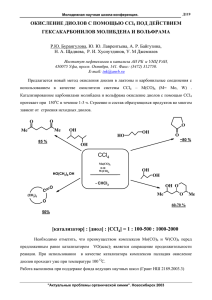
ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ВОЕННОЕ
advertisement

ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ВОЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ «ВОЕННО-МЕДИЦИНСКАЯ АКАДЕМИЯ ИМ. С.М.КИРОВА» МИНИСТЕРСТВА ОБОРОНЫ РОССИЙСКОЙ ФЕДЕРАЦИИ На правах рукописи ЛЕБЕДЕВ Виктор Андреевич ЦЕНТРАЛЬНЫЕ ЭФФЕКТЫ ПОЛИПРЕНОЛСОДЕРЖАЩИХ ПРЕПАРАТОВ 14.03.06 – фармакология, клиническая фармакология Диссертация на соискание ученой степени кандидата биологических наук НАУЧНЫЙ РУКОВОДИТЕЛЬ: доктор медицинских наук профессор П.Д. ШАБАНОВ Санкт-Петербург 2014 2 СОДЕРЖАНИЕ стр. ВВЕДЕНИЕ ……………………………………………………………….. 5 Глава 1. РОЛЬ И ФУНКЦИИ ПОЛИПРЕНОЛОВ В ОРГАНИЗМЕ (АНАЛИТИЧЕСКИЙ ОБЗОР) ……………………………………..…… 13 Глава 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ …………….. 40 2.1. Выбор животных и их содержание …… …………………………… 40 2.2. Вживление электродов в дофаминергические структуры мозга ….. 41 2.3. Методы самораздражения мозга у крыс ……………………………. 43 2.4. Исследование поведения крыс в «открытом поле» ………………… 45 2.5. Исследование функциональной асимметрии мозга с помощью метода ротации …………………………………………………………… 47 2.6. Исследование поведения крыс в приподнятом крестообразном лабиринте …………………………………….……….. 49 2.7. Исследование агрессии в тесте «чужак-резидент» ………………... 50 2.8. Исследование антидепрессантной активности в тесте Порсолта …. 51 2.9. Моделирование экспериментального подострого гепатоза ………. 52 2.10. Биохимическое исследование крови ……………………………… 53 2.11. Гистологическое исследование печени ……….………………….. 54 2.12. Биохимическое исследование мозга ………………………………. 55 2.13. Фармакологические вещества, используемые для анализа двигательных и эмоциональных форм поведения ……………………… 56 2.14. Статистическая обработка полученных материалов …………….. 57 Глава 3. РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ ………. 59 3.1. Исследование поведенческих эффектов полипренолов …………... 59 3.1.1. Влияние ропрена на двигательную активность и поведение крыс в «открытом поле» ………………………………………………….. 59 3.1.2. Влияние полипренолов на двигательную активность и стереотипное (ротационное) поведение крыс …………………………… 61 3 3.1.3. Влияние полипренолов на поведение крыс в приподнятом крестообразном лабиринте …………………………………………….… 62 3.1.4. Влияние полипренолов на показатели агрессии в тесте «чужакрезидент» у крыс …………………………………………………………. 64 3.1.5. Влияние полипренолов на показатели депрессии в тесте Порсолта у крыс …………………………………………………………… 65 3.1.6. Влияние полипренолов на систему мозгового подкрепления в тесте самостимуляции латерального гипоталамуса у крыс ………..… 66 3.1.7. Заключение …………………………………………………………. 70 3.2. Исследование поведенческих и нейрохимических эффектов полипренолов при подостром гепатозе ………………………………….. 71 3.2.1. Характеристика экспериментальной модели …………………….. 71 3.2.2. Влияние полипренолов и адеметионина на выживаемость крыс, подвергнутых воздействию CCl4 …………………….………………….. 75 3.2.3. Влияние полипренолов и адеметионина на поведение крыс, подвергнутых воздействию CCl4 …………………….………………….. 76 3.2.4. Влияние полипренолов и адеметионина на биохимические показатели крови крыс, подвергнутых воздействию CCl4 ……….………………….. 82 3.2.5. Влияние полипренолов и адеметионина на морфологическую картину печени на модели подострого гепатоза, вызванного CCl4 у крыс ……….………………………………………………………... 85 3.2.6. Влияние полипренолов и адеметионина на обмен моноаминов в структурах мозга крыс, подвергнутых воздействию CCl4 ……….…….. 89 Глава 4. ОБСУЖДЕНИЕ ПОЛУЧЕННЫХ РЕЗУЛЬТАТОВ …………… 98 5. ЗАКЛЮЧЕНИЕ…………………………………………………………. 120 6. ВЫВОДЫ………………………………………………………………… 123 7. НАУЧНО-ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ …………………… 125 8. ЛИТЕРАТУРА…………………………………………………………… 126 4 СПИСОК СОКРАЩЕНИЙ 5-ГИУК 5-гидрокси-3-индолуксусная кислота CCl4 четыреххлористый углерод АЛТ аланинтрансаминаза АСТ аспартаттрансаминаза ГВК гомованилиновая кислота ДА дофамин ДОФУК 3,4-диоксифенилуксусная кислота КОМТ катехол-О-метилтрансфераза ЛПНП липопротеиды низкой плотности НА норадреналин СЕР серотонин ЭДТА этилендиаминтетрауксусная кислота 5 ВВЕДЕНИЕ Актуальность исследования Современная фармакология требует открытия и изучения новых нейропротекторов и средств защиты печени, в том числе растительного происхождения. К их числу может быть отнесен и ропрен, получаемый из нейтральной части хвои сосны и ели. И хотя ропрен зарегистрирован и позиционирован, прежде всего, как гепатопротектор, у него обнаружены нейропротекторные свойства при ишемии и токсическом поражении головного мозга (Шабанов П.Д, Султанов В.С., 2012). Ропрен представляет собой смесь полипренолов, природных линейных полимеров, содержащих 8-18 изопреновых единиц, и имеет следующую структурную формулу: СН3 │ Н – (СН2–С =СН–СН2)n–ОН, где n – число изопреновых звеньев от 8 до 18. Препарат разрешен к практическому перименению (регистрационный номер ЛСР-001521/07), его получают по способу, описанному в патенте RU2238291. Ропрен производства ОАО «Фармацевтическая фабрика», СанктПетербург, рассматривается как концентрат полипренолов (с содержанием суммы полипренолов 95%), он рекомендован в качестве гепатопротекторного средства растительного происхождения (Код АТХ05ВА). Показаниями к применению ропрена являются жировая дистрофия печени различной этиологии, гепатит, цирроз печени (в комплексном лечении), токсические поражения печени (алкогольные, наркотические, лекарственные). Использование ропрена стимулирует репаративно-регенерационные процессы в печени, восстанавливает нарушенный гомеостаз, выводит токсины, способствует повышению активности антиоксидантной системы организма. Наряду с этим отмечено стимулирующее действие ропрена на уровень 6 неспецифической резистентности организма (нейтрофильного и моноцитарного звена). В ходе лечения больных с патологией печени было установлено, что ропрен обладает еще иммуномодулирующим действием, оказывает нормализующее влияние на измененный статус организма, особенно на клеточное звено иммунитета, о чем свидетельствует повышение активных Т- лимфоцитов и индекса Тхелперы/Тсупрессоры. Применение ропрена снижает уровень аллергической напряженности организма, что подтверждается снижением циркулирующих иммунных комплексов и эозинофилов крови (Свидерский В.Л. и др., 2006). Вместе с тем, ропрен – это высококонцентрированный препарат полипренолов, применяемый как гепатопротектор в достаточно высоких дозах. Имеются сведения, что малые дозы полипренолов могут проявлять и нейропротекторную активность, особенно при хронических ишемических повреждениях головного мозга (Свидерский В.Л. и др., 2007). Степень разработанности темы Центральные эффекты ропрена и вообще полипренолов изучены недостаточно. Более того, нет точных сведений о мишенях и интимных механизмах действия полипренолов на центральную нервную систему (ЦНС) и высшие функции мозга. Не изучено влияние полипренолов на нейромедиаторные процессы в клетках головного мозга, по-видимому, во многом определяющих психоактивирующее действие препарата. Все указанное позволило сформулировать цель и задачи настоящей работы. Цель и задачи исследования Целью исследования было изучение поведенческих, антитоксических и нейропротекторных эффектов полипренолов у крыс. 7 Задачи исследования: 1) Исследовать влияние полипренолов на поведение крыс в тестах «открытое поле», приподнятый крестообразный лабиринт, «чужак-резидент», ротации, вынужденной беспомощности Порсолта, самостимуляции латерального гипоталамуса. 2) Исследовать поведение, состояние системы крови и морфологию печени крыс с экспериментальным гепатозом и токсической энцефалопатией, вызванной подострым введением четыреххлористого углерода. 3) Исследовать нейропротекторные эффекты полипренолов на модели подострого гепатоза и токсической энцефалопатии, вызванной четыреххлористым углеродом у крыс. 4) Исследовать влияние полипренолов на метаболизм моноаминов (дофамина и серотонина) в двигательных и эмоциогенных структурах головного мозга крыс. Научная новизна Отсутствие четких представлений о системных эффектах полипренолсодержащих препаратов и интимных механизмах их действия позволили автору сосредоточить основное внимание на изучении поведенческого спектра эффектов ропрена и выяснении его нейропротекторных свойств в модели экспериментального токсического поражения головного мозга. Показано, что полипренолы в широком диапазоне доз (2,2-4,3-11,6 мг/кг) обладают умеренными психоактивирующими свойствами, антидепрессантной активностью, способностью изменять подкрепляющие свойства головного мозга. Указанные особенности действия полипренолов на ЦНС связаны, по-видимому, с изменением обмена моноаминов, в частности дофамина, в головном мозге. В действии препарата можно выделить, по крайней мере, два направления: первое связано с эффектами на эмоциональную сферу (депрессивность, подкрепление), второе – с двигательными эффектами препарата (в основном, 8 нормализация разных видов двигательной активности, сниженной вследствие токсических воздействий на мозг). Традиционно эти виды фармакологической активности связывают с влиянием на нейромедиаторные системы мозга. Действительно, полипренолы, применяемые в сравнительно низких дозах (2,2-4,3 мг/кг) повышают активность мезолимбической (прилежащее ядро) и нигростриатной (стриатум) дофаминергических систем у крыс с подострым гепатозом и токсической энцефалопатией. Этот эффект не связан с гепатопротекторными свойствами полипренолов. Данное положение подтверждается фактом, что гепатопротектор адеметионин (гептрал) таким действием не обладает. Сказанное позволяет отнести полипренолы к перспективным современным средствам с выраженным центральным психоактивирующим действием и способностью защищать печень и другие внутренние органы от действия токсикантов типа четыреххлористого углерода. Теоретическая и практическая значимость В результате проведенных исследований получены новые данные о системных нейропротекторных, гепатопротекторных, антитоксических и эмоциогенных свойствах полипренолсодержащих препаратов, в частности, малых доз ропрена. В модельных опытах на крысах, у которых воспроизводили подострый гепатоз введением четыреххлористого углерода доказано, что полипренолы в дозах 2,2-4,3-11,6 мг/кг оказывает нормализующее действие на двигательную и исследовательскую активность, а также эмоциональность крыс, нарушенную CCl4. Препарат сравнения адеметионин (гептрал 300 мг/кг такого действия не оказывает). Полипренолы проявляют умеренное антиагрессивное действие, превышающее таковое для адеметионина, включая высокую степень защиты (защитного поведения) у экспериментальных животных. Полипренолы нормализуют показатели крови, нарушенные введением CCl4. Гепатопротекторное действие полипренолов заключается в нормализации морфологического повреждения печени при эксперименталь- 9 ном гепатозе (белковой и жировой дистрофии), вызванном CCl4. Максимальный гепатопротекторный эффект регистрируется при введении ропрена в дозе 11,6 мг/кг и гептрала в дозе 300 мг/кг (данные сопоставимы статистически). В противоположность адеметионину полипренолы в дозах 2,2-4,3-11,6 мг/кг оказывают умеренное психоактивирующее действие, в основе которого лежит их влияние на активность дофаминергических систем головного мозга: у крыс с подострым гепатозом и токсической энцефалопатией полипренолы повышают активность мезолимбической (прилежащее ядро) и нигростриатной (стриатум) дофаминергических систем, тогда как адеметионин таким действием не обладает. В целом продемонстрирована фармакологическая близость гепатопротекторных эффектов полипренолов (особенно в дозе 11,6 мг/кг) и адеметионина, но принципиальное отличие в их центральных эффектах. Изучение этих и родственных с ними вопросов во многом может приблизить нас к пониманию механизмов действия полипренолсодержащих препаратов и к разработке конкретных рекомендаций по их использованию в неврологической практике. Методология и методы исследования Методология исследования состояла в изучении у крыс поведенческих эффектов полипренолов растительного происхождения в разных дозах (2,24,3-11,6-13,4 мг/кг), оцененных в батарее тестов («открытое поле», приподнятый крестообразный лабиринт, ротационный тест, «чужак-резидент», тест Порсолта, самостимуляция латерального гипоталамуса), моделировании подострого гепатоза с энцефалопатией введением CCl4 с последующей оценкой гепато- и нейропротекторных эффектов полипренолов в сравнении с адеметионином на основании поведенческих тестов, биохимии крови, морфологических изменений в печени и нарушенеий обмена моноаминов (дофамина и серотонина) в головном мозге. Исследования выполнены с соблюдением всех правил доказательной медицины. 10 Основные положения, выносимые на защиту: 1. Полипренолы в дозах 4,3-8,6-13,4 мг/кг при их курсовом введении (5-7 дней) умеренно активируют двигательную активность, проявляют анксиогенное действие, подавляют индивидуальное поведение и оказывают двухфазный эффект (подавление-активация) на поведение самостимуляции в камере Скиннера у интактных крыс. 2. В модели экспериментального подострого гепатоза с энцефалопатией, вызванного четыреххлористым углеродом, полипренолы в дозах 2,2-4,311,6 мг/кг нормализуют двигательную и исследовательскую активность, а также эмоциональность крыс, проявляют умеренное антиагрессивное действие, при этом препарат сравнения адеметионин 300 мг/кг такого действия не оказывает. 3. Полипренолы в дозах 2,2-4,3-11,6 мг/кг, так же как и препарат сравнения адеметионин, нормализуют показатели крови, нарушенные введением CCl4, и восстанавливают нормальную морфологическую структуру печени при токсическом подостром гепатозе (антитоксическое действие). Эффекты полипренолов 11,6 мг/кг и адеметионина 300 мг/кг приблизительно равны. 4. Полипренолы в дозах 2,2-4,3-11,6 мг/кг оказывает центральное психоактивирующее действие, в основе которого лежит его влияние на активность дофаминергических систем головного мозга. Это выявляется в модели подострого гепатоза с токсической энцефалопатией, когда полипренолы повышают активность мезолимбической (прилежащее ядро) и нигростриатной (стриатум) дофаминергических систем. Адеметионин таким действием не обладает. Степень достоверности и апробация материалов исследования Степень достоверности определяется большим количеством экспериментальных животных (306 крыс), рандомизацией и формированием групп 11 сравнения и активного контроля, адекватными поведенческими, токсикологическими, морфологическими, биохимическими и фармакологическими методами исследования, длительными сроками наблюдения и корректными методами статистической обработки. Реализация результатов Материалы исследования используются в лекционном курсе кафедры фармакологии Военно-медицинской академии имени С.М.Кирова, кафедры терапии, нервных болезней и психиатрии Института медицинского образования Новгородского государственного университета им. Ярослава Мудрого. Работа выполнена в соответствии с плановыми научно-исследовательскими разработками Военно-медицинской академии им. С.М. Кирова. Материал диссертации вошел в грантовые разработки Российского фонда фундаментальных исследований РАН (РФФИ №10-04-00473). Апробация результатов Материалы, вошедшие в диссертацию, доложены на научной конференции Российской ассоциации психонейроэндокринологии (СанктПетербург, 2010), международных конференциях «Актуальные проблемы химии природных соединений» (Ташкент, 2010) и «Новые технологии в медицине и экспериментальной биологии» (Рио-де-Жанейро, 2011), 4-ом съезде фармакологов России (Казань, 2012). По теме диссертации опубликованы 3 статьи (все в журналах, рекомендованных ВАК РФ) и 4 тезиса. Апробация диссертации прошла на совместном заседании кафедр фармакологии и НИО обитаемости НИЦ Военно-медицинской академии им. С.М.Кирова. Личный вклад автора Личный вклад автора осуществлялся на всех этапах работы и состоял в планировании экспериментов, их непосредственном выполнении, обработке полученных результатов, обсуждении результатов, написании статей и тезисов, написании диссертации и автореферета. Участие автора в 12 выполнении, сборе и анализе – 95%, статистической обработке – 100%, в написании статей и тезисов – 90%, написании диссертации и автореферата – 95%. Структура и объем диссертации Диссертация состоит из введения, главы обзора литературы, материалов и методов исследования, главы результатов собственных исследований (включающей 2 раздела), обсуждения результатов, выводов, практических рекомендаций, списка литературы. Работа изложена на 141 страницах машинописного текста, иллюстрирована 32 рисунками и 9 таблицами. Библиографический указатель содержит 158 наименований, в том числе 62 отечественных и 96 иностранных. 13 ОСНОВНОЕ СОДЕРЖАНИЕ РАБОТЫ Глава 1. РОЛЬ И ФУНКЦИИ ПОЛИПРЕНОЛОВ В ОРГАНИЗМЕ (обзор литературы) Синтез таких известных антиоксидантов как витамины А, Е, каротиноиды, убихиноны идет через образование и использование изопреновых звеньев. Главным структурным звеном этих соединений является изопрен С5Н8. Изопреноидный путь обмена осуществляется через ацетилкоэнзим А, мевалонаткиназу, полипренолы (долихолы), сквален и далее может приводить к образованию циклических производных – холестерина, стероидов, гормонов, либо к длинноцепочечным линейным полиенам – каротина, каротиноидов, витамина А, либо к построению таких хиноновых соединений, как витамин К2 и убихинон. Участвуя в изопреноидном пути обмена, полипренолы, как транспортные липидсодержащие молекулы, обладают разносторонней биологической активностью. Изопрениловые молекулы убихинона и менахинона являются наиболее важными, так как они являются дыхательными коферментами в системах переноса электронов, как у растений, так и у животных. 1.1. Химия полипренолов Полипренолы, или как их еще называют полиизопреноиды, представляют собой большую группу соединений – линейных 1,5-полиенов с различной конфигурацией тризамещенных связей С≡С с α-изопреновым звеном. Биологически они являются высоко функционально активными веществами. Полипренолы относятся к соединениям с общей структурной формулой (рис. 1) − Н(С5Н8)n ОН, где n=5-24. Большинство изопреновых остатков имеют цис-замещенную двойную связь, и только 2 или 3 в ω-конце цепи представлены транс-конфигурациеей, 14 возрастающей до ди(транс) или три(транс)поли(цис)преноидных алкоголей. В этом отношении они отличаются от полипренолов, таких как соланенол, которые являются предшественниками боковой цепи пластохинонов, убихинонов и менахинонов. Некоторые поли(цис)преноидные алкоголи содержат один или, по крайней мере, небольшое число насыщенных изопреновых остатков. Долихолы (от греч. δολικοσ – длинный) имеют насыщенный αостаток, то есть являются 2,3-дигидрополи(цис)изопреноидными алкоголями. Они представляют собой очень гидрофобные длинные молекулы с n в верхней части ряда, как правило, от 18 до 24. СН3 СН3 СН3 │ │ │ СН3 – С =СН – СН2 – (СН2-С =СН – СН2–)n-2 – СН2 – С = СН – СН2 –ОН ω-остаток α-остаток Рис 1. Общая структура полиизопреноидных спиртов (полипренолов). Термин поли(цис)изопреноидные алкоголи часто сокращается до поли(цис)пренолов или полипренолов в зависимости от контекста. Само содержание полипренолов в ткани не всегда отражает концентрацию соединения в его функциональной форме, которой является монофосфат. Поэтому часто необходимо измерять фосфатазную активность или скорость метаболизма свободных полипренолов, чтобы более точно оценить их функциональную активность. Полипренолы относятся к природным биорегуляторам (физиологически активны), для которых установлены коферментные функции мембраноактивных участников транспорта гидрофильных частиц в процессе биосинте- 15 за полисахаридов, гликопротеидов, пептидогликанов и углеводсодержащих биополимеров. Все бактерии содержат ди(транс)поли(цис)пренолы с 10-12 изопреновыми остатками. Их главный компонент ундекапренол иногда называют бактопренолом. В организме бактерий он присутствует, как правило, в форме фосфоундекапренола или его гликозилированных производных. В частности, такие структуры присутствуют в золотистом стафилококке, Staphylococcus aureus, а также в лактобактериях, Lactobacilli. В Staphylococcus aureus, находящегося в стационарной фазе, свыше 90% ундекапренола (0,03 моль/г клеток) неэстерифицировано (Hemming F.W., 1985). Зеленые листья высших растений содержат три(транс)поли(цис)пренолы, в которых n колеблется в пределах 10-13 изопреновых остатков, обычно большинство компонентов в смеси составляют 11-12 остатков. Например, в работах, выполненных на листьях фикуса, Ficus elastica, большинство полипренолов было представлено фикапренолом-11 и фикапренолом-12. Концентрация этих спиртов в листьях обычно возрастает по мере старения листьев. Иглы сосны разных видов накапливают три(транс)поли(цис)пренолы с колебаниями изопреновых остатков от 13 до 21, которые представлены главным образом ациловыми эфирами. Нефотосинтезирующие ткани высших растений иногда содержат мало отличающиеся поли(цис)пренолы с малым числом изопреновых единиц (6-9), обычно в форме эфиров жирных кислот. Например, в древесной ткани серебристой березы, Betula verricosa, содержатся бетулапренолы-6-9 (Hemming F.W., 1985). Большинство эукариотических клеток содержат 2,3- дигидрополи(цис)пренолы, которые называют долихолами. Почти все они включают 2 транс-остатка в ω-конце цепи. Суммарные данные по распределению долихолов в организме позвоночных представлены в табл. 1. Видно, что наиболее изучено распределение долихолов в организме человека и крысы. Концентрация долихолов резко колеблется в зависимости от органа или 16 ткани. У человека содержание долихолов в тканях в 10-50 раз превышает таковое в организме крысы. Из тканей человека, наиболее богатых долихолами, необходимо выделить гонады (яички), гипофиз и надпочечники, то есть именно те органы, которые осуществляют гормональный контроль функциями. В организме крысы концентрация долихолов наиболее высока в селезенке, поджелудочной железе и печени. Если в человеческом организме содержание долихолов колеблется весьма выражено, то у крысы оно мало отличается от органа к органу. Достаточно высокие концентрации долихолов обнаружены в клетках печени свиньи, причем неэстерифицированные долихолы локализованы главным образом в митохондриях, а эстерифицированные с жирными кислотами – в грубой ядерной фракции. Радиоактивный [3H]долихол, введенный крысам внутривенно, обнаруживается главным образом в печени, на митохондриальной внешней мембране, в меньших количествах – в легких, желудочно-кишечном тракте и селезенке. При введении крысам радиоактивного долихола, смешанного с подсолнечным маслом, сыворотка крови и печень содержали по одной третьей части общей радиоактивности каждая, менее всего – в легких и селезенке. При внутрижелудочковом введении содержание долихола в различных отделах мозга крыс снижалось в ряду: мозжечок > средний мозг > серое вещество > ствол мозга > белое вещество. Таблица 1 Распределение долихолов в организме животных и человека (Hemming F.W., 1985, с изменениями) Животное Орган Концентрация мкг/г массы Бык Щитовидная железа 200 Гипофиз 130-170 17 Курица Человек Яичники 164-316 Печень 182 Кровь > 0,5 Яички 3226 Гипофиз 1400 Надпочечник 1273 Печень 1226 Щитовидная железа 1145 Поджелудочная железа 943 Простата 268 Сердце 262 Почки 240 Селезенка 161 Легкие 82 Свинья Печень 69-129 Кролик Печень 10-40 Крыса Селезенка 106 Поджелудочная железа 26 Печень 23 Головной мозг 17 Легкие 14 Тонкая кишка 14 Почки 13 Яички 11 Сердце 8 Мышцы 4 Кровь <2 Костный мозг <2 18 Кожа <2 1.2. Биосинтез полипренолов Принципиальная схема синтеза полипренолов у прокариотов и эукариотов сходна (Holstein SA, Hohl RJ., 2004). В клетках эукариотов синтез поли(цис)пренолов, как и убихинонов, локализован главным в митохондриях и представлен на рис. 2. Однако ряд этапов биосинтеза требует более подробных пояснений. В частности, этапы 7 и 8 локализованы на внутренней мембране митохондрий, хотя этап 5, связанный с долихолсинтетазой, осуществляется на внешней мембране митохондрий. Этапы 9 и 10 идут на гладком эндоплазматическом ретикулуме. Утилизация фарнезилпирофосфата на этапе 9 происходит в цитоплазме, так же, как и этап 2. Важно отметить, что стереохимия изопреновых остатков полипренолов определяется свойствами ферментов, катализирующих этапы 4, 5 и 7. Рис. 2. Биосинтез дифосфополи(цис)пренолов и связь с другими путями синтеза изопреноидов (Hemming F.W., 1985) х и n означают число изопреновых остатков в каждой молекуле. 19 1.3. Роль полипренолов в переносе сахаров (гликозилировании). Как уже отмечалось выше, фосфополи(цис)пренолы выполняют функцию коферментов в нескольких реакциях гликозилирования. Ферменты, которые участвуют в этом процессе, относятся к категории связанных с мембраной гликозилтрансфераз, сама система достаточна проста. Переносимый сахар присутствует в форме растворенного в воде дифосфата сахара, выступающего в роли донора. Он переносится из цитоплазмы связанной с мембраной трансферазой, которая катализирует его перенос на монофосфополи(цис)пренол, также связанный с мембраной. Поскольку эта реакция предусматривает обмен одного сахар-1-фосфата на другой, реакция обратима и потенциал переноса фосфата сахара остается в этом связанном с липидом промежуточном продукте. Гликозилтрансфераза использует этот гидрофобный донор в качестве субстрата, катализирующего перенос остатка сахара на специфический акцептор для связывания гликозилированием и высвобождает монофосфополи(цис)пренол для следующего взаимодействия с трансферазой (McTaggart SJ., 2006; Hemming F.W., 1985). Выше мы рассмотрели общую схему участия фосфополи(цис)пренолов в гликозилировании (переносе сахаров). Однако эта схема вполне переносима на комплекс полисахаридов бактериальных клеток, где фосфополи(цис)пренолы также выполняют роль коферментов, обеспечивая формирование пептидогликанов (главного компонента бактериальной клетки) как у грам-положительных, так и у грам-отрицательных бактерий. Кроме того, фосфополи(цис)пренолы обеспечивают N-гликозилирование белков (через остаток аспарагина), О-гликозилирование у растений (олигоманнозильную группу на серин и треонин), а также гликозилирование (маннозилирование) ретинола, который также является одним из полипренолов – аллильным изопреноидным алкоголем (Burda P., Aebi M., 1999). Схематически Nгликозилирование, С- и О-манозилирование в листьях растений представлены на рис. 3. 20 Рис. 3. Схематическое представление путей N-гликозилирования, С- и Оманозилирования (А) Предшественник олигосахарида структуры Glc3Man8GlcNAc2 (глюкозаманноза-N-ацетилглюкозамин) расположен на внешней и внутренней сторонах листков эндоплазматического ретикулума (ER). Олигосахарид переносится на протеин с помощью олигосахаридтрансферазы. Число требуемых молекул долихолфосфата показано под рисунком. (В) Белки модифицируются с помощью якорей гликозилфосфатидилинозитола (GPI), С- или Оманнозилирования (обозначены как а, b, с соответственно) на внутренней поверхности листков эндоплазматического ретикулума (ER) (Denecke J., Kranz C., 2009). Таким образом, представленные данные подчеркивают универсальный характер участия полипренолов в гликозилировании (переносе сахаров), что является важнейшей биохимической реакцией, без которой не мыслимо осуществление ни одной функции организма – будь то бактерия, растение, животное или человек. 21 1.4. Функциональная роль полипренолов в организме В организме животных роль поли(цис)пренолов выполняют их 2,3дигидропроизводные – долихолы, представляющие собой длинноцепочечные поли(цис)изопреновые спирты. Синтез долихолов осуществляется из мевалоната посредством мевалонаткиназы (см. выше рис. 1). Долихолы регулируют N-гликозилирование белков. Активной является фосфорилированная форма долихолов (Adair W.L., Keller R.K., 1982). Долихолы располагаются внутри фосфолипидного бислоя мембран, но их точное расположение неизвестно, так как их движение и распределение варьирует в зависимости от геометрии мембраны. Предполагается, что они определяют и модифицируют текучесть, стабильность и проницаемость мембран. Долихол может действовать как радикальный поглотитель образующихся на мембране перекисных липидов. Некоторые данные свидетельствуют, что, будучи эндогенным липофильным поглотителем, долихол вместе с витамином Е, возможно, является интегральным элементом антиокислительного механизма клеточных мембран. Полагают, что долихол может взаимодействовать с витамином Е, образовывая хорошо отлаженную цепь по перемещению свободных радикалов, а нарушения в работе этой цепи влекут за собой молекулярно-деструктивные процессы в патогенезе многих заболеваний человеческого организма (Владимиров Ю.А., 2000; Bergamini E. et al., 2004). В качестве примеров приведем ряд фактов, свидетельствующих о развитии клеточных повреждений вследствие активации перекисного окисления липидов (ПОЛ). В клетках и, в особенности, в субклеточных органеллах митохондриях, где дыхательная цепь генерирует основную массу активных форм кислорода, окисление тиоловых групп продуктами ПОЛ приводит к появлению в мембранах различных дефектов, повышающих ее проницаемость (пор). Под действием разности электрических потенциалов на мембранах сквозь такие поры в клетки входят ионы натрия, а в митохондрии – ионы 22 калия. Эти процессы повышают осмотическое давление внутри клеток и митохондрий и вызывают их набухание, что еще больше повреждает мембраны. Генерализация ПОЛ постепенно делает мембраны более твердыми, хрупкими и проницаемыми («протекающими»), лишая их способности нормально реагировать с окружающей средой (Тодоров И.Н., Тодоров Г.И., 2003). Продукты ПОЛ могут также поражать ионтранспортные ферменты, такие как Са2+-АТФазы, окисляя тиоловые группы их активного центра. Эти события ведут к замедлению откачивания ионов кальция из клетки и одновременно к ускорению входа кальция в клетку, что сопровождается повышением внутриклеточной концентрации ионов кальция и повреждению клетки. Под влиянием ПОЛ мембраны митохондрий делаются проницаемыми для ионов водорода и кальция, в результате чего эти органеллы теряют способность к синтезу АТФ, и клетка оказывается в условиях энергетического голода (Владимиров Ю.А., 2000). Эти примеры можно продолжить, но для повествования представляется более важным рассмотреть участие полипренолов в процессах регуляции ПОЛ и антиоксидантной защиты. 1.5. Фармакологическое значение полипренолов и их производных Прежде всего, вернемся к публикации Е. Бергамини и соавторов (Bergamini E. et al., 2004), работающих в Пизе (Италия). Эти авторы подметили, что уровень долихолов, как и холестерина, с возрастом неуклонно растет. Низкокалорийная диета несколько замедляет связанные с возрастом изменения в распределении долихолов в организме. Более того, авторы представили доказательства, что долихолы могут выступать как поглотители активных форм кислорода и нейтрализаторы ПОЛ мембран клеток.. Описывая этот процесс, они подчеркивают, что долихолы взаимодействуют с ненасыщенными жирными кислотами и витамином Е (наиболее мощным биологическим антиоксидантом). По сути, долихолы предохраняют от прогрессирования клеточных нарушений, приводящих к атеросклерозу и нейродегенеративным 23 заболеваниям (Farooqui T, 2009; Eckert GP, 2009). Задолго до исследований E. Bergamini и соавторов (2004) аналогичное наблюдение было выполнено R.K.Pullatkat и H. Reha (1982). Они выявили, что за время жизни уровень долихолов в головном мозге увеличивается почти в 15 раз: от 18 мкг/г ткани у 6-летних детей до 263 мкг/г ткани у 68-летних пожилых людей. Следовательно, с возрастом происходит интенсификация образования и накопления долихолов, которые, по-видимому, осуществляют защитные функции, нейтрализуя некоторые химически агрессивные соединения, например, активные формы кислорода. По мнению R.R.Kurup и P.A.Kurup (2003), в результате изопреноидного обмена могут образовываться три главных метаболита: 1) дигоксин – эндогенный ингибитор мембранной Na+-К+-АТФазы (речь идет об эндогенных дигиталисоподобных соединениях, имеющих видовую специфичность) ; 2) убихинон – мембранный антиоксидант и вещество, способствующее прохождению транспорта электронов через мембрану и улавливанию свободных радикалов, и 3) долихол, осуществляющий N-гликозилирование сахаров и белков. Этот вывод авторы сделали на основании большого клинического материала, полученного в процессе изучения различных форм патологии, связанных с нарушениями изопреноидного обмена, где в качестве маркеров они изучали не только изменения уровня в сыворотке крови дигоксина, убихинона и долихола, но и магния, а также активность 3-гидрокси-3-метилглутарилКoA-редуктазы, Na+-К+-АТФазы, соотношение холестерин/фосфолипиды, кальций/магний, уровень лизосомальных ферментов, свободных радикалов при различных заболеваниях. Как отмечают эти авторы, в клинической практике нарушения метаболизма изопреноидного пути наблюдаются довольно часто при различных нейродегенеративных заболеваниях ЦНС, включая болезнь Альцгеймера, при хронических заболеваниях внутренних органов, например, хронической эмфиземе легких, идиопатическом фиброзе легких, саркоидозе легких, бронхиальной астме, язвенном колите, алкогольном цир- 24 розе печени и гепатолентикулярной дегенерации, пептических язвах и других заболеваниях желудочно-кишечного тракта, дегенеративных заболеваниях нервно-мышечной системы, остеопорозе, остеоартрите, спондилезе, бесплодии. Кроме того, R.R.Kurup и P.A.Kurup (2003) уверены, что нарушения обмена изопреноидов встречаются у больных при тромбозе сосудов и закупорке артерий, иммунодефицитных состояниях (ревматизм, стрептококковая инфекция), эритроматозной волчанке, зависимости от еды (булимия и анорексия), болезни Крейтцфельда-Якобса и других патологиях. R.R.Kurup и P.A.Kurup опубликовали более десятка работ в Международном журнале нейронаук (2003), в каждой из которых подчеркивают значение полипренолов в развитии указанной патологии. Действительно, многие из этих наблюдений подтверждаются. Например, в исследовании Н.Я.Григорьевой и А.М.Моисеенкова (Grigor’eva N.Ya., Moiseenkov A.M., 1989) продемонстрирована антиульцерогенная активность многих полипренолов и их производных, что стало предметом изобретения. Результаты других исследований показали важную роль долихолов при нарушении холестеринового обмена (Bentinger M. et al., 2008), когда наблюдается высокое накопление холестерина и липидов низкой и очень низкой плотности в сосудах и других органах, что взаимосвязано с повышением концентрации долихолфосфата в результате ингибирования транспорта нейтрального спирта долихола из эндоплазматического ретикулума в лизосомы. Они играют, по-видимому, важную роль в поддержании правильного липидного состава мембран (Jones P.H., 2003). У пациентов с болезнью Альцгеймера содержание долихолов в 10 различных областях мозга оказалось на 20-50% меньше, чем у здоровых людей (Parentini I, 2005; Cole SL, 2006;Hooff G.P. et al., 2008, 2010). При нарушении синтеза долихолов (расстройствах гликозилирования), происходящего на эндоплазматическом ретикулуме или в цитоплазме, возможны серьезные врожденные дефекты. Так, в работе E. Morava и соавторов 25 (2010) описан врожденный мозжечково-глазной синдром, в состав которого входит мозжечковая атаксия, анемия, ихтиоформный дерматит, дисфункция печени, нарушения свертывания крови и прогрессирующие офтальмологические признаки в форме гипоплазии глазного нерва, отслойки сетчатки, врожденной катаракты и глаукомы. Причина данной патологии не ясна. Исследуя 12 пациентов из 9 семей с признаками мозжечковой атаксии и врожденной патологией глаз, авторы выявили наличие у пациентов (детей) мутации гена SRD5A3, кодирующего стероидную 5α-редуктазу типа 3. Это приводит к нарушениям N-гликозилирования сахаров и белков, то есть синтезу долихолов. Данное исследование еще раз подтвердило важность правильного синтеза долихолов для обеспечения здоровья человека. Синтез долихолов может нарушаться на разных уровнях (Chojnacki T, Dallner G, 1988).. Во-первых, чаще всего это происходит на уровне образования мевалоновой кислоты. Так действуют статины, блокирующие активность 3-гидрокси-3-метилглутарил-КoA-редуктазы. К ним относится большая группа гиполипидемических средств, применяемых для лечения атеросклероза (аторвастатин, симвастатин, правастатин, ловастатин, флувастатин, розувастатин). Эта группа препаратов квалифицируется как наиболее активная из всех гиполипидемических препаратов (Jones P.H. et al., 2003). Однако статины крайне редко рассматриваются с позиции возможного торможения долихолов. Единственным связующим звеном между долихолами и холестерином является факт взаимосвязанного снижения содержания обоих сложных спиртов в организме под влиянием лечения статинами (Espenshade PJ, Hughes AL., 2007). Второй мишенью действия лекарственных веществ в цепочке биосинтеза долихолов является фарнезилфосфат (Mo H., Elson C.E., 2004). Именно это звено является привлекательным для создания противоопухолевых средств, учитывая, что одним из путей превращения фарнезилфосфата является его трансформация в геранилгеранилфосфат, осуществляющий N- 26 гликозилирование рецепторов фактора роста. Действительно, препараты, угнетающие мевалонатпирофосфатдекарбоксилазу (фенилацетат натрия, фенилбутират натрия) показали высокую противоопухолевую эффективность в доклинических и клинических (фазы I-II) исследованиях. Средств угнетения фарнезилпротеинтрансферазы (следующего этапа синтеза долихолов) оказалось больше, чем испытанных на предыдущем этапе. Ими стали типифарниб и экспериментальные соединения под шифрами R115777, SCH66336, BMS214662, L-778,123. Эти соединения показали разные результаты, но в целом антибластомная активность их не вызывала сомнений. Другими исследованиями было показано, что соединения, влияющие на фарнезилпротеинтрансферазу, обладают антивирусной активностью в отношении вируса гепатита D (Einav S., Glenn J.S., 2003). И, наконец, еще одним перспективным антибластомным средством является периллил алкоголь, также угнетающий фарнезилпротеинтрансферазу. Экспериментальные и клинические исследования подтвердили его эффективность в качестве средства лечения и профилактики онкологических заболеваний (Mo H., Elson C.E., 2004). В зависимости от количества изопреновых единиц, заместителей, насыщенности двойных связей, эти соединения способны проявлять различное иммуномодулирующее и метаболическое действие на организм. Иммуномодулирующее действие полипренолов на организм, как предполагают, определяется способностью этих веществ стимулировать образование нейтрофилов и активировать макрофаги ретикулоэндотелиальной системы. Некоторые синтетические полипренолы были запатентованы японской фирмой «Эйсан» в качестве препаратов для профилактики и лечения болезней, обусловленных иммунодефицитом (бронхиальная астма, ревматоидный артрит и др.), а также в сочетании с антибиотиками при различных инфекционных заболеваниях для повышения иммунитета. Пониженный уровень долихолов наблюдается при остром ревматизме и других иммунодефицитных состояниях (Buhaescu I., Izzedine H., 2007). 27 Иммуномодулирующее действие полипренолов было доказано рядом авторов на модели вируса гриппа и при нейроинфекциях. Эффективность полипренолов выявлена при профилактическом введении их за двое суток до аэрогенного инфицирования мышей вирусом гриппа А. Кроме того, имеются данные, указывающие на то, что у животных, зараженных вирусом Сендай, регистрируется более высокий уровень интерферонов в ткани легких при введении им дигидрогептапренола, одного из производных полипренолов. У некоторых из полипренолов, полученных как путем синтеза, так и выделенных из растительного сырья (гингко билоба, сосна и др.) выявлена противоязвенная активность (Grigor’eva N.Ya., Moiseenkov A.M., 1989), некоторые из этого класса соединений обладают антитромботической активностью (четвертичные моноаммониевые соли диаминов). Синтезированные различными способами полипренолы могут обладать также противоопухолевой активностью (например, при гепатоканцерогенезе) (Holstein SA, 2010). Их низкая токсичность делает этот класс соединений перспективным при лечении различных видов опухолей (Mo H., Elson C.E., 2004). Большую перспективу представляют изопреноиды, созданные путем химического или микробиологического синтеза (Klein-Marcuschamer D. et al., 2007). Среди них выделяются средства растительного происхождения, влияющие на мевалонатный путь (каротиноиды, моно-, ди- и тритерпены, токоферолы, гиббереллины, производные хлорофилла, стеролы) (Kirby J., Keastling J.D., 2009). Наконец, полиизопреноидные спирты, кислоты и их амиды способны восстанавливать функцию печени, что позволило запатентовать эти соединения как средства для лечения заболеваний печени, вызванных воспалением, перерождением, а также нарушениями в жировом и углеводном обмене. 28 1.6. Гепатопротекторы синтетического и растительного происхождения. Гепатопротектор ропрен Многообразие нарушений функций печени существенно осложняет задачу медикаментозной коррекции таких состояний и требует применения фармакологических средств широкого спектра действия. К тому же следует иметь в виду, что в патогенезе самых различных заболеваний печени важную роль играют иммунологические сдвиги, сопровождающие патологию этого органа. В соответствии с современными принципами лечения заболеваний печени, программа комплексной терапии такой патологии включает два основных направления (Подымова С.Д., 1998). Первое представляет этиотропную терапию, направленную на подавление патологического возбудителя, его элиминацию и санацию организма. Второе направление соответствует патогенетической терапии, имеющей целью адекватную фармакологическую коррекцию универсальных, мультифакторных и разнесенных во времени звеньев патогенеза. При этом нужно отметить, что универсальность основных звеньев патогенеза различных поражений печени позволяет при всей полиэтиологичности данной патологии использовать достаточно близкую патогенетическую терапию, основу которой могут составлять лекарственные средства с направленным действием на печеночные клетки. Основной целью этиотропной терапии хронических гепатитов является удаление из организма этиологического фактора и продукция адекватного иммунного ответа (Подымова С.Д., 1998). Комплексная терапия таких больных должна учитывать тип возбудителя, степень активности патологического процесса, ведущего клинического синдрома, наличие признаков цирротической стадии, а при наличии вируса – фазы вирусной инфекции – репликации или интеграции (Оковитый С.В. и др., 2002). В этом отношении ведущее значение имеет система интерферонов, обладающих прямой и обратной связью с иммунной системой организма. Противовирусное действие интерферонов 29 осуществляется через клеточный синтез нуклеиновых кислот с помощью ряда ферментов и ингибиторов, приводящих к деградации чужеродной генетической информации. Среди патогенетических средств лечения заболеваний печени традиционно выделяют группу гепатопротекторов растительного происхождения (Пастушенков Л.В. и др., 1990; Пастушенков Л.В., Лесиовская Е.Е., 1995). В методической работе П.Д.Шабанова (2002) рассмотрены вопросы использования таких гепатопротекторов для лечения заболеваний печени в основном токсического и алкогольного происхождения. Дана классификация гепатопротекторов. Подробно описаны механизмы действия гепатопротекторов растительного происхождения. Специальный раздел посвящен использованию нового гепатопротектора растительного происхождения маргали в комплексном лечении паренхиматозных поражений печени. На большом фактическом материале (более 100 больных с алкогольными гепатитами и циррозами) доказана эффективность клинического применения маргали, используемого как гепатопротектор в течение 3-недельного курса лечения. У больных, получавших лечение с помощью комплексного препарата маргали, отмечено значительное улучшение самочувствия, нормализация биохимических показателей печени, иммунного статуса, уменьшение размеров желчного пузыря. Сделан вывод о возможности включения препарата маргали в комплексную терапию гепатитов и циррозов алкогольного происхождения. Однако современная гепатология требует открытия и изучения новых гепатопротекторных средств, в первую очередь растительного происхождения. К их числу отнесен и ропрен, полученный из нейтральной части хвои сосны и ели. Ропрен представляет собой группу полипренолов, точнее смесь полипренолов, содержащих 8-18 изопреновых единиц и имеет следующую структурную формулу: 30 СН3 │ Н – (-СН2-С =СН-СН2-)n-ОН, где n – число изопреновых звеньев от 8 до 18. Лекарственный препарат ропрен зарегистрирован Минздавсоцразвития РФ (ЛСР-001521/07 от 12.07.2007 г.), его получают из хвои сосны и ели по способу, описанному в патенте (RU2238291). Ропрен производства ОАО «Фармацевтическая фабрика СанктПетербурга» (ГаленоФарм), представляет собой концентрат полипренолов (с содержанием суммы полипренолов 95%), рекомендован в качестве гепатопротекторного средства растительного происхождения (Код АТХ05ВА). Показаниями к применению ропрена являются жировая дистрофия печени различной этиологии, гепатит, цирроз печени (в комплексном лечении), токсические поражения печени (алкогольные, наркотические, лекарственные). Использование ропрена стимулирует репаративно-регенерационные процессы в печени, восстанавливает нарушенный гомеостаз, выводит токсины, способствует повышению активности антиоксидантной системы организма. Наряду с этим отмечено стимулирующее действие ропрена на уровень неспецифической резистентности организма (нейтрофильного и моноцитарного звена). В ходе лечения больных с патологией печени было установлено, что ропрен обладает еще иммуномодулирующим действием, оказывает нормализующее влияние на измененный статус организма, особенно на клеточное звено иммунитета, о чем свидетельствует повышение активных Т- лимфоцитов и индекса Тхелперы/Тсупрессоры. Применение ропрена снижает уровень аллергической напряженности организма, что подтверждается снижением циркулирующих иммунных комплексов и эозинофилов крови. Ассортимент лекарственных средств, применяемых в комплексной терапии заболеваний печени и желчевыводящих путей, насчитывает более 1000 31 наименований. Среди такого многообразия препаратов выделяют сравнительно небольшую группу оказывающих избирательное защитное действие на печень – гепатопротекторов. Их действие направлено на восстановление гомеостаза в печени, повышение устойчивости органа к действию патогенных факторов, нормализацию функциональной активности и стимуляцию репаративно-регенерационных процессов в печени. Основные требования к идеальному гепатопротектору можно представить в следующем виде: • препарат должен достаточно полно абсорбироваться; • обязательно наличие эффекта «первого прохождения» через печень; • у препарата должна быть выраженная способность связывать или предупреждать образование высокоактивных повреждающих соединений; • наличие противовоспалительных свойств; • наличие способности подавлять процесс фиброгенеза; • препарат должен стимулировать регенерацию печени; • у соединения должен быть естественный метаболизм при патологии печени; • важна экстенсивная энтерогепатическая циркуляция; • невысокая токсичность. К сожалению, на сегодняшний день ни один из использующихся в медицинской практике гепатопротекторов не удовлетворяет в полной мере этим требованиям, хотя в последние годы арсенал современных гепатозащитных средств и расширился как за счет появления синтетических препаратов, так и новых природных средств. К основным группам гепатопротекторов следует отнести: • препараты растительного и животного происхождения (в первую очередь препараты из расторопши пятнистой, ропрен, эссенциале); 32 • стероидные и нестероидные синтетические анаболические средства (оротат калия, метилурацил, пентоксил, станозолол и др.); • препараты, восстанавливающие целостность мембран гепатоцитов (эссенциале, тропофар); • антиоксиданты витамин Е (aльфа-токоферол), селенит натрия, супероксиддисмутаза; • простагландины и факторы роста (простенон, илопрост, человеческий фактор роста, рекомбинантный человеческий фактор роста); • средства, влияющие на активность микросомальных ферментов печени (диэтилтиокарбамат, метирапона гидротартрат и пиперанил бутоксид); • витамины и гормоны: цианкобаламин (витамин В12), фолиевая кислота (витамин Вc), пиридоксин (витамин В6), липоевая кислота и её препараты (липамид, тиоктацид), глюкокортикоиды, глюкагон; • адаптогены (препараты лимонника, заманихи, аралии, женьшеня, родиолы, элеутерококка, пантокрин, получаемый из пантов марала и рантарин из пантов северного оленя, дибазол (бендазол), бемитил, этомерзол (томерзол); • препараты различных групп: S-аденозил-L-метионин (гептрал), урсодеокисихолевая кислота, производные никотиновой кислоты, изопропоксигерматран, даларгин, убихинон-10, оксикоричные кислоты, антагонисты кальция. Остановимся более подробно на препаратах растительного и животного происхождения. Эта группа средств традиционна для лечения заболеваний печени и прочно занимает одно из ведущих мест в терапии поражений печени инфекционного и неинфекционного происхождения. Лечебное действие многих видов лекарственных трав, применяющихся в настоящее время в народной и научной медицине, связано с наличием в них различных 33 биологически активных веществ: алкалоидов, гликозидов, кумаринов, сапонинов, эфирных масел, витаминов и т. д. В основе действия препаратов растительного происхождения на процессы регенерации печени лежит либо прямое активирующее влияние на синтез РНК и белка, либо вторичный опосредованный эффект, связанный с наличием у данных средств противомикробной, противовоспалительной, желчегонной и антиоксидантной активности. У многих препаратов первичное и вторичное действие могут сочетаться. Препараты растительного происхождения нашли широкое применение в клинической гепатологии (Соколов С.Я., Замотаев И.П., 1990). Наиболее часто в настоящее время используются лечебные средства, созданные на основе расторопши пятнистой: силибинин, силибор, легалон (силибин, силимарин). Силибор, содержащий флаволигнаны (силибинин, силидианин, силихристин) и флавонолы, оказывает отчетливое гепатозащитное действие при вирусном гепатите, повреждениях печени, механической желтухе, однако полного восстановления функциональной активности митохондриальных и микросо- мальных ферментных систем при использовании этого препарата все же не происходит (Машковский М.Д., 2008; Оковитый С.В., 2002). Достаточно высокую гепатозащитную активность при экспериментальных отравлениях проявляет индивидуальный флаволигнан силибинин (силимарин). Препарат дает антиоксидантный эффект и подавляет перекисное окисление полиненасыщенных жирных кислот в составе мембран фосфолипидов, усиливает репаративные процессы. Улучшая функцию мембран и течение метаболических процессов в повреждённой паренхиме печени, эти препараты препятствуют развитию гидропической и жировой дистрофии, некрозов, стимулируют желчеобразование, синтез альбуминов и факторов свертывания крови. В то же время имеются работы, доказывающие, что силибор, карсил и легалон в низких дозах обладают слабым гепатопротективным и выраженным холеретиче- 34 ским действием, а в высоких дозах угнетают секреторно-экскреторную функцию печени. В механизме действия комплексного растительного препарата ЛИВ-52 также имеют значение ослабление перекисного окисления липидов и нормализация соотношения отдельных фосфолипидных фракций в мембранах печеночных клеток, в частности уменьшается количество гепатотоксичного лизолейцина. Установлено позитивное влияние на регенерацию печени суммы фенольных соединений арники горной и арники облиственной, препарата солянки холмовой (лохеин), высушенных извлечений из травы пастушьей сумки, шлемника байкальского, экстракта родиолы розовой, горечавки желтой, настоя эффективность травы при астрагала гепатите, шерстистоцветкового. вызванном Показана тетрахлорметаном, терпеноидных кумаринов (производных гальбановой ряда кислоты), которые ингибируют перекисное окисление липидов, улучшают углеводный обмен, восстанавливают скорость секреции желчи. В лечении гелиотринового гепатита с успехом был использован новый анаболический препарат, относящийся к группе фитоэкдистероидов и представляющий собой природное соединение, получаемое из корней и соцветий ряда видов левзеи. В качестве действующего начала этот препарат содержит экдистерон. Действие его подобно действию стероидных гормонов, но в отличие от последних, он не обладает тимолитическим действием. Кроме этого, у него полиферментных обнаружена систем способность повышать мембран стабильность митохондрий печени, мембраностабилизирующая активность. Особый интерес для клинической гепатологии представляют антиоксиданты флавоноидной природы. Новое производное цианиданола – катерген – представляет собой полусинтетический флавоноид растительного происхождения, гепатопротективное действие которого доказано на 35 основании предупреждения и уменьшения токсического влияния на печень таких ядов как Д-галактозамин, тетрахлорметан (ТХМ), фаллоидин. Эффект препарата обусловлен тем, что он может связывать свободные радикалы, высвобождаемые многими гепатотоксическими веществами. Катерген также стимулирует синтез АТФ в печени, что облегчает протекание биохимических реакций, связанных с затратами макроэргов. Помимо этого он обладает мембраностабилизирующим действием, уменьшая проницаемость клеточных мембран для низкомолекулярных водорастворимых соединений (Оковитый С.В. и др., 2002). Перспективно изучение и использование в качестве гепатопротекторов веществ, являющихся продуктами пчеловодства. Обнаружено репаративное действие у фенольного гидрофобного препарата прополиса, содержащего в своем составе комплекс фенольных соединений (флавоны, флавоноиды, фенолкарбоновые кислоты и др.) и обладающего умеренной антиоксидантой активностью, желчегонным и противовоспалительным действием. Установлено замедление образования продуктов перекисного окисления липидов при гепатите, вызванном тетрахлорметаном, спиртовым экстрактом пчелиной пыльцы. При Д-галактозаминовом гепатите показана активность препарата эплира, содержащего комплекс фосфолипидов и каротиноидов лечебной грязи. Среди средств, стимулирующих процессы регенерации, есть ряд препаратов гидролизаты животного ткани высокомолекулярных происхождения. печени. Такие компонентов, Обычно препараты иногда это экстракты состоят обладают из или многих ферментативной активностью, нередко содержат низкомолекулярные вещества. Механизм их репаративного действия сложен, может иметь много различных звеньев, часть из которых представляет прямое активирующее влияние на процессы регенерации, а часть – опосредованное. 36 Активация синтеза белка препаратами животного происхождения может осуществляться за счет действия входящих в них веществ пептидной или нуклеиновой природы, обладающих соответствующими регулирующими свойствами. Стимуляция репаративных процессов некоторыми препаратами объясняется также включением их компонентов в синтезируемые соединения. При этом по субстратному механизму усиливается новообразование ферментов, участвующих в синтетических процессах. Хотя, разумеется, возможны и другие механизмы действия средств животного происхождения. В современной гепатологии из средств этой группы нашли применение такие препараты как витогепат, сирепар, вигератин и тропофар. Тропофар содержит полипептидные компоненты клеточных мембран гепатоцитов, экстрагированных из печени крупного рогатого скота. Как показали экспериментальные исследования и клинические наблюдения, эффект тропофара основан на его способности уменьшать лечебный повреждение структурных компонентов мембран гепатоцитов, стимулировать процессы протеинсинтеза, предупреждать цитолиз гепатоцитов (Оковитый С.В. и др., 2002). Сирепар и витогепат представляют собой сочетание гидролизатов печени крупного рогатого скота и комплекса витаминов пластического обмена. Эти препараты при отравлении парацетамолом ослабляют продукцию диеновых конъюгатов и малонового диальдегида, снижают проницаемость мембран гепатоцитов для поступления в кровь аланинаминотрансферазы, аспартатаминотрансферазы, щелочной фосфатазы, усиливают секрецию желчи. Близкий по своему составу к этим препаратам вигератин содержит ещё и панкреатин – препарат поджелудочной железы. Все перечисленные выше лекарственные средства эффективны при хроническом гепатите, циррозе печени и токсическом поражении органа. Гепатопротекторная и антиоксидантная активность этих средств, повидимому, обусловлена наличием в их составе токоферолов, флавоноидов, 37 цианокобаламина и других биологически активных веществ, оказывающих многостороннее положительное действие на печень. В последние годы в качестве гепатопротектора стал активно применяться адеметионин (гептрал). Адеметионин представляет собой биологически активное вещество, входящее в состав всех тканей и жидких сред организма. Он участвует в реакциях трансаминирования как донор метильных групп, а также является предшественником тиоловых соединений (цистеин, таурин, глутатион и др.). Адеметионин обладает антидепрессивной активностью, которая проявляется постепенно, начиная с конца 1-й недели лечения. У больных с ипохондрической депрессией наблюдается регрессия симптомов заболевания. Препарат обладает также гепатопротекторной активностью. При его длительном применении отмечается улучшение показателей функции печени. У больных с алкогольным циррозом отмечено повышение содержания глутамина в печени, повышение концентрации цистеина и таурина в плазме и понижение метионина в сыворотке, что свидетельствует о нормализации метаболических реакций в печени (Бурбелло А.Т., Шабров А.В., 2007). Анализируя все представленные данные, можно заключить, что существующие лекарственные средства для стимуляции процессов регенерации в печени не полностью удовлетворяют потребности клинической практики. Некоторые препараты обладают нежелательными побочными эффектами (стероидные анаболические средства). Другие пока мало изучены в гепатологии и дороги (гепатопоэтины). Многие средства, хотя и вполне безопасны, но не всегда достаточно эффективны (витамины, нестероидные анаболические средства). Лекарственные вещества – экстракты животного и растительного происхождения, содержащие высокомолекулярные вещества, – несут потенциальную опасность развития аллергических реакций. Поэтому актуальным остаётся поиск новых высокоэффективных и безопасных гепатопротекторов и активаторов регенераторных процессов в 38 печени (Шабанов П.Д., Калишевич С.Ю., 1998; Шабанов П.Д. и др., 2008, 2011). Предпочтение здесь следует отдавать средствам, стимулирующим синтез нуклеиновых кислот и белка. Перспективно и изучение препаратов, обладающих иммуномодулирующими свойствами. К ним можно отнести и новый препарат ропрен, получаемый путем хроматографической очистки из хвои ели Picea abies (L.) Karst. По сути, он представляет собой природный биорегулятор, или транспортный липид, который в печени быстро метаболизируется в долихол, участвующий в гликозилировании мембранных белков и образовании гликопротеинов. Наибольшая скорость метаболизма долихолов наблюдается в печени, почках, селезенке. Выведение долихолов с желчью считается главным путем выведения их из организма. Таким образом, ропрен может активно компенсировать нарушения метаболизма изопреноидов в организме. В клинической практике нарушения метаболизма изопреноидного пути наблюдаются довольно часто при различных нейродегенеративных заболеваниях ЦНС, органов желудочно- кишечного тракта, иммунодефицитных состояниях, при таких заболеваниях как язвенный колит, алкогольный цирроз печени, гепатоцеребральная дегенерация печени, пептические язвы и заболевания пищеварительной системы, дегенеративные заболевания нервно-мышечной системы (остеопороз, остеоартрит, спондилез). Кроме того, подобные нарушения отмечены у больных с тромбозом сосудов и закупоркой артерий, а также при иммунодефицитных состояниях, включая ревматоидный артрит и другие системные заболевания соединительной ткани. И, наконец, следует отметить, что долихолфосфатный цикл является необходимым метаболическим звеном в процессах регенерации, дифференциации и пролиферации клеток (Scherer M.G., Waechter C.J., 1984; Sakakihara Y., Volpe J.J., 1985; Elmberger P.G. et al., 1987). Все указанное позволило сформулировать цель второй части настоящей работы – изучение действия полипренолов (разных доз ропрена) при токсическом поражении печени и головного мозга, вызванного 39 четыреххлористым углеродом, на функциональное состояние печени, поведение животных и метаболизм моноаминов в мозге крыс в сравнении с известным гепатопротектором адеметионином (гептралом). 40 Глава 2. МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ Методология экспериментального изучения различных компонентов двигательных и эмоциональных форм поведения у грызунов сводится, в первую очередь, к исследованию поведения животных в «открытом поле», агрессивности, игрового поведения, функциональной асимметрии, реакций самораздражения мозга через вживленные электроды в разных вариантах (педальная электростимуляция в камере Скиннера, самораздражение мозга в челночной камере) или условной реакции предпочтения места (условнорефлекторное подкрепление), использованию исследуемого фармакологического вещества в различных дозах для поведенческого (этологического) анализа. Ниже приведено описание исследуемых препаратов и методических приемов, использованных для изучения дофаминзависимых форм поведения крыс. 2.1. Выбор животных и их содержание Опыты выполнены на 306 крысах самцах Вистар массой 200-220 г, выращенных в группе по 5 особей в стандартных пластмассовых клетках в условиях вивария кафедры фармакологии Военно-медицинской академии им. С.М.Кирова. Температура воздуха поддерживалась в пределах 20-22°C, относительная влажность – 50–70%. Температура и влажность воздуха регистрировались ежедневно. На карантине животных содержали в течение 10 дней. В течение этого времени проводили ежедневный осмотр каждого животного (поведение и общее состояние), дважды в день животных наблюдали в клетках (заболеваемость и смертность). Животных содержали при свободном доступе к воде и пище. Все опыты проведены в осенне-зимний период. 41 2.2. Вживление электродов в дофаминергические структуры мозга Стереотаксические операции по вживлению электродов в мозг крысам проводили под нембуталовым наркозом (50 мг/кг) с использованием стереотаксического прибора фирмы «Medicor», Венгрия. Рис. 4. Последовательные этапы проведения операции по вживлению электродов в мозг крысы Билатерально в латеральное гипоталамическое ядро вживляли нихромовые монополярные электроды в стеклянной изоляции (диаметр электрода 0,25 мм, длина оголенного кончика 0,25-0,30 мм, его толщина 0,12 мм) по следующим координатам: АР = 2,5 мм назад от брегмы, SD = 2,0 мм лате- 42 рально от сагитального шва, Н = 8,4 мм от поверхности черепа (König K.P., Klippel A.A., 1963). Индифферентный электрод из нихромовой проволоки закрепляли на черепе животного. Все электроды коммутировались на микроразъеме, который фиксировался на черепе самотвердеющей пластмассой (рис. 4). Поведенческие эксперименты начинали не ранее 10 дней после операции. По окончании всех опытов производили морфологический контроль локализации кончиков электродов на серии фронтальных срезов мозга, которые окрашивали по методу Ниссля, предварительно производили коагуляцию через вживленные электроды током силой 1 мА в течение 30 с (рис. 5). Рис. 5. Фронтальный срез головного мозга крысы Стрелкой указана область вживления электрода в латеральный гипоталамус. 43 2.3. Методы самораздражения мозга у крыс Использовали классический вариант изучения самораздражения мозга в виде педальной самостимуляции в камере Скиннера (рис. 6). Рис. 6. Реакция педальной самостимуляции у крыс в камере Скиннера Два верхних фото демонстрируют нативную методику самостимуляции, нижнее – схематическое ее изображение. Через 10 дней после вживления электродов в мозг крыс обучали нажимать на педаль в камере Скиннера для получения электрического раздражения мозга (прямоугольные импульсы отрицательной полярности, длительностью 1 мс, с частотой 100 Гц, в течение 0,4 с, пороговые значения тока в режиме «фиксированных пачек»). Для повторного раздражения животное было 44 вынуждено вновь нажимать на педаль. Частота и длительность нажатий регистрировались автоматически. Анализировали частоту и время каждого нажатия на педаль. На основании этих результатов вычисляли коэффициент «рассогласования» (Лебедев А.А., Шабанов П.Д., 1992) по формуле: Т1 - Т2 К = ---------, где Т1 + Т2 К – коэффициент «рассогласования»; Т1 – время нажатия педали после окончания стимуляции в случае длительных нажатий, по длительности больших, чем 0,4 сек; Т2 – время от момента отжатия педали до окончания стимуляции. Коэффициент «рассогласования» принимает значения от -1 до +1 и показывает долю активации положительной и отрицательной подкрепляющей фазы самостимуляции (Шабанов П.Д. и др., 2002). Если данный коэффициент принимает положительные значения, то это означает, что крыса продолжала нажимать на педаль даже после того, как раздражение мозга прекратилось. При отрицательных значениях коэффициента «рассогласования» крыса заканчивала нажимать на педаль раньше, чем прекратилась стимуляция мозга. Учитывая, что реакцию самостимуляции можно рассматривать как одновременное включение положительного и отрицательного механизмов подкрепления, сдвиг в сторону увеличения и снижения коэффициента позволяет говорить как об изменении частоты самостимуляции, так и об изменениях подкрепляющих свойств мозга. Поэтому, как дополнительный критерий изменения подкрепляющих свойств стимуляции, коэффициент «рассогласования» является удобным показателем для оценки действия фармакологических препаратов. Исследуемый препарат полипренолов вводили на 3-й день эксперимента и далее в течение 7 дней после стабилизации реакции при использовании фиксированного значения силы тока. Регистрировали число нажатий на педаль и коэффициент «рассогласования» в течение 10 мин эксперимента, за- 45 тем производили внутрибрюшинную инъекцию препарата, и через 30 мин регистрировали те же показатели (число нажатий на педаль и коэффициент «рассогласования») за 15-минутный интервал времени. При изучении подкрепляющих свойств электрической стимуляции мозга использовался свободный режим подкрепления, когда электрическая стимуляция мозга длится все время нажатия педали ( Петров Е.С., 1990). Использование данного режима предполагает повышенный уровень нагрузки на подкрепляющие механизмы головного мозга и дает возможность выявлять отрицательную эмоциональную составляющую реакции самостимуляции, которая обычно следует после 0,5 с от начала раздражения и заставляет животное отжимать педаль (как бы избегать ее) (Вартанян Г.А., Петров Е.С., 1989). 2.4. Исследование поведения крыс в «открытом поле» Свободную двигательную активность животных исследовали в тесте «открытого поля» (Пошивалов В.П., 1977; Михеев В.В., Шабанов П.Д., 2007), представляющего собой круглую площадку диаметром 80 см, ограниченную по окружности непрозрачными бортами высотой 30 см (рис. 7). По всей площади открытого поля равномерно расположены 16 отверстий (норок), диаметром 3 см каждая, предназначенных для выявления видоспецифического компонента исследовательской активности у грызунов (норковый рефлекс). Освещенность открытого поля равнялась 100 лк. Во время опыта экспериментальный вольер находился в специальной звукоизолированной комнате. Продолжительность одного опыта составляла 3 минуты. На основании поведенческого атласа для грызунов (Пошивалов В.П., 1978; Михеев В.В., Шабанов П.Д., 2007) выбирался ряд элементарных двигательных актов и поз, совокупность которых характеризует целостное поведение в «открытом поле». 46 Рис. 7. «Открытое поле» для изучения поведения крыс и основные паттерны их поведения 47 Исходя из требований регистрации и математической обработки, каждому отдельному элементарному акту присваивался определенный номер (код): 0 – «локомоция» (поступательное движение тела в горизонтальной плоскости); 1 – «обнюхивание» (принюхивание и повороты головы без существенных изменений координат корпуса в горизонтальной и вертикальной плоскостях). Этот акт может осуществляться в позах «сидя», «стоя», которые трудно различимы без потери его основного биологического значения, поэтому при регистрации не разделялся в зависимости от позы, в которой он появлялся); 2 – «вертикальная стойка» (стойка на задних лапах в центре открытого поля); 3 – груминг (все разновидности этой реакции); 4 – «неподвижность» (покой, сидение, визуально определяемая неподвижность животного обычно в позе «сидя» с подогнутыми конечностями и сгорбленной спиной); 5 – «движение на месте» (изменение координат головы и корпуса в пределах условной окружности, центром которой являются задние конечности животного, координаты которых существенно не меняются. Достигается переступанием передних конечностей при опоре на задние); 6 – «заглядывание в норку» (норковый рефлекс); 7 – «стойка на стенку» (вертикальная стойка на задних лапах с упором передними на стенку вольера). Регистрация актов производилась нажатием экспериментальной клавиши, соответствующей определенному поведенческому акту, на клавишном устройстве. Полученные данные обрабатывали математически с использованием персонального компьютера. 2.5. Исследование функциональной асимметрии мозга с помощью метода ротации Число ротаций определяли в полусфере диаметром 30 см (рис. 8) через 30 мин после введения фенамина (2,5 мг/кг внутрибрюшинно) за два последовательных периода по 10 мин, используя для анализа средние значения 48 (определяемые за 10 мин). Регистрировали число полных вращений на 360о отдельно вправо и влево. В дальнейшем вычисляли разность между числом предпочитаемых и непредпочитаемых вращений. Если эта разность достигала пяти, считали что животное удовлетворяет критерию ротации. Рис. 8. Исследование ротационного поведения у крыс в ротометре Фиксировали также число неполных ротаций (от 90о до 360о), которые связаны со спонтаной двигательной активностью и отражают снижение чувствительности к фенамину (Carter C.J., Pycock C., 1979). Для оценки чувствительности животного к фенамину в последние 10 сек каждой минуты тестирования в ротометре определяли также показaтель стереотипии по следующей шкале: 0 – сидит спокойно; 1 – обычная (нормальная) двигательная активность; 2 – прерывистая активность, включая частые принюхивания и стойки; 3 – непрерывная стереотипная активность с обнюхиванием и стойками; 4 – отчетливые стереотипные движения головой, двигательная активность падает; 5 – то же, крыса грызет и лижет пол; 6 – преобладают последние формы поведения (Сlark D., White F., 1987). 49 2.6. Исследование поведения крыс в приподнятом крестообразном лабиринте Поведение крыс в приподнятом лабиринте исследовали в установке, представлявшей крестообразный приподнятый лабиринт, который состоял из двух открытых рукавов 50х10 см и двух закрытых рукавов 50х10 см с отрытым верхом, расположенных перпендикулярно относительно друг друга (рис. 9). Высота над полом 1 м. Животное помещали в центр лабиринта. Путем нажатия соответствующей клавиши этографа, связанного с компьютером, фиксировали время пребывания в закрытых и открытых рукавах, время свешивания в отрытых рукавах и выглядывания из закрытых рукавов. Продолжительность теста составляла 5 минут. Рис. 9. Приподнятый крестообразный лабиринт для крыс (общий вид и схема) 50 2.7. Исследование агрессии в тесте «чужак-резидент» Агрессивность изучали у половозрелых крыс самцов в тесте чужакрезидент в соответствии с описанием этологического атласа (Пошивалов В.П., 1986). Рис. 10. Тест «чужак-резирент» у крыс Верхние снимки демонстрируют поведение животных, нижний рисунок – характерную позу угрозы у крыс Смысл методики состоит в том, что к крупному самцу, постоянно находящемуся в клетке (резиденту) подсаживают более мелкое животное (чу- 51 жака). Регистрируют число поведенческих проявлений агрессивности и защиты, а также общее число поведенческих актов, описывающих взаимоотношение двух особей крыс. Изучение внутривидового взаимодействия производили в тесте «чужак-резидент» следующим образом (рис. 10). Подопытное животное – «резидент» в течение часа находилось в клетке размерами 20x36x20 см, после чего к нему подсаживали на 5 мин второе животное – «чужака». «Чужаками» являлись крысы-самцы (30 животных) массой 170-180 г, т.е. заведомо меньших размеров, чем «резиденты», что создавало условия для зоосоциального доминирования последних. В процессе 5-минутного совместного пребывания «резидента» и «чужака», помещаемого в клетку только на время опыта, регистрировали этограмму поведения «резидента» – общее число, последовательность и длительность всех элементарных актов и поз, образующих внутривидовую общительность, агрессию, защиту и индивидуальное поведение. Общительность включала в себя следующие дискретные акты: приближение, следование за партнером, обнюхивание партнера, груминг загривка или тела, наползание или подползание под партнера. Агрессия проявлялась в виде вертикальных или боковых стоек (угроза) или атаки. Социальная пассивность выражалась различными актами индивидуального поведения: локомоцией, обнюхиванием, аутогрумингом, движениями на месте, вертикальными стойками, неподвижностью. 2.8. Исследование антидепрессантной активности в тесте Порсолта Плавательный тест «отчаяния» Порсолта (Porsolt R.D., 1978) предусматривает оценку двигательной активности крыс, помещенных в стеклянный цилиндр диаметром 20 см и высотой 40 см, на 1/3 заполненный водой с температурой 27±1оС (рис. 11). 52 Рис. 11. Плавательный тест Порсолта Животное помещают в цилиндр на 6 мин, регистрируют время активного и пассивного плавания и время иммобилизации. Увеличение активного плавания и уменьшение времени иммобилизации рассматривают как антидепрессантный эффект (Андреева Н.И., 2002). 2.9. Моделирование экспериментального подострого гепатоза Экспериментальный подострый гепатоз (токсическое поражение печени) моделировали внутрибрюшинным введением крысам 50%-ного раствора ССl4. Для приготовления матричного раствора брали 50 мл 100%-ного раствора ССl4 добавляли к нему 50 мл стерильного оливкового масла, перемешивали и в дальнейшем вводили внутрибрюшинно по 0,4 мл/крысу 50%-ного раствора ССl4 (это соответствовало 1000 мг/кг ССl4) на 1-й и 4-й дни опыта. В связи с нарастающей гибелью экспериментальных животных (более 50%) дальнейшее введение ССl4 не производили. В целом, каждая крыса получила 2000 мг/кг ССl4 за опыт. 53 В соответствии с протоколом исследования все животные были разделены на 7 групп, каждая из которых включала 10-14 крыс: Группа 1 – получала внутрибрюшинно (в/бр) физиологический 0,9%ный раствор хлорида натрия; Группа 2 – получала в/бр раствор ССl4 + физиологический раствор 14 дней; Группа 3 – получала в/бр ССl4 + полипренолы (2,2 мг/кг) в/бр ежедневно 14 дней; Группа 4 – получала в/бр раствор ССl4 + полипренолы (4,3 мг/кг) в/бр ежедневно 14 дней; Группа 5 – получала в/бр раствор ССl4 + полипренолы (11,6 мг/кг) в/бр ежедневно 14 дней; Группа 6 – получала в/бр раствор ССl4 + адеметионин раствор 300 мг/кг в/бр на основе лиофилизата ежедневно 14 дней; Группа 7 – получала в/бр раствор ССl4 + адеметионин раствор 300 мг/кг в/бр на основе таблеток ежедневно 14 дней. Тестирование поведения начинали на 14-й день через 0,5-1 ч после последнего введения исследуемых веществ. Поведенческие тесты проводили в течение 2 последовательных дней. Забой животных осуществляли на 16-й день опыта. После декапитации крыс производили взятие крови, печени и мозга (стриатум, прилежащее ядро и префронтальная кора). В случае выпадения животных из опыта (гибель вследствие токсического действия ССl4) группы дополняли новыми животными, опыт в этом случае смещался во времени, но его схема сохранялась. 2.10. Биохимическое исследование крови Для биохимического анализа кровь всех животных забирали в сухие стерильные пробирки на 16-й день опыта. Материал центрифугировали при 54 3000 об./мин. Сыворотку отбирали для дальнейшего биохимического анализа на общий белок, глюкозу, холестерол, креатинин, мочевину, α-амилазу, лактатдегидрогеназу, триглицериды, общий билирубин, аспартат- аминотрансферазу (АСТ), аланин-аминотрансферазу (АЛТ), связанный билирубин, щелочную фосфатазу, мочевую кислоту, креатин-фосфокиназу (КФК), фосфор, липопротеиды низкой плотности и γ-глютамилтрансферазу. Материал исследовали на биохимическом анализаторе Sapphire 400 (производства фирмы Sapphire, Япония) с использованием реактивов системы Randox (RANDOX Laboratories Ltd., United Kingdom). 2.11. Гистологическое исследование печени После окончания поведенческих исследований на 16-й день опыта животных забивали путем декапитации, извлекали печень и ткань печени крыс фиксировали в 10%-ном растворе нейтрального формалина в течение 24 ч, после чего материал проходил стандартную обработку в спиртах нарастающей концентрации (70-95%), ксилоле и парафине для изготовления гистологических препаратов с толщиной серийных парафиновых срезов 3-5 мкм. Для микроскопического исследования срезы окрашивались гематоксилином и эозином. Морфологическое исследование гистологических препаратов проводили при помощи светооптического микроскопа Leica DM LS (Япония) в пяти полях зрения при увеличении 200. Микрофотографирование проводили при помощи цифровой фотокамеры Leica DC320 (Япония). Масштаб патологических изменений в дольке, учитывая ее строение в виде шестигранника, оценивался как: поражение 1/3, 1/2, 2/3 и 3/4 дольки. Патогистологическая оценка изменений в печени была проведена по трехбалльной системе: 55 1 балл – слабо выраженные изменения в виде белковой гиалиново- капельной и мелкокапельной жировой дистрофии в цитоплазме гепатоцитов, расположенных в периферических отделах долек (перипортально) с локализацией ее в отдельных дольках в незначительном количестве гепатоцитов. 2 балла – умеренно выраженные изменения. Наблюдалась дискомплексация (нарушение) балочного строения в периферических отделах долек в отдельных полях зрения с поражением 1/3-1/2 протяженности балок в дольке, появлялась мелкокапельная жировая дистрофия гепатоцитов перипортально. В центральных отделах долек гепатоциты не имели дистрофических изменений, слабо выраженная лимфоцитарная инфильтрация портальных трактов. 3 балла – выраженные изменения, которые характеризовались поражением 2/3-3/4 гепатоцитов долек и локализовались перипортально. Развивались гиалиново-капельная белковая дистрофия и мелкокапельное ожирение гепатоцитов, дискомплексация балочного строения, слабо выраженная лимфоцитарная инфильтрация портальных трактов, центральные отделы долек были сохранны. 2.12. Биохимическое исследование мозга После декапитации структуры мозга извлекали на льду и замораживали в жидком азоте. Пробы гомогенизировали в 10 объемах 0,1 М HCIО4 и центрифугировали при 15 000 g в течение 10 мин. Содержание моноаминов и их метаболитов определяли методом высокоэффективной жидкостной хроматографии (ВЭЖХ) с электрохимической детекцией (Ротенко А.А. и др., 2006). Надосадочную жидкость в количестве 20 мкл наносили на аналитическую колонку Zorbax C18. Стандартные исследуемые растворы медиаторов и их метаболитов (Sigma, США) приготовляли на основе бидистиллированной воды. Норадреналин (НА), дофамин (ДА), серотонин (СЕР), а также 3,4диоксифенилуксусную (ДОФУК), гомованилиновую (ГВК), 5- 56 гидроксииндолуксусную кислоты (5-ГИУК) разделяли с использованием в качестве подвижной фазы 0,1 молярного цитратно-фосфатного буфера, содержащего 0,3 ммоля октансульфоната натрия, 0,1 ммоля ЭДТА и 10% ацетонитрила (рН 3,2). Определение моноаминов и их метаболитов осуществляли на стеклоуглеродном электроде при потенциале +0,7 мВ против Ag/AgCl электрода сравнения. Скорость потока подвижной фазы составляла 0,8 мл/мин. Содержание моноаминов и их метаболитов в гомогенатах различных отделов мозга определяли методом ВЭЖХ относительно содержания стандартных растворов, приготовленных выше указанным способом, как описано в работе (Ротенко А.А. и др., 2006). 2.13. Фармакологические вещества, используемые для анализа двигательных и эмоциональных форм поведения В 1-ой серии исследований (поведение) для нейрофармакологического анализа были использованы следующие фармакологические агенты (табл. 1): Ропрен в виде 25%-ного масляного раствора в дозах 0,5 мл/кг (что соответствовало 4,3 мг/кг), 1 мл/кг (8,6 мг/кг) и 1,5 мл/кг (13,4 мг/кг), а также непрямой адреномиметик фенамин (1-2,5 мг/кг) для инициации стереотипного поведения. Выбор доз ропрена основывался на рекомендациях разработчика и ранее полученных результатах исследований на грызунах, а фенамина – на предпочтительном использовании указанных доз в поведенческих экспериментах (Андреева Н.И., 2000). Все вещества вводили внутрибрюшинно в виде курса в течение 5-7 дней, каждая инъекция за 40-60 мин до начала опыта. В качестве контроля использовали введение 0,9%-ного раствора хлорида натрия. Во 2-ой серии исследований (подострый гепатоз) для фармакологического анализа были использованы следующие фармакологические агенты: ропрен масляный раствор, приготовленный на основе 25%-ного масляного 57 раствора, в дозах 0,5 мл (что соответствовало 2,2 мг/кг), 0,75 мл (4,3 мг/кг) и 1,2 мл (11,6 мг/кг), а также синтетический гепатопротектор гептрал (Sаденозил-L-метионин) в дозе 300 мг/кг. Выбор доз основывался на предпочтительном использовании указанных доз в поведенческих экспериментах (Андреева Н.И., 2002; Шабанов П.Д., 2008). Все вещества вводили внутрибрюшинно в виде курса в течение 14 дней. Тестирование поведения начинали на 14-й день через 0,5-1 ч после последнего введения исследуемых веществ. Поведенческие тесты проводили в течение 2 последовательных дней. Забой животных осуществляли на 16-й день опыта. После декапитации крыс производили взятие крови, печени и мозга (стриатум, прилежащее ядро и префронтальная кора). Приготовление матричного раствора ропрена состояло в следующем. Брали 4 мл 25%-ного масляного раствора ропрена, добавляли 496 мл стерильного оливкового масла, перемешивали и в дальнейшем вводили по 0,50,75-1,2 мл (группы 3-4-5) внутрибрюшинно ежедневно в течение 14 дней в одно и то же время (10.00-11.00). Ропрен вводили за 1 ч до ССl4, если его инъекция совпадала с введением токсиканта. Раствор гептрала (S-аденозил-L-метионина) готовили следующим образом. Брали стандартный раствор гептрала (5 мл растворителя добавляли в пузырек с лиофилизатом) и вводили по 0,75 мл/крысу внутрибрюшинно в течение 14 дней ежедневно (группа 6). Либо 20 таблеток гептрала растворяли в 100 мл 0,9%-ного физиологического раствора, вводили по 0,75 мл/крысу внутрибрюшинно в течение 14 дней ежедневно (группа 7). 2.14. Статистическая обработка полученных материалов Выборка для каждой группы животных составила не менее 10-12 крыс. Полученные результаты обрабатывали статистически с использованием t- 58 критерия Стьюдента, непараметрического критерия U Вилкоксона-МаннаУитни и однофакторного дисперсионного анализа по методу ANOVA с последующим Newman-Keuls post-hoc анализом. Данные обрабатывали на персональном компьютере с использованием стандартного набора программ Statistika for Windows 6.0. 59 Глава 3. РЕЗУЛЬТАТЫ СОБСТВЕННЫХ ИССЛЕДОВАНИЙ 3.1. Исследование поведенческих эффектов полипренолов Первый раздел исследований предусматривал изучение влияния полипренолов в 3 дозах (4,3 мг/кг, 8,6 мг/кг и 13,4 мг/кг) при его курсовом введении (5-7 дней, 1 инъекция в сутки) на разные виды поведения крыс: поведение в открытом поле, в приподнятом крестообразном лабиринте, в тесте «чужак-резидент», в ротометре (стереотипное поведение), в тесте Порсолта, в тесте самостимуляции латерального гипоталамуса. 3.1.1. Влияние ропрена на двигательную активность и поведение крыс в «открытом поле» Исследование влияния курсового введения полипренолов (5 дней) на поведение крыс в «открытом поле» показало, что независимо от дозы (4,38,6-13,4 мг/кг) полипренолы умеренно активировали горизонтальную, не влияя на вертикальную двигательную активность (табл. 2). Полипренолы подавляли (в 3-4 раза) норковый рефлекс (исследовательская активность), умеренно снижали груминг (только в дозе 13,4 мг/кг) и общую эмоциональность животных (в дозах 4,3 и 13,4 мг/кг, но не 8,6 мг/кг), оцененную по болюсам дефекаций. В дозе 8,6 мг/кг полипренолы повышали число болюсов дефекации, что указывает на умеренное повышение эмоциональности при использовании данной дозы препарата, Таким образом, в тесте «открытого поля» полипренолы не проявляют дозозависимого эффекта. В целом, полипренолы растормаживают (активируют) горизонтальную двигательную активность при умеренном снижении всех других поведенческих паттернов (прежде всего, исследовательской активности и эмоциональности). 60 Таблица 2 Влияние полипренолов (4,3-8,6-13,4 мг/кг) на поведение крыс в «открытом поле» (время поведенческих паттернов, сек) Препараты 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы 8,6 мг/кг Полипренолы 13,4 мг/кг Поведенческие акты ВертиИсследоваГруминг кальные ние норок стойки Болюсы дефекаций 17,0±2,2 6,7±0,9 4,7±0,6 3,8±0,5 1,0±0,1 24,0±3,1* 8,0±1,0 1,0±0,1** 3,0±0,4 0,3±0,1** 25,0±3,3* 5,3±0,7 1,5±0,2** 4,3±0,6 1,5±0,2* 25,7±3,3* 5,5±0,7 1,5±0,2** 2,6±0,3* 0,5±0,1* Пересеченные квадраты Примечание. *р<0,05, **р<0,01 в сравнении с соответствующим контролем. 3.1.2. Влияние полипренолов на двигательную активность и стереотипное (ротационное) поведение крыс Одним из наиболее типичных дофаминзависимых форм поведения является ротационное и стереотипное поведение. Ротационное поведение, как правило, инициируется введением фенамина, непрямого дофаминомиметика. Поскольку в предварительных исследованиях в тесте «открытого поля» нами не были выявлены дозозависимые эффекты полипренолов, в данном тесте использовали наиболее высокую дозу препарата – 13, 4 мг/кг. Данные этой серии исследований представлены в табл. 3. Из таблицы видно, что фенамин (2,5 мг/кг) в среднем в 4 раза повышал число вращений как влево, так и вправо (р<0,01). Полипренолы (13,4 мг/кг) при курсовом введении в течение 5 дней умеренно подавляли вращение вле- 61 во и не влияли на показатели вращения вправо. Во всех случаях полипренолы не вызывали стереотипного поведения, характерного для активации дофаминергической системы мозга. Таблица 3 Влияние полипренолов (13,4 мг/кг) на ротационное поведение крыс Препараты 0,9% раствор NaCl (контроль) Фенамин 2,5 мг/кг Полипренолы 13,4 мг/кг Сумма полных оборотов Сумма полных оборотов влево Вправо 2,2±0,3 1,7±0,2 8,3±1,1** 1,5±0,2* 6,8±0,9** 1,8±0,2 Примечание. *р<0,05, **р<0,01 в сравнении с соответствующим контролем. Таким образом, в отличие от фенамина полипренолы (13,4 мг/кг) умеренно подавляют ротационное поведение крыс в ротометре и не вызывают стереотипий. Это указывает на возможное участие дофаминергической системы мозга (ее умеренное подавление) в эффектах полипренолов. 3.1.3. Влияние полипренолов на поведение крыс в приподнятом крестообразном лабиринте Приподнятый крестообразный лабиринт позволяет оценить анксиолитические (противотревожные) или анксиогенные (тревожные) свойства фармакологических препаратов, оцениваемые по времени пребывания в затемненных и освещенных рукавах лабиринта. Увеличение времени пребывания в освещенных рукавах лабиринта и повышение числа выглядываний и свешиваний с открытых площадок лабиринта квалифицируется как анксиолитический (противотревожный) эффект, а уменьшение этих показателей – как анксиогенный (тревожный) эффект. 62 Таблица 4 Влияние полипренолов (4,3-8,6-13,4 мг/кг) на поведение крыс в приподнятом крестообразном лабиринте Препараты 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы 8,6 мг/кг Полипренолы 13,4 мг/кг Время в открытых рукавах (мин) Поведенческие акты Время в закрытых рукавах (мин) Число выглядываний Число свешиваний 0,4±0,1 4,6±0,2 10,2±1,3 6,2±0,8 0,4±0,1 4,6±0,2 5,7±0,7* 3,7±0,5* 0,5±0,1 4,5±0,2 3,2±0,4** 10,0±1,4* 0,2±0,1* 4,8±0,3 2,1±0,3** 3,3±0,4* Примечание. *р<0,05, **р<0,01 в сравнении с соответствующим контролем. Исследование влияния полипренолов (4,3-8,6-13,4 мг/кг) при курсовом введении в течение 5 дней на поведение крыс в приподнятом крестообразном лабиринте показало (табл. 4), что только в дозе 13,4 мг/кг полипренолы вдвое уменьшали время пребывания в освещенных рукавах лабиринта. Вместе с тем, зависимо от дозы полипренолы в 2-5 раз снижали число выглядываний из темных рукавов и в 2 раза снижалит число свешиваний с открытых платформ лабиринта (в дозах 4,3 и 13,4 мг/кг, но не 8,6 мг/кг). В дозе 8,6 мг/кг полипренолы повышали число выглядываний из затемненных рукавов лабиринта, однако данный показатель не является основным, а рассматривается как вспомогательный при оценке анксиолитического или анксиогенного действия фармакологических веществ. Таким образом, полипренолы проявляют умеренную анксиогенную активность, которая наиболее стабильно воспроизводится при введении препарата в дозе 13,4 мг/кг. 63 3.1.4. Влияние полипренолов на показатели агрессии в тесте «чужак-резидент» у крыс Агрессивное поведение контролируется преимущественно холинергической и дофаминергической системами мозга. Влияние полипренолов при курсовом введении в течении 5 дней на показатели системы агрессия-защита в тесте «чужак-резидент» показало, что препарат вне зависимости от дозы существенно (в 3,5-7 раз) подавлял проявления общительности и паттерны индивидуального поведения. При этом в дозе 13,4 мг/кг полипренолы растормаживали систему агрессии и защиты (табл. 5). Таблица 5 Влияние полипренолов (4,3-8,6-13,4 мг/кг) на показатели агрессии и защиты у крыс Препараты 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы 8,6 мг/кг Полипренолы 13,4 мг/кг Поведенческие акты, усл. ед. ИндивидуОбщительальное поАгрессия ность ведение Защита 65,8±8,6 77,3±10,1 0,0±0,0 0,0±0,0 9,8±1,3*** 14,3±1,8*** 0,0±0,0 0,0±0,0 11,1±1,4*** 17,8±2,3*** 0,0±0,0 0,0±0,0 13,8±1,8*** 21,3±2,8*** 0,8±0,1** 0,2±0,1* Примечание. *р<0,05, **р<0,01, ***р<0,001 в сравнении с соответствующим контролем. В последнем случае у контрольных животных практически не отмечено проявление агрессии, хотя после введения полипренолов в наибольшей изучаемой дозе показатели агрессии умеренно повышались (p<0,01). Параллельно этому активировалась и система защиты. Это закономерно, поскольку 64 система защиты, как правило, активируется при повышении показателей агрессивности. В то же время существуют лекарственные средства, например, прямой дофаминомиметик апоморфин, которые избирательно повышают только систему агрессии, не влияя на систему защиты. Таким образом, полипренолы в тесте «чужак-резидент» не оказывают дозозависимого эффекта, значимо подавляя индивидуальное поведение и общительность (коммуникативность) животных. Видимого антиагрессивного действия препарат не оказывает. В дозах 4,3 и 8,6 мг/кг полипренолы не оказывают агрессивного/антиагрессивного действия, а в большой дозе (13,4 мг/кг) проявляют незначительное агрессивное действие с одновременной активацией системы защиты. 3.1.5. Влияние полипренолов на показатели депрессии в тесте Порсолта у крыс Тест Порсолта используется для оценки уровня депрессии у крыс. Он считается достаточно информативным и специфичным для оценки антидепрессантного действия. В условиях наших экспериментов полипренолы при курсовом введении в течение 5 дней существенно (в 4-5 раз) увеличивал время иммобилизации животных, мало меняя показатели активного и пассивного плавания (табл. 6). Время иммобилизации является основным показателем, по которому оценивают антидепрессантное действие препарата. Увеличение этого показателя говорит о наличии депрессантных (подавляющих ЦНС) свойств, а уменьшение, напротив, – о проявлении антидепрессантных свойств. Только в дозе 13,4 мг/кг полипренолы умеренно уменьшали время активного плавания, в дозе 4,3 мг/кг – пассивного плавания, а в дозе 8,6 мг/кг – не меняли времени активного и пассивного плавания крыс. 65 Таблица 6 Влияние полипренолов (4,3-8,6-14,3 мг/кг) на поведение в тесте Порсолта у крыс Препараты Активное плавание Показатель, с Пассивное Иммобилизация плавание 0,9% раствор NaCl 18,5±2,4 369,5±18,0 2,8±0,4 (контроль) Полипренолы 15,3±2,0 262,7±47,2* 12,1±1,6** 4,3 мг/кг Полипренолы 18,8±2,4 353,3±45,9 15,8±2,1** 8,6 мг/кг Полипренолы 14,3±1,9* 346,2±45,0 17,1±2,2** 13,4 мг/кг Примечание. *р<0,05, **р<0,01 в сравнении с соответствующим контролем. Следовательно, полипренолы оказывают депрессантный эффект на поведение в тесте Порсолта. Степень выраженности депрессантного эффекта полипренолов может рассматриваться как умеренная. 3.1.6. Влияние полипренолов на систему мозгового подкрепления в тесте самостимуляции латерального гипоталамуса у крыс Для изучения систем мозгового подкрепления обычно используют реакцию самостимуляции (показатель первично подкрепляющих свойств, или безусловного подкрепления) или условную реакцию предпочтения места (показатель вторично подкрепляющих свойств, или условного подкрепления). Реакция самораздражения латерального гипоталамуса представляет собой жестко детерминированную реакцию, легко воспроизводимую и трудно поддающуюся изменению. Лишь одна группа препаратов – психостимуляторы (фенилалкиламины, сиднонимины, в меньшей степени кофеин) – стабильно активируют реакцию самостимуляции. Все другие фармакологиче- 66 ские средства либо не влияют на нее, либо угнетают (например, нейролептики, транквилизаторы и т.д.). Самостимуляцию гипоталамуса регистрировали последовательно после каждого из 7 дней введения порлипренолов в дозах 4,3-8,6-13,4 мг/кг (табл. 7). В первые 2 дня исследования (введения полипренолов) препарат либо не влиял (13,4 мг/кг), либо умеренно (на 18-28%) угнетал реакцию самостимуляции в дозах 4,3-8,6 мг/кг. Начиная с 3-го дня введения полипренолы (8,6 и 13,4 мг/кг) на 27-39% увеличивали реакцию самостимуляции, оказывая типичный психоактивирующий эффект. Этот положительный эффект проявлялся и на 4-5 дни введения полипренолов в дозах 8,6 и 13,4 мг/кг с максимумом (+39%) на 3-й день. В то же время на 3-й день введения полипренолы в дозе 4,3 мг/кг выражено подавляли реакцию самостимуляции (-34% против -11% в контроле); подавление данной реакции прослеживалось в течение всех 5-ти дней эксперимента, когда вводили полипренолы в данной дозе (то есть, на 3-ий, 4-ый и 5-ый дни). Следует отметить, что психоактивирующий эффект полипренолов, проявляющийся после их введения в дозах 8,6 и 14,3 мг/кг, последовательно уменьшался (на 4-й день на 15%, на 5-й день – на 12-16%). На 6-й и 7-й дни анализировали действие полипренолов на реакцию самостимуляции в камере Скиннера на фоне введения психостимулятора фенамина. Сам фенамин (1 мг/кг) дает четкую реакцию активации самостимуляции на 35-37%. На фоне введения полипренолов и независимо от их дозы фенамин сохранил свой психоактивирующий эффект, повышая число нажатий на педаль на 27-45%, то есть в том интервале, который характерен (типичен) для самого фенамина. Таким образом, действие полипренолов на реакцию самостимуляции зависит от продолжительности его введения. В первые дни введения полипренолы, как правило, умеренно подавляет реакцию самостимуляции, затем эффект сменяется на умеренный психоактивирующий, который по мере вве- 67 Таблица 7 Влияние полипренолов (4,3-8,6-13,4 мг/кг) и фенамина (1 мг/кг) на показатели самостимуляции латерального гипоталамуса у крыс Препараты 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы 8,6 мг/кг Полипренолы 13,4 мг/кг 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы 8,6 мг/кг Полипренолы 13,4 мг/кг 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы 8,6 мг/кг Полипренолы 13,4 мг/кг 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы Число нажатий на педаль за 10 мин До ввеПосле введения дения 1-й день 161±12 147±16 (+10%) 122±15 148±21 (-18%) 113±20* 158±12 (-28%) 160±18 165±21 (-3%) 2-й день 152±12 143±23 (-6%) 136±19* 188±23 (-28%) 172±21 169±20 (+2%) 157±13 171±22 (-8%) 3-й день 161±11 144±24 (-11%) 155±17 170±23 (-34%) 232±22* 182±19 (+27%) 244±22* 176±19 (+39%) 4-й день 171±11 154±24 (-11%) 121±26* 182±17 (-34%) 191±23 246±18* Коэффициент «рассогласования» До введеПосле ния введения 0,21±0,02 0,20±0,01 0,17±0,01 0,23±0,03 0,21±0,04 0,19±0,01 0,14±0.4 0,18±0,02 0,34±0,03 0,26±0,05 0,19±0,06 0,16±0,02 0,14±0,3 0,16±0,02 0,21±0,12 0,17±0,02 0,14±0,03 0,18±0,06 0,21±0,02 0,20±0,01 0,25±0,04 0,14±0,03* 0,20±0,09 0,18±0,5 0,16±0,03 0,18±0,06 0,15±0,02 0,21±0,03* 0,19±0,05 0,21±0,03 68 8,6 мг/кг Полипренолы 13,4 мг/кг 209±23 0,9% раствор NaCl (контроль) Полипренолы 4,3 мг/кг Полипренолы 8,6 мг/кг Полипренолы 13,4 мг/кг 142±22 0,9% раствор NaCl (контроль) Фенамин 1 мг/кг 166±17 Полипренолы 4,3 мг/кг + фенамин 1 мг/кг Полипренолы 8,6 мг/кг + фенамин 1 мг/кг Полипренолы 13,4 мг/кг + фенамин 1 мг/кг 0,9% раствор NaCl (контроль) Фенамин 1 мг/кг Полипренолы 4,3 мг/кг + фенамин 1 мг/кг Полипренолы 8,6 мг/кг + фенамин 1 мг/кг Полипренолы 13,4 мг/кг + фенамин 1 мг/кг 205±31 211±13 228±21 185±19 (+15%) 241±19 (+15%) 5-й день 156±17 (+10%) 170±24 (-17%) 244±24 (+16%) 255±15 (+12%) 6-й день 155±22 (-7%) 253±21* (+37%) 0,13±0,01 0,19±0,04* 0,13±0,04 0,18±0,03 0,22±0,05 0,19±0,04 0,16±0,11 0,22±0,01* 0,20±0,04 0,22±0,01 0,15±0,01 0,13±0,04 0,19±0,02 0,13±0,04* 217±25 275±19* (+27%) 0,25±0,03 0,14±0,04* 220±19 280±35* (+27%) 0,21±0,11 0,19±0,10 201±21 292±17* (+45%) 0,24±0,06 0,12±0,02* 0,16±0,01 0,14±0,04 0,18±0,02 0,12±0,04* 7-й день 176±17 165±22 (-6%) 180±21 243±19* (+35%) 212±30 288±34* (+36%) 0,24±0,02 0,22±0,04 211±22 294±26* (+39%) 0,12±0,04 0,23±0,06 221±15 303±19* (+37%) 0,26±0,05 0,23±0,06 Примечание. *р<0,05 в сравнении с данными до введения препаратов. 69 дения препарата снижается. Кроме того, полипренолы не проявляют функционального антагонизма с психостимулятором фенамином, на фоне их введения фенамин в полной мере проявляет свой психоактивирующий эффект, активируя самостимуляцию на тот порядок, который характерен для введения самого фенамина. 3.1.7. Заключение Изучали влияние полипренолов на дофамин-зависимые формы поведения крыс самцов Вистар (поведение в открытом поле, в приподнятом крестообразном лабиринте, в тесте «чужак-резидент», в ротометре, в тесте Порсолта, в тесте самостимуляции латерального гипоталамуса). Полипренолы использовали в дозах 4,3-8,6-13,4 мг/кг при их курсовом введении (5-7 дней). В «открытом поле» полипренолы не проявляли дозозависимого эффекта, растормаживая (активируя) горизонтальную двигательную активность при умеренном снижении всех других поведенческих паттернов (прежде всего, исследовательской активности и эмоциональности). В ротационном тесте полипренолы только в большой дозе (13,4 мг/кг) в отличие от фенамина (непрямого дофаминомиметика) умеренно уменьшали число вращений в ротометре и не вызывали стереотипий. Полипренолы проявляли умеренную анксиогенную (тревожную) активность, которая наиболее стабильно воспроизводится при введении препарата в дозе 13,4 мг/кг. В тесте «чужак-резидент» полипренолы значимо подавляли индивидуальное поведение и общительность (коммуникативность) животных. Видимого антиагрессивного действия препарат не выявил. В тесте Порсолта полипренолы (4,3-8,6-13,4 мг/кг) оказывали депрессантный эффект на поведение. В дозе 13,4 мг/кг полипренолы проявляли максимальное действие. Степень выраженности депрессантного эффекта полипренолов может рассматриваться как умеренная. Действие полипренолов на реакцию самостимуляции в камере Скиннера зависело от про- 70 должительности их введения. В первые дни введения полипренолы, как правило, умеренно подавляли реакцию самостимуляции, затем эффект сменяется на умеренный психоактивирующий, который по мере введения препарата снижался. Кроме того, полипренолы не проявляли функционального антагонизма с психостимулятором фенамином, и на фоне их введения фенамин в полной мере развертывал свой психоактивирующий эффект. Сделано предположение, что полипренолы может проявлять свойства парциального агониста (агониста-антагониста) рецепторов дофамина. 71 3.2. Исследование поведенческих и нейрохимических эффектов полипренолов при подостром гепатозе Исследование предусматривало изучение влияния полипренолов в 3 дозах (2,2 мг/кг, 4,3 мг/кг и 11,6 мг/кг) при их курсовом введении (14 дней, 1 инъекция в сутки) на разные виды поведения крыс с подострым гепатозом: поведение в открытом поле, в приподнятом крестообразном лабиринте, в тесте «чужак-резидент», в тесте Порсолта, в тесте самостимуляции латерального гипоталамуса. 3.2.1. Характеристика экспериментальной модели Двукратное внутрибрюшинное введение 0,4 мл/крысу 50%-ного раствора ССl4 (это соответствовало 1000 мг/кг ССl4) на 1-й и 4-й дни опыта приводило к развитию явного токсического действия гепатотропного яда. Летальность крыс, не защищенных гепатопротекторами, после первого введения ССl4 составила около 40%, после второго введения – увеличилась до 50% (ЛД50). В связи с нарастающей гибелью экспериментальных животных дальнейшее введение ССl4 не производили. В целом, каждая крыса получила 2000 мг/кг ССl4 за опыт. В крови крыс, забитых на 16-й день опыта, регистрировали признаки поражения внутренних органов: печени, миокарда, поджелудочной железы, почек. В частности, у крыс, получавший ССl4, в 2 раза возрастала активность аспартатаминотрансферазы (с 123,1±14,4 до 248,6±10,4 МЕ/л), в 4 раза – лактатдегидрогеназы (с 22,8±9,3 до 88,7±49,7 МЕ/л), умеренно повышалась активность α-амилазы (с 1708,8±40,9 до 1930,8±35,4 МЕ/л), снижалась активность γ-глутамилтрансферазы (с 2,33±0,83 до 0,98±0,97 МЕ/л), общего и непрямого билирубина, мочевой кислоты, липопротеидов низкой плотности. 72 При гистологическом исследовании печени у крыс был выявлен экспериментальный гепатоз (дистрофия печени), который характеризовался развитием следующих гистологических проявлений: • гиалиново-капельной белковой дистрофией; • мелкокапельной жировой дистрофией; • слабо выраженной лимфоцитарной инфильтрацией портальных трактов. Гистологическая картина в группах лабораторных животных распределилась следующим образом. В группе 1 (крысы, получавшие физиологический раствор) печень имела нормальное гистологическое строение (рис. 12). В печеночных дольках радиальное расположение балок из гепатоцитов в виде радиально отходящих от центральной вены к портальным трактам тяжей, гепатоциты с розовой цитоплазмой и синими ядрами, местами цитоплазма имела различную оптическую плотность, связанную со степенью накопления гликогена. Дистрофические и некротические изменения отсутствовали, наблюдалась незначительная лимфоидная инфильтрация портальных трактов, в которых имелись триады – артерия, вена и желчный проток, окруженные незначительным количеством соединительнотканной стромы. Синусоиды в дольках не расширены, наблюдалось полнокровие центральных вен. В группе 2 (крысы, получавшие CCl4 + физиологический раствор) на фоне нарушения балочного строения на большем протяжении в печени имелась паренхиматозная дегенерация в виде белковой гиалиново-капельной дистрофии, которая поражала гепатоциты всех печеночных долек на 1/3 – 2/3 протяженности балок, местами захватывала дольки очагово в виде мелкокапельной жировой дистрофии, локализовалась перипортально (рис. 13). В трактах наблюдалась слабо выраженная мононуклеарная инфильтрация, некроз гепатоцитов отсутствовал. В соответствие с балльной оценкой степени патогистологических изменений можно было констатировать 2-3 бальное поражение печени крыс данной группы – гистологические изменения соответ- 73 ствовали картине умеренной и выраженной токсической паренхиматозной дистрофии печени (гепатозу). Рис. 12. Печень крысы, получавшей физиологический раствор (слева) Нормальное гистологическое строение ткани. Окраска гематоксилином и эозином. Увеличение 200. Рис. 13. Печень крысы, получавшей CCl4 + физиологический раствор (справа) Дискомплексация балочного строения, умеренно выраженные и выраженные проявления дистрофии гепатоцитов (↑). Окраска гематоксилином и эозином. Увеличение 200. Таким образом, двукратное введение CCl4 в дозе 2000 мг/кг приводило к развитию подострого гепатоза (дистрофии печени), регистрирующегося при исследовании даже через 15 дней после первого введения гепатотропного яда. На данной модели можно адекватно оценивать действие гепатопротекторных средств. 74 3.2.2. Влияние полипренолов и адеметионина на выживаемость крыс, подвергнутых воздействию CCl4 Двукратное внутрибрюшинное введение CCl4 в суммарной дозе 2000 мг/кг вызывало в группе, получавшей физиологический раствор, гибель 50% крыс (табл. 8). Таблица 8 Влияние полипренолов и адеметионина на выживаемость крыс, подвергнутых воздействию CCl4 Группа крыс Число крыс в Число погиб- группе ших крыс (%) 1. Контроль (физиологический раствор) 2. CCl4 + физ. раствор 10 0 (0%) 14 7 (50%) 3. CCl4 + полипренолы 2,2 мг/кг 12 5 (42%) 4. CCl4 + полипренолы 4,3 мг/кг 14 6 (43%) 5. CCl4 + полипренолы 11,6 мг/кг 14 7 (50%) 6. CCl4 + адеметионин 300 мг/кг (на основе лиофилизата) 7. CCl4 + адеметионин 300 мг/кг (на основе таблеток) 14 6 (43%) 10 10 (100%) Полипренолы в дозах 2,2-4,3-11,6 мг/кг достоверно не снижали гибель крыс, подвергнутых воздействию CCl4. Процент погибших животных в этих группах составлял 42-50%. Гибель крыс приходилась на второй день после инъекции CCl4, то есть на 2-й и 5-й день опыта. Адеметионин 300 мг/кг, приготовленный на основе лиофилизата для инъекций, также не снижал процента гибели животных. Адеметионин 300 мг/кг, приготовленный на основе таблеток и вводимый внутрибрюшинно, приводил к гибели всех крыс (100%). Последняя крыса в этой группе погибла на 7-й день опыта. Поэтому в дальнейшем эта группа в анализ результатов не вошла. 75 3.2.3. Влияние полипренолов и адеметионина на поведение крыс, подвергнутых воздействию CCl4 3.2.3.1. Влияние полипренолов и адеметионина на поведение крыс в тесте «открытое поле», характеризующем двигательную и исследовательскую активность и эмоциональность животных Двукратное введение CCl4 (2000 мг/кг) с оценкой поведения через 15 дней после первого введения токсиканта приводило к существенному нарушению поведения крыс в «открытом поле» (рис. 14). Так, в группе крыс, получавшей CCl4, регистрировали снижение горизонтальной (в 3 раза) и исследовательской (в 2,5 раза) активности, практически подавление груминговых реакций (снижение более чем в 5 раз) и повышение эмоциональности (увеличение болюсов дефекации в 2 раза). Вертикальная активность при этом существенно не менялась. Полипренолы при курсовом введении (14 дней) в целом нормализовали поведение крыс, при этом максимальные их эффекты отмечены при введении препарата в дозах 2,2 и 4,3 мг/кг, но не 11,6 мг/кг. Именно в этих дозах полипренолы восстанавливали горизонтальную двигательную (пересечение квадратов) и исследовательскую (норковый рефлекс) активность, нормализовали эмоциональность животных. На груминговые реакции полипренолы не влияли. Адеметионин 300 мг/кг практически не менял ни одного из исследованных показателей. Таким образом, полипренолы, в отличие от гептрала, оказывают восстанавливающее (нормализующее) действие на поведение в открытом поле крыс, подвергнутых воздействию CCl4. При этом максимальный эффект оказывали малые (2,2 мг/кг) и средние (4,3 мг/кг) дозы полипренолов. 76 квадраты Физ. р-р + CCl4 45 норки физиологический раствор 8 физиологический раствор Физ. р-р + CCl4 7 40 ропрен 0.5 + CCl4 35 30 25 20 15 ропрен 0.5 + CCl4 6 ропрен 0.75 + CCl4 5 ропрен 1.2 + CCl4 3 ропрен 0.75 + CCl4 4 ропрен 1.2 + CCl4 2 10 гептрал р-р + CCl4 5 0 гептрал табл + CCl4 1 гептрал р-р + CCl4 1 0 1 физиологический раствор стойки Физ. р-р + CCl4 10 9 8 7 6 5 4 3 2 1 0 грумминг ропрен 0.5 + CCl4 10 ропрен 0.75 + CCl4 8 ропрен 0.75 + CCl4 6 ропрен 1.2 + CCl4 4 ропрен 1.2 + CCl4 гептрал р-р + CCl4 1 болю сы физиологический раствор Физ. р-р + CCl4 14 12 ропрен 0.5 + CCl4 гептрал табл + CCl4 гептрал р-р + CCl4 2 0 1 гептрал табл + CCl4 физиологический раствор Физ. р-р + CCl4 2,5 ропрен 0.5 + CCl4 2 ропрен 0.75 + CCl4 1,5 1 ропрен 1.2 + CCl4 0,5 гептрал р-р + CCl4 0 1 гептрал табл + CCl4 Рис. 14. Влияние полипренолов и адеметионина на поведение в «открытом поле» крыс, подвергнутых воздействию CCl4 3.2.3.2. Влияние полипренолов и адеметионина на поведение крыс в тесте «приподнятый крестообразный лабиринт», характеризующем анксиолитические или анксиогенные свойства исследуемых препаратов Вторым поведенческим тестом был приподнятый крестообразный лабиринт. Этот тест позволяет оценить анксиолитические (противотревожные) или анксиогенные (тревожные) свойства фармакологических препаратов, оцениваемые по времени пребывания в затемненных и освещенных рукавах 77 лабиринта. Увеличение времени пребывания в освещенных рукавах лабиринта и повышение числа выглядываний и свешиваний с открытых площадок лабиринта квалифицируется как анксиолитический (противотревожный) эффект, а уменьшение этих показателей – как анксиогенный (тревожный) эффект. Двукратное введение CCl4 (2000 мг/кг) с оценкой поведения через 14 дней после первого введения токсиканта существенно нарушало поведение крыс в приподнятом крестообразном лабиринте (рис. 15). У крыс, получавших CCl4, время пребывания в освещенных (открытых) рукавах лабиринта снижалось в 2,5 раза, число выглядываний из закрытых рукавов – в 4 раза, а число свешиваний с открытых рукавов платформы – в 10 раз. Это указывает на мощное анксиогенное действие токсиканта. В данном тесте ни полипренолы (2,2-4,3-11,6 мг/кг), ни адеметионин в исследуемых дозах не проявляли значимого анксиолитического (противотревожного) действия. время в откры ты х рукавах физиологический раствор Физ. р-р + CCl4 120 ропрен 0.5 + CCl4 100 80 ропрен 0.75 + CCl4 60 ропрен 1.2 + CCl4 40 гептрал р-р + CCl4 20 0 гептрал табл + CCl4 1 свешивания Физ. р-р + CCl4 12 ропрен 0.5 + CCl4 10 8 ропрен 0.75 + CCl4 6 ропрен 1.2 + CCl4 4 гептрал р-р + CCl4 2 0 вы гляды вания физиологический раствор 1 гептрал табл + CCl4 физиологический вопрос Физ. р-р + CCl4 25 20 ропрен 0.5 + CCl4 15 ропрен 0.75 + CCl4 10 ропрен 1.2 + CCl4 5 гептрал р-р + CCl4 0 1 гептрал табл + CCl4 Рис. 15. Влияние полипренолов и адеметионина на поведение в приподнятом крестообразном лабиринте крыс, подвергнутых воздействию CCl4 78 Таким образом, полипренолов и адеметионина при курсовом применении в течение 14 дней не оказывают противотревожного действия в тесте приподнятого крестообразного лабиринта. 3.2.3.3. Влияние полипренолов и адеметионина на поведение крыс в тесте «чужак-резидент», характеризующем агрессивные или защитные формы поведения Следующим поведенческим тестом было изучение агрессии в тесте «чужак-резидент» у крыс. Агрессивное поведение контролируется преимущественно холинергической и дофаминергической системами мозга. Двукратное введение CCl4 (2000 мг/кг) с оценкой поведения через 15 дней после первого введения токсиканта вдвое снижало общительность крыс с умеренным растормаживанием системы «агрессия-защита» в тесте «чужакрезидент». Индивидуальное поведение животных при этом не менялось (рис. 16). Полипренолы (2,2-4,3-11,6 мг/кг) при курсовом введении в течение 14 дней нормализовали показатели коммуникативности и индивидуального поведения, а также снижали повышенную агрессивность с компенсацией системы защиты. Адеметионин также восстанавливал общительность, умеренно снижал агрессивность и полностью подавлял систему защиты. Таким образом, полипренолы и адеметионин в тесте «чужак-резидент» проявляют умеренное антиагрессивное действие, при этом полипренолы по эффективности превышают адеметионин, включая высокую степень защиты (защитного поведения), а адеметионин подавлял этот показатель поведения. 79 коммуникативное физиологический раствор Физ. р-р + CCl4 200 180 160 140 120 100 80 60 40 20 0 ропрен 0.5 + CCl4 индивидуаль ное 80 агрессивное физиологический раствор Физ. р-р + CCl4 4,5 4 3 ропрен 0.75 + CCl4 2,5 2 1 гептрал р-р + CCl4 0,5 0 1 гептрал р-р + CCl4 20 0 гептрал табл + CCl4 1 физиологический раствор защитное Физ. р-р + CCl4 2,5 ропрен 0.5 + CCl4 2 ропрен 0.75 + CCl4 1,5 1 ропрен 1.2 + CCl4 0,5 гептрал р-р + CCl4 ропрен 1.2 + CCl4 1,5 ропрен 1.2 + CCl4 40 ропрен 0.5 + CCl4 3,5 ропрен 0.75 + CCl4 60 гептрал р-р + CCl4 гептрал табл + CCl4 ропрен 0.5 + CCl4 100 ропрен 1.2 + CCl4 1 Физ. р-р + CCl4 120 ропрен 0.75 + CCl4 физиологический раствор гептрал табл + CCl4 0 гептрал табл + CCl4 1 Рис. 16. Влияние полипренолов и адеметионина на поведение в тесте «чужакрезидент» крыс, подвергнутых воздействию CCl4 3.2.3.4. Влияние полипренолов и адеметионина на поведение крыс в плавательном тесте Порсолта, характеризующем антидепрессантное действие препаратов Последним поведенческим тестом был плавательный тест Порсолта. Тест Порсолта используется для оценки уровня депрессии у крыс. Он считается достаточно информативным и специфичным для оценки антидепрессантного действия. Двукратное введение CCl4 (2000 мг/кг) с оценкой поведения через 14 дней после первого введения токсиканта в 4 раза подавлял активное поведение, умеренно снижал пассивное поведение и вдвое повышал иммобилизацию крыс (рис. 17). Последний показатель (обездвиженность, или иммобилизация) является основным показателем теста и в данном случае 80 указывает на появление у отравленных крыс типичного депрессантного поведения. Полипренолы во всех исследованных дозах (2,2-4,3-11,6 мг/кг) практически не меняли активного и пассивного плавания крыс, но восстанавливали иммобилизационное поведение вплоть до контрольного уровня. Адеметионин 300 мг/кг усиливал антиобездвиживающее действие CCl4, не влияя на другие показатели теста Порсолта. активное физиологический раствор Физ. р-р + CCl4 120 ропрен 0.5 + CCl4 100 80 ропрен 0.75 + CCl4 60 ропрен 1.2 + CCl4 40 гептрал р-р + CCl4 20 0 гептрал табл + CCl4 1 пассивное Физ. р-р + CCl4 300 ропрен 0.5 + CCl4 250 200 ропрен 0.75 + CCl4 150 ропрен 1.2 + CCl4 100 гептрал р-р + CCl4 50 0 иммобилизационное физиологический раствор 1 гептрал табл + CCl4 физиологический раствор Физ. р-р + CCl4 80 70 ропрен 0.5 + CCl4 60 50 40 ропрен 0.75 + CCl4 30 ропрен 1.2 + CCl4 20 гептрал р-р + CCl4 10 0 1 гептрал табл + CCl4 Рис. 17. Влияние полипренолов и адеметионина на поведение в тесте Порсолта крыс, подвергнутых воздействию CCl4 Таким образом, полипренолы в дозах 2,2-4,3-11,6 мг/кг проявляют определенный антагонизм по отношению к CCl4 в плавательном тесте Порсолта. Адеметионин подобным действием не обладает, усиливая действие четыреххлористого углерода в тесте Порсолта. 81 3.2.4. Влияние полипренолов и адеметионина на биохимические показатели крови крыс, подвергнутых воздействию CCl4 Результаты биохимических тестов показали, что по таким показателям как холестерин, креатинин и мочевина исследованные группы крыс не отличались (табл. 9), что указывает на отсутствие влияния CCl4 и исследованных соединений (полипренолов и адеметионина) на метаболизм данных биологических субстратов. При этом выявлено умеренное снижение среднего показателя по общему белку в группе животных, которые подвергались воздействию CCl4 и получали полипренолы в дозе 2,2 мг/кг (p < 0,05). По-видимому, полипренолы в этой дозе на фоне действия CCl4 усиливают метаболизм белка в данной группе животных. Достоверное повышение уровня глюкозы в группе животных, которые получали физиологический раствор и CCl4, свидетельствует о неспецифическом влиянии четырёххлористого углерода на метаболизм полисахаридов, либо за счёт нарушения функции печеночной паренхимы, либо вследствие функциональных нарушений со стороны поджелудочной железы о чём свидетельствует достоверное повышение уровня α-амилазы в данной группе крыс (p < 0,05). Значительное повышение уровня лактатдегидрогеназы у крыс, которые получали полипренолы и адеметионин, свидетельствует об индукции синтеза данного фермента в печёночной паренхиме указанными препаратами (возможно, не только в печени, но и других внутренних органах и мышцах). Снижение уровня билирубина в группах животных, подвергшихся воздействию CCl4, может указывать на изменение уровня хилезности сыворотки или степени эмульгации предшественников билирубина под влиянием четырёххлористого углерода. Значительное повышение уровня аспартатаминотрансферазы (АСТ) в эксперименте свидетельствует о повреждающем влиянии CCl4 на печеночную паренхиму, однако, уровень аспартатаминотрансфе- 82 Таблица 9 Влияние полипренолов и адеметионина на течение хронического гепатоза, вызванного CCl4, у крыс Биохимический показатель Общий белок, г/л Глюкоза, ммоль/л Холестерол, ммоль/л Креатинин, мкмоль/л Мочевина, ммоль/л α-амилаза, МЕ/л Лактатдегидрогеназа, МЕ/л Триглицериды, ммоль/л Общий билирубин, мкмоль/л АСТ, МЕ/л АЛТ, МЕ/л Непрямой билирубин, мкмоль/л Щелочная фосфатаза, МЕ/л Мочевая кислота, мкмоль/л Креатинкиназа, МЕ/л Фосфор, ммоль/л ЛПНП, ммоль/л γ-глютамилтрансфераза, МЕ/л Контроль (физ. раствор) ССl4 + физ. раствор ССl4 + полипренолы 2,2 мг/кг ССl4 + полипренолы 4,3 мг/кг 72,45±2,0 70,42±1,47 65,18±2,08* 70,14±0,65 4,85±0,33 6,34±0,46* 4,18±0,44 5,40±0,21 1,87±0,11 1,66±0,09 1,58±0,04 1,87±0,13 45,21±0,95 44,34±1,10 43,82±2,47 44,08±1,08 8,13±0,27 1708,83±40,89 8,14±0,38 1930,8±35,4* 8,52±0,43 2001,6±54,6* 8,84±0,35 1719,6±144,3 22,76±9,32 88,74±49,68* 133,68±50,73** 1659,86±156,75** 0,27±0,008 0,34±0,03 0,39±0,02* 0,43±0,02* 20,0±3,5 123,1±14,4 95,9±3,4 11,24±1,35* 248,62±10,94* 99,92±8,23 7,66±1,49** 213,54±17,81* 95,22±6,45 12,88±1,95* 198,52±19,97* 119,56±18,30* 14,2±1,5 7,17±0,61* 6,08±0,98* 9,06±1,13* 531,45±40,15 437,8±63,5 656,12±87,95* 604,48±49,97 320,0±49,75 165,4±17,2* 159,6±19,55* 214,2±15,3* 4541,0±527,9 3,11±0,13 0,19±0,04 5183,4±402,80 2,62±0,12* 0,12±0,01* 5358,8±354,4 2,12±0,07* 0,17±0,01 5315,8±520,0 2,43±0,10* 0,18±0,02 2,33±0,83 0,98±0,97* 2,40±0,84 1,56±0,90* Примечание. *р<0,05; **p<0,01 по отношению к группе контроля. 83 Таблица 9 (продолжение) Влияние полипренолов и адеметионина на течение хронического гепатоза, вызванного CCl4, у крыс Биохимический показатель Общий белок, г/л Глюкоза, ммоль/л Холестерол, ммоль/л Креатинин, мкмоль/л Мочевина, ммоль/л α-амилаза, МЕ/л Лактатдегидрогеназа, МЕ/л Триглицериды, ммоль/л Общий билирубин, мкмоль/л АСТ, МЕ/л АЛТ, МЕ/л Непрямой билирубин, мкмоль/л Щелочная фосфатаза, МЕ/л Мочевая кислота, мкмоль/л Креатинкиназа, МЕ/л Фосфор, ммоль/л ЛПНП, ммоль/л γ-глютамилтрансфераза, МЕ/л ССl4 + полипренолы 11,6 мг/кг ССl4 + адеметионин 300 мг/кг (лиофилизат) Контроль (физ.раствор) ССl4 + физ.раствор 72,45±2,0 70,42±1,47 67,4±0,91 74,16±1,22 4,85±0,33 6,34±0,46* 4,64±0,72 5,58±0,48 1,87±0,11 1,66±0,09 1,81±0,06 1,91±0,17 45,21±0,95 44,34±1,10 44,98±3,17 58,52±13,11 8,13±0,27 1708,83±40,89 8,14±0,38 1930,8±35,4* 7,65±0,61 1989,0±7,97* 11,06±2,79 1424,2±234,16 22,76±9,32 88,74±49,68* 0,27±0,008 0,34±0,03 0,37±0,01* 0,37±0,03* 20,0±3,5 123,1±14,4 95,9±3,4 11,24±1,35* 248,62±10,94* 99,92±8,23 6,80±0,58** 136,80±47,79 99,38±5,89 14,84±2,21* 212,12±11,68* 102,70±4,79 14,2±1,5 7,17±0,61* 6,06±0,50* 47,76±25,16** 531,45±40,15 437,8±63,5 528,82±40,58 505,8±103,4 320,0±49,75 165,4±17,2* 169,8±12,16* 271,40±38,41 4541,0±527,9 3,11±0,13 0,19±0,04 5183,4±402,80 2,62±0,12* 0,12±0,01* 5036,80±445,16 2,15±0,48* 0,18±0,01 4589,20±721,59 2,58±0,12* 0,13±0,01* 2,33±0,83 0,98±0,97* 0,50±0,30** 1,52±1,11 1315,52±253,22** 1838,90±140,54** Примечание. *р<0,05; **p<0,01 по отношению к группе контроля. 84 разы в группе животных с максимально полученной дозой полипренолов (11,6 мг/кг), который не отличался от контрольных величин (интактный контроль), свидетельствует о выраженных гепатопротекторных свойствах данного вещества. Пониженный уровень мочевой кислоты, которая является конечным продуктом пуринового обмена, в экспериментальных группах может свидетельствовать либо о торможении дезаминирования аминопуринов и/или окисления оксипуринов, либо об ускорении выведения мочевой кислоты в организме. Достоверное снижение уровня фосфора в сыворотке экспериментальных животных может указывать на изменение вводно-электролитного обмена в результате воздействия четырёххлористого углерода. Кроме того, отмечено нормализующее действие полипренолов (2,2-4,3 мг/кг) и адеметионина (300 мг/кг) на активность γ-глутамилтрансферазы, одного из основных маркеров поражения печени. Под влиянием адеметионина более чем в 3 раза возрастал уровень непрямого билирубина, что может указывать на нарушение им эвакуационной функции желчевыводящей системы печени. Таким образом, полученные результаты биохимического анализа указывают, что полипренолы во всех дозах, но особенно в дозе 11,6 мг/кг и адеметионин в дозе 300 мг/кг обладают выраженными гепатопротекторными свойствами. 3.2.5. Влияние полипренолов и адеметионина на морфологическую картину печени на модели подострого гепатоза, вызванного CCl4 у крыс Введение полипренолов зависимо от дозы (2,2-4,3-11,6 мг/кг) защищало печень от токсического действия гепатотропного яда CCl4. 3.2.5.1. Лечение полипренолами в дозе 2,2 мг/кг Так, в группе крыс, получавшие CCl4 + полипренолы 2,2 мг/кг (группа 3), в печени на фоне нарушения балочного строения имелась паренхиматоз- 85 ная дистрофия в виде белковой гиалиново-капельной формы, которая поражала гепатоциты всех печеночных долек на 2/3 их протяженности балок, местами захватывала дольки очагово в виде мелкокапельной жировой дистрофии, локализовалась перипортально, некроз гепатоцитов не наблюдался. Рис. 18. Печень крысы, получавшей CCl4 + физ. раствор (вверху в центре) Дискомплексация балочного строения, умеренно выраженные и выраженные проявления дистрофии гепатоцитов. Окраска гематоксилином и эозином. Увеличение 200. Рис. 19. Печень крысы, получавшей CCl4 + полипренолы 2,2 мг/кг (слева) Дискомплексация балочного строения, выраженные проявления дистрофии гепатоцитов. Окраска гематоксилином и эозином. Увеличение 200. Рис. 20. Печень крысы, получавшей CCl4 + полипренолы 4,3 мг/кг (справа) Дискомплексация балочного строения, умеренно выраженные и выраженные проявления дистрофии гепатоцитов. Окраска гематоксилином и эозином. Увеличение 200. 86 В трактах определялась слабо выраженная мононуклеарная инфильтрация. В соответствие с балльной оценкой степени патогистологических изменений можно было констатировать 3 бальное поражение печени крыс данной группы – гистологические изменения соответствовали картине выраженной токсической паренхиматозной дистрофии печени (рис. 19). 3.2.5.2. Лечение полипренолами в дозе 4,3 мг/кг В группе крыс, получавших CCl4 + полипренолы 4,3 мг/кг в печени еще не наблюдалось долек, имевших нормальное гистологическое строение. Дискомплексация балочного строения, сопровождавшаяся дистрофическими изменениями в виде белковой гиалиново-капельной и мелкокапельной жировой дистрофии, захватывала 1/2 – 3/4 протяженности гепатоцитов в дольках. Некроз гепатоцитов отсутствовал, лимфоидная инфильтрация портальных трактов носила слабо выраженный характер. Эти изменения соответствовали 2–3 бальному поражению печени и картине умеренной и выраженной паренхиматозной токсической дистрофии печени (рис. 20). 3.2.5.3. Лечение полипренолами в дозе 11,6 мг/кг В группе крыс, получавших CCl4 + полипренолы 11,6 мг/кг, в печени также не наблюдалось долек, имевших нормальное гистологическое строение. Дискомплексация балочного строения, сопровождавшаяся белковой гиалиново-капельной и мелкокапельной жировой дистрофией, захватывала 1/3 – 1/2 протяженности гепатоцитов в дольках. Некроз гепатоцитов отсутствовал, лимфоидная инфильтрация портальных трактов носила слабо выраженный характер. Эти изменения соответствовали 2-бальному поражению печени и картине умеренно выраженной паренхиматозной токсической дистрофии печени (рис. 21). 87 Рис. 21. Печень крысы, получавшей CCl4 + полипренолы 11,6 мг/кг (слева) Дискомплексация балочного строения, умеренные проявления дистрофии гепатоцитов. Окраска гематоксилином и эозином. Увеличение 200. Рис. 22. Печень крысы, получавшей CCl4 + адеметионин 300 мг/кг (справа) Дискомплексация балочного строения, слабо выраженные проявления дистрофии гепатоцитов. Окраска гематоксилином и эозином. Увеличение 200. 3.2.5.4. Лечение адеметионином в дозе 300 мг/кг И, наконец, в группе крыс, получавших CCl4 + адеметионин 300 мг/кг, в печени регистрировали слабо выраженное поражение долек. В дольках наблюдалась дискомплексация балочного строения, сопровождавшаяся белковой гиалиново-капельной дистрофией на 1/3 протяженности гепатоцитов балок, в одном случае на 2/3 протяженности. Некроз гепатоцитов отсутствовал, лимфоидная инфильтрация портальных трактов носила слабо выраженный характер. Эти изменения соответствовали слабо выраженной паренхиматозной токсической дистрофии печени и преимущественно 1-бальному поражению печени (рис. 22). Таким образом, после двукратного введения CCl4 (2000 мг/кг) в печени крыс наблюдалась морфологическая картина умеренной и выраженной белковой и жировой паренхиматозной дистрофии печени (гепатоза). В группе крыс, подвергшихся введению гепатопротекторов (полипренолов и адеме- 88 тионина), морфологическая картина умеренной и выраженной белковой и жировой паренхиматозной дистрофии печени уменьшалась (улучшалась). При этом наименьшие проявления паренхиматозной дистрофии выявлены в группе крыс, подвергшихся введению адеметионина и полипренолов в дозе 11,6 мг/кг, что указывает на их гепатопротекторный эффект. 3.2.6. Влияние полипренолов и адеметионина на обмен моноаминов в структурах мозга крыс, подвергнутых воздействию CCl4 Посылкой настоящего исследования явилось допущение, что при экспериментальном подостром гепатозе, вызванном CCl4, может развиваться токсическая энцефалопатия, приводящая к изменению обмена биогенных аминов в головном мозге. С этой целью с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) с электрохимической детекцией анализировали обмен моноаминов в 3 структурах мозга (прилежащем ядре, стриатуме и коре), имеющим самое непосредственное отношение к двигательной (стриатум), эмоциональной (прилежащее ядро) и когнитивной (кора головного мозга) активности. Примеры хроматограмм со стандартами представлены на рис. 23. 89 НА ДА ДОФ УК 5-ГИУК СЕР ГВК Рис. 23. Примеры хроматограмм стандартных растворов норадреналина (НА), дофамина (ДА), серотонина (СЕР), а также 3,4-диоксифенилуксусной (ДОФУК), гомованилиновой (ГВК), 5-гидроксииндолуксусной кислот (5ГИУК) по 6 нг каждого По оси ординат – концентрация исследуемого интермедиата (моль), по оси абсцисс – время (мин). Особенности хроматограмм гомогенатов из структур головного мозга представлены на рис. 24 (прилежащее ядро), рис. 25 (стриатум) и рис. 26 (фронтальная кора). Схема метаболизма дофамина: Дофамин (ДА) → 3-метокситриптамин (3-МТ) → 3,4-дигидроксифенилуксусная кислота (ДОФУК) → гомованилиновая кислота (ГВК). Схема метаболизма серотонина: Серотонин (СЕР) → 5-гидрокси-3индолуксусная кислота (5-ГИУК). 90 ДА НА ДОФ УК 5-ГИУК СЕР ГВК Рис. 24. Хроматограмма гомогената из прилежащего ядра крысы Буквенные обозначения, как на рис. 23. ДА ДОФУК НА 5-ГИУК ГВК Рис. 25. Хроматограмма гомогената из стриатума крысы Буквенные обозначения, как на рис. 23. СЕР 91 НА ДА ДОФУК СЕР 5-ГИУК Рис. 26. Хроматограмма гомогената из фронтальной коры мозга крысы Буквенные обозначения, как на рис. 23. Анализ показал, что введение четыреххлористого углерода и исследуемых препаратов (полипренолов и адеметионина) существенно не влияло на уровень медиаторов норадреналина, серотонина, а также 5- гидроксиуксусной кислоты (5-ГИУК) в прилежащем ядре, стриатуме и фронтальной коре, но меняло их обмен (метаболизм). В частности, однофакторный дисперсионный анализ показал различия в уровне гомованилиновой кислоты (ГВК) и отношения ГВК/ДА в прилежащем ядре между экспериментальными группами (группы 3 и 4, р=0,0009 и р=0,0457, соответственно). Итак, введение CCl4 умеренно снижало уровень ГВК. Полипренолы в дозах 2,2 и 4,3 мг/кг достоверно (приблизительно в 2 раза) увеличивали содержание ГВК в прилежащем ядре по сравнению с обеими контрольными группами (то есть, между группами 3 и 4 и группой 1, р < 0,01, а также группой 2, р < 0,05 соответственно). Более наглядно это представлено на рис. 27. 92 DA ACB DA DOPAC 9 8 7 6 5 4 3 2 1 0 HVA 1.5 1.0 * * 1.0 0.5 0.5 g1 g2 g3 g4 g5 g6 0.0 0.0 g1 g2 g3 g4 g5 g6 g1 g2 DOPAC/DA g3 g4 g5 g6 g5 g6 HVA/DA 0.25 0.125 0.20 0.100 0.15 0.075 0.10 0.050 0.05 0.025 0.00 0.000 g1 g2 g3 g4 g5 g6 g1 g2 g3 g4 Рис. 27. Показатели обмена дофамина в прилежащем ядре крыс По оси ординат – уровень медиатора или метаболита (нмоль/г ткани); по оси абсцисс (латинскими буквами) – номер экспериментальной группы Однофакторный дисперсионный анализ показал также различия в уровне ДОФУК и отношении ГВК/ДА в стриатуме между экспериментальными группами (группы 3 и 4 и группами 1 и 2, р=0,0412 и р=0,0390, соответственно). Полипренолы в дозах 2,2 и 4,3 мг/кг снижал содержание ДОФУК в стриатуме по сравнению с контрольной группой (р < 0,05), что представлено на рис. 28. 93 DA STR DOPAC DA HVA 2 1.5 10.0 7.5 1.0 * * 1 5.0 0.5 2.5 0 0.0 g1 g2 g3 g4 g5 g1 g6 g2 g3 g4 g5 g6 0.0 g1 g2 DOPAC/DA g3 g4 g5 g6 g5 g6 HVA/DA 0.2 0.2 0.1 0.1 0.0 0.0 g1 g2 g3 g4 g5 g6 g1 g2 g3 g4 Рис. 28. Показатели обмена дофамина в стриатуме крыс По оси ординат – уровень медиатора или метаболита (нмоль/г ткани); по оси абсцисс (латинскими буквами) – номер экспериментальной группы. Таким образом, полученные данные свидетельствуют о разнонаправленном влиянии полипренолов на уровень ГВК и ДОФУК в прилежащем ядре и стриатуме соответственно, что в свою очередь может указывать на активацию мезолимбической дофаминергической системы (ответственна за подкрепление) и снижение активности нигростриатной дофаминергической системы (ответственна за двигательную активность). Однако, учитывая общую тенденцию к снижению отношения ДО- ФУК/ДА и увеличению отношения ГВК/ДА в обеих исследованных структурах можно думать об ускоренном обмене дофамина в мозге под влиянием 94 полипренолов, поскольку ДОФУК является промежуточным метаболитом дофамина по отношению к ГВК, и/или смещению метаболизма дофамина в сторону КОМТ-зависимого пути (как показано на рис. 29). Таким действием обладают некоторые нейролептики (включая атипичные), проявляющие свойства антагонистов рецепторов дофамина. Рис. 29. Схема метаболизма дофамина В то же время следует отметить, что адеметионин никаким образом не влиял на обмен дофамина и норадреналина в головном мозге, но несколько ускорял обмен серотонина во фронтальной коре, умеренно повышая в ней уровень 5-гидроксиуксусной кислоты (рис. 30). Прежде всего, с помощью однофакторного дисперсионного анализа были найдены различия в индексе активности серотонинергической системы 5-ГИУК/СЕР во фронтальной коре 95 между экспериментальными группами (группами 6 и 2, р=0,0157). При этом адеметионин (р < 0,05) достоверно увеличивал соотношение 5-ГИУК/СЕР по сравнению с группами, получавшими четыреххлористый углерод, а полипренолы в исследуемых дозах лишь проявили тенденцию к повышению данного соотношения. Это может свидетельствовать о нормализующем влиянии обоих исследованных препаратов (адеметионина и полипренолов) на активность серотонинергической системы во фронтальной коре. Подобный эффект отмечен и в работе (Ротенко А.А. и др., 2006), когда при хроническом гепатозе, вызванном четыреххлористым углеродом, авторы регистрировали повышение уровня серотонина в целом мозге крыс. SER Frontal cortex SER HIAA 3 2 2 1 1 0 0 g1 g2 g3 g4 g5 g6 g1 g2 g3 g4 g5 g6 HIAA/SER * 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0.0 g1 g2 g3 g4 g5 g6 Рис. 30. Показатели обмена серотонина во фронтальной коре мозга крыс По оси ординат – уровень медиатора или метаболита (нмоль/г ткани); по оси абсцисс (латинскими буквами) – номер экспериментальной группы. 96 Следовательно, гепатопротекторных можно свойств, заключить, обладают что полипренолы выраженным помимо центральным действием, которое заключается в умеренной психоактивации, главным образом за счет изменения активности дофаминергических систем мозга. 97 Глава 4. ОБСУЖДЕНИЕ ПОЛУЧЕННЫХ РЕЗУЛЬТАТОВ В первую часть исследования было включено изучение влияния полипренолов в дозах 4,3-8,6-13,4 мг/кг после курсового введения препарата в течение 5-7 дней на ряд поведенческих тестов, позволяющих охарактеризовать психотропный спектр активности препарата. Часть из тестов (ротационный тест, агрессивность в методике «чужак-резидент», двигательная активность в «открытом поле», самостимуляция латерального гипоталамуса) могут рассматриваться как типичные для анализа дофаминзависимого поведения, поскольку данные реакции в основном контролируются дофаминергической системой мозга (Шабанов П.Д. и др., 2002, 2004, 2006, 2008; Salamone JD, Correa M., 2012). Остановимся более подробно на этих формах поведения. Дофаминзависимые формы поведения представляют собой наиболее типичные поведенческие паттерны, воспроизводимые при активации дофаминергической системы мозга (Отеллин В.А., 1990; Стрельцов В.Ф., 2003, 2009; Curtis J.T., Wang Z., 2005; Comai S, et. al., 2012). Они достаточно разнообразны, поскольку в головном (среднем) мозге дофаминергические нейроны формируют унилатерально восходящие проекции, среди которых условно выделяют четыре функциональных системы (Björklund A., Hökfelt T., 1984; Раевский К.С. и др., 1996; Угрюмов М.В., 1998; Lieu C.A, Subramanian T., 2012): 1) нигростриатную (формируется клетками группы А9, иннервирующими хвостатое ядро и неостриатум); 2) мезолимбическую (формируется клетками группы А10, иннервирующими прилежащее ядро, обонятельные бугорки, центральное и базолатеральное ядра миндалины, перегородку, дорсолатеральную часть интерстициального ядра конечной полоски, фронтальную, цингулярную энториальную кору и вентральные отделы полосатого тела); 3) мезокортикальную (формируется клетками группы А10, иннервирую- 98 щими вентромедиальную область покрышки, и клетками группы А9, иннервирующими префронтальную, переднюю цингулярную, энториальную и пириформную кору, а также глубокие слои фронтальной коры) и 4) тубероинфундибулярную (формируется клетками групп А12-А14, иннервирующими срединное возвышение, ножки гипофиза, заднюю и среднюю доли гипофиза, дорсальные и передние отделы гипоталамуса). Часто мезолимбическую и мезокортикальную системы функционально объединяют в единую мезокортиколимбическую ДА-ергическую систему (Лебедев А.А., Шабанов П.Д., 1992; Шабанов П.Д. и др., 1999, 2000, 2002; Amano T. et al,. 2012 ). Первые доказательства существования рецепторов дофамина в мозге и связь их с системой аденилатциклазы были получены в 1972 г. В 1978 г. было доказано, что рецепторы дофамина в нервной системе представлены, по крайней мере, двумя дискретными популяциями, одна из которых связана с системой 3’5’-циклического аденозинмонофосфата (цАМФ), другая независима от него. В частности, было продемонстрировано, что в гипофизе дофамин тормозит секрецию пролактина, но не стимулирует формирования цАМФ, и что нейролептик сулпирид, будучи антагонистом дофамина, не влияет на дофамин-чувствительную аденилатциклазу в аденогипофизе (Kebabian J., Calne D., 1979). Дж. Кебабян и Д. Калне (Kebabian J., Calne D., 1979), суммировав эти данные, предложили подразделить рецепторы дофамина на D1, связанные со стимуляцией аделатциклазы, и D2, не связанные с этим эффектом. В дальнейшем это разделение рецепторов дофамина на два подтипа было подтверждено фармакологически, биохимически, физиологически и морфологически. Дуалистическая концепция дофаминергической передачи была и остается основополагающей до настоящего времени. Тем не менее, методами клонирования выделены и охарактеризованы еще три подтипа рецепторов дофамина: D3, D4 и D5/1b. Однако оказалось, что выделенные подтипы рецепторов могут быть принципиально отнесены к первоначальным двум семействам рецепторов дофамина. В настоящее время следует считать 99 признанной классификацию рецепторов дофамина с выделением двух семейств – D1, объединяющего D1 и D5/1b подтипы рецепторов, и D2, объединяющего D2, D3 и D4 подтипы (Missale C. et al., 1998). У млекопитающих D1b рецептор, первоначально названный так благодаря высокой гомологии с D1 рецептором, чаще выделяют как D5 рецептор дофамина (de Manzano Ö. et al., 2013). Возвращаясь к дофаминзависимым формам поведения, следует подчеркнуть, что в любом поведенческом акте они присутствуют (Febo M., Ferris CF., 2007). Вследствие этого выделяют два блока дофаминзависимого поведения: 1) двигательный (связан с активацией нигростриарной системы мозга), который проявляется усилением двигательной активности, появлением агрессивности, повышением ротационного (вращательного) компонента поведения и стереотипий (Cools R., 2011) и 2) эмоционально-мотивационный (связан с активацией мезокортиколимбической системы мозга), проявляющийся усилением мотивации пищевого и полового поведения и активацией реакции самостимуляции лимбических структур мозга и, прежде всего, латерального гипоталамуса, через который проходит передний медиальный дофаминергический пучок (Sagara H. et al., 2010). Именно это и послужило отправной точкой в выборе методик исследования, вошедших в первый раздел настоящей работы, целью которого было оценить возможное влияние препарата полипренолов на дофаминзависимые формы поведения у грызунов (крыс). Различные эмоции, как и другие функциональные состояния, лежащие в основе многочисленных поведенческих актов, возникают в результате установления определенных взаимосвязей между разными структурами мозга, в первую очередь между корой больших полушарий и подкорковыми центрами (Менделевич В.Д., 2006; Менделевич В.Д., Зобин М.Л., 2012; Göderqvist S. et al., 2012). Начиная с 1920-х гг., были проведены нейрофизиологические эксперименты с раздражением структур мозга 100 бодрствующих животных (кошки) электрическим током, позволившие установить ряд важных закономерностей, касающихся структурно- функциональной организации эмоций (Симонов П.В., 1987, 2004; Вартанян Г. А., Петров Е.С., 1989; Шабанов П.Д. и др., 2002; Сапронов Н.С., 2005; Göderqvist S. et al., 2012). Во-первых, стало ясно, что одним из важнейших центров регуляции эмоционального поведения является подбугорная область мозга, или гипоталамус. Во-вторых, эмоциональные функции гипоталамуса контролируются передним мозгом (корой больших полушарий). И, наконец, в-третьих, эмоциональные реакции могут проявляться только в том случае, когда сохранены нервные связи между гипоталамусом и нижележащими структурами среднего мозга. В дальнейшем более углубленное изучение нейрофизиологами функций головного мозга показало, что гипоталамус является важным, но не единственным центром, регулирующим эмоциональное поведение животных (Воеводин Е.Е., 2007; Aragona B.J.et. al., 2007) Реакцию ярости у кошек удавалось воспроизвести при раздражении некоторых отделов среднего мозга даже в тех случаях, когда средний мозг был хирургически отделен от всех вышележащих образований центральной нервной системы. эмоциональные Эти центры эксперименты среднего мозга продемонстрировали, обеспечивают что координацию двигательных компонентов эмоциональной реакции, но сами по себе без регулирующего воздействия высших эмоциональных центров генерировать эмоциональные состояния не способны. Постепенно стало ясно, что эмоциональное поведение целостного организма определяется сложным взаимодействием многих центров головного мозга, составляющих определенную систему, получившую название «лимбической» (рис. 31). 101 Рис. 31. Лимбическая система мозга человека Центральным образованием, своеобразным распределительным центром эмоций, является гипоталамус. Проводящими путями гипоталамус связан с передневентральным ядром таламуса и через него с поясной извилиной, особой зоной коры больших полушарий, ответственной за восприятие и осознание эмоций (Carlezon WA, Jr, Thomas MJ., 2009). Эфферентные пути, несущие импульсацию к двигательным (соматическим) и вегетативным центрам среднего, продолговатого и спинного мозга, проходят через гиппокамп и мамиллярные тела гипоталамуса. В целом рефлекторная регуляция эмоциональных состояний выглядит следующим образом (Симонов П. В., 1987; Шабанов П. Д., Калишевич С. Ю., 1998; Morikawa H, Paladini C.A., 2011). Дистантные рецепторы (обоняние, слух, зрение) по своим проводящим путям приносят в кору больших полушарий информацию о постоянно изменяющейся внешней среде (Morava E.A. et al., 2011) В соответствующие корковые зоны поступает также тактильная информация и импульсация от интерорецепторов, локализованных во внутренних органах. Анализ полученной информации завершается формированием афферентной 102 модели предполагаемых результатов действия организма (акцептор результата действия по П. К. Анохину). Обширные связи корковых зон с группами ядер гипоталамуса и поступление в эти структуры нервной импульсации вызывает формирование определенного эмоционального состояния. Через связи гипоталамуса с поясной извилиной возникает осознание положительной или отрицательной эмоции, а благодаря эфферентным связям поясной извилины осуществляются двигательные и вегетативные компоненты эмоционального поведения (Конорский Ю., 1970; Цикунов С.Г., 2008; Cagiano R. et al., 2008). Эмоциональное поведение может модифицироваться или трансформироваться в зависимости от совпадения или несовпадения афферентной модели предполагаемых результатов и реального результата действия организма (Вальдман А. В. и др., 1988; Шабанов П. Д., Калишевич С. Ю., 1998; Шабанов П.Д. и др., 2004; DeLeon K.R., 2005; Glimcher P.W. et al., 2011). В 1950-е гг. появились новые возможности изучения нейрофизиологических механизмов эмоций с помощью самораздражения (самостимуляции) структур мозга электрическим током у животных ( золотые рыбки, крысы, кошки, обезьяны). В лаборатории Дж. Олдса было показано, что в зависимости от локализации электродов животное (чаще крыса) самостимулировало определенные зоны мозга, названные стартзонами, с частотой до 5000 стимулов в 1 ч и до 200 тысяч раздражений за 20 ч непрерывного эксперимента, после чего, как правило, наступало его полное изнеможение (Симонов П. В., 1987; Вальдман А. В. и др., 1988; Nathaniel T.I., 2009). Раздражение мозга осуществлялось животным, даже если на пути к контакту оно должно было преодолеть серьезные препятствия, например решетчатый пол, через который пропускали надпороговые значения электрического тока. В течение сеанса самораздражения животное отказывалось от пищи, не реагировало на появление в камере посторонних предметов, шума, вспышек света. Самцы крыс не обращали никакого 103 внимания на появление в камере самок. Иначе говоря, самораздражение становилось основным смыслом жизнедеятельности животного в данной ситуации. Напротив, даже одиночное раздражение центров отрицательных эмоций (стоп-зон) вызывало резко негативную реакцию. Животное стремилось избежать приближения к контактам, а при условии, когда животное, например обезьяна, было лишено возможности избежать раздражения из-за жесткой фиксации в специальном аппарате, оно астенизировалось, отказывалось от пищи, у него выпадали волосы, развивались функциональные расстройства сердечно-сосудистой деятельности, дисфункции желудочно-кишечного тракта. Эксперименты с самораздражением эмоциональных центров позволили картировать мозг с выделением положительных и отрицательных зон. В это же время возникают представления о подкрепляющих системах мозга как структурно-функциональной основе эмоционального поведения. Этому способствовали успехи развития нейроморфологии и функциональной нейрохимии, доказавших, что подкрепляющие системы мозга представляют собой специализированные проводящие пути в центральной нервной системе, медиаторами в которых были идентифицированы преимущественно катехоламины (норадреналин, дофамин) и серотонин. Действительно, наложение карт зон самостимуляции и топографии моноаминергических путей головного мозга дало перекрытие этих зон более чем на две трети (Вальдман А. В. и др., 1988; Вартанян Г. А., Петров Е. С., 1989; Blum K., 2012). С 1980-х гг. изучение подкрепляющих систем головного мозга идет в основном по пути уточнения и детализации участия отдельных морфофункциональных образований и звеньев моноаминергических систем в феномене самораздражения мозга и других поведенческих моделях (самовведение доказательства, веществ, что предпочтение существенную места роль в и т. д.). Появляются реализации механизмов подкрепления играют эндогенные опиоиды (β-эндорфин, энкефалины) и 104 опиоидные рецепторы мозга. Остается неясным вопрос о долевом (или преимущественном) участии дофамина (ДА), норадреналина, серотонина, глутамата, опиоидов функционировании в механизмах подкрепляющих эмоционального систем мозга поведения (Шабанов и П. Д., Калишевич С. Ю., 1998 Blum K., 2012). В ряде работ, выполненных в лаборатории П.Д.Шабанова (Шабанов П.Д. и др., 2002, 2004, 2006, 2008, 2011) рассматривается и обосновывается гипотеза, что в основе внутримозгового механизма подкрепления лежит единый интегрированный механизм изменения градиента эмоциональности в сторону его возрастания или убывания (концепция эмоционального градиента). При этом состояние единого механизма внутримозгового подкрепления определяется его связью с конкретной потребностью организма в данный момент, которая в своем психофизиологическом выражении имеет, как правило, отрицательную эмоциональную окраску. Оценка вероятности удовлетворения самых разных потребностей при выделении доминирующей потребности создает особенности и специфику указанного градиента, обусловливает вовлечение конкретных механизмов его осуществления. Роль морфофункционального субстрата подкрепления играет мезокортиколимбическая система мозга. С нейрохимических позиций в нем участвуют различные нейромедиаторные системы, включая систему дофамина (ДА), норадреналина, серотонина, ГАМК, глутамата, опиоидов. Такая многосложная, нейробиологической иерархически основой построенная реализации система указанного является механизма подкрепления (Van den Oever M.C. et al., 2012) Многочисленными работами (Вартанян Г. А., Петров Е. С., 1989; Lebedev A. A. et al., 1993; LeMoal M., Simon H., 1991; Panchenko G. N. et al., 1998; McBride W. J., Murphy J. M., Ikemoto S., 1999; Koob G.F., 2011) показано, что реакция самостимуляции реализуется при участии целого ряда структур лимбико-диэнцефального комплекса и коры головного мозга и 105 включении многих нейромедиаторных систем. Основное внимание наших исследований уделялось роли мезокортиколимбической ДА-ергической системы в механизмах Мезокортиколимбическая внутримозгового система включает подкрепления. вентральную область покрышки, миндалевидный комплекс, медиальный передний мозговой пучок, прилежащее ядро и медиальную префронтальную кору (Дробленков А.В. , 2006) (рис. 32). Рис. 32. Мезокортиколимбическая дофаминергическая система мозга крысы (Шабанов П.Д. и др., 2002) С помощью нейрофармакологического анализа в опытах на крысах было показано синергичное включение D1 и D2-подобных семейств рецепторов ДА в реализацию указанного механизма (Petrov E.S., Lebedev A.A., 1997; Panchenko G.N. et al., 1998; Шабанов П.Д. и др., 2002). Было доказано, что ДА-ергический механизм является определяющим в реализации поведения самостимуляции у крыс (Tritsch N.X., Sabatini B.L.., 2012). Кроме того, продемонстрирована модулирующая роль адренорецепторов, рецепторов ГАМК и глутамата, опиоидных рецепторов, а также стимуляторов и ингибиторов синтеза серотонина при действии 106 агонистов ДА на реакцию самостимуляции (Шабанов П. Д., Лебедев А. А., 1994; Aragona B.J., Wang Z.., 2007: Guarraci F.A., 2008). При этом выявлены особенности выработки, пороговые значения, частотные характеристики реакции самостимуляции основных структур мезокортиколимбической системы (Петров Е.С., 1990; Petrov E.S., Lebedev A.A., 1997). Во время реакции самостимуляции наблюдались элементы мотивационного поведения: грызение и лизание педали, фрагменты полового поведения (эрекция). Интересно отметить, что разность пороговых значений силы тока для реакции самостимуляции и наблюдаемых паттернов поведения увеличивалась при локализации электродов в каудо-ростральном (заднепереднем) направлении и достигала наибольших значений в медиальной префронтальной коре (Вартанян Г.А., Петров Е.С., 1989; Petrov E.S., Lebedev A.A., 1997). Проявление мотивационных элементов поведения оказалось зависимым от силы тока стимуляции, что согласуется с экспериментами П. В. Симонова (1987), показавшего аналогичную закономерность для стимуляции структур лимбической системы мозга, в частности гиппокампа и миндалины. По-видимому, значимость подкрепления определяется тонкой регуляцией именно мотивационных систем. П.В. Симонов (1987) выделяет специальный блок структур (миндалину и гипоталамус), которые в наибольшей степени регулируют мотивационные аспекты интегрированного эмоционального ответа. Кроме того, можно думать и о преимущественном включении определенных нейрохимических систем (или рецепторов в пределах нейрохимической системы) в регуляцию данного ответа организма. Так, в экспериментах Е.С. Петрова и А.А. Лебедева (1997) при использовании низких значений силы тока, когда фрагменты поведения проявлялись в большей степени, антагонист D1-рецепторов ДА SCH23390 (0,05 мг/кг) блокировал вызванную фенамином (1 мг/кг) активацию реакции самостимуляции. При высоких значениях силы тока введение SCH23390 не вызывало подобного эффекта (Petrov E.S., Lebedev A.A., 1997). 107 В эти же годы были исследованы особенности включения мнестических механизмов мозга в реализацию подкрепляющих эффектов фенамина (Лебедев А.А., Шабанов П.Д., 1992, Лебедев А.А. и др., 1995). Фенамин относят к непрямым адреномиметикам, основным механизмом действия которого является усиление высвобождения ДА и норадреналина из пресинаптических терминалей, вследствие чего происходит активация постсинаптических рецепторов ДА и адренорецепторов. Фенамин часто используют в виде средства-анализатора для решения вопросов, связанных с активацией ДА-ергической системы мозга. В отношении реакции самораздражения гипоталамуса фенамин в диапазоне доз от 0,5 до 5 мг/кг неизменно проявляет стимулирующий эффект. В дозе 1 мг/кг фенамин увеличивает реакцию самостимуляции на 35-40%. При сравнении безусловных (первичных) и условных (вторичных) подкрепляющих эффектов фенамина с помощью реакции самостимуляции и условной реакции предпочтения места были показаны разные дозозависимые характеристики. Была выявлена дозозависимая диссоциация эффектов фенамина (Лебедев А. А., Шабанов П. Д., 1992). Она проявлялась в том, что фенамин в меньшей дозе (1 мг/кг) вызывал бόльший стимулирующий эффект в отношении реакции самостимуляции, чем в отношении условного предпочтения места, и наоборот, в бόльшей дозе (5 мг/кг) препарат в меньшей степени активировал реакцию самостимуляции, чем предпочтение места. Несоответствие эффектов разных доз фенамина при исследовании реакции самостимуляции и условного предпочтения места можно объяснить различной физиологической сущностью применяемых методов, а именно: в основе действия вещества при самостимуляции лежит активация эмоционально-мотивационных механизмов, тогда как условное предпочтение места детерминируется как эмоциональными, так и мнестическими компонентами условнорефлекторного поведения (Шабанов П.Д., Бородкин Ю.С., 1989; Burbassi S., Cervo L., 2008). Повышение частоты реакции 108 самостимуляции наблюдается непосредственно после введения фенамина и представляет собой отражение подкрепляющих свойств таких искусственных безусловных раздражителей как электрическая стимуляция мозга, участвующих в естественной регуляции положительных эмоциональных состояний (Хананашвили М.М., 1983; Вальдман А.В. и др., 1988). Вторичноподкрепляющее действие фенамина наблюдается после ряда сочетаний препарата с окружающей обстановкой, но без его введения в последний день эксперимента (Alcaro, A. et al., 2007). При повторных сочетаниях фармакологического препарата с обстановочными стимулами между ними возникает ассоциация, и сигналы обстановки становятся условными сигналами, вызывающими условнорефлекторную реакцию. Хотя давно сложились представления о том, что энграмма памяти делокализована, тем не менее, передние отделы новой коры и гиппокамп являются именно теми структурами, которые в бόльшей степени вовлекаются в процесс формирования памятного следа (Шабанов П.Д., Бородкин Ю.С., 1989; Преображенская Л.А., 1991; Mameli M., Lüscher C., 2011; Parkes S.L., Westbrook R.F., 2010). По-видимому, можно думать и о преимущественном включении определенных нейрохимических систем или их отдельных компонентов в формирование отсроченных реакций, ассоциированных с подкрепляющим действием препарата или иного биологически значимого стимула (Bardo MT, Bevins RA., 2000). Так, условная реакция предпочтения места наблюдается при сочетаниях обстановки как с электрической стимуляцией положительных эмоциогенных зон головного мозга, так и c подачей пищи (Вартанян Г.А., Петров Е.С., 1989; Лебедев А.А., Шабанов П.Д., 1992; Мещеров Ш.К., 2001; Mameli M., Lüscher C., 2011). Таким образом, реализация единого механизма внутримозгового подкрепления зависит как от включения эмоционально-мотивационных, так и мнестических компонетов. Предполагается, что использование различных методов исследования (реакция самостимуляции в различных модификациях, 109 условная реакция предпочтения места), преимущественно связанных с активностью того или иного компонента, позволяет выявлять нейрохимические звенья обеспечения деятельности подкрепляющих систем мозга. Реализация механизма внутримозгового подкрепления обеспечивается взаимосвязанной работой ряда нейрохимических систем мозга (ДА-, норадреналин-, серотонин-, ГАМК-, глутаматергической, опиоидной) (Елисеева А.П., 2005). Степень вовлечения нейрохимической системы зависит от ее включения в функциональный компонент единого механизма подкрепления (Balfour DJ., 2009; Koob G.F., 2011). Предполагается, что патологические изменения функционирования подкрепляющих систем мозга связаны с нарушением и с разобщением связи между описанными компонентами. В этих условиях может наблюдаться дисбаланс нейрохимических систем, изменение чувствительности рецепторов, синтеза медиаторов и их выделения из пресинаптической терминали (Шабанов П.Д. и др., 2002, 2004, 2006, 2008, 2011; Любимов А.В., 2012; Tritsch N.X., Sabatini B.L., 2012; Gorwood P. et al., 2012). Возвращаясь к обсуждению полученных данных, напомним, что в тесте «открытого поля» полипренолы не проявляли дозозависимого эффекта. При этом полипренолы растормаживали (активировали) горизонтальную двигательную активность при умеренном снижении всех других поведенческих паттернов (прежде всего, исследовательской активности и эмоциональности). Именно двигательная активность контролируется преимущественно дофаминергической системой, тогда как исследовательская активность (норковый рефлекс), груминг и эмоциональные проявления в значительной степени зависят от свойств серотонинергической и холинергической систем мозга (Avena N.M., Rada P.V., 2012). В ротационном тесте, типичном для изучения дофаминзависимого поведения грызунов, полипренолы только в максимальной выбранной нами дозе (13,4 мг/кг) в отличие от фенамина (непрямого дофаминомиметика) уме- 110 ренно уменьшали число вращений в ротометре и не вызывали стереотипий (повторяющихся крупноразмашистых кивков головой, повторных непрекращающихся вращений только в одну сторону, перебирание лапами по типу гребли, навязчивого грызения). Это указывает на возможное участие нигростриарной части дофаминергической системы мозга (ее умеренное подавление в виде блокады постсинаптических рецепторов дофамина) в эффектах полипренолов. Данный эффект важен для проявления свойств атипичного нейролептика (умеренный нейролептический, антипсихотический, миорелаксирующий, вегетотропный и потенцирующий действие депримирующих средств эффекты). Кроме того, полипренолы проявляли умеренную анксиогенную (тревожную) активность, которая наиболее стабильно воспроизводится при введении пренолов в дозе 13,4 мг/кг. Этот вид активности характерен для ряда фармакологических средств, оказывающих судорожное действие (судорожные агенты в субсудорожных дозах, психостимуляторы типа амфетамина, сиднокарба, кофеина). Подобный эффект может проявляться и у атипичных нейролептиков, он обычно рассматривается как растормаживающий (Шабанов П.Д., 2008; Dalley J.W., Roiser J.P., 2012). В тесте «чужак-резидент» полипренолы также не оказывали дозозависимого эффекта, значимо подавляя индивидуальное поведение и общительность (коммуникативность) животных. В дозе 13,4 мг/кг пренолы влияли на все исследуемые показатели данного теста. Видимого антиагрессивного действия пренолы не выявили. В тесте Порсолта полипренолы (4,3-8,6-13,4 мг/кг) оказывали депрессантный эффект на поведение. В дозе 13,4 мг/кг пренолы проявляли максимальное в данных опытах действие. Степень выраженности депрессантного эффекта полипренолов может рассматриваться как умеренная. Это также подтверждает, что направленность действия больших доз пренолов депримирующая (подавляющая функции центральной нервной системы). 111 Весьма любопытные результаты получены при изучении влияния полипренолов на реакцию самостимуляции в камере Скиннера. При этом действие полипренолов на данную реакцию зависело от продолжительности его введения. В первые дни введения полипренолы, как правило, умеренно подавляли реакцию самостимуляции, затем эффект сменяется на умеренный психоактивирующий, который по мере введения препарата снижался. Кроме того, полипренолы не проявляли функционального антагонизма с психостимулятором фенамином (амфетамином) и на фоне его введения фенамин в полной мере развертывал свой психоактивирующий эффект. Данные факты позволяют полагать, что полипренолы влияют на мезокортиколимбическую дофаминергическую систему мозга (нейрохимическую основу подкрепления), модулируя ее, то есть, либо подавляя (преимущественно в первые 2 дня введения и, особенно, в дозе 4,3 мг/кг), либо активируя (в дозах 8,6 и 13,4 мг/кг на 3-5 дни введения). Однако данный эффект, по-видимому, не связан с влиянием на пресинаптические механизмы выделения медиатора из пресинаптической терминали, а также с механизмом обратного захвата ими дофамина (отсутствие функционального антагонизма с фенамином, способствующим высвобождению дофамина из пресинаптических депо и блокирующего его обратный захват). Не исключена в этом случае и частичная блокада постсинаптических рецепторов дофамина, когда полипренолы могут проявлять свойства парциального агониста (агониста-антагониста) (Carboni E. et al., 2004) Подобные свойства проявляет ряд фармакологических препаратов опиоидного типа действия (пентазоцин, буторфанол, налбуфин), которые являются агонистами каппа- и дельта-опиоидных рецепторов, но антагонистами мю-опиоидных рецепторов (Звартау Э.Э., 1988). Это не препятствует им проявлять аналгетические свойства, но блокировать эффекты морфина и аналогов, являющихся типичными агонистами мю-рецепторов (Шабанов П.Д., 2008). Из дофаминергических средств аналогичным действием обладает апо- 112 морфин. В дозе 0,5 мг/кг у грызунов он активирует пресинаптические D2рецепторы дофамина, по механизму ауторегуляции блокируя высвобождение дофамина из пресинаптических депо и оказывая, по сути, антагонистическое действие на постсинаптическую мембрану. В дозе 5 мг/кг (в 10 раз больше) апоморфин активирует постсинаптические D1- и D2-рецепторы дофамина (Шабанов П.Д. и др., 2002; Afonso V.M.et al. 2009; Pare D, Duvarci S., 2012), выявляя типичный дофаминомиметический эффект (повышение двигательной активности, агрессии, усиление ротационного поведения, появление стереотипий). Кроме того, в малых дозах (как правило, в 50-100 раз меньше, чем терапевтические) антагонисты рецепторов могут проявлять свойства агонистов. В этом случае в качестве первой фазы действия возникает возбуждение рецепторов с последующей их блокадой. Такой тип действия характерен для барбитуратов (противосудорожных), транквилизаторов бензодиазепинового ряда, нейролептиков, аналгетиков и представителей других классов фармакологических веществ (Adams D.B.. 2006). Все представленные данные позволяют сделать предварительное заключение о возможном двухфазном действии полиролов, проявляющемся в виде парциального агонизма в отношении рецепторов дофамина. Для уточнения этого вопроса (подтверждения либо снятия) необходимо провести исследования эффектов полипренолов в дозах 1-2 мг/кг и 20-25 мг/кг (разница в один порядок) с использованием дополнительного фармакологического анализа, включающего введение агонистов и антагонистов разных подтипов рецепторов дофамина. Выбранный диапазон доз, использованный в наших исследованиях (4,3-8,6-13,4 мг/кг), не позволил сделать однозначный вывод относительно дофаминергического механизма действия полипренолов, что было в дальнейшем исследовано дополнительно в модели подострого токсического гепатоза с энцефалопатией, вызванной четыреххлористым углеродом. Современные биохимические исследования в значительной степени подверглись диверсификации в зависимости от направлений и потребностей 113 клинической медицины. Однако по-прежнему существует стандартный перечень биохимических параметров, которые позволяют проводить эффективный мониторинг метаболических изменений организма. Несомненно, что к таким параметрам относится исследование на глюкозу, что позволяет выявить нарушения в метаболизме полисахаридов, исследования на липиды, включая оценку уровня холестерина и триглицеридов, исследование на αамилазу, как показатель функциональных нарушений со стороны поджелудочной железы. Особое место в батарее биохимических тестов занимает оценка уровня в плазме крови печеночных ферментов (АЛТ, АСТ) и билирубина. Соотнесение полученных значений с референтными значениями позволяет выявить изменения в паренхиме печени. Уровень лактатдегидрогеназы (МВ–фракция) в плазме крови позволяет судить о структурно- функциональных изменениях миокарда. В связи с этим во второй части настоящего исследования оценивали в плазме крови крыс такие показатели как общий белок, глюкозу, холестерин, креатинин, мочевину, α-амилазу, лактатдегидрогеназу, триглицериды, общий билирубин, аспартатаминотрансферазу (АСТ), аланинаминотрансферазу (АЛТ), связанный билирубин, щелочную фосфатазу, мочевую кислоту, креатинфосфокиназу (КФК), фосфор, липопротеиды низкой плотности и γ– глютамилтрансферазу. Изучение указанных показателей позволило охарактеризовать использованную в настоящем исследовании модель подострого гепатоза (дистрофии печени) у крыс, воспроизводимую 2-кратным внутрибрюшинным введением четыреххлористого углерода (суммарная доза 2000 мг/кг). Введение указанной дозы CCl4 в контрольной группе (активный контроль) приводило к гибели 50% крыс, что указывает на достаточную токсичность данного гепатотропного яда. У выживших животных, которые в течение 2 последующих недель наблюдались, отмечено существенное изменение в поведении, которое проявлялось снижением горизонтальной двигательной и исследовательской 114 активности, подавлением груминговых реакций, повышением эмоциональности. Кроме указанных изменений у таких животных отмечена высокая тревожность (анксиогенность) в приподнятом крестообразном лабиринте, снижение коммуникабельности (общительности), растормаживание системы «агрессия-защита» в тесте «чужак-резидент», снижение уровня активного и пассивного плавания, а также повышение иммобилизации в тесте на депрессивность Порсолта. В крови крыс, забитых на 16-й день опыта, регистрировали признаки поражения внутренних органов: печени, поджелудочной железы, почек. В частности, у крыс, получавший ССl4, в 2 раза возрастала активность аспартатаминотрансферазы, в 4 раза – лактатдегидрогеназы, умеренно повышалась активность α-амилазы, снижалась активность γ-глутамилтрансферазы, уровни общего и непрямого билирубина, мочевой кислоты, липопротеидов низкой плотности. При гистологическом исследовании печени у крыс был выявлен экспериментальный гепатоз (дистрофия печени), который характеризовался развитием гиалиново-капельной белковой дистрофии, мелкокапельной жировой дистрофии и слабо выраженной лимфоцитарной инфильтрацией портальных трактов. В группе крыс, получавших CCl4, на фоне нарушения балочного строения на большем протяжении в печени имелась паренхиматозная дистрофия в виде белковой гиалиново-капельной, которая поражала гепатоциты всех печеночных долек на 1/3 – 2/3 их протяженности балок, местами захватывала дольки очагово в виде мелкокапельной жировой дистрофии, локализовалась перипортально. В трактах наблюдалась слабо выраженная мононуклеарная инфильтрация, некроз гепатоцитов отсутствовал. В соответствие с балльной оценкой степени патогистологических изменений можно было констатировать 2-3-бальное поражение печени крыс данной группы – гистологические изменения соответствовали картине умеренной и выраженной токсической паренхиматозной дистрофии печени (гепатозу). 115 В головном мозге CCl4 не вызывал значимых отклонений в содержании и обмене медиаторов дофамина, норадреналина и серотонина в прилежащем ядре, стриатуме и фронтальной коре головного мозга, проявляя тенденцию к уменьшению содержания медиаторов. Аналогичные данные были получены А.А. Ротенко и соавторами (2006), которые также не отмечали изменений в содержании норадреналина, дофамина и серотонина, а также их метаболитов в головном мозге крыс при экспериментальном гепатозе, вызванном 5кратным введением внутрь четыреххлористого углерода (1 г/кг). Не регистрировали они и изменений в обмене исследованных медиаторов. Данное исследование предусматривало изучение влияния полипренолов в дозах 2,2-4,3-11,6 мг/кг в сравнении с адеметионином в дозе 300 мг/кг на модели гепатоза с развитием печеночной энцефалопатии после курсового введения препаратов в течение 14 дней на ряд поведенческих тестов. Эти тесты позволяют охарактеризовать психотропный спектр активности препаратов, биохимический спектр показателей крови, морфологическое исследование печени и обмен моноаминов в головном мозге. По разным тестам выявлено, что полипренолы в исследованных дозах нормализуют поведение крыс, нарушенное введением CCl4, улучшают показатели крови и печени у крыс с подострым гепатозом и оказывают психоактивирующее действие на головной мозг, в основе которого лежит изменение активности мезолимбической и нигростриатной дофаминергических систем. Интересно отметить, что в поведенческих тестах наибольшую активность полипренолы проявили в малой (2,2 мг/кг) и средней (4,3 мг/кг) дозах, тогда как гепатопротекторные их свойства были более выражены в самой большой из изученных доз пренолов (11,6 мг/кг). Последние были сопоставимы с действием классического гепатопротектора адеметионина (гептрала), который использовался в дозе 300 мг/кг (!). Не совсем понятно, но адеметионин, приготовленный на основе официнальных таблеток препарата и введенный внутрибрюшинно (аналог внутривенного 116 введения у человека) не только оказался неактивным, но и усугублял токсическое действие CCl4, что привело к 100%-ной гибели всех крыс данной группы на 7-й день эксперимента. Возвращаясь к особенностям действия полипренолов в разных дозах, следует отметить, что их поведенческие эффекты более выражены в дозах 2,2 и 4,3 мг/кг, то есть в малых и средних. Этому соответствует и действие полипренолов на метаболизм дофамина в головном мозге. Именно в этих дозах (но не в большой – 11,6 мг/кг) полипренолы активировали обмен дофамина в мезолимбической системе (в прилежащем ядре) и замедляли в нигростриатной (стриатуме), ответственной за двигательную активность. Действительно, это полностью перекликается с поведенческими данными (соответствует им), в которых показано снижение внешней двигательной активности под влиянием CCl4 и ее восстановление после курсового применения полипренолов. Следует напомнить, что в первом разделе наших исследованиях было оценено влияние полипренолов на дофаминзависимые формы поведения (см. главу 3.1). Так, в тесте «открытого поля» полипренолы (4,3-8,6-13,4 мг/кг) не проявляли дозозависимого эффекта, но растормаживали (активировали) горизонтальную двигательную активность при умеренном снижении всех других поведенческих активности и паттернов (прежде всего, исследовательской эмоциональности). Именно двигательная активность контролируется преимущественно дофаминергической системой, тогда как исследовательская активность (норковый рефлекс), груминг и эмоциональные проявления в значительной степени зависят от свойств серотонинергической и холинергической систем мозга. Сходные данные были получены и в данной части исследований, в которой полипренолы восстанавливали двигательную и исследовательскую подавленную в результате применения CCl4. активность, 117 В ротационном тесте, типичном для изучения дофаминзависимого поведения грызунов, полипренолы (13,4 мг/кг) в отличие от фенамина (непрямого дофаминомиметика) умеренно уменьшали число вращений в ротометре и не вызывали стереотипий. Это указывает на возможное участие нигростриатной части дофаминергической системы мозга (ее умеренное подавление в виде блокады постсинаптических рецепторов дофамина) в эффектах полипренолов. Данные были подтверждены в настоящем исследовании, в котором показано замедление обмена дофамина в стриатуме (главной структуре нигростриатной системы). Кроме того, полипренолы проявляли умеренную анксиогенную (тревожную) активность, которая наиболее стабильно воспроизводится при введении пренолов в дозе 13,4 мг/кг. Этот вид активности характерен для ряда фармакологических средств, оказывающих судорожное действие (судорожные агенты в субсудорожных дозах, психостимуляторы типа амфетамина, сиднокарба, кофеина). В настоящем исследовании полипренолы также не проявляли анксиолитической активности на фоне действия CCl4. В тесте «чужак-резидент» полипренолы также не оказывали дозозависимого эффекта, значимо подавляя индивидуальное поведение и общительность (коммуникативность) животных. В высокой дозе 13,4 мг/кг полипренолы влияли на все исследуемые показатели данного теста. Видимого антиагрессивного действия пренолы не выявили. В данном исследовании в отличие от предыдущего (глава 3.1), полипренолы восстанавливали общительность и индивидуальное поведение, сниженное CCl4, оптимизировали систему «агрессия-защита». В тесте Порсолта полипренолы оказывали депрессантный эффект на поведение интактных крыс (без патологии). Степень выраженности депрессантного эффекта пренолов была квалифицирована как умеренная. Во втором разделе настоящего исследования полипренолы существенно не влияли на активное и пассивное поведение крыс в плавательном тесте, но восстанавли- 118 вали иммобилизационный эффект, сниженный введением CCl4, что указывает на наличие у них антидепрессантных свойств. Таким образом, полипренолы обладают как гепатопротекторным, так и центральным действием. Гепатопротекторный эффект полипренолов (11,6 мг/кг) сопоставим с таковым адеметионина (300 мг/кг), что подтверждается как биохимическими показателями крови, так и морфологическим исследованием печени. В дозах 2,2 и 4,3 мг/кг полипренолы проявляют гепатопротекторный эффект, но менее выраженный. В то же время именно в этих дозах отмечены наиболее яркие эффекты пренолов на поведение и обмен моноаминов в головном мозге. Адеметионин (300 мг/кг) практически не оказывает центрального действия по поведенческим тестам и лишь незначительно ускоряет обмен серотонина во фронтальной коре мозга. Все вышеуказанное позволяет заключить, что полипренолы относятся к перспективным современным средствам с выраженным центральным психоактивирующим действием и способностью защищать печень и другие внутренние органы от действия токсикантов типа четыреххлористого углерода. Центральное действие полипренолов в значительной степени определяется их влиянием на активность дофаминергических систем головного мозга. 119 5. ЗАКЛЮЧЕНИЕ В первой части диссертационного исследования изучали влияние полипренолов (приготовленных на основе 95%-ного концентрата, или ропрена) на дофаминзависимые формы поведения крыс самцов Вистар (поведение в открытом поле, в приподнятом крестообразном лабиринте, в тесте «чужакрезидент», в ротометре, в тесте Порсолта, в тесте самостимуляции латерального гипоталамуса). Полипренолы использовали в дозах 4,3-8,6-13,4 мг/кг при их курсовом введении (5-7 дней). В «открытом поле» полипренолы не проявляли дозозависимого эффекта, растормаживая (активируя) горизонтальную двигательную активность при умеренном снижении всех других поведенческих паттернов (прежде всего, исследовательской активности и эмоциональности). В ротационном тесте полипренолы только в большой дозе (13,4 мг/кг) в отличие от фенамина (непрямого дофаминомиметика) умеренно уменьшали число вращений в ротометре и не вызывали стереотипий. Полипренолы проявляли умеренную анксиогенную (тревожную) активность, которая наиболее стабильно воспроизводится при введении препарата в дозе 13,4 мг/кг. В тесте «чужак-резидент» полипренолы значимо подавляли индивидуальное поведение и общительность (коммуникативность) животных. Видимого антиагрессивного действия пренолы не выявили. В тесте Порсолта полипренолы (4,3-8,6-13,4 мг/кг) оказывали депрессантный эффект на поведение. В дозе 13,4 мг/кг пренолы проявляли максимальное действие. Степень выраженности депрессантного эффекта полипренолов может рассматриваться как умеренная. Действие пренолов на реакцию самостимуляции в камере Скиннера зависело от продолжительности их введения. В первые дни введения полипренолы, как правило, умеренно подавляли реакцию самостимуляции, затем эффект сменяется на умеренный психоактивирующий, который по мере введения препарата снижался. Кроме того, полипренолы не проявляли функционального антагонизма с психостимулятором фенамином, и на фоне 120 их введения фенамин в полной мере развертывал свой психоактивирующий эффект. Сделано предположение, что полипренолы могут проявлять свойства парциального агониста (агониста-антагониста) рецепторов дофамина. Цель исследования второй части диссертации состояла в оценке гепатопротекторного и нейропротекторного действия полипренолов на модели подострого гепатоза, вызванного CCl4 у крыс. Подострый гепатоз (дистрофию печени) у крыс моделировали 2-кратным внутрибрюшинным введением CCl4 (2 инъекции по 1 г/кг с интервалом в 3 дня). Полипренолы (2,2-4,3-11,6 мг/кг) вводили внутрибрюшинно в течение 2 недель. После курса полипренолов оценивали поведение животных, биохимические показатели крови, морфологию печени и обмен моноаминов в головном мозге. Препаратом сравнения служил гептрал, или адеметионин (300 мг/кг), приготовленный на основе лиофилизата (сухого порошка) и таблеток. Введение гепатотропного яда приводило к гибели 50% крыс. У выживших животных отмечали существенное изменение в поведении по тестам «открытого поля», приподнятого крестообразного лабиринта, «чужакрезидент», Порсолта. В крови крыс, забитых на 16-й день опыта, регистрировали признаки поражения внутренних органов: печени, поджелудочной железы, почек. При гистологическом исследовании печени у крыс был выявлен экспериментальный гепатоз (дистрофия печени), который характеризовался развитием гиалиново-капельной белковой дистрофии, мелкокапельной жировой дистрофии и слабо выраженной лимфоцитарной инфильтрацией портальных трактов. В головном мозге CCl4 не вызывал значимых отклонений в обмене дофамина, норадреналина и серотонина в прилежащем ядре, стриатуме и фронтальной коре головного мозга. По разным тестам выявлено, что полипренолы в исследованных дозах (2,2-4,3-11,6 мг/кг) нормализуют поведение крыс, нарушенное введением CCl4, улучшают показатели крови и печени у крыс с подострым гепатозом и оказывают психоактивирующее действие на головной мозг, в основе которого лежит изменение активности мезо- 121 лимбической и нигростриарной дофаминергических систем (активация обмена дофамина в прилежащем ядре и замедление в стриатуме). В поведенческих тестах наибольшую активность полипренолы проявили в малой (2,2 мг/кг) и средней (4,3 мг/кг) дозах, тогда как гепатопротекторные свойства пренолов были выражены во всех исследованных дозах (2,2-4,3-11,6 мг/кг), но максимально в дозе 11,6 мг/кг. Последние были сопоставимы с действием лиофилизата адеметионина (300 мг/кг). Адеметионин лиофилизат практически не проявил центральных эффектов. Адеметионин на основе таблеток усугублял токсическое действие CCl4, что приводило к 100%-ной гибели всех экспериментальных животных к 7-му дню исследования. Следовательно, полипренолы обладают выраженным гепатопротекторным и нейропротекторным эффектом в широком диапазоне доз. 122 6. ВЫВОДЫ 1. Полипренолы в дозах 4,3-8,6-13,4 мг/кг при их курсовом введении (5-7 дней) активируют горизонтальную двигательную активность при умеренном снижении всех других поведенческих паттернов (прежде всего, исследовательской активности и эмоциональности), мало влияют на тревожность, подавляют индивидуальное поведение и общительность (коммуникативность) животных, не выявляют антиагрессивного действия и оказывают умеренный депрессантный эффект на поведение. 2. Действие полипренолов на реакцию самостимуляции в камере Скиннера зависит от продолжительности их введения. В первые дни введения они умеренно подавляют реакцию самостимуляции, затем эффект сменяется на умеренный психоактивирующий, который по мере введения препарата снижается. Кроме того, полипренолы не проявляют функционального антагонизма с психостимулятором фенамином, и на фоне их введения фенамин в полной мере развертывал свой психоактивирующий эффект. 3. Экспериментальный подострый гепатоз, вызванный CCl4, является удобной моделью для оценки как гепатопротекторных, так и нейропротекторных свойств, поскольку CCl4 поражает печень и вызывает экспериментальную энцефалопатию у крыс. 4. В поведенческих тестах ропрен в дозах 2,2-4,3-11,6 мг/кг оказывает нормализующее действие на двигательную и исследовательскую активность, а также эмоциональность крыс, нарушенную CCl4. Адеметионин 300 мг/кг такого действия не оказывает. 5. Полипренолы и адеметионин проявляют умеренное антиагрессивное действие, при этом полипренолы по эффективности превышают адеметионин, включая высокую степень защиты (защитного поведения), в то время как адеметионин подавляет этот показатель поведения. 123 6. Полипренолы в дозах 2,2-4,3-11,6 мг/кг нормализуют большинство показателей крови, нарушенных введением токсиканта CCl4. Адеметионин умеренно нарушает эвакуаторную функцию печени, о чем свидетельствует повышение в крови непрямого билирубина. 7. Гепатопротекторное действие полипренолов 2,2-4,3-11,6 мг/кг и адеметионина 300 мг/кг заключается в нормализации морфологического повреждения печени при экспериментальном гепатозе (белковой и жировой дистрофии), вызванном CCl4. Максимальный гепатопротекторный эффект регистрируется при введении полипренолов в дозе 11,6 мг/кг и адеметионина в дозе 300 мг/кг. 8. Полипренолы в дозах 2,2-4,3-11,6 мг/кг оказывают выраженный нейропротекторный эффект, в основе которого лежит их способность повышают активность мезолимбической (прилежащее ядро) и нигростриатной (стриатум) дофаминергических систем в модели подострого гепатоза и токсической энцефалопатии у крыс. Адеметионин таким действием не обладает. 124 7. НАУЧНО-ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ Полученные данные позволяют использовать полипренолсодержащие препараты (ропрен) не только в качестве гепатопротекторов, но и для защиты других внутренних органов (поджелудочная железа, миокард, почки) от действия токсикантов. В малых дозах (2,2-4,3 мг/кг) полипренолы обладают выраженным нейропротекторным действием, что не свойственно другим гепатопротекторам, например, адеметионину или препаратам расторопши пятнистой. Способность полипренолов нормализовать обмен моноаминов (дофамина и серотонина) в головном мозге может лежать в основе других их центральных эффектов, прежде всего, возможной антидепрессантной активности. 125 8. ЛИТЕРАТУРА 1. Андреева, Н.И. Методические указания по изучению антидепрессантной активности фармакологических веществ /Н.И Андреева // Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ / Под ред. В.П.Фисенко. М.: Ремедиум, − 2002. − С. 121-125. 2. Бурбелло, А.Т. Современные лекарственные средства: Клинико- фармакологический справочник практического врача. 4-е изд., перераб. и доп. /А.Т.Бурбелло, А.В.Шабров -М.: ОЛМА Медиа групп, −2007. − С.189. 3. Вальдман, А.В. Психо-фармакологические и медико-правовые аспекты токсикоманий. /А.В.Вальдман, Э. А.Бабаян, Э. Э. Звартау − М.: Медицина, − 1988. − 320 с. 4. Вартанян, Г.А Эмоции и поведение. /Г.А.Вартанян, Е. С. Петров −Л.: Наука, −1989. − 150 с. 5. Владимиров, Ю.А. Свободные радикалы в биологических системах /Ю.А. Владимиров // Сорос. образ. журн. − 2000. − Т.6, №12. − С.13-19. 6. Воеводин, Е.Е. Кортиколибериновые механизмы подкрепления и их модуляция нейропептидами и наркогенами: /Е.Е. Воеводин − Автореф. дис. … канд. мед. наук. СПб.: ВМедА, −2007. − 24 с. 7. Дробленков, А.В. Краткий микроскопический атлас ядерных и корковых центров мезокортиколимбической и некоторых других дофаминэргических систем головного мозга крысы /А.В. Дробленков − СПб.: СПбГПМА, −2006. −34 с. 8. Елисеева, А.П. Значение серотонинергической системы для формирования подкрепляющих механизмов мозга в онтогенезе у крыс /А.П. Елисеева.- Автореф. дисс. … канд. мед. наук. СПб.: ВМедА, − 2005. − 24 с. 126 9. Звартау, Э.Э. Методология изучения наркотоксикомании /Э.Э. Звартау // Итоги науки и техники. Сер. Наркология. М.: ВИНИТИ, −1988.- Т. 1. − С. 1-166. 10. Конорский, Ю. Интегративная деятельность мозга. /Ю.Конорский - М.: Мир. − 1970. − 420 с. 11. Лебедев, А.А. Эффекты дофаминергических средств на самостимуляцию латерального гипоталамуса и обмен дофамина в мозге крыс-изолянтов с разрушением вентральной области покрышки. /А.А.Лебедев, И. В.Лосева, П. Д.Шабанов // Журн. высш. нервн. деят. −1995. − Т. 45, Вып. 2. − С. 395-401. 12. Лебедев, А.А. Сопоставление реакции самостимуляции и условного предпочтения места при введении фенамина у крыс /А.А.Лебедев, П. Д.Шабанов // Журн. высш. нервн. деят. − 1992. − Т. 42, Вып. 4. − С. 692-698. 13. Любимов, А.В. Участие структур расширенной миндалины в подкрепляющем действии наркогенов / А.В. Любимов //Автореф. дис. … канд. мед. наук. СПб.: ВМедА, −2012. − 22 с. 14. Машковский, М.Д. Лекарственные средства: Пособие для врачей. 15-е изд. /М.Д.Машковский − М.: Новая волна, −2008. -С.526-528. 15. Менделевич, В.Д. Наркомания и наркология в России в зеркале общественного мнения и профессионального анализа. /В.Д.Менделевич Казань: Медицина, − 2006. − 262 с. 16. Менделевич, В.Д. Аддиктивное влечение. /В.Д.Менделевич, М.Л.Зобин М.: МЕДпресс-информ, − 2012. − 264 с. 17. Мещеров, Ш.К. Значение формирования дофаминергических систем мозга в онтогенезе для реализации эффектов психостимуляторов. / Ш.К. Мещеров - Автореф. дисс. … канд. мед. наук. СПб., − 2001. − 24 с. 127 18. Мещеров, Ш.К. Фармакологическая коррекция последствий социальной изоляции. / Ш.К. Мещеров − Автореф. дис. … д-ра мед. наук. СПб.,2004. − 48 с. 19. Михеев, В.В. Фармакологическая асимметрия мозга. /В.В.Михеев, П.Д.Шабанов − СПб.: Элби-СПб, − 2007. − 368 с. 20. Оковитый, С.В. Экспериментально-клиническая оценка применения препарата метаболического действия бемитила при хронической патологии печени /С.В. Оковитый // Мед. акад. журн. − 2002. − Т. 2. № 2. − С. 44-52. 21. Оковитый, С.В.. Гепатопротекторный эффект бемитила у больных с хроническими алкогольными поражениями печени /С.В.Оковитый, О.В.Иванова, П.Д. Шабанов // Наркология. − 2002. − № 3. − С. 19-23. 22. Отеллин, В.А. Медиаторные системы головного мозга: субстрат межнейронных связей, мишени фармакологических воздействий и объемы трансплантация /В.А. Отеллин // Актуальные вопросы биологии и медицины. Фундаментальные и прикладные проблемы / Под ред. Н.П. Бехтеревой. Л.: АМН СССР. −1990. − №2. −С. −74-85. 23. Пастушенков, Л.В. Фармакотерапия с основами фитотерапии. Часть II. /Л.В.Пастушенков, Е.Е.Лесиовская − СПб.: СПбХФИ, −1995. − 464 с. 24. Пастушенков, Л.В. Лекарственные растения: использование в народной медицине и быту. /Л.В.Пастушенков, А.Л.Пастушенков, В.Л. Пастушенков − Л.: Лениздат, −1990. − 184 с. 25. Петров, Е.С. Изучение нейробиологических основ сложных безусловных рефлексов в Физиологическом отделе им. И.П. Павлова. Итоги последних лет /Е.С. Петров // Физиол. журн. СССР. − 1990. − Т. 76, №12. − С. 1669-1680. 26. Подымова, С.Д. Болезни печени. /С.Д. Подымова − М.: Медицина, − 1998. − 256 с. 128 27. Пошивалов, В.П. Патологические последствия социальной изоляции у людей и животных: обзор литературы и собственные экспериментальные наблюдения. /В.П.Пошивалов - М., − 1977. − 34 с. Деп. в ВИНИТИ, № 2357-77. 42 с. 28. Пошивалов, В.П. Последствия зоосоциальной изоляции в зависимости от индивидуальных особенностей животных /В.П.Пошивалов // Журн. высш. нервн. деят. −1978. − Т. 28. − С. 438-455. 29. Пошивалов, В.П. Экспериментальная психофармакология агрессивного поведения. /В.П.Пошивалов - Л.: Наука, -1986.- 184 с. 30. Пошивалов, В.П. Этологический атлас для фармакологических исследований на лабораторных грызунах. /В.П.Пошивалов -Деп. в ВИНИТИ, № 3164-78. М., −1978. − 43 с. 31. Преображенская, Л.А. Эмоции в инструментальном поведении животных. /Л.А. Преображенская − М.: Наука, − 1991. 32. Раевский, К.С. Дофаминергические системы мозга: рецепторная гетерогенность, функциональная роль, фармакологическая регуляция. /К.С. Раевский, Т.Д. Сотникова, Р.Р. Гайнетдинов // Успехи физиол. наук. − 1996. − Т. 27. № 4. − С. 3-29. 33. Ротенко, А.А. Влияние хронического введения лекарственного препарата «гептрал» и аминокислоты метионина на содержание моноаминов и S-аденозилметионина в мозге крыс с гепатозом /А.А.Ротенко, С.Ф.Дугин, В.В.Крылин, З.С. Агаджанян // Совр. Наукоемкие технологии. −2006. − №4. − С.16-19. 34. Сапронов, Н.С. Гипоталамо-гипофизарно-надпочечниковая система и мозг. /Н.С. Сапронов − СПб.: Элби-СПб, −2005. − 512 с. 35. Свидерский, В.Л. // Анализ действия полипренольного препарата "Ропрен" и холинальфосцератного препарата "Глиатилин" на мембраносвязанные и "растворимые" формы холинэстераз и моноаминоксидазы мозга и сыворотки крови крыс при тетрахлорметановой модельной системе 129 печеночной энцефалопатии /В.Л. Свидерский., В.С.Султанов, В.И. Рощин. и др. // Докл. Акад. наук. − 2007. − T.412. № 3. − C.412-416. 36. Свидерский, В.Л. Сравнительное исследование действия полипренольного препарата "Ропрен" из хвойных растений на ключевые ферменты холинергического и моноаминергического типов нервной передачи / В.Л.Свидерский, А.Е Хованских., Е.В. Розенгарт и др. // Докл. Акад. наук. − 2006. − Т.408. № 3. − C.414-417. 37. Симонов, П.В. Мотивированный мозг. / П.В.Симонов − М.: Наука, − 1987. − 240 с. 38. Симонов, П.В. Эмоции, потребности, поведение. Избранные труды. Т.1. / П.В.Симонов − М: Наука, − 2004. 39. Соколов, С.Я. Справочник по лекарственным растениям. М.: Медицина, 1990. Соринсон С.Н. Вирусные гепатиты. /С.Я.Соколов, И.П. Замотаев − СПб.: Теза, − 1998. − 208 с. 40. Стрельцов, В.Ф. Значение гормональных механизмов в действии психостимуляторов на подкрепляющие системы мозга. / В.Ф. Стрельцов Автореф. дис. … канд. мед. наук. − СПб.: ВМедА, −2003. −23 с. 41. Стрельцов, В.Ф. Фармакология кортиколибериновых механизмов подкрепления и зависимости. / В.Ф.Стрельцов Автореф. дис. … д-ра мед. наук. − Смоленск: СГМА, −2009. − 46 с. 42. Тодоров, И.Н. Стресс, старение и их биохимическая коррекция. / И.Н.Тодоров, Г.И.Тодоров − М.: Наука, − 2003. − 479 с. 43. Угрюмов, М.В. Дифференцировка дофаминергических нейронов in situ, in vitro и в трансплантате /М.В.Угрюмов // Рос. Физиол. журн. им. И. М. Сеченова. −1998. −Т. 84. № 10. −С. 1019-1028. 44. Хананашвили, М.М. Патология высшей нервной деятельности. / М.М. Хананашвили − М.: Медицина, − 1983. − 287 с. 130 45. Цикунов, С.Г. Системные механизмы регуляции адаптивного поведения. // Теоретические и прикладные аспекты акмеологии. Вып.2. / С.Г. Цикунов / Под ред Ю.А. Шаронова. − СПб.: СПбПиА, − 2008. − С. 205-221. 46. Шабанов, П.Д. Влияние препарата ропрен на дофамин-зависимые формы поведения у крыс (заключительный отчет). /П.Д. Шабанов СПб.: ВМедА, −2008. −45 с. 47. Шабанов, П.Д. Маргали в комплексном лечении заболеваний печени: Методические рекомендации для врачей. /П.Д. Шабанов СПб.: ВМедА, 2002. −24 с. 48. Шабанов, П.Д. Наркология: Руководство для врачей. Изд. 2-е, перераб. и доп. /П.Д. Шабанов М.: Геотар-медиа, −2012. −824 с. 49. Шабанов, П.Д. Основы наркологии. /П.Д. Шабанов −СПб.: Лань, −2002. 560 с. 50. Шабанов, П.Д. Психофармакология. /П.Д. Шабанов −СПб.: Элби-СПб, 2008. 368 с. 51. Шабанов, П.Д. Психофармакология. /П.Д. Шабанов −СПб.: Элби-СПб, 2008. 416 с. 52. Шабанов, П.Д. Нарушения памяти и их коррекция. /П.Д.Шабанов, Ю.С. Бородкин −Л.: Наука, 1989. 150 с. 53. Шабанов, П.Д. Метаболические корректоры гипоксии. /П.Д.Шабанов, И.В. Зарубина, В.Е Новиков., В.Н. Цыган −СПб.: Н-Л, −2010. − 916 с. 54. Шабанов, П.Д. Биология алкоголизма. / П.Д. Шабанов, С.Ю. Калишевич −СПб: Лань, −1998. −372 с. 55. Шабанов, П.Д. Функциональное маркирование состояния социальной изолированности с помощью аналога меланостатина алаптида у крыс. /П.Д. Шабанов, А. А. Лебедев, А. Д. Ноздрачев // ДАН. −1999. −Т. 368, № 2. − С. 283-285. 56. Шабанов, П.Д. Блокада серотонинергических рецепторов мозга диэтиламидом лизергиновой кислоты препятствует облегчающему эффекту 131 фенамина на самостимуляцию крыс с разрушением медиальной префронтальной коры /П.Д. Шабанов, А.А. Лебедев // Журн. высш. нервн. деят. −1994. −Т. 44, Вып. 6. −С. 1124-1129. 57. Шабанов, П.Д. Дофаминергический и серотонинергический компоненты реакции самостимуляции латерального гипоталамуса крыс с разрушением медиальной префронтальной коры / П.Д. Шабанов, А.А. Лебедев // Физиол. журн. им. И.М.Сеченова. −1994. −Т. 80, № 1. −С. 19-25. 58. Шабанов, П.Д. Значение системы кортиколиберина и дофамина в миндалине для подкрепляющих эффектов опиатов и опиоидов на самостимуляцию латерального гипоталамуса у крыс /П.Д. Шабанов, А.А. Лебедев, А.В. Любимов, В.А. Корнилов // Эксперим. и клин. фармакол. −2011. −Т. 74, №7. −С. 3-8. 59. Шабанов, П.Д. Дофамин и подкрепляющие системы мозга. /П.Д. Шабанов, А.А. Лебедев, Ш.К. Мещеров −СПб.: Лань, −2002. −208 с. 60. Шабанов, П.Д. Зоосоциальное поведение млекопитающих. /П.Д. Шабанов, А.А. Лебедев, В.В. Русановский. − СПб.: Элби-СПб, −2006. − 160 с. 61. Шабанов, П.Д. Синдром социальной изоляции. /П.Д. Шабанов, Ш.К. Мещеров, А.А .Лебедев − СПб.: Элби-СПб, −2004. − 208 с. 62. Шабанов П.Д. Оценка анксиолитических и антидепрессантных эффектов полипренолов после витального психогенного стресса у крыс / П.Д.Шабанов, В.С.Султанов, В.А.Лебедев и др. // Инновации в современной фармакологии. Матер. IV съезда фармакологов России. – Казань; М.: Фолиум, 2012. – С.198-199. 63. Adair, W.L. Dolichol metabolism in rat liver. Determination of the subsellular distribution of dolichyl phosphate and its site and rate of de novo biosynthesis /W.L. Adair, R.K. Keller // J. Biol. Chem. −1982. −Vol.257. №15. −P.89908996. 64. Adams, D.B. Brain mechanisms of aggressive behavior: an updated review. /D.B. Adams // Neurosci. Biobehav. Rev. −2006− Vol. 30, № 3. − P. 304–18. 132 65. Afonso, V.M. Amphetamine pretreatment facilitates appetitive sexual behaviors in the female rat. /V.M. Afonso, D. Mueller, J. Stewart, J.G. Pfaus // Psychopharmacology. −2009 −Vol. 205, № 1. − P. 35–43. 66. Alcaro, A. Behavioral functions of the mesolimbic dopaminergic system: an affective neuroethological perspective. /A. Alcaro, R. Huber, J. Panksepp // Brain Res. Rev. − 2007 − Vol. 56, № 2. − P 283–321. 67. Amano, T. Role of amygdala subnuclei in fear expression and extinction learning. / T. Amano // Nihon. Yakurigaku Zasshi. − 2012. − Vol. 140, № 1. − P. 14-8. 68. Aragona, B.J.. Amphetamine reward in the monogamous prairie vole. /B.J. Aragona, J.M Detwiler., Z. Wang // Neurosci Lett. − 2007 − Vol. 418, −P. 190–194. 69. Aragona, B.J.. Opposing regulation of pair bond formation by cAMP signaling within the nucleus accumbens shell. /B.J.Aragona, Z Wang. // J. Neurosci. −2007.− Vol. 27, P. 13352–13356. 70. Avena, N.M.. Cholinergic modulation of food and drug satiety and withdrawal. / N.M. Avena, P.V. Rada // Physio.l Behav. − 2012 −Vol. 106, №3. − P. 332-336. 71. Balfour, D.J. The neuronal pathways mediating the behavioral and addictive properties of nicotine. /D.J. Balfour // Handb. Exp. Pharmacol. −2009 −Vol. 192, −P. 209–233. 72. Bardo, M.T. Conditioned place preference: what does it add to our preclinical understanding of drug reward? /M.T. Bardo, R.A. Bevins. // Psychopharmacology. −2000 −Vol. 153, № 1. − P. 31–43. 73. Bentinger, M. Polyisoprenoid epoxides stimulate the biosynthesis of Coenzyme Q and inhibit cholesterol synthesis /M. Bentinger, M. Teckle, K. Brismar et al. // J. Biol. Chem. −2008. −Vol.283. №21. −P.14645-14653. 133 74. Bergamini, E. Ageing and oxidative stress: a role for dolichol in the antioxidant machinery of cell membranes /E. Bergamini, R. Bizzarri, G. Cavallini et al. // J. Alzheimers Dis. −2004. V−ol.6. №2. − P.129-135. 75. Björklund, A. Classical transmitters in the CNS. /A. Björklund, T. Hökfelt Oxford-Amsterdam-New York: Elsevier, −1984. − 463 p. 76. Blum, K. The addictive brain: all roads lead to dopamine. /K Blum, A.L. Chen, J. Giordano, J. Borsten, T.J. Chen, M. Hauser, T. Simpatico, J. Femino, E.R. Braverman, D. Barh //J. Psychoactive Drugs. −2012 −Vol. 44, № 2. −P. 134-43 77. Buhaescu, I. Mevalonate pathway: A review of clinical and therapeutic implications /I. Buhaescu, H. Izzedine // Clin. Biochem. −2007. − Vol.40. −P.575584. 78. Burbassi, S. Stimulation of serotonin (2C) receptors influences cocaineseeking behavior in response to drug-associated stimuli in rats. /S. Burbassi, L. Cervo, // Psychopharmacology (Berl) − 2008. −№ 1. −P. 15–27. 79. Burda, P. The dolichol pathway of N-linked glycosylation /P. Burda, M. Aebi // Biochim. Biophys. Acta. −1999. − Vol.1426. −P.239-257. 80. Cagiano, R. Effects on rat sexual behaviour of acute MDMA (ecstasy) alone or in combination with loud music./ R. Cagiano, I. Bera, R. Sabatini, P. Flace, D. Vermesan, H. Vermesan, S.I. Dragulescu, L Bottalico., L. Santacroce // Eur. Rev. Med. Pharmacol. Sci. −2008 −Vol. 12, № 5. P. 285–92. 81. Carboni, E. Dopamine and drug addiction: the nucleus accumbens shell connection. /E. Carboni, V. Valentini, D. Lecca // Neuropharmacology. − 2004 −Vol. 47, Suppl. 1. −P. 227–241. 82. Carey, R. J. Unilateral 6-hydroxydopamine lesion of dopamine neurons produced bilateral sels-stimulation deficits /R. J. Carey // Behav. Brain Res. −1982. −Vol. 6, № 2. −P. 101-114. 134 83. Carlezon, W.A. Biological substrates of reward and aversion: a nucleus accumbens activity hypothesis. /W.A. Carlezon., M.J. Thomas //Neuropharmacology. −2009 −Vol. 56, Suppl. 1. −P. 122–132. 84. Carter, C.J. The effect of 5,7-dihydroxy tryptamine lesions of extrapiramidal and mesolimbic sites on spontaneus motor behavior and amphetamine stereotypy /C.J. Carter, C.J. Pycock // Arch.Pharmacol. −1979. −Vol. 308. −P. 5154. 85. Chojnacki, T. The biological role of dolichol /T. Chojnacki, G. Dallner // Biochem.J. −1988. −Vol. 251, − P. 1–9. 86. Clark, D. D1 dopamine receptor – the search for a function /D. Clark, F. J. White // Synapse. −1987. − Vol. 1. −P. 347-388. 87. Cole, S.L., Vassar R. Isoprenoids and Alzheimer's disease: a complex relationship. /S.L. Cole, R.Vassar //Neurobiol. Dis. −2006. −№ 22. −P. 209–222. 88. Comai, S. The psychopharmacology of aggressive behavior: a translational approach: part 1: neurobiology. S. Comai, M. Tau, G. Gobbi // J. Clin. Psychopharmacol. −2012 −Vol. 32, № 1. −P. 83-94. 89. Cools, R. Dopaminergic control of the striatum for high-level cognition. /R. Cools // Curr. Opin. Neurobiol. −2011 −V.21 № 3: −P.402-407. 90. Curtis, J.T. Ventral tegmental area involvement in pair bonding in male prairie voles. /J.T. Curtis, Z. Wang // Physiol Behav. −2005 −Vol. 86, № 3. −P. 338–346. 91. Dalley, J.W. Dopamine, serotonin and impulsivity. /J.W. Dalley, J.P. Roiser // Neuroscience. −2012 −Vol. 2, № 215. −P. 42-58. 92. De Manzano, Ö.. Individual differences in the proneness to have flow experiences are linked to dopamine D2-receptor availability in the dorsal striatum. / Ö.de Manzano, S. Cervenka, A. Jucaite, O. Hellenäs, L. Farde, F Ullén // Neuroimage. −2013 −Vol. 67, −P. 1-6. 93. DeLeon, K.R. Adolescent cocaine exposure and offensive aggression: involvement of serotonin neural signaling and innervation in male Syrian ham- 135 sters. /K.R. DeLeon, J.M. Grimes, D.F. Connor, R.H. Melloni. // Behav. Brain. Res. −2005 −Vol. 133, № 2. −P. 211–220. 94. Denecke, J. Hypoglycosylation due to dolichol metabosism defects / J.Denecke, C.Kranz // Biochim. Biophys. Acta. −2009. −Vol. 1792. −P.888895. 95. Eckert, G.P. Regulation of the brain isoprenoids farnesyl- and geranylgeranylpyrophosphate is altered in male Alzheimer patients. /G.P.Eckert, G.P.Hooff, D.M. Strandjord, U. Igbavboa, D.A. Volmer, W.E. Muller., W.G. Wood // Neurobiol Dis. −2009 −Vol. 35, −P. 251–257. 96. Einav, S. Phenylation inhibitors: a novel class of antiviral agents /S. Einav, J.S. Glenn // J. Antimicrob. Ther. −2003. −Vol. 52. −P.883-886. 97. Elmberger, P.G. In vivo and in vitro synthesis of dolichol and other main mevalonate products in various organs of the rat /P.G Elmberger., A Kalen., E.L Appelkvist., G. Dallner // Eur. J. Biochem. −1987. −V.168. −P.1-11. 98. Espenshade, P.J. Regulation of sterol synthesis in eukaryotes /P.J. Espenshade, A.L. Hughes // Annu. Rev. Genet. −2007. −Vol. 41, −P. 401–427. 99. Farooqui, T. Aging: an important factor for the pathogenesis of neurodegenerative diseases. /T. Farooqui, A.A. Farooqui // Mech Ageing Dev. −2009 −Vol. 130, −P. 203–215. 100. Febo, M. Development of cocaine sensitization before pregnancy affects subsequent maternal retrieval of pups and prefrontal cortical activity during nursing. /M. Febo, C.F Ferris // Neuroscience. −2007 −Vol. 148, № 2. −P. 400– 412. 101. Francis, D.D. Frequency of maternal licking and grooming correlates negatively with vulnerability to cocaine and alcohol use in rats. /D.D Francis., M.J. Kuhar // Pharmacol. Biochem. Behav. −2007 −Vol. 90, № 3. −P 497– 500. 136 102. Glimcher, P.W. Understanding dopamine and reinforcement learning: the dopamine reward prediction error hypothesis. /P.W. Glimcher // Proc. Natl. Acad. Sci. USA. −2011 − V. 13, № 108. Suppl. 3. −P. 15647-15654. 103. Göderqvist, S.. Dopamine, working memory, and training induced plasticity: implications for developmental research. /S. Göderqvist, S. Bergman Nutley, M. Peyrard-Janvid., H. Matsson, K. Humphreys, J. Kere, T. Klingberg // Dev. Psychol. −2012 −V.48, № 3 −P. 836-843. 104. Gorwood, P. Simonneau M. Genetics of dopamine receptors and drug addiction. /P. Gorwood, Y. Le Strat, N. Ramoz, C. Dubertret, J.M. Moalic, // Hum. Genet. −2012 −Vol. 131, № 6. −P. 803-822. 105. Grigor’eva, N.Ya. Physiological activity of polyisoprenoids (review of patent data) /N.Ya. Grigor’eva, A.M. Moiseenkov // Pharm. Chem. Journ. −1989. − Vol.23. −P.103-113. 106. Guarraci, F.A. Intracranial infusions of amphetamine into the medial preoptic area but not the nucleus accumbens affect paced mating behavior in female rats. /F.A. Guarraci, R.J. Frohardt, D Hines., E. Navaira, J. Smith, L. Wampler // Pharmacol. Biochem. Behav. −2008 −Vol. 89, № 3. −P. 253–62. 107. Gurkovskaya, O. V. Reinforcing properties of dexamethasone /O. V. Gurkovskaya, A. A. Lebedev, P. D. Shabanov // Psychoneuroendocrinology. −1997. −Vol. 22, Suppl. 2. −P. S205. 108. Hemming, F.W. Glycosyl phosphopolyprenols /F.W. Hemming // Glycolipids. New comprehensive biochemistry. Vol. 10 / Ed. By H.Wiegandt. Amsterdam: Elsevier, −1985. − P.261-305. 109. Holstein, S.A. Isoprenoids: re−markable diversity of form and function./ S.A. Holstein., R.J. Hohl //Lipids. −2004. − Vol. 39, −P.293–309. 110. Holstein, S.A. Differential activities of thalidomide and isoprenoid biosynthetic pathway inhibitors in multiple myeloma cells. /S.A. Holstein, H. Tong, R.J. Hohl // Leuk Res. −2010. Vol. 34, −P. 344–351. 137 111. Hooff, G.P. Modulation of cholesterol, farnesylpyrophosphate, and geranylgeranylpyrophosphate in neuroblastoma SH-SY5Y-APP695 cells: Impact on amyloid beta-protein production /G.P.Hooff, I .Peters, W.G. Wood et al. // Mol. Neurobiol. 2−v010. − Vol.41. P.341-350. 112. Hooff, G.P. Isoprenoid quantitation in human brain tissue: a validated HPLCfluorescence detection method for endogenous farnesyl- (FPP) and geranylgeranylpyrophosphate (GGPP) /G.P. Hooff, D.A. Volmer, W.G. Wood, W.E. Muller, G.P. Eckert // Anal Bioanal. Chem. −2008 −Vol. 392, −P. 673–680. 113. Hooff, G.P. Isoprenoids, small GTPases and Alzheimer’s disease /G.P. Hooff, W.G. Wood, W.E. Muller, G.P. Eckert // Biochim. Biophys. Acta. −2010. −Vol.1801. −P.896-905. 114. Jones, P.H. Comparison of the efficacy and safety of rosuvastin versus atorvastatin, simvastatin, and pravastatin across doses (STELLAR trial) /P.H. Jones, M.H. Davidson, E.A. Stein et al. // Amer. J. Cardiol. −2003. −Vol. 92. −P.152-160. 115. Kebabian, J. W. Multiple receptors for dopamine /J. W. Kebabian, D−. B. Calne // Nature. −1979. −Vol. 277. −P. 93-96. 116. Kirby, J. Biosynthesis of plant isoprenoids: perspectives for microbial engineering /J. Kirby, J.D. Keasling // Ann. Rev. Plant. Biol. −2009. −Vol.60. −P.335-355. 117. Klein-Marcuschamer, D. Engineering microbial cell factories for biosynthesis of isoprenoid molecules: beyond lycopene /D. Klein-Marcuschamer, P.K. Ajikumar, G. Stephanopoulos // Trends Biotechnol. −2007. − Vol.25. №9. −P.417-424. 118. König, K.P. A stereotaxic atlas of the forebrain and lower parts of the brain stem. /K.P. König, A.A. Klippel– Baltimore, 1963. – 214 р. 119. Koob, G.F. Reward, addiction, withdrawal to nicotine. /G.F .Koob // Annu. Rev. Neurosci. −2011 −Vol. 34, −P. 105-130. 138 120. Kurup, R.K. Endogenous hypodigoxinemia-related immune deficiency syndrome /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1287-1303. 121. Kurup, R.K. Hypothalamic digoxin and hemispheric chemical dominance: relation to alcoholic addiction, alcoholic cirrhosis, and acquired hepatocerebral degeneration /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1105-1125. 122. Kurup, R.K. Hypothalamic digoxin and hemispheric chemical dominance – relation to the pathogenesis of senile osteoporosis, degenerative osteoarthritis, and spondylosis /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1341-1359. 123. Kurup, R.K. Hypothalamic digoxin, cerebral chemical dominance, and regulation of gastrointestinal/hepatic function /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. − 2003. −Vol. 113. −P.1175-1195. 124. Kurup, R. Hypothalamic digoxin, hemispheric chemical dominance, and mesenteric artery occlusion /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003−. Vol. 113. −P. 1741-1760. 125. Kurup, R.K. Hypothalamic digoxin, hemispheric chemical dominance, and oncogenesis: evidence from multiple myeloma /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. − Vol. 113. −P. 1719-1740. 126. Kurup, R.K. Hypothalamic digoxin, hemispheric chemical dominance, and interstitial lung disease /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1427-1443. 127. Kurup, R.K. Hypothalamic digoxin, hemispheric chemical dominance, and chronic bronchitis emphysema /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1241-1258. 128. Kurup, R.K. Hypothalamic digoxin, hemispheric chemical dominance, and inflammatory bowel disease /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1221-1240. 139 129. Kurup, R.K. Hypothalamic digoxin, hemispheric chemical dominance, and Alzheimer's disease /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P.1361-1381. 130. Kurup, R.K. Hypothalamic digoxin-mediated model for Parkinson's disease /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 515-536. 131. Kurup, R.K.Isoprenoid pathway dysfunction in human male infertility /R.K. Kurup, P.A. Kurup // Arch. Androl. −2003. −Vol. 49. −P. 117-127. 132. Kurup, R.K. Schizoid neurochemical pathology-induced membrane Na(+)-K+ ATPase inhibition in relation to neurological disorders /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1705-1717. 133. Kurup, R.K. Hypothalamic digoxin, hemispheric chemical dominance, and peptic ulcer disease /R.K. Kurup, P.A. Kurup // Int. J. Neurosci. −2003. −Vol. 113. −P. 1395-1412. 134. Lebedev, A. A.. Dopaminergic mode of action for melanostatine analogue in animal model of social isolation /A. A. Lebedev, G. N. Panchenko, P. D. Shabanov // Neuroendocrinology Letts. −1993. −Vol. 15, № 4. −P. 320. 135. Le Moal, M. Mesocorticolimbic dopaminergic network: functional and regulatory roles /M .Le Moal , H. Simon // Physiol. Rev. −1991. −Vol. 71. − P. 155-232. 136. Lieu, C.A. The interhemispheric connections of the striatum: Implications for Parkinson's disease and drug-induced dyskinesias. /C.A. Lieu, T. Subramanian // Brain. Res. Bull. −2012 −Vol. 87, № 1. −P. 1-9. 137. Mameli, M. Synaptic plasticity and addiction: learning mechanisms gone awry. /M. Mameli, C. Lüscher // Neuropharmacology. −2011 −V. 61, №.7. −P. 1052-1059. 138. McBride, W. J. Localization of brain reinforcement mechanisms: intracranial self-administration and intracranial place-conditioning studies /W. J. McBride, J. M. Murphy, S. Ikemoto // Behav. Brain Res. −1999. − Vol. 101. −P. 129-152. 140 139. McTaggart, S. J. Isoprenylated proteins. /S. J. McTaggart // Cell. Mol. Life Sci. −2006. −Vol. 63, −P. 255–267. 140. Missale, C. Dopamine receptors: from structure to function /C. Missale, S. R. Nash, S. W. Robinson et al. // Physiol. Rev. −1998. −Vol. 78. −P. 189-225. 141. Mo, H.. Studies of the isoprenoid-mediated inhibition of mevalonate synthesis applied cancer chemotherapy and chemoprevention /H. Mo, C.E. Elson // Exp. Biol. Med. −2004. −Vol.229. −P.567-585. 142. Morava, E. A novel cerebello-ocular syndrome with abnormal glycosylation due to abnormalities in dolichol metabolism /E. Morava, R.A. Wevers, V. Cantagrel et al. // Brain. −2010. −Vol.133. −P.3210-3220. 143. Morikawa, H. Dynamic regulation of midbrain dopamine neuron activity: intrinsic, synaptic, and plasticity mechanisms. /H. Morikawa, C.A. Paladini // Neuroscience. −2011 −V. 15, № 198. −P. 95-111. 144. Nathaniel, T.I. Drug-seeking behavior in an invertebrate system: evidence of morphine-induce reward, extinction an reinstatementin crayfish. /T.I.Nathaniel, J. Panksepp, R.Huber // Behav. Brain Res. −2009 −№ 1. −P. 331–338. 145. Panchenko, G. N. Comparison of the effects of dopamine agonists on selfstimulation of the hypothalamus with lesioning of mesolimbic brain structures in rats reared in conditions of social isolation /G. N.Panchenko, A. A. Lebedev, P. D. Shabanov // Neurosci. Behav. Physiol. −1998. −Vol. 28, № 2. −P. 130-135. 146. Pare, D. Amygdala microcircuits mediating fear expression and extinction. D. Pare, S. Duvarci // Curr. Opin. Neurobiol. −2012. − Vol. 22, № 4. −P. 717723. 147. Parentini, I. Accumulation of dolichol in older tissues satisfies the proposed criteria to be qualified a biomarker of aging. /I. Parentini, G. Cavallini, A Donati., Z Gori., E. Bergamini // J.Gerontol.A Biol.Sci.Med.Sci. −2005 −Vol. 60, −P.39–43. 141 148. Parkes, S.L. The basolateral amygdala is critical for the acquisition and extinction of associations between a neutral stimulus and a learned danger signal but not between two neutral stimuli. /S.L. Parkes, R.F. Westbrook // J. Neurosci. −2010. −Vol. 30, № 38. −P. 12608-12618. 149. Petrov, E.S. Dopamine and reinforcing system of the brain / E.S. Petrov, A.A. Lebedev // Neurosci. Behav. Physiol. −1997. −Vol. 27, № 3. −P. 309311. 150. Phillips, A. G. Neuroanatomical bases of intracranial self-stimulation: untangling the gordian knot /A. G. Phillips, H. C. Fibiger // Neuropharmacological Bases of Reward / Eds. Liebman J. and Cooper S.J. New York: Oxford Univ. Press, −1989. −P. 66-105. 151. Porsolt, R.D. Behavioural despair in rats: a new model sensitive to antidepressant treatments /R.D. Porsolt., G. Anton, N. Blavet // Eur. J. Pharmacol. −1978. −V.47. −P. 379-391. 152. Pullarkat, R.K. Accumulation of dolichols in brains of elderly /R.K. Pullarkat, H. Reha // J. Biol. Chem. −1982. −Vol. 257. № 11. −P. 5991-5993. 153. Sagara, H. Evaluation of motivational effects induced by intracranial selfstimulation behavior. /Sagara H., Sendo T., Gomita Y. // Acta Med Okayama. −2010 −V. 64, № 5. −P. 267-275. 154. Sakakihira, Y. Dolichol in Human brain: Regional and developmental aspects /Y. Sakakihira, J.J. Volpe // J. Neurochemistry. −1985. −V.44. −P.1535-1540. 155. Salamone, J.D. The mysterious motivational functions of mesolimbic dopamine. /J.D Salamone, M. Correa //Neuron. −2012. −Vol. 76, № 3− P. 470485. 156. Scherer, M.G. Brain dolichyl pyrophosphate: Solubilization, characterization, and differentiazion from dolichyl monophosphate phosphatase activity /M.G. Scherer, C.J. Waechter // J. Biol. Chem. −1984. −V.259. №23. −P.1458014585. 142 157. Tritsch, N.X. Dopaminergic modulation of synaptic transmission in cortex and striatum. /N.X. Tritsch, B.L. Sabatini // Neuron. −2012 −Vol. 76, № 1. −P. 33-50. 158. Van den Oever, M.C. The synaptic pathology of drug addiction. /M.C. Van den Oever, S. Spijker, A.B. Smit // Adv. Exp. Med. Biol. −2012 −Vol. 119, № 4. −P. 303-812.