организма спортсменов к действию гипоксии / Т.В. Ласукова, С.В. Низкодубова, Е.Ю. Мухтобарова // Вестник ТГПУ. – 2013. – № 12. – С. 215–221. 70. Лебедев, В.П. Значение серотонинергической системы мозга для развития транскраниальной электроанальгезии / В.П. Лебедев, А.Б. Савченко, В.А. Отеллин // Физиол. журн. СССР им. И.М. Сеченова. – 1995. – Т. 81. – № 10. – С. 36–43. 71. Лебедев, В.П. Влияние транскраниальной электростимуляции в анальгетическом режиме на сомато-симпатические рефлексы / В.П. Лебедев [и соавт.] // Физиол. журн. им. И.М. Сеченова. – 1992. – Т. 78. – № 11. – С. 40–50. 148 72. Лебедев, В.П. Об участии опиоидного и неопиоидного звеньев антиноцицептивной системы в физиологическом механизме транскраниальной электроанальгезии / В.П. Лебедев, А.Б. Савченко, А.В. Красюков // Синтез, фармакологические и клинические аспекты новых обезболивающих средств : тез. докл. науч. конф. – Н. Новгород, 1991. – С. 18–19. 73. Лебедев, В.П. Об опиатном механизме транскраниальной электроанальгезии у крыс и мышей / В.П. Лебедев, А.Б. Савченко, Н.В. Петряевская // Физиол. журн. СССР. – 1988. – Т. 74. – № 9. – С. 1249–1256. 74. Лебедев, В.П. Влияние активации структур мозга на процессы репарации / В.П. Лебедев [и соавт.] // Регуляция иммунного гомеостаза : тез. докл. 4 Всесоюзн. симп. – Л., 1986. – С. 147–148. 75. Лебедев, В.П. Адаптация мозга к болевым раздражениям: угнетение и дезинтеграция соматических и вегетативных реакций на боль с помощью неинвазивной электрической стимуляции опиоидных механизмов мозгового ствола / В.П. Лебедев [и соавт.] // Адаптивные компенсаторные процессы в головном мозге. – М., 1986. – С. 74–76. 76. Лебедев, В.П. Участие опиоидных и других медиаторных механизмов в регуляторных функциях антиноцицептивной системы мозга при ее транскраниальной активации // 15 съезд Всесоюзн. физиол. о-ва им. И.П. Павлова : тез. докл. – Кишинёв, 1986. – Т. 1. – С. 162–163. 77. Левичкин, В.Д. Влияние ТЭС-терапии на показатели системы про/антиоксиданты у крыс с экспериментальным ишемическим инсультом / В.Д. Левичкин [и соавт.] // Современные проблемы науки и образования. – 2014. – № 2. – С. 332. 78. Лилли, Р. Патогистологическая техника и практическая гистохимия. – М. : Мир, 1969. – 643 с. 79. Маслов, Л.Н. Опиоидные рецепторы. Состояние проблемы и перспективы / Л.Н. Маслов, Ю.Б. Лишманов, Г.Н. Смагин // Экспериментальная и клиническая фармакология. – 2002. – Т. 65. – № 2. – С. 70–75. 149 80. Маслов, Л.Н. Пептидные агонисты опиатных рецепторов – новая группа препаратов антиишемического, антиаритмического действия? / Л.Н. Маслов, Р.С. Карпов, Ю.Б. Лишманов [и соавт.] // Клиническая фармакология и терапия. – 2002. – Т. 11. – № 3. – С. 60–65. 81. Мельников, К. Н. Калиевые ионные каналы клеточных мембран / К.Н. Мельников [и соавт.] // Обзоры по клинической фармакологии и лекарственной терапии. – 2009. – Т. 7. – № 1. – С. 3–27. 82. МКБ 10- Международная классификация болезней 10-го пересмотра // Всемирная организация здравоохранения, 2007. 83. Мольберг, А.А. Цитокины как медиаторы нейроиммунных взаимодействий / А.А. Мольберг, Т.В. Гришина // Успехи физиологических наук. – 2006. – Т. 37. – № 1. – С. 18–27. 84. Нечайкина, О.В. Влияние β-эндорфина на сократительную активность изолированных лимфатических сосудов крысы / О.В. Нечайкина, С.Г. Петунов, А.С. Радилов // Регионарное кровообращение и микроциркуляция. – 2013. – Т. 12. – № 3. – С. 64–70. 85. Олейник, А.А. Рецепторы и механизмы реализации нейротропных эффектов цитокинов и факторов роста / А.А. Олейник, Р.С. Вастьянов // Успехи физиологических наук. – 2008. – Т. 39. – № 2. – С. 47–57. 86. Онуфриев, М.В. Нейроиммунные аспекты повреждения мозга после фокальной ишемии / М.В. Онуфриев [и соавт.] // Нейрохимия. – 2014. – Т. 31. – № 1. – С. 84–90. 87. Пермяков, А.В., Витер В.И., Неволин Н.И. Судебно-медицинская гистология. Руководство для врачей. – Ижевск–Екатеринбург : Экспертиза, 2003. – 214 с. 88. Пешков, М.В. Метод гистологической проводки тканей с использованием изопропанола и минерального масла / М.В. Пешков, И.И. Дыгало // Архив патологии. – 2009. – Т. 71. – № 3. – С. 39–41. 89. Повещенко, А.Ф. Цитокины – факторы нейроэндокринной регуляции / А.Ф. Повещенко, В.В. Абрамов, В.А. Козлов // Успехи физиологических наук. – 2007. – Т. 38. – № 3. – С. 40–46. 150 90. Полунина, А.Г. Опиоидные системы головного мозга: нейроанатомия и физиологические характеристики / А.Г. Полунина, Е.А. Брюн // Наркология. – 2013. – Т. 12. – № 7. – С. 83–91. 91. Пшенникова, М.Г. Врожденная эффективность стресс- лимитирующих систем, как фактор устойчивости к стрессорным повреждениям / М.Г. Пшенникова // Успехи физиологических наук. – 2003. – Т. 34. – № 3. – С. 55–67. 92. Реброва, О.Ю. Статистический анализ медицинских данных. Применение пакета прикладных программ STATISTICA. – М. : МедиаСфера, 2006. – 312 с. 93. Редько, А.Н. Региональные особенности первичной инвалидности взрослого населения Краснодарского края вследствие цереброваскулярных болезней / А.Н. Редько, Р.К. Карипиди, В.В. Завьялова // Кубанский научный медицинский вестник. – 2013. – № 5. – С. 161–164. 94. Рекомендации по ведению больных с ишемическим инсультом и транзиторными ишемическими атаками (ESO), 2008. 95. Розенблюм, А.С. Применение транскраниальной электростимуляции для лечения больных с нейросенсорной тугоухостью / А.С. Розенблюм [и соавт.] // Транскраниальная электростимуляция: эксперим.-клинич. исслед. / под ред. Д.П. Дворецкого. – СПб., 1998. – С. 454–461. 96. Рубцовенко, А.В. Иммунотропные эффекты транскраниальной электростимуляции : автореф. дис. … канд. мед. наук. – Краснодар, 1996. – 21 с. 97. Рычкова, С.В. Транскраниальная электростимуляция: обоснованность применения и клиническая эффективность / С.В. Рычкова // Физиотерапия, бальнеология и реабилитация. – 2006. – № 5. – С. 44–47. 98. Рычкова, С.В. Влияние транскраниальной электростимуляции (ТЭС) опиоидных структур головного мозга на процессы репарации слизистой желудка у крыс // Механизмы регуляции физиологических функций: тез. докл. науч. конф. – СПб., 1992. – С. 144–145. 151 99. Рычкова, С.В. Влияние транскраниальной электростимуляции на развитие экспериментальной язвы у крысы / С.В. Рычкова, В.А. Александрова // Хроническое воспаление и заболевания органов пищеварения : тез. докл. науч.- практ. конф. – Харьков, 1991. – С. 1–28. 100. Савченко, А.Б. Центральный анальгетический и периферические эффекты транскраниальной электростимуляции : автореф. дис. … канд. биол. наук. – Л., 1994. – 22 с. 101. Самотруева, М.А. Пути реализации нейро-иммунно-эндокринных взаимодействий / М.А. Самоструева, Д.Л. Теплый, И.Н. Тюренков // Естественные науки. – 2009. – № 4. – С. 112–130. 102. Сеин, О.Б. Иммунотропные эффекты опиоидных пептидов / О.Б. Сеин [и соавт.] // Вестник Курской государственной сельскохозяйственной академии. – 2009. – № 5. – С. 74–77. 103. Сепиашвили, Р.И. Иммунная система мозга и спинномозговой жидкости / Р.И. Сепиашвили // Аллергология и иммунология. – 2013. – Т. 14. – № 4. – С. 241–253. 104. Силачев, Д.Н. Сравнение магнитно-резонансной томографии и трифенилтетразолиевого выявления повреждений головного мозга как методов исследования экспериментальной фокальной ишемии / Д.Н. Силачев // Бюллетень экспериментальной биологии и медицины. – 2009. – Т.147. – № 2. – С.223–226. 105. Скворцова, В.И. Анализ медико-организационных мероприятий по профилактике инсультов и реабилитации постинсультных состояний на современном этапе / В.И. Скворцова, Г.С. Алексеева, Н.Ю. Трифонова // Электронный научный журнал «Социальные аспекты здоровья населения». URL : http://vestnik.mednet.ru/content/view/453/30/ от 19.03.2013 года. С. 1–8. 106. Соколова, Л.П. Адаптация к стрессу — основополагающий фактор жизнеспособности и когнитивной активности / Л.П. Соколова // Клиническая геронтология. – 2011. – Т. 17. – № 5–6. – С. 16–20. 152 107. Соленкова, Н.В. Участие Катф-каналов в реализации антиаритмического эффекта стимуляции мю- и дельта-опиатных рецепторов при острой ишемии, реперфузии и постинфарктном кардиосклерозе : автореф. дис. … канд. мед. наук. – Томск, 2002. – 22 с. 108. Трофименко, А.И. Динамика цитокинового статуса у крыс при моделировании ишемического инсульта / А.И. Трофименко [и соавт.] // Современные проблемы науки и образования. – 2014. – № 2. – С. 326. 109. Трофименко, А.И. Визуализация очага ишемии головного мозга у крысы тетразолиевым методом / А.И. Трофименко [и соавт.] // Международный журнал прикладных и фундаментальных исследований. – 2013. – № 5. – С. 99. 110. Трофименко, А.И. Моделирование церебральной ишемии посредством коагуляции средней мозговой артерии у крыс / А.И. Трофименко [и соавт.] // Фундаментальные исследования. – 2012. – № 2. – С. 215–218. 111. Супрун, Э.В. Метаболическая церебропротекция антагониста рецепторов интерлейкина-1 при экспериментальном гемморагическом инсульте // Архив клiнiчноi та експериментальноi медицини. – 2009. – Т. 18. – № 2. – С. 164–169. 112. Тиликин, В.С. Влияние ТЭС-терапии на динамику интерлейкина 4, 6, 10 у больных с острым пиелонефритом / В.С. Тиликин [и соавт.] // Фундаментальные исследования. – 2012. – № 4. – С. 129–132. 113. Титова, Ж.В. Роль цитокиновой сети в механизмах нейроиммунного взаимодействия (обзор литературы) / Ж.В. Титова, Г.М. Бодиенкова // Бюллетень Восточно-Сибирского научного центра СО РАМН. – 2013. – № 2–1 (90). – С. 171–175. 114. Фирсов, А.А. Современный взгляд на патогенез и интенсивную терапию острой церебральной ишемии / А.А. Фирсов, A.A. Смирнов, Т.А. Усанова [и соавт.] // Архивъ внутренней медицины. – 2012. – № 3. – С. 29–33. 115. Хана-Мурад, А.Х. Вторичное повреждение при мозговом инсульте и возможность восстановления функций мозга (роль цитокинов, нейротрофи- 153 ческих факторов, адгезивных молекул) / А.Х. Хана-Мурад, Л.И. Павлинова, А.А. Мокрушин // Нейрохимия. – 2007. – Т. 24. – № 2. – С. 121–131. 116. Хныченко, Л.К. Стресс и его роль в развитии патологических процессов / Л.К. Хныченко, Н.С. Сапронов // Обзоры по клинической фармакологии и лекарственной терапии. – 2003. – Т. 2. – № 3. – С. 2–15. 117. Цаканова, Г.В. Сравнительный анализ системы антиоксидантов и интенсивности процесса перекисного окисления липидов при сахарном диабете 2-го типа и ишемическом инсульте, отягощенном и не отягощенном сахарным диабетом 2-го типа / Г.В. Цаканова, В.А. Айвазян, А.С. Бояджян [и соавт.] // Бюллетень экспериментальной биологии и медицины. – 2011. – Т. 151. – № 5. – С. 496–499. 118. Чейдо, М.А. µ-опиоидергическая иммуномодуляция: нейрохимические основы / М.А. Чейдо, Г.В. Идова // Успехи физиологических наук. – 2009. – Т. 40. – № 2. – С. 12–25. 119. Черешнев, В.А. β-эндорфин эндогенный регулятор иммунных процессов / В.А. Черешнев, С.В. Гейн // Российский физиологический журнал им. Сеченова. – 2009. – Т. 95. – № 12. – С. 1279–1290. 120. Шапоренко, Р.В. ТЭС-терапия в лечении язвенной болезни желудка и двенадцатиперстной кишки у детей подросткового возраста : автореф. дис. … канд. мед. наук. – Ростов-на-Дону, 2008. – 18 с. 121. Шаравьева, И.Л. Влияние блокады мю-, дельта-опиатных рецепторов на функции иммунной системы при различных вариантах стресса : автореф. дис. … канд. биол. наук. – Екатеринбург, 2011. – 24 с. 122. Шток, В.Н. Справочник по формулированию клинического диагноза болезней нервной системы / В.Н. Шток, О.С. Левин. – М. : Мед. информ. агентство, 2006. – 358 с. 123. Adams, H.P. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment / H.P. Adams, B.H. Bendixen, L.J. Kappelle [et al.] // Stroke. – 1993. – V. 24. – № 1. – P. 35–41. 154 124. Adibhatla, R.M. Integration of cytokine biology and lipid metabolism in stroke / R.M. Adibhatla, R. Dempsey, J.F. Hatcher // Frontiers in bioscience. – 2008. – V. 13. – P. 1250–1270. 125. Adibhatla, R.M. Secretory phospholipase A2 is us regulated by TNF-α and IL-1 α/β after transient focal cerebral ischemia in rat / R.M. Adibhatla, J.F. Hatcher // Brain Research. – 2007. – P. 199–205. 126. Aggarwal, A. Cerebral ischemic stroke: sequels of cascade / A. Aggarwal, P. Aggarwal, M. Khatak [et al.] // International journal of pharma and bioscience. – 2010. – V. 1. – № 3. – P. 1–24. 127. Akhter, N. Delta-opioid receptors attenuate TNF-α induced MMP-2 secretion from human ONH astrocytes / N. Akhter, M. Nix, Y. Abdul [et al.] // Investigative ophthalmology & Visual Science. – 2013. – V. 54. – № 10. – P. 6605– 6611. 128. Alzheimer, C. Molecular and cellular biology of neuroprotection in the CNS. – Kluwer Academic / Plenum Publishers. – 2002. – 513 p. 129. Amantea, D. Post-ischemic brain damage: pathophysiology and role of inflammatory mediators / D. Amantea, G. Nappi, G. Bernardi [et al.] // J. the FEBS. – 2008. – V. 276. – P. 13–26. 130. Anderson, T.R. Spreading depression: imaging and blockade in the rat neocotical brain slice / T.R. Anderson, R.D. Andrew // J. Neurophysiol. – 2002. – V. 88. – P. 2713–2725. 131. Becker, K.J. Targeting the central nervous system inflammatory response in ischemic stroke / K.J. Becker // Curr. Opin. Neurol. – 2001. – V. 14. – № 3. – P. 349–353. 132. Benakis, C. Inflammation and stroke / C. Benakis, L. Hirt, R.A. Du Pasquier // Kardiovaskulare Medizin. – 2009. – V. 12. – № 5. – P. 143–150. 133. Bidlack, J.M. Detection and function of opioid receptors on cells from the immune system / J.M. Bidlack // Clinical and diagnostic laboratory immunology. – 2000. – V. 7. – № 5. – P. 719–723. 155 134. Bodnar, R.J. Endogenous opiates and behavior / R.J. Bodnar // Peptides. – 2011. – V. 32. – № 12. – P. 25–22. 135. Boehncke, S. Endogenous µ-opiod peptides modulate immune response towards malignant melanoma / S. Boehncke, K. Hardt, D. Schadendorf [et al.] // Experimental dermatology. – 2010. – V. 20. – P. 24–28. 136. Borlongan, V.C. Hibernation-like state induced by an opioid peptide protects against experimental stroke / V.C. Borlongan, T. Hayashi, P.R. Oeltgen [et al.] // DMC Biology. – 2009. – P. 1–10. 137. Borner, C. Mechanisms of opioid-mediated inhibition of human T cell receptor signaling / C. Borner, B. Warnick, M. Smida [et al.] // J. Immunol. – 2009. – V. 183. – № 2. – P. 882–889. 138. Bose, A. The Penumbra System: a mechanical device for the treatment of acute stroke due to thromboembolism / A. Bose, H. Henkes, K. Alfke [et al.] // Am. J. Neuroradiol. – 2008. – V. 29. – № 7. – P. 1409–1413. 139. Boutin, H. Role of IL-1α and IL-1β in ischemic brain damage // H. Boutin, R.A. Le Feuvre, R. Horai [et al.] // The Journal of neuroscience. – 2001. – V. 21. – № 15. – P. 5528–5534. 140. Carr, D.J.J. The role endogenous opioids and their receptors and the immune system / D.J.J. Carr // Proc. Soc. Exp. Biol. Med. – 1991. – V. 198. – № 2. – P. 710–720. 141. Ceulemans, A.G. The dual role of the neuroinflammatory response after ischemic stroke: modulatory effects of hypothermia / A.G. Ceulemans, T. Zgavc, R. Kooijman [et al.] // J. Neuroinflammation. – 2010. – V. 74. – № 7. – P. 1–18. 142. Choi, S. Inflammatory cytokines IL-1β and TNF-α regulate p75NTR expression in CNS neurons and astrocytes by distinct cell-type-specific signalling mechanisms / S. Choi, W.J. Friedman // ASN Neuro. – 2009. – V. 1(2). – P. 113– 123. 143. Church, L.D. Primer: inflammasomes and interleukin 1β in inflammatory desorders / L.D. Church, P.G. Cook, M.F. Mc Dermott // Nature clinical practice rheumatology. – 2008. – V. 4. – № 1. – P. 34–42. 156 144. Clark, W.M. The rtPA (alteplase) 0- to 6-hour acute stroke trial: results of a double-blind, placebo-controlled, multicenter study. Thrombolytic therapy in acute ischemic stroke study investigators / W.M. Clark, G.W. Albers, K.P. Madden [et al.] // Stroke. – 2000. – V. 31(4). – P. 811–816. 145. Clarkson, A.N. The biology and pathology of hypoxia-ischemia: an update / A.N. Clarkson, B.A. Sutherland, I. Appleton // Arch. Immunol. Ther.Exp. – 2005. – V. 53. – P. 213–225. 146. Cojocaru, I.M. Expression of IL-6 activity in patients with acute ischemic stroke / I.M. Cojocaru, M. Cojocaru, R. Tanasescu [et al.] // Rom. Intern. J. Med. – 2009. – V. 47. – № 4. – P. 393–396. 147. Del Zoppo, G.J. Inflammation after stroke: is it harmful? / G.J. del Zoppo, K.J. Becker, J.M. Hallenbeck // Arch. Neurol. – 2001. – V. 58(4). – P. 669– 672. 148. Di Carlo, A. Human and economic burden of stroke / A. Di Carlo // Age ageing. – 2009. – V. 38(1). – P. 4–5. 149. Di Napoli, M. Systemic inflammation, blood pressure, and stroke outcome / M. Di Napoli, F. Papa // J. Clin. Hypertens (Greenwich). – 2006. – V. 8(3). – P. 187–194. 150. Dinarello, C.A. Interleukin-1 in the pathogenesis and treatment of inflammatory diseases / C.A. Dinarello // Blood. – 2011. – V. 117. – № 14. – P. 3720–3732. 151. Dinarello, C.A. Immunological and inflammatory functions of the interleukin-1 family annual review of immunology / C.A. Dinarello // Annu. Rev. Immunol. – 2009. – V. 27. – P. 519–550. 152. Dravolina, O.A. Decrement in operant performance produced by NMDA receptor antagonists in the rat: tolerance and cross-tolerance / O.A. Dravolina, E.E. Zvartau, A.Y. Bespalov // Pharmacology biochemistry and behavior. – 2000. – V. 65. – № 4. – P. 611–620. 153. Durukan, A. Acute ischemic stroke: overview of major experimental rodent models, pathophysiology and therapy of focal cerebral ischemia / A. Du- 157 rukan, T. Tatlisumak // Pharmacol. Biochem. Behav. – 2007. – V. 87(1). – P. 179–197. 154. Eskandari, F. Neural immune pathways and their connection to inflammatory diseases / F. Eskandari, J.I. Webster, M.S. Esther // Arthritis research & therapy. – 2003. – V. 5. – № 6. – P. 251–265. 155. Faas, M.M. Monocyte intracellular cytokine production during human endotoxaemia with or without a second in vitro LPS challenge: effect of RWJ-67657, a p38 MAP-kinase inhibitor, on LPS-hyporesponsiveness / M.M. Faas, H. Moes, J.W. Fijen [et al.] // Clin. Exp. Immunol. – 2002. – V. 127(2). – P. 337–343. 156. Fabricius, M. Cortical spreading depression and peri-infarct depolarization in acutely injured human cerebral cortex / M. Fabricius, S. Fuhr, R. Bhatia [et al.] // Brain. – 2006. – V. 29(Pt 3). – P. 778–790. 157. Faden, A.I. Neuroprotection: challenges and opportunities / A.I. Faden, B. Stoica // Arch. Neurol. – 2007. – V. 64(6). – P. 794–800. 158. Feng, Y. Effect of delta-opioid receptor over-expression on cortical expression of GABAA- receptor α1-subunit in hypoxia / Y. Feng, X. He, Y. Yang [et al.] // Chinese journal of physiology. – 2011. – V. 54. – № 2. – P. 118–123. 159. Fichtner-Feigi, S. IL-13 signaling through the IL-13 alpha 2 receptor is involved in induction of TGF-beta1 production and fibrosis/ S. Fichtner-Feigi [et al.] // Nat. Med. – 2006. – V. 12. – P. 99–106. 160. Finley, M.J. Opioid and nociceptin receptors regulate cytokine and cytokine receptor expression / M.J. Finley, C.M. Happel, D.E. Kaminsky [et al.] // Cell Immunol. – 2008. – V. 252(1–2). – P. 146v154. 161. Fisher, M. New perspectives on developing acute stroke therapy / M. Fisher, R. Ratan // Ann. Neurol. – 2003. – V. 53(1). – P. 10–20. 162. Fryer, R.M. Ischemic preconditioning in rats: role of mitochondrial K (ATP) channel in preservation of mitochondrial function / R.M. Fryer, J.T. Eells, A.K. Hsu [et al.] // Am. J. Physiol. Heart Circ. – 2000. – V. 278(1). – P. 305–312. 158 163. Galea, J. The role of inflammation and interleukin-1 in acute cerebrovascular disease / J. Galea, D. Brough // Journal of inflammation research. – 2013. – V. 6. – P. 121–128. 164. Garlanda, C. The interleukin-1 family: back the future / C. Garlanda, C.A. Dinarello, A. Mantovani // Immunity. – 2013. – V. 39. – № 6. – P. 1003–1018. 165. Garsia, J.B.S. Opioids and the immune system: clinical relevance / J.B.S. Garsia, M. G. de Melo Cardoso, M.C. Dos-Santos // Rev. Bras. Anestesiol. – 2012. – V. 62. – № 5. – P. 709–718. 166. Gidday, J.M. Cerebral preconditioning and ischemic tolerance / J.M. Gidday // Nat. Rev. Neurosci. – 2006. – V. 7(6). – P. 437–448. 167. Ginsberg, M.D. Adventures in pathophysiology of brain ischemia: penumbra, gene expression, neuroprotectiom / M.D. Ginsberg // Stroke. – 2003. – V. 34. – № 5. – P. 101–103. 168. Govindaswami, M. Delta2-specific opioid receptor agonist and hibernating woodchuck plasma fraction provide ischemic neuroprotection / M. Govindaswami, S.A. Brown, J. Yu [et al.] // Acad. Emerg. Med. – 2008. – V. 15. – № 3. – P. 250–257. 169. Graham, S.H. Programmed cell death in cerebral ischemia / S.H. Graham, J. Chen // J. Cereb. Blood Flow Metab. – 2001. – V. 21(2). – P. 99–109. 170. Graulich, J. Acute neuronal injury after hypoxia is influenced by the reoxygenation mode in juvenile hippocampal slice cultures / J. Graulich, U. Hoffmann, R.F. Maier [et al.] // Brain Res. Dev. – 2002. – V. 137(1). – № 30. – P. 35–42. 171. Green, H.F. A role for interleukin-1β in determining the lineage fate of embryonic rat hippocampal neural precursor cells / H.F. Green [et al.] // Mol. Cell Neurosci. – 2012. – V. 49(3) – P. 311–321. 172. Gross, G.L. The role of mitochondrial KATP channels in cardioprotection / G.L. Gross // Basic. Res. Cardiol. – 2000. – V. 95(4). – P. 280–284. 173. Happel, C. DAMGO-induced expression of chemokines and chemokine receptors: the role of TGF-beta1 / C. Happel, A.D. Steele, M.J. Finley [et al.] // J. Leukoc. Biol. – 2008. – V. 83(4). – P. 956–963. 159 174. Heuschmann, P.U. Incidence of stroke in Europe at the beginning of the 21-st century. European Registers of Stroke (EROS) Investigators / P.U. Heuschmann, A. Di Carlo, Y. Bejot [et al.] // Stroke. – 2009. – V. 40(5). – P. 1557–1563. 175. Hietbrink, F. Trauma: the role of the innate immune system / F. Hietbrink, L. Koenderman, G. Rijkers [et al.] // World J. Emerg. Surg. – 2006. – V. 20. – P. 1–15. 176. Hirasawa, H. Basic concept and definition of SIRS and sepsis-present consideration and future perspectives / H. Hirasawa, S. Oda, K. Matsuda [et al.] // Nihon Rinsho. – 2004. – V. 62. – № 12. – P. 2177–2183. 177. Hossmann, K.A. The two pathophysiologies of focal brain ischemia: implications for translational stroke research / K.A. Hossmann // J. Cereb. Blood Flow Metab. – 2012. – V. 32. – № 7. – P. 1310–1316. 178. Huang, Y. Neuron-specific effects of interleukin-1β are mediated by a novel isoform of the IL-1 receptor accessory protein / Y. Huang [et al.] // J. Neurosci. – 2011. – V. 31. – № 49. – P. 18048–18059. 179. Husain, S. Opioid receptor activation: suppression of ischemia/reperfusion-induced production of TNF-α in the retina / S. Husain, G.I. Liou, C.E. Crosson // Investigative ophthalmology & Visual Science. – 2011. – V. 52. – № 5. – P. 2577–2583. 180. Husain, S. Opioid receptor-activation: retina protected from ischemic injury / S. Husain, D.E. Potter, C.E. Crosson // Investigative ophthalmology & Visual Science. – 2009. – V. 50. – № 8. – P. 3853–3859. 181. Hutchinson, M.R. Exploring the neurommunopharmaccology of opioids: an integrative review of mechanisms of central immune signaling and their implications for opioid analgesia / M.R. Hutchinson, Y. Shavit, P.M. Grace [et al.] // Pharmacol. Rev. – 2011. – V. 63. – № 3. – P. 772–810. 182. Ishihara, K. IL-6 in autoimmune disease and chronic inflammatory proliferative disease / K. Ishihara, T. Hirano // Cytokine Growth Factor Rev. – 2002. – V. 13. – P. 357–368. 160 183. Iwaszkiewicz, K. S. Targeting peripheral opioid receptor promote analgesic and anti-inflammatory actions / K.S. Iwaszkiewicz, J.J. Schneider, S. Hua // Frontiers in pharmacology. – 2013. – V. 4. – P. 1–7. 184. Iwata, M. Effects of delta-opioid receptor stimulation and inhibition on hippocampal survival in a rat of forebrain ischemia / M. Iwatai, S. Inouei, M. Kawaguchi [et al.] // British journal of anaesthesia. – 2007. – V. 99. – № 4. – P. 538–546. 185. Jauch, E.C. Guidelines for the early management of patients with acute ischemic stroke: a guideline for healthcare professionals from the American Heart Association / American Stroke Association / E.C. Jauch, J.L. Saver, H.P. Adams [et al.] // Stroke – 2013 – V. 44(3). – P. 870–947. 186. Jeong, H.K. Inflammatory response are not sufficient to cause delayed neuronal death in ATP-induced acute brain injury / H.K. Jeong, K.M. Ji, B. Kim [et al.] // PLoS ONE. – 2010. – V. 5. – P. 1–14. 187. Johansson, A. Cortisol axis abnormalities early after stroke- relationships to cytokines and leptin / A. Johansson, B. Ahren, B. Nasman [et al.] // Journal of intimal medicine. – 2000. – V. 247. – P. 179–187. 188. Jones, S. Directing transition from innate to acquired immunity: defining a role for IL-6 / S. Jones // J. Immunol. – 2005. – V. 175. – P. 3463–3468. 189. Jonsdottir, I.H. Neuropeptides and they interaction with exercise and immune function / I.H. Jonsdottir // Immunology and cell biology. – 2000. – V. 78. – P. 562–570. 190. Jordan, J. Inflammation as therapeutic objective in stroke / J. Jordan, T. Segura, M.F. Galindo [et al.] // Current Pharmaceutical design. – 2008. – V. 14. – P. 3549–3564. 191. Joy, M.L.G. Current density in sections through rabbit brain / M.L.G. Joy [et al.] // SMR 2 ׳nd Meeting Proceeding. – 1994. – V. 3. – P. 1404. 192. Joy, M.L.G. Low frequency current density imaging in rabbits / M.L.G. Joy [et al.] // Ann. Biomed. Engin. – 1993. – V. 21, suppl. 1. – P. 56. 161 193. Kaste, M. Neuroprotection in brain ischemia: an update / M. Kaste, H.J. Castillo, S. de Compostela // Cerebrovascular diseases. – 2006. – V. 21. – Suppl. 2. – P. 48–54. 194. Kawalec, M. Neuroprotective potential of biphalin, multireceptor opioid peptide, against excitotoxic injuri in hippocampal organotypic culture / M. Kawalec, J.E. Kowalczuk // Neurochem. Res. – 2011. – V. 36. – P. 2091–2095. 195. Klijn, C.J. Management of acute ischaemic stroke: new guidelines from the American Stroke Association and European Stroke Initiative / C.J. Klijn, G.J. Hankey // Lancet Neurol. – 2003. – V. 2(11). – P. 698–701. 196. Konsman, J.P. Perivascular production and action pro-inflammatory cytokines and brain pathology / J.P. Konsman, B. Drukarch, A.M. van Dam // Clinical science. – 2007. – V. 112. – P. 1–25. 197. Kwan, K.Y. Cytokine levels cerebrospinal fluid and delayed ischemic deficits in patients with aneurysmal subarachnoid hemorrhage/ K.Y. Kwan, B.H. Jeon // J. Korean Med. Sci. – 2001. – V. 16. – P. 774–780. 198. Lansberg, M.G. Risk factors of symptomatic intracerebral hemorrhage after tPA therapy for acute stroke / M.G. Lansberg, V.N. Thijs, R. Bammer [et al.] // Stroke. – 2007. – V. 38. – № 8. – P. 2275–2278. 199. Lasukova, T.V. Opioid system and adaptation to hypoxia / T.V. Lasukova, S.V. Nizkodubova, A.S. Tayurskaya // TSPU Bulleein. – 2011. – V. 8. – P. 109– 114. 200. Lay, A.Y. Microglia in cerebral ischemia: molecular actions and interactions / A.Y. Lay, K.G. Todd // Can. J. Physiol. Pharmacol. – 2006. – V. 84. – P. 49–59. 201. Lebedev, V. P. Devices for non-invasive transcranial electrostimulation of the brain endorphinergic system: application for improvement of human psychophysiological status / V. P. Lebedev [et al.] // Proceedings of 7-th Int. Workshop on Functional Electrostimulation. – Vienna, 2001. – P. 131–135. 202. Letterio, J.J. Murine models define the role of TGF-beta as a master regulator of immune cell function / J.J. Letterio // Cytokine Growth Factor Rev. – 2000. – V. 11(1–2) – P. 81–87. 162 203. Leys, D. Facilities available in European hospitals treating stroke patients. Executive Committee of the European Stroke Initiative / D. Leys, E.B. Ringelstein, M. Kaste // Stroke. – 2007. – V. 38(11). – P. 2985–2991. 204. Liu, N. Expression of IL-10 and TNF-α in rats with cerebral infarction after transplantation with mesenchymal stem cells / N. Liu, R. Chen, H. Du [et al.] // Cellular & Molecular immunology. – 2009. – V. 6. – № 3. – P. 207–213. 205. Lloyd-Jones, D. Heart disease and stroke statistics — 2009 update: a report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. American Heart Association Statistics Committee and Stroke Statistics Subcommittee / D. Lloyd-Jones, R. Adams, M. Carnethon [et al.] // Circulation. – 2009. – V. 119(3). – P. 480–486. 206. Lopez, A.D. Measuring the global burden of disease and epidemiological transitions: 2002–2030 / A.D. Lopez, C.D. Mathers // Ann. Trop. Med. Parasitol. – 2006. – V. 100(5–6). – P. 481–499. 207. Lee, Y. Therapeutically targeting neuroinflammation and microglia after acute ischemic stroke / Y. Lee, S.R. Lee, S.S. Choi [et al.] // Biomed. Res. Int. – 2014. – P. 241–297. 208. Machelska, H., Stein, K. Immune mechanisms of pain and analgesia. – Plenum Publishers. – 2003. – 166 p. 209. Maddahi, A. Cerebral ischemia induces microvascular pro- inflammatory cytokine expression via the MEK/ERK pathway / A. Maddahi, L. Edvinsson // Journal of neuroinflammation. – 2010. – V. 7. – № 14. – P. 1–13. 210. Manning, N.W. Acute ischemic stroke: time, penumbra, and reperfusion / N.W. Manning, B.C. Campbell, T.J. Oxley [et al.] // Stroke. – 2014. – V. 45(2). – P. 640–644. 211. Marik, P.E. The effect of aging on circulating levels of proinflammatory cytokines during septic shock. North American Sepsis Trial II / P.E. Marik, G.P. Zaloga // J. Am. Geriatr. Soc. – 2001. – V. 49(1). – P. 5–9. 212. Markus, R. Hypoxic tissue in ischemic stroke: persistence and clinical consequences of spontaneous survival / R. Markus, D.C. Reutens, S. Kazui [et al.] // Brain. – 2004. – V. 127(Pt 6). – P. 1427–1436. 163 213. Mattson, M.P. Neurodegenerative disorders and ischemic brain diseases / M.P. Mattson, W. Duan, W.A. Pedersen [et al.] // Apoptosis. – 2001. – V. 6(1–2). – P. 69–81. 214. Mc Coll, B.W. Systemic inflammatory stimulus potentiates the acute phase and CXC chemokine responses to experimental stroke and exacerbates brain damage via Interleukin-1 and neutrophil-dependent mechanisms / B.W. Mc Coll, N.J. Rothwell, S.M. Allan // The Journal of neuroscience. – 2007. – V. 27. – № 16. – P. 4403–4412. 215. Moskowitz, M.A. The science of stroke: mechanisms in search of treatments / M.A. Moskowitz, E.H. Lo, C. Iadecola // Neuron. – 2010. – V. 67(2). – № 29. – P. 181–198. 216. Nabe, T. Regulatory role of antigen-induced interleukin-10, produced by CD4 (+) T cells, in airway neutrophilia in a murine model for asthma / T. Nabe [et al.] // Eur. J. Pharmacol. – 2012. – V. 677. – № 1–3. – P. 154–162. 217. Nyberg, F. Opioid peptides in cerebrospinal fluid-methods for analysis and their significance in the clinical perspective / F. Nyberg // Frontiers in Bioscience. – 2004. – V. 9. – P. 3510–3525. 218. Osuchowski, M.F. Circulating cytokine/inhibitor profiles reshape the understanding of the SIRS/CARS continuum in sepsis and predict mortality / M.F. Osuchowski, K. Welch, J. Siddiqui [et al.] // J. Immunol. – 2006. – V. 177(3). – P. 1967–1974. 219. Papanicolaou, D. A. The pathophysiologic roles of interleukin-6 in human disease / D.A. Papanicolaou [et al.] // Ann. Intern. Med. – 1998. – V. 128. – № 2. – P. 127–137. 220. Paul-Pletzer, K. Toclizumab: blocade of interleukin-6 signaling pathway as a therapeutic strategy for inflammatory disorders / K. Paul-Pletzer // Drugs Today (Barc.). – 2006. – V. 42(9). – P. 559–576. 221. Perry, L. Screening for dysphagia and aspiration in acute stroke: a systematic review / L. Perry, C.P. Love // Dysphagia. – 2001. – V. 16(1). – P. 7–18. 164 222. Pinard, E. Discovery of (R)-1-[2-hydroxy-3-(4-hydroxy-phenyl)propyl]-4-(4-methyl-benzyl)-piperidin-4-ol: a novel NR1/2B subtype selective NMDA receptor antagonist / E. Pinard, A. Alanine, A. Bourson [et al.] // Bioorg. Med. Chem. Lett. – 2001. – V. 20. – № 11. – P. 2173–2176. 223. Popa-Wagner, A. Premate cellular proliferation following cortical infarct in aged rats / A. Popa-Wagner, I. Badan, R. Vintilescu [et al.] // Rom. J. of Morphology and Embryology. – 2006. – V. 47. – № 3. – P. 215–228. 224. Priller, J. Targeting gene-modified hematopoietic cells to the central nervous system: use of green fluorescent protein uncovers microglial engraftment / J. Priller, A. Flugel, T. Wehner // Nat. Med. – 2001. – V. 7(12). – P. 1356–1361. 225. Quan, N. Connecting cytokines and brain: a review of current issues / N. Quan, M. Herkenham // Histology and Histopathology. – 2002. – V. 17. – P. 273–288. 226. Quintana, A. Differential role of tumor necrosis factor receptors in mouse brain inflammatory responses in cryolesion brain injury / A. Quintana, M. Giralt, S. Rojas [et al.] // Journal of Neuroscience research. – 2005. – V. 82. – P. 701–716. 227. Rehni, A.K. Possible involvement of insulin, endogenous opioids and calcitonin gene related peptide in remote ischemic preconditioning of brain / A.K. Rehni, N. Singh, S. A. Jaggi // The pharmaceutical society of Japan. – 2007. – V. 127. – № 6. – P. 1013–1020. 228. Rittner, H.L. Pain in the immune system / H.L. Rittner, A. Brack, C. Stein // British journal of anaesthesia. – 2008. – V. 101. – № 1. – P. 40–44. 229. Rohl, L. Correlation between diffusion- and perfusion-weighted MRI and neurological deficit measured by the Scandinavian Stroke Scale and Barthel Index in hyperacute subcortical stroke (< or = 6 hours) / L. Rohl, J. Geday, L. Ostergaard [et al.] // Cerebrovasc. Dis. – 2001. – V. 12(3). – P. 203–213. 230. Rother, J. Neuroprotection does not work! / J. Rother // Stroke. – 2008. – V. 39(2). – P. 523–524. 165 231. Schilling, M. Predoninant phagocytic activity of resident microglia over hematogenous macrophages following transient focal cerebral ischemia: an investigation using green fluorescent protein transgenic bone marrow chimeric mice / M. Schilling, M. Besselmann, M. Muller [et al.] // Experimental neurology. – 2005. – V. 196. – P. 290–297. 232. Sharma, S. Bioactive peptides: a review / S. Sharma, R. Singh, S. Rana // Int. J. BIOautomation. – 2011. – V. 15. – P. 223–250. 233. Shenhar-Tsarfaty, S. Early signaling of inflammation in acute ischemic stroke: clinical and rheological implications / S. Shenhar-Tsarfaty, E. Ben Assayag, I. Bova [et al.] // Thrombosis research. – 2008. – V. 122. – P. 167–173. 234. Simmons, M. L. Endogenous opioid regulation of hippocampal function / M.L. Simmons, C. Chavkin // Int. Rev. Neurobiol. – 1996. – V. 39. – P. 145– 196. 235. Smith, W.S. Pathophysiology of focal cerebral ischemia: a therapeutic perspective / W.S. Smith // J. Vasc. Interv. Radiol. – 2004. – V. 15. – P. 3–12. 236. Sinning, J.M. Systemic inflammatory response syndrome predicts increased mortality in patients after transcatheter aortic valve implantation / J.M. Sinning [et al.] // Eur. Heart J. – 2012. – V. 33(12). – P. 1459–1468. 237. Stanimirovic, D. Inflammatory mediators of cerebral endothelium: a role in ischemic brain inflammation / D. Stanimirovic, K. Saton // Brain Pathol. – 2000. – V. 10(1). – P. 113–126. 238. Staples, M. Delta opioid receptor and its peptide: a receptor ligand neuroprotection / M. Staples, S. Acosta, N. Taijiri [et al.] // International journal molecular sciences. – 2013. – V. 15. – P. 17410–17419. 239. Strle, K. Interleukin-10 in the brain / K. Strle, J.H. Zhou, W.H. Shen [et al.] // Crit. Rev. Immunol. – 2001. – V. 21(5). – P. 427–449. 240. Su, D.S. Dose-dependent neuroprotection of delta opioid peptide [D-Ala2, D-Leu5] encephalin in neuronal death and retarded behavior induced by forebrain ischemia in rats / D.S. Su, Z.H. Wang, Y.H. Zhao [et al.] // Neuroscience Letters. – 2007. – V. 423. – P. 113–117. 166 241. Tanaka, T. Interleukin-6; pathogenesis and treatment of autoimmune inflammatory diseases / T. Tanaka, M. Narazaki, K. Masuda [et al.] // Inflammation and regeneration. – 2013. – V. 33. – № 1. – P. 54–65. 242. Tanasescu, R. An immunological approach to cerebral ischemia (I). Immune cell and adhesion molecul / R. Tanasescu, A. Nicolau, M. Ticmeanu [et al.] // Rom. J. Intern. Med. – 2008. – V. 46. – № 1. – P. 3–8. 243. Tissue plasminogen activator for acute ischemic stroke. The National Institute of Neurological Disorders and Stroke rt-PA Stroke Study Group // N. Engl. J. Med. – 1995. – V. 333(24). – № 14. – P. 1581–1587. 244. Tomita Y. Moment analysis of mocroflow histogram in focal ischemic lesion to evaluate microvascular derangement after small pial arterial occlusion in rats / Y. Tomita, M. Tomita, I. Schiszler [et al] // J. Cereb. Blood Flow Metab. – 2002. – V. 22(6). – P. 663–669. 245. Tschaikowsky, K. Coincidence of pro- and anti-inflammatory responses in the early phase of severe sepsis: Longitudinal study of mononuclear histocompatibility leukocyte antigen-DR expression, procalcitonin, C-reactive protein, and changes in T-cell subsets in septic and postoperative patients / K. Tschaikowsky, M. Hedwig-Geissing, A. Schiele [et al.] // Crit. Care Med. – 2002. – V. 30(5). – P. 1015–1023. 246. Tuttolomondo, A. Inflammatory cytokines in acute ischemic stroke / A. Tuttolomondo, D. Di Raimondo, R. Di Sciacca [et al.] // Current pharmaceutical design. – 2008. – V. 14. – P. 3574–3589. 247. Umegaki, M. Peri-infarct depolarizations reveal penumbra-like conditions in striatum / M. Umegaki,Y. Sanada ,Y. Waerzeggers [et al.] // J. Neurosci. – 2005. – V. 25(6). – № 9. – P. 1387–1394. 248. Upadhyay, R.K. Drug delivery systems, CNS protection, and the blood brain barrier / R.K. Upadhyay // BioMed Research International. – 2014. – V. 20. – № 14. – P. 1–37. 249. Venet, F. Regulatory T cell populations in sepsis and trauma / F. Venet, C.S. Chung, G. Monneret [et al.] // J. Leukoc. Biol. – 2008. – V. 83(3). – P. 523–535. 167 250. Viel, J.J. Age- and concentration-dependent neuroprotection and toxicity by TNF in cortical neurons from beta-amyloid / J.J. Viel, D.Q. Mc Manus, S.S. Smith // J. Neurosci. Res. – 2001. – V. 64(5). – P. 454–465. 251. Vila, N. Levels of anti-inflammatory cytokines and neurological worsening in acute ischemic stroke / N. Vila, J. Castillo, A. Davalos [et al.] // Stroke. – 2003. – P. 671–675. 252. Vogelgesang, A. Functional status of peripheral blood T-cells in ischemic stroke patients / A. Vogelgesang, V.E.L. May., U. Grunvald [et al.] // PLoS ONE. – 2010. – V. 5. – P. 1–9. 253. Wang, Q. The inflammatory response in stroke / Q. Wang, X.N. Tang, M.A. Yenari // J. Neuroimmunol. – 2007. – V. 184(1–2). – P. 53–68. 254. Wang-Fischer, Y. Manual of stroke models in rats. – CRC Press Taylor&Francis Group. – 2009. – 352 p. 255. Wen, Y.D. Inflammatory mechanism in ischemic neuronal injury / Y.D. Wen, H.L. Zhang, Z.H. Qin // Neuroscience bulletin. – 2006. – V. 22(3). – P. 171–182. 256. Witt, K.A. CNS drug delivery: opioid peptides and the blood-brain barrier / K.A. Witt, T.P. Davis // The AAPS Journal. – 2006. – V. 8. – № 1. – P. 76–88. 257. Wood, J.D. Function of opioids in the enteric nervous system / J.D. Wood, J.J. Galligan // Neurogastroenterol. Motil. – 2004. – Suppl.2. – P. 17–28. 258. Wood, P.L. Neuroinflammation. Mechanisms and management. – Humana Press Inc. – 2003. – 419 p. 259. Yang, Y. δ-opioid receptor activation attenuates oxidative injury in the ischemic rat brain / Y. Yang, X. Xia, Y. Zhang [et al.] // BMC Biology. – 2009. – V. 55. – № 7. – P. 1–9. 260. Yu A.C.H. Expression of interleukin-1 alpha, tumor necrosis factor alpha and interleukin-6 genes in astrocytes under ischemic injury / A.C.H. Yu, L.T. Lau // Neurochemistry international. – 2000. – V. 36. – P. 369–377. 168 261. Yu, D.H. Pathophysiologic and Immunologic Changes in a Canine Endotoxemia Over a Period of 24 Hours / D.H. Yu, B. Kim, J. Park // J. Vet. Med. Sci. – 2011. – V. 74(5). – P. 537–544. 262. Zaremba, J. Contribution of tumor necrosis factor alpha to the pathogenesis of stroke / J. Zaremba // Folia Morphol. – 2000. – V. 59. – № 3. – P. 137–143. 263. Zhang, H. Endogenous opioids upregulate brain-derived neurotrophic factor mRNA through delta- and micro-opioid receptors independent of antidepressant-like effects / H. Zhang [et al.] // Eur. J. Neurosci. – 2006. – V. 23. – № 4. – P. 984–994. 264. Zhang, M. K. Effects of naoshuningon the plasma of TNF-α, IL-1β and ICAM-1 levels in a rat brain ischemia-reperfusion injuri model / M. Zhang, S. Shen, R. Liu [et al.] // Sci. Res. Essays. – 2011. – V. 6. – № 4. – P. 923–927. 265. Zhu, Y. Transforming growth factor-β1 increases bad phosphorylation and protects neurons against damage / Y. Zhu, G.Y. Yang, B. Ahlemeyer [et al.] // The Journal of Neuroscience. – 2002. – V. 22. – № 10. – P. 3898–3909. 266. Zuidema, M. Y. Iscemia/reperfusion injury: the role of immune cells / M.Y. Zuidema, C. Zhang // World Journal of Cardiology. – 2010. – V. 2(10). – P. 325–332. 169 ПРИЛОЖЕНИЯ 170 Приложение 1 Список опубликованных работ аспиранта кафедры общей и клинической патофизиологии Трофименко Артема Ивановича *1. Трофименко, А.И. Моделирование церебральной ишемии посредством перевязки средней мозговой артерии у крыс / А.И. Трофименко, А.Х. Каде, С.А. Занин, А.В. Бурлаков // Кубанский научный медицинский вестник. – 2011. – №4 (127). – С. 107–110. 2. Трофименко А.И. Получение модели ишемического инсульта посредством коагуляции средней мозговой артерии у крыс / А.И. Трофименко, А.Х. Каде, С.А. Занин, Ю.В. Дынько // Вестник МУЗ ГБ № 2. – 2012. – № 1 (19); URL: http://vestnik.kmldo.ru/pdf/12/01/06.pdf (дата обращения: 23.04.2012). *3. Трофименко, А.И. Особенности электрокардиограммы у крыс с моделью церебральной ишемии, вызванной посредством коагуляции правой средней мозговой артерии / А.И. Трофименко, А.Х. Каде, В.П. Лебедев, С.А. Занин, Т.Б. Заболотских, В.Д. Левичкин, С.Н. Мерзлякова, М.В. Колпаков // Кубанский научный медицинский вестник. – 2012. – № 2 (131) – С. 175–179. *4. Трофименко, А.И. Моделирование церебральной ишемии посредством коагуляции средней мозговой артерии у крыс / А.И. Трофименко, А.Х. Каде, В.П. Лебедев, С.А. Занин, В.В. Мясникова // Фундаментальные исследования. – 2012. – № 2. – С. 215–218. *5. Трофименко, А.И. Характеристика сдвигов в системе про- /антиоксиданты у крыс с моделью острой локальной церебральной ишемии / А.И. Трофименко, В.Д. Левичкин, И.И. Павлюченко, А.Х. Каде, О.С. Охременко, С.А. Занин // Фундаментальные исследования. – 2013. – № 9 (ч. 4). – С. 683– 686. 171 6. Трофименко, А.И. Визуализация очага ишемии головного мозга у крысы тетразолиевым методом / А.И. Трофименко, А.Х. Каде, С.А. Занин, В.Д. Левичкин, А.Ю. Туровая, С.П. Вчерашнюк // Международный журнал прикладных и фундаментальных исследований. – 2013. – № 5. – С. 99. *7. Трофименко, А.И. Динамика цитокинового статуса у крыс при моделировании ишемического инсульта / А.И. Трофименко, А.Х. Каде, В.Д. Левичкин, Ф.А. Нехай, С.А. Занин // Современные проблемы науки и образования. – 2014. – № 2; URL: http://www.science-education.ru/116-12557 (дата обращения: 31.03.2014). *8. Трофименко, А.И. Влияние ТЭС-терапии на показатели системы про-/антиоксиданты у крыс с экспериментальным ишемическим инсультом / А.И. Трофименко, В.Д. Левичкин, Е.И. Ременякина, И.И. Павлюченко, А.Х. Каде, С.А. Занин // Современные проблемы науки и образования. – 2014. – № 2; URL: http://www.science-education.ru/116-12557 (дата обращения: 28.07.2014). 9. Трофименко, А.И. Активность свободно-радикального окисления и эндорфинергической системы у крыс с моделью острой локальной церебральной ишемии / А.И. Трофименко, В.Д. Левичкин, И.И. Павлюченко, А.Х. Каде, О.С. Охременко, Ф.А. Нехай, С.А. Занин // Современные наукоемкие технологии. – 2014. – № 4. – С. 126– 127. 10. Трофименко, А.И. Про-/антиоксидантная активность плазмы крови у крыс с моделью острой локальной церебральной ишемии / А.И. Трофименко, В.Д. Левичкин, И.И. Павлюченко, А.Х. Каде, О.С. Охременко, Ф.А. Нехай, С.А. Занин // Современные наукоемкие технологии. – 2014. – № 4. – С. 142–143. 11. Трофименко, А.И. Влияние ТЭС-терапии на динамику трансформирующего фактора роста-β при острой локальной церебральной ишемии в эксперименте у крыс / А.И. Трофименко, А.Х. Каде, В.Д. Левичкин, Ф.А. Нехай, С.А. Занин // Международный журнал прикладных и фундаментальных исследований. – 2014. – № 5 (ч. 1). – С. 136–137. 172 12. Трофименко, А.И. Влияние ТЭС-терапии на динамику интерлейкина10 при острой локальной церебральной ишемии в эксперименте у крыс / А.И. Трофименко, А.Х. Каде, В.Д. Левичкин, Ф.А. Нехай, С.А. Занин // Международный журнал прикладных и фундаментальных исследований. – 2014. – № 5 (ч. 1). – С. 137–138. *13. Трофименко, А.И. Динамика уровня β-эндорфина при моделировании ишемического инсульта у крыс / А.И. Трофименко, А.Х. Каде, Ф.А. Нехай, В.П. Лебедев, В.Д. Левичкин, С.А. Занин // Кубанский научный медицинский вестник. – 2014. – №3 (145). – С. 115–118. * - работа опубликована в журнале, включенном ВАК при Министерстве образования и науки Российской Федерации в Перечень ведущих рецензируемых научных журналов и изданий 173 Приложение 2 174 Приложение 3