70
advertisement

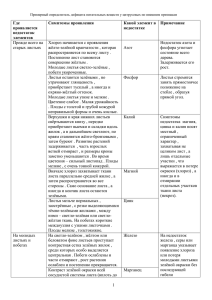
С. В. Мацкова, В. П. Дедков число видов 50 40 30 20 10 0 70 42 34 4 естественная флора апофитная флора адвентивная флора Рис. Соотношение флор в сквере на Московском проспекте Фактором, обеспечивающим возможность развития на нарушенных местах синантропных видов — как аборигенных, так и адвентивных, — является нарушение целостности растительного покрова. При этом характер зарастания зависит не только от особенностей экотопа, но и от флористического потенциала данного района, то есть наличия в окружающей флоре видов, способных заселять нарушенные участки [2]. Список литературы 1. Инструкция о порядке государственного учета объектов растительного мира, расположенных на землях населенных пунктов, и обращения с ними: Постановление Министерства жилищно-коммунального хозяйства Республики Беларусь. № 40, от 29 декабря 2004 г. 2. Антипина Г. С. Урбанофлора Карелии: Монография. Петрозаводск, 2002. Об авторах С. В. Мацкова — ассист., РГУ им. И. Канта centaurea@inbox.ru. В. П. Дедков — д-р биол. наук, проф., РГУ им. И. Канта dedkov@albertina.ru УДК 581.1+591.1+ 581.4 Е. А. Коршунов ФАКТОРЫ ФОРМИРОВАНИЯ ПРОВОДЯЩЕЙ СИСТЕМЫ ВИДОВ СЕМЕЙСТВА БОБОВЫХ В статье предлагается объяснение механизма жилкования. Описываются дифференциация клеток, формирование жилок и некоторые надгенные факторы, влияющие на этот процесс. The article focuses on a venation mechanism. It also presents cells’ differentiation and some genetic factors influencing this process. Установлено, что пространственная организация структурных элементов мезофила листа (форма, количество и размер клеток и хлоропластов) имеет важное значение в формировании адаптивных реакций вида, Вестник РГУ им. И. Канта. 2008. Вып. 1. Естественные науки. С. 70—76. 70 Факторы формирования проводящей системы видов семейства бобовых 71 поскольку определяет такие жизненные функции растительного организма, как фотосинтез, транспирацию и газообмен [1]. Структура жилкования листа предопределена его видовой принадлежностью и способна выступать как критерий идентификации. Давно существует мнение, что образование определенных структур, в частности листьев, обеспечивает специфическая генетическая программа [2] или каскадное включение группы программ [3]. К настоящему времени выявлен ряд гомеозисных генов, участвующих в процессах морфогенеза [4]. В то же время даже у листьев, взятых для сравнения с одного растения и с одного яруса, расположение проводящих элементов отличается. На первый взгляд это кажется непонятным, поскольку «похожесть» «предопределена» генетически, однако этот вопрос возможно объяснить с физиологической точки зрения. У большинства двудольных проводящие пучки образуют разветвленную систему, в которой от более крупных жилок ответвляются более мелкие. Такое жилкование называют сетчатым. Часто самая крупная (средняя) жилка тянется вдоль оси листа и вместе с прилегающей основной тканью выступает над его нижней поверхностью в виде гребня. В отличие от этого у большинства однодольных множество жилок примерно равного размера проходит вдоль листа параллельно друг другу. Такое жилкование называется параллельным. При этом продольные жилки связаны между собой более мелкими, что образует сложную сеть [5]. Поскольку более заметные различия в жилковании отдельных листьев прослеживаются на двудольных, в данной статье их развитию будет уделено основное внимание. Проводящие пучки, более или менее полностью погруженные в мезофилл, называют мелкими жилками, а сопровождающиеся выступами на нижней стороне листа — крупными жилками. Главную роль в отводе ассимилятов из клеток мезофилла играют первые. С увеличением размеров жилок их связь с мезофиллом ослабевает, и они все больше окружаются нефотосинтезирующими тканями гребней. Следовательно, функция сбора ассимилятов постепенно вытесняется главным образом транспортной [5]. Участки мезофила, ограниченного самыми мелкими жилками (жилками последних порядков), называются ареолами [6]. Установлено, что число ареол, приходящихся на 1 мм2, не зависит от площади листа [7—9], однако имеются данные, что деятельность жилок связана с относительно определенным количеством клеток, которые жилки способны «обслуживать» [10; 11]. Жилки содержат ксилему и флоэму, которые имеют, как правило, первичное происхождение (средние, а иногда и другие крупные жилки у некоторых двудольных способны к вторичному росту). Окончания жилок у двудольных часто содержат только трахеальные элементы, хотя до их концов могут доходить как ксилемные, так и флоэмные элементы. Обычно ксилема находится на верхней стороне листа, флоэма — на нижней [5]. Нанесение сахара и ауксина в агаре на поверхность каллюса выявило важное значение сахара для дифференциации флоэмы. Меняя концентрацию сахара, можно изменить соотношение ксилемы и флоэмы; низкая концентрация благоприятна для дифференциации ксилемы, а высокая — для дифференциации флоэмы. Средняя концентрация, которая, вероятно, преобладает в нормально растущих растениях, способствует в одинаковой степени дифференциации обеих тканей, обычно с камбием между ними, если в качестве материала используются двудольные [12]. 71 Е. А. Коршунов 72 Вначале листовой зачаток состоит только из меристематических клеток, но очень скоро меристематическими остаются только краевые клетки, причем верхушка листа прекращает расти раньше других частей. Меристематические клетки листа удлиняются в двух направлениях, образуя листовую пластинку. Однако в листе происходит и несколько поперечных делений, что ведет к увеличению его толщины [13]. По мере удлинения клиновидного листового примордия прокамбиальный тяж все дальше проникает в него, образуя систему крупных жилок, отходящих от главной жилки. У верхушки листа закладываются более мелкие жилки, которые развиваются по направлению к его основанию без перерывов, переходя в более крупные. Таким образом, верхушка листа первой приобретает полную систему жилкования, что отражает общую последовательность развития этого органа «сверху вниз». Эта схема отражает основное распределение проводящих пучков. Появление различий определяется тем, что в процессе онтогенеза происходит не только нарастание, надстройка новых частей, но и их изменение. Каждый последующий орган не является точным повторением ранее возникшего, так как новые органы закладываются на качественно иных участках растения и в новых условиях внешней среды. Эта особенность онтогенеза высших растений находит отражение в периодичности, цикличности как морфологических, так и физиологических особенностей [5]. Радиальные (паренхимные) периферии транспортных систем флоэмы и ксилемы могут формироваться сходно — как эндоплазматические сети, развивающиеся во встречных направлениях [14]. Эндоплазматические сети, в начале функционирующие как внутриклеточные, в дальнейшем посредством плазмодесм формируют межклеточные сети. Зависимость формирования эндоплазматической сети от фотосинтеза и транспирации ассимилятов наблюдается в клетках мезофила и флоэмы развивающихся листьев [15]. Трахеальные элементы включаются в сеть апопласта, а ситовидные — в сеть симпласта. Таким образом, не только радиальная периферия, но и осевые элементы обеих тканей, ксилемы и флоэмы, возникают как производные эндоплазматической сети [16]. Морфология растения зависит от формы его клеток, в свою очередь определяющейся направлением растяжения. Растяжение клеток регулируется свойствами клеточной стенки, в частности ориентацией целлюлозных микрофибрилл. В продольных стенках удлиняющейся растительной клетки микрофибриллы целлюлозы, отложенные последними, обычно перпендикулярны оси удлинения. Хотя расположение отложенных ранее микрофибрилл может быть иным, именно от ориентации волокон в самой внутренней ламелле клеточной стенки зависит направление роста клетки. До сих пор точно неизвестно, каким образом определяется тонкая структура всех компонентов клеточной стенки, однако направление отложения внутренних (самых молодых, новых) микрофибрилл целлюлозы коррелирует с ориентацией кортикальных микротрубочек. Возможно, в этих процессах принимают участие актиновые компоненты кортекса клетки [17]. Строгий контроль за плоскостями деления клеток возможен лишь в том случае, если ткань растения и составляющие ее клетки обладают 72 Факторы формирования проводящей системы видов семейства бобовых 73 определенной структурой полярности, которая впоследствии может быть либо закреплена, либо видоизменена. Хотя структурная основа подобной полярности у высших растений еще не выявлена, у низших растений можно найти хорошо изученные примеры. В полярности клеток могут участвовать асимметрично расположенные ионные каналы и белки-носители, присутствующие в плазматической мембране, которые могут генерировать внутриклеточные потоки ионов. Ключом к пониманию механизма полимеризации, например зиготы бурой водоросли Fucus, находящейся на ранней стадии развития, служит небольшой электрический ток, проходящий через эту клетку. Этот ток генерируется и поддерживается отчасти благодаря пассивному переносу ионов Ca2+ из другого участка, что, вероятно, отражает асимметричное распределение кальциевых каналов и насосов в плазматической мембране. Такой ток способен перемещать путем электрофореза внутриклеточные компоненты, обладающие высоким зарядом; кроме того, имеются данные, что одним из результатов подобной поляризации является накопление секреторных пузырьков у базального полюса, за которым следуют локализованные отложения специфических сульфатированных полисахаридов в этом участке клеточной стенки [18]. Полярность у растений находит отражение в закономерном снижении или возрастании по длине побега и корня концентрации разных веществ — физико-химические градиенты, в интенсивности физиологических процессов (дыхание, фотосинтез, транспирация и др.) — физиологические градиенты. Кроме того, полярность обуславливается анатомическим строением (размер, форма, строение клеток и их органоидов) — анатомическими градиентами. Все градиенты в растительном организме взаимосвязаны и взаимоподчинены [19]. Не обладая полярностью, зигота Fucus становится поляризованной в ответ на внешние стимулы, например градиенты освещения или силы тяжести. Вероятно, эти факторы приводят к изначальной неравномерности распределения ионов кальция в яйцеклетке. По-видимому даже небольшие сдвиги в гомогенном распределении ионов кальция вызывают значительные последствия, поскольку быстро усиливаются благодаря механизму с обратной связью. Элементы цитоскелета участвуют в соединении кальциевых каналов с фибриллами клеточной стенки путем образования на месте будущего базального полюса трансмембранных мостиков [18]. Способность реагировать на внешние факторы является общим свойством цитоскелета любой клетки, в частности его актиновых и тубулиновых структур. Некоторые пространственно-ориентированные сигналы, по-видимому, способны оказывать прямое поляризующее воздействие на структуру и свойства цитоскелета. К таким факторам относятся гравитация, электрические поля, изменение осмотических и ионных градиентов, механические воздействия, в том числе межклеточные контакты. Быстрое искажение полярности существующего в конкретный момент времени цитоскелета в дальнейшем сопровождается более медленной его перестройкой и закреплением новой морфофизиологической оси полярности клетки. С другой стороны, поляризующие сигналы могут влиять на цитоскелетный комплекс через изменение состояния рецепторов и ионных каналов, через локальные изменения концентрации вторичных посредников, и в первую очередь ци- 73 Е. А. Коршунов 74 топлазматического кальция. Возможно, подобным образом на структуру цитоскелета могут влиять фитогормоны [17]. Только благодаря поляризации гормональных продуктов внутри органа происходит корреляционное взаимодействие между отдельными его частями, выражающееся в неравномерном распределении процессов клеточного растяжения [19]. Такие ауксины, как ИУК и НУК, — обязательные участники координации процессов морфогенеза, оказывающие влияние на деление, растяжение и дифференциацию клеток. Эти гормоны необходимы прежде всего для инициации репликации ДНК. Ауксины стимулируют секрецию ионов водорода (протонов). Это приводит к понижению внеклеточного рН, а тем самым и к разрыхлению клеточной стенки; возможно, в этом процессе участвует фермент с низким оптимумом рН. Кроме того, нужно, чтобы внутри клетки поддерживался высокий осмотический потенциал и было достаточно воды, которая входила бы в клетку и создавала тургорное давление [20]. К фитогормонам, синтезированным в других органах и передвигающимся по проводящей системе, чувствительны лишь некоторые клетки — «клетки мишени», имеющие специфические гормоносвязующие белки-рецепторы, что подтвердилось обнаруженными в последнее десятилетие рецепторами, локализованными на плазматических мембранах и внутри клеток, предположительно на стенках эндоплазматического ретикулума [3]. Достаточно давно исследователи задаются вопросом, является ли изменеенное расположение компонентов цитоскелета причиной или следствием гормоно-индуцированного изменения направления роста, и получают неоднозначный ответ на него — в зависимости от выбранной ими биологической системы и длительности наблюдения. Но по крайней мере в некоторых работах показано, что это, по-видимому, явления, не связанные прямой причинной связью [21]. У многоклеточных организмов морфогенез происходит таким образом, что в конечном счете путь дифференциации и специализации каждой клетки как бы предопределен: тотипотентные клетки, т. е. клетки, обладающие полной генетической информацией, претерпевают специализацию по существенно различным путям [22]. Любой шаг дифференциации может рассматриваться как выбор между возможностями. В настоящее время принято считать, что положение клетки является основным фактором, определяющим этот выбор [23; 10; 17]. Очевидно, что в регуляции морфогенеза принимают участие как генетические, морфогенетические и метаболические системы [24], так и системы надгенной регуляции, которые включают или создают условия реализации генетических механизмов. Поскольку надгенная регуляция связана с физическими предпосылками: освещением, наличием питательных веществ, влажностью и др., а также действием физических законов: градиентное перемещение веществ по растению, давление массы клеток, полярное перемещение ионов и т. д. — и проявляется неравномерно на отдельных участках растения, то подобную регуляцию удобно представить в терминах поля. Поле — это любое явление, нарушающее внутреннее равновесие (т. е. приводящее к полярности) в отдельной клетке, популяции клеток, органе или организме и вызывающее включение генетических и реали- 74 Факторы формирования проводящей системы видов семейства бобовых 75 зацию надгенных программ, способствующих ориентированному делению, росту (растяжению) клеток, контролю и поддержанию формы, а также это группа явлений, определяющих место в ткани (органе), где происходит дифференциация клеток [25]. Таким образом, поле или сочетание полей выступают как условие реализации морфогенеза. Чаще всего происходит совместное действие полей, однако в некоторых случаях существует иерархия, когда действие одного из полей превалирует над остальными и/или обусловливает их возникновение. Как правило, это относится к росту и развитию одного органа, а также к дифференциации и специализации отдельной клетки. Отсюда вопрос о механизме дифференциации — это вопрос о факторах, побуждающих гены к их экспрессии. Кратко дифференциацию отдельной клетки в клетку проводящей системы можно представить следующим образом. Все клетки поляризованы, и всякий раз, когда в популяции за счет делений накапливается их критическое число, создаются специфические условия: количественно-позиционное воздействие окружающих клеток на клетку, которой предстоит дифференцироваться, усиление тока метаболитов через плазмодесмы, повышение содержания CO2 (ведет к понижению рН). По-видимому, включается механизм, заставляющий клетку приобрести качества «клетки-мишени», и она становится чувствительной к присутствию гормона. Происходит полярное передвижение иона Са2+, микротрубочки пространственно ориентируются, на плазмолемме обозначаются направления мембранных перестроек. Активируются ранее синтезированные ферменты, которые при нормальных условиях находятся в лизосомах. Как только в определенной зоне популяции складываются «подходящие» условия, мембраны лизосом разрушаются и ферменты выплескиваются в цитоплазму. Ферменты гидролизуют полимеры и мембраны; в клетках возрастает концентрация осмотически деятельных веществ; в такие клетки усиленно поступает вода, и они растягиваются. Из клеток первичной меристемы образуются клетки прокамбия. По ходу дальнейшего растяжения внутренних клеток процессы лизиса усиливаются, и некоторые клетки прокамбия теряют все внутреннее содержимое, из них образуются членики сосудов, входящих в состав ксилемы. У других клеток, расположенных рядом с члениками сосудов, лизируются ядро, тонопласт, синтезирующая белок система и другие внешние структуры. Цитоплазма в таких клетках тонким слоем выстилает внутренние стенки клетки. В клетках сохраняются митохондрии, но АТФ не синтезируется. Эти членики щитовидных трубок составляют основу флоэмы. Флоэма и ксилема образуют проводящую систему растений [26]. Сходство и различие в жилковании листьев возможно объяснить наличием двух влияющих на жилкование групп систем регуляции. Первая группа — это «консервативные» системы, определяющие внутривидовое сходство жилкования: сюда входят такие системы регуляции, как генетическая и количественно-позиционная. Вторая — реактивная группа, которая состоит из двух подсистем: первой — метаболической и гормональной; они связаны с генетической, но находятся в большей степени под воздействием внешних факторов; и второй подсистемой — трофической и электро-физиологической, которые постоянно находятся под воздействием факторов света, гравитации, механического давления и т. д. [27]. И, таким образом, различия в жилковании связаны с изменяющимся в онтогенезе воздействием реактивной системы. 75 Е. А. Коршунов Список литературы 76 1. Головина Е. Ю., Мясоедова М. Ю. Морфофизиология специализированных побегов многолетних травянистых растений // Программа и тезисы докладов всероссийского совещания. Сыктывкар, 2000. С. 59—60. 2. Боннер Дж. Развитие // Биохимия растений. М., 1968. С. 520—530. 3. Романов Г. А. Рецепторы фитогормонов // Физиология растений. 2002. Т. 49. № 4. С. 615—625. 4. Хавкин Э. Е. Генетическая регуляция морфогенеза растений // Физиология растений. 1998. Т. 45. № 5. С. 763—777. 5. Рейвн П., Эверт Р., Айкхорн С. Современная ботаника. М., 1990. Т. 2. 6. Лотова А. И. Морфология и анатомия высших растений. М., 2001. 7. Коршунов Е. А. Структурные характеристики листьев бобовых растений. СПб., 2001. 8. Коршунов Е. А. Факторы жилкования горошка мышиного. СПб., 2002. 9. Коршунов Е. А. Система жилкования листьев чины лесной // Труды Аристотелевской академии формы. СПб., 2002. Т. 11. 10. Бурень В. М., Недорезков В. Д. Онтогенез пшеницы в числах и схемах. СПб., 2001. 11. Коршунов Е. А. Развитие и морфометрия некоторых бобовых // Сборник научных трудов педагогического университета им. Герцина. СПб., 2002. 12. Эсау Е. Анатомия растений. М., 1979. 13. Кефели В. И. Рост растений. М., 1984. 14. Гамалей Ю. В. Эндоплазматическая сеть растений. Происхождение, структура и функции. СПб., 1994. 15. Гамалей Ю. В. Отток фотоассимилятов в природных и экспериментальных условиях //Физиология растений. 1996. Т. 43. № 3. С. 328—343. 16. Гамалей Ю. В. Надклеточная организация растений //Физиология растений. 1997. Т. 44. № 6. С. 819—846. 17. Медведев С. С., Маркова М. В. Цитоскелет и полярность растений // Физиология растений. 1998. Т. 45. № 2. С 185—197. 18. Альбертс Б., Брейн Д., Льюис Д. и др. Молекулярная биология клетки. М., 1994. Т. 3. 19. Лебедев С. И. Физиология растений. 2-е изд., перераб. и доп. М., 1982. 20. Грин Н., Стаут У. Тейлор Д. Биология / Пер. с англ. 2-е изд., стереотип. Т. 2. М., 1996. 21. Клячко Н. Л. Фитогормоны и цитоскелет // Физиология растений. 2003. Т. 50. № 3. С. 475—480. 22. Гродзинский Д. М. Биофизика растений. Киев, 1972. 23. Нагл В. Роль хромосом в дифференцировке // Современные достижения молекулярной биологии хромосом и клеток / Пер. с англ. Алма-Ата, 1989. 24. Конарев В. Г. Морфогенез и молекулярно-биологический анализ растений. Изд. 2-е, доп. СПб., 2001. 25. Коршунов Е. А. К вопросу о биологическом поле // Материалы международного научно-практического семинара. Калининград, 2005. 26. Бурень В. М. Формирование организма на земле // Труды Аристотелевской академии формы. Т. 1. СПб.; Пушкин, 1998. 27. Коршунов Е. А. Проводящие системы листочков одного листа караганы древовидной // Научно-информационный бюллетень ВНИИР им. Вавилова. 2003. № 242. [Электрон. ресурс]. Режим доступа: kea_1000@mail.ru. Об авторе Е. А. Коршунов — сотрудник агрофирмы «Бауэр Тэхникс», kea_1000@mail.ru. 76