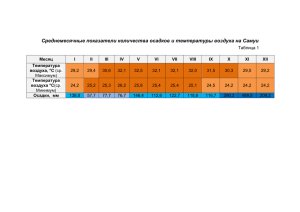
ВИДОВОЕ РАЗНООБРАЗИЕ И ЭКОЛОГИЯ ПЧЁЛ СЕМЕЙСТВА
advertisement

1 Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования «Кемеровский государственный университет» На правах рукописи Яковлева Светлана Николаевна ВИДОВОЕ РАЗНООБРАЗИЕ И ЭКОЛОГИЯ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE (HYMENOPTERA, APOIDEA) КУЗНЕЦКО-САЛАИРСКОЙ ПРОВИНЦИИ 03.02.08 – экология (биология) Диссертация на соискание учёной степени кандидата биологических наук Научный руководитель: д-р биол. наук, профессор Н.И. Еремеева Кемерово – 2014 2 ОГЛАВЛЕНИЕ ВВЕДЕНИЕ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ГЛАВА 1. ИСТОРИЯ ИЗУЧЕНИЯ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ГЛАВА 2. ПРИРОДНО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА КУЗНЕЦКО-САЛАИРСКОЙ ПРОВИНЦИИ . . . . . . . . . . . . . . . . ГЛАВА 3. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ . . . . . . . . . . . ГЛАВА 4. ВИДОВОЕ РАЗНООБРАЗИЕ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE КУЗНЕЦКО-САЛАИРСКОЙ ПРОВИНЦИИ. . . . . . 4.1. Аннотированный список видов пчёл семейства Megachilidae Кузнецко-Салаирской провинции . . . . . . . . . . . 4.2. Видовое разнообразие и численное обилие пчёл семейства Megachilidae Кузнецко-Салаирской провинции . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4.3. Зоогеографическая характеристика фауны пчёл семейства Megachilidae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ГЛАВА 5. ЭКОЛОГИЧЕСКАЯ ДИФФЕРЕНЦИАЦИЯ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE КУЗНЕЦКОСАЛАИРСКОЙ ПРОВИНЦИИ . . . . . . . . . . . . .. . . . . . . . . . . . . . . 5.1. Биотопические группы . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5.2. Трофические группы . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ГЛАВА 6. ОСОБЕННОСТИ ЛАНДШАФТНО-БИОТОПИЧЕСКОЙ ДИФФЕРЕНЦИАЦИИ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE КУЗНЕЦКО-САЛАИРСКОЙ ГОРНОЙ ОБЛАСТИ. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6.1. Особенности распределения пчёл семейства Megachilidae по орографическим регионам КузнецкоСалаирской провинции . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6.2. Топические комплексы мегахилид. . . . . . . . . . . . . . . . . . . . 6.3. Пчёлы сем. Megachilidae в антропогенных местообитаниях. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6.4. Пчёлы сем. Megachilidae в городской среде . . . . . . . . . . . . ВЫВОДЫ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ . . . . . . . . . . . . . . . . . . . . . . . . . . . СПИСОК ЛИТЕРАТУРЫ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . стр. 3 9 21 37 46 46 120 132 144 144 157 172 172 181 193 198 208 210 211 3 ВЕДЕНИЕ Актуальность темы исследования. Megachilidae – одно из наиболее обширных и высокоорганизованных семейств пчёл, насчитывающее в мировой фауне более 3140 видов 74 родов 2 подсемейств [Michener C., 2007]. Данная группа распространена повсеместно, её представители встречаются на всех континентах, кроме Антарктиды. Наибольшее разнообразие мегахилиды демонстрируют в тропиках и пустынях [Попов В.В, 1948; Мариковская Т.П., 1972; Радченко В.Г., Песенко Ю.А., 1994]. В Палеарктике семейство представлено главным образом подсемейством Megachilinae (36 родов из 71 известных); второе подсемейство – Fideliinae – является преимущественно афротропической группой и на территории Палеарктики представлено только 1 родом Pararhophites Friese, 1898 [Попов В.В., 1939; Радченко В.Г., Песенко Ю.А., 1994; Whitehead V., Eardley C., 2003; Michener C., 2007]. Пчёлы семейства Megachilidae энтомофильных растений. Многие являются авторы эффективными опылителями отмечают значительную роль мегахилид в повышении урожайности посевной люцерны, плодовых культур, подсолнечника [Попов В.В., 1950, 1951, 1956; Никифорук К.С., 1957; Пономарева А.А., 1960, 1962; Песенко Ю.А., 1971, 1975; Delaplane K., Mayer D., 2000; Kemp W., Bosch J., 2000; Maccagnani B. et al., 2003; Ladurner E. et al., 2004; Monzon V. et al., 2004; Shebl M. et al., 2009]. Они отличаются широкой пластичностью гнездостроительного поведения, способностью развиваться и выводить потомство в искусственных условиях. Благодаря этим этологическим и биологическим особенностям мегахилиды являются наиболее перспективной группой пчёл в отношении промышленного разведения [Bohart G., 1962, 1964, 1970; Песенко Ю.А., 1976, 1982 а; Волошина Т.А., 1984; Ромасенко Л.П., 1990; Ladurner E. et al., 1999; Иванов С.П., Фатерыга А.В., 2006]. Степень разработанности темы исследования. На территории России фауна и экология мегахилид достаточно подробно изучены лишь в европейской части [Левченко Т.В., 2010], Восточной Сибири и Дальнем Востоке страны 4 [Прощалыкин М.Ю., 2011]. Специальных исследований видового разнообразия и экологии пчёл сем. Megachilidae на территории Западной Сибири и КузнецкоСалаирской провинции до настоящего времени не проводилось. Изучению этой группы посвящено лишь несколько публикаций [Кузьмин П.М., Молчанов А.Е., 1983; Романькова Т.Г., 1983; Сарычева З.Н., Тарасенко Н.Д., 1989; Romankova T., 2003; Прощалыкин М.Ю., 2013 б], некоторые данные по фауне этого семейства имеются также в работах С.Д. Лаврова [1927], А.А. Пономаревой [1967], Д.В. Панфилова [1968], П.С. Шумаковой с соавт. [1982], О.Л. Конусовой и В.В. Янюшкина [2000], Ю.Н. Данилова [2006], М.Ю. Прощалыкина [2013 а]. Наиболее полные данные о фауне мегахилид опубликованы для Курганской и Томской областей [Кузьмин П.М., Молчанов А.Е., 1983, 1987; Конусова О.Л., Янюшкин В.В., 2000]; для остальных территорий имеются лишь отрывочные сведения. Особенности населения и экологии мегахилид в Кузнецко-Салаирской провинции рассмотрены в единичных обзорных работах, посвящённых главным образом фауне пчёл отдельных территорий провинции [Сидоров Д.А., 2005; Еремеева Н.И., Сидоров Д.А., 2006; Еремеева Н.И. и др., 2009]. Таким образом, имеющиеся данные не позволяют составить целостного представления о фауне и особенностях экологии пчёл сем. Megachilidae на исследуемой территории. Целью работы является изучение видового разнообразия и экологических особенностей пчёл семейства Megachilidae в Кузнецко-Салаирской провинции. Для достижения цели были поставлены следующие задачи: 1. Установить видовое разнообразие, таксономический и ареалогический состав, структуру доминирования пчёл семейства Megachilidae в КузнецкоСалаирской провинции. 2. Составить аннотированный список видов с данными по экологическим особенностям, биологии и распространению мегахилид Кузнецко-Салаирской провинции. 3. Выявить особенности и закономерности распределения мегахилид по орографическим регионам и типичным биотопам Кузнецко-Салаирской провинции. 5 4. Изучить трофические связи гнездостроящих видов пчёл семейства Megachilidae на исследуемой территории. 5. Выявить особенности видового разнообразия и экологии пчёл сем. Megachilidae в условиях городской среды на примере урбанизированных территорий Кемеровской области. Научная новизна. Впервые проведён эколого-фаунистический обзор пчёл семейства Megachilidae в Кузнецко-Салаирской провинции. Изучена экологическая дифференциация мегахилид, выявлены основные экологические группы видов. Проанализировано распространение пчёл орографическим регионам приуроченность особенности в регионах, населения исследуемой установлены мегахилид на сем. Megachilidae по территории, трофические антропогенно биотопическая связи. Изучены преобразованных и урбанизированных территориях Кузнецко-Салаирской провинции. Составлен аннотированный список видов пчёл семейства Megachilidae Кузнецко-Салаирской провинции, включающий данные по распространению, экологическим и биологическим особенностям (фенология, особенности гнездования, трофические предпочтения). В результате исследований на территории Кузнецко-Салаирской провинции выявлено 49 видов мегахилид из 12 родов 1 подсемейства. Из них 1 род и 6 видов указаны впервые для Западной Сибири, 13 видов – впервые для КузнецкоСалаирской провинции. Теоретическая и практическая значимость работы. Изучение видового разнообразия и экологических особенностей пчёл сем. Megachilidae необходимо для наиболее рационального использования биологических ресурсов, выявления роли мегахилид в естественных и преобразованных экосистемах, а также охраны редких видов в условиях возрастающего антропогенного пресса. Полученные в результате исследований данные могут быть использованы для дальнейших экологических исследований естественных и преобразованных экосистем Кузнецко-Салаирской провинции, при составлении списков видов животных, каталогов по фауне пчёл Сибири, разработке рекомендаций о включении редких 6 видов в Красную книгу Кемеровской области и практических мер сохранения биоразнообразия пчёл сем. Megachilidae. Данные об экологических особенностях мегахилид можно использовать для привлечения их в антропогенные ландшафты в качестве эффективных опылителей культурных растений. Сведения о населении мегахилид на урбанизированных территориях могут служить основой для разработки систем биоиндикации степени антропогенной нагрузки. Результаты работы используются при изучении курсов «Общая экология», «Экология популяций и сообществ», «Экология городских экосистем», «Биоиндикация окружающей среды», «Экологический мониторинг», «Введение в биоразнообразие» на биологическом факультете КемГУ (направление «Экология и природопользование»). Методология и методы исследования. В работе были использованы стандартные методики, широко используемые в экологических исследованиях пчелиных: - методы сбора и первичной обработки материала (сбор преимущественно индивидуальный, сачком или пинцетом с цветущих растений или на лету; унифицированная монтировка и этикетирование); - определение материала и кормовых растений, составление аннотированного списка изученных видов; - выделение классов обилия (расчет верхнего и нижнего пределов численного обилия классов); - методы оценки видового разнообразия (расчет индексов видового богатства, меры доминирования); - методы сравнения ряда местообитаний (коэффициент ЧекановскогоСъёренсена); - методы графического представления результатов. Основные положения, выносимые на защиту. 1) Пчёлы семейства Megachilidae характеризуются высокой экологической пластичностью, которые являются предпосылкой успешного освоения ими антропогенно преобразованных территорий. 7 2) Население пчёл сем. Megachilidae Кузнецко-Салаирской провинции формируется главным субдоминантных) образом видов с за счёт широкой массовых (доминантных биотопической и и трофической приуроченностью. Степень достоверности и апробация результатов. Достоверность результатов исследований определена применением традиционных методических подходов и классических методов экологии, а также статистической обработкой данных и анализом полученных результатов. Достоверность определения видов обусловлена исследованием обширного материала и типовых экземпляров значительного числа видов. Материалы по результатам исследований представлены на XXХIII и XXХIV Международных научных конференций студентов, аспирантов и молодых ученых (Кемерово, 2006, 2007), XIII съезде Русского энтомологического общества (Краснодар, 2007), III Международной конференции «Биоразнообразие, проблемы экологии Горного Алтая и сопредельных территорий: настоящее, прошлое, будущее» (Горно-Алтайск, 2013), X Международной научной конференции «Наука и образование» практической (Белово, конференции 2014), «Экология VI XXI Межрегиональной века: актуальные научновопросы образования, науки и производства» (г. Кемерово, 2014). Публикации. По теме диссертационного исследования опубликовано 6 печатных работ, из них 3 статьи в журналах из перечня, рекомендованного ВАК РФ. Структура и объём диссертации. Текст диссертационной работы изложен на 242 страницах текста и состоит из введения, 6 глав, выводов, практических рекомендаций и списка литературы. В работу включено 52 рисунка и 13 таблиц. Список литературы содержит 280 источников, из которых 64 – на иностранных языках. Благодарности. Автор глубоко благодарен и признателен научному руководителю, доктору биологических наук, профессору кафедры зоологии и экологии Кемеровского государственного университета Н.И. Еремеевой за 8 помощь, чёткую координацию и содействие при выполнении работы, критические замечания, ценные советы, корректировку, а также предоставленные для работы материалы и безграничное терпение. Автор выражает благодарность сотрудникам кафедры зоологии и экологии КемГУ Д.А. Сидорову, С.Л. Лузянину, А.В. Коршунову, а также О.Л. Конусовой (Томск, ТГУ) за поддержку, ценные советы, замечания, помощь при сборе материала и предоставленные коллекции. Особая благодарность – сотрудникам отдела систематики перепончатокрылых насекомых Зоологического института РАН (Санкт-Петербург): д.б.н. Ю.А. Песенко – за поддержку, ценные советы, предоставленные литературные источники и возможность работы со сравнительным материалом коллекций ЗИН РАН, куратору коллекций ЗИН к.б.н. Ю.В. Астафуровой – за советы и помощь при работе с коллекциями, д.б.н. С.А. Белокобыльскому – за доброжелательность и поддержку. Глубокая признательность выражается специалистам к.б.н. Т.В. Левченко (Москва, Государственный Дарвиновский музей) и к.б.н. М.Ю. Прощалыкину (Владивосток, Биолого-почвенный институт РАН) – за помощь в поиске литературных источников, полезные советы и критические замечания. Автор благодарен также сотрудникам кафедры ботаники КемГУ – зав. кафедрой, к.б.н. Л.Н. Ковригиной, ведущему инженеру И.В. Тарасовой, и сотрудникам ИЭЧ СО РАН (Кемерово) к.б.н. С.А. Шереметовой и к.б.н. Т.Е. Буко – за помощь в определении кормовых растений и геоботаническом описании участков. 9 ГЛАВА 1. ИСТОРИЯ ИЗУЧЕНИЯ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE Семейство Megachilidae вызывает значительный интерес у исследователей, являясь одним из наиболее молодых и высокоспециализированных семейств пчёл [Попов В.В., 1954 б]. Наличие морфологически однородных, но значительно различающихся по экологическим особенностям групп обусловливает трудность изучения мегахилид и делает семейство интересным в первую очередь для экологов [Благовещенская Н.Н., 1983]. Разномасштабные исследования пчёл семейства Megachilidae проводились во многих регионах мира. К настоящему моменту наиболее полно изучена фауна мегахилид на территории Неарктики [Ромасенко Л.П., 1984]. Для Палеарктики также имеется большое число обзорных работ, ревизий, а также публикаций, посвящённых изучению систематики и таксономии отдельных групп мегахилид, описанию новых видов, изучению распространения и т. д. (работы Г. Зандена, К. Варнке, Б. Ткальцу и др.), но изучена эта территория крайне неравномерно [Давыдова Н.Г., 2003]. В пределах Палеарктики наиболее детальные исследования видового состава и экологических особенностей пчёл сем. Megachilidae проводили на территории Западной и Центральной Европы. Фаунистические списки опубликованы для большинства стран и областей к западу от Польши, Чехии и Словении. Однако в Восточной Европе комплексными исследованиями охвачено лишь небольшое число регионов [Левченко Т.В., 2010 а]. Здесь фауна пчёл, в том числе мегахилид, подробно изучена на территории Литвы [Монсявичюс В.Г., 1988, 1989; Moncevičius V., 1995, 2004]. Фауна пчёл Украины изучена в отношении семейств Colletidae, Andrenidae и Megachilidae [Осичнюк А.З., 1970, 1977; Радченко В.Г., 1982; Ромасенко Л.П., 1989, 1990 а]; исследования видового состава, биологии и экологии пчёл сем. Megachilidae в последние годы проводили в Крыму [Иванов С.П., Фатерыга А.В., 2006, 2007; Иванов С.П. и др., 2007; Радченко В.Г. и др., 2009]. 10 В России фауна пчёл, несмотря на длительную историю изучения, изучена очень неравномерно. Большинство регионов остаются неохваченными масштабными исследованиями. Кроме того, малоизученными остаются биология видов пчёл, их трофические связи и поведение, а также особенности антропогенного влияния на фауну пчёл [Левченко Т.В., 2010 а]. Достаточно подробно изучена фауна пчёл европейской части России. На данной территории пчёл изучали А.С. Скориков, В.В. Попов, Д.В. Панфилов, Ю.А. Песенко [Левченко Т.В., 2010 а]. Опубликованы данные о видовом составе мегахилид Башкирии [Никифорук К.С., 1957], степей Нижнего Дона (Ростовская область) [Песенко Ю.А., 1971], Удмуртии [Ситдиков А.А., 1981, 1986], Среднего Поволжья [Благовещенская Н.Н., 1983, 1994; Попова Л.М., 1990], Пермского края [Лыков В.А., 2007, 2008]. В работах представлены данные о стациальной приуроченности некоторых видов пчёл сем. Megachilidae, их трофических связях и особенностях гнездования. Авторы отмечают важную роль мегахилид как опылителей культурных растений. Относительно недавно опубликованы данные о фауне пчёл Московской области [Левченко Т.В., 2006 а, 2008, 2009 а, б, в, 2010 а, б]. Изучены трофические связи пчёл, их стациальное распределение, дана зоогеографическая характеристика фауны пчёл данной территории. Ряд работ [Левченко Т.В., 2006 б, 2007 а] посвящён пчёлам сем. Megachilidae. Азиатская часть России в отношении как видового состава мегахилид, так и экологических особенностей данной группы пчёл остаётся очень неравномерно изученной. Наиболее полные комплексные исследования фауны и экологии пчелиных проводили на территории Восточной Сибири и Дальнего Востока России [Прощалыкин М.Ю., 2011]. В пределах Восточной Сибири фауна мегахилид достаточно хорошо изучена лишь на территории Забайкалья (Забайкальский край, Бурятия и частично Иркутская область), и в последние годы – в Якутии [Давыдова, Песенко, 2002; Прощалыкин, 2009 а]. Практически неизученной остаётся фауна мегахилид обширной территории Красноярского края и республики Тыва [Прощалыкин, 2013 в]. 11 Первые сведения о фауне пчёл Восточной Сибири опубликованы в конце XIX века Ф. Моравицем и О. Радошковским [Прощалыкин М.Ю., 2013 б]. Данные о видовом составе мегахилид Забайкалья известны из работ Н.Р. Кокуева [1927] и Т. Коккерелла [Cockerell T., 1928]. Дальнейшему изучению фауны мегахилид посвящены работы Т.Г. Романьковой [1983, 1984, 1994]. В 1992 году опубликован первый список видов пчёл сем. Megachilidae Забайкалья, включающий 36 видов [Романькова Т.Г., 1992]. В первой части четвёртого тома «Определителя насекомых Дальнего Востока России» для Забайкалья указывается 11 видов мегахилид [Романькова Т.Г., 1995]; в пятой части четвёртого тома «Определителя» число видов видов мегахилид, указанных для Забайкалья, увеличилось до 15 [Прощалыкин М.Ю., 2009 б]. К настоящему времени опубликована серия обобщающих работ по фауне пчёл Забайкалья [Прощалыкин М.Ю., 2007 а, 2009 а, 2010; Proshchalykin, M. Yu., 2007 и др.]. Для исследуемой территории указан 51 вид мегахилид из 11 родов. Однако, М.Ю. Прощалыкин [2009 б] отмечает, что данные по биологии, поведению и хозяйственному значению пчёл данного региона практически отсутствуют. Подробно исследована фауна пчёл сем. Megachilidae в Якутии. Опубликован список видов, включающий 27 видов пчёл сем. Megachilidae из 6 родов [Давыдова Н.Г., Песенко Ю.А., 2002]. В дальнейшем Н.Г. Давыдовой [2003, 2010] проведён анализ географических ареалов пчёл Якутии, изучена их зональная приуроченность, особенности фенологии и трофические связи. Исследования фауны пчёл тундровой зоны продолжено Л.В. Шелоховской [2009]. Большое число работ посвящено изучению фауны мегахилид Дальнего Востока России. В связи с большой площадью территории данного региона и труднодоступностью многих районов она изучалась крайне неравномерно [Прощалыкин М.Ю., 2005]. С 80-х годов исследования фауны мегахилид данного региона (на территории Приморского края) проводились Т.Г. Романьковой [1983, 1984, 1986]. экологических Впоследствии особенностях Романьков А.В., 1986, 1993]. опубликованы пчёл этого данные семейства о биологических [Романькова и Т.Г., 12 В течение последнего десятилетия опубликована большая серия работ по фауне пчёл Дальнего Востока России [Proshchalykin, M., 2003, 2004, 2009; Romankova T., 2003; Прощалыкин М.Ю., 2003, 2005, 2006 а, б, в, г, 2007 а, б, в, г, д, 2008, 2011, 2012; Proshchalykin M., Lelej A., 2004; Прощалыкин М.Ю. и др., 2004; Proshchalykin M., Kupiaskaya A., 2005; Игнатенко Е.В., Прощалыкин М.Ю., 2005]. Исследованиями охвачены территории Амурской области, Еврейской автономной области, Приморского и Хабаровского краёв, о-вов Сахалин и Монерон, а также Курильских островов. Проведены комплексные исследования видового состава пчёл на данной территории, а также их экологических особенностей, включая трофические связи и фенологию. Опубликованы списки видов мегахилид для Курильских островов (6 видов), острова Сахалин (17 видов), Еврейской автономной области (29 видов), Амурской области (47 видов). Установлено, что фауна пчёл сем. Megachilidae Дальнего Востока России в целом насчитывает 65 видов из 12 родов, из них 5 видов являются эндемиками данной территории [Прощалыкин М.Ю., 2008, 2012]. Кроме того, 2 вида мегахилид обнаружены на Шантарских островах Охотского моря [Kupianskaya A. et al., 2011]. В целом фауна пчёл Забайкалья и Дальнего Востока России насчитывает 70 видов 12 родов пчёл сем. Megachilidae [Прощалыкин М.Ю., 2007 а]. Малоизученной в отношении фауны пчёл, в том числе мегахилид, является территория Западной Сибири [Левченко Т.В., 2010 а]. Регионы с действительно хорошо изученной фауной всех групп пчёл здесь практически отсутствуют. Данные о видовом составе, биологии и экологии пчёл имеются не для всех территорий. Списки видов пчёл разных семейств опубликованы лишь для нескольких областей – Курганской (Anthophoridae, Megachilidae), Томской (Andrenidae, Colletidae), Кемеровской (Apidae – род Bombus, Andrenidae, Megachilidae). По исследуемым регионам имеется лишь небольшое число публикаций, включающих данные о фауне и экологии пчёл сем. Megachilidae. Для остальных групп пчёл на данных территориях имеются немногочисленные предварительные данные, некоторые группы остаются полностью неизученными. 13 Первые фаунистические исследования на территории Западной Сибири датируются началом XVIII века. В экспедиции Д.Г. Мессершмидта по территории современной Тюменской области (1720–1727 годы) была собрана первая коллекция насекомых. Большой вклад в изучение энтомофауны данного региона внёс В.И. Мочульский, посетивший г. Тобольск в 1839 году. Во второй половине XIX века в изучении энтомофауны Западной Сибири активное участие принимали как русские учёные, так и специалисты-энтомологи из Европы в составах многочисленных экспедиционных отрядов. Начало планомерных инвентаризационных фаунистических работ в Западной Сибири положила экспедиция М.Д. Рузского (1896 г.). Активное изучение энтомофауны на данной территории было прервано Первой мировой войной, а затем революционными событиями до начала 1920-х гг. [Ломакин Д.И., Толстиков А.В., 2006]. Впоследствии детальное изучение фауны насекомых, в том числе пчёл, в Западной Сибири продолжено исследованиями В.В. Внуковского [Wnukowskij W., 1926, 1927; Wnukowsky W., 1929, 1936] и профессора С.Д. Лаврова [1927]. В.В. Внуковский в своих работах приводит данные о видовом составе пчёл бывшей Томской современных губернии Кемеровской (исследования и Томской проводились областей и на территориях Алтайского края). Для окрестностей г. Томска автором указано 4 вида пчёл сем. Megachilidae, для территории Каменского района Алтайского края – 5 видов [Wnukowskij W., 1927; Wnukowsky W., 1936]. Одновременно были опубликованы данные о составе энтомофауны г. Омска и его окрестностей [Лавров С.Д., 1927]. На основе материалов, собранных в результате десятилетних исследований автора, составлен список видов насекомых Омского округа. На исследуемой территории автор отмечает 44 вида пчёл, в том числе 8 видов мегахилид, относящихся к 4 родам. Дополнения к указанной работе Лаврова опубликованы В.В. Внуковским в 1930 году [Внуковский В.В., 1930]. Список видов пчёл Омского округа дополнен 4 видами; отмечен еще один вид пчёл р. Megachile. Изучение фауны пчёл на территории Омской области продолжается под руководством А.М. Бывальцева. К настоящему времени фауна 14 пчёл этого региона, за исключением шмелей, остаётся недостаточно изученной [Белова К.А., 2012; Проскурякова А.А., 2012; Прощалыкин М.Ю., 2013 а; Белова К.Е., Телегина Я.Р., 2013]. В 1948–1953 годах продолжены исследования в отношении фауны и экологии пчёл Алтайского края, в рамках изучения пчелиных степной зоны и юга лесостепи Западной Сибири. Комплексное исследование проведено в юго-западной части Кулундинской степи [Шумакова П.С. и др., 1982]. Изучали видовой состав опылителей посевов бобовых культур и дикорастущих бобовых в составе травостоя сопредельных участков. Всего было обнаружено 60 видов пчёл, среди них 9 видов мегахилид из 5 родов. Установлена общая продолжительность лёта пчелиных данного региона, а также определены сроки лёта для некоторых видов. Отмечены различия в привлекательности разных видов бобовых растений для пчёл разных видов, проявление приуроченности ряда видов пчёл к посевным или дикорастущим бобовым. Одновременно (1948–1950 годы) на юге Западной Сибири изучение пчёлмегахилид проводили на территории Восточно-Казахстанской области (г. Усть-Каменогорск и его окрестности) [Пашина Л.С., 1955, 1967]. В рамках исследования биологии и экологии пчёл рода Megachile Казахстана изучали гнездование Megachile bombycina. Описано строение гнезда и используемые для его постройки материалы, а также представлена схема годового цикла данного вида мегахилид. Отмечена также роль мегахилид в опылении посевной люцерны. Обобщённые данные о видовом составе пчёл семейства Megachilidae на территории Сибири приводит Д.В. Панфилов [1968]. Автор отмечает, что в составе пчелиных западносибирской тайги мегахилиды составляют в среднем 5–7 % населения в каждой местности (3–4 вида). В 80-х годах XX века продолжено изучение фауны пчёл Томской области. Е.М. Гришина [1980] отмечает на исследуемой территории около 70 видов пчёл. Кроме этого, опубликованы данные о видовом составе пчёл сем. Megachilidae и Anthophoridae Курганской области [Кузьмин П.М., Молчанов А.Е., 1983]. На исследуемой территории было обнаружено 42 вида мегахилид из 9 родов, 15 указаны наиболее посещаемые растения для каждого вида. Также были отмечены фенологические особенности пчёл, выделено 4 периода и 12 фаз лёта пчёл на протяжённости лётного сезона. На основе собранного материала также проведено исследование гнезд пчёл и их типизация для 45 видов пчелиных, в том числе 15 видов мегахилид [Молчанов А.Е., Кузьмин П.М., 1985]. В 1987 году опубликована работа, посвящённая проблемам охраны колоний пчелиных [Кузьмин П.М., Молчанов А.Е., 1987]. Охране энтомофауны Курганской области посвящены также работы А.Е. Молчанова [1989], Н.А. Уткина [1993], В.А. Балахоновой [2009]. Проанализирован видовой состав и современное состояние популяций видов насекомых исследуемого района. В результате проделанной работы, к настоящему моменту 2 вида пчёл мегахилид (Anthidium septemspinosum, Osmia aurulenta) включены в Красную книгу Курганской области как виды редкие и сокращающиеся в численности (категории II и III соответственно) [Красная книга Курганской области, 2012]. Территорию Курганской области можно считать наиболее изученной в отношении фауны пчёл сем. Megachilidae в Западной Сибири. К настоящему времени достаточно подробно изучен видовой состав различных групп пчёл в Томской области [Конусова О.Л., 1997, 1998, 2000 а, б, 2002; Конусова О.Л., Гришаев Л.В., 2006; Конусова О.Л. и др., 2007]. Опубликованы данные о видовом составе мегахилид южной тайги Томского Приобья (обнаружено 29 видов), дана экологическая характеристика фауны района исследований, в том числе и урбанизированных территорий [Конусова О.Л., Янюшкин В.В., 2000]. Выявлены массовые, обычные и редкие виды, установлены трофические связи пчёл. Дополнительные данные о трофических связях мегахилид для юга Томской области приводят А.С. Прокофьев, О.Л. Конусова и Т.Н. Беляева [2004], исследовавшие видовой состав опылителей очитков. Авторы установили трофические предпочтения для 7 видов мегахилид, относящихся к родам Chelostoma, Hoplitis, Megachile и Coelioxys. Работу по изучению и привлечению диких пчелиных Новосибирской области в 80-х годах проводили А.Г. Красноперов, Н.Р. Богатырёв, В.С. Гребенников 16 [Бывальцев А.М., 2010]. Изучением трофических связей пчёл-мегахилид здесь занимались С.М. Сарычев [1988], З.Н. Сарычева и Н.Д. Тарасенко [1989]. К настоящему моменту фауна пчёл Новосибирской области наиболее полно изучена в отношении представителей рода Bombus. Видовой состав мегахилид Новосибирской области в целом не изучен [Прощалыкин М.Ю., 2013 б]. Современные предварительные данные о видовом составе и экологии пчёлмегахилид Алтайского края опубликованы в работе Ю.Н. Данилова [2006]. Автор приводит список видов пчёл, отловленных в окрестностях г. Барнаул. Наибольшее число отмеченных на данной территории видов (13 видов из 9 родов) относится к семейству Megachilidae. Автор отмечает, что, несмотря на интерес, который местная фауна пчёл может представлять для специалистов, детальных исследований на территории Алтайского края в целом не проводилось. М.Ю. Прощалыкин [2013 а, б] опубликовал данные об известных с территории Сибири и Дальнего Востока России видах пчёл трибы Anthidiini (всего автор отмечает 10 видов пчёл данной трибы), а также о новых для Сибири находках пчёл, в том числе мегахилид. Проведённые исследования охватывают в том числе такие малоизученные территории Сибири, как республики Хакасия и Тыва, Красноярский край. В 2014 г. опубликованы первые данные о фауне пчёл ХантыМансийского автономного округа [Levchenko Т.V., Tomkovich K.P., 2014]. Кузнецко-Салаирская провинция расположена в юго-восточной части Западной Сибири. В её пределах находятся следующие административные территории: Кемеровская область (занимает большую часть провинции), часть Новосибирской области (бассейн р. Иня), Алтайского края (юго-западный склон Салаирского кряжа), крайний юго-запад Республики Алтай (бассейн р. Лебедь) и западная окраина Хакасии (Восточная часть кузнецкого Алатау и Абаканский хребет) [Михайлов Н.И., 1961; Вдовин В.В., 1969]. К настоящему времени большая часть имеющихся данных о видовом составе пчёл сем. Megachilidae касается в основном территории Кемеровской области. Первые сведения о видовом составе пчёл Кемеровской области опубликованы В.В. Внуковским в 1926 году [Wnukowskij W., 1926]. Исследования проводились в 17 северной части Кузнецкого района, в окрестностях с. Кольчугино (современный г. Ленинск-Кузнецкий) и деревень Тараданово (Крапивинский район), Морозово, Титово (Промышленновский район) и Старые Черви (Кемеровский район) в типичных лесостепных ландшафтах и на остепнённых участках. Для исследованной территории было указано 6 видов пчёл из 5 родов, среди них мегахилид не отмечено. Возобновлены исследования фауны и экологии пчёл в Кузнецко-Салаирской провинции относительно недавно. Сбор пчёл продолжен сотрудниками кафедры зоологии и экологии Кемеровского государственного университета в 1980-х годах Опубликованные на основе обработанного материала данные в основном касаются пчёл-апид трибы Bombini. Достаточно подробно освещены особенности населения, видовое разнообразие и экологические особенности этих пчёл во всех орографических регионах Кузнецко-Салаирской провинции [Еремеева Н.И. и др., 2003, 2004; Еремеева Н.И., Лузянин С.Л., 2005, 2008 а, б; Лузянин С.Л., 2006 а, б, в]. Кроме того, опубликованы предварительные данные по фауне и экологии отдельных семейств пчёл, таких как Colletidae [Еремеева Н.И. и др., 2006; Конусова О.Л. и др., 2007], Halictidae [Еремеева Н.И. и др., 2007; Яковлева С.Н., 2006], Apidae (за исключением рода Nomada) [Еремеева Н.И., Сидоров Д.А., 2005; Сидоров Д.А., Яковлева С.Н., 2006; Сидоров Д.А., Еремеева Н.И., 2008], Andrenidae [Сидоров Д.А., 2006, 2008; Сидоров Д.А., Еремеева Н.И., 2010]. В процессе исследований велось активное сотрудничество с О.Л. Конусовой (Томский государственный университет). Подробное изучение видового состава и экологических особенностей пчёл семейства Megachilidae в Кузнецко-Салаирской провинции проводится сотрудниками Кемеровского госуниверситета Н.И. Еремеевой, Д.А. Сидоровым и С.Н. Яковлевой с 2005 года. За это время было опубликовано несколько работ [Сидоров Д.А., 2005; Еремеева Н.И., Сидоров Д.А., 2006; Еремеева Н.И. и др., 2009, 2010, 2011; Яковлева С.Н., 2011, 2012, 2013, 2014], содержащих информацию как о видовом составе мегахилид исследуемой территории, так и об экологических особенностях пчёл данной группы. 18 Первые данные по фауне пчёл рода Megachile Кузнецко-Салаирской провинции и сопредельных территорий опубликованы Д.А. Сидоровым [2005]. Предварительный список включает 13 видов пчёл данного рода. Автор также приводит данные по распространению пчёл рода Megachile на исследуемой территории. В дальнейших исследованиях Н.И. Еремеева и Д.А. Сидоров [2006] отметили в Кузнецко-Салаирской провинции 36 видов мегахилид, относящихся к 10 родам (в том числе ошибочно указанные Anthidium diadema, A. florentinum, Osmia coerulescens, Coelioxys haemorrhoa). Рассмотрено распространение мегахилид по орографическим регионам провинции, отмечено наибольшее видовое и численное обилие мегахилид в Кузнецкой котловине. Для 27 видов приводятся данные о трофических связях. Отмечено, что мегахилиды посещают растения 13 семейств, отдавая предпочтение бобовым и астровым. Комплексные исследования населения и экологии пчёл были проведены в Горной Шории [Еремеева Н.И. и др., 2009]. На данной территории обнаружено 16 видов мегахилид, исследованы особенности их распределения по основным типам биотопов Горной Шории. Отмечена приуроченность пчёл данного семейства в основном к суходольным и лесным разнотравно-злаковым лугам. Таким образом, до начала исследований автора сведения по фауне и экологическим особенностям пчёл семейства Megachilidae в Кузнецко- Салаирской провинции имели неполный, отрывочный характер. Собранный материал был обработан частично, не для всех экземпляров мегахилид была установлена видовая принадлежность. Для большинства видов мегахилид оставались неизвестными трофические связи, биотопическая приуроченность. Дальнейшее изучение фауны и экологии пчёл сем. Megachilidae в КузнецкоСалаирской провинции было продолжено на территориях Кузнецкой котловины, Кузнецкого Алатау и Горной Шории. В результате был опубликован предварительный список видов мегахилид Кузнецко-Салаирской провинции, включающий 42 вида, принадлежащих к 11 родам 4 триб подсем. Megachilinae. Наибольшее число отмеченных видов принадлежит к трибам Megachilini и Osmiini [Яковлева С.Н., 2011]. 19 Наиболее полно изучена фауна мегахилид Кузнецкой котловины [Яковлева С.Н., 2012]. На исследуемой территории установлено обитание 43 видов 13 родов 4 триб 1 подсемейства (за исключением ошибочно указанных A. florentinum и H. turcestanica). 9 видов были отмечены впервые для Кузнецко-Салаирской провинции, 2 вида – впервые для Западной Сибири. Уделено внимание изучению фауны мегахилид ранее малоизученных горных территорий Кузнецко-Салаирской провинции [Еремеева и др., 2010; Яковлева С.Н., 2013]. Наименее изученной остаётся территория Салаирского кряжа, что объясняется недостаточностью исследований в этом регионе. Во всех орографических регионах Кузнецко-Салаирской провинции проведены исследования с целью выявления редких видов перепончатокрылых насекомых [Еремеева Н.И. и др., 2011]. Авторы приводят список видов, включающий 23 видов пчёл, в числе которых 2 вида мегахилид (M. argentata, M. circumcincta); один вид мегахилид – H. turcestanica – указан ошибочно. Кроме того, отмечают места обнаружения редких видов на исследуемой территории. К настоящему моменту правильность определенного материала проверена по эталонным коллекциям Зоологического института РАН, из фаунистического списка исключены ошибочно определённые виды. Список видов мегахилид Кузнецко-Салаирской провинции увеличился до 49 видов 12 родов 5 триб. В настоящее время исследования фауны и экологических особенностей пчёл семейства Megachilidae в Кузнецко-Салаирской провинции продолжаются. Таким образом, история изучения пчёл на территории Сибири началась в конце XIX века с работ Ф. Моравица и О. Радошковского [Прощалыкин М.Ю., 2013]. Первые данные по фауне мегахилид Западной Сибири были опубликованы В.В. Внуковским [Wnukowskij W., 1926, 1927; Wnukowsky W., 1929, 1936] и С.Д. Лавровым [1927]. Дальнейшее изучение Западной Сибири в отношении фауны пчёл, в том числе мегахилид, шло неравномерно. К настоящему времени наиболее изучен видовой состав пчёл сем. Megachilidae на территориях Томской [Гришина Е.М., 1980; Конусова О.Л., 1997; Конусова О.Л., Янюшкин В.В., 2000; Прокофьев А.С. и др., 2004], Курганской [Кузьмин П.М., Молчанов А.Е., 1983, 20 1987; Молчанов А.Е., Кузьмин П.М., 1989] и Кемеровской [Сидоров Д.А., 2005; Еремеева Н.И., Сидоров Д.А., 2006; Еремеева Н.И. и др., 2009, 2010, 2011; Яковлева С.Н., 2011, 2012, 2013] областей. Для этих регионов Западной Сибири в целом известно около 70 видов мегахилид 15 родов. Неизученной остается фауна мегахилид Новосибирской и Омской областей Западной Сибири [Прощалыкин М.Ю., 2013 а, б]. Подробное изучение фауны и экологии мегахилид Кузнецко-Салаирской провинции ведется с 2005 года. К началу исследований автора для данной территории было известно 32 вида пчёл данного семейства, относящихся к 10 родам. Дальнейшее изучение фауны и экологии мегахилид продолжено на территориях Кузнецкой котловины, Кузнецкого Алатау и Горной Шории. 21 ГЛАВА 2. ПРИРОДНО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА КУЗНЕЦКО-САЛАИРСКОЙ ПРОВИНЦИИ 2.1. Географическое положение, рельеф, гидрография, климат, почвы, растительность Кузнецко-Салаирская провинция Кузнецко-Алтайской области Алтае- Саянской горной страны находится в юго-восточной части Западной Сибири. С севера и запада она ограничена соседними участками Западно-Сибирской равнины (Кулундинская, Бийско-Барнаульская, Чулымо-Енисейская котловины), на востоке примыкает к Минусинской котловине. Южная часть граничит с горными массивами окрестностей Телецкого озера и долиной р. Бия [Михайлов Н.И., 1961; Гвоздецкий Н.А., 1968; Вдовин В.В., 1969; Давыдова М.И. и др., 1989]. По геоморфологическим признакам Кузнецко-Салаирская провинция отчётливо разделяется на две части – горную и равнинную. Горный рельеф характерен для двух третей территории провинции: горные хребты в виде подковы с запада, юга и востока окаймляют расположенную в середине Кузнецкую котловину [Михайлов Н.И., 1961; Давыдова М.И. и др., 1989; Соловьёв Л.И., 2010]. Таким образом, исследуемая территория включает в себя 4 орографических региона (Кузнецкая котловина, Салаирский кряж, Кузнецкий Алатау и Горная Шория), отличающихся особенностями рельефа, геологическим строением и происхождением [Михайлов Н.И., 1961; Лаврентьев А.И., 1967; Гвоздецкий Н.А., Михайлов Н.И., 1978] (рис. 2.1). В основном территория Кузнецко-Салаирской провинции расположена ниже 800 м над уровнем моря, но некоторые хребты восточной части достигают высоты 2000 м. Значительная разность высот поверхности определяет разнообразие природных условий [Гвоздецкий Н.А., Михайлов Н.И., 1978; Вдовин В.В., 1988]. 22 Рисунок 2.1. Орографическая схема Кузнецко-Салаирской провинции с указанием границ орографических регионов (приводится по: Гвоздецкий Н.А., Михайлов Н.И., 1978, с изменениями) Горы на территории нижнепалеозойскими Кузнецко-Салаирской кристаллическими провинции известняками, сложены зеленоватыми песчаниками, сланцами, доломитами и вулканическими туфами. Интенсивные процессы складкообразования здесь происходили в эпоху салаирской фазы каледонской складчатости. В среднем палеозое приподнятые участки выступали в виде островов среди морей, занимавших тогда Западно-Сибирскую равнину, 23 Кузнецкую и Минусинскую котловины. Затем, уже в начале четвертичного времени, после длительной эпохи континентального выравнивания, невысокие горы подверглись молодым дифференцированным поднятиям, и их амплитуда на востоке области была более значительной, чем на западе [Гвоздецкий Н.А., Михайлов Н.И., 1978; Давыдова М.И. и др., 1989]. Большинство рек Кузнецко-Салаирской провинции принадлежит к системе правых притоков Оби (Томь, Чулым, Чумыш, Иня, Бердь). Юго-восточная часть провинции относится к бассейнам р. Лебедь (правый приток р. Бия) и Абакан (левый приток р. Енисей). Как правило, они начинаются в горах, где характеризуются быстрым течением и большой прозрачностью воды. Из типов питания преобладают снеговое (40–50 %) и дождевое (30–40 %). Основная часть годового стока приходится на весенние и летние месяцы. Климат Кузнецко-Салаирской провинции в целом заметно отличается от соседних горных территорий более теплым летом и большим количеством осадков, равномерно распределенных по сезонам года. Специфику климата данной территории определяют небольшая высота и особенности рельефа с преобладанием горных массивов, протянувшихся в субмеридиональном направлении [Гвоздецкий Н.А., Михайлов Н.И., 1978]. Климат Кузнецко-Салаирской провинции резко континентальный; амплитуды перепада годовых и суточных температур увеличиваются при продвижении с запада на восток по мере увеличения высоты рельефа. Погода характеризуется значительной изменчивостью. Зима морозная, характеризуется устойчивыми низкими температурами, периодически сопровождающимися ветрами. Температура января в среднем составляет около -20 °С. Снежный покров устанавливается во второй половине октября и держится до апреля. Его толщина может достигать нескольких метров на западном макросклоне Кузнецкого Алатау. Лето тёплое (средние температуры июля составляют 16,9–18,5 °С), с увеличением высоты рельефа – умеренно прохладное, характеризуется значительными перепадами суточных температур, особенно в горах [Хлонов Ю.П., 1979; Шульгин В.Н., 1997]. 24 Территория Кузнецко-Салаирской провинции характеризуется очень неравномерным распределением осадков, что связано главным образом с особенностями рельефа. Годовое количество осадков на равнинных территориях составляет 400–600 мм, в горных регионах – 600–1000 мм, где они в основном приурочены к летнему времени года [Шварёв Ю.О., 1963]. От особенностей рельефа и режима увлажнения зависит распределение типов почв на исследуемой территории. Почвы Кузнецко-Салаирской провинции сформированы на однородных отложениях – лёссовидных карбонатных суглинках или покровных бескарбонатных глинах. Как правило, почвы здесь характеризуются значительной мощностью и богаты гумусом. Маломощные почвы встречаются только в высокогорьях, а также на восточном макросклоне Кузнецкого Алатау [Куминова А.В., 1950; Гвоздецкий Н.А., Михайлов Н.И., 1978]. Почвообразующие породы в горных территориях представлены в основном образом бурыми бескарбонатными глинами и тяжёлыми суглинками небольшого возраста в подножьях горных систем, а также пролювиально-делювиальными отложениями на возвышенных участках [Ильиных Н.И., 1970; Ковалёва С.Р. и др, 1974; Трофимов С.С., 1975]. Территория Кузнецко-Салаирской провинции располагается на границе лесостепного и степного широтных поясов растительности. Благодаря горным массивам, занимающим значительную часть территории провинции, здесь хорошо выражена также высотная поясность [Куминова А.В., 1950; Красноборов И.М. и др., 2001; Лащинский Н.Н. и др., 2007]. В растительном покрове Кузнецко-Салаирской провинции выделяют 5 основных поясов растительности: степной, лесостепной, подтаёжный, горнотаёжный, альпийско-тундровый. Климатически зональным для Кузнецко- Салаирской провинции поясом растительности является лесостепь. Значительное распространение лесов на этой территории обусловлено развитием горных хребтов, окаймляющих котловину. Распределение растительности в КузнецкоСалаирской провинции характеризуется уменьшением ксерофитности при продвижении от центра региона к его периферии с увеличением абсолютных 25 высот [Куминова А.В., 1950; Гвоздецкий Н.А., Михайлов Н.И., 1978; Красноборов И.М. и др., 2001]. Степной пояс на исследуемой территории достаточно развит только на северозападном пониженном участке Кузнецкой котловины, где под влиянием Салаирского кряжа формируется так называемый эффект «дождевой тени». В настоящее время значительная часть степных фитоценозов Кузнецко-Салаирской провинции уничтожена в результате развития сельскохозяйственной деятельности и горнодобывающей промышленности. Лучше всего сохранились каменистые степи, которые встречаются на повышениях рельефа (на юго-западных и восточных склонах Тарадановского и Караканского хребтов и предгорьях Салаирского кряжа) [Куминова А.В., 1950; Матысюк Н.Т., 1980]. Лесостепная растительность (берёзово-осиновые колки, с повышением рельефа переходящие в берёзовые, осиново-берёзовые и смешанные леса) наиболее характерна для территории Кузнецкой котловины. Территория лесостепи в настоящее время значительно расширена за счёт вырубки лесов и расширения площади населённых пунктов. Кроме того, большинство луговых и степных фитоценозов безлесных участков также сильно трансформированы в результате распашки земель и интенсивного выпаса скота. Средняя залесенность лесостепи к настоящему времени составляет не более 10 % [Куминова А.В., 1950; Красноборов И.М. и др., 2001]. Почти половину территории Кузнецко-Салаирской провинции занимают таёжные леса, развитые главным образом в горных регионах. Ведущая роль здесь принадлежит черневой тайге, которая характерна для низко- и среднегорий Горной Шории и Кузнецкого Алатау. Она характеризуется слабо развитыми моховым покровом и лесной подстилкой; в древостое преобладают пихта и осина, подлесок образован различными кустарниками (черемуха, рябина, калина) и высокотравьем [Куминова А.В., 1950]. 26 2.2. Кузнецкий Алатау Кузнецкий Алатау представляет сложный по своему строению массив, окаймляющий Кузнецкую котловину с востока. Это плоскогорье со средними высотами 400–800 м, ориентированное с северо-запада на юго-восток, протянувшееся более чем на 500 км и расчленённое глубокими долинами горных рек на отдельные среднегорные массивы неправильных очертаний. На юго- западе Кузнецкий Алатау сливается с возвышенностями Горной Шории, на юговостоке продолжается Абаканским хребтом [Воскресенский С.С., 1962; Ильичёв А.И. и др., 1995]. На периферии Кузнецкий Алатау постепенно приобретает мягкоочерченный, холмисто-увалистый рельеф. На восток отходят постепенно снижающиеся хребты и массивы, вдающиеся в пространства Минусинской котловины. Местами граница между этими территориями выражена чётко, в виде уступа до 400 м высотой. На севере, постепенно теряя высоту, превращается в пологие увалы и переходит в Западно-Сибирскую низменность. На западе в сторону Кузнецкой котловины Кузнецкий Алатау обрывается чередой уступов высотой 300–400 м [Колобков М.Н., 1956; Дубинкин С.Ф., 1967]. Массив Кузнецкого Алатау обладает большим многообразием форм рельефа, что в значительной мере является результатом его сложной геологической истории и неоднородности состава пород, слагающих хребет. Основная часть Кузнецкого Алатау сформировалась в эпоху каледонской складчатости. Горы сложены известняками, кварцитами, кремнистыми и глинистыми сланцами протерозоя и нижнего палеозоя. Наименьшие абсолютные высоты отмечены в северной части (300–500 м над ур. м.), наибольшие – в южной части (г. Верхний Зуб, 2179 м над ур. м.). Наиболее возвышенные участки связаны с выходами трудно размываемых горных пород. Современный рельеф создан в неогенантропогеновое время в результате поднятия и расчленения разновозрастных поверхностей выравнивания [Додин А.Л., 1948; Михайлов Н.И., 1976; Гвоздецкий Н.А., Михайлов Н.И., 1978; Седельников В.П., 1979]. 27 Кузнецкий Алатау, особенно его западный склон, отличается густой гидрографической сетью. Реки западного склона принадлежат бассейну Томи, их долины более узкие и глубокие, русла имеют больший уклон по сравнению с реками восточного склона. Это связано с более крутым спадом высот Кузнецкой котловины по сравнению с Минусинской. Крупнейшие водотоки Кузнецкого Алатау – реки Белый и Чёрный Июс, Томь в верхнем течении, Кия, Верхняя, Нижняя и Средняя Терси. Восточный склон Кузнецкого Алатау характеризуется обилием небольших озёр, занимающих дно бессточных впадин [Додин А.Л., 1948; Куминова А.В., 1950]. Климат Кузнецкого Алатау резко континентальный, с преобладанием отрицательных температур. Зима здесь продолжительная, средние температуры января составляют -15 °С в средних высотах; лето короткое и холодное. Продолжительность периода с среднесуточными температурами выше 10 °С составляет 60–70 дней, выше 15 °С среднесуточная температура не поднимается. Кузнецкий Алатау характеризуется максимальным во всей Западной Сибири количеством осадков, выпадающим на западных склонах (2100–2500 мм), на восточных склонах всего около 500 мм в год [Шварёв Ю.О., 1963]. Выпадение большей части осадков связано с зимним временем года, вследствие чего формируется снеговой покров мощностью в несколько метров, сохраняющийся от 6 до 9 месяцев. Для высокогорий Кузнецкого Алатау характерны сильные юго-западные хребтов, преграждающих ветры, обусловленные расположением основной путь движения воздушных масс [Хлонов Ю.П., 1979; Седельников В.П., 1979; Вдовин В.В., 1988]. Согласно почвенно-географическому районированию Алтае-Саянской горной страны Кузнецкий Алатау отнесен к циклонической почвенной провинции [Петров Б.Ф., 1952]. Почвы высокогорий и вершин гольцов представлены горнотундровым перегнойным типом. Выше лесного пояса расположены также горнотундровые и горно-луговые почвы. Для них характерны фитоценозы, сформированные главным образом зелёными мхами [Ильиных Н.И., 1970]. 28 Около снежников встречаются горно-луговые альпийские почвы, фитоценозы на них представлены альпийскими лужайками и субальпийскими лугами у верхней границы лесного пояса луговинами. Под отмечены горные дерново-луговые субальпийские почвы [Ильиных Н.И., 1970; Ковалёва С.Р. и др., 1974]. Горно-таёжные иллювиально-гумусовые почвы развиваются у верхней границы подгольцовых кедровников, в подпоясе кедрово-пихтовой тайги – горнотаёжные бурые; на пологих склонах и в подножиях горных массивов распространены глубокооподзоленные почвы [Ковалёва С.Р. и др., 1974]. Западный макросклон Кузнецкого Алатау характеризуется серыми лесными и дерново-подзолистыми почвами, распространёнными за пределами черневой тайги в зоне вторичных лиственных лесов. При спуске в сторону Кузнецкой котловины начинают преобладать выщелоченные и оподзоленные чернозёмы, приуроченные к лесостепному типу растительности [Трофимов С.С., 1975]. В Кузнецком Алатау хорошо развит высокогорный пояс растительности, расположенный выше 1500 м над ур. м. В его пределах выделяют подгольцовый и гольцовый подпояса с преобладающими луговым и тундровым типами растительности соответственно. Тундровая растительность, как правило, приурочена к участкам между курумами, субальпийские луга – к нижней части высокогорного пояса. Там же встречаются небольшие участки лесной растительности с доминированием пихты и кедра сибирского. Верхние части высокогорий характеризуются небольшими участками альпийской луговой растительности. Между берёзовое криволесье лесным и высокогорнымм поясами встречается с доминированием берёзы извилистой, а также кустарниковые и болотные фитоценозы [Куминова А.В., 1950; Шумилова Л.В., 1962; Седельников В.П., 1979]. 2.3. Горная Шория Южную часть Кузнецко-Салаирской провинции занимает Горная Шория. Это типичная среднегорная страна с преобладанием сильно расчленённых долинами рек и ручьёв пологих водоразделов, основная масса которых имеет высоты, не 29 превышающие 1200 метров. Горная Шория расположена на стыке трёх горных систем. На юге этот регион примыкает к Алтаю, на северо-востоке – к Кузнецкому Алатау, на западе сливается с холмистой равниной приобского плато, а на северо-западе через Ненинско-Чумышский дол постепенно смыкается с Салаирским кряжем. Горная Шория занимает площадь около 30 тыс. км2, что составляет почти треть территории Кемеровской области. Она протянулась на 170 км с севера на юг и на 100 км с запада на восток [Колобков М.Н., 1956; Воскресенский С.С., 1962; Мытарев А.А., 1973]. Горы в Шории сформированы нижнепалеозойскими осадочными породами, собранными в крупные складки, разорванные сбросами и надвигами и сильно метаморфизированные. Современный рельеф Горной Шории образовался в результате длительного разрушения пород и постепенного поднятия всей территории. Абсолютные высоты массива, как правило, не превышают 1000– 1200 м, однако встречаются вершины, которые сохранили прежние черты высокогорного рельефа и имеющие высоты 1600–1800 м – Мустаг, Патын, Куль-Тайга. Горная Шория является сложной горной системой в форме платообразной возвышенности, глубоко расчленённой эрозионными долинами, разделёнными плоскими водоразделами и короткими массивами разной протяжённости. В отличие от Кузнецкого Алатау, для Горной Шории характерен длинный и пологий западный макросклон большинства хребтов и короткий и крутой восточный [Воскресенский С.С., 1962; Вдовин В.В., 1988; Шеметов Г.А., Андреева О.С., 1997]. Абаканский хребет и Бийская грива представляют собой наиболее крупные и протяжённые массивы Горной Шории. Они ограничивают её с востока и юга. Шорский хребет, тянущийся с юго-запада на северо-восток от долины р. Кондома до долины р. Мрассу, разделяет горную страну на две части. Северо-западная низкогорный часть характеризуется средними абсолютными высотами порядка 600–750 м и сглаженными извилистыми водоразделами. Низкогорный рельеф понижается в направлении с юго-востока на северо-запад до 180–350 м, постепенно выравнивается и приобретает черты гривисто-увалистого. 30 Юго-восточная часть Горной Шории представлена главным образом хребтами, вытянутыми с севера на юг и образующими систему параллельных и продольных речных долин. Её среднегорный рельеф характеризуется значительной расчленённостью. Абсолютные высоты водоразделов здесь достигают 1200 м, глубина долин может превышать 400 м [Михайлов Н.И., 1961]. Гидрологическая сеть Горной Шории формируется типичными горными водотоками – реками Мрассу в юго-восточной части и Кондомой в северозападной. Обе реки являются крупными левыми притоками р. Томь и характеризуются быстрым течением и порожистыми, валунистыми руслами. Долины рек широкие, имеют пологие склоны, что является характерной особенностью Горной Шории по сравнению с другими горными орографическими регионами Кузнецко-Салаирской провинции [Воскресенский С.С., 1962]. Климат Горной Шории континентальный, с большим перепадом среднегодовых и среднесуточных температур. Температура января составляет в среднем около -19 °С, июля – 16 °С. Осадков выпадает 800–1300 мм в год, большая часть их приурочена к тёалому весеннему и летнему периоду (до 900 мм) [Михайлов Н.И., 1961]. Распределение осадков характеризуется неравномерностью: в низкой северо-западной части они более обильны, чем в юго-восточной части, характеризующейся большими высотами [Шульгин В.Н., 1997]. Преобладающими типами почв являются дерново-подзолистые и, в меньшей степени, подзолистые почвы. Дерново-подзолистые почвы распространены преимущественно в нижней части таежного пояса, а подзолистые – в верхней его части [Ковалёва С.Р. и др., 1974]. Главной особенностью растительного покрова Горной Шории является преобладание черневой тайги на высотах 300–800 м над ур. м. и широкое развитие пихтово-кедровой тайги в пределах 800–1000 м над ур. м. Нижняя граница черневой тайги плавно переходит в пояс лесостепи, а верхняя граница пихтовокедровой тайги на отдельных гольцах смешивается с участками альпийских и 31 субальпийских лугов, подгольцовыми кедровниками или лесотундровым криволесьем [Куминова А.В., 1950; Шумилова Л.В., 1962]. 2.4. Салаирский кряж Салаирский кряж представляет собой асимметричную, скорее холмистую, чем горную, систему. Главный хребет и отроги ярко выражены лишь в центральной части кряжа. Несмотря на утрату черт горного рельефа, он достаточно чётко обособлен от соседних орографических регионов – Горной Шории, Кузнецкой котловины, Западно-Сибирской низменности. Кряж вытянут в субмеридиональном направлении и по форме напоминает дугу, выпуклой часть обращённую в северо-восточном направлении. Длина кряжа достигает 300 км, ширина – около 100 км, общая площадь составляет 22,5 тыс. км2 [Воскресенский С.С., 1962; Вдовин В.В., Малолетко А.М., 1969; Лащинский Н.Н., Ревякина М.П., 1991]. Восточные границы Салаирского кряжа проходят по выходам палеозойского фундамента в долине р. Чумыш на юго-востоке, а северо-восточная граница четко выражена Тырганским уступом высотой в 100–200 м, круто обрывающимся в сторону Кузнецкой котловины [Ильичёв А.И., Соловьёв Л.И., 1994]. От массивов Горной Шории кряж отделён долиной р. Неня. Пологий западный склон Салаирского кряжа постепенно сливается с Бийско-Барнаульской котловиной на юго-западе. По мере продвижения на север кряж сглаживается, от соседних Кузнецкой котловины и Западно-Сибирской низменности отграничен долиной р. Чема. Салаирский кряж кристаллическими образован известняками, в раннем песчаниками, Кембрии. туфами, Горы сложены гранитами. В формировании Салаирского кряжа отсутствовал этап вторичного орогенеза, который претерпели хребты Кузнецкого Алатау и Горной Шории. С этим связаны низкие средние абсолютные высоты кряжа, не превышающие 420–450 м. Наибольшие высоты характерны для южной и центральной частей Салаира (г. Кивда, 618 м). Кряжу свойственна расчленённость густой сетью рек и логов; 32 особенно это касается восточного склона. В рельефе преобладают длинные отлогие склоны, широкие долины рек и уплощённые междуречья, которые придают рельефу кряжа характер эрозионного плато. На восточном макросклоне кряжа отмечены также карстовые формы рельефа, сформировавшиеся, повидимому, в юре и начале третичного периода [Воскресенский С.С., 1962; Дубинкин С.Ф., 1967; Малолетко А.М., 1972; Мытарев А.А., 1973; Ковалёва С.Р. и др., 1974; Гвоздецкий Н.А., Михайлов Н.И., 1978; Вдовин В.В., 1988]. Реки северной части Салаирского кряжа относятся к бассейну р. Иня, центральной и южной – к бассейнам Берди и Чумыша. Озёра в этом орографическом регионе встречаются редко и характеризуются главным образом старичным происхождением [Куминова А.В., 1950; Хлонов Ю.П., 1979; Лащинский Н.Н., 2009]. Салаирский кряж характеризуется резкой континентальностью климата. Для января характерны средние температуры около -20 °С (минимальные – -50 °С) , для июля – 19 °С (могут подниматься до 40 °С). Среднегодовое количество осадков изменяется в пределах 400–600 мм. Как и Кузнецкий Алатау, Салаирский кряж также представляет собой крупное препятствие для движения воздушных масс, переносимых западными атлантическими ветрами. Однако вследствие значительно меньших относительных высот и более выровненного характера рельефа на Салаирском кряже наблюдается гораздо меньший контраст количества осадков, выпадающих на западном и восточном склонах. Для западных склонов зарегистрировано от 450 до 550 мм осадков в год, для восточных склонов отмеченное количество осадков снижается до 400 мм. Снеговой покров достигает мощности 80–90 см, иногда до 1,5 м в отдельных понижениях [Шварёв Ю.О., 1963; Ковалёва С.Р. и др., 1974]. Почвы этого орографического региона характеризуются глубокой оподзоленностью, причиной чего служит формирование их на лёссовидных суглинках как материнской породе. Этому способствовало также формирование почв в условиях значительной расчленённости рельефа и влажного резко континентального климата. Глубокооподзоленные почвы черневой тайги Салаира 33 относят к южно-таёжному высотному поясу дерново-подзолистых почв Предалтайской провинции. Почвы обладают кислой реакцией; в составе гумуса преобладают подвижные агрессивные фракции гуминовых и фульвокислот [Ковалёва С.Р. и др., 1974; Лащинский Н.Н., Ревякина М.П., 1991]. В средней и нижней частях пологих склонов Салаирского кряжа (подтаёжный пояс) формируются серые лесные почвы [Ковалёва С.Р. и др., 1974; Трофимов С.С., 1975]. В пределах Салаирского кряжа растительный покров сформирован осиновопихтовыми черневыми лесами, берёзово-осиновыми высокотравными, а также парковыми берёзово-лиственно-сосновыми травянистыми лесами. Леса первого типа расположены главным образом в средней части кряжа, вторые характерны для западного пологого склона. Берёзово-сосновые леса расположены в в крайней восточной части кряжа, где протянулись в виде узкой полосы. Характерной чертой разрастающиеся на Салаирских прогалинах, черневых полянах и лесов являются вырубках обильно многочисленные кустарники[Куминова А.В., 1950; Крылов Г.В., 1961; Вдовин В.В., 1988; Лащинский Н.Н., 2009]. 2.5. Кузнецкая котловина Кузнецкая котловина представляет собой обширный межгорный прогиб площадью около 30 тыс. км2. Она имеет форму неправильного прямоугольника; вытянута с северо-запада на юго-восток на расстояние около 400 км при наибольшей ширине 120 км. С северо-востока Кузнецкая котловина ограничена хребтами Кузнецкого Алатау, с юго-запада – Салаирским кряжем, с юга – Горной Шорией. В северной части котловина через Томь-Колыванскую складчатую зону сливается с Западно-Сибирской равниной. Слагается палеозойскими осадочными породами [Файнер Ю.Б., 1969; Вдовин В.В., 1988]. В целом территория Кузнецкой котловины представляет собой слабо всхолмленную возвышенную эрозионную равнину, слабо наклонённую к Западно-Сибирской равнине и расчленённую густой сетью пологосклонных 34 долин и балок, а также увалами и грядами. Северо-западная часть котловины является наиболее глубокой и ровной, абсолютные высоты здесь составляют в среднем около 200 м [Фомичёв В.Д., 1940; Колобков М.Н., 1956]. Вдоль северной части долины р. Томь отметки дна котловины немногим превышают 100 м над уровнем моря. В южной части преобладает холмисто-увалистый рельеф [Файнер Ю.Б., 1969; Гвоздецкий Н.А., Михайлов Н.И., 1978]. В центральной части котловины расположена так называемая Мелафировая подкова, представляющая собой крутую, открытую к северо-востоку дугу, образованную рядом невысоких горных хребтов, сложенных мезозойскими базальтами (Караканскими, Кайлотскими и Нарыкскими горами, Тарадановским увалом и Салтымаковский хребтом). Максимальные высоты здесь – от 479 м на Караканском до 734 м на Салтымаковском хребте [Воскресенский С.С., 1962]. Абсолютные высоты на территории Кузнецкой котловины возрастают при продвижении на юго-запад, к Кузнецкому Алатау и Горной Шории. Здесь они достигают 400–500 м, в восточной части рельеф приобретает низкогорный характер, а в западной и центральной частях – холмисто-увалистый [Фомичёв В.Д., 1940; Мытарев А.А., 1973; Гвоздецкий Н.А., Михайлов Н.И., 1978; Вдовин В.В., 1988]. Кузнецкая котловина характеризуется обилием ручьёв и рек. Основной водосборный бассейн формируют р. Томь и её притоки Уса, Мрассу, Кондома, Верхняя, Средняя и Нижняя Терси, Тайдон и другие. Вторая крупная река Кузнецкой котловины – Иня – берет начало от рек, стекающих по внутренним склонам гор Мелафировой подковы [Хлонов Ю.П., 1979]. Формирование Кузнецкой котловины завершилось в эпоху верхнепалеозойской складчатости. Большая её часть занята девонскими морскими отложениями, чередующимися с более или менее мощными пластами каменного угля. Залежи каменных углей Кузнецкой котловины характеризуются доступностью для разработки, а сам уголь – высокими качественными показателями, что к настоящему времени привело кзначительным нарушениям почвенного покрова, рельефа и естественных ландшафтов Кузнецкой котловины в 35 процессе интенсивной угледобычи [Михайлов Н.И., 1961; Гвоздецкий Н.А., Михайлов Н.И., 1978]. Климат Кузнецкой котловины достаточно сухой и тёплый летом и холодный зимой. Этот орографический регион отличается несколько более тёплым климатом и большей влажностью по сравнению с лесостепной полосой ЗападноСибирской равнины [Куминова А.В., 1950; Михайлов Н.И., 1961]. Среднегодовая температура изменяется от 0,5 °С до 1,5 °С; морозы зимой могут достигать -50 °С. В отдельные летние дни температура может повышаться до 38 °С и выше. Наибольшее разнообразие климатических условий складывается в центре восточной части Кузнецкой котловины, в районе хребтов Мелафировой подковы [Рутковская Н.В., Попова К.И., 1967; Хлонов Ю.П., 1979; Вдовин В.В., 1988]. Увлажнение территории котловины умеренное, довольно режимное. Более половины годовой нормы осадков приходится на тёплое время года [Михайлов Н.И., 1961]. Самым засушливым является северо-западный участок котловины, который находится в дождевой тени Салаирского кряжа. Этот район характеризуется максимальным получаемым количество тепла, по сравнению с другими территориями Кузнецко-Салаирской провинции. Распределение снежного покрова в котловине неравномерно; в конце зимы его мощность, в зависимости от рельефа, составляет от 25 до 90 см [Трофимов С.С., 1975]. Основная часть почвенного покрова Кузнецкой котловины складывается из четырёх основных типов почв: чернозёмы, лугово-чернозёмные, луговые, серые лесные. Из них зональным типом почв для Кузнецкой котловины являются чернозёмы. В основном встречаются оподзоленные и выщелоченные суглинистые формы чернозёмов с хорошо выраженной зернистой структурой. Они характерны для ровных и низких участков Кузнецкой котловины, которые заняты степными ландшафтами. Чернозёмы солонцеватого и осолоделого типа занимают незначительную площадь и встречаются в районе Присалаирского понижения рельефа. Серые лесные почвы отмечены в лесостепной зоне и, как правило, приурочены к колочным березнякам и осиново-берёзовым лесам. На террасах речных долин и в поймах рек встречаются лугово-чернозёмные а также луговые 36 почвы, которые, кроме того, характерны для депрессионных элементов рельефа и приурочены к лесостепи [Трофимов С.С., 1975; Гвоздецкий Н.А., Михайлов Н.И., 1978]. В Кузнецкой котловине развита лесостепная растительность, представленная берёзово-осиновыми колками, чередующимися с обширными массивами суходольных лугов и переходящими в предгорьях в берёзовые, осиновоберёзовые и смешанные леса. Для пониженных участков Кузнецкой котловины, расположенных в бассейне р. Ини, характерны разнотравно-дерновинно-злаковые степи, почти целиком распаханные. Естественная растительность на данной территории сохранилась лишь на небольших участках [Куминова А.В., 1950; Красноборов И.М. и др., 2001]. 37 ГЛАВА 3. МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ Материалом для выполнения настоящей работы послужили сборы автора, проводившиеся в ходе полевых работ в период с мая по сентябрь 2006–2012 гг. на территории Кузнецко-Салаирской провинции Алтае-Саянской горной страны. Исследованы естественные и антропогенно трансформированные местообитания пчёл сем. Megachilidae в равнинных и горных орографических регионах (Кузнецкой котловине, Горной Шории и Кузнецком Алатау). Помимо собственных сборов, в работе также использованы коллекционные фонды кафедры зоологии и экологии Кемеровского государственного университета, коллекции, любезно предоставленные Н.И. Еремеевой, С.Л. Лузяниным, Д.А. Сидоровым, Д.В. Сущевым, А.В. Д.А. Коршуновым, Ефимовым, Н.С. В.Б. Тепловой, Ильяшенко, С.В. Блиновой, Ю.А. Песенко, Ю.В. Астафуровой. Исследования охватывали типичные биотопы Кузнецко-Салаирской провинции – каменистые и луговые степи, остепнённые, суходольные, лесные и пойменные луга, деградированные луга и участки с рудеральной растительностью; сосновые, сосново-берёзовые, берёзово-пихтовые и осиновопихтовые леса, черневую тайгу. Кроме того, исследованы антропогенные ландшафты разной степени преобразованности. Изучение видового состава производили в 72 точках и экологических особенностей мегахилид Кузнецко-Салаирской провинции (рис. 3.1). Принадлежность точек сбора материала к орографическим регионам определяли по преобладающему типу растительности. Для каждой точки сбора указана её административно-территориальная приуроченность. 38 Рисунок 3.1. Карта-схема Кузнецко-Салаирской провинции с указанием точек сбора материала Примечание. Римскими цифрами обозначены орографические регионы: I – Кузнецкая котловина, II – Кузнецкий Алатау, III – Горная Шория. Чёрными точками обозначены границы орографических регионов. Точки сбора материала 1. Горная Шория Новокузнецкий р-н: 1 – окр. д. Сары-Чумыш; Таштагольский р-н: 2 – окр. п. Шерегеш (4 км В г. Мустаг), 3 – окр. г. Мустаг (г. Зелёная), 4 – Подкатунские скалы, 5 – 6 км ЮЗ п. Мундыбаш, 6 – окр. д. Клёпочный (ср. теч. р. Мундыбаш), 7 – окр. д. Кондома, 8 – окр. п. Усть-Анзас, 9 – 2 км ЮЗ п. Усть-Кабырза, 10 – окр. п. Каз, 11 – пр. б. р. Кондома (52° 53,866' с. ш., 87° 13,460' в. д.), 39 12 – пр. б. р. Кондома (52° 54,529' с. ш., 87° 14,332' в. д.), 13 – пр. б. р. Кондома (52° 50,675' с. ш., 87° 09,756' в. д.), 14 – пр. б. р. Кондома (52° 69,768' с. ш., 87° 04,017' в. д.), 15 – пр. б. р. Кондома (52° 59,767' с. ш., 87° 04,019' в. д.), 16 – 11 км С от устья р. Антроп. 2. Кузнецкий Алатау Крапивинский р-н: 17 – окр. д. Междугорное, 18 – окр. биостанции КемГУ «Ажендарово», 19 – окр. г. Зеленогорск; Новокузнецкий р-н: 20 – 5 км СВ г. Новокузнецк, 21 – 4 км ЮЮВ г. Чёрный Ворон (верх. теч. р. Верх. Терсь), 22 – 8 км ССВ г. Соловей (ср. теч. р. Верх. Терсь), 23 – 5 км ЮВ п. Загадное (ср. теч. р. Верх. Терсь); Междуреченский р-н: 24 – 9 С от ст. Лужба; Тисульский р-н: 25 – окр. оз. Б. Берчикуль, 26 – окр. с. Городок, 27 – 10 км С п. Полуторник, 28 окр. – п. Макаракский; Тяжинский р-н: 29 – окр. с. Преображенка; Чебулинский р-н: 30 – окр. п. Чумай, 31 – окр. д. Кураково; Хакасия, Аскизский р-н: 32 – 11 км СЗ п. Балыксу. 3. Кузнецкая котловина Беловский р-н: 33 – окр. г. Белово, 34 – окр. с. Беково, 35 – окр. п. Краснобродский, 36 – п. Старобачаты, 37 – Караканский хребет, 38 – 4 км ЮВ с. Уроп; Гурьевский р-н: 39 – окр. г. Гурьевск, 40 – с. Сосновка; Кемеровский р-н: 41 – г. Кемерово, 42 – окр. д. Подъяково, 43 – окр. д. Силино, 44 – окр. с. Балахонка, 45 – окр. п. Кедровка, 46 – окр. д. Мазурово, 47 – окр. п. Новостройка, 48 – д. Сухая Речка, 49 – окр. п. Пионер, 50 – д. Мамаевка, 51 – п. Пригородный, 52 – окр. д. Барановка, 53 – окр. д. Осиновка, 54 – окр. п. Мозжуха, 55 – окр. п. Зеленовский, 56 – 5 км СВ д. Шевели, 57 – 5 км ССЗ д. Старочервово, 58 – с. Креково; Ленинск-Кузнецкий р-н: 59 – г. Ленинск-Кузнецкий, 60 –окр. с. Красное; 61 – окр. п. Мирный, 62 – окр. с. Шабаново; Прокопьевский р-н: 63 – г. Прокопьевск, 64 – 4 км СВ г. Прокопьевск, 65 – 5 км СВ д. Большая Талда, 66 – окр. г. Киселёвск; Промышленновский р-н: 67 – пгт Промышленная, 68 – с. Тарасово, 69 –окр. с. Журавлёво; Топкинский р-н: 70 – г. Топки, 71 – окр. д. Малый Корчуган, 72 – окр. д. Цыпино. 40 При сборе материала использовали стандартные методики [Попов В.В., 1950; Фасулати К.К., 1971; Бубнова Т.В., 1988; Дунаев Е.А, 1997]. Отлов проводили безвыборочно, в основном методом индивидуального сбора всех замеченных пчёл пинцетом или энтомологическим сачком с цветков, листьев или поверхности почвы около гнёзд или источников воды, а также отловом пчёл в полёте сачком. При сборе отмечали вид посещаемого растения для определения трофических предпочтений пчёл, погодные условия, точное время отлова, биотоп, в котором отловлена пчела. Собранный материал помещали на ватные слои до дальнейшей камеральной обработки. По окончанию полевого сезона весь собранный материал монтировали на энтомологические булавки. Под наколотый экземпляр помещали этикетку из плотной бумаги с датой, местом и обстоятельствами сбора, фамилией сборщика. Сведения по видовому составу, биотопической приуроченности, трофическим предпочтениям, срокам лёта, полученные при сборе материала, вводили в электронную базу данных (MS Excel). Для определения видовой принадлежности мегахилид использованы ключи из «Определителя насекомых европейской части СССР» [Осычнюк А.З. и др., 1978] и «Определителя насекомых Дальнего Востока России» [Прощалыкин М.Ю., 2007 в]. При работе с отдельными группами мегахилид использовали также монографии и ревизии отечественных и зарубежных авторов, включающие комментарии, определительные таблицы и описания таксонов [Попов В.В., 1932; Michener, C. D., 1948; Erlandsson S., 1955; Noskiewicz J., 1961; Zanden G., 1977, 1986, 1988, 1995, 1997; Eickwort G. C., 1980; Warncke K., 1991, 1992 а, b; Griswold T.L., Michener C.D., 1997; Banaszak, J., Romasenko L., 1998; Přidal A., Tkalců B., 2001; Pachinger B., 2004; Guler Y., Cağatay N., 2006]. Видовая принадлежность более 90 % мегахилид определена автором. Помощь в определении пчёл оказывал доцент каф. зоологии и экологии КемГУ, к.б.н. Сидоров Д.А. Для подтверждения и уточнения видовой идентификации использовали материалы (в том числе типовые) коллекции ЗИН РАН (СанктПетербург). 41 Классификация и номенклатура пчёл сем. Megachilidae приводится в соответствии с системой Ч. Миченера [2000]. Классификация трибы Osmiini дана с учетом точки зрения S. Ungricht et al. [2008] и C. J. Praz et al. [2008], согласно которой род Hoplosmia сводится к рангу подрода р. Osmia. Номенклатура осмиин приводится по работе Ungricht et al., 2008. Экология и биология пчёл сем. Megachilidae (сезон лёта имаго, трофические связи, особенности гнездования и приуроченность к определенному типу местообитания) описаны на основе литературных данных [Erlandsson S., 1955; Монсявичюс, В.С., 1989; Gogala, A., 1991, 1994, 1999; Радченко B.Г., Песенко Ю.А., 1994; Banaszak J., Romasenko L., 1998] и собственных исследований автора. При составлении аннотированного списка видов пчёл сем. Megachilidae КузнецкоСалаирской провинции использованы также данные, полученные сотрудниками каф. зоологии и экологии КемГУ д.б.н., профессором Н.И. Еремеевой и к.б.н. Д.А. Сидоровым [Сидоров Д.А., 2005; Еремеева Н.И., Сидоров Д.А., 2006]. При описании взаимоотношений пчёл с цветковыми растениями использовали общепринятую терминологию. Под трофическими (лектическими [Michener С.D., 2007]) связями понимали связи гнездостроящих видов пчёл с цветковыми растениями, с которых осуществляется сбор пыльцы и / или нектара для выкармливания потомства [Радченко В.Г., Песенко Ю.А., 1994]. По типу трофических связей пчёл разделяли на две группы – полилектичные (polilectic) и олиголектичные (oligolectic) виды [Robertson C., 1928]. Всего на исследуемой территории выделены 4 трофические группы мегахилид – широкие полилекты с широкого круга растений), узкие (самки этих видов собирают пыльцу полилекты (самки собирают корм с немногих видов растений, принадлежащих к разным семействам), широкие олиголекты (для сбора пыльцы посещают растения преимущественно одного семейства), узкие олиголекты (посещают растения исключительно одного семейства, одного или ряда близких родов) [Радченко В.Г., Песенко Ю.А., 1994]. Видовое определение посещаемых пчёлами растений проводили по «Определителю растений Кемеровской области» [Красноборов и др., 2001]. Виды, 42 сложные в определении, идентифицировали по собранным гербарным образцам с помощью специалистов кафедры ботаники Кемеровского государственного университета – зав. кафедрой, к.б.н. Л.Н. Ковригиной и ведущего инженера И.В. Тарасовой; а также сотрудников Кузбасского ботанического сада (ИЭЧ СО РАН) – к.б.н. С.А. Шереметовой и к.б.н. Т.Е. Буко. В результате исследований биотопической приуроченности в КузнецкоСалаирской провинции выделено 4 экологические группы пчёл сем. Megachilidae – луговая, лугово-степная, лугово-лесная и эвритопная. Типы природных биотопов выделены на основании работ А.В. Куминовой [1950], А.Г. Воронцова [1973], Г.Г. Павловой [1980], А.В. Ронгинской [1988]. Помощь в геоботаническом описании участков оказали специалисты кафедры ботаники Кемеровского госуниверситета и сотрудники Кузбасского ботанического сада. Изучение пчёл сем. Megachilidae в антропогенных ландшафтах и городской среде проводили на примере городов Кемерово, Ленинск-Кузнецкий, Киселёвск, Прокопьевск и их окрестностей, а также в Кемеровском, Ленинск-Кузнецком и Прокопьевском районах. Антропогенные местообитания классифицировали по схеме, предложенной И.И. Рахимовым и Р.А. Уленговым [2004]. Биотопы в пределах антропогенных ландшафтов разделили на 3 группы по степени антропогенной трансформации: слабо изменённые, изменённые и преобразованные. К слабо изменённым ландшафтам относили лесопарки, овраги, водоёмы, нетронутые участки луговых и лесных местообитаний, сохранившиеся на территориях, подвергающихся антропогенному воздействию. В г. Кемерово к этой категории относится сосновый бор – сохранённый участок барзасской тайги в Рудничном районе на правом берегу р. Томь. К изменённым (нарушенным) местообитаниям относили парки, кладбища, агроценозы, деградированные луга, сады, рудеральные участки, пустыри, лесополосы; к преобразованным – дороги и их обочины, сельские и городские застройки. 43 При изучении населения мегахилид на урбанизированных территориях использовали классификацию видов насекомых по степени их устойчивости к факторам городской среды, предложенную В.Б. Бейко [1992]. На исследуемой территории выделено 3 группы пчёл-мегахилид – урбофильные (увеличивающие численность на урбанизированных территориях по сравнению с природными местообитаниями), урботолерантные (в равной мере встречаются в природных и антропогенных биотопах, в том числе урбанизированных) и урбофобные (избегают урбанизированных и преобразованных человеком местообитаний). При выделении ареалогических групп и описании типа распространения видов пчёл-мегахилид, обитающих в Кузнецко-Салаирской провинции, рассматривали долготные и широтные составляющие ареала. На основании системы, предложенной К.Б. Городковым [1984], выделяли долготные группы ареалов. Всего по долготному признаку выделено 4 ареалогические группы (голарктическая, транспалеарктическая, западно- центрально-палеарктическая и центрально-восточно-палеарктическая). Широтные группы выделяли по М.Г. Сергееву [1986]. На исследуемой территории по широтному признаку выделены следующие группы – полизональная, бореальная, суббореальная, субаридная. При распределении видов мегахилид по классам обилия использовали методику расчёта, предложенную Ю.А. Песенко [1982 б]. На основании расчётов верхнего и нижнего пределов численного обилия классов были выделены следующие классы обилия: доминанты – с численным обилием от 6,06 % и более от общих сборов мегахилид на исследованной территории; субдоминанты – от 1,45 % до 6,05 %; редкие – от 0,37 % до 1,44 %; очень редкие – с численным обилием 0,36 % и менее. Статистическую обработку данных проводили с использованием стандартных методов математического анализа [Песенко Ю.А., 1982 б]. Для оценки видового разнообразия пчёл сем. Megachilidae КузнецкоСалаирской провинции применяли индекс Шеннона Ish' и показатель PIE' (probability of interspecific encounter – вероятность межвидовых встреч). 44 Индекс Шеннона вычисляли по формуле Ish' = - Σpi*log(pi), где pi – доля i-го вида в выборке. Обычно значения индекса Шеннона лежат в пределах от 1,5 до 3,5. Чем он больше, тем больше видовое разнообразие сообщества [Лебедева Н.В., 2002]. Индекс разнообразия PIE' выводится из меры «концентрации» Симпсона λ' по формуле 1 – λ'. Эта величина изменяется в пределах 0 ≤ PIE' ≤ 1 и является удобным показателем видового разнообразия (показывает, насколько оно приближено к максимальному) [Песенко Ю.А., 1982 б; Розенберг Г.С., 2007]. При сравнении фаун мегахилид орографических регионов Кузнецко- Салаирской провинции использованы следующие показатели: индекс Шеннона и вероятность межвидовых встреч PIE', которые признаются лучшими при сравнительной оценке фаун [Андреев А.В., 2002]. Кроме того, использован индекс полидоминантности Вильямса (1 / λ'), который многие авторы считают лучшей мерой оценки типа распределения видовых обилий, когда число видов превышает 10 [Розенберг Г.С., 2007]. Индекс Шеннона использовали также для сравнения видового разнообразия пчёл сем. Megachilidae в типовых биотопах орографических регионов КузнецкоСалаирской провинции. В качестве меры фаунистического сходства при сравнении фаун пчёл сем. Megachilidae разных орографических регионов Кузнецко-Салаирской провинции взят индекс Чекановского-Съёренсена, чувствительный к объему выборок [Песенко Ю.А., 1982 б]: Ksc = 2c a+b , 45 где а, b – число видов в разных фаунах / выделах; с – число общих видов для двух фаун / выделов. Пределы коэффициента – от 0 до 1, Кsc =1 означает полное сходство фаун / выделов (абсолютное совпадение списков), Кsc = 0 означает, что они не имеют ни одного общего вида. Для построения диаграмм использованы возможности программы MS Excel. Общий объём обработанного материала, собранного на территории КузнецкоСалаирской провинции, составил 1507 экземпляров пчёл сем. Megachilidae. Представленные в работе результаты получены соискателем лично. Достоверность результатов проведённых исследований обусловлена чётким планированием работы, использованием большого массива источников информации, применением традиционных методических подходов, классических методов экологии, а также статистической обработкой экспериментальных данных и анализом полученных результатов. По материалам исследования опубликовано 6 печатных работ; из них одна выполнена в соавторстве с сотрудниками кафедры зоологиии экологии КемГУ Н.И. Еремеевой, С. В. Блиновой, С. Л. Лузяниным, Д. А. Сидоровым [Еремеева Н.И. и др., 2011]. 46 ГЛАВА 4. ВИДОВОЕ РАЗНООБРАЗИЕ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE КУЗНЕЦКО-САЛАИРСКОЙ ПРОВИНЦИИ 4.1. Аннотированный список видов пчёл сем. Megachilidae КузнецкоСалаирской провинции Аннотированный список включает 49 видов пчёл сем. Megachilidae, относящихся к 12 родам, 5 трибам, 1 подсемейству. Для каждого вида указаны экологические и биологические особенности, данные по распространению, а также материал, собранный на территории Кузнецко-Салаирской провинции. Экология и биология пчёл сем. Megachilidae (сезон лёта, трофические связи, особенности гнездования и приуроченность к определённому типу местообитания) описаны на основе литературных данных и собственных исследований автора. Территории распространения мегахилид в пределах биогеографических регионов перечислены с запада на восток; сначала указано распространение в России, потом за её пределами. Наиболее подробные данные по распространению видов мегахилид даны для территории Сибири. В описании материала, собранного на территории Кузнецко-Салаирской привинции, названия физико-географических регионов и административных районов расположены административного в алфавитном района точки порядке. сбора В пределах материала каждого перечислены хронологическом порядке согласно дате сбора. В аннотированном списке использованы следующие сокращения: PА – Палеарктика КСП – Кузнецко-Салаирская провинция NА – Неарктика ГШ – Горная Шория NT – Неотропическая область КА – Кузнецкий Алатау AU – Австралийская область КК – Кузнецкая котловина в 47 обл. – область, г. – город, п. – посёлок, д. – деревня, пгт – посёлок городского типа, окр. – окрестности, верх. – верхний, ср. – средний, нижн. – нижний, теч. – течение. С – северный /севернее СВ – северо-восток Ю – южный / южнее СЗ – северо-запад В – восточный / восточнее ЮВ – юго-восток З – западный / западнее ЮЗ – юго-запад Коллекторы: НЕ – Н.И. Еремеева, ДС – Д.А. Сидоров, СЯ – С.Н. Яковлева, СЛ – С.Л. Лузянин, НТ – Н.С. Теплова, СБ – С.В. Блинова, ДВ – Д.В. Сущёв, ДЕ – Д.А. Ефимов, ВИ – В.Б. Ильяшенко, АК – А.В. Коршунов. Виды мегахилид, впервые указанные для территории Западной Сибири, отмечены символом *, для Кузнецко-Салаирской провинции – **. Подсемейство MEGACHILINAE Триба LITHURGINI Lithurgus Latreille, 1825 Пчёлы от мелкого до крупного размера (9–19 мм). Чёрные, коренастые. Покрыты серовато-белыми волосками. Распространены повсеместно. Предпочитают зоны с тропическим, тёплым и умеренным климатом. Летние формы [Michener С., 1983, 2007]. В Палеарктике род представлен 1 подродом Lithurgus, включающим 7 видов. Гнездятся в сухой древесине и деревянных сооружениях. Имеют тесные трофические связи с растениями сем. Asteraceae [Zanden, G., 1977, 1986; Осычнюк А.З. и др., 1978; Banaszak J. et al., 2001; Pachinger, B., 2004]. В Кузнецко-Салаирской провинции встречается 1 вид. 48 Lithurgus (Lithurgus) fuscipennis Lepeletier, 1841 Экология и биология Сезон лёта: июль – август. Трофические связи: является олиголектом на Asteraceae. Предпочитает посещать растения родов Carduus, Centaurea, Cirsium [Песенко Ю.А., 1971; Banaszak J., Romasenko L., 1998; Gogala A., 1999]. На территории КСП отмечено посещение цветков Arctium tomentosum. Гнездовое поведение: гнездится в мягкой гниющей древесине. Гнёзда линейноветвистые, содержат от 2 до 8 ячеек [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006]. Местообитание: в КСП встречается на остепнённых и каменистых участках, а также на деградированных разнотравно-злаковых лугах и агроценозах. Отмечена приуроченность к селитебным зонам [Еремеева Н.И., Сидоров Д.А., 2006]. Распространение Западно-центрально-палеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Омская обл. [Лавров С.Д., 1927], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Алтайский край [Данилов Ю.Н., 2006]; Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]). Южная, Центральная и Восточная Европа, Северная Африка, Турция, Кавказ, Средняя Азия, Западный Казахстан [Banaszak J., Romasenko L., 1998]. Материал ГШ: Таштагольский р-н, 11 км С от устья р. Антроп, 31.VII.2011 – 3♀, 2♂ (ДС). КК: Кемеровский р-н, г. Кемерово, 17.VII.1996 – 4♀♀ (НЕ), окр. п. Кедровка, 08.VIII.2003 – 3♀♀ (ДС); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 16.VII.2012 – 5♀♀, 2♂♂, 17.VII.2012 – 3♂♂, 20.VII.2012 – 2♀♀, 6♂♂, 22.VII.2012 – 2♀♀, 28.VII.2012 – 2♀♀ (СЯ), 2 км ЮВ с. Красное, 12.VII.2004 – 1♀, 4♂♂, 13.VII.2004 – 5♀♀, 2♂♂, 14.VII.2004 – 5♂♂ (ДС). Прокопьевский р-н, 49 г. Киселёвск, 17.VII.2010 – 4♂♂ (ДС); Топкинский р-н, г. Топки, 15.VIII.2012 – 6♂♂ (СЯ). Точки сбора: указаны на рис. 4.1.1. Рисунок 4.1.1. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции 50 Триба OSMIINI Chelostoma Latreille, 1809 Мелкие и средние пчёлы (5–15 мм). Тело стройное, цилиндрическое, чёрное, покрыто короткими бледными волосками. Брюшная щётка самок имеет беловатую окраску [Осычнюк А.З. и др., 1978]. Весенне-летние и летние формы. Большинство является олиголектами; палеарктические виды трофически тесно связаны с растениями, принадлежащими к родам Campanula и Ranunculus. Гнёзда размещают в готовых полостях в сухой древесине, иногда в полых стеблях растений [Banaszak J. et al., 2001; Прощалыкин М.Ю., 2007 в]. Род насчитывает 55 видов из 6 подродов. Представители распространены по всей Голарктике. На территории Палеарктики встречаются 43 вида [Eickwort G., 1980; Zanden, G., 1988; Прощалыкин М.Ю., 2012]. В Западной Сибири отмечено 2 вида. На территории Кузнецко-Салаирской провинции обнаружен 1 вид. Chelostoma (Gyrodromella) rapunculi (Lepeletier, 1841) Экология и биология Сезон лёта: июнь – июль. Трофические связи: является олиголектом на Campanula [Banaszak J., Romasenko L., 1998; Gogala F., 1999; Guler Y., Cağatay N., 2006]. В КСП посещает Campanula altaica, C. glomerata, C. sibirica и C. trachelium. Отмечено также посещение Convolvulus arvensis (Convolvulaceae), Veronica longifolia (Scrophulariaceae), Hedysarum gmelinii и Trifolium repens (Fabaceae). Гнездовое поведение: гнездятся в полых стеблях растений, а также готовых отверстиях в стволах деревьев и стенах деревянных построек. Гнёзда линейные, имеют по 3–7 ячеек, отделённых друг от друга перегородками из смеси земли и мелких камешков [Banaszak J., Romasenko L., 1998]. Местообитание: на территории КСП встречается на суходольных разнотравно-злаковых лугах, а также на залежах. остепнённых и 51 Распространение Голарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Республика Алтай, Алтайский край [Прощалыкин М.Ю., 2013 б], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Красноярский край, Иркутская обл. [Прощалыкин М.Ю., 2013 б], Бурятия [Proshchalykin М., 2007]); Дальний Восток России [Proshchalykin М., 2004]. Европа, Северная Африка, Турция, Кавказ, Казахстан, Туркменистан. NA. Северо-Восток США [Eickwort G., 1980; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005; Guler Y., Cağatay N., 2006]. Материал ГШ: Таштагольский р-н, Подкатунские скалы, 07.VII.2005 – 5♂♂ (ДС), 2 км ЮЗ п. Усть-Кабырза, 04.VII.2008 – 2♂♂ (СЯ), 14.VII.2008 – 3♀♀, 2♂♂ (ДС), окр. п. Каз, 29.VI.2010 – 3♂♂ (ДС). КА: Крапивинский р-н, окр. биостанции КемГУ «Ажендарово», 10.VI.2008 – 1♀ (ДС); Новокузнецкий р-н, 8 км ССВ г. Соловей (ср. теч. р. Верх. Терсь), 26.VII.1998 – 3♀♀ (НЕ); Тисульский р-н, 10 км С п. Полуторник, 07.VII.2009 – 6♀♀ (ДС); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 09.VII.2004 – 1♂ (НЕ), 11.VII.2009 – 3♂♂ (ДС). КК: Кемеровский р-н, г. Кемерово, 30.VI.1999 – 1♀ (НЕ), с. Креково, 30.VI.2012 – 1♀ (ДЕ); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 12.VII.2006 – 1♀, 20.VII.2012 – 20♀♀, 21.VII.2012 – 12♂♂, 25.VII.2012 – 6♂♂ (СЯ), окр. г. Ленинск-Кузнецкий, 15.VII.2009 – 1♂ (СЯ); Промышленновский р-н, 6 км СЗ с. Журавлёво, 23.VI.2000 – 1♀ (НЕ), 4 км СЗ с. Журавлёво, 30.VI.2006 – 4♀♀ (СЯ), 02.VII.2006 – 1♀ (ДС), 06.VII.2011 – 1♂ (СЯ); 08.VII.2011 – 1♂ (ДС). Точки сбора: указаны на рис. 4.1.1. 52 Hoplitis Klug, 1807 Мелкие и средние пчёлы, иногда крупные (6–18 мм). Стройные, реже коренастые, чёрные, изредка с красными пятнами. Опушение бледное. Весенне-летние или летние формы с продолжительным сезоном лёта. Полилектичные или олиголектичные виды, более или менее тесно связаны с Fabaceae, Lamiaceae, а также с растениями родов Campanula и Echium. Гнездовое поведение разнообразно. Большинство видов устраивает гнёзда в готовых полостях в сухой древесине, пустых раковинах брюхоногих моллюсков и полых стеблях растений. Несколько видов сооружают свои гнёзда на поверхности различных субстратов (так называемые пчёлы-каменщицы) [Осычнюк А.З. и др., 1978; Banaszak J. et al., 2001]. Род включает около 330 видов из 27 подродов. Представители данного рода широко распространены в Голарктическом регионе. В Палеарктике отмечено 282 вида 20 подродов [Прощалыкин М.Ю., 2007 в, 2012]. На территории Западной Сибири отмечено 9 видов. В Кузнецко-Салаирской провинции встречаются 6 видов. **Hoplitis (Alcidamea) acuticornis (Dufour et Perris, 1840) Экология и биология Сезон лёта: июнь – июль. Трофические связи: является олиголектом на Fabaceae [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. В КСП посещает виды рода Lathyrus (L. gmelinii, L. pisiformis, L. sylvestris), отмечен также на Oxytropis campanulata и Vicia cracca. Гнездовое поведение: самки располагают гнёзда в сухих стеблях, проделывая ходы в мягкой сердцевине. Гнёзда линейного типа. Полости ячеек отделены друг от друга поперечными перегородками, изготовленными из спрессованных частиц сердцевины стебля [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Местообитание: в КСП обитает на суходольных разнотравно-злаковых лугах. Распространение Западно-центрально-палеарктический полизональный вид. 53 PA. Россия: европейская часть [Лыков В.А., 2008]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Алтайский край [Данилов Ю.Н., 2006], Кемеровская обл. [Яковлева С.Н., 2011]). Канарские о-ва, Южная Европа, Центральная и Восточная Европа (южные регионы), Турция, Кавказ, Западный Казахстан [Warncke K., 1991; Banaszak J., Romasenko L., 1998]. Материал КА: Крапивинский р-н, 3 км Ю биостанции КемГУ «Ажендарово», 02.VII.2009 – 2♀ (ДС). КК: Беловский р-н, Караканский хребет, 12.VI.2006 – 4♀♀ (СЯ), 02.VI.2008 – 8♀♀, 03.VI.2008 – 2♀♀ (ДС); Кемеровский р-н, г. Кемерово, 07.VI.2006 – 1♀ (СЯ), 5 км СВ д. Шевели, 16.VII.2009 – 4♀♀ (ДС); Прокопьевский р-н, 4 км СВ г. Прокопьевск, 23.VI.2006 – 1♂; Промышленновский р-н, 4 км СЗ с. Журавлёво, 28.VI.2006 – 2♀ (ДС). Точки сбора: указаны на рис. 4.1.1. Hoplitis (Alcidamea) claviventris (Thomson, 1872) Экология и биология Сезон лёта: июнь – август. Трофические связи: полилект; предпочитает посещать цветки растений сем. Fabaceae [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999]. В КСП отмечено посещение Astragalus daticus, Trifolium repens, виды рода Vicia. Из других семейств растений отмечен на Asteraceae (Leontodon autumnalis, Tragopogon orientalis), Viburnaceae (Viburnum opulus) и Geraniaceae (Geranium pratense). Гнездовое поведение: гнёзда, как правило, располагают в сердцевине сухих стеблей и ветвей кустарников, проделывая в ней тоннели. Иногда размещают гнёзда и в готовых отверстиях в сухой древесине. Тип гнезда – линейный. Ячеек от 4 до 8, они отделены друг от друга поперечными перегородками из лиственной мастики (пережёванные листья) [Banaszak J., Romasenko L., 1998]. 54 Местообитание: на территории КСП вид встречается на суходольных разнотравно-злаковых лугах, а также на опушках сосново-берёзовых и сосновоосиновых лесов. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]). Европа (до 66° с. ш.), Кавказ, Казахстан, Монголия [Warncke K., 1991; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998;]. Материал ГШ: Таштагольский р-н, окр. п. Каз, 06.VII.2010 – 1♀ (ДС), пр. б. р. Кондома (52° 69,768' с. ш., 87° 04,017' в. д.), 12.VI.2011 – 2♂ (ДС). КА: Крапивинский р-н, окр. биостанции КемГУ «Ажендарово», 07.VIII.2006 – 1♂ (СЯ); Тисульский р-н, 10 км С п. Полуторник, 02.VII.2009 – 3♀♀, 04.VII.2009 – 2♀♀, 10.VII.2009 – 2♀♀ (ДС); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 03.VII.2004 – 1♀, 2♂♂ (НЕ). КК: Кемеровский р-н, г. Кемерово, 18.VI.1998 – 3♀♀, 5♂♂, 01.VII.1999 – 1♀, 17.VI.2004 – 2♂♂ (НЕ); Прокопьевский р-н, ЮВ окр. г. Киселёвск, 23.VI.2006 – 1♀ (ДС); Промышленновский р-н, 4 км СЗ с. Журавлёво, 14.VI.1999 – 1♂, 30.VI.2006 – 4♀♀, 03.VII.2006 – 2♀♀, 04.VII.2006 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.2. Hoplitis (Alcidamea) leucomelana (Kirby, 1802) Экология и биология Сезон лёта: июнь – август. Трофические связи: полилект, предпочитает Fabaceae (Astragalus, Lotus, Medicago, Trifolium), нередко также посещает цветки губоцветных [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J.et al., 2001]. 55 Рисунок 4.1.2. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции На территории КСП посещает растений из семейств Crassulaceae (Sedum hybridum), Fabaceae (Lathyrus sylvestris, Trifolium repens, Vicia sylvatica), Rosaceae (Filipendula ulmaria, Rubus caesius), Lamiaceae (Phlomis tuberosa, Prunella vulgaris), Asteraceae (Aster alpines). Гнездовое поведение: самки устраивают гнёзда линейного типа в сердцевине сухих стеблей и ветвей кустарников, в которых самостоятельно проделывают ходы. Гнездо содержит от 2 до 16 ячеек, разделённых перегородками из лиственной мастики; ход от последней ячейки до входного отверстия обычно 56 заполнен маленькими кусочками сердцевины стебля, мелкими камешками и высохшими частями насекомых [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Местообитание: в КСП отловлен на суходольных злаково-разнотравных лугах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Алтай [Proshchalykin M., 2007], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Тыва, Хакасия, Красноярский край [Прощалыкин М.Ю., 2013 а, в], Иркутская обл., Бурятия, Забайкальский край, Якутия [Романькова Т.Г., 1995; Давыдова Н.Г., Песенко Ю.А., 2002; Proshchalykin M., 2007]); Дальний Восток России [Proshchalykin M., 2004]. Европа (до 57° с. ш.), Северная Африка, Турция, Средняя Азия [Warncke K., 1991; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, окр. п. Тельбес, 02.VII.2003 – 6♀♀ (ДС), пр. б. р. Кондома (52° 54,529' с. ш., 87° 14,332' в. д.), 08.VI.2011 – 3♀♀ (ДС). КА: Крапивинский р-н, 3 км Ю биостанции КемГУ «Ажендарово», 02.VIII.2009 – 1♀ (ДС); Тисульский р-н, 10 км С п. Полуторник, 04.VII.2009 – 4♂♂ (ДС); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу,08.VII.2004 – 1♀, 2♂♂, 09.VII.2004 – 5♀♀, 2♂♂ (ДС). КК: Беловский р-н, Караканский хребет (северная часть), 03.VI.2007 –1♀; Кемеровский р-н, 5 км СВ д. Шевели, 16.VII.2009 – 2♀♀, 1♂ (ДС); Прокопьевский р-н, ЮВ окр. г. Киселёвск, 23.VI.2006 – 3♀♀ (ДС); Промышленновский р-н, 4 км СЗ с. Журавлёво, 03.VII.2006 – 1♀ (СЯ). Точки сбора: указаны на рис. 4.1.2. 57 Hoplitis (Alcidamea) mitis (Nylander, 1852) Экология и биология Сезон лёта: июнь – июль. Моновольтинный [Gogala A., 1999]. Трофические связи: олиголектичный вид, трофически связан с растениями рода Campanula [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001]. В КСП отмечено посещение видов Campanula altaica и C. sibirica. Кроме того, на исследуемой территории представители данного вида пчёл посещают цветки Aster alpinus (Asteraceae) и Dracocephalum nutans (Lamiaceae). Гнездовое поведение: гнездятся в готовых полостях в почве, редко в отверстиях в отмершей древесине. Стенки ячеек сооружают из кусочков листьев, склеенных лиственной мастикой [Banaszak J., Romasenko L., 1998]. Местообитание: в КСП приурочен главным образом к остепнённым разнотравно-злаковым лугам и степным участкам, изредка встречается на суходольных лугах. Распространение Транспалеарктический полизональный вид. PA. Россия: Западная Сибирь (Кемеровская обл.) [Еремеева Н.И., Сидоров Д.А., 2006]; Восточная Сибирь (Красноярский край [Прощалыкин М.Ю., 2013 б], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]). Юго-Западная, Центральная и Восточная Европа, Средняя Азия [Warncke K., 1991; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КК: Беловский р-н, Караканский хребет, 12.VI.2006 – 3♀♀, 9♂♂ (ДС, СЯ); Кемеровский р-н, г. Кемерово, 10.VI.2006 – 4♀♀ (СЯ); Промышленновский р-н, 6 км СЗ с. Журавлёво, 26.VI.2002 – 1♀ (НЕ), 4 км СЗ с. Журавлёво, 27.VI.2006 – 2♀♀ (ДС), 28.VI.2006 – 4♀♀ (ДС), 29.VI.2006 – 7♀♀ (СЯ), 30.VI.2006 – 2♀♀ (ДС), 02.VII.2006 – 1♀ (СЯ), 04.VII.2006 – 1♀ (СЯ), 08.VII.2006 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.2. 58 Hoplitis (Alcidamea) tridentata (Dufour et Perris, 1840) Экология и биология Сезон лёта: июнь – август. Моновольтинный [Gogala A., 1999]. Трофические связи: олиголект на Fabaceae [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001; Matache I., Ban C., 2006]. Из указанного семейства растений в КСП предочитает посещать Vicia amoena, V. cracca, V. sylvatica, Lathyrus pratensis, L. sylvestris, L. tuberosus, Trifolium repens. Отмечен также на Nonea rossica (Boraginaceae). Гнездовое поведение: гнёзда линейного типа. Самки сооружают их в сердцевине сухих стеблей кустарников, в которой самостоятельно проделывают ходы. Ячейки разделены перегородками из лиственной мастики [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006]. Местообитание: на территории КСП обнаружен на суходольных разнотравнозлаковых лугах, остепнённых участках, отмечен также на пойменном лугу. Распространение Западно-центрально-палеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]). Южная, Центральная и Восточная Европа, Турция, Кавказ, Средняя Азия [Warncke K., 1991; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, 6 км ЮЗ п. Мундыбаш, 03.VII.2005 – 1♀, 3♂♂ (ДС). КА: Крапивинский р-н, территория биостанции КемГУ «Ажендарово», 29.VI. 2002 – 2♀♀, 3 км Ю биостанции «Ажендарово», 02.VIII.2009 – 1♀ (ДС). КК: Беловский р-н, 4 км ЮВ с. Уроп, 23.VII.2006 – 7♀♀ (ДС); Кемеровский р-н, г. Кемерово, 12.VII.1996 – 1♀ (НЕ), окр. д. Подъяково, 10.VII.1998 – 3♀♀ (НТ); Ленинск-Кузнецкий р-н, окр. с. Шабаново, 14.VII.2004 – 1♂ (НЕ), г. Ленинск-Кузнецкий, 29.VII.2006 – 6♀♀, 17.VII.2012 – 3♀♀, 18.VII.2012 – 1♀, 59 30.VII.2012 – 2♀♀ (СЯ); Прокопьевский р-н, 5 км СВ д. Большая Талда, 18.VII.2004 – 4♀♀ (ДС), ЮВ окр. г. Киселёвск, 23.VI.2006 – 1♀, 3♂♂ (ДС); Промышленновский р-н, 4 км СЗ с. Журавлёво, 29.VI.2006 – 2♂♂ (СЯ). Точки сбора: указаны на рис. 4.1.3. Рисунок 4.1.3. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции 60 Hoplitis (Monumetha) tuberculata (Nylander, 1848) Экология и биология Сезон лёта: июнь – июль. Трофические связи: вид полилектичен, чаще посещает цветки Rosaceae [Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Шелоховская Л.В. и др., 2011]. Гнездовое поведение: неизвестно. Местообитание: в КСП вид обнаружен в лесных биотопах – сосново-берёзовых лесах и на лесных лугах. Распространение Транспалеарктический полизональный вид. PA. Россиия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Ханты-Мансийский автономный округ [T.V. Levchenko, K.P. Tomkovich, 2014], Омская обл. [Лавров С.Д., 1927], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Алтай [Banaszak J., Romasenko L., 1998; Proshchalykin M., 2007], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Иркутская обл., Бурятия, Забайкальский край [Proshchalykin M., 2007], Якутия [Романькова Т.Г., 1995]); Дальний Восток России [Proshchalykin M., 2004]. Северная Европа (между 68° и 56° с. ш.), Центральная и Восточная Европа, Монголия [Warncke K., 1991; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, окр. п. Тельбес, 06.VII.2003 – 2♀♀ (ДС). КА: Новокузнецкий р-н, 5 км ЮВ п. Загадное (ср. теч. р. Верх. Терсь), 21.VI.2008 – 3♀♀ (ДС); Тисульский р-н, окр. с. Городок, 29.VI.1999 –1♀ (НЕ), окр. п. Макаракский, 06.VII.2010 –3♀♀ (СЯ). Точки сбора: указаны на рис. 4.1.3. 61 Osmia Panzer, 1806 Полностью чёрные, иногда с металлическим блеском пчёлы. Средней величины (7–15 мм), обильно опушённые. Самки обычно имеют бледную или чёрную брюшную щётку. Ранневесенние, весенне-летние и летние формы, полилекты или олиголекты, имеющие более или менее тесные трофические связи с растениями семейств Fabaceae, Asteraceae и Lamiaceae. Гнездовое поведение значительно различается у разных видов. Большинство из них размещает гнёзда в готовых отверстиях в мёртвой сухой древесине, стеблях и ветвях растений, пустых раковинах моллюсков и галлах, в трещинах камней, бетонных и кирпичных стен, иногда в почве [Осычнюк и др., 1978; Иванов С.П., Фатерыга А.В., 2006]. Представители рода Osmia широко распространены по всей Голарктике, всего род насчитывает около 320 видов из 19 подродов. Из них 188 видов отмечено на территории Палеарктики [Banaszak J. et al., 2001; Прощалыкин М.Ю., 2012]. На территории Кузнецко-Салаирской провинции отмечено 6 видов. *Osmia (Helicosmia) leaiana (Kirby, 1802) Экология и биология Сезон лёта: июнь. Трофические связи: олиголект на растениях сем. Asteraceae (Centaurea, Cirsium, Taraxacum, Leontodon) [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Гнездовое поведение: самки размещают гнёзда в готовых полостях – отверстиях, проделанных насекомыми в мёртвой древесине, в полых стеблях и отверстиях в каменных сооружениях. Гнёзда линейные, ячейки отделены друг от друга перегородками, изготовленными из лиственной мастики [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001]. Местообитание: в КСП отмечен на галечной террасе в зарослях прибрежной растительности. Распространение Транспалеарктический полизональный вид. 62 PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл. [Яковлева С.Н., 2013]; Восточная Сибирь (Иркутская обл., Забайкальский край) [Romankova T., 2003], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]; Дальний Восток России [Proshchalykin M., 2004]. Европа, Турция, Кавказ, Казахстан, Таджикистан [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Прощалыкин М.Ю., 2012]. Материал ГШ: Таштагольский р-н, пр. б. р. Кондома (52° 50,675' с. ш., 87° 09,756' в. д.), 10.VI.2011 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.3. Osmia (Hoplosmia) spinulosa (Kirby, 1802) Экология и биология Сезон лёта: июль – август. Трофические связи: олиголект на растениях сем. Asteraceae (посещает растения родов Hieracium, Centaurea, Inula, Anthemis, Carduus, Cichorium, Tragopogon) [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001]. В КСП отловлен на цветках Inula salicina (Asteraceae), Sedum hybridum (Crassulaceae), Ranunculus repens (Ranunculaceae). Гнездовое поведение: самки размещают гнёзда в пустых раковинах брюхоногих моллюсков р. Helix. Тип гнезда – линейный; ячейки отделены друг от друга поперечными перегородками [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006]. Местообитание: в КСП обитает на суходольных и лесных разнотравнозлаковых лугах. Распространение Западно-центрально-палеарктический полизональный вид. PA. Россия: европейская часть [Лыков В.А., 2008]; Западная Сибирь (Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Алтайский край, респ. Алтай [Прощалыкин М.Ю., 2013 б], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 63 2006]); Восточная Сибирь (Хакасия) [Прощалыкин М.Ю., 2013 б], Южная, Центральная и Восточная Европа, Кавказ, Средняя Азия, [Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, окр. п. Тельбес, 02.VII.2003 – 2♀♀ (ДС), Подкатунские скалы, 06.VII.2003 – 5♀♀ (НЕ), 13.VII.2005 – 10♀♀ (ДС), 3 км ЮЗ п. Усть-Кабырза, 03.VII.2008 – 6♀♀ (НЕ, ДС), 05.VII.2008 – 4♀♀ (СЯ), 3♂♂, 14.VII.2008 – 8♀♀ (ДС). КК: Топкинский р-н, окр. д. Малый Корчуган, 11.VIII.2003 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.4. *Osmia (Melanosmia) parietina Curtus, 1828 Экология и биология Сезон лёта: июнь – июль. Трофические связи: полилектичен, предпочитает Fabaceae и Asteraceae [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001]. На территории КСП отмечено посещение растений из сем. Crassulaceae (Sedum hybridum), Fabaceae (Trifolium repens, Vicia cracca), Rosaceae (Rubus idaeus, Potentilla chrysantha), Geraniaceae (Geranium pratense). Гнездовое поведение: самки устраивают гнёзда в готовых отверстиях в сухой древесине [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Местообитание: на территории европейской части России приурочен к лесной зоне [Осычнюк и др., 1978]; в КСП данный вид отмечен на суходольных и лесных разнотравно-злаковых лугах, встречается также в осиново-пихтовых лесах. Распространение Транспалеарктический полизональный вид. 64 Рисунок 4.1.4. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Ханты-Мансийский автономный округ [T.V. Levchenko, K.P. Tomkovich, 2014], Кемеровская обл. [Яковлева С.Н., 2012]); Дальний Восток России [Proshchalykin M., Kupianskaya A., 2005]. Северная, Центральная и Восточная Европа, Южная Европа [Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, 2 км ЮЗ п. Усть-Кабырза, 03.VII.2008 – 1♀ (ДС), 4.VII.2008 – 2♀♀ (ДС), 12.VII.2008 – 1♀ (СЯ); пр. б. р. Кондома (52° 59,768' с. ш., 65 87° 04,017' в. д.), 12.VI.2011 – 8♀♀ (ДС). КА: Новокузнецкий р-н, 5 км ЮВ п. Загадное (ср. теч. р. Верх. Терсь), 21.VI.2008 – 5♀♀, 22.VI.2008 – 4♀♀, 24.VI.2008 – 2♀♀, 26.VI.2008 – 3♀♀ (ДС); Тисульский р-н, 10 км С п. Полуторник, 09.VII.2009 – 1♀ (ДС); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 08.VII.2004 – 2♀♀ (ДС). КК: Беловский р-н, Караканский хребет (сев. отрог), 02.VI.2008 – 1♀ (ДС); Промышленновкий р-н, 4 км СЗ с. Журавлёво, 03.VII.2006 – 4♀♀ (ДС). Точки сбора: указаны на рис. 4.1.4. **Osmia (Melanosmia) pilicornis Smith, 1846 Экология и биология Сезон лёта: июнь – июль. Трофические связи: вид полилектичен, посещает цветки Pulmonaria, Lotus [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001]. В КСП предпочитает Asteraceae (Anthemis subtinctoria), Fabaceae (Astragalus daticus, Lathyrus pisiformis, Oxytropis campanulata, Vicia amoena, V. megalotropis, V. multicaulis), Lamiaceae (Dracocephalum nutans, Phlomis tuberosa). Гнездовое поведение: самки размещают гнёзда в пустых раковинах брюхоногих моллюсков [Banaszak J., Romasenko L., 1998] и в древесине [Gogala A., 1999]. Местообитание: вид приурочен к лесным участкам [Осычнюк А.З. и др., 1978]. В КСП обитает на остепнённых разнотравно-злаковых лугах, суходольных лугах, отмечен также в сосново-берёзовых лесах. Распространение Западно-центрально-палеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Кемеровская обл. [Яковлева С.Н., 2011]). Северная, Центральная и Восточная Европа [Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. 66 Материал ГШ: Таштагольский р-н, окр. п. Каз, 04. VII. 2010 – 5♀♀ (ДС), 06.VII.2010 – 1♀ (СЯ). КК: Беловский р-н, Ю часть Караканского хребта, 11. VI.2006 –2♀♀, 12.VI.2006 – 3♀♀ (ДС), 4 км ЮВ с. Уроп, 23.VII.2006 – 3♀♀ (ДС), С отрог Караканского хребта, 01.VI.2008 – 2♀♀, 02.VI.2008 – 5♀♀, 03.VI.2008 – 5♀♀; Кемеровский р-н, окр. д. Подъяково, 08.VII.2007 – 1♀; Прокопьевский р-н, г. Прокопьевск, 16.VII.2007 – 4♀♀ (ДС); Промышленновский р-н, 4 км СЗ с. Журавлёво, 30.VI.2006 – 4♀♀ (ДС), 01.VII.2006 – 2♀♀ (СЯ), 29.V.2009 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.4. **Osmia (Melanosmia) uncinata Gerstaecker, 1869 Экология и биология Сезон лёта: май – июнь. Трофические связи: вид полилектичен, предпочитает посещать цветки растерий сем. Fabaceae, Asteraceae и Lamiaceae [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. В КСП отмечен на Chorispora sibirica (Brassicaceae). Гнездовое поведение: самки размещают гнёзда в готовых полостях, в щелях коры деревьев, используют проделанные другими насекомыми отверстия в мёртвой сухой древесине [Stoeckhert F., 1933; Banaszak J., Romasenko L., 1998; Banaszak et al., 2001]. Местообитание: вид приурочен к лесной зоне [Осычнюк А.З. и др., 1978]. В КСП обнаружен на опушке соснового бора. Распространение Транспалеарктический бореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Ханты-Мансийский автономный округ [T.V. Levchenko, K.P. Tomkovich, 2014]), Восточная Сибирь (Иркутская обл., Забайкальский край [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России 67 [Романькова Т.Г., 1984]. Северная, Восточная и Центральная Европа, Восточная Азия [Banaszak J., Romasenko L., 1998]. Материал КК: Кемеровский р-н, г. Кемерово, сосновый бор, 25.V.2006 – 2♀♀ (СЯ). Точки сбора: указаны на рис. 4.1.5. Рисунок 4.1.5. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции 68 Osmia (Neosmia) bicolor (Schrank, 1781) Экология и биология Сезон лёта: май – июль. Трофические связи: полилект [Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. В КСП отмечен на растениях из семейств Boraginaceae (Pulmonaria mollis), Crassulaceae (Sedum hybridum), Liliaceae (Gagea granulosa), Ranunculaceae (Pulsatilla patens), Salicaceae (Salix caprea). Гнездовое поведение: самки располагают линейные гнёзда в пустых раковинах брюхоногих моллюсков. Ячейки разделяют перегородками из лиственной мастики. Чтобы замаскировать гнездо, покрывают раковину хвоей и мелкими щепками [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Местообитание: на территории КСП представители данного вида приурочены к остепнённым и суходольным разнотравно-злаковым лугам. Распространение Западно-центрально-палеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]). Европа (до 61° с. ш.), Кавказ, Средняя Азия [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, окр. п. Тельбес, 03.VII.2003 – 3♀♀ (ДС). КА: Тисульский р-н, окр. п. Макаракский, 03.VII.2007 – 1♀ (СЯ), 10 км С п. Полуторник, 04.VII.2009 – 1♀ (ДС). КК: Кемеровский р-н, г. Кемерово, 13.V.1998 – 1♀ (НЕ), 26.V.1998 – 1♀ (ДВ), 15.V.1999 – 2♀♀ (НЕ), 07.V.2001 – 4♀ (НЕ), 05.V.2004 – 7♀♀ (ДС), 03.VI.2004 – 1♀ (НЕ), 20.V.2012 – 4♀♀ (СЯ), 25.V.2012 – 3♀♀ (СЯ), 04.VI.2012 – 1♀, 05.VI.2012 – 8♀♀, окр. д. Силино, 18.V.1999 – 3♀♀, окр. п. Мозжуха, 14.V.2008 – 1♀ (ДС), 16.V.2008 – 1♀ (ДС), 15.V.2010 – 2♀♀ (АК). 69 Точки сбора: указаны на рис. 4.1.5. Триба ANTHIDIINI Anthidiellum Cockerell, 1904 Мелкие (6–8 мм) коренастые пчёлы. Чёрные, с жёлтыми пятнами и коротким светлым опушением. Летние формы. Трофически связаны с растениями из семейств Fabaceae и Asteraceae. Сооружают гнёзда в виде свободных ячеек из смолистого вещества на поверхности камней, либо прикрепляют их к ветвям деревьев [Осычнюк А.З. и др., 1978]. Небольшой род, насчитывающий 44 вида из 6 подродов. Представители данного рода широко распространены на территории Палеарктики (6 видов) и Австралийского региона. В России встречается 1 вид номинативного подрода [Banaszak J. et al., 2001; Прощалыкин М.Ю., 2007 в]. **Anthidiellum (Anthidiellum) strigatum (Panzer, 1805) Экология и биология Сезон лёта: июнь – июль. Трофические связи: олиголект на Fabaceae [Песенко Ю.А., 1971; Осычнюк А.З. и др., 1978; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Matache I., Ban C., 2006]. Кроме растений сем. Fabaceae (Trifolium repens, Vicia cracca), в КСП отмечено также посещение цветков Apiaceae (Heracleum dissectum), Crassulaceae (Sedum hybridum), Geraniaceae (Geranium pratense), Scrophulariaceae (Rhinanthus aestivalis), Fabaceae (Trifolium hybridum). Гнездовое поведение: сооружают гнёзда на поверхности скал и камней, или прикрепляют их к ветвям кустарников. Ячейки продолговато-овальной формы, формируются из смолы [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006]. Местообитание: на территории КСП данный вид приурочен главным образом к сосново-берёзовым лесам и лесным разнотравно-злаковым лугам. 70 Распространение Голарктический полизональный вид. PA. Россия: Европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Новосибирская обл. [Прощалыкин М.Ю., 2013 а], Алтайский край [Wnukowskij, W., 1927], Кемеровская обл. [Яковлева С.Н., 2011]); Восточная Сибирь (респ. Тыва [Прощалыкин М.Ю., 2013 а], Бурятия, Забайкальский край [Proshchalykin M., 2007]); Дальний Восток России [Proshchalykin M., 2004]. Европа (до 63° с. ш.), Северная Африка, Турция, Кавказ, Средняя Азия, Корея [Monsevićius V., 1995, Banaszak J., Romasenko L., 1998; Banaszak J.et al., 2001; Guler Y., Cağatay N., 2006]. Материал ГШ: Таштагольский р-н, 2 км ЮЗ п. Усть-Кабырза, 02.VII.2008 – 1♀, 04.VII.2008 – 7♀♀, 06.VII.2008 – 3♀♀ (ДС), 10.VII.2008 – 3♀♀, 12.VII.2008 – 1♀ (СЯ). КК: Промышленновский р-н, 4 км СЗ с. Журавлёво, 29.VI.2006 – 3♀♀ (СЯ), 03.VII.2006 – 4♀♀, 05.VII.2006 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.5. Anthidium Fabricius, 1804 Крупные и средние пчёлы, длиной от 10 до 16 мм. Чёрные, с яркими жёлтыми, красновато-жёлтыми или беловатыми пятнами и редким светлым опушением. Полилекты и олиголекты, имеют более или менее тесные трофические связи с растениями сем. Fabaceae, Asteraceae, Lamiaceae. Летние формы. Гнездятся в готовых полостях в различных субстратах, ячейки облицовывают пухом растений [Осычнюк А.З. и др., 1978; Прощалыкин М.Ю., 2007 в]. Род насчитывает около 100 видов из 7 подродов. Ареал охватывает все континенты, за исключением Австралии и тропиков Ориентальной области. В Палеарктике встречается около 90 видов [Michener C., 1948; Banaszak J. et al., 2001; Прощалыкин М.Ю., 2012]. На территории Западной Сибири отмечено 4 вида. В Кузнецко-Салаирской области встречается 3 вида. 71 Anthidium (Anthidium) punctatum Latreille, 1809 Экология и биология Сезон лёта: июнь – июль. Трофические связи: полилект; предпочитает цветки растений сем. Fabaceae (Lotus, Trifolium), Lamiaceae (Thymus), Asteraceae (Centaurea, Cirsium) [Осычнюк и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999]. На территории КСП отловлен на Trifolium pratense. Гнездовое поведение: гнездятся в готовых норках в песчаной почве и в щелях между камнями. Гнёзда линейные, содержат 1–3 ячейки овальной формы, состоящие из хлопкоподобного вещества [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Местообитание: в КСП обнаружен на суходольном и лесном разнотравнозлаковых лугах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл. [Яковлева С.Н., 2013], Алтайский край [Прощалыкин М.Ю., 2013 а]); Восточная Сибирь (респ. Тыва, Хакасия, Красноярский край [Прощалыкин М.Ю., 2013 а, в], Иркутская обл., Бурятия, Забайкальский край [Proshchalykin M., 2007; Прощалыкин М.Ю., 2012], Якутия [Романькова Т.Г., 1995]); Дальний Восток России [Proshchalykin M., 2004]. Европа, Северная Африка, Кавказ, Центральная Азия, Северный Казахстан, Монголия [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998]. Материал ГШ: Таштагольский р-н, 2 км ЮЗ п. Усть-Кабырза, 03.VII.2008 – 2♂♂ (ДС). КК: Беловский р-н, окр. г. Белово, 26.VII.2005 – 3♀♀, 1♂ (НТ); Прокопьевский р-н, 5 км СВ д. Большая Талда, 18.VII.2004 – 2♂♂ (ДС). Точки сбора: указаны на рис. 4.1.6. 72 Рисунок 4.1.6. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Anthidium (Anthidium) manicatum (Linnaeus, 1758) Экология и биология Сезон лёта: июнь – июль. Трофические связи: полилект. Предпочитает растения сем. Lamiaceae и Fabaceae [Песенко Ю.А., 1971; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J.et al., 2001; Guler Y., Cağatay N., 2006; Matache I., Ban C., 2006]. В КСП зарегистрирован на Asteraceae (Chamomilla recutita), Crassulaceae (Sedum hybridum). 73 Гнездовое поведение: гнездятся в готовых полостях в различных субстратах (в том числе используют норки других насекомых в почве, полые стебли растений, отверстия в древесине и камне). Гнёзда линейные, реже линейно-разветвлённые. Ячейки крупные [Banaszak J., Romasenko L., 1998; Matache I., Ban C., 2006]. Местообитание: в КСП встречается на лесных и пойменных лугах. Распространение Голарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Новосибирская обл., респ. Алтай [Прощалыкин М.Ю., 2013 а], Кемеровская обл. [Еремеева Н.И., Сидоров Д. А., 2006]); Восточная Сибирь (респ. Тыва, Хакасия [Прощалыкин М.Ю., 2013 а, в], Бурятия [Proshchalykin M., 2007]). Европа (севернее 64° с.ш.), Канарские острова, Северная Африка, Турция, Кавказ, Средняя Азия. NA. Северная Америка, Канада. NT. Южная Америка (Перу, Бразилия, Суринам, Парагвай, Уругвай, Аргентина). AU. Новая Зеландия [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J.et al., 2001; Guler Y., Cağatay N., 2006; Strange J.P. et al., 2011]. Материал ГШ: Таштагольский р-н, Подкатунские скалы, 07.VII.2005 – 1♀ (ДС), окр. п. Тельбес, 03.VII.2009 – 1♀ (НЕ), 11 км С от устья р. Антроп, 30.VII.2011 – 1♀ (ДС). КК: Кемеровский р-н, г. Кемерово, 17.VII.1996 – 3♀♀ (НЕ), 01.VII.1997 – 2♀♀, 1♂ (НЕ), 23.VI.1999 – 1♀ (НЕ), 03.VII.1999 – 4♀♀, 02.VII.2004 – 1♂ (СБ), окр. д. Подъяково, 01.VII.2001 – 1♀ (НЕ). Точки сбора: указаны на рис. 4.1.6. Anthidium (Anthidium) septemspinosum Lepeletier, 1841 Экология и биология Сезон лёта: июль – август. Трофические связи: предпочитает посещать растения из сем. Lamiaceae 74 (Phlomis tuberosa), Asteraceae (Centaurea jacea), Fabaceae (Vicia), Lythraceae (Lythrum salicaria) [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. В КСП установлены тесные трофические связи с растениями сем. Fabaceae (Medicago falcate, Vicia cracca). Отмечено также посещение Lamiaceae (Phlomis tuberosa). Гнездовое поведение: гнездятся в готовых полостях в различных субстратах [Banaszak J., Romasenko L., 1998]. Местообитание: на территории КСП приурочен к суходольным разнотравнозлаковым и разнотравно-злаково-бобовым лугам, а также к остепнённым участкам. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Омская обл. [Лавров С.Д., 1927], Новосибирская обл. [Прощалыкин М.Ю., 2013 а], Алтайский край [Шумакова П.С. и др., 1982], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (респ. Тыва, Хакасия, Красноярский край [Прощалыкин М.Ю., 2013 а, в], Иркутская обл., Бурятия, Забайкальский край [Proshchalykin M., 2007]); Дальний Восток России [Proshchalykin M., 2004]. Южная Европа, Центральная и Восточная Европа (частично), Северный Казахстан, Монголия, Корея, Япония [Banaszak J., Romasenko L., 1998]. Материал КА: Крапивинский р-н, г. Зеленогорск, 13.VII.2007 – 5♀♀ (НЕ). КК: Беловский р-н, окр. г. Белово, 23.VIII.2007 – 4♂♂ (ДС); Гурьевский р-н, с. Сосновка, 11.VII.2005 – 1♀ (НЕ); Кемеровский р-н, г. Кемерово, 17.VII.1996 – 2♂♂, 10.VIII.1998 – 1♀, 1♂ (НЕ), 13.VIII.1998 – 1♂ (НЕ), 06.VII.1999 – 1♂ (НЕ), 02.VII.2005 – 1♀ (ДС), д. Мамаевка, 02.IX.2005 – 1♀ (ДС); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 28.VII.2006 – 2♀♀, 29.VII.2006 – 16♀♀, 01.VIII.2006 – 8♀♀, 3♂♂, 12.VII.2012 – 3♀♀, 2♂♂, 26.VII.2012 – 4♀♀ (СЯ); окр. с. Красное, 13.VII.2004 – 1♂ (НЕ); Промышленновский р-н, пгт Промышленная, 75 28.VIII.2005 – 3♂♂ (НЕ); Топкинский р-н, окр. г. Топки, 05.VIII 2004 – 1♂, окр. д. Цыпино, 29.VII.2003 – 1♂ (НЕ); Юргинский р-н, окр. п. Юргинский, 15.VII.2004 – 1♀ (СБ). Точки сбора: указаны на рис. 4.1.6. Bathanthidium Mavromoustakis, 1953 Мелкие пчёлы, с жёлтыми пятнами на брюшке. Гнёзда строят в готовых полостях, используя древесную смолу. Включает летние формы. Небольшой род, распространённый исключительно в Азии. Включает 8 видов [Прощалыкин М.Ю., 2007 в, Прощалыкин М.Ю., 2012]. На территории КузнецкоСалаирской области встречается 1 вид. *Bathanthidium (Stenanthidiellum) sibiricum (Eversmann, 1852) Экология и биология Сезон лёта: июнь – июль. Трофические связи: отловлен на Rosaceae. Гнездовое поведение: самки размещают гнёзда в готовых полостях [Прощалыкин М.Ю., 2007 в]. Местообитание: в КСП обнаружен на суходольных разнотрано-злаковых лугах. Распространение Центрально-восточно-палеарктический полизональный вид. PA. Россия: Западная Сибирь (Алтайский край [Прощалыкин М.Ю., 2013 а], Кемеровская Забайкальский обл. [Яковлева край) С.Н., [Proshchalykin 2012]); M., Восточная 2007]; Сибирь Дальний (Бурятия, Восток России [Proshchalykin M., 2004]. Северо-Восточный Китай, Корея [Proshchalykin M., 2007]. Материал КК: Кемеровский р-н, г. Кемерово, 29.VI.2004 – 2♀♀ (НЕ), 04.VII.2006 – 2♀♀, 25.VII.2006 – 1♀ (СД), с. Креково, 24.VI.2012 – 1♀, 01.VII.2012 – 3♂♂ (ДЕ); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 13.VII.206 – 2♀♀ (СЯ). 76 Точки сбора: указаны на рис. 4.1.7. Рисунок 4.1.7. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Stelis Panzer, 1806 Мелкие (3–8 мм) и средние (9–12 мм) клептопаразитические пчёлы. Преимущественно чёрные, с редким опушением из светлых волосков. Вершинные части тергитов часто светлые или имеют красновато-бурую окраску. Весенние, весеннее-летние и летние формы. Паразитируют в гнёздах пчёл из родов Anthidium, Anthidiellum, Lithurgus, Chelostoma, Heriades, Hoplitis, Osmia, 77 Anthocopa, Chalicodoma, Ceratina [Noskiewicz, J. , 1961; Осычнюк А.З. и др., 1978; Прощалыкин М.Ю., 2007 в]. Представители данного рода широко распространены на всех континентах, кроме Австралии. Род насчитывает около 100 видов из 6 подродов. Для территории Палеарктики отмечено около 30 видов [Banaszak J. et al., 2001; Прощалыкин М.Ю., 2012]. Для Западной Сибири отмечено 4 вида. В КузнецкоСалаирской провинции встречается 3 вида. Stelis (Stelis) punctulatissima (Kirby, 1802) Экология и биология Сезон лёта: июнь – июль. Виды-хозяева: является клептопаразитом у видов Hoplitis adunca, Osmia aurulenta, O. fulviventris, Anthidium manicatum, Chalicodoma parietina, Anthophora furcata [Осычнюк А.З. и др.. 1978; Warncke K., 1992 b; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001; Matache I., Ban C., 2006]. Местообитание: в КСП встречается на суходольных разнотравно-злаковых лугах. Распространение Западно-центрально-палеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Новосибирская обл. [Прощалыкин М.Ю., 2013 а], Алтайский край [Данилов Ю.Н., 2006], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]). Европа (до 63° с. ш.), Северная Африка, Кавказ, Северный Казахстан [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КК: Беловский р-н, окр. с. Беково, 18.VII.2001 – 2♀♀ (НТ); Кемеровский р- н, окр. д. Подъяково, 08.VII.2005 – 2♀♀ (НЕ); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 13.VII.2006 – 3♀♀ (СЯ). Точки сбора: указаны на рис. 4.1.7. 78 Stelis (Stelis) simillima Morawitz, 1876 Экология и биология Сезон лёта: июнь – июль. Виды-хозяева: паразитирует у Lithurgus fuscipennis [Warncke K., 1992 b; Banaszak J., Romasenko L., 1998]. Местообитание: в КСП встречается на суходольных разнотравно-злаковых лугах. Распространение Западно-центрально-палеарктический субаридный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]). Южная, Центральная и Восточная Европа, Северная Африка, Кавказ, Закавказье, Туркестан [Banaszak J., Romasenko L., 1998]. Материал КК: Кемеровский р-н, окр. д. Подъяково, 28.VI.2000 – 1♂ (НТ), окр. п. Пионер, 15.VII.2005 – 5♀♀, 4♂♂ (НЕ); Ленинск-Кузнецкий р-н, окр. г. Ленинск-Кузнецкий, 15.VII.2009 – 4♂♂, 13.VII.2010 – 3♂♂, 16.VII.2012 – 1♂, 17.VII.2012 – 1♂, 22.VII.2012 – 4♂♂ (СЯ). Точки сбора: указаны на рис. 4.1.7. **Stelis (Stelis) ornatula (Klug, 1807) Экология и биология Сезон лёта: июнь – июль. Виды-хозяева: является клептопаразитом у видов Hoplitis claviventris, H. leucomelana, H. tridentata, Osmia maritima, Ceratina cucurbitina [Осычнюк А.З. и др., 1978; Warncke K., 1992 a; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001]. Местообитание: в КСП отловлен на деградированных разнотравно-злаковых лугах. 79 Распространение Транспалеарктический полизональный вид. PA. Россия: Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Кемеровская обл. [Яковлева С.Н., 2011]); Восточная Сибирь (Красноярский край [Прощалыкин М.Ю., 2013 а], Иркутская обл. [Прощалыкин М.Ю., 2012], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 2004]. Европа (между 62° и 40° с. ш.), Северная Африка, Иран, Туркестан [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КК: Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 20.VII.2005 – 2♀♀, 29.VII.2006 – 2♀♀ (СЯ). Точки сбора: указаны на рис. 4.1.8. Trachusa Panzer, 1804 Коренастые пчёлы с брюшком округлой формы, средней величины (11–12 мм). Самки полностью чёрные, с серовато-жёлтым опушением; у самцов наличник, боковые части лица и мандибулы жёлтые. Олиголекты на Fabaceae. Летние формы. Гнездятся в песчаной почве, небольшими колониями. Ячейки гнёзд облицовывают кусочками листьев, склеенных сосновой камедью. Обитают на всех континентах, кроме Австралии. Род включает 43 вида 11 подродов, в России встречается 1 вид [Осычнюк А.З. и др., 1978; Banaszak J. et al., 2001; Прощалыкин М.Ю., 2007 в]. Trachusa (Trachusa) byssina (Panzer, 1798) Экология и биология Сезон лёта: июнь – август. Трофические связи: олиголект на Fabaceae. Встречается на растениях родов Medicago, Trifolium, Lotus, Vicia, Astragalus [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J.et al., 2001; Ban C., 2005]. В КСП трофически связан с Lathyrus pratensis, Vicia cracca. 80 Рисунок 4.1.8. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Гнездовое поведение: гнездится в песке и песчаной почве. Гнёзда обычно линейно-ветвистого типа, расположены близко друг к другу и образуют небольшие скопления. Ячеек обычно от 1 до 4, они формируются из вырезанных полосками кусочков листьев, которые покрыты смолой, собираемой с сосны [Радченко В.Г., Песенко Ю.А., 1994; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Ban C., 2005; Прощалыкин М.Ю., 2007 в]. Местообитание: В европейской части России – лесная зона [Осычнюк А.З. и др., 1978]. В. С. Монсявичус [Monsevićius V., 1995] отмечает, что на территории Литвы данный вид более всего тяготеет к агрикультурным ландшафтам, где 81 преобладают лёгкие почвы, пригодные для гнездования. На территории КСП встречается на суходольных и лесных разнотравно- злаковых лугах, отмечен также в сосново-берёзовых лесах и на остепнённом лугу. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (респ. Тыва, Хакасия, Красноярский край [Прощалыкин М.Ю., Забайкальский край [Proshchalykin M., 2013 2007]); а, в], Дальний Иркутская Восток обл., России [Proshchalykin M., 2007]. Южная Европа (частично), Северная, Центральная и Восточная Европа, Турция, Кавказ, Закавказье, Северная Азия [Осычнюк и др., 1978; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005]. Материал ГШ: Таштагольский р-н, 6 км ЮЗ п. Мундыбаш, 11.VII.2005 – 1♂ (ДС), 2 км ЮЗ п. Усть-Кабырза, 04.VII.2008 – 5♀♀ (ДС), 05.VII.2008 – 1♂ (СЯ), 06.VII.2008 – 2♀♀ (СЯ), пр. б. р. Кондома (52° 59,768' с. ш., 87° 04,017' в. д.), 12.VI.2011 – 1♀ (ДС). КА: Крапивинский р-н, 3 км Ю биостанции КемГУ «Ажендарово», 02.VIII.2009 – 7♀♀, 03.VIII.2009 – 1♀, 05.VIII.2009 – 1♂ (ДС). КК: Беловский р-н, Караканский хребет (сев. отрог), 02.VI.2008 – 4♀♀ (ДС); Ленинск-Кузнецкий р-н, окр. с. Шабаново, 03.VII.1998 – 3♀♀ (НЕ); Промышленновский р-н, 6 км СЗ с. Журавлёво, 28.VI.2000 – 2♀♀ (СЯ), 07.VII.2006 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.8. 82 Триба DIOXYINI Aglaoapis Cameron, 1901 Мелкие (7–10 мм) и средней величины (10–12 мм) чёрные пчёлы с редким сероватым опушением. Клептопаразитические виды, паразитируют у представителей родов Chalicodoma, Megachile и Hoplitis. Род включает 3 вида, широко распространённых на территории Европы, Малой Азии, Кавказа и Северной Африки [Прощалыкин М.Ю., 2007 в]. На территории Западной Сибири встречается 1 вид. Aglaoapis tridentatus (Nylander, 1848) Экология и биология Сезон лёта: июнь – июль. Виды-хозяева: клептопаразит видов Hoplitis adunca, H. ravouxi, H. anthocopoides, Chalicodoma parietina, Megachile argentata [Песенко Ю.А., 1971; Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J.et al., 2001]. Местообитание: в КСП обнаружен на остепнённых участках. Распространение Транспалеарктический суббореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл.) [Еремеева Н.И., Сидоров Д.А., 2006]; Восточная Сибирь (Тыва, Хакасия [Прощалыкин М.Ю., 2013 б, в], Иркутская обл., Бурятия, Забайкальский край) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Европа (до 62 °с. ш.), Турция, Кавказ, [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КК: Ленинск-Кузнецкий р-н, окр. с. Шабаново, 04.VII.1998 – 1♂ (НЕ); Промышленновский р-н, 6 км СЗ с. Журавлёво, 29.VI.2002 – 3♀♀ (НЕ). Точки сбора: указаны на рис. 4.1.8. 83 Триба MEGACHILINI Coelioxys Latreille, 1809 Паразитические пчёлы. Обычно чёрные, иногда с красновато-бурыми пятнами, склериты грубо скульптурированные. Коренастые, размеры варьируются от 2–9 до 14–22 мм. Глаза большинства видов опушены волосками. Вершина брюшка у самок сильно вытянута и заострена, у самцов шестой тергит брюшка с развитыми шипами. Обычно летние и весенне-летние формы [Осычнюк А.З. и др., 1978; Прощалыкин М.Ю., 2006 в]. Являются клептопаразитами пчёл сем. Megachilidae – паразитируют у представителей родов Chalicodoma, Megachile, некоторых видов рода Osmia, Anthocopa, Hoplitis, редко у нескольких видов сем. Anthophoridae. Представители рода распространены на всех континентах, кроме Австралии. Описано боее 310 видов 15 подродов, на территории Палеарктики зарегестрировано около 150 видов [Michener C., 2000; Banaszak J. et al., 2001; Прощалыкин М.Ю., 2006 в, 2007 в]. Для территории Западной Сибири отмечено 13 видов, из них 10 встречается в Кузнецко-Салаирской провинции. *Coelioxys (Allocoelioxys) afra Lepeletier, 1841 Экология и биология Сезон лёта: июнь – июль. Виды-хозяева: является клептопаразитом у Megachile pilidens, M. argentata, M. apicalis [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Guler Y., Cağatay N., 2006], Местообитание: в КСП отловлен на остепнённом разнотравно-злаковом лугу. Распространение Транаспалеарктический субаридный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл.) [Яковлева С.Н., 2012]; Восточная Сибирь (Бурятия) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004; Proshchalykin M., Lelej A., 2004]. Южная, Центральная и Восточная Европа, 84 Северная Африка, Турция, Кавказ, Средняя Азия, Китай [Warncke K., 1992 a; Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001 Guler Y., Cağatay N., 2006]. Материал КК: Кемеровский р-н, 5 км ССЗ д. Старочервово, 17.VII.2009 – 1♂ (ДС). Точки сбора: указаны на рис. 4.1.9. Рисунок 4.1.9. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции 85 Coelioxys (Allocoelioxys) emarginata Forster, 1853 Экология и биология Сезон лёта: июль – август. Виды-хозяева: предположительно Megachile leucomalla [Warncke K., 1992 a]. Местообитание: в КСП обнаружен на суходольном разнотравно-злаковом лугу и в смешанном лесу. Распространение Транспалеарктический субаридный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл.) [Еремеева Н.И., Сидоров Д.А., 2006]; Восточная Сибирь (Тыва [Прощалыкин М.Ю., 2013 в], Бурятия [Proshchalykin M., 2007]); Дальний Восток России [Proshchalykin M., 2004; Прощалыкин М.Ю., 2006 в]. Южная, Центральная и Восточная Европа, Турция, Туркестан, Китай [Warncke K., 1992 a; Ozbek H., Zanden G., 1994; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, окр. п. Тельбес, 08.VII.2003 – 1♀ (СД). КК: Кемеровский р-н, 8 км ЮВ г. Кемерово (с/о «Маручак»), 26.VIII.1999 – 1♀ (НЕ); Прокопьевский р-н, 5 км СВ г. Прокопьевск, 16.VIII.2005 – 1♂ (СД). Точки сбора: указаны на рис. 4.1.9. Coelioxys (Boreocoelioxys) alata Forster, 1853 Экология и биология Сезон лёта: июль – август. Виды-хозяева: паразитирует у Megachile ligniseca, Anthophora furcata [Erlandsson S., 1955; Осычнюк и др., 1978; Gogala A., 1999; Banaszak J. et al., 2001]. Местообитание: в КСП обнаружен на разнотравно-злаковых лугах, а также в берёзовом лесу. Распространение Транспалеарктический бореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь 86 (Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Иркутская обл. [Романькова Т.Г., 1995], Забайкалье [Proshchalykin M., 2007]); Дальний Восток России [Proshchalykin M., 2004; Прощалыкин М.Ю., 2006 в]. Центральная и Восточная Европа [Warncke K., 1992 a; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КА: Крапивинский р-н, окр. биостанции КемГУ «Ажендарово», 07.VIII.2006 – 1♀ (ДС); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 05.VII.2004 – 1♀ (ДС). КК: Кемеровский р-н, 3 км ЮЗ д. Барановка, 26.VIII.2007 – 1♀, окр. п. Зеленовский, 19.VII.2008 – 2♀♀ (ДС). Точки сбора: указаны на рис. 4.1.9. Coelioxys (Boreocoelioxys) inermis (Kirby, 1802) Экология и биология Сезон лёта: июль – август. Виды-хозяева: является клептопаразитом Megachile centuncularis, M. alpicola, M. versicolor и Anthocopa papaveris [Осычнюк А.З. и др., 1978; Banaszak J. et al., 2001; Gogala A., 1999; Прощалыкин М.Ю., 2006 в; Matache I., Ban C., 2006]. Местообитание: в КСП встречается в различных биотопах: на деградированных лугах и залежах, остепнённых, суходольных и лесных разнотравно-злаковых лугах, осиново-берёзовых и сосново-берёзовых лесах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Ханты-Мансийский автономный округ [T.V. Levchenko, K.P. Tomkovich, 2014], Омская обл. [Прощалыкин М.Ю., 2013 б], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Бурятия, Забайкальский край) [Proshchalykin M., 2007]; Дальний Восток России 87 [Proshchalykin M., 2004]. Европа (до 66°с. ш.), Северная Африка, Кавказ, Турция, Туркестан [Warncke K., 1992 a; Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Новокузнецкий р-н, окр. д. Сары-Чумыш, 09.VII.2002 – 2♂♂, 11.VII.2002 – 1♂ (НЕ); Таштагольский р-н, окр. п. Шерегеш, 4 км В г. Мустаг, 21.VII.1999 – 2♂♂, окр. г. Мустаг (г. Зелёная) (НЕ), 30.VII.1999 – 1♂ (ДВ), окр. п. Тельбес, 03.VII.2003 – 1♂ (НЕ), 2 км ЮЗ п. Усть-Кабырза, 03.VII.2008 – 3♀♀, 3♂♂ (ДС), 06.VII.2008 – 3♂♂, 10.VII.2008 – 1♂ (СЯ), пр. б. р. Кондома (52° 53,866' с. ш., 87° 13,460' в. д.), 08.VI.2011 – 1♂, пр. б. р. Кондома (52° 55,675' с. ш., 87° 09,756' в. д.), 10.VI.2011 – 3♀♀,1♂, пр. б. р. Кондома (52° 59,768' с. ш., 87° 04,017' в. д.), 12.VI.2011 – 1♂. окр. п. Каз, 30.VI.2012 – 1♀ (ДС). КА: Крапивинский р-н, окр. биостанции КемГУ «Ажендарово», 14.VII. 2006 – 1♀, 09.VIII.2006 – 2♀♀ (СЯ); Новокузнецкий р-н, 4 км ЮЮВ г. Чёрный Ворон (верх. теч. р. Верх. Терсь), 20.VII.1998 – 4♀♀, 23.VII.1998 – 1♀ (НЕ), 5 км ЮВ с. Загадное (ср. теч. р. Верх. Терсь), 21.VI.2008 – 2♂♂ (ДС); Тисульский р-н, окр. п. Макаракский, 01.VII.2010 – 1♀, 1♂ (СЯ); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 03.VII.2006 – 1♂ (НЕ), 06.VII.2004 – 2 ♀♀, 2♂♂, 08.VII.2004 – 1 ♀, 2♂♂ (СД). КК: Кемеровский р-н, г. Кемерово, 12.VII.2000 – 3♀♀ (НЕ), 28.VII.2006 – 1♀, 29.VII.2006 – 3♀♀, 30.VII.2006 – 1♀ (СЯ), 10.VIII.2008 – 1♀ (СЯ), 05.VII.2012 – 1♀(СЯ), 12.VII.2012 – 1♀, 14.VII.2012 – 2♀♀ (ДС), 26.VII.2012 – 1♀ (СЯ); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 10.VII.2005 – 2♀♀, 27.VII.2006 – 4♀♀, 30.VII.2006 – 4♀♀, 15.VII.2012 – 2♀♀, 22.VII.2012 – 1♀, 23.VII.2012 – 2♀♀, 25.VII.2012 – 3♀♀, 28.VII.2012 – 3♀♀ (СЯ); Промышленновский р-н, 4 км СЗ с. Журавлёво, 03.VII.2006 – 1♂ (ДС), 04.VII.2006 – 1♂ (СЯ), 08.VII.2006 – 1 ♀, 2♂♂ (СЯ); Топкинский р-н, окр. д. Малый Корчуган, 10.VIII.2003 – 3♀♀ (ДС). Точки сбора: указаны на рис. 4.1.10. 88 Рисунок 4.1.10. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Coelioxys (Boreocoelioxys) mandibularis Nylander, 1848 Экология и биология Сезон лёта: июнь – август. Виды-хозяева: является клептопаразитом Megachile circumcincta, M. centuncularis, M. brevicornis, M. argentata, M. versicolor, M. pyrenaea и Anthocopa papaveris и у некоторых Osmia [Erlandsson S., 1955; Осычнюк и др., 1978; Banaszak J. et al., 2001; Gogala A., 1999]. Местообитание: в КСП отловлен на остепнённых разнотравно-злаковых лугах, 89 лесных лугах, а также в сосново-берёзовых и пихтово-берёзовых лесах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская чать [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Алтайский край [Данилов Ю.Н., 2006], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Тыва, Хакасия, Красноярский край [Прощалыкин М.Ю., 2013 а, в], Бурятия, Забайкальский край [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 2004; Прощалыкин М.Ю., 2006 в]. Северная, Восточная и Центральная Европа, Турция, Кавказ, Казахстан [Warncke K., 1992 a; Ozbek H., Zanden G. 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Новокузнецкий р-н, окр. д. Сары-Чумыш, 04.VII.2002 – 1♂ (НЕ); Таштагольский р-н, 6 км ЮЗ п. Мундыбаш, 15.VII.2005 – 4♀♀ (ДС), 2 км ЮЗ п. Усть-Кабырза, 04.VII.2008 – 3♂♂ (ДС), пр. б. р. Кондома (52° 55,675' с. ш., 87° 09,756' в. д.), 10.VI.2011 – 3♂♂, пр. б. р. Кондома (52° 59,768' с. ш., 87° 04,017' в. д.), 12.VI.2011 – 2♂♂ (ДС). КА: Крапивинский р-н, территория биостанции КемГУ «Ажендарово», 10.VIII.2009 – 2♂♂ (ДС); Тисульский р-н, окр. оз. Большой Берчикуль, 07.VIII.1997 – 1♀ (ДВ); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 08.VII.2004 – 2♂♂ (ДС). КК: Беловский р-н, 4 км ЮВ с. Уроп, 23.VII.2006 – 2♀♀ (ДВ), Прокопьевский р-н, 5 км СВ г. Прокопьевск, 23.VI.2006 – 1♂ (ДС). Точки сбора: указаны на рис. 4.1.10. Coelioxys (Boreocoelioxys) rufescens Lepeletier & Serville, 1825 Экология и биология Сезон лёта: июль – август. Виды-хозяева: клептопаразит Megachile circumcincta, M. willughbiella, 90 M. argentata, Trachusa byssina, Anthophora furcata, A. quadrimaculata, A. bimaculata, A. plagiata, A. parietina [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001; Ban C., 2005; Прощалыкин М.Ю., 2006 в]. Местообитание: на территории КСП встречается на суходольных разнотравнозлаковых лугах, в том числе деградированных, а также на лесных лугах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Красноярский край, Иркутская обл. [Прощалыкин М.Ю., 2013 б], Бурятия, Забайкальский край [Proshchalykin M., 2007], Якутия [Романькова Т.Г., 1995]); Дальний Восток России [Proshchalykin M., 2007]. Европа (до 65° с. ш.), Северная Африка, Турция, Иран, Средняя Азия, Китай [Warncke K., 1992 a; Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J. et al., 2001; Ban C., 2005]. Материал ГШ: Новокузнецкий р-н, окр. д. Сары-Чумыш, 11.VII.2002 – 2♀♀ (НЕ); Таштагольский р-н, 2 км ЮЗ п. Усть-Кабырза, 03.VII.2008 – 3♀♀ (ДС). КА: Крапивинский р-н, территория биостанции КемГУ «Ажендарово», 15.VII.2006 – 1♀ (ВИ), 10.VIII.2009 – 1♀ (ДС); Новокузнецкий р-н, 4 км ЮЮВ г. Чёрный Ворон (верх. теч. р. Верх. Терсь), 23.VII.1998 – 2♀♀ (НЕ). КК: Беловский р-н, окр. п. Краснобродский, 03.VIII.2003 – 1♀ (НЕ); Гурьевский р-н, Ю окр. г. Гурьевск, 02.VII.2007 – 2♀♀ (СЛ); Кемеровский р-н, окр. г. Кемерово, 10.VIII.2000 – 2♀♀ (НЕ), окр. д. Мазурово, 29.VII.2004 – 1♂ (НЕ), г. Кемерово, 21.VII.2005 – 2♀♀ (ДВ), окр. д. Осиновка, 20.VII.2007 – 1♀ (ДС), окр. п. Зеленовский, 26.VII.2008 – 2♀♀ (ДС), с. Креково, 30.VI.2012 – 1♀ (ДЕ); Ленинск-Кузнецкий р-н, окр. с. Красное, 12.VII.2004 – 4♀♀ (ДС); Прокопьевский р-н, 5 км СВ д. Большая Талда, 18.VII.2004 – 1♂ (ДС). 91 Точки сбора: указаны на рис. 4.1.10. Coelioxys (Coelioxys) conoidea (Illiger, 1806) Экология и биология Сезон лёта: июль – август. Виды-хозяева: паразитирует у Megachile lagopoda, M. maritima, Anthophora plagiata, A. quadrimaculata, A. borealis, A. fulvitarsiss, A. furcata, Chalicodoma ericetorum [Erlandsson S., 1955; Осычнюк А.З. и др., 1978; Прощалыкин М.Ю., 2006 в]. Местообитание: тяготеет к агроценозам и другим человеком ландшафтам, где преобладают лёгкие почвы [Monsevićius V., 1995]. На территории КСП обнаружен на суходольных разнотравно-злаковых лугах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Омская обл. [Лавров С.Д., 1927], Алтайский край [Wnukowskij, W., 1927], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Тыва, Красноярский край [Прощалыкин М.Ю., 2013 б, в], Бурятия, Якутия [Романькова Т.Г., 1995; Proshchalykin M., 2007]); Дальний Восток России [Proshchalykin M., 2004]. Европа (до 62° с. ш.), Северная Африка, Турция, Кавказ, Средняя Азия, Китай [Warncke K., 1992 a; Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КК: Прокопьевский р-н, 5 км СВ г. Прокопьевск, 09.VIII.2005 – 1♂ (ДС); Тяжинский р-н, окр. с. Преображенка, 10.VII.2005 –2♀♀ (НЕ). Точки сбора: указаны на рис. 4.1.11. 92 Рисунок 4.1.11. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Coelioxys (Coelioxys) elongata Lepeletier, 1841 Экология и биология Сезон лёта: июнь – сентябрь. Виды-хозяева: клептопаразит Megachile circumcincta, M. centuncularis, M. argentata, M. ligniseca, M. willughbiella [Erlandsson S., 1955; Осычнюк и др., 1978; Banaszak J. et al., 2001; Matache I., Ban C., 2006]. Местообитание: в КСП отмечен на суходольных и лесных разнотравнозлаковых лугах. 93 Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Бурятия, Забайкальский край) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004; Прощалыкин М.Ю., 2006 в]. Европа (до 63° с. ш), Северная Африка, Турция, Кавказ, Иран [Warncke K., 1992 a; Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КА: Новокузнецкий р-н, 5 км ЮВ п. Загадное (ср. теч. р. Верх. Терсь), 21.VI.2008 – 2♀ (ДС); Чебулинский р-н, окр. п. Чумай, 02.IX.2006 – 4♀♀ (ДС). КК: Промышленновский р-н, 4 км СЗ с. Журавлёво, 29.VI.2006 – 4♀♀ (ДС). Точки сбора: указаны на рис. 4.11. *Coelioxys (Coelioxys) lanceolata Nylander, 1852 Экология и биология Сезон лёта: июнь – июль. Виды-хозяева: паразит Megachile nigriventris [Erlandsson S., 1955; Осычнюк А.З. и др., 1978]. Местообитание: в КСП отловлен на суходольном разнотравно-злаковом лугу. Распространение Транспалеарктический бореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл.) [Яковлева С.Н., 2012]; Восточная Сибирь (Иркутская обл. [Romankova T., 2003], Забайкалье [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 2004; Proshchalykin M., Kupianskaya A., 2005]. Европа [Cockerell, T. D., 1928; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. 94 Материал КА: Тисульский р-н, окр. п. Макаракский, 08.VII.2007 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.11. *Coelioxys (Coelioxys) quadridentata (Linnaeus, 1761) Экология и биология Сезон лёта: июнь – июль. Виды-хозяева: клептопаразит Megachile circumcincta, M. willughbiella, M. argentata, Trachusa byssina, Anthophora furcata, A. plagiata, A. parietina [Erlandsson S., 1955; Осычнюк А.З. и др., 1978; Gogala A., 1999; Banaszak J. et al., 2001]. Местообитание: в КСП приурочен к суходольным разнотравно-злаковым лугам и остепнённым участкам. Также отловлен в пихтово-берёзовом лесу. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл.) [Яковлева С.Н., 2011]; Восточная Сибирь (Иркутская обл., Бурятия, Забайкальский край, Якутия) [Романькова Т.Г., 1995; Proshchalykin M., Kupianskaya A., 2005; Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004; Прощалыкин М.Ю. и др., 2004]. Европа (между 66° и 40° с. ш.), Турция, Средняя Азия [Warncke K., 1992 a; Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КА: Тисульский р-н, окр. п. Макаракский, 03.VII.2007 – 2♂♂ (СЛ), 10 км С п. Полуторник, 02.VII.2009 – 1♂, 08.VII.2009 – 3♂♂ (ДС); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 07.VII.2004 – 2♂♂ (ДС). КК: Беловский р-н, Караканский хребет, 12.VI.2006 – 3♀♀ (СЯ). Точки сбора: указаны на рис. 4.1.12. 95 Рисунок 4.1.12. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Megachile Latreille, 1802 Мелкие (7–9 мм), средние и крупные (до 17 мм) коренастые пчёлы, полностью чёрные, иногда имеются красные пятна. Обильно опушены белыми, желтоватосерыми или красновато-бурыми волосками, прилегающими или отстоящими. Данный род включает весенне-летние и летние формы с продолжительным периодом лёта. Виды характеризуются более или менее тесными трофическими связями с растениями сем. Fabaceae, Asteraceae и Lamiaceae. Гнёзда размещают в готовых полостях в различных субстратах, ячейки выстилают кусочками листьев, 96 бересты, лепестков [Осычнюк А.З. и др., 1978]. Представители рода Megachile широко распространены по всему миру. Род включает более 1100 видов из 54 подродов. Для территории Палеарктики известно около 300 видов [Banaszak J. et al., 2001]. В Западной Сибири отмечено 19 видов, из которых 15 встречается на территории Кузнецко-Салаирской провинции. Megachile (Eumegachile) bombycina Radozskovski, 1874 Экология и биология Сезон лёта: июнь – август. Трофические связи: олиголект, посещает цветки растений сем. Asteraceae (Leontodon, Carduus, Centaurea, Helianthus), иногда встречается на Lamiaceae [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Matache I., Ban C., 2006]. В КСП отмечен на Cirsium heterophyllum (Asteraceae). Из других семейств растений посещает также Rosaceae (Filipendula vulgaris) и Scrophulariaceae (Veronica spicata). Гнездовое поведение: самки располагают гнёзда линейного типа в готовых отверстиях в сухой древесине или полых стеблях. Гнёзда располагаются рядом друг с другом, образуя скопления. Гнездо может содержать от 6 до 8 ячеек, разделённых поперечными перегородками. Стенки ячеек стеклоподобные, сформированы из овальных и округлых кусочков листьев, склеенных лиственной мастикой [Banaszak J., Romasenko L., 1998; Matache I., Ban C., 2006]. Местообитание: в КСП приурочен к остепнённым и суходольным разнотравнозлаковым лугам. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Омская обл. [Лавров С.Д., 1927], Новосибирская обл. [Прощалыкин М.Ю., 2013 б], Томская 97 обл. [Конусова О.Л., Янюшкин В.В., 2000], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Бурятия, Забайкальский край) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Европа, Средняя Азия, Казахстан [Пашина Л.С., 1967; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, 11 км С от устья р. Антроп, 31.VII.2011 – 2♀♀. КА: Крапивинский р-н, территория биостанции КемГУ «Ажендарово», 10.VIII.2006 – 1♂ (ДС), окр. г. Зеленогорск, 19.VII.2008 – 2♀♀ (НЕ); Новокузнецкий р-н, 8 км ССВ г. Соловей (ср. теч. р. Верх. Терсь), 20.VII.1998 – 1♂ (НЕ), 4 км ЮЮВ г. Чёрный Ворон (верх. теч. р. Верх. Терсь), 16.VII.1998 – 2♀♀ (НЕ). КК: Беловский р-н, п. Старобачаты, 15.VII.2005 – 1♀ (ДС); Кемеровский рн, окр. п. Новостройка, 25.VII.2004 – 3♀♀ (НЕ), окр. д. Барановка, 03.VIII.2006 – 1♂ (НЕ), окр. п. Зеленовский, 19.VII.2008 – 5♀♀ (НЕ), окр. с. Креково, 01.VII.2012 – 1♂ (ДЕ); Прокопьевский р-н, 5 км СВ д. Большая Талда, 18.VII.2004 – 2♂♂ (ДС); Промышленновский р-н, 6 км СЗ с. Журавлёво, 23.VI.2000 – 1♂ (НЕ), 4 км СЗ с. Журавлёво, 28.VII.2006 – 1♂ (ДС); Топкинский р-н, г. Топки, 15.VIII.2010 – 3♀♀ (ДС). Точки сбора: указаны на рис. 4.1.12. Megachile (Eutricharaea) argentata (Fabricius, 1793) Экология и биология Сезон лёта: июнь – июль. Трофические связи: вид полилектичен, предпочитает Fabaceae [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006]. В КСП зарегистрирован на Dipsacaceae (Scabiosa ochroleuca). Гнездовое поведение: гнездятся в трещинах камней, полых стеблях растений, в почве (главным образом, песчаной). Гнёзда располагают близко одно к другому, образуя большие скопления. Тип гнезда – линейный, редко линейно-ветвистый, 98 с 1–5 ячейками. Ячейки стеклоподобные, изготовлены из кусочков листьев, обычно используют листья берёзы (Betula). Ход от последней ячейки до входного отверстия заполнен песком [Осычнюк А.З. и др., 1978; Banaszak, J. Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006]. Местообитание: в КСП отловлен на остепнённом участке. Распространение Голарктический суббореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Кемеровская обл. [Еремеева Н.И. и др., 2011]); Восточная Сибирь (Бурятия, Забайкальский край) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Европа (до 63° с. ш.), Северная Африка, Турция, Кавказ. NA. Северная Америка [Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998]. Материал КК: Кемеровский р-н, 5 км ССЗ д. Старочервово, 17.VII.2009 – 2♂♂ (ДС). Точки сбора: указаны на рис. 4.1.12. Megachile (Eutricharaea) rotundata (Fabricius, 1787) Экология и биология Сезон лёта: июль – август. Трофические связи: полилект. Посещает цветки растений сем. Asteraceae (Cirsium, Helianthus, Carduus, Centaurea), Fabaceae (Medicago, Onobrychis) и Lamiaceae (Thymus) [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Guler Y., Cağatay N., 2006; Matache I., Ban C., 2006]. На территории КСП отмечено посещение данным видом цветков растений из сем. Asteraceae, Lamiaceae и Rosaceae. Гнездовое поведение: самки устраивают гнёзда в готовых полостях в различных субстратах, с лёгкостью занимают искусственные гнёзда. Тип гнезда – линейный. Ячейки стеклоподобные, в количестве от 5 до 20, не имеющие собственных стенок, лишь разделённые поперечными перегородками [Bosch J. 99 et al., 1993; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006]. Местообитание: в КСП приурочен к остепнённым участкам. Распространение Голарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Омская обл. [Лавров С.Д., 1927], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Алтайский край [Данилов Ю.Н., 2006], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А.. 2006]); Восточная Сибирь (Бурятия, Забайкальский край, Якутия) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Европа, Северная Африка, Кавказ, Турция, Средняя Азия. NA. Северная Америка. NT. Южная Америка (Чили, Аргентина). AU. Австралия, Новая Зеландия [Ozbek H., Zanden G., 1994; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Guler Y., Cağatay N., 2006]. Материал КА: Крапивинский р-н, окр. д. Междугорное, 23.VIII.2003 – 3♀♀ (ДС). КК: Беловский р-н, окр. п. Краснобродский, 03.VIII.2003 – 2♂♂ (НЕ); Кемеровский р-н, 5 км ССЗ д. Старочервово, 17.VII.2009 – 4♂♂ (ДС); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 30.VII.2006 – 6♀♀, 01.VIII.2008 – 1♀ (СЯ). Точки сбора: указаны на рис. 4.1.13. Megachile (Megachile) alpicola Alfken, 1924 Экология и биология Сезон лёта: июнь – август. Трофические связи: вид полилектичен, предпочитает Fabaceae (Medicago, Lotus, Vicia, Trifolium) и Asteraceaea (Leontodon, Cirsium, Carduus, Cichorium, Crepis, Tanacetum) [Banaszak J., Romasenko L,, 1998; Gogala A., 1999; Banaszak J. et al., 2001]. 100 Рисунок 4.13. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции В КСП представители данного вида отмечены на Rosaceae (Potentilla argentea, P. bifurca, Rubus idaeus), Papaveraceae (Chelidonium majus) и Scrophulariaceae (Rhinanthus). Гнездовое поведение: самки устраивают гнёзда в готовых полостях в различных субстратах. Гнёзда линейного типа. Стенки ячеек формируют из кусочков листьев [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Местообитание: в КСП отловлен на суходольных и остепнённых, а также лесных разнотравно-злаковых лугах. 101 Распространение Транспалеарктический бореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл.) [Еремеева Н.И. и др., 2009]; Восточная Сибирь (Иркутская обл., Бурятия, Забайкальский край, Якутия) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Европа, Турция, Корея, Япония [Ozbek H., Zanden G.,1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, 2 км ЮЗ п. Усть-Кабырза, 06.VII.2008 – 1♂ (ДС). КА: Крапивинский р-н, территориия биостанции КемГУ «Ажендарово», 10.VIII.2009 – 1♀ (ДС); Новокузнецкий р-н, 5 км ЮВ п. Загадное (ср. теч. р. Верх. Терсь), 21.VI.2008 – 5♂♂ (ДС); Тисульский р-н, 10 км С п. Полуторник, 02.VII.2009 – 3♀♀, 2♂♂, 10.VII.2009 – 1♂ (СЯ). КК: Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 13.VII.2006 – 4♀♀ (СЯ), окр. п. Мирный, 23.VII.2003 – 5♀♀ (НЕ), окр. с. Шабаново, 15.VII.2008 – 2♂♂ (НЕ), окр. с. Красное, 12.VII.2004 – 1♀ (НЕ), 14.VII.2004 – 1♀ (ДС); Промышленновский р-н, 4 км СЗ с. Журавлёво, 29.VI.2006 – 1♀ (ДС), 02.VII.2006 – 2♀♀ (СЯ). Точки сбора: указаны на рис. 4.1.13. **Megachile (Megachile) centuncularis (Linnaeus, 1758) Экология и биология Сезон лёта: июнь – июль. Трофические связи: полилект, предпочитает Fabaceae (Medicago, Astragalus, Lathyrus, Lotus, Trifolium), Asteraceaea (Carduus, Leontodon, Cichorium, Hieracium, Crepis, Inula, Cirsium) [Осычнюк А.З. и др., 1978; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 200; Matache I., Ban C., 2006]. На территории КСП отловлен на Achillea millefolium (Asteraceae). 102 Гнездовое поведение: самки располагают гнёзда в готовых полостях в различных субстратах. Тип гнезда – линейный, строение ячеек типичное для представителей рода Megachile. Гнездо включает от 3 до 15 ячеек, обычно ход от крайней ячейки до входного отверстия заполнен кусочками листьев. Представители данного вида легко заселяют искусственные места гнездования [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Matache I., Ban C., 2006; Иванов С.П., 2010]. Местообитание: в КСП обитает на суходольных разнотравно-злаковых лугах. Распространение Голарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Омская обл. [Внуковский В.В., 1930], Алтайский край [Шумакова П.С. и др., 1982], Кемеровская обл. [Яковлева С.Н., 2013]); Восточная Сибирь (Иркутская обл. [Романькова Т.Г., 1983], Бурятия, Забайкальский край, Якутия [Давыдова Н.Г., Песенко Ю.А., 2002; Proshchalykin M., 2007]); Дальний Восток России [Proshchalykin M., 2004]. Европа (до 63° с. ш.), Северная Африка, Турция, Кавказ. NA. Северная Америка [Шумакова П.С. и др., 1982; Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, 2 км ЮЗ п. Усть-Кабырза, 05.VII.2008 – 2♀♀ (ДС), окр. п. Тельбес, 03.VII.2003 – 4♀♀ (НЕ), 04.VII.2003 – 2♀♀ (НЕ), 05.VII.2003 – 3♀♀ (ДС). Точки сбора: указаны на рис. 4.1.13. Megachile (Megachile) genalis Morawitz, 1880 Экология и биология Сезон лёта: июнь – август. Трофические связи: олиголект на Asteraceae [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. 103 В КСП отловлен на Cirsium heterophyllum, Arctium tomentosum, Sonchus arvensis (Asteraceae). Отмечено также посещение цветков Lathyrus pratensis (Fabaceae) и Leonurus tataricus (Lamiaceae). Гнездовое поведение: самки размещают линейные гнёзда в полых сухих стеблях и цветоносах растений рода Allium («стрелках» лука). Гнездо может содержать от 1 до 15 ячеек, стеклоподобных, отделённых от стенок полости стебля слоем из кусочков листьев. Пространство от крайней ячейки до выхода заполняется кусочками листьев [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L, 1998; Gogala A., 1999; Banaszak J. et al., 2001; Hausl-Hofstatter U., 2007; Иванов С.П., 2010]. Местообитание: в КСП обитает на остепнённых и суходольных разнотравнозлаковых лугах, встречается также в сосново-березовых лесах. Распространение Голарктический суббореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Алтайский край [Данилов Ю.Н., 2006], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Дальний Восток России [Proshchalykin M., 2004]. Южная и Центральная Европа, Кавказ, Западный Казахстан, Монголия, Китай (провинция Уйгур), Япония [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КА: Крапивинский р-н, окр. биостанции КемГУ «Ажендарово», 05.VII.1996 – 1♂ (ДС), 12.VIII.2006 – 3♀♀, 2♂♂ (СЯ), 03.VIII.2009 – 2♀♀ (ДС); Новокузнецкий р-н, 4 км ЮЮВ г. Чёрный Ворон (верх. теч. р. Верх. Терсь), 16.VII.1998 – 1♀, 20.VII.1998 – 1♀ (НЕ). КК: Гурьевский р-н, 7 км ЮЗ г. Гурьевск, 02.VII.2007 – 2♂♂ (СЛ); Кемеровский р-н, 17 км Ю г. Кемерово (с/о «Маручак»), 20.VIII.2005 – 3♀♀ (НЕ), Ю окр. д. Подъяково, 10.VIII.1995 – 1♂ (НЕ), п. Пригородный, 23.VII.2006 – 1♀ (НЕ); Промышленновский р-н, 4 км СЗ с. Журавлево, 28.VI.2006 – 5♀♀ (ДС), 30.VI.2006 – 4♀♀, 6♂♂ (СЯ), 04.VII.2006 – 1♂ (ДС). 104 Точки сбора: указаны на рис. 4.1.14. Рисунок 4.14. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Megachile (Megachile) lapponica Thomson, 1872 Экология и биология Сезон лёта: июнь – август. Трофические связи: предпочитает посещать растения сем. Fabaceae Lamiaceae [Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005]. и 105 На территории КСП, кроме вышеуказанных семейств растений, отмечен на Campanulaceae (Campanula sibirica), Onagraceae (Chamerion angustifolium) и Rosaceae (Geum, Potentilla, Rubus). Из Fabaceae предпочитает Medicago falcata и Vicia amoena. Гнездовое поведение: самки располагают гнёзда в готовых отверстиях в отмершей сухой древесине [Banaszak J., Romasenko L., 1998; Ban C., 2005]. Местообитание: в КСП встречается главным образом на остепнённых и суходольных разнотравно-злаковых лугах, реже – на лесных разнотравнозлаковых лугах и в сосново-берёзовых лесах. Распространение Голарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З.и др., 1978]; Северный Урал [Proshchalykin M., 2007]; Западная Сибирь (Ханты-Мансийский автономный округ [T.V. Levchenko, K.P. Tomkovich, 2014], Томская обл. [Wnukowsky W., 1936], респ. Алтай [Прощалыкин М.Ю., 2013 б], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Иркутская обл., Бурятия, Забайкальский край [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 2004]. Северная Европа, Восточная и Центральная Европа (часть), Корея, Япония [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005]. Материал ГШ: Таштагольский р-н, окр. п. Шерегеш, 4 км В г. Мустаг, 21.VII.1999 – 2♂♂ (НЕ), окр. г. Мустаг (г. Зелёная), 11.VII.2001 – 1♂ (НЕ), 06.VII.2003 – 1♀ (ДС), ср. теч. Р. Мундыбаш, окр. д. Клёпочный, 07.VIII.2007 – 1♀, окр. д. Кондома, 08.VIII.2007 – 2♀♀, (ДС), 2 км ЮЗ п. Усть-Кабырза, 05.VII.2008 – 1♀, 1♂ (СЯ), 14.VII.2008 – 1♀, п. Усть-Анзас, 04.VIII.2012 – 1♂ (СД). КА: Крапивинский р-н, территория биостанции КемГУ «Ажендарово», 10.VIII.2006 – 2♂♂ (ДС); Новокузнецкий р-н, 5 км ЮВ п. Загадное (ср. теч. р. Верх. Терсь), 20.VI.2008 – 2♂♂, 21.VI.2008 – 2♂♂ (ДС); Тисульский р-н, 10 км С п. Полуторник, 07.VII.2009 – 1♂ (СЯ), 08.VII.2009 – 1♂ (ДС), 09.VII.2009 – 4♂♂ 106 (ДС); Хакасия, Аскизский р-н, КК: Кемеровский р-н, 11 км СВ п. Балыксу, 07.VII.2004 – 2♂♂ (ДС). п. Кедровка, 03.VIII.2004 – 2♀♀ (ДС); Ленинск-Кузнецкий р-н, окр. с. Шабаново, 04.VII.1998 – 5♀♀ (ДЕ); 5 км СВ г. Прокопьевск, 26.VII.2007 – 1♀ (ДС); Прокопьевский р-н, Промышленновский р-н, 4 км СЗ с. Журавлёво, 02.VII.2006 – 6♀♀ (СЯ), 03.VII.2006 – 4♀♀, 3♂♂ (ДС), 04.VII.2006 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.14. Megachile (Megachile) ligniseca (Kirby, 1802) Экология и биология Сезон лёта: июнь – август. Моновольтинный [Gogala A., 1999]. Трофические связи: полилектичный вид, предпочитает Asteraceae (Cirsium, Carduus, Centaurea) [Осычнюк А.З. и др., 1978; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001; Ban C., 2005; Matache I., Ban C., 2006]. В КСП также отмечено посещение особями данного вида растений сем. Asteraceae (Arctium tomentosum, Carduus crispus, Cirsium heterophyllum, C. vulgare, Crepis sibirica, Leontodon autumnalis, Solidago virgaurea), а также Fabaceae (Lathyrus pratensis, Trifolium hibridum), Geraniaceae (Geranium pratense), Lamiaceae (Prunella vulgaris, Leonurus tataricus), Lythraceae (Lythrum virgatum), Onagraceae (Chamerion angustifolium) и Scrophulariaceae (Veronica incana, V. longifolia). Гнездовое поведение: гнездятся в мёртвой древесине, используя готовые полости. Гнездо содержит 6–8 ячеек. Структура гнезда и используемые материалы типичны для представителей рода Megachile [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Ban C., 2005; Matache I., Ban C., 2006]. Местообитание: на территории КСП встречается на суходольных, остепнённых и лесных разнотравно-злаковых лугах, в том числе деградированных, на залежах, а также в берёзовых и берёзово-осиновых лесах. В Хакасии отмечен в пихтовоосиновых лесах. 107 Распространение Транспалеарктический суббореальный вид. PA. Росcия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Томская обл. [Wnukowsky W., 1936; Конусова О.Л., Янюшкин В.В., 2000], Алтайский край [Wnukowskij W., 1927], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Иркутская обл., [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 2004]. Северная, Центральная и Восточная Европа, Кавказ, Северо-Восточный Китай, Япония [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005]. Материал ГШ: Таштагольский р-н, 4 км В г. Мустаг, 21.VII.1999 – 1♂ (НЕ), окр. п. Шерегеш, окр. г. Мустаг (г. Зелёная), 29.VII.1999 – 3♀♀ (НЕ), 6 км ЮЗ п. Мундыбаш, 13.VII.2005 – 4♂♂ (ДС, СЯ), Подкатунские скалы, 13.VII.2005 – 10♀♀ (ДС, СЯ), 17.VII.2007 – 1♂, 01.VIII.2007 – 1♂, 2 км ЮЗ п. Усть-Кабырза, 02.VII.2008 – 1♂ (ДС), 04.VII.2008 – 7♂♂ (СЯ), 05.VII.2008 – 1♀,1♂ (СЯ), 06.VII.2008 – 1♂ (ДС), 07.VII.2008 – 1♂ (ДС), 09.VII.2008 – 1♀ (ДС), 10.VII.2008 – 2♀♀ (СЯ), 11.VII.2008 – 7♂♂ (СЯ), 12.VII.2008 – 1♂ (СЯ), 14.VII.2008 – 2♂♂ (ДС), п. Усть-Анзас, 03.VII.2008 – 3♂♂, 04.VIII.2010 – 2♀♀ (ДС), 11 км С от устья р. Антроп, 31.VII.2011 – 2♀♀ (ДС). КА: Крапивинский р-н, 3 км Ю биостанции КемГУ «Ажендарово», 02.VIII.2009 – 7♀♀, 11♂♂ (ДС), окр. биостанции КемГУ «Ажендарово», 03.VIII.2009 – 1♀, 09.VIII.2009 – 9♀♀, 10.VIII.2009 – 1♀, 2♂♂ (ДС); Новокузнецкий р-н, 5 км СВ г. Новокузнецк, 18.VII.1998 – 2♂♂ (НЕ), 4 км ЮЮВ от г. Чёрный Ворон (верх. теч. р. Верх. Терсь), 23.VII.1998 – 1♂ (НЕ), 8 км ССВ от г. Соловей (ср. теч. р. Верх. Терсь), 18.VII.1998 – 1♂, 26.VII.1998 – 1♂, 29.VII.1998 – 4♀♀ (НЕ); Хакасия, Аскизский р-н, 11 км СВ п. Балыксу, 03.VII.2004 – 4♂♂ (НЕ), 07.VII.2004 – 7♀♀, 3♂♂ (ДС), 08.VII.2004 – 1♀ (ДС). КК: Беловский р-н, окр. г. Белово, 15.VIII.2005 – 7♀♀, 21.VIII.2007 – 6♀♀ (ДС), п. Старобачаты, 15.VII.2005 – 8♀♀, 17.VIII.2006 – 2♀♀ (ДС), 108 4 км ЮВ с. Уроп, 20.VII.2006 – 1♀ (ДС); Гурьевский р-н, Ю окр. г. Гурьевск, 21.VIII.2007 – 2♀♀ (ДС); Кемеровский р-н, окр. г. Кемерово, 28.VII.2000 – 1♂, 20.VII.2006 – 4♀♀, 28.VII.2007 – 8♀♀ (НЕ), окр. д. Барановка, 03.VIII.2006 – 1♂, 05.VII.2008 – 5♀♀, 1♂ (НЕ), окр. п. Зеленовский, 19.VII.2008 – 10♀♀ (НЕ), 3♂♂, окр. п. Мамаевка, 10.VIII.2012 – 4♀♀ (ДС), окр. п. Мозжуха, 06.VIII.2006 – 1♂ (АК), п. Новостройка, 15.VII.2009 – 1♀ (НЕ), д. Осиновка, 27.VII.2008 – 4♀♀ (НЕ), Ю окр. д. Подъяково, 05.VII.1992 – 1♂, 15.VI.2010 – 11♀♀, 09.VII.2010 – 8♀♀ (НТ); г. Кемерово, сосновый бор, 23.VIII.2010 – 3♂♂; Ленинск-Кузнецкий р-н, окр. г. Ленинск-Кузнецкий, 15.VII.2012 – 7♀♀, 20.VII.2012 – 5♀♀, 22.VII.2012 – 5♀♀, 23.VII.2012 – 4♀♀, 25.VII.2012 – 1♀, 26.VII.2012 – 2♀♀, 27.VII.2012 – 1♀ (СЯ), окр. с. Шабаново, 05.VII.1998 – 1♂ (НЕ); Промышленновский р-н, 4 км СЗ с. Журавлёво, 25.VI.2002 – 1♂ (НЕ), 26.VI.2002 – 1♂ (НЕ), 05.VII.2006 – 2♂♂ (ДС), с. Тарасово, 16.VII.2005 – 8♀♀ (НЕ); Топкинский р-н, г. Топки, 15.VIII.2010 – 1♀ (ДС), окр. д. Цыпино, 14.VII.2007 – 6♀♀ (НЕ); Яшкинский р-н, музей-заповедник «Томская Писаница», 15.VII.2009 – 1♀ (СЛ). Точки сбора: указаны на рис. 4.1.15. Megachile (Megachile) melanopyga Costa, 1863 Экология и биология Сезон лёта: июнь – июль. Трофические связи: полилект. Обычно посещает цветки растений сем. Asteraceae [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Гнездовое поведение: гнездится в травяных кочках [Gogala A., 1999]. Местообитание: в КСП отмечен на суходольных и остепнённых участках. Распространение Транспалеарктический полизональный вид. 109 Рисунок 4.1.15. Точки сбора M. ligniseca на территории Кузнецко-Салаирской провинции PA. Россия: Западная Сибирь (Алтайский край [Данилов Ю.Н., 2006], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Бурятия) [Proshchalykin M., 2007]. Южная, Восточная и Центральная Европа, Турция, Кавказ, Северо-Восточный Китай, Корея, Япония [Ozbek H., Zanden G.,1994; Banaszak J., Romasenko L., 1998]. Материал ГШ: Таштагольский р-н, окр. п. Тельбес, 14.VII.2003 – 3♂♂ (ДС), устье р. Антроп, 27.VII.2009 – 1♂ (ДС), Подкатунские скалы, 07.VII.2005 – 1♂ (ДС). 110 КК: Кемеровский р-н, окр. Г. Кемерово, 21.VII.2005 – 2♂♂ (ДВ), Мозжуха, 15.VII.2006 – 3♂♂ (АК), 17.VII.2006 – 1♂ (АК), Промышленновский, 13. VII.2008 – 1♂ (ДС); Промышленновский р-н, окр. п. п. 6 км СЗ д. Журавлево, 26.VI.2002 – 1♂ (НЕ). Точки сбора: указаны на рис. 4.1.14. Megachile (Megachile) versicolor Smith, 1844 Экология и биология Сезон лёта: июнь – август. Трофические связи: полилект. Предпочитает цветки растений сем. Asteraceae (Leontodon, Crepis, Centaurea, Hieracium, Aster, Cichorium, Cirsium) и Fabaceae (Trifolium, Lathyrus, Medicago) [Осычнюк А.З. и др., 1978; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001; Ban C., 2005; Matache I., Ban C., 2006]. На территории КСП отмечено посещение цветков Arctium tomentosum, Chamomilla recutita, Leontodon autumnalis, Centaurea scabiosa (Asteraceae), Lathyrus pratensis, Medicago falcata (Fabaceae), Potentilla argentea (Rosaceae), Veronica longifolia (Scrophulariaceae), Sedum hybridum (Crassulaceae). Гнездовое поведение: самки устраивают гнёзда в полых стеблях растений и полостях в сухой древесине. Тип гнезда, структура и строительные материалы – типичные для представителей данного рода [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Ban C., 2005; Matache I., Ban C., 2006; Иванов С.П., 2010]. Местообитание: в КСП отмечен на остепнённых и суходольных разнотравнозлаковых лугах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Ханты-Мансийский автономный округ [T.V. Levchenko, K.P. Tomkovich, 2014], Алтайский край [Шумакова П.С. и др., 1982], Кемеровская обл. [Еремеева Н.И. и др., 2009]); 111 Восточная Сибирь (Иркутская обл., Забайкальский край [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 2004]. Европа (до 65° с. ш.), Турция, Западный и Северный Казахстан [Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005]. Материал ГШ: Таштагольский р-н, 11 км С от устья р. Антроп, 31.VII.2011 – 2♀♀, ср. теч. Р. Мундыбаш, окр. д. Клёпочный, 07.VIII.2007 – 1♀, 6 км ЮЗ п. Мундыбаш, 11.VII.2005 – 5♀♀ (ДС), 2 км ЮЗ п. Усть-Кабырза, 09.VII.2008 – 1♀, 13.VII.2008 – 1♀, 14.VII.2008 – 1♀ (ДС), окр. д. Кондома, 07.VIII.2007 – 1♀ (ДС). КА: Крапивинский р-н, окр. биостанции «Ажендарово», 07.VIII.2009 – 2♀♀, 13.VIII.2009 – 2♀♀ (ДС). КК: Гурьевский р-н, Ю окр. г. Гурьевск, 28.VIII.2007 – 1♀ (ДС); Кемеровский р-н, г. Кемерово, 12.VII.1996 – 1♀ (НЕ); Ленинск-Кузнецкий р-н, г. Ленинск-Кузнецкий, 15.VII.2006 – 7♀♀ (СЯ), 18.VII.2006 – 5♀♀ (СЯ), окр. с. Красное, 13.VII.2004 – 2♀♀ (ДС); Прокопьевский р-н, окр. с. Лучшево, 31.VII.2008 – 1♀ (ДС), г. Прокопьевск, Промышленновский р-н, 4 км СЗ 02.VIII.1999 – 3♀♀ (НЕ); с. Журавлёво, 30.VI.2006 – 1♀ (ДС), 04.VII.2006 – 3♀♀ (СЯ), 08.VII.2011 – 3♀♀ (ДС), 10.VII.2011 – 2♀♀ (ДС). Точки сбора: указаны на рис. 4.1.16. Megachile (Xanthosarus) analis Nylander, 1852 Экология и биология Сезон лёта: июнь – июль. Трофические связи: вид полилектичен [Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. В КСП посещает цветки растений из семейств Crassulaceae (отловлен на Sedum hybridum), Fabaceae (Astragalus daticus, Lathyrus pratensis,Onobryhis arenaria, Oxytropis campanulata, Vicia amoena), Lamiaceae (Dracocephalum ruyschiana, Phlomis tuberosa). 112 Рисунок 4.1.16. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции Гнездовое поведение: гнёзда размещают в почве, на глубине 2,5–5 см, используя готовые полости либо самостоятельно выкапывая норки. Гнездо линейного типа, состоит из одного хода, расположенного горизонтально, параллельно поверхности почвы. Ячейки цилиндрической формы, состоят из тончайших полосок коры сосны (Pinus) и берёзы (Betula), или из полосок коры и кусочков листьев. Гнездо включает 2–5 ячеек [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998]. Местообитание: на территории КСП встречается главным образом на 113 суходольных и остепнённых разнотравно-злаковых лугах, отмечен также на степных и каменистых участках. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Иркутская обл., Бурятия, Забайкальский край [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 2004]. Центральная и Северная Европа, Южная Европа (часть), Турция, Кавказ, Туркменистан, Корея [Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал ГШ: Таштагольский р-н, окр. п. Тельбес, 03.VII.2003 – 1♂ (НЕ), 05.VII.2003 – 3♀♀ (ДС). КА: Чебулинский р-н, окр. д. Кураково, 29.VI.2008 – 5♀♀, 10.VII.2008 – 1♂ (СЛ). КК: Беловский р-н, Караканский хребет, 12.VI.2006 – 2♂♂ (СЯ), 4 км ЮВ с. Уроп, 20.VII.2006 – 2♀♀ (ДС), 21.VII.2006 – 1♀ (СЯ); Кемеровский р-н, окр. с. Балахонка, 11.VI.2001 – 1♀ (НЕ), Ю окр. д. Подъяково, 01.VII.2010 – 4♀♀ (НТ); Ленинск-Кузнецкий р-н, окр. г. Ленинск-Кузнецкий, 25.VII.2012 – 1♀, 1♂, 26.VII.2012 – 1♂ (СЯ), окр. с. Красное, 14.VII.2004 – 2♂♂ (ДС); Прокопьевский р-н, 4 км СВ г. Прокопьевск, 23.VI.2006 – 5♂♂, 25.VI.2006 – 4♂♂, 5 км СВ г. Прокопьевск, 14.VII.2006 – 1♀, 26.VII.2007 – 1♀ (ДС); Промышленновский р-н, 6 км СЗ с. Журавлёво, 27.VI.2000 – 1♂ (НЕ), 4 км СЗ с. Журавлёво, 30.VI.2006 – 1♀, 02.VII. 2006 – 1♂, 05.VII.2006 – 1♀, 08.VII.2011 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.16. 114 **Megachile (Xanthosarus) circumcincta (Kirby, 1702) Экология и биология Сезон лёта: июнь – июль. Трофические связи: олиголектичный вид, посещает цветки Fabaceae [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. В КСП отмечен также на Lamiaceae (Phlomis tuberosa). Гнездовое поведение: гнездятся в почве, выкапывая норки или используя готовые полости и отверстия. Гнездо линейного типа, содержит 3–5 ячеек. Строение ячейки – типичное для представителей рода. Вход в гнездо заполняется почвой или песком [Banaszak J., Romasenko L., 1998; Gogala A., 1999]. Местообитание: в КСП встречается на суходольных разнотравно-злаковых лугах. Распространение Голарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Томская обл. [Конусова О.Л., Янюшкин В.В., 2000], Алтайский край [Шумакова П.С. и др., 1982], Кемеровская обл. [Еремеева Н.И. и др., 2011]); Восточная Сибирь (Иркутская обл., Бурятия, Забайкальский край, Якутия [Cockerell T. D., 1928; Давыдова Н.Г., Песенко Ю.А., 2002; Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Европа, Турция, Монголия [Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001]. Материал КК: Беловский р-н, Караканский хребет (сев. отрог), 02.VI.2008 – 3♀♀ (ДС), 4 км ЮВ с. Уроп, 20.VII.2006 – 5♀♀ (ДС, СЯ), 21.VII.2006 – 2♀♀ (ДС, СЯ), 22.VII.2006 – 2♀♀ (СЯ). Точки сбора: указаны на рис. 4.1.16. 115 Megachile (Xanthosarus) lagopoda (Linnaeus, 1761) Экология и биология Сезон лёта: июнь – август. Трофические связи: вид полилектичен, посещает главным образом цветки растений сем. Asteraceae (Carduus, Centaurea, Helianthus, Senecio) [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001; Ban C., 2005; Guler Y., Cağatay N., 2006; Matache I., Ban C., 2006]. На территории КСП отловлен на Centaurea. Гнездовое поведение: полостях. Гнёзда гнёзда размещают в песчаной почве, в готовых линейного типа или линейно-ветвистые, строительные материалы и структура гнезда типичная для представителей данного рода. Ячеек от 2 до 12 [Осычнюк А.З. и др., 1978; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Ban C., 2005; Matache I., Ban C., 2006]. Местообитание: в КСП представители данного вида приурочены к суходольным разнотравно-злаковым лугам и залежам. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983], Алтайский край [Wnukowskij, W., 1927], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (респ. Хакасия [Прощалыкин М.Ю., 2013 б], Бурятия, Забайкальский край) [Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Восточная и Центральная Европа, Северная Африка, Турция, Кавказ, Средняя Азия, Япония [Ozbek H., Zanden G.,1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005; Guler Y., Cağatay N., 2006; Hausl-Hofstatter U., 2007]. Материал ГШ: Таштагольский р-н, 6 км ЮЗ п. Мундыбаш, 03.VII.2005 – 2♀♀ (ДС). КК: Беловский р-н, окр. г. Белово, 21.VIII.2007 – 4♀♀, 23.VIII.2007 – 1♂ (ДС); Гурьевский р-н, окр. г. Гурьевск, 20.VIII.2007 – 3♀♀ (ДС); Кемеровский 116 р-н, д. Сухая Речка, 15.VI.2005 – 1♀ (НЕ), Ленинск-Кузнецкий р-н, г. ЛенинскКузнецкий, 29.VII.2006 – 5♀♀ (СЯ), 30.VII.2006 – 2♀♀, 1♂ (СЯ); Прокопьевский р-н, 5 км СВ д. Большая Талда, 18.VII.2004 – 1♀, 2 км СВ г. Прокопьевск, 02.VIII.1999 – 1♂, 30.VII.2004 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.17. Рисунок 4.17. Точки сбора отдельных видов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции 117 Megachile (Xanthosarus) maakii Radozskovski, 1874 Экология и биология Сезон лёта: июнь – июль. Трофические связи: трофические предпочтения – Fabaceae [Banaszak J., Romasenko L., 1998]. На территории КСП отмечено посещение Vicia megalotropis, V. sylvatica, Trifolium pratense, из других семейств – Potentilla argentea и P. chrysantha (Rosaceae). Гнездовое поведение: самки устраивают гнёзда в готовых полостях в различных субстратах. Местообитание: в КСП приурочен к суходольным разнотравно-злаковым лугам, а также остепнённым участкам. Распространение Транспалеарктический бореальный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Кемеровская обл.) [Еремеева Н.И., Сидоров Д.А., 2006]; Восточная Сибирь (Иркутская обл., Бурятия, Якутия) [Романькова, 1995; Proshchalykin M., 2007]; Дальний Восток России [Proshchalykin M., 2004]. Юго-Восточная, Восточная и Северная Европа, Монголия [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998]. Материал ГШ: Таштагольский р-н, окр. п. Каз, 29.VI.2010 – 2♀♀, 1♂, 04.VII.2010 – 2♀♀ (ДС), 6 км ЮЗ п. Мундыбаш, 03.VII.2005 – 2♂♂ (ДС), окр. п. Тельбес, 12.VII.2003 – 4♀♀, 1♂ (ДС), 2 км ЮЗ п. Усть-Кабырза, 03.VII.2008 – 1♀, 04.VII.2008 – 1♀ (ДС), пр. б. р. Кондома (52° 55,675' с. ш., 87° 09,756' в. д.), 10.VI.2011 – 1♀ (ДС), пр. б. р. Кондома (52° 59,768' с. ш., 87° 04,017' в. д.), 12.VI.2011 – 2♀♀ (ДС), 13.VI.2011 – 3♀♀ (ДС). КА: Междуреченский р-н, 9 км от ст. Лужба, 11.VII.1997 – 1♂ (НЕ); Хакасия, Аскизский р-н, 11 км СЗ п. Балыксу, 10.VII.2004 – 2♀♀ (НЕ). КК: Беловский р-н, Караканский хребет, 12.VI.2006 – 1♀ (СЯ); Гурьевский р-н, 7 км ЮЗ г. Гурьевск, 02.VII.2007 – 2♀♀ (СЛ); Прокопьевский р-н, 118 ЮВ окр. г. Киселёвск, 23.VI.2006 – 3♀♀ (ДС); Промышленновский р-н, 4 км СЗ с. Журавлёво, 12.VII.2006 – 1♂ (СЯ). Точки сбора: указаны на рис. 4.1.17. Megachile (Xanthosarus) willughbiella (Kirby, 1702) Экология и биология Сезон лёта: июнь – август. Трофические связи: полилект. Предпочитает Asteraceae (Carduus, Cirsium, Centaurea, Tragopogon) и Fabaceae (Lotus, Trifolium, Lathyrus, Lupinus) [Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Gogala A., 1999; Banaszak J. et al., 2001; Ban C., 2005]. На территории КСП отмечено посещение особями данного вида растений из сем. Crassulaceae (Sedum hybridum), Fabaceae (Lathyrus pratensis, Medicago falcate, Vicia multicaulis, V. cracca, Trifolium pratense), Onagraceae (Chamerion angustifolium), Primulaceae (Lysimachia vulgaris), Ranunculaceae (Delphinium sp., Ranunculus polyanthemos), Rosaceae (Fragaria vesca), Geranium pretense (Geraniaceae). Гнездовое поведение: для размещения гнёзд используют полости в мёртвой древесине и стеблях растений. Гнёзда линейные; ячейки стеклоподобные, количество их может варьироваться от 6 до 8 [Banaszak J., Romasenko L., 1998; Gogala A., 1999; Ban C., 2005]. Местообитание: в КСП представители данного вида встречаются, как правило, на суходольных и остепнённых разнотравно-злаковых лугах. Распространение Транспалеарктический полизональный вид. PA. Россия: европейская часть [Осычнюк А.З. и др., 1978]; Западная Сибирь (Ханты-Мансийский автономный округ [T.V. Levchenko, K.P. Tomkovich, 2014], Кемеровская обл. [Еремеева Н.И., Сидоров Д.А., 2006]); Восточная Сибирь (Иркутская обл. [Cockerell T. D., 1928], Бурятия [Proshchalykin M., 2007], Якутия [Давыдова Н.Г., Песенко Ю.А., 2002]); Дальний Восток России [Proshchalykin M., 119 2004]. Европа, Турция, Кавказ, Япония [Ozbek H., Zanden G., 1994; Monsevićius V., 1995; Banaszak J., Romasenko L., 1998; Banaszak J. et al., 2001; Ban C., 2005]. Материал ГШ: Таштагольский р-н, окр. п. Каз, 20.VI.2010 – 4♂♂ (ДС), р. Мундыбаш, окр. п. Клёпочный, 07.VIII.2007 – 1♂ (ДС), ср. теч. 6 км ЮЗ п. Мундыбаш, 07.VII.2005 – 1♂ (СЯ), окр. п. Тельбес, 04.VII.2003 – 2♀♀ (ДС), окр. п. Темир-Тау, 04.VIII.2007 – 2♂♂, 2 км ЮЗ п. Усть-Кабырза, 03.VII.2008 – 7♀♀ (ДС, СЯ), 06.VII.2008 – 1♂ (ДС), 10.VII.2008 – 1♂ (СЯ), 11.VII.2008 – 1♀ (ДС), 12.VII.2008 – 4♀♀, 2♂♂ (СЯ), окр. п. Шерегеш, 3 км СВ г. Мустаг, 27.VII.1999 – 3♀♀ (НЕ), 30.VII.1999 –1♀ (ДВ), пр. б. р. Кондома (52° 59,767' с. ш., 87° 04,019' в. д.), 12.VI.2011 – 1♂ (ДС), пр. б. р. Кондома (52° 59,768' с. ш., 87° 04,017' в. д.), 12.VI.2011 – 2♂♂ (ДС), 11 км С от устья р. Антроп, 31.VII.2011 – 2♂♂ (ДС). КА: Крапивинский р-н, территория биостанции КемГУ «Ажендарово», 10.VIII.2008 – 3♀♀, 1♂ (ДС); Тисульский р-н, окр. п. Макаракский, 05.VII.2007 – 4♂♂, 09.VII.2007 – 2♂♂ (ДС), 10 км С п. Полуторник, 04.VII.2009 – 2 ♀♀, 2♂♂ (СЯ). КК: Беловский р-н, Караканский хребет (сев. отрог), 02.VI.2008 – 2♀♀ (ДС); Гурьевский р-н, окр. г. Гурьевск, 28.VIII.2007 – 1♀ (ДС); Кемеровский р-н, г. Кемерово, 03.VI.2004 – 3♂♂ (НЕ), 21.VI.2007 – 1♂ (ДС), 23.VI.2010 – 1♂ (СЯ); Прокопьевский р-н, 5 км СВ д. Большая Талда, 18.VII.2004 – 2♂♂ (ДС), СВ окр. г. Прокопьевск, 28.VII.2006 – 1♀ (ДС); Промышленновский р-н, 6 км СЗ с. Журавлёво, 24.VI.2000 –1♀ (НЕ), 26.VI.2000 – 4♀♀ (НЕ), 4 км СЗ с. Журавлёво, 28.VI.2006 – 1♂ (СЯ), 30.VI.2006 – 2 ♀♀, 2♂♂ (СЯ), 30.VI.2006 – 1♀ (ДС), 02.VII.2006 – 5♀♀, 1♂ (ДС, СЯ), 08.VII.2006 – 1♀ (ДС). Точки сбора: указаны на рис. 4.1.17. 120 4.2. Видовое разнообразие и численное обилие пчёл сем. Megachilidae Кузнецко-Салаирской провинции В результате проведённых исследований в фауне Кузнецко-Салаирской провинции выявлено 49 видов пчёл семейства Megachilidae, относящихся к 12 родам подсемейства Megachilinae: Lithurgus (1 вид), Chelostoma (1), Hoplitis (6), Osmia (6), Anthidiellum (1), Anthidium (3), Bathanthidium Trachusa (1), Aglaoapis (1), Coelioxys (10), Megachile (1), Stelis (3), (15). Из них 1 род (Bathanthidium Mavromoustakis, 1953) и 6 видов (12 % от общего числа) указаны впервые для фауны Западной Сибири: Bathanthidium sibiricum, Coelioxys afra, C. lanceolata, C. quadridentata, Osmia leaiana, O. parietina. Для КузнецкоСалаирской провинции впервые указаны 13 видов (табл. 4.1). Таблица 4.1 – Видовой состав и численное обилие (%) пчёл семейства Megachilidae Кузнецко-Салаирской провинции Род Подрод Вид 1 2 3 Lithurgus Lithurgus s. str. Chelostoma Hoplitis Gyrodromella Alcidamea Osmia Monumetha Helicosmia Hoplosmia Melanosmia Neosmia Anthidiellum Anthidium Anthidiellum s. str. Anthidium s. str. Bathanthidium Stelis Stenanthidiellum Stelis s. str. Lithurgini L. fuscipennis Osmiini Ch. rapunculi H. acuticornis** H. claviventris H. leucomelana H. mitis H. tridentata H. tuberculata O. leaiana* O. spinulosa O. parietina * O. pilicornis** O. uncinata** O. bicolor Anthidiini A. strigatum** A. manicatum A. punctatum A. septemspinosum B. sibiricum * S. punctulatissima S. simillima Численное обилие, % 4 Класс обилия 5 3,79 сд 5,33 1,72 2,26 1,99 2,62 2,89 0,36 сд сд сд сд сд сд ор 0,09 ор 2,62 2,17 2,53 сд сд сд 0,09 ор 3,07 сд 1,63 0,91 0,27 3,88 0,54 0,36 1,63 сд р ор сд р ор сд 121 Окончание таблицы 4.1. 1 2 3 Trachusa Trachusa s. str. Aglaoapis Aglaoapis s. str. Coelioxys Allocoelioxys Boreocoelioxys Coelioxys s. str. Megachile Eumegachile Eutricharaea Megachile s. str. Xanthosarus S. ornatula** T. byssina Dioxyini A. tridentatus Megachilini C. afra * C. emarginata C. alata C. inermis C. mandibularis C. rufescens C. conoidea C. elongata C. lanceolata * C. quadridentata * M. bombycina M. argentata M. rotundata M. alpicola M. centuncularis** M. genalis M. lapponica M. ligniseca M. melanopyga M. versicolor M. analis M. circumcincta** M. lagopoda M. maakii M. willughbiella 4 0,09 1,99 5 ор сд 0,27 ор 0,09 0,27 0,35 6,23 1,44 1,98 0,27 0,45 0,09 0,54 1,63 0,09 0,99 1,91 0,91 2,44 3,52 18,61 ор ор ор д р сд ор р ор р сд ор р сд р сд сд д 0,72 р 2,44 2,81 0,72 1,63 1,89 4,88 сд сд р сд сд сд Примечание: * – вид впервые указан для Западной Сибири. ** – вид впервые указан для Кузнецко-Салаирской провинции. д – доминант, сд – субдоминант, р – редкий, ор – очень редкий. Индекс видового разнообразия (вероятность межвидовых встреч) PIE' населения пчёл сем. Megachilidae Кузнецко-Салаирской провинции равен 0,941 (биоразнообразие приближается к максимальному). Индекс разнообразия Megachilidae представлено Шеннона Ish' для исследуемой территории равен 1,438. На исследованной территории семейство единственным подсемейством Megachilinae, объединяющим 5 триб (Lithurgini, Osmiini, Anthidiini, Dioxyini, Megachilini). Наибольшее число родов мегахилид, обитающих в Кузнецко-Салаирской провинции, относится к трибе Anthidiini 122 (включает 5 родов), значительно уступают ей трибы Osmiini и Megachilini (включают 3 и 2 рода соответственно). В состав триб Lithurgini и Dioxyini на исследованной территории входит по 1 роду, каждый из которых включает 1 вид. На исследуемой территории наиболее богата видами триба Megachilini. К двум родам этой трибы (Megachile и Coelioxys) относится половина видов мегахилид, обнаруженных на исследуемой территории (25 видов, 52 %). Уступают ей по числу видов трибы Osmiini (13 видов, 26 %) и Anthidiini (9; 18 %). На долю триб Lithurgini и Dioxyini приходится 5 % от общего числа видов (рис. 4.2.1). 100% 90% 80% 70% Lithurgini 60% Dioxyni 50% Osmiini 40% Anthidiini 30% Megachilini 20% 10% 0% 1 2 3 Рисунок 4.2.1. Родовое (1), видовое (2) и численное (3) обилие триб пчёл сем. Megachilidae в Кузнецко-Салаирской провинции, % По численному обилию в фауне мегахилид на исследованной территории также выделяются представители трибы Megachilini. К данной трибе относится 57 % от общего числа отловленных особей. Менее представлены в сборах трибы Osmiini (27,7 % особей) и Anthidiini (11,3 %). На долю триб Lithurgini и Dioxyini приходится 3,8 и 0,3 % соответственно от общего числа отловленных мегахилид. 123 Наибольшим видовым богатством в фауне Кузнецко-Салаирской провинции отличается род Megachile – на исследованной территории обнаружено 15 видов (31 % от их общего числа) (рис. 4.2.2, а). Lithurgus 2% Chelostoma 2% Trachusa 2% Hoplitis 12% Megachile 31% Osmia 12% Anthidiellum 2% Aglaoapis 2% Anthidium 6% Coelioxys 21% Stelis 6% Bathanthidium 2% а Chelostoma 5,33 % Lithurgus 3,79 % Trachusa 1,99 % Hoplitis 11,84 % Megachile 45,19 % Osmia 10,57 % Anthidiellum 1,63 % Aglaoapis 0,27 % б Coelioxys 11,71 % Stelis 2,08 % Anthidium 5,06 % Bathanthidium 0,54 % Рисунок 4.2.2. Видовое (а) и численное (б) обилие родов пчёл сем. Megachilidae в Кузнецко-Салаирской провинции, % 124 Род Megachile доминирует и по численному обилию – к нему принадлежит 45 % от общего числа мегахилид, отловленных на данной территории (рис. 4.2.2, б). На территории Кузнецко-Салаирской провинции род Megachile представлен 4 подродами, включающими от 1 до 7 видов (табл. 4.1, рис. 4.2.3). Eumegachile Eutricharaea Xanthosarus Megachile s. str. 0 5 10 15 Видовое обилие, % 20 25 30 35 Численное обилие, % Рисунок 4.2.3. Видовое и численное обилие подродов рода Megachile в Кузнецко-Салаирской провинции (% от общих сборов) Наибольшее число видов данного рода относится к подродам Megachile s. str. (7 видов) и Xanthosarus (5). Эти же подроды характеризуются наибольшими значениями суммарного численного обилия видов. К подроду Megachile s. str. принадлежит более 30 % отловленных на территории Кузнецко-Салаирской провинции особей пчёл-мегахилид и 67 % представителей данного рода. К подроду Xanthosarus относятся 12 % от общих сборов мегахилид и 26 % отловленных представителей рода Megachile. Подроды Eutricharaea и Eumegachile на исследуемой территории представлены двумя и одним видом соответственно, их доля от общих сборов пчёл сем. Megachilidae составляет менее 3 % (6 % от числа отловленных представителей данного рода). Вклад представителей различных подродов в видовое и численное обилие р. Megachile показан на рис. 4.2.4. 125 а б Рисунок 4.2.4. Вклад подродов Megachile s. str., Xanthosarus, Eutricharaea, Eumegachile в видовое (а) и численное (б) обилие р. Megachile в КузнецкоСалаирской провинции, % Наибольшим численным обилием среди видов рода Megachile отличается M. ligniseca (подрод Megachile s. str.). Представители этого вида составляют 18 % от общих сборов мегахилид и 41 % от числа отловленных представителей рода. Значительный вклад в численное обилие представителей данного рода вносит также M. (Xanthosarus) willughbiella (4,8 % от общего числа отловленных особей мегахилид, 11 % от отловленных представителей р. Megachile). Численное обилие остальных видов данного рода не превышает 4 % от общих сборов. Наиболее малочисленным в сборах является M. argentata (менее 0,1 %). К роду Coelioxys относится 10 видов пчёл сем. Megachilidae, что составляет 20 % видового богатства мегахилид Кузнецко-Салаирской провинции. На данный род приходится 11,7 % от числа отловленных представителей сем. Megachilidae. На исследуемой территории данный род представлен 3 подродами (табл. 4.1, рис. 4.2.5). Подроды Coelioxys s. str. и Boreocoelioxys включают по 4 вида. По наибольшим значениям суммарного численного обилия видов выделяется подрод Boreocoelioxys. К нему относится 85 % отловленных представителей рода и 10 % от общих сборов мегахилид. Представители подрода Coelioxys s. str. составляют 11,5 % от отловленных представителей рода и 1,3 % от общих сборов мегахилид. 126 Coelioxys s. str. Boreocoelioxys Allocoelioxys 0 2 4 Видовое обилие, % 6 8 10 12 Численное обилие, % Рисунок 4.2.5. Видовое и численное обилие подродов рода Coelioxys в Кузнецко-Салаирской провинции (% от общих сборов) Подрод Allocoelioxys, включающий на территории провинции 2 вида, вносит незначительный вклад в численное обилие мегахилид (менее 0,5 % от общих сборов и 3 % от отловленных представителей р. Coelioxys). На рис. 4.2.6 показана роль подродов в формировании видового и численного обилие р. Coelioxys. а б Рисунок 4.2.6. Вклад подродов Coelioxys s. str., Boreocoelioxys и Allocoelioxys в видовое (а) и численное (б) обилие р. Coelioxys в Кузнецко-Салаирской провинции, % 127 Численное обилие рода Coelioxys обеспечивает главным образом C. inermis (представители данного вида составляют 53 % от собранных представителей рода и 6 % от числа отловленных особей мегахилид). Также по численному обилию выделяются C. rufescens (17 и 2 % соответственно) и C. mandibularis (12 и 1,4 %). Численное обилие каждого из остальных видов не превышает 0,55 % от общих сборов. Наименее представлены в сборах представители видов C. afra и C. lanceolata (доля каждого не превышает 0,1 % от общих сборов мегахилид на исследуемой территории). Триба Osmiini уступает по числу отловленных особей, составляющему 27,6 % от общих сборов (рис. 4.2.1). К трём родам данной трибы относится четверть видов мегахилид, обнаруженных в Кузнецко-Салаирской провинции. Роды Hoplitis и Osmia включают по 6 видов, род Chelostoma представлен 1 видом. Суммарное численное обилие представителей рода Hoplitis составляет 11,8 % от общих сборов (рис. 4.2.2, а, б). На исследуемой территории род представлен 2 подродами – Alcidamea (включает 5 видов) и Monumetha (1 вид) (табл. 4.1, рис. 4.2.7). Monumetha Alcidamea 0 2 4 Видовое обилие, % 6 8 10 12 14 Численное обилие, % Рисунок 4.2.7. Видовое и численное обилие подродов рода Hoplitis в Кузнецко-Салаирской провинции (% от общих сборов) 97 % отловленных представителей рода Hoplitis относится к подроду Alcidamea; их суммарное численное обилие составляет 11,5 % от общего сбора мегахилид. Подрод Monumetha представлен единственным видом H. tuberculata и составляет менее 0,4 % от числа отловленных мегахилид. 128 Наибольшее численное обилие из видов рода Hoplitis демонстрируют H. tridentata и H. mitis. На эти виды приходится 2,8 и 2,6 % соответственно от общего числа собранных мегахилид и 24 и 22 % от отловленных представителей рода. Редки в сборах представители вида H. tuberculata (0,36 % от сборов). Род Osmia составляет 12 % видового богатства мегахилид исследуемой территории и 10,5 % от общих сборов пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции (рис. 4.2.2, а, б). Этот род включает в себя 6 видов из четырёх подродов (рис. 4.2.8). Подрод Melanosmia включает 3 вида и характеризуется наибольшим численным обилием (4,8 % от общих сборов, 45,3 % от числа собранных представителей рода). Подрод Neosmia включает 1 вид O. bicolor, численное обилие которого составляет 3 % от общих сборов и 29 % от числа отловленных представителей рода. Данный вид является наиболее многочисленным в сборах пчёл р. Osmia на исследуемой территории. Helicosmia Hoplosmia Melanosmia Neosmia 0 1 2 3 Видовое обилие, % 4 5 6 7 Численное обилие, % Рисунок 4.2.8. Видовое и численное обилие подродов рода Osmia в КузнецкоСалаирской провинции (% от общих сборов) Подрод Hoplosmia с единственным видом O. spinulosa составляет 2,6 % в сборах мегахилид и 24,7 % в сборах представителей рода. Самым малочисленным в сборах является подрод Helicosmia, представленный 129 на исследуемой территории единственным видом O. leaiana (0,9 % от представителей рода Osmia, 0,09 % от общих сборов мегахилид). Вклад представителей различных подродов в видовое и численное обилие р. Osmia показан на рис. 4.2.9. а б Рисунок 4.2.9. Вклад подродов Melanosmia, Neosmia, Hoplosmia и Helicosmia в видовое (а) и численное (б) обилие р. Osmia в Кузнецко-Салаирской провинции, % Род Chelostoma на территории Кузнецко-салаирской провинции представлен 1 подродом Gyrodromella с единственным видом Ch. rapunculi. Представители данного вида составляют 5,3 % от общего числа особей мегахилид, отловленных на исследуемой территории (рис. 4.2.2, б). Триба Anthidiini в Кузнецко-Салаирской провинции представлена наибольшим числом родов (рис. 4.2.1). Большая часть видов (3 вида) относится к роду Anthidium, представленному на исследуемой территории номинативным подродом. Численное обилие представителей рода составляет 5 %. Наибольший вклад в численное обилие представителей рода вносит A. septemspinosum (3,8 % от общих сборов, 76,8 % от отловленных представителей рода Anthidium). Роды Anthidiellum, Bathanthidium и Trachusa в Кузнецко-Салаирской провинции включают по 1 виду и их суммарное численное обилие составляет 4 % от числа отловленных на исследуемой территории мегахилид. 130 Представители клептопаразитического рода Stelis составляют 2 % от общего числа отловленных пчёл-мегахилид. Данный род на исследуемой территории представлен 3 видами, наибольшим численным обилием из которых выделяется S. simillima (1,6 % от общих сборов). Суммарное численное обилие остальных видов рода невелико (0,45 %). На рис. 4.2.10 показана роль разных родов в формировании видового и численного обилия трибы Anthidiini. а б Рисунок 4.2.10. Вклад родов Anthidium, Anthidiellum, Bathanthidium, Trachusa и Stelis в видовое (а) и численное (б) обилие трибы Anthidiini в КузнецкоСалаирской провинции, % Триба Lithurgini на исследуемой территории представлена единственным родом Lithurgus, включающим 1 вид. На него приходится 3,8 % численного обилия пчёл сем. Megachilidae, отловленных в Кузнецко-Салаирской провинции. Триба Dioxyini на данной территории также включает 1 вид (Aglaoapis tridentatus) с численным обилием 0,27 %. При оценке относительного обилия фауны пчёл сем. Megachilidae по логарифмической шкале (Песенко, 1982 б) в Кузнецко-Салаирской провинции выделены следующие классы обилия: доминантные виды, субдоминантные виды, редкие и очень редкие виды (рис. 4.3.1). 131 70 65,25 60 51 50 40 30 26,5 24,84 18,4 20 10 7,22 4,1 2,69 0 Доминанты Субдоминанты Видовое обилие, % Редкие Очень редкие Численное обилие, % Рисунок 4.3.1. Видовое и численное обилие различных классов обилия пчёл сем. Megachilidae в Кузнецко-Салаирской провинции, % К классу доминантных видов (более 6,06 % от общего объёма сборов мегахилид) на территории Кузнецко-Салаирской провинции отнесены 2 вида: Megachile ligniseca и Coelioxys inermis. Доминантные виды составляют 4,1 % от общего числа видов мегахилид исследуемой территории; их суммарное численное обилие 24,84 % от общих сборов. К субдоминантным (от 1,45 % до 6,05 %) относятся 25 видов: Lithurgus fuscipennis, Chelostoma rapunculi, Hoplitis acuticornis, H. claviventris, H. leucomelana, H. mitis, H. tridentatа, Osmia spinulosa, O. parietina, O. pilicornis, O. bicolor, Anthidiellum strigatum, Anthidium septemspinosum, Stelis simillima, Trachusa byssina, Coelioxys rufescens, M. bombycina, M. alpicola, M. genalis, M. lapponica, M. versicolor, M. analis, M. lagopoda, M. maakii, M. willughbiella. Они составляют 51 % от общего числа видов и 65,25 % от общего числа отловленных особей. К редким видам мегахилид (от 0,37 % до 1,44 %) отнесены 9 видов: Anthidium manicatum, Bathanthidium sibiricum, Coelioxys mandibularis, 132 C. elongata, C. quadridentata, M. rotundata, M. centuncularis, M. melanopyga, M. circumcincta. Их вклад в видовое богатство мегахилид исследуемой территории составляет 18,4 %, суммарное численное обилие – 7,22 % от объема сборов. К очень редким (0,36 % и менее) отнесены 13 видов (26,5 % от общего числа видов): H. tuberculata, O. leaiana, O. uncinata, Anthidium punctatum, S. punctulatissima, Stelis ornatula, Aglaoapis tridentatus , C. afra, C. emarginata, C. alata, C. conoidea, C. lanceolata, M. argentata. Суммарное численное обилие видов данного класса – 2,69 % от числа особей мегахилид, отловленных в Кузнецко-Салаирской провинции. Более половины видов (61,5 %), относимых к этой категории, являются видами-клептопаразитами. 4.3. Зоогеографическая характеристика фауны пчёл семейства Megachilidae При выделении ареалогических групп видов пчёл-мегахилид, обитающих в Кузнецко-Салаирской провинции рассматривали долготные [Городков К.Б., 1984] и широтные [Сергеев М.Г., 1986] составляющие ареала. При рассмотрении долготной составляющей ареала выявлено 4 ареалогические группы по типам голарктического распространения (табл. 4.2). Подавляющее число видов мегахилид исследуемой территории (31 вид, 63,2 % от их общего числа), принадлежит к транспалеарктической ареалогической группе (Hoplitis claviventris, H. leucomelana, H. mitis, H. tuberculata, Osmia parietina, O. leaiana, O. uncinata, Anthidium punctatum, Anthidium septemspinosum, Stelis ornatula, Trachusa byssina, Aglaoapis tridentatus, Coelioxys afra, C. alata, C. conoidea, C. elongata, C. emarginata, C. inermis, C. lanceolata, C. mandibularis, C. rufescens, C. quadridentata, Megachile alpicola, M. analis, M. bombycina, M. lagopoda, M. ligniseca, M. maakii, M. versicolor, M. willughbiella, M. melanopyga). 133 Таблица 4.2 – Ареалогические группы пчёл семейства Megachilidae Кузнецко-Салаирской провинции Вид 1 Долготная группа 2 Широтная группа 3 ЗЦП П Г ЗЦП ТП ТП ТП ЗЦП ТП ЗЦП ТП ЗЦП ЗЦП ТП ТП П П П П П П П П П П П П Б Г Г* ТП ТП ЦВП ТП ЗЦП ЗЦП ТП П П П П П П П СА П ТП СБ ТП ТП ТП ТП ТП ТП ТП ТП ТП ТП ТП ТП Г ТП Г СА Б П П СА П Б П П П Б П СБ П П Lithurgini L. fuscipennis Osmiini Ch. rapunculi H. acuticornis H. claviventris H. leucomelana H. mitis H. tridentata H. tuberculata O. bicolor O. parietina O. pilicornis O. spinulosa O. leaiana O. uncinata Anthidiini Anhidiellum strigatum A. manicatum A. punctatum A. septemspinosum B. sibiricum S. ornatula S. punctulatissima S. simillima T. byssina Dioxyini A. tridentatus Megachilini C. afra C. alata C. conoidea C. elongata C. emarginata C. inermis C. lanceolata C. mandibularis C. rufescens C. quadridentata M. alpicola M. analis M. argentata M. bombycina M. centuncularis 134 Окончание таблицы 4.2 1 2 3 M. circumcincta Г П M. genalis Г СБ M. lagopoda ТП П M. lapponica Г П M. ligniseca ТП СБ M. maakii ТП Б M. rotundata Г* П M. versicolor ТП П M. willughbiella ТП П M. melanopyga ТП П Примечание: долготные группы: Г – голарктическая, ТП – транспалеарктическая, ЗЦП – западно-центрально-палеарктическая, ЦВП – центрально-восточно-палеарктическая; широтные группы: П – полизональная, Б – бореальная, СБ – суббореальная, СА – субаридная. * – виды, интродуцированные в другие зоогеографические регионы. Транспалеарктическая ареалогическая группа характеризуется максимальным численным обилием, составляющим 64,3 % от общего числа отловленных особей мегахилид (рис. 4.4.1). 70 63,2 64,31 60 50 40 30 20 18,36 16,54 16,3 18,61 10 2,04 0,54 0 Г ТП Видовое обилие, % ЗЦП ЦВП Численное обилие, % Рисунок 4.4.1. Видовое и численное обилие долготных ареалогических групп пчёл сем. Megachilidae в Кузнецко-Салаирской провинции, % Обозначения: Г – голарктическая, ТП – транспалеарктическая, ЗЦП – западно-центральнопалеарктическая, ЦВП – центрально-восточно-палеарктическая. 135 К этой группе принадлежат оба вида-доминанта, отмеченных на исследуемой территории, более половины видов-субдоминантов (60 %), 44 % редких видов и подавляющее большинство очень редких видов мегахилид (84 %). Доминанты составляют 38 % численного обилия данной группы, субдоминанты – 53 %, редкие – 5,5 % , очень редкие виды – 3,5 %. К голарктической ареалогической группе относится 9 видов, что составляет 18,4 % от общего числа видов мегахилид, обнаруженных на исследуемой территории, и 16,5 % от общих сборов (Chelostoma rapunculi, Anhidiellum strigatum, A. manicatum, Megachile argentata, M. centuncularis, M. circumcincta, M. genalis, M. lapponica, M. rotundata). В эту ареалогическую группу входит 16 % всех субдоминантных видов, обитающих на территории Кузнецко-Салаирской провинции, 44 % редких видов и 7,7 % очень редких видов. Вклад субдоминантов в численное обилие группы составляет 78 %, редких видов – 21,3 %, очень редких видов – 0,7 %. Большинство голарктических видов пчёл сем. Megachilidae случайно были завезены в Северную Америку из Европы человеком с различными строительными и иными материалами, что возможно вследствие способности данной группы пчёл гнездиться в различных субстратах. Представители двух голарктических видов мегахилид в настоящее время интродуцированы также в Неотропический и Австралийский зоогеографические регионы в качестве эффективных опылителей люцерны и других культур (A. manicatum и M. rotundata) [Радченко В.Г., Песенко Ю.А., 1994]. Западно-центрально-палеарктическая ареалогическая группа представлена 8 видами (16,3; 18,6) (Lithurgus fuscipennis, Hoplitis acuticornis, H. tridentata , Osmia bicolor, O. pilicornis, O. spinulosa, Stelis punctulatissima, S. simillima). К настоящему времени данные виды (за исключением O. spinulosa) не отмечены восточнее Кузнецко-Салаирской провинции на территории России и Палеарктики. 136 Эта ареалогическая группа включает в себя 28 % всех субдоминантных видов, обнаруженных на исследуемой территории, и 7,7 % очень редких видов. Субдоминанты составляют 98 % численного обилия данной ареалогической группы, очень редкие виды – 2 %. К центрально-восточно-палеарктической группе принадлежит только 1 вид (Bathanthidium sibiricum) (2; 0,5), относящийся к категории редких видов. Западнее Кузнецко-Салаирской провинции данный вид отмечен только на территории Алтайского края [Прощалыкин М.Ю., 2013]. Проведённые исследования позволили расширить границы ареалов 2 видов на восток (указаны крайние восточные точки нахождения): H. acuticornis – Крапивинский р-н, окр. биостанции КемГУ «Ажендарово», 54° 45' с.ш., 87° 02' в.д. (ранее – Томское Приобье [Конусова О.Л., Янюшкин В.В., 2000]); O. pilicornis – Таштагольский р-н, окр. п. Каз, 53° 09' с.ш., 87° 42' в.д. (ранее – Курганская обл. [Кузьмин П.М., Молчанов А.Е., 1983]). При анализе широтного распространения мегахилид выявлено 4 широтные группы ареалов (полизональная, бореальная, суббореальная и субаридная). Наибольшая часть видов мегахилид Кузнецко-Салаирской провинции относится к широтной ареалогической группе полизональных видов. В состав этой группы входит 37 видов пчёл сем. Megachilidae, что составляет 75,5 % от общего числа видов, и 72,3 % от числа отловленных особей (Lithurgus fuscipennis, Chelostoma rapunculi, Hoplitis acuticornis, H. claviventris, H. leucomelana, H. mitis, H. tridentata, H. tuberculata, Osmia bicolor, O. parietina, O. pilicornis, O. spinulosa, O. leaiana, Anhidiellum strigatum, Anthidium manicatum, A. punctatum, A. septemspinosum, Bathanthidium sibiricum, Stelis ornatula, S. punctulatissima, Trachusa byssina, Coelioxys conoidea, C. elongata, C. inermis, C. mandibularis, C. rufescens, C. quadridentata, Megachile analis, M. bombycina, M. centuncularis, M. circumcincta, M. lagopoda, M. lapponica, M. rotundata, M. melanopyga) (рис. 4.4.2). M. versicolor, M. willughbiella, 137 80 70 60 50 40 30 20 10 0 75,5 72,3 21,4 10,2 8,2 4,33 Полизональная Бореальная Видовое обилие, % Суббореальная 6,1 1,9 Субаридная Численное обилие, % Рисунок 4.4.2. Видовое и численное обилие (%) широтных ареалогических групп пчёл сем. Megachilidae в Кузнецко-Салаирской провинции. Северная граница их распространения проходит в зоне тундры или лесотундры, на юге представители этой группы могут проникать в зону полупустынь и южнее. К ареалогической группе полизональных видов относится один из двух отмеченных на исследуемой территории видов-доминантов (C. inermis), 84 % всех субдоминантных видов, 66,6 % редких видов 46 % очень редких видов. Доминанты составляют 8,6 % численного обилия данной ареалогической группы, субдоминанты – 79,6 %, редкие виды – 7,3 %, очень редкие виды – 4,5 %. Группа бореальных видов представлена 5 видами (10,2 % от общего числа видов; 4,33 % от общих сборов): Osmia uncinata, Coelioxys alata, C. lanceolata, Megachile alpicola, M. maakii). Распространение видов этой группы на севере совпадает с распространением полизональных, на юге эти виды не встречаются южнее лесостепной зоны в центральной Палеарктике и зоны широколиственных лесов в западной и восточной Палеарктике. 138 К этой группе принадлежат 8 % видов-субдоминантов, отмеченных на исследуемой территории, и 23 % очень редких видов мегахилид. Субдоминанты составляют 87,5 % численного обилия данной группы, очень редкие виды – 12,5 %. В широтную группу суббореальных видов включены 4 вида (8,2 % от общего их числа, 21,4 % от общих сборов) (Aglaoapis tridentatus, Megachile argentata, M. genalis, M. ligniseca). Виды этой группы не выходят за пределы таёжной зоны на севере и лесостепной зоны – на юге. В данную ареалогическую группу входит один из двух доминантных видов Кузнецко-Салаирской провинции (M. ligniseca), один субдоминантный вид (M. genalis) и 2 вида из категории очень редких. Вклад вида-доминанта в численное обилие ареалогической группы составляет 87 %, вида-субдоминанта – 11,4 %, очень редких видов – 1,6 %. Cубаридная широтная ареалогическая группа представлена 3 видами (6,1 % от общего числа) и характеризуется наименьшим суммарным численным обилием (1,9 % от общих сборов) (Coelioxys afra, C. emarginata, Stelis simillima). Представители этих видов не проникают севернее лесостепи. Эта ареалогическая группа включает в себя 1 субдоминантный вид (Stelis simillima) и 2 вида из категории очень редких. Субдоминантный вид составляют 82 % численного обилия субаридной группы, очень редкие виды – 18 %. Сопоставление фауны мегахилид Кузнецко-Салаирской провинции с фаунами других территорий. К настоящему времени в пределах Западной Сибири наиболее изучен видовой состав мегахилид Курганской и Кемеровской областей [Кузьмин П.М., Молчанов А.Е., 1983; Сидоров Д.А, 2005; Еремеева Н.И., Сидоров Д.А., 2006; Яковлева С.Н., 2011, 2012]. Кроме того, предварительные данные по фауне пчёл сем. Megachilidae известны также для тайги Томского Приобья [Конусова О.Л., Янюшкин В.В., 2000], частично – для Алтайского края (юго-восточная часть Кулундинской степи [Шумакова П.С. и др., 1982], Каменский район [Wnukowskij, W., 1927] и окрестности г. Барнаул [Данилов Ю.Н., 2006]), окрестностей г. Омска [Лавров С.Д., 1927], 139 Новосибирской области и республики Алтай [Прощалыкин М.Ю., 2013 а, б]. Сопредельные с Кузнецко-Салаирской провинцией территории (Томская и Новосибирская области, Алтайский и Красноярский края, республики Алтай и Хакасия) в отношении видового состава пчёл сем. Megachilidae остаются недостаточно изученными, 80 % видов, отмеченных на данных территориях являются общими с фауной Кузнецко-Салаирской провинцией. На основе литературных данных и результатов собственных исследований установлено, что на территории изученных регионов Западной Сибири в целом обитает не менее 70 видов пчёл сем. Megachilidae, принадлежащих к 14 родам (табл. 4.3). Таблица 4.3 – Распределение видов пчёл сем. Megachilidae, отмеченных на территории Западной Сибири (по данным: Лавров С.Д., 1927; Wnukowskij, W., 1927; Кемеровская область 5. 6. 8. 9. 10. 11. 12. 13. 14. 15. Республика Алтай 4. Алтайский край 2. 3. Новосибирская область 1. 2 Lithurgus fuscipennis Chelostoma proximum rapunculi Heriades truncorum Hoplitis acuticornis adunca claviventris laevifrons leucomelana mitis nitidula papaveris robusta serrilabris Томская область 1 Виды Омская область № Курганская область Внуковский В.В., 1930; Шумакова П.С. и др., 1982; Кузьмин П.М., Молчанов А.Е., 1983; Конусова О.Л., Янюшкин В.В., 2000; Данилов Ю.Н., 2006; Яковлева С.Н., 2011, 2012; Прощалыкин М.Ю., 2013 а, б) Территории 3 4 5 6 7 8 9 + + + + + + + + + + + + + + + + + + + + + + + + + + + 140 Продолжение таблицы 4.3. 1 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30. 31. 32. 33. 34. 35. 36. 37. 38. 39. 40. 41. 42. 43. 44. 45. 46. 47. 48. 49. 50. 51. 52. 53. 54. 55. 2 tridentata tuberculata Osmia aurulenta bicolor bidentata leaiana parietina pilicornis spinulosa uncinata Antihidiellum strigatum Anthidium florentinum manicatum punctatum septemspinosum Bathanthidium sibiricum Icteranthidium laterale Stelis ornatula minima punctulatissima simillima Trachusa byssina Aglaoapis tridentatus Coelioxys afra acuminata alata aurolimbata brevis conoidea elongata emarginata inermis lanceolata mandibularis rufescens quadridentata Megachile alpicola analis apicalis argentata 3 + + 4 + + + 5 + + 6 7 8 + + + + + + + + + + + + + + + + + 9 + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + 141 Окончание таблицы 4.3. 1 56. 57. 58. 59. 60. 61. 62. 63. 64. 65. 66. 67. 68. 69. 70. 2 bombycina centuncularis circumcincta derasa ericetorum genalis lagopoda lapponica ligniseca maakii maritima melanopyga rotundata versicolor willughbiella Всего: 3 + + + 4 + + 5 + 6 + 7 8 + + + + + + + + + + + + + + + + + + + + + 42 9 22 + + + + + + 8 26 7 9 + + + + + + + + + + + + 49 По числу известных видов мегахилид фауна Западной Сибири уступает фауне европейской части России, насчитывающей около 100 видов [Никифорук К.С., 1957; Песенко Ю.А., 1971; Осычнюк А.З. и др., 1978; Ситдиков А.А., 1981; Левченко Т.Ю., 2010] и сходна с фаунами территорий изученной части Восточной Сибири (Якутия и Забайкалье) и Дальнего Востока России, насчитывающими 55 и 65 видов мегахилид соответственно [Давыдова Н.Г., Песенко Ю.А., 2002; Прощалыкин М.Ю., 2009 а, 2012]. Однако по видовому составу фауна мегахилид Западной Сибири проявляет большее сходство с фауной европейской части России (индекс сходства Чекановского-Съеренсена равен 0,72), чем с фауной восточно-азиатской части (индекс сходства менее 0,6). Фауна мегахилид Кузнецко-Салаирской провинции по числу видов и по видовому составу проявляет наибольшее сходство с фаунами регионов европейской части России – Удмуртии и Московской области (коэффициент сходства равен 0,7 и 0,69 соответственно) и фауной Забайкалья (0,7) (табл. 4.4). Это даёт возможность предположить, что фауна пчёл сем. Megachilidae КузнецкоСалаирской провинции по видовому составу занимает промежуточное положение между фаунами европейской и восточно-азиатской частей России. 142 Таблица 4.4 – Число видов пчёл сем. Megachilidae фаун регионов России и их сходство с фауной Кузнецко-Салаирской провинции Число видов пчёл сем. Megachilidae Число видов, общих с Кузнецко-Салаирской провинцией Московская область Удмуртия Башкирия Курганская область Забайкалье Якутия 60 48 59 42 51 27 38 34 31 27 35 21 Дальний Восток России 65 37 Регион России Индекс сходства ЧекановскогоСъеренсена 0,69 0,70 0,65 0,59 0,70 0,55 0,65 Таким образом, на территории Кузнецко-Салаирской провинции установлено обитание 49 видов пчёл семейства Megachilidae, относящихся к 1 подсемейству, 5 трибам и 12 родам. Из них для Западной Сибири впервые указаны 6 видов и 1 род, для Кузнецко-Салаирской провинции – 14 видов. В фауне мегахилид Кузнецко-Салаирской провинции по видовому и численному обилию значительно преобладают представители трибы Megachilini. К данной трибе принадлежат более половины видов (25 видов, 51 %), отмеченных на исследуемой территории, и 57 % от общего числа отловленных особей. Наибольшее видовое обилие демонстрируют роды Megachile (включает 15 видов, 30 % от общего числа видов) и Coelioxys (10 видов, 11,7 %). Род Megachile наиболее представлен и по суммарному численному обилию особей (45 % от числа отловленных мегахилид). Наибольшим численным обилием среди на исследуемой территории характеризуется M. ligniseca (подрод Megachile s. str.) – 18 % от общих сборов и 41 % от числа отловленных представителей рода. По классам обилия виды мегахилид на территории Кузнецко-Салаирской провинции распределяются следующим образом: к классу доминантных видов (с численным обилием более 6,06 % от общего объёма сборов мегахилид) отнесены 2 вида (Megachile ligniseca, Coelioxys inermis); субдоминантных (1,45–6,05 %) – 25 видов; редких (0,37–1,44 %) – 9 видов; очень редких (0,36 % и менее) – 13 видов. 143 При рассмотрении ареалогического состава фауны выявлено 4 долготные и 4 широтные ареалогические группы видов. Среди долготных групп преобладают виды с транспалеарктическим распространением (63,2 % от общего числа видов, 64,3 % от общих сборов), среди широтных – полизональные виды (75,5; 72,3). По числу видов и по видовому составу фауна мегахилид Кузнецко-Салаирской провинции проявляет наибольшее сходство с фаунами регионов европейской части России – Удмуртии и Московской области (индекс сходства 0,7 и 0,69 соответственно) и фауной Забайкалья (0,7). 144 ГЛАВА 5. ЭКОЛОГИЧЕСКАЯ ДИФФЕРЕНЦИАЦИЯ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE КУЗНЕЦКО-САЛАИРСКОЙ ПРОВИНЦИИ 5.1. Биотопические группы В результате проведённого исследования на территории Кузнецко-Салаирской провинции выявлено 4 биотопические группы мегахилид (рис. 5.1.1). а б Рисунок 5.1.1. Видовое (а) и численное (б) обилие пчёл семейства Megachilidae различных биотопических групп в Кузнецко-Салаирской провинции, % На территории Кузнецко-Салаирской провинции как по видовому (70 %), так и по суммарному численному (62 %) обилию особей преобладает группа луговых видов. К ней относится 34 вида пчёл сем. Megachilidae (табл. 5.1). Биотопы, предпочитаемые представителями этой группы, по типу растительности представляют собой различные типы лугов. Из них наиболее предпочитаемыми для мегахилид являются суходольные разнотравно-злаковые луга (на них встречается 43 вида мегахилид), остепнённые разнотравно-злаковые луга (19 видов) и лесные разнотравно-злаковые луга (16 видов). 144 Таблица 5.1 – Биотопические группы пчёл семейства Megachilidae Кузнецко-Салаирской провинции природные Вид 1 Луговые виды L. fuscipennis Ch. rapunculi H. acuticornis H. tridentata II III IV V VI VII VIII IX X XI XII XIII XIV XV XVI 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 + + + O. parietina O. spinulosa O. leaiana O. uncinata A. strigatum A. manicatum A. punctatum A. septemspinosum B. sibiricum S. ornatula + + + + S. punctulatissima S. simillima T. byssina C. alata C. conoidea C. elongata C. inermis + + ++ ++ ++ ++ ++ ++ ++ ++ + ++ ++ + ++ ++ ++ ++ ++ ++ ++ ++ ++ Числ. обилие, % 18 19 20 21 22 23 + + + + + 5 3,79 Ш + + 6 3 6 5,33 1,72 2,89 Ш Ш Ш + 3 2 1 1 4 4 2,17 2,62 0,09 0,09 1,63 0,91 Ш У У У Ш Ш 2 6 5 2 0,27 3,88 0,54 0,09 У Ш Ш У 3 3 0,36 1,63 У Ш + + + + + 5 5 1 2 + + 10 1,99 0,34 0,27 0,45 6,23 Ш Ш У У Ш антропогенные I XVII XVIII XIX + + + + + + + ++ + + + + ++ + + + + + + + ++ + + + + + + + + + + Широта биотопической приуроченности Всего Характерные биотопы в Кузнецко-Салаирской провинции + + + 145 Продолжение таблицы 5.1. 1 2 3 4 C. lanceolata C. rufescens + + M. alpicola M. bombycina M. centuncularis M. circumcincta M. genalis ++ M. lagopoda M. lapponica ++ + + M. maakii M. melanopyga M. versicolor M. willughbiella Лугово-степные виды H. mitis ++ O. bicolor O. pilicornis A. tridentatus C. afra C. emarginata M. analis M. argentata M. rotundata + + 5 ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ + + ++ + ++ ++ ++ ++ + ++ + + ++ ++ ++ + + 6 7 8 9 1 22 0,09 23 У 5 1,98 Ш 5 5 1,91 1,63 Ш Ш 1 0,91 У 1 6 0,72 2,44 У Ш 4 6 1,63 3,52 Ш Ш 3 1,89 Ш 4 0,72 Ш + + 3 3 2,44 4,88 Ш Ш + + + + + + 6 6 2,62 3,07 Ш Ш 5 2 2,53 0,27 Ш У 1 0,09 У 4 0,27 Ш 5 1 2,81 0,09 Ш У 3 0,99 Ш 10 11 12 13 14 15 16 17 18 19 20 21 + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + + 146 Окончание таблицы 5.1. 1 Лугово-лесные виды H. claviventris H. leucomelana 2 3 4 5 ++ ++ 6 7 8 9 22 23 5 2,26 Ш 3 1,99 Ш 2 5 0,36 1,44 У Ш 0,54 10 18,61 Ш Ш 10 11 12 13 14 15 16 17 18 19 20 21 ++ ++ + + + + H. tuberculata + ++ C. mandibularis + ++ ++ + + Эвритопные виды C. quadridentata + + + M. ligniseca + + ++ ++ + + + ++ + + 5 11 19 43 1 4 5 2 16 5 2 2 11 3 1 3 7 21 26 Всего: 3 Примечание. Природные биотопы: I – каменистая степь, II – разнотравно-луговая степь, III – остепнённый разнотравно-злаковый луг, IV – суходольный разнотравно-злаковый луг, V – высокотравный суходольный разнотравно-злаковый луг, VI – суходольный злаковоразнотравно-бобовый луг, VII – пойменный разнотравно-злаковый луг, VIII – пойменный разнотравно-осоковый луг, IX – лесной разнотравнозлаковый луг, X – высокотравный лесной разнотравно-злаковый луг, XI – берёзовый лес, XII – осиново-берёзовый лес, XIII – сосновоберёзовый лес, XIV – пихтово-берёзовый лес, XV – осиново-пихтовый лес, XVI – черневая тайга. Антропогенные биотопы: XVII – слабо изменённые, XVIII – измененные (нарушенные), XIX – преобразованные. «++» – биотопы, наиболее предпочитаемые определённым видом мегахилид. 147 Эти биотопы характеризуются благоприятным сочетанием природных условий (низкая влажность, высокая степень прогрева почвы и воздуха, большое разнообразие кормовых растений). Благоприятны условия для гнездования видов, устраивающих гнезда в полых стеблях растений и норках в сухой почве. Суходольные луга распространены в Кузнецко-Салаирской провинции повсеместно, на их долю приходится 75 % всей луговой площади провинции [Куминова А.В., 1950], что также объясняет высокое видовое и численное обилие мегахилид – представителей группы луговых видов. Анализ таксономического состава показал, что половину как видового, так и суммарного численного обилия луговых видов мегахилид составляют представители трибы Megachilini (рис. 5.1.2). К этой группе относятся 6 из 10 видов рода Coelioxys, отмеченных на исследуемой территории, и 11 из 15 видов рода Megachile. Кроме того, в состав группы входят все виды мегахилид трибы Anthidiini (гнездостроящие и паразитические), отмеченные на исследуемой территории. а б Рисунок 5.1.2. Представленность разных родов мегахилид в видовом (а) и суммарном численном обилии (б) биотопической группы луговых видов, % Примечание: одинаковой штриховкой отмечены роды в составе одной трибы. 148 Подавляющее большинство луговых видов на территории КузнецкоСалаирской провинции относятся к классу субдоминантов (19 видов) – A. strigatum, A. septemspinosum, Ch. rapunculi, C. rufescens, H. acuticornis, H. tridentata, L. fuscipennis, M. alpicola, M. bombycina, M. genalis, M. lagopoda, M. lapponica, M. maakii, M. versicolor, M. willughbiella, O. parietina, O. spinulosa, S. simillima, T. byssina. Они составляют 56 % видового и около 80 % суммарного численного обилия биотопической группы (рис. 5.1.3). 100% 90% 80% 70% 60% Очень редкие 50% Редкие 40% 30% Субдоминанты 20% Доминанты 10% 0% 1 2 Рисунок 5.1.3. Видовое (1) и численное (2) обилие мегахилид различных классов обилия в составе группы луговых видов, % К группе луговых видов также относится один доминантный вид – C. inermis, вклад которого в численное обилие луговых видов составляет 10,5 %. Из числа редких на территории Кузнецко-Салаирской провинции видов мегахилид к луговым относятся 6 видов (A. manicatum, B. sibiricum, C. elongata, M. centuncularis, M. circumcincta, M. melanopyga). Они составляют 17,6 % видового и 7 % численного обилия группы. 149 Очень редкие виды исследуемой территории в группе луговых видов представлены 8 видами мегахилид (A. punctatum, C. alata, C. conoidea, C. lanceolata, O. leaiana, O. uncinata, S. ornatula, S. punctulatissima). Вклад этих видов в видовое обилие группы составляет 23,5 %, в численное обилие – 2,7 %. Анализ относительного численного обилия видов мегахилид в пределах биотопической группы показал, что по численности среди луговых видов преобладают C. inermis, Ch. rapunculi и M. willughbiella (составляют 10, 8,6 и 7,9 % соответственно от суммарного численного обилия группы). Кроме того, высокое относительное численное обилие демонстрируют A. septemspinosum (6,26 %), L. fuscipennis (6,11 %), M. lapponica (5,7 %). Наиболее редкими видами в луговых биотопах являются O. leaiana, O. uncinata, S. ornatula, C. lanceolata (численность каждого не превышает 0,15 % от общего числа особей мегахилид группы луговых видов). Лугово-степная группа представлена меньшим числом видов мегахилид (9 видов, 18,4 % видового обилия); суммарное численное обилие её представителей составляет 12,7 % от числа всех отловленных особей (табл. 5.1). Представители этой группы предпочитают луговые и остепнённые местообитания – растительные сообщества с преобладанием в травостое многолетних ксерофитов, характеризующиеся недостаточным увлажнением и часто выраженным ритмом вегетации. Эти биотопы, благодаря редкому травостою, характеризуются благоприятными условиями для гнездования мегахилид, размещающих гнёзда в лёгкой, хорошо прогреваемой почве (H. mitis, M. analis, M. argentata), а в период вегетации (весна) привлекают пчёл с ранним началом сроков лёта своей богатой флорой. Однако эти местообитания, приуроченные к степному поясу, на исследуемой территории встречаются только в пределах Кузнецкой котловины и испытывают значительные преобразования в результате хозяйственной деятельности человека [Куминова А.В., 1950; Матысюк Н.Т., 1980]. 150 Таксономический состав группы лугово-степных видов показан на рис. 5.1.4. Наибольшее число видов в составе этой группы принадлежит трибе Megachilini, однако основной вклад в численное обилие группы вносят представители трибы Osmiini (наибольшим численным обилием характеризуются представители рода Osmia). а б Рисунок 5.1.4. Представленность разных родов мегахилид в видовом (а) и суммарном численном обилии (б) биотопической группы лугово-степных видов, % Примечание: одинаковой штриховкой отмечены роды в составе одной трибы. Особенностью таксономического состава данной группы на территории Кузнецко-Салаирской провинции является приуроченность к остепнённым участкам всех представителей подродов Eutricharaea (M. argentata и M. rotundata) и Allocoelioxys (C. afra и C. emarginata), отмеченных на территории области. Небольшое число видов в данной группе, вероятно, связано с постоянным сокращением площадей ненарушенных остепнённых территорий в результате антропогенного воздействия. 151 Из числа лугово-степных видов мегахилид Кузнецко-Салаирской провинции 4 вида относятся к субдоминантам (H. mitis, M. analis, O. bicolor, O. pilicornis) и 4 – к очень редким видам (A. tridentatus, C. afra, C. emarginata, M. argentata). Доля видов каждого класса обилия составляет 44,4 % видового обилия лугово-степной группы. Численное обилие видовсубдоминантов составляет 86,5 % от всех отловленных представителей лугово-степных видов, численное обилие очень редких видов – 5,6 % (рис. 5.1.5). 100% 90% 80% 70% 60% Очень редкие 50% 40% Редкие 30% 20% Субдоминанты 10% 0% 1 2 Рисунок 5.1.5. Видовое (1) и численное (2) обилие мегахилид различных классов обилия в составе группы лугово-степных видов, % Один вид (M. rotundata) относится к категории редких видов; его вклад в видовое и численное обилие этой биотопической группы составляет 11,2 и 7,9 % соответственно. Биотопическая группа лугово-лесных видов насчитывает 4 вида мегахилид (8,2 % от числа видов, отмеченных на территории КузнецкоСалаирской провинции) (табл. 5.1). Численное обилие представителей этой группы составляет 6,1 % от числа всех отловленных особей. Кроме луговых 152 местообитаний, представители лугово-лесных видов встречаются в лесных биотопах, отдавая предпочтение сосново-берёзовым и пихтово-берёзовым лесам. Леса в целом занимают более половины территории Кузнецко-Салаирской провинции, но распределены неравномерно; наибольшей лесистостью характеризуются горные территории южной и восточной частей провинции (орографические регионы Горная Шория и Кузнецкий Алатау). Несмотря на широкую представленность лесных биотопов на исследуемой территории и доступность разнообразных мест для гнездования и материалов для постройки ячеек (смола, хвоя, кусочки коры), ни один из видов мегахилид Кузнецко-Салаирской провинции не предпочитает леса другим типам биотопов. Повышенная влажность и небольшое разнообразие энтомофильной флоры по сравнению с луговыми и остепнёнными участками снижают привлекательность лесных типов биотопов для мегахилид, приуроченных в большинстве случаев к аридным территориям. Основу таксономического состава группы лугово-лесных видов на исследуемой территории составляют представители рода Hoplitis (3 вида); по численному обилию также преобладают пчёлы этого рода (76 % от отловленных представителей группы). По классам обилия группа представлена субдоминантными (H. leucomelana, H. claviventris; видовое и численное обилие составляет 50 и 70 % соответственно), редким (C. mandibularis; 25 и 23,9 %) и очень редким (H. tuberculata; 25 и 6,1 %) видами (рис.5.1.6). Биотопическая группа эвритопных видов мегахилид на территории Кузнецко-Салаирской провинции включает всего 2 вида (C. quadridentata, M. ligniseca). Указанные виды в равной мере встречаются в биотопах различных типов, не отдавая явного предпочтения ни одному из них. 153 100% 90% 80% 70% 60% Очень редкие 50% 40% Редкие 30% Субдоминанты 20% 10% 0% 1 2 Рисунок 5.1.6. Видовое (1) и численное (2) обилие мегахилид различных классов обилия в составе группы лугово-лесных видов, % Основной вклад в численное обилие этой экологической группы создает доминантный M. ligniseca (97,2 % от суммарного обилия представителей группы). Отсутствие у этого вида явной приуроченности к определенному типу биотопов связана главным образом со способностью гнездиться в различных готовых полостях, присущей многим видам мегахилид. Второй вид (C. quadridentata) относится к классу редких, его численное обилие составляет 2,8 %. Эвритопность представителей этого вида, вероятно, частично объясняется широким спектром видов-хозяев. В результате изучения биотопической приуроченности пчёл сем. Megachilidae на территории Кузнецко-Салаирской провинции разделены на 2 группы. Представители первой группы характеризуются широкой биотопической приуроченностью. Эта группа включает 34 вида (70 % видового обилия мегахилид исследуемой территории). 154 Представители видов этой группы встречаются в биотопах более чем одного типа, в том числе и в антропогенно преобразованных местообитаниях. Широкой эвритопные биотопической виды приуроченностью Кузнецко-Салаирской характеризуются провинции (M. все ligniseca и C. quadridentata), большинство лугово-степных видов (6 из 9 видов, 66 %), почти все лугово-лесные виды (3 из 4 видов, 75 %) и более половины луговых видов (67 %). Анализ классов численного обилия видов с широкой биотопической приуроченностью показывает, что эту группу составляют главным образом доминантные и субдоминантные виды мегахилид, доля редких и очень редких видов с широкой биотопической приуроченностью приходится в основном на клептопаразитов. Из табл. 5.1 видно, что наиболее приуроченность демонстрируют эвритопный широкую M. биотопическую ligniseca и луговой C. inermis, каждый из которых отмечен в биотопах 10 типов. Эти виды встречаются в природных и антропогенных ландшафтах. Вид C. inermis предпочитает биотопы лугового типа (остепнённые и суходольные разнотравно-злаковые луга, лесные луга), к которым тяготеет вслед за видами-хозяевами (M. alpicola, M. centuncularis, M. versicolor). Кроме того, этот вид встречается в осиново-берёзовых и сосново-берёзовых лесах. Местообитания антропогенного типа (агроценозы, территории сельской и городской застройки) C. inermis, вероятно, также осваивает вслед за видамихозяевами, находящими в указанных биотопах кормовую базу и большое разнообразие мест для гнездования. Для M. ligniseca, самого распространённого вида мегахилид на исследуемой территории, наиболее предпочитаемыми биотопами являются суходольные разнотравно-злаковые луга, разнотравно-злаковые лесные луга и таёжные местообитания, кроме того, он встречается на степных и остепнённых участках, берёзовых и осиново-берёзовых Антропогенные местообитания привлекают M. ligniseca наличием мест для гнездования (деревянные лесах. прежде всего постройки), а также 155 разнообразием кормовых растений, в том числе рудеральных. Группа видов с узкой биотопической приуроченностью насчитывает 15 видов мегахилид. В основном это луговые виды (73 % от видового обилия группы); кроме того, 3 лугово-степных вида (A. tridentatus, C. afra, M. argentata) и 1 лугово-лесной вид (H. tuberculata). Представители группы, как правило, встречаются в биотопах только одного, редко двух типов; на антропогенных территориях не встречаются. Большинство видов с узкой биотопической приуроченностью относится к классам очень редких (73 % видового обилия) и редких видов Только 1 вид (O. spinulosa) в составе данной группы (20 %). является субдоминантом. Наиболее узкая биотопическая приуроченность характерна для 8 видов мегахилид – O. leaiana, O. uncinata, C. conoidea, C. afra, C. lanceolata, M. argentata, M. centuncularis, M. circumcincta. Эти виды отмечены в биотопах только 1 типа. Возможно, это объясняется малой долей особей указанных видов в сборах. Из всех природных биотопов Кузнецко-Салаирской провинции наиболее привлекательными для пчёл сем. Megachilidae являются суходольные разнотравно-злаковые луга. На местообитаниях этого типа отмечены 43 вида мегахилид (87 % видового обилия исследуемой территории). Кроме того, мегахилид привлекают также лесные и остепнённые луга. В этих биотопах отловлены 16 и 19 видов мегахилид (32 и 38 % соответственно). Следует отметить, что в целом антропогенные ландшафты в КузнецкоСалаирской провинции привлекают больше видов пчёл сем. Megachilidae, чем большинство природных биотопов. На территориях, относящихся к категориям изменённых (нарушенных) и преобразованных человеком отмечено 21 и 26 видов мегахилид соответственно (43 % и 53 % видового обилия). Эти местообитания по числу отмеченных видов уступают лишь суходольным разнотравно-злаковым лугам. 156 Наименее привлекательными для мегахилид местообитаниями на исследуемой территории являются высокотравные суходольные разнотравнозлаковые луга, пойменные разнотравно-осоковые луга, а также большая часть лесных биотопов (берёзовые, осиново-берёзовые и осиново-пихтовые леса). В биотопах этого типа были отмечены по 1–2 вида мегахилид. 5.2. Трофические группы На территории Кузнецко-Салаирской провинции выделены 4 трофические группы мегахилид (рис. 5.2.1). а б Рисунок 5.2.1. Видовое (а) и численное (б) обилие пчёл сем. Megachilidae разных трофических групп в Кузнецко-Салаирской провинции, % К широким полилектам на исследуемой территории относится 18 видов мегахилид (табл. 5.2). Широкие полилекты собирают пыльцу с широкого круга растений. В Кузнецко-Салаирской провинции представители этой группы преобладают как по видовому, так и по суммарному численному обилию (53 и 61 % соответственно). Более половины широких полилектов относятся к роду Megachile (11 видов); кроме того, к ним принадлежит большая часть видов р. Osmia (4 из 6). Из антидиин в трофическую группу входит только 1 вид – A. manicatum, остальные представители трибы имеют более узкие трофические предпочтения. 157 1 I II III IV V VI VII VIII IX X XI XII XIII XIV XV XVI XVII XVIII XIX XX XXI XXII XXIII XXIV 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 Широкие полилекты A. manicatum H. claviventris H. leucomelana O. bicolor O. parietina O. pilicornis O. uncinata M. alpicola M. analis M. argentata M. centuncularis M. genalis + M. lapponica M. ligniseca M. melanopyga * M. rotundata M. versicolor M. willughbiella Узкие полилекты A. septemspinosum *+ + + * * + * *+ * + *+ + + *+ * *+ *+ * * * * *+ *+ + + + + * * * * + * + * * * + *+ *+ * * + * * + *+ + * *+ * * + + * * + * + * *+ * *+ *+ *+ * + *+ + + * + + * * * * + * * + + *+ * + + * + + * + * + * + * *+ + + + + * * Собственные данные Семейства растений Вид Литературные данные Таблица 5.2 – Трофические группы пчёл семейства Megachilidae Кузнецко-Салаирской провинции 26 27 3 1 3 3 2 1 5 4 1 8 8 1 2 4 1 5 5 5 2 4 4 5 3 3 4 3 1 1 3 5 7 2 3 6 3 2 158 Окончание таблицы 5.2. 1 2 Широкие олиголекты L. fuscipennis + A. strigatum A. punctatum H. acuticornis H. tridentata H. tuberculata O. spinulosa O. leaiana M. bombycina M. circumcincta M. maakii M. lagopoda Узкие олиголекты T. byssina Ch. rapunculi H. mitis По литературным данным, всего: Собственные данные, всего: 3 4 5 6 7 *+ * * 8 + + *+ * *+ + 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 1 3 4 1 1 1 3 1 3 1 1 1 1 5 1 1 2 1 3 3 2 2 1 1 1 1 1 4 5 *+ + *+ * *+ *+ *+ * * *+ *+ * + * + * + * + + + *+ + *+ + *+ + *+ + + + + 1 20 1 2 2 1 2 1 2 23 1 13 1 5 1 - 1 3 1 3 - - 5 - 2 15 2 - 3 1 9 - 1 20 2 9 1 1 3 1 1 - 2 10 1 1 4 1 Примечание. Семейства растений: I – Apiaceae, II – Asteraceae, III – Boraginaceae, IV – Brassicaceae, V – Campanulaceae, VI – Convolvulaceae, VII – Crassulaceae, VIII – Cucurbitaceae, IX – Dipsacaceae, X – Fabaceae, XI – Geraniaceae, XII – Lamiaceae, XIII – Liliaceae, XIV – Lythraceae, XV – Onagraceae, XVI – Papaveraceae, XVII – Primulaceae, XVIII – Malvaceae, XIX – Ranunculaceae, XX – Rosaceae, XXI – Rubiaceae, XXII – Salicaceae, XXIII – Scrophulariaceae, XXIV – Viburnaceae. «*» по литературным данным (Adolph, 1934; Песенко, 1971; Anasiewicz, 1973; Gogala, 1991, 1994, 1999; Радченко, Песенко, 1994; Banaszak, Romasenko, 1998; Ban, 2005; Амолин, 2008; Прощалыкин, 2009 а; Игнатенко, 2012). « +» – семейства растений, посещаемые мегахилидами в Кузнецко-Салаирской провинции 159 По классам обилия внутри трофической группы мегахилиды распределяются следующим образом: к широким полилектам относится 1 доминант (M. ligniseca), 11 субдоминантов (H. claviventris, H. leucomelana, O. bicolor, O. parietina, O. pilicornis, M. alpicola, M. analis, M. genalis, M. lapponica, M. versicolor, M. willughbiella), 4 редких (A. manicatum, M. centuncularis, M. melanopyga, M. rotundata) и 2 очень редких для Кузнецко-Салаирской провинции вида (O. uncinata, M. argentata) (рис. 5.2.2). 100% 90% 80% 70% Очень редкие Редкие Субдоминанты Доминанты 60% 50% 40% 30% 20% 10% 0% 1 2 Рисунок 5.2.2. Видовое (1) и численное (2) обилие мегахилид различных классов обилия в составе трофической группы широких полилектов, % Большинство широких полилектов относится к луговым видам (10 видов, 55,5 % видового обилия) (рис. 5.2.3). Кроме того, в эту группу входят 5 лугово-степных и 2 лугово-лесных вида (составляют 28 и 11 % видового обилия соответственно), и единственный эвритопный вид гнездостроящих мегахилид (Megachile ligniseca) Кузнецко-Салаирской провинции (5,5 %). По численному обилию среди широких полилектов преобладает эвритопный M. ligniseca (35,6 % суммарного численного обилия трофической группы). Наименьшее численное обилие демонстрируют лугово-лесные виды (8 %). 160 а б Рисунок 5.2.3. Соотношение видового (а) и численного (б) обилия мегахилид разных биотопических групп в составе трофической группы широких полилектов, % Большинство видов – широких полилектов относятся к группе видов мегахилид с широкой биотопической приуроченностью (16 видов, 89 % видового обилия). Наиболее широкая трофическая приуроченность (по числу посещаемых семейств растений) на исследуемой территории свойственна M. ligniseca (посещает растения из 7 семейств, предпочитает Asteraceae) и M. willughbiella (6 семейств, предпочитает Fabaceae) (табл. 5.2.). Для таких полилектичных видов, как M. argentata, M. melanopyga и O. uncinata, трофические связи установлены только на основе литературных данных, и на территории Кузнецко-Салаирской провинции остаются неясными. Трофическая группа узких полилектов включает 1 вид мегахилид (A. septemspinosum), составляющий 3 % видового обилия гнездостроящих мегахилид и 4,5 % численного обилия. Самки пчёл этого вида собирают пыльцу с немногих видов растений разных семейств. На исследуемой территории вид предпочитает семейства Fabaceae (Vicia cracca) и Lamiaceae (Phlomis tuberosa). В Кузнецко-Салаирской провинции A. septemspinosum относится к видам- 161 субдоминантам, входит в состав биотопической группы луговых видов и характеризуется широкой биотопической приуроченностью. К широким олиголектам на территории Кузнецко-Салаирской провинции относятся 12 видов мегахилид, предпочитающих для сбора пыльцы посещать растения преимущественно одного семейства. Видовое и суммарное численное обилие широких олиголектов составляет 35 и 22,4 % соответственно. В районе исследований представители этой трофической группы предпочитают посещать растения сем. Fabaceae (A. strigatum, A. punctatum, H. acuticornis, H. tridentata, M. maakii), Asteraceae (L. fuscipennis, O. spinulosa, M. bombycina, M. lagopoda), Lamiaceae (M. circumcincta). Для O. leaiana и H. tuberculata типы трофических связей приведены по литературным данным, касающихся других территорий; в Кузнецко-Салаирской провинции остаются неизвестными. Большая часть видов – широких олиголектов (8 видов) на исследуемой территории относится к категории субдоминантов. Один вид является редким (M. circumcincta), и 3 вида (A. punctatum, H. tuberculata, O. leaiana) – очень редкими. По численности в составе группы преобладают L. fuscipennis, составляющий 20 % численного обилия группы, и H. tridentata (15 %). Минимальный вклад в численное обилие группы вносит Большинство широких олиголектов являются O. leaiana (0,5 %). луговыми видами (11 видов, 91 %), и один только 1 вид относится к лугово-лесным (H. tuberculata). Особенностью данной трофической группы является наибольшее число видов с узкой биотопической приуроченностью (42 % видового и 21 % суммарного численного обилия группы). 162 Группа узких олиголектов на исследуемой территории включает 3 вида пчёл сем. Megachilidae (8,8 % видового обилия, 11,5 % численного обилия отловленных особей гнездостроящих видов). Представители этой группы для сбора пыльцы посещают, как правило, растения исключительно одного семейства или рода. В Кузнецко-Салаирской провинции узкие олиголекты отдают предпочтение растениям из сем. Fabaceae (T. byssina) и рода Campanula (Ch. rapunculi, H. mitis). Данные о трофических связях этих видов в Кузнецко-Салаирской провинции совпадают с литературными [Радченко В.Г., Песенко Ю.А., 1998; Прощалыкин М.Ю., 2009 а]. Все три вида в составе трофической группы на исследуемой территории являются субдоминантами; по численности среди узких олиголектов преобладает Ch. rapunculi (54 % суммарного численного обилия). Минимальным численным обилием в сборах характеризуется T. byssina (20 %). Оба этих вида относятся к луговым видам, а H. mitis – к луговостепным. Все три вида характеризуются широкой биотопической приуроченностью. Из табл. 5.2 и рис. 5.2.4 видно, что наибольшее число видов пчёл сем. Megachilidae в Кузнецко-Салаирской провинции предпочитают посещать растения из трёх семейств: Fabaceae (на цветках растений данного семейства отмечено 20 видов гнездостроящих мегахилид, 57 % видового обилия), Asteraceae (15 видов, 43 %) и Rosaceae (10 видов, 28 %). Предпочтение особенностями растений строения из первых двух лабио-максиллярного семейств объясняется комплекса мегахилид, относящихся к условной группе «длиннохоботковых» пчёл [Прощалыкин М.Ю., 2009 а]. Данная особенность позволяет мегахилидам добывать нектар из цветков с глубоким трубчатым венчиком, недоступный для большинства пчёл и других насекомых. Растения сем. Rosaceae не имеют ярко выраженных приспособлений к каким-либо агентам опыления, и, вероятно, привлекают мегахилид большим количеством пыльцы и доступного нектара [Тахтаджян А.Л., 1980]. 163 Рисунок 5.2.4. Привлекательность различных семейств растений для пчёл сем. Megachilidae (по числу отмеченных на них видов мегахилид) в Кузнецко-Салаирской провинции, % от видового обилия Полный спектр кормовых растений мегахилид в Кузнецко-Салаирской провинции представлен в табл. 5.3. На исследуемой территории пчёлы сем. Megachilidae посещают растения из 21 семейства, 58 родов и 80 видов. Из сем. Asteraceae на исследуемой территории мегахилиды посещают 19 видов растений из 18 родов, из сем. Fabaceae – 18 видов 8 родов, Rosaceae – 9 видов 5 родов. Наибольшее число посещаемых растений относится к группам ценных кормовых (23 вида, преимущественно Fabaceae) и лекарственных растений (22 вида, главным образом Asteraceae и Lamiaceae). 164 Scrophulariaceae Viburnaceae Всего: 1 1 2 3 1 1 16 1 1 Рудеральные 2 1 4 Синантропные Декоративные 8 Пищевые 1 2 Прочие 1 Лекарственные 2 19 2 4 1 1 1 18 1 6 1 1 1 1 1 4 9 1 1 4 1 80 Кормовое 2 18 2 1 1 1 1 8 1 5 1 1 1 1 1 3 5 1 1 2 1 58 Число видов растений по группам Медоносы Apiaceae Asteraceae Boraginaceae Campanulaceae Convolvulaceae Crassulaceae Dipsacaceae Fabaceae Geraniaceae Lamiaceae Liliaceae Lythraceae Onagraceae Papaveraceae Primulaceae Ranunculaceae Rosaceae Rubiaceae Salicaceae Всего видов растений Семейства растений Всего родов растений Таблица 5.3 – Кормовой спектр пчёл сем. Megachilidae в КузнецкоСалаирской провинции 1 2 5 1 1 1 1 2 1 1 5 23 3 1 1 22 1 2 1 3 1 1 2 15 1 5 2 1 6 Анализ данных показывает, что наибольшим числом видов растений, посещаемых мегахилидами, характеризуются семейства Fabaceae, Asteraceae, Rosaceae (рис. 5.2.5). Наименьшее число посещаемых мегахилидами видов растений (по 1 виду) на исследуемой территории включают семейства Convolvulaceae, Crassulaceae, Dipsacaceae, Liliaceae, Lythraceae, Papaveraceae, Primulaceae, Rubiaceae, Salicaeae. 165 Рис. 5.2.5. Привлекательность различных семейств растений (по числу посещаемых видов) для пчёл сем. Megachilidae в Кузнецко-Салаирской провинции На цветках растений вышеперечисленных семейств отмечено по 1 виду мегахилид, за исключением более привлекательного Crassulaceae (посещают 9 видов мегахилид). Малая привлекательность остальных, вероятно, обусловлена «общедоступностью» нектара и пыльцы для насекомых самых разнообразных групп (Convolvulaceae, Dipsacaceae, Rubiaceae и др.); ранним периодом цветения (Salicaceae – отмечено единственное посещение O. bicolor – видом с ранним началом лёта); малым числом видов растений в семействе (Dipsacaceae, Lythraceae, Papaveraceae – на исследуемой территории насчитывают всего по 2 вида). Частота посещаемости мегахилидами растений наиболее привлекательных семейств отражена на рис. 5.2.6. По числу зарегистрированных посещений цветков установлено, что наибольшей привлекательностью для пчёл сем. Megachilidae характеризуются растения из следующих семейств: Fabaceae – на растения, принадлежащие к этому семейству приходится 166 около трети (32 %) всех зарегистрированных посещений мегахилидами кормовых растений в Кузнецко-Салаирской провинции. Семейство Fabaceae на исследуемой территории насчитывает 14 родов и более 70 видов растений. Характеризуется приобретённым в ходе коэволюции с пчелиными особым строением цветка, предупреждающим проникновение в него неэффективных опылителей [Тахтаджян А.Л., 1980]. Рисунок 5.2.6. Частота посещаемости мегахилидами растений наиболее привлекательных семейств в Кузнецко-Салаирской провинции, % Самым посещаемым растением этого семейства в Кузнецко-Салаирской провинции является Горошек мышиный (Vicia cracca), являющийся обычным во всех регионах провинции. На него приходится 30 % всех посещений мегахилидами цветков бобовых растений и 9,5 % от всех зарегистрированных посещений кормовых растений. Наиболее часто на цветках бобовых отмечали представителей видов A. septemspinosim (17 % случаев) и O. pilicornis (12 %). Asteraceae – 28,7 % посещений. В Кузнецко-Салаирской провинции семейство насчитывает 58 родов и около 180 видов (самое крупное семейство 167 растений по числу видов в Кузнецко-Салаирской провинции). Наиболее предпочитаемыми растениями из этого семейства для мегахилид являются Бодяк разнолистный (Cirsium heterophyllum) – 12 % от всех посещений астровых и 3,5 % от общего числа отмеченных посещений кормовых растений; Девясил иволистный (Inula salicina) (11 и 3,2 %); Василёк шероховатый (Centaurea scabiosa) (9,5 и 2,7 %). Эти виды широко распространены в большинстве биотопов по всему исследуемому региону. Чаще других видов мегахилид растения этого семейства посещает M. ligniseca (35 % случаев) и O. spinulosa (11 %). Campanulaceae – 12,4 % посещений. На исследуемой территории растения данного семейства привлекают преимущественно мегахилид – узких олиголектов, отдающих предпочтение растениям из р. Campanula (отмечено посещение 4 видов из 10). Самый посещаемый вид растений в семействе – Колокольчик сибирский (Campanula sibirica), распространённый в большинстве типичных биотопов Кузнецко-Салаирской провинции. На этот вид приходится 66 % от всех посещений мегахилидами растений из сем. Campanulaceae и 8 % от общего числа зарегистрированных посещений мегахилидами растений на исследуемой территории. В 80 % случаев посещения цветков растений данного рода на них был отмечен Ch. rapunculi. Для Rosaceae и Lamiaceae отмечено 6,4 и 6,7 % посещений мегахилидами соответственно. Явного предпочтения растений указанных семейств не выявлено ни у одного вида мегахилид. На территории Кузнецко-Салаирской провинции сем. Rosaceae насчитывает 78 видов 21 рода; сем. Lamiaceae – 45 видов 20 родов. Наиболее привлекательными для пчёл растениями являются широко распространённые Зопник клубненосный (Phlomis tuberosa) (Lamiaceae) – 37 % от всех посещений растений семейства и 2,5 % от общего числа посещений; и Лапчатка серебристая (Potentilla argentea) (Rosaceae) – 27 и 1,7 %. 168 На сем. Crassulaceae приходится 4,7 % от всех зарегистрированных посещений растений пчёлами-мегахилидами. На исследуемой территории семейство представлено 3 родами и 7 видами растений, приуроченными в большистве случаев к каменистым местообитаниям, скалам, осыпям, а также альпийским лугам и высокогорным тундрам. Посещение мегахилидами отмечены для 1 вида – Очитка гибридного (Sedum hybridum), широко распространённого в Кузнецко-Салаирской провинции. Доля посещений мегахилидами каждого из остальных семейств растений на исследуемой территории не превышает 1,5 %. Наиболее широким спектром видов посещаемых растений характеризуются представители рода Megachile – M. ligniseca (отмечен на 18 видах растений 17 родов), M. willughbiella (11 видов 10 родов) и M. lapponica (10 видов 9 родов). Таким образом, на территории Кузнецко-Салаирской провинции виявлено 4 биотопические группы мегахилид. Из них по видовому (69,3 %) и по суммарному численному (62 %) обилию особей преобладает группа луговых видов, что связано как с особенностями экологии пчёл, так и с ландшафтной дифференциацией исследуемой территории. В составе этой биотопической группы преобладают представители трибы Megachilini (к ней относится 17 из 34 луговых видов). Максимальным численным обилием среди луговых видов характеризуются C. inermis, Ch. rapunculi и M. willughbiella (10, 8,6 и 7,9 % соответственно от суммарного численного обилия группы). Наиболее редки O. leaiana, O. uncinata, S. ornatula, C. lanceolata. Лугово-степные виды значительно уступают луговым по видовому и численному обилию (18,4 и 12,7 % соответсвенно). Большинство видов в составе этой группы принадлежит трибе Megachilini (5 из 9 видов); основной вклад в численное обилие группы вносят представители трибы Osmiini (65 %). По численности в составе группы преобладают субдоминантные O. bicolor, O. pilicornis, H. mitis, M. analis). Наиболее редки A. tridentatus, C. afra, C. emarginata, M. argentata. 169 Для видов лугово-лесной группы характерно небольшое видовое и наименьшее численное обилие (4 вида, 8,2 % видового и 6,1 % численного обилия). В составе группы по численности преобладают субдоминанты H. leucomelana, H. claviventris и редкий C. mandibularis, а наименьшее численное обилие демонстрирует очень редкий H. tuberculata. Группа эвритопных видов характеризуется минимальным видовым обилием и включает 2 вида мегахилид – доминантный M. ligniseca и редкий C. quadridentata. По численному обилию представители этой группы уступают только луговым видам за счёт M. ligniseca, обитающего повсеместно и являющегося самым массовым видом на исследуемой территории. По числу посещаемых биотопов пчёлы-мегахилиды Кузнецко-Салаирской провинции разделены на 2 группы. Группа мегахилид с широкой биотопической приуроченностью включает 34 вида пчёл (70 % видового обилия мегахилид исследуемой территории), встречающихся в биотопах более чем 1 типа, в том числе в антропогенных. Эту группу составляют главным образом доминантные и субдоминантные виды мегахилид, доля редких и очень редких видов приходится в основном на клептопаразитов. Группа видов с узкой биотопической приуроченностью насчитывает 15 видов мегахилид (30,5 % видового обилия). Её представители, как правило, встречаются в биотопах только одного (редко двух) типов и на антропогенных территориях не встречаются. Большинство видов этой группы относится к классам редких и очень редких видов. Наиболее широкую биотопическую приуроченность демонстрируют эвритопный M. ligniseca и луговой C. inermis (отмечены в биотопах 10 типов), самая узкая биотопическая приуроченность характерна для 8 видов мегахилид – O. leaiana, O. uncinata, C. conoidea, C. afra, C. lanceolata, M. argentata, M. centuncularis, M. circumcincta (отмечены в биотопах 1 типа). Наиболее привлекательными из всех биотопов для пчёл сем. Megachilidae являются суходольные разнотравно-злаковые луга (на них отмечено 87 % 170 видового обилия мегахилид исследуемой территории). Установлено, что на исследуемой территории пчёлы сем. Megachilidae посещают растения из 21 семейства, 58 родов и 80 видов. Из них наиболее привлекательными для большинства видов являются растения из семейств Fabaceae, Asteraceae и Rosaceae. По широте спектра кормовых растений на исследуемой территории выделено 4 трофические группы мегахилид – широкие полилекты (18 видов, 53 % видового и 61 % численного обилия гнездостроящих видов), узкие полилекты (1 вид, 3 и 4,4 %), широкие олиголекты (12 видов, 35 и 22,4 %), узкие олиголекты (3 вида, 8,8 и 11,5 5). Основу каждой из выделенных трофических групп пчёл-мегахилид составляют луговые виды. Наиболее широкой трофической приуроченностью (по числу посещаемых семейств растений) на исследуемой территории характеризуются M. ligniseca (посещает растения из 7 семейств, предпочитает Asteraceae) и M. willughbiella (посещает растения из 6 семейств, предпочитает Fabaceae). M. ligniseca и M. willughbiella характеризуются также самым широким спектром видов посещаемых растений (отмечены на 18 видах растений 17 родов и 11 видов 10 родов соответственно). Для 4 видов гнездостроящих мегахилид (B. sibiricum, O. leaiana, O. uncinata, M. melanopyga) трофические связи на территории КузнецкоСалаирской провинции не установлены. 171 ГЛАВА 6. ОСОБЕННОСТИ ЛАНДШАФТНОБИОТОПИЧЕСКОЙ ДИФФЕРЕНЦИАЦИИ ПЧЁЛ СЕМЕЙСТВА MEGACHILIDAE КУЗНЕЦКО-САЛАИРСКОЙ ПРОВИНЦИИ 6.1. Особенности распределения пчел семейства Megachilidae по орографическим регионам Кузнецко-Салаирской провинции Орографические регионы Кузнецко-Салаирской провинции значительно различаются разнообразием по особенностям представленных рельефа, климатическим условиями, типов ландшафтов и растительности, вследствие чего изучение распределения фауны пчёл сем. Megachilidae по орографическим регионам представляет большой интерес. Сравнение фауны пчёл сем. Megachilidae равнинных (Кузнецкая котловина) и горных (Кузнецкий Алатау, Горная Шория) территорий Кузнецко-Салаирской провинции показало значительную неравномерность распределения населения мегахилид по орографическим регионам КузнецкоСалаирской провинции (рис. 6.1.1, табл. 6.1). Рисунок 6.1.1. Видовое и численное обилие пчёл сем. Megachilidae орографических регионов Кузнецко-Салаирской провинции, % 172 Таблица 6.1 – Видовой состав и численное обилие (%) пчёл семейства Megachilidae в орографических регионах Кузнецко-Салаирской провинции Численное обилие, % Вид 1 Кузнецкая котловина Кузнецкий Алатау Горная Шория КузнецкоСалаирская провинция Класс обилия 2 3 4 5 6 0,83 3,79 сд Lithurgini 1. Lithurgus fuscipennis 3,90 – Osmiini 2. Chelostoma rapunculi 6,00 4,51 4,50 5,33 сд 3. Hoplitis acuticornis 2,70 0,58 – 1,72 сд 4. H. claviventris 2,24 3,93 0,84 2,26 сд 5. H. leucomelana 0,90 6,18 2,07 1,99 сд 6. H. mitis 4,35 – – 2,62 сд 7. H. tridentata 4,05 1,12 1,25 2,89 сд 8. H. tuberculata – 1,68 0,42 0,36 ор 9. Osmia bicolor 4,65 1,12 0,42 3,07 сд – – 0,42 0,09 ор 11. O. parietina 0,29 7,30 3,73 2,17 сд 12. O. pilicornis 3,75 – 1,25 2,53 сд 13. O. spinulosa 0,14 – 11,58 2,62 сд 14. O. uncinata 0,14 – – 0,09 ор 10. O. leaiana Anthidiini 15. Anthidiellum strigatum 0,59 – 5,79 1,63 Сд 16. Anthidium manicatum 1,19 – 0,83 0,91 Р 17. A. punctatum 0,29 – 0,41 0,27 ор 18. A. septemspinosum 6,00 1,12 – 3,88 сд 19. Bathanthidium sibiricum 0,89 – – 0,54 р 20. Stelis punctulatissima 0,59 – – 0,36 ор 21. S. simillima 2,70 – – 1,63 сд 22. S. ornatula 0,14 – – 0,09 ор 23. Trachusa byssina 1,19 3,37 3,31 1,99 сд – 0,27 ор Dioxyini 24. Aglaoapis tridentatus 0,43 – 173 Окончание таблицы 6.1. 1 2 3 4 5 6 Megachilini 25. Coelioxys afra 0,14 – – 0,09 ор 26. C. alata 0,29 1,12 – 0,35 ор 27. C. conoidea 0,44 – – 0,27 ор 28. C. elongata 0,59 0,56 – 0,45 р 29. C. emarginata 0,29 – 0,41 0,27 ор 30. C. inermis 6,00 6,18 6,61 6,23 д – 0,56 – 0,09 ор 32. C. mandibularis 0,29 2,25 3,72 1,44 р 33. C. quadridentata 0,29 2,25 – 0,54 р 34. C. rufescens 1,94 2,25 1,24 1,98 сд 35. Megachile alpicola 1,64 5,05 0,41 1,91 сд 36. M. analis 4,35 – 0,82 2,81 сд 37. M. argentata 0,14 – – 0,09 ор 38. M. bombycina 1,79 2,25 0,82 1,63 сд 39. M. centuncularis 0,59 – 2,48 0,91 р 40. M. circumcincta 1,19 – – 0,72 р 41. M. genalis 2,71 3,93 – 2,44 сд 42. M. lagopoda 2,25 – 0,82 1,63 сд 43. M. lapponica 2,10 7,86 4,54 3,52 сд 44. M. ligniseca 17,74 25,28 17,35 18,61 д 45. M. maakii 0,45 1,12 5,78 1,89 сд 46. M. melanopyga 0,60 – 1,65 0,72 р 47. M. rotundata 1,50 0,56 – 0,99 р 48. M. versicolor 3,00 2,25 4,96 2,44 сд 49. M. willughbiella 2,56 5,62 10,74 4,88 сд 31. C. lanceolata Всего видов 46 26 30 49 Примечание: д – доминантный, сд – субдоминантный, р – редкий, ор – очень редкий. На территории Кузнецкой котловины обнаружено 46 видов пчёл сем. Megachilidae (94 % от общего числа видов в Кузнецко-Салаирской провинции); суммарное численное обилие отловленных здесь пчёл составляет 60 % от общих сборов. Горных регионы значительно уступают 174 котловине по видовому и численному обилию мегахилид. В Горной Шории отмечено 30 видов (59 % видового обилия), в Кузнецком Алатау – 26 видов (53 %). Численное обилие пчёл, отловленных в горных регионах, составляет 21,8 % для Горной Шории и 16 % для Кузнецкого Алатау. Кузнецкая котловина. Наибольшее число видов мегахилид обнаружено на территории Кузнецкой котловины (46 видов). Из них 11 видов (22 % от общего числа видов) на исследуемой территории отмечены только в пределах этого орографического региона (табл. 6.1). Численное обилие мегахилид здесь в 2,8–3 раза больше, чем в горных орографических регионах (Горной Шории и Кузнецком Алатау). связано с наличием и преобладанием предпочитаемых мегахилидами котловины типичны Вероятно, такое распределение биотопов. остепнённые и в Кузнецкой Для степные котловине пониженных участков местообитания, для повышенных – лесостепь, где берёзовые и берёзово-осиновые колки чередуются с массивами суходольных лугов и луговых степей [Гвоздецкий Н.А., Михайлов Н.И., 1978]. В состав видов, не обнаруженных в других регионах провинции, входят представители лугово-степной (4 вида) и луговой (7 видов) групп; большинство из них (8 видов) характеризуется узкой биотопической приуроченностью. Кузнецкая котловина выделяется по числу надвидовых таксонов мегахилид. Здесь встречаются представители всех триб и родов, отмеченных в Кузнецко-Салаирской провинции (12 родов, 5 триб). Только в этом орографическом регионе обнаружены представители родов Aglaoapis, Bathanthidium (включают по 1 виду) и Stelis (3 вида), относящиеся к луговой и лугово-степной биотопическим группам. Остальные трибы в этом регионе представлены наиболее полно по сравнению с горными регионами. Только в Кузнецкой котловине встречаются все виды антидиин, обитающих в Кузнецко-Салаирской провинции. 175 Основу населения мегахилид Кузнецкой котловины составляют представители трибы Megachilini (рис 6.1.2). К родам этой трибы (Coelioxys и Megachile) в этом регионе относится около половины видов мегахилид (52 %). Численное обилие представителей трибы Megachilini в Кузнецкой котловине составляет 53 % от числа отловленных здесь мегахилид. Рисунок 6.1.2. Соотношение триб сем. Megachilidae по видовому (а) и численному (б) обилию в разных орографических регионах КузнецкоСалаирской провинции, % Наибольшее численное обилие на территории Кузнецкой котловины демонстрируют M. ligniseca (17 % численного обилия) и Ch. rapunculi и C. inermis (по 6 %), наименьшее – O. spinulosa, O. uncinata, C. afra, M. argentata (менее 0,15 %). В целом по видовому составу и численному обилию мегахилид в Кузнецкой котловине преобладают луговые (33 вида, 72 % видового и 59 % численного обилия) и лугово-степные (9 видов, 19,5 и 27 %) виды. 176 В Кузнецкой котловине не обнаружены 3 вида мегахилид – лугово-лесной H. tuberculata и малочисленные в сборах O. leaiana и C. lanceolata. Горная Шория. При переходе от центральной равнинной части провинции к периферии с увеличением высоты рельефа изменяется тип растительности, и режим увлажнения изменяется в сторону повышения влажности. Вследствие этого, видовой состав мегахилид горных территорий значительно беднее. В Горной Шории отмечено 30 видов пчёл сем. Megachilidae (61 % от общего числа видов в Кузнецко-Салаирской провинции). Численное обилие мегахилид составляет 21,8 % от общих сборов. Только на территории Горной Шории обнаружен очень редкий O. leaiana. Горная Шория уступает Кузнецкой котловине по числу отмеченных триб и родов мегахилид (4 трибы, 9 родов). По видовому и численному обилию в Горной Шории представители трибы Megachilini (рис 6.1.2). преобладают К ней относится половина отмеченных здесь видов мегахилид, их суммарное численное обилие составляет 62,8 %. По численному обилию в Горной Шории преобладают M. ligniseca (17,3 % от общего числа особей), O. spinulosa (11,6), M. willughbiella (10,7), C. inermis (6,6). Наиболее редки A. punctatum, H. tuberculata, O. bicolor, O. leaiana, M. alpicola (по 0,4 %). Численность таких видов, как A. strigatum O. spinulosa, M. centuncularis, M. maakii, M. versicolor и M. willughbiella в Горной Шории резко увеличивается по сравнению с численностью в других орографических регионах и провинции в целом, что вероятно, связано как с уменьшением числа особей других видов, так и с наличием благоприятных биотопов. В видовом составе мегахилид Горной Шории преобладают луговые виды (21 вид, 70 % от числа видов, отловленных в этом орографической регионе). Возрастает роль лугово-лесных видов по сравнению с Кузнецкой котловиной (4 вида, 13,3 %). По численному обилию выделяются группы луговых (73 %) и эвритопных (17 %) видов. 177 В число видов мегахилид, отсутствующих в Горной Шории, входят луговые (A. septemspinosum, B. sibiricum, C. alata, C. conoidea, C. elongata, C. lanceolata, C. quadridentata, H. acuticornis, M. circumcincta, M. genalis, O. uncinata, S. punctulatissima, S. simillima, S. ornatula) и лугово-степные (A. tridentatus, C. afra, H. mitis, M. argentata, M. rotundata) виды. Кузнецкий Алатау. По видовому обилию пчёл сем. Megachilidae Кузнецкий Алатау значительно уступает другим орографическим регионам. Здесь отмечено 26 видов мегахилид (53 % от общего числа видов). Численное обилие пчёл этого семейства также невелико и составляет 16 % от общих сборов. По числу отмеченных триб и родов мегахилид Кузнецкий Алатау также уступает Кузнецкой котловине и Горной Шории. Здесь обитают представители 3 триб (отсутствуют Lithurginini и Dioxyini) и 7 родов (кроме представителей указанных триб, отсутствуют Anthidiellum, Bathanthidium, Stelis). Триба Antidiini на территории этого региона наименее представлена и включает всего 2 вида (A. septemspinosum и T. byssina). По видовому и численному обилию в Кузнецком Алатау преобладают представители трибы Megachilini (рис 6.1.2). К ней относится более половины отмеченных здесь видов мегахилид (61 %), их суммарное численное обилие составляет 69 %, что превышает соответствующие показатели в других орографических регионах. Вероятно, это объясняется широкой биотопической приуроченностью и широкой полилектичностью большинства видов этой трибы. Только в этом регионе обнаружен очень редкий C. lanceolata. По численному обилию в Кузнецком Алатау преобладают эвритопный M. ligniseca (25% от сборов в этом регионе) и луговой O. parietina (7,3 %). Численность представителей таких видов, как H. leucomelana, H. tuberculata, M. alpicola и M. ligniseca возрастает по сравнению с численностью в других орографических регионах и провинции в целом. 178 В видовом составе мегахилид преобладают луговые (18 видов, 69 % видового обилия) и лугово-лесные (4 вида, 15 %) виды. По численному обилию выделяются группы луговых (56 %) и эвритопных (55 %) видов. Лугово-лесные виды в Кузнецком Алатау демонстрируют максимальное численное обилие по сравнению с другими регионами (14 %), луговостепные – минимальное (1,6 %), что связано с типом растительности. В этом орографическом регионе преобладает горная тайга, горные луговые степи представлены фрагментарно по южным склонам [Королюк А.Ю., Макунина Н.И., 2000]. В Кузнецком Алатау отсутствуют луговые (A. strigatum, A. manicatum, A. punctatum, B. sibiricum, C. conoidea, L. fuscipennis, O. leaiana, O. spinulosa, O. uncinata, M. centuncularis, M. circumcincta, M. lagopoda, M. melanopyga, S. punctulatissima, S. simillima, S. оrnatula) и лугово-степные (A. tridentatus, C. afra, C. emarginata, H. mitis, O. pilicornis, M. analis, M. argentata) виды. Сравнительные данные по видовому богатству и видовому разнообразию мегахилид разных орографических регионов Кузнецко-Салаирской провинции приведены в табл. 6.2. Таблица 6.2 – Показатели видового разнообразия пчёл сем. Megachilidae в разных орографических регионах Кузнецко-Салаирской провинции Показатель Число видов (видовое богатство) Индекс Симпсона λ' Индекс видового разнообразия PIE' Индекс полидоминантности Вильямса Индекс Шеннона Ish' Мера Кузнецкая котловина Горная Шория Кузнецкий Алатау 46 30 26 0,057 0,076 0,097 0,943 0,924 0,903 17,5 13,1 10,3 1,795 1,245 1,203 «концентрации» Симпсона (λ') является чувствительным индикатором доминирования одного или нескольких видов (отражает обилие обычных видов) [Шитиков В.К., Розенберг Г.С., 2005]. По мере увеличения 179 индекса Симпсона разнообразие уменьшается. Для оценки видового разнообразия удобно использовать обратное ему по смыслу выражение (1 – λ') – показатель вероятности видовых встреч PIE', изменяющийся в пределах от 0 до 1 [Песенко Ю.А., 1982]. Наибольшее видовое разнообразие пчёл сем. Megachilidae установлено для территории Кузнецкой котловины. Показатель PIE' для этого орографического региона равен 0,943. Для Горной Шории и Кузнецкого Алатау индекс PIE' равен 0,924 и 0,903 соответственно. Во всех орографических регионах видовое разнообразие пчёл сем. Megachilidae приближается к максимальному. Индекс Шеннона также показывает, что разнообразие среди исследуемых регионов максимальное видовое свойственно Кузнецкой котловине (Ish' = 1,795). Горные орографические регионы характеризуются меньшим разнообразием (Ish' равен 1,245 для Горной Шории и 1,203 для Кузнецкого Алатау). Индекс полидоминантности Вильямса (1/λ') позволяет произвести оценку типа распределения видовых обилий [Розенберг Г.С., 2007] и показывает вероятное число массовых в исследуемом орографическом регионе видов. Максимальное значение индекс полидоминантности принимает для фауны мегахилид территории Кузнецкой котловины (17,5), минимальное – для фауны Кузнецкого Алатау (10,3). Наибольшее фаунистическое сходство демонстрируют Кузнецкая котловина и Горная Шория. Индекс Чекановского-Съеренсена Кsc для этих территорий равен 0,74 (имеют 28 общих видов). Индексы фаунистического сходства для пар Кузнецкая котловина – Кузнецкий Алатау и Горная Шория – Кузнецкий Алатау равны 0,66 и 0,64 (24 и 18 общих видов соответственно). Из 49 видов пчёл семейства Megachilidae во всех орографических регионах встречаются 17 (35 % от общего числа видов): Ch. rapunculi, C. inermis, C. mandibularis, C. rufescens, H. claviventris, H. leucomelana, H. tridentata, O. parietina, O. bicolor, M. alpicola, M. bombycina, M. lapponica, M. ligniseca, M. maakii, M. versicolor, M. willughbiella, T. byssina. 180 Их суммарное численное обилие составляет 65 % от общих сборов мегахилид в Кузнецко-Салаирской провинции. Эти виды характеризуются широкой биотопической приуроченностью, и относятся к классам доминантов (C. inermis и M. ligniseca) и субдоминантов (за исключением редкого C. mandibularis). Максимальное численное обилие большинство указанных видов демонстрируют в горных регионах, что связано, вероятно, с уменьшением числа особей других видов; только Ch. rapunculi, O. bicolor и H. tridentata имеют большее численное обилие на территории Кузнецкой котловины (6, 4,65 и 4,05 % соответственно). Более равномерно по всем орографическим регионам распределяется C. inermis (6–6,6 % численного обилия). На территории только одного орографического региона встречается 13 видов мегахилид (26,5 % от общего числа видов, 7 % от числа отловленных особей). Из них 11 видов встречается только в Кузнецкой котловине (A. tridentatus, B. sibiricum, C. afra, C. conoidea, H. mitis, O. uncinata, M. argentata, M. circumcincta, S. ornatula, S. punctulatissima, S. simillima). Вид C. lanceolata отмечен только на территории Кузнецкого Алатау, O. leaiana – только в Горной Шории. Большинство этих видов относится к классу очень редких, за исключением субдоминантных H. mitis и S. simillima и редких B. sibiricum и M. circumcincta. Остальные виды (19 видов, 38 % видового обилия, 28 % численного обилия) встречаются в двух орографических регионах, причём на территории только горных регионов отмечен один вид H. tuberculata. Остальные виды обитают на территории Кузнецкой котловины и одного из горных регионов. Из них 11 видов встречаются в Кузнецкой котловине и Горной Шории, 7 видов – в Кузнецкой котловине и Кузнецком Алатау. 181 6.2. Топические комплексы мегахилид 6.2.1. Кузнецкая котловина В Кузнецкой котловине создаются наиболее благоприятные условия для обитания пчёл сем. Megachilidae, благодаря тому, что центральная часть занята типичной лесостепью, сочетающей мелколиственные травяные леса, луговые степи и остепнённые луга. В связи с особенностями рельефа, климата и геологического строения, на территории котловины встречаются разнообразные типы растительности, часто резко контрастирующие друг с другом [Гуляева А.Ф., Климов А.В., 2013]. Вследствие этого, в Кузнецкой котловине исследованы топические комплексы мегахилид в наибольшем числе типичных биотопов (12) (табл. 6.3). Таблица 6.3 – Видовой состав и численное обилие пчёл сем. Megachilidae в различных биотопах Кузнецкой котловины (%) Виды 1 Lithurgini L. fuscipennis Биотопы VI VII 7 8 I 2 II 3 III 4 IV 5 V 6 0,60 – – 2,85 0,45 – – – – – 0,15 – – – – – – 0,15 – – – 0,60 – 0,45 – 0,30 – – 0,15 0,15 – – 2,55 – 2,41 – 0,30 – – 5,70 2,55 1,64 0,75 0,90 4,05 1,79 0,29 3,15 0,14 0,14 – – – – – – – – – – – – – – – – – – – – – – – 0,15 – – – – – 0,15 – – – – – – – 0,15 – 0,89 0,29 4,80 0,74 0,59 1,35 0,09 0,89 0,29 – 0,15 – Всего % VIII 9 IX 10 X 11 XI 12 12 13 – – – – – – 3,90 – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – 0,60 0,15 0,15 – – – – – – – – – – – – – – – – – 6,00 2,70 2,24 0,90 4,35 4,05 4,65 0,29 3,75 0,14 0,14 – – – – – – – 0,05 – – – – 1,05 – – 1,35 – – – 0,15 – – – – – – – – – – – 0,15 – – – – 0,15 0,15 – – – – – – – – – – – – – – – – 0,29 – – – – – – – 0,15 – – – – – – – – – 0,59 1,19 0,29 6,00 0,89 0,59 2,70 0,14 1,19 – – – – – – – – 0,44 14 Osmiini Ch. rapunculi H. acuticornis H. claviventris H. leucomelana H. mitis H. tridentata O. bicolor O. parietina O. pilicornis O. spinulosa O. uncinata Antidiini A. strigatum A. manicatum A. punctatum A.septemspinosum B. sibiricum S. punctulatissima S. simillima S. ornatula T. byssina Dioxyini A. tridentatus 182 Окончание таблицы 6.3 1 Megachilini C. afra C. alata C. conoidea C. elongata C. emarginata C. inermis C. mandibularis C. quadridentata C. rufescens M. alpicola M. analis M. argentata M. bombycina M. centuncularis M. circumcincta M. genalis M. lagopoda M. lapponica M. ligniseca M. maakii M. melanopyga M. rotundata M. versicolor M. willughbiella Всего видов Всего особей, % Индекс Шеннона Ish' 2 3 4 5 6 7 8 9 10 11 12 13 14 – – – – – – – – – – 0,15 – – – – – – – 0,15 – 0,15 – – – 6 0,14 – – – – – – 0,29 – – 0,30 0,14 – – – – – – – 0,15 0,15 1,50 – – 12 – – – – – 0,30 0,29 – – 0,15 1,20 – 0,15 – – 0,75 – 0,60 0,30 – – – 0,15 1,05 18 – – 0,44 0,59 0,29 5,55 – – 1,64 1,49 2,40 – 1,49 0,59 1,19 1,36 1,50 0,90 15,79 0,30 0,30 – 2,85 1,51 37 – – – – – – – – 0,30 – – – – – – – – – – – – – – – 3 – – – – – – – – – – – – 0,15 – – – 0,15 – – – – – – – 4 – – – – – – – – – – – – – – – – – – – – – – – – 1 – – – – – – – – – – – – – – – – – – – – – – – – 1 – – – – – – – – – – 0,15 – – – – – – – – – – – – – 3 – 0,29 – – – – – – – – – – – – – – – – 0,75 – – – – – 2 – – – – – 0,15 – – – – 0,15 – – – – 0,60 – 0,60 – – – – – – 9 – – – – – – – – – – – – – – – – 0,60 – 0,75 – – – – – 2 0,14 0,29 0,44 0,59 0,29 6,00 0,29 0,29 1,94 1,64 4,35 0,14 1,79 0,59 1,19 2,71 2,25 2,10 17,74 1,49 4,32 10,95 73,76 0,8 2,7 0,15 0,15 0,45 1,04 2,84 1,35 100 0,69 0,91 1,01 1,32 0,37 0,44 0,00 0,00 0,47 0,25 0,86 0,29 0,45 0,60 1,50 3,00 2,56 46 Примечание: I – каменистая степь, II – луговая степь, III – разнотравно-злаковый остепнённый луг, IV – разнотравно-злаковый суходольный луг, V – деградированный разнотравно-злаковый суходольный луг, VI – злаково-разнотравно-бобовый суходольный луг, VII – разнотравно-злаковый пойменный луг, VIII – разнотравно-осоковый пойменный луг, IX – разнотравно-злаковый лесной луг, X – берёзовый лес, XI – сосново-берёзовый лес, XII – рудеральные участки. Наибольшее видовое (80,4 %) и численное (73,8 %) обилие пчёл сем. Megachilidae в Кузнецкой котловине установлено для суходольных разнотравно-злаковых лугов, характерных для повышенных участков котловины. Биотопы этого типа характеризуются максимальным видовым разнообразием мегахилид (Ish' = 1,32). Здесь встречаются 38 видов мегахилид (табл. 6.3). В биотопах только этого типа в котловине обнаружены субдоминантные H. tridentata, O. parietina, O. spinulosa, редкие C. elongata, 183 M. centuncularis, M. circumcincta, и очень редкие A. punctatum, C. conoidea, C. emarginata, O. uncinata, S. punctulatissima. Из отмеченных на территории Кузнецкой котловины видов в биотопах этого типа не обнаружены только 9 видов (A. strigatum, A. tridentatus, C. afra, C. alata, C. mandibularis, C. quadridentata, M. argentata, M. lapponica, M. rotundata). Подавляющее число видов (24) в Кузнецкой котловине демонстрируют максимальное численное обилие на суходольных разнотравно-злаковых лугах, по сравнению с другими типами A. septemspinosum, B. sibiricum, Ch. rapunculi, биотопов (A. manicatum, C. inermis, C. rufescens, H. acuticornis, H. claviventris, H. leucomelana, L. fuscipennis, M. alpicola, M. analis, M. bombycina, M. genalis, M. lagopoda, M. lapponica, M. ligniseca, M. maakii, M. melanopyga, M. versicolor, M. willughbiella, O. pilicornis, S. ornatula, T. byssina). Наибольшим численным обилием в биотопах рассматриваемого типа характеризуется триба Megachilini (54 % численного обилия) – за счёт представителей рода Megachile. Половину отловленных особей этого рода составляет M. ligniseca, преобладающий по численности на разнотравнозлаковых суходольных лугах Кузнецкой котловины (составляет 15,8 % от общих сборов в Кузнецкой котловине и 21 % от сборов в биотопах указанного типа). Среди других видов высокое численное обилие демонстрируют Ch. rapunculi (5,7; 7,7 %) и C. inermis (5,5; 7,4 %). Наиболее редок на разнотравно-злаковых суходольных лугах S. ornatula (0,09; 0,1 %). Экологическая структура населения пчёл сем. Megachilidae суходольных разнотравно-злаковых лугов Кузнецкой котловины показана на рис. 6.2.1. В биотопах этого типа как по видовому (78,4 %), так и численному (63,2 %) обилию существенно преобладают луговые виды, на долю видов из других биотопических групп приходится менее четверти видового и менее половины численного обилия. 184 а б Рисунок 6.2.1. Соотношение разных биотопических групп по видовому (а) и численному (б) обилию в населении пчёл сем. Megachilidae разнотравнозлаковых суходольных лугов Кузнецкой котловины, % Остепнённые разнотравно-злаковые луга уступают суходольным разнотравно-злаковым как по числу обнаруженных видов мегахилид (18 видов, 39 % видового обилия Кузнецкой котловины), так и по численному обилию отловленных особей (10,95 %). Только в биотопах этого типа в котловине обнаружен редкий C. mandibularis. Максимальным численным обилием на остепнённых разнотравнозлаковых лугах характеризуется триба Osmiini (50 %); вклад трибы Megachilini в численное обилие составляет 40 %. По численности в биотопах рассматриваемого типа в Кузнецкой котловине преобладают H. mitis (2,55 % от общих сборов в котловине и 23 % от сборов на остепнённых лугах) и О. bicolor (2,41; 22 %), которые демонстрируют на остепнённых разнотравно-злаковых лугах максимальное численное обилие по сравнению с другими типами биотопов. Наиболее редкими на остепнённых разнотравно-злаковых лугах являются A. tridentatus, A. strigatum, Ch. rapunculi, H. acuticornis, M. bombycina, M. versicolor, T. byssina (по 0,15 % от общих сборов в котловине и 1,4 % от сборов на остепнённых лугах). 185 Экологическая структура населения пчёл сем. Megachilidae для остепнённых разнотравно-злаковых лугов Кузнецкой котловины показана на рис. 6.2.2. а б Рисунок 6.2.2. Соотношение разных биотопических групп по видовому (а) и численному (б) обилию в населении пчёл сем. Megachilidae остепнённых разнотравно-злаковых лугов Кузнецкой котловины, % Вклад лугово-степных видов в видовое обилие мегахилид по сравнению с суходольными разнотравно-злаковыми лугами увеличился до 27,8 %. По численному обилию на остепнённых разнотравно-злаковых лугах значительно преобладают лугово-степные виды (59 %), доля видов из других биотопических групп значительно снижена. Луговые степи на территории Кузнецкой котловины также привлекают большое число видов мегахилид (12 видов, 26 % видового обилия исследуемого региона). Однако по численному обилию мегахилид биотопы этого типа значительно уступают разнотравно-злаковым суходольным и остепнённым лугам (здесь отловлено всего 4,32 % особей). Только в биотопах этого типа в Кузнецкой котловине отмечены очень редкие C. quadridentata, M. rotundata и очень редкие C. afra и M. argentata. Основу населения мегахилид луговых степей составляют представители трибы Megachilini (их численное обилие составляет 62 % от всех 186 отловленных здесь особей). Максимальным численным обилием в биотопах этого типа характеризуются M. rotundata (составляет 1,5 % от общих сборах в котловине и 35 % численного обилия луговых степей). Вклад остальных видов в численное обилие невелик. По видовому и численному обилию на луговых степях преобладают представители биотопической группы лугово-степных видов (7 видов, 58 % видового и 77 % численного обилия). В экологической структуре населения мегахилид луговых степей отсутствуют лесные виды. Остальные типы биотопов на территории Кузнецкой малопривлекательны для мегахилид. Меньше всего видов котловины (по 1 виду) обнаружено на разнотравно-злаковых и разнотравно-осоковых пойменных лугах, где в связи с повышенной влажностью создаются не вполне благоприятные условия обитания для этих пчёл. 6.2.2. Кузнецкий Алатау В Кузнецком Алатау преобладает горнотаёжный тип растительности; кроме того, для растительности характерно поясное распределение. В этом орографическом регионе исследовано 9 типичных групп биотопов (табл. 6.4). Максимальное видовое (92,3 %) и численное (69,5 %) обилие мегахилид на территории Кузнецкого Алатау, как и в Кузнецкой котловине, установлено для суходольных разнотравно-злаковых лугов. Здесь отмечено 24 вида пчёл сем. Megachilidae. Индекс видового разнообразия Ish' для этого типа биотопов составляет 1,14 (наибольший показатель среди биотопов Кузнецкого Алатау). Треть видов мегахилид (A. septemspinosum, C. lanceolata, H. acuticornis, O. bicolor, M. bombycina, M. rotundata, M. versicolor, M. willughbiella, T. byssina) в этом орографическом регионе встречаются в биотопах только этого типа. Из отмеченных на территории Кузнецкого Алатау видов на суходольных разнотравно-злаковых лугах не обнаружены только 2 вида – H. tuderculata и C. elongata. 187 Таблица 6.4 – Видовой состав и численное обилие пчёл сем. Megachilidae в различных биотопах Кузнецкого Алатау (%) Виды 1 Osmiini Ch. rapunculi H. acuticornis H. claviventris H. leucomelana H. tridentata H. tuderculata O. bicolor O. parietina Биотопы V VI 6 7 I 2 II 3 III 4 IV 5 3,93 0,58 2,77 5,60 0,56 – 1,12 1,16 – – 0,58 – – – – – – – – – 0,56 – – – – – – – – 1,68 – 6,14 – – – – – – – – 1,12 3,37 – – – – – – 0,56 – 1,55 0,58 1,67 1,67 1,12 2,52 2,25 2,77 5,55 20,08 0,56 0,56 2,25 5,62 24 – – 0,58 – – – – – – 0,58 – 0,58 – – – – 4 – – 0,58 – – – – – – 0,58 – – 0,56 – – – 4 69,50 2,32 1,14 0,60 Всего, % VII 8 VIII 9 IX 10 – – – – – – – – – – – – – – – – – – 0,58 0,58 – – – – 0,58 – – – – – – – 4,51 0,58 3,93 6,18 1,12 1,68 1,12 7,30 – – – – – – – – – – 1,12 3,37 – 0,56 1,73 – – – – – – – – – – – – – 4 0,56 – 0,58 – – – 1,13 2,53 – – 2,31 – – – – – 5 – – 0,58 – – – – – – – – – – – – – 1 – – – – 0,58 0,58 – – – – – – – – – – 2 – – – – – – – – – – – 4,62 – – – – 3 – – 0,58 – – – – – – – – – – – – – 2 1,12 0,56 6,18 0,56 2,25 2,25 2,25 5,05 2,25 3,93 7,86 25,28 1,12 0,56 2,25 5,62 2,28 10, 11 7,11 0,58 1,16 5,78 1,16 100 0,59 0,46 0,62 0 0,30 0,27 0,3 11 Antidiini A. septemspinosum T. byssina Megachilini C. alata C. elongata C. inermis C. lanceolata C. mandibularis C. quadridentata C. rufescens M. alpicola M. bombycina M. genalis M. lapponica M. ligniseca M. maakii M. rotundata M. versicolor M. willughbiella Всего видов Всего особей, % Индекс Шеннона Ish' 26 Примечание: I – суходольный разнотравно-злаковый луг, II – деградированный суходольный разнотравно-злаковый луг, III – пойменный разнотравно-злаковый луг, IV – лесной разнотравно-злаковый луг, V – лесной высокотравный луг, VI – осиновоберёзовый лес, VII – пихтово-берёзовый лес, VIII – черневая тайга, IX – рудеральные участки. Наибольшее численное обилие на суходольных разнотравно-злаковых лугах по сравнению с другими местообитаниями в Кузнецком Алатау демонстрируют эвритопный M. ligniseca, луговые Ch. rapunculi, C. quadridentata, M. genalis, M. lapponica и лугово-лесные H. claviventris, H. leucomelana, C. mandibularis. 188 Наибольшим численным обилием в биотопах рассматриваемого типа характеризуется триба Megachilini (49 % численного обилия). Из них 40 % отловленных особей составляют преобладающего по численности представители M. ligniseca, на суходольных разнотравно-злаковых лугах Кузнецкого Алатау (составляет 20 % от общих сборов в этом орографическом регионе и 29 % от сборов в исследуемом типе биотопов). Высокое численное обилие демонстрируют также M. willughbiella (5,62; 8 %) и H. leucomelana (5,60; 8 %). Редки на суходольных разнотравно-злаковых лугах H. acuticornis, H. tridentata, C. alata, M. maakii, M. rotundata (0,56–0,58 % от общих сборов в Кузнецком Алатау и 0,8 % от особей, отловленных в биотопах этого типа). Из рис. 6.2.3 видно, что на суходольных разнотравно-злаковых лугах Кузнецкого Алатау по видовому и численному обилию преобладает биотопическая группа луговых видов (70,8 и 51, 2 % соответственно). а б Рисунок 6.2.3. Соотношение разных биотопических групп по видовому (а) и численному (б) обилию в населении пчёл сем. Megachilidae разнотравнозлаковых суходольных лугов Кузнецкого Алатау, % Вклад лугово-лесных и эвритопных видов мегахилид в видовое и численное обилие по сравнению с биотопами Кузнецкой котловины 189 значительно возрастает (12,6 и 14 %, 8,3 и 32,3 % соответственно). Вклад лугово-степных видов минимален и составляет 8,3 % видового и 2,5 % численного обилия. Лесные разнотравно-злаковые луга характеризуются достаточно высоким численным обилием пчёл сем. Megachilidae (10 % от сборов в Кузнецком Алатау), однако существенно уступают суходольным лугам по числу обнаруженных видов (здесь отмечено всего 4 вида мегахилид). Из них два вида встречаются только в биотопах указанного типа (H. tuberculata и C. elongata), а остальные демонстрируют на лесных разнотравно-злаковых лугах максимальное численное обилие по сравнению с другими биотопами, где они встречаются (C. inermis, O. parietina). Максимальной численностью на лесных разнотравно-злаковых лугах характеризуется O. parietina (6,14 % от сборов в Кузнецком Алатау и 61 % от сборов в биотопах этого типа). Наиболее редок C. elongata (0,56; 5,5 %). Большее число видов пчёл сем. Megachilidae отмечено на лесных высокотравных лугах (5 видов – C. alata, C. inermis, C. rufescens, M. alpicola, M. lapponica). По численному обилию мегахилид этот тип биотопов уступает разнотравно-злаковым лесным лугам – здесь отловлено 7 % пчёл. Остальные типы биотопов в Кузнецком Алатау малопривлекательны для мегахилид, численное обилие отловленных в них особей составляет от 0,58 до 5,78 %, число отмеченных видов – 1–4. Наименьшее число видов отмечено в осиново-берёзовых лесах (1 вид – C. inermis). 6.2.3. Горная Шория Этот орографический регион характеризуется преобладанием осиновопихтовых лесов (не менее 90 % площади). В Горной Шории исследовано население мегахилид в наименьшем числе типов биотопов – 7 (табл. 6.4). 190 Таблица 6.4 – Видовой состав и численное обилие пчёл сем. Megachilidae в различных биотопах Горной Шории (%) Виды 1 Lithurgini L. fuscipennis Osmiini Ch. rapunculi H. claviventris H. leucomelana H. tridentata H. tuberculata O. bicolor O. leaiana O. parietina O. pilicornis O. spinulosa I 2 II 3 III 4 Биотопы IV 5 V 6 VI 7 VII 8 0,83 – – – – – – 0,83 3,93 0,84 2,07 0,68 – 0,42 0,42 1,25 1,25 10,43 – – – – – – – – – – 0,57 – – – – – – 1,24 – 1,15 – – – 0,57 – – – – – – – – – – – – – – – – – – – – 0,42 – – – – – – – – – – – – 1,24 – – 4,50 0,84 2,07 1,25 0,42 0,42 0,42 3,73 1,25 11,58 1,72 0,83 – 1,66 – – – – 4,07 – 0,41 1,65 – – – – – – – – – – – – – – – – 5,79 0,83 0,41 3,31 – 3,31 – – 0,41 0,82 0,82 2,48 0,82 4,54 12,75 5,78 1,65 4,96 10,33 25 – – – – – – – – – – 2,30 – – – – 1 – 3,30 1,86 1,24 – – – – – – – – – – – 9 – – 1,86 – – – – – – – – – – – – 2 – – – – – – – – – – 2,30 – – – – 1 – – – – – – – – – – – – – – 0,41 2 0,41 – – – – – – – – – – – – – – 2 0,41 6,61 3,72 1,24 0,41 0,82 0,82 2,48 0,82 4,54 17,35 5,78 1,65 4,96 10,74 75,01 2,30 15,49 2,42 2,30 0,83 1,65 100 1,17 0 0,86 0,23 0 0,30 0,24 Всего, % 9 Ahthidiini A. strigatum A. manicatum A. punctatum T. byssina Megachilini C. emarginata C. inermis C. mandibularis C. rufescens M. alpicola M. analis M. bombycina M. centuncularis M. lagopoda M. lapponica M. ligniseca M. maakii M. melanopyga M. versicolor M. willughbiella Всего видов Всего особей, % Индекс Шеннона Ish' 30 Примечание: I – суходольный разнотравно-злаковый луг, II – деградированный суходольный разнотравно-злаковый луг, III – лесной разнотравно-злаковый луг, IV – лесной высокотравный луг, V – берёзово-осиновый лес, VI – берёзово-пихтовый лес, VII – осиново-пихтовый лес. Наибольшее число видов пчёл сем. Megachilidae в Горной Шории отмечено на суходольных разнотравно-злаковых лугах (25 видов, 83 % 191 видового обилия этого орографического региона). Указанный тип биотопов характеризуется также максимальным численным обилием особей (75 %). Разнотравно-злаковые суходольные луга Горной Шории характеризуются максимальным видовым разнообразием по сравнению с другими типами биотопов этого орографического региона (индекс Шеннона равен 1,17). Более половины видов мегахилид в Горной Шории встречаются только на суходольных разнотравно-злаковых лугах (16 видов – A. manicatum, H. claviventris, H. leucomelana, L. fuscipennis, O. bicolor, O. leaiana, O. pilicornis, M. alpicola, M. analis, M. bombycina, M. centuncularis, M. lagopoda, M. lapponica, M. maakii, M. melanopyga, M. versicolor). Из отмеченных в Горной Шории видов в биотопах этого типа не встречаются только 5 видов (A. punctatum, H. tuberculata, C. emarginata, C. inermis, C. rufescens). Большинство видов (за исключением A. strigatum) демонстрируют на суходольных разнотравно-злаковых лугах максимальное численное обилие в этом орографическом регионе. Наибольшим численным обилием в биотопах указанного типа характеризуются M. ligniseca (составляет 12,7 % численного обилия отловленный в Горной Шории особей мегахилид и 17 % от сборов на суходольных разнотравно-злаковых лугах), O. spinulosa (10,4; 14 %) и M. willughbiella (10,3; 13,7 %). Из рис. 6.2.4 видно, что основу населения мегахилид исследуемого типа биотопов в Горной Шории составляют луговые виды мегахилид, преобладающие по видовому (76 %) и численному (82,5 %) обилию. Лесные разнотравно-злаковые луга Горной Шории уступают суходольным разнотравно-злаковым как по числу обнаруженных видов мегахилид (9 видов, 30 % видового обилия), так и по численному обилию отловленных особей (15,5 %). Только в биотопах этого типа в Горной Шории встречаются A. punctatum и C. rufescens. 192 а б Рисунок 6.2.4. Соотношение разных биотопических групп по видовому (а) и численному (б) обилию в населении пчёл сем. Megachilidae разнотравнозлаковых суходольных лугов Горной Шории, % Максимальным численным обилием на лесных разнотравно-злаковых лугах характеризуются представители видов A. strigatum (4 % от общих сборов в Горной Шории и 26 % от особей, отловленных в биотопах этого типа), и C. inermis (3,3; 21 %). Редко встречаются Ch. rapunculi (0,57; 3,6 %) и A. punctatum (0,4; 2,5 %). Большая часть видов, встречающихся в этом биотопе (за исключением лугово-лесного C. mandibularis), относится к биотопической группе луговых видов. В остальных исследованных типах биотопов Горной Шории число обнаруженных видов пчёл сем. Megachilidae не превышает 2. Наименее привлекательными для мегахилид в этом орографическом регионе являются деградированные суходольные луга и берёзово-осиновые леса (здесь встречается только 1 вид – эвритопный M. ligniseca). 193 6.3. Пчёлы сем. Megachilidae в антропогенных местообитаниях На территории подвержены Кемеровской сильной области антропогенной естественные нагрузке. экосистемы Бурное развитие горнодобывающей промышленности влечёт за собой катастрофическое изменение природных систем. Площадь нарушенных земель здесь превышает 100 тыс. га; около 30 % территории области по всем признакам соответствуют зоне экологического бедствия [Шереметова С.А., 2012]. По степени возрастания антропогенной нагрузки на исследуемой территории выделено 3 группы антропогенных ландшафтов. К слабо изменённым ландшафтам относятся такие местообитания, как лесопарки, водоёмы и овраги, сохранившиеся на антропогенно-изменённых территориях [Рахимов И.И., Уленгов Р.А., 2004]. В пределах г. Кемерово к данному типу ландшафтов также относится сосновый бор – сохранившийся участок барзасской тайги, расположенный на правом берегу р. Томь. В целом в ландшафтах этого типа в Кузнецко-Салаирской провинции отмечено 8 видов пчёл сем. Megachilidae – A. septemspinosum, B. sibiricum, C. inermis, H. acuticornis, H. mitis, L. fuscipennis, O. bicolor, M. versicolor (табл. 6.5). Среди них только H. acuticornis не заходит в местообитания с более высокой антропогенной трансформацией. Эти виды относятся к луговой и лугово-степной группам и характеризуются широкой биотопической приуроченностью. Небольшое число видов мегахилид, характерных для слабо изменённых территорий, вероятно связано с низкой привлекательностью этих участков по сравнению с естественными местообитаниями (в первую очередь, в отношении кормовой базы). Тем не менее, лесопарки и другие малоизмененные природно-территориальные комплексы предоставляют пчелам относительно пригодные места гнездования и кормовую базу и являются важным местообитанием диких пчёл в антропогенной среде [Пантова А.Ю., 2013]. Численное обилие представителей указанных видов составляет 26,2 % от числа особей, отловленных на антропогенно изменённых территориях. 194 Таблица 6.5 – Пчёлы сем. Megachilidae в антропогенно преобразованных биотопах Кузнецко-Салаирской провинции Биотоп Вид Биотопическая группа 1 Урбофильные виды A. manicatum A. septemspinosum Ch. rapunculi C. inermis H. tridentata L. fuscipennis O. bicolor M. ligniseca M. versicolor Урботолерантные виды B. sibiricum C. alata C. emarginata C. rufescens H. claviventris H. mitis O. pilicornis M. alpicola M. bombycina M. genalis M. lagopoda M. lapponica M. rotundata M. willughbiella S. ornatula S. punctulatissima S. simillima T. byssina Урбофобные виды A. tridentatus A. strigatum A. punctatum C. afra C. elongata C. conoidea C. lanceolata C. mandibularis C. quadridentata H. acuticornis H. leucomelana H. tuberculata слабо изменённые изменённые (нарушенные) преобразованные I II III IV V VI VII VIII IX 2 3 4 5 6 7 8 9 10 11 Л Л Л Л Л Л ЛС Э Л – + – – – + + – + – + – + – + + – + – – – + – + – + – – + – + – + + + + – – + + + + – + – – – – + – – – – – + + + + + + – + + + + + + + + + + + – – – – + – – – – Л Л ЛС Л ЛЛ ЛС ЛС Л Л Л Л Л ЛС Л Л Л Л Л – – – – – + – – – – – – – – – – – – + – – – – + – – – – – – – – – – – – – – – + + – – – – + – – – – – – – – + + + – – – – + + + – + + – – + – – – – – – – – – – – – + – – – + – – – – + – + + + + – – + – + – – – – – – + + + + – – – + + + + + + + – + + + + + – + + – – + + + – – + + – + + + – – – – – – – – + – + – – + – – – – ЛС Л Л ЛС Л Л Л ЛЛ Э Л ЛЛ ЛЛ – – – – – – – – – – – – – – – – – – – – – + – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – – 195 Окончание таблицы 6.5 1 O. parietina O. spinulosa O. leaiana O. uncinata M. analis M. argentata M. centuncularis M. circumcincta M. maakii M. melanopyga 2 Л Л Л Л ЛС ЛС Л Л Л Л 3 – – – – – – – – – – 4 – – – – – – – – – – 5 – – – – – – – – – – 6 – – – – – – – – – + 7 – – – – – – – – – – 8 – – – – – – – – – – 9 – – – – – – – – – – 10 – – – – – – – – – – 11 – – – – – – – – – – Примечание. Антропогенные местообитания: I – сохранившиеся луговые биотопы в черте города, II – лесопарки, сосновые бор (г. Кемерово), III – деградированные луга, IV – агроценозы, V – рудеральные местообитания, VI – обочины дорог, VII – сельская застройка, VIII – городская застройка, IX – отвалы шахт. Биотопические группы: Л – луговая, ЛС – лугово-степная, ЛЛ – лугово-лесная, Э – эвритопная. В группу нарушенных (изменённых) ландшафтов включены деградированные луга, агроценозы, рудеральные участки, парки, сады и др. [Рахимов И.И., Уленгов Р.А., 2004]. В местообитаниях этого типа встречаются 22 вида мегахилид, что обусловлено обилием кормовых растений (в том числе культурных и рудеральных). Численное обилие представителей этих видов составляет 31,3 % от числа особей, отловленных на антропогенно изменённых территориях. Для обитания пчёл в агроландшафтах важным условием является наличие в них длительно цветущего набора культурных и дикорастущих растений, а также сохранение от распашки многолетних поселений пчёл, гнездящихся в почве. Существенное негативное влияние на биоту агроландшафтов оказывает комплекс агротехнических мероприятий [Ченикалова Е.В., 2005]. Местами сохранения и размножения диких пчелиных (своеобразными рефугиумами) в агроценозах часто выступают облесенные овраги, пустыри, полосы отчуждения шоссейных дорог и др. [Сапаев Е.А., 1993]. Городские парки как места обитания пчелиных характеризуются повышенной антропогенной нагрузкой в виде стрижки газонов, вытаптывания, неравномерностью распределения кормовых растений, что ведёт к нестабильности кормовой базы [Богатырёв Н.Р., 1985]. 196 Все отмеченные характеризуются в изменённых широкой местообитаниях биотопической виды мегахилид приуроченностью (за исключением S. ornatula и S. punctulatissima, предпочитающих луговые местообитания). В антропогенных местообитаниях только этого типа отмечены виды S. ornatula и M. melanopyga. К преобразованным местообитаниям отнесены дороги, городские и сельские застройки [Рахимов И.И., Уленгов Р.А., 2004]. В этих местообитаниях отмечены представители 26 видов пчёл сем. Megachilidae. Их них 5 видов (A. manicatum, O. pilicornis, S. simillima, T. byssina, M. willughbiella) обнаружены в антропогенных ландшафтах только этого типа. Остальные виды встречаются также в изменённых и слабо изменённых местообитаниях. Большинство видов пчёл сем. Megachilidae, отмеченных на преобразованных участках, являются представителями луговой группы, только O. pilicornis относится к лугово-степным видам. Многих пчёл преобразованные местообитания привлекают, вероятнее всего, наличием удобных мест для постройки гнёзд, а также кормовой базой, включающей большое число культурных растений (огороды частного сектора, городские клумбы). Отмечено, что обочины дорог зачастую являются местом для гнездования пчелиных [Чернышев В.Б. и др., 2010]. Кроме того, высокое видовое разнообразие мегахилид в преобразованных местообитаниях связано с отсутствием пестицидного пресса [Сапаев Е.А., 1993; Ченикалова Е.В., 2005]. Численное обилие представителей видов, отмеченных в местообитаниях преобразованного типа, составляет 42,5 % от числа особей, отловленных на антропогенно изменённых территориях. Во всех трёх типах изменённых ландшафтов встречаются 6 видов пчёл сем. Megachilidae (L. fuscipennis, O. bicolor, A. septemspinosum, B. sibiricum, C. inermis, M. versicolor). Как правило, это луговые виды (за исключением лугово-степного O. bicolor), по численному обилию относящиеся к 197 категориям доминантных (C. inermis) и субдоминантных видов. В изменённых местообитаниях двух типов (нарушенных и преобразованных) отмечены 14 видов мегахилид. Один вид (H. mitis) встречается в местообитаниях слабо изменённых и преобразованных человеком. Одним из факторов, способствующих приспособлению мегахилид к видоизменённым и преобразованным местообитанием, является широкая экологическая пластичность, свойственная представителям данного семейства. Она выражается в способности самок многих видов сооружать ячейки гнёзд в любых готовых полостях разнообразных субстратов, включая искусственные материалы, деревянные и каменные постройки и др. Кроме того, многие виды мегахилид являются политрофами или широкими олиготрофами, что позволяет им использовать в качестве кормовой базы любые доступные растения, в том числе произрастающие в черте города на клумбах, в скверах и парках, огородах и садах. 6.4. Пчёлы сем. Megachilidae в городской среде Антропогенная нагрузка прямо и косвенно влияет на видовой состав и состояние популяций перепончатокрылых насекомых, особенно пчелиных. В условиях городской среды население пчёл существенно отличается от природных местообитаний и зависит от рельефа, развитости растительного покрова, состава и стабильности кормовой базы и наличия пригодных мест для сооружения гнёзд [Бейко В.Б., Березин М.В., 1992, 2000; Бейко В.Б., 2000; Конусова О.Л., Янюшкин В.В., 2000]. Исследования пчёл в условиях городской среды на территории России проводились в Москве [Бейко В.Б., Березин М.В., 1992, 2000; Бейко В.Б. 2000; Левченко Т.В., 2010], Казани [Сапаев Е.А., 1989, 1993], Перми [Лыков В.А., 1992, 1993, 1995], республике Башкортостан [Курмакаев Е.В., 2003] и других территориях. Большинство исследований касаются пчёл трибы Bombini, удобных для использования в качестве индикаторов степени урбанизированности ландшафтов. На территории Западной Сибири 198 подобные исследования проводили в г. Новосибирске [Богатырёв Н.Р., 1985; Бывальцев А.М., 2009, 2010], Томске [Конусова О.Л., 2000 б; Конусова О.Л., Янюшкин В.В., 2000], Кемерово [Еремеева Н.И., 2002; Еремеева Н.И., Лузянин С.Л., 2008; Еремеева Н.И. и др., 2010]. По степени устойчивости к воздействию факторов городской среды на территории Кузнецко-Салаирской провинции выделены 3 экологические группы мегахилид (рис. 6.4.1). Рисунок 6.4.1. Соотношение пчёл сем. Megachilidae разных групп устойчивости к воздействию факторов городской среды по видовому обилию, % Урбофильные виды. Группа характеризуется минимальным видовым обилием и насчитывает всего 9 видов пчёл сем. Megachilidae (18 % всех видов мегахилид исследуемой территории) (табл. 6.5). Отмечается, что снижение видового многообразия пчёл на урбанизированных территориях связано со слабой озеленённостью и нарушенностью местообитаний видов, гнездящихся в почве [Конусова О.Л., 2000 а]. У видов мегахилид урбофильной группы в условиях высоких 199 антропогенных нагрузок возрастает численность [Бейко В.Б., Березин М.В., 1992; Еремеева Н.И. и др., 2010]. К ним в Кузнецко-Салаирской провинции относятся преимущественно экологически пластичные виды, значительно повышающие численность на относительно нетронутых участках природных экосистем в черте города, рудеральных территориях и пустырях в связи со сниженной конкуренцией со стороны других видов (H. tridentata, L. fuscipennis, M. ligniseca, M. versicolor, O. bicolor). Для O. bicolor важным условием, позволяющим поддерживать высокую численность в урбанизированных ландшафтах г. Кемерово является обилие раковин пресноводных моллюсков на берегах р. Томь в городской черте. Известно, что именно дефицит мест для гнездования является для этого вида лимитирующим фактором в урбанизированных ландшафтах [Красная книга г. Москвы…, 2011]. Литературные данные [Лыков В.А., 1993: Конусова О.Л., 2000 а] также подтверждают, что L. fuscipennis входит в число фоновых видов мегахилид во всех городских биотопах. Кроме того, к этой группе относятся те виды мегахилид, которые способны заселять постройки человека (A. manicatum, A. septemspinosum). Способность большинства видов пчёл рода Anthidium заселять городские местообитания подтверждается во многих работах [Сапаев Е.А., 1989; Лыков В.А., 1992; Амолин А.В., 2008; Левченко Т.В., 2010]. Из рис. 6.4.2 видно, что если в естественных местообитаниях представители разных экологических групп пчёл сем. Megachilidae вносят примерно равный урбанизированных урбофильных видов вклад в суммарное территориях мегахилид численной исследуемого значительно обилие, региона превышает то на численность численность урботолерантных и урбофобных видов и составляет 68 % от общего числа отловленных особей. 200 Рисунок 6.4.2. Соотношение численности разных групп мегахилид в естественных (чёрный цвет) и урбанизированных (серый цвет) ландшафтах, %. Анализ экологической структуры урбофильной группы мегахилид показывает, что большинство видов в её составе относятся к луговым (7 из 9 видов, 78 %), и являются полилектами (62 %) или широкими олиголектами (25 %). Интересна урбофильность узкого олиголекта Ch. rapunculi, проявляющаяся в резком увеличении численности этого вида в городских поселениях в связи с высаживанием на клумбы колокольчиков (Campanula) в декоративных целях. Данные об урбофильности Ch. rapunculi в Кемеровской области противоречат результатам исследований, проведённых в г. Москве и Московской области. В.Б. Бейко [2000] отмечает, что на территории Москвы полностью исчезли пчёлы-мегахилиды рода Chelostoma в связи с исчезновением основных кормовых растений. Однако исследования фауны пчёл в г. Томске подтверждают, что Ch. rapunculi входит в группу наиболее массовых видов пчёл городской среды и отмечен в числе видов, наиболее устойчивых к антропогенному воздействию [Конусова О.Л., Янюшкин В.В., 201 2000]. Увеличение численности представителей р. Chelostoma на урбанизированной территории выявлено также в г. Пермь [Лыков В.А., 1992, 1993]. Все урбофильные виды пчёл сем. Megachilidae на исследуемой территории характеризуются широкой биотопической приуроченностью. К урбофильным видам в Кузнецко-Салаирской провинции относятся оба вида-доминанта (C. inermis, M. ligniseca); 6 видов относятся к категории субдоминантов, за исключением редкого A. manicatum. По численности среди урбофильных видов преобладают M. ligniseca (25 % отловленных особей) и A. septemspinosum (15 %). Преобладание особей этих видов связано с разнообразием экологических и микроклиматических условий в городах [Сапаев Е.А., 1993]. Наиболее распространённый в Кузнецко-Салаирской провинции эвритопный M. ligniseca является самым экологически пластичным видом пчёл сем. Megachilidae на исследуемой территории. Он встречается в максимальном числе биотопов и посещает наибольшее число видов кормовых растений по сравнению с представителями других видов мегахилид. Однако литературные данные не содержат прямых сведений о приуроченности M. ligniseca к городским местообитаниям. [1989] отмечает, что представители этого вида Е.А. Сапаев встречается на урбанизированных территориях единично (г. Казань). Урботолерантные виды. В составе этой группы насчитывается 18 видов пчёл сем. Megachilidae (37 % видового обилия мегахилид Кузнецко- Салаирской провинции). Они в равной мере встречаются в природных и антропогенных биотопах разной степени преобразованности, в том числе в урбанизированных (рис 6.4.2). В городах Кемеровской области, как правило, тяготеют к участкам частного сектора, огородам (C. rufescens, M. alpicola, M. rotundata), пустырям и рудеральным участкам, заросшим свалкам (M. lagopoda, виды рода Stelis), а также малозатронутым участкам естественных природных местообитаний, сохранившимся в черте города (луга, сосновый бор в г. Кемерово) (B. sibiricum, C. alata, C. emarginata, 202 H. claviventris, H. mitis, T. byssina, M. bombycina, M. genalis, M. lapponica, M. willughbiella, O. pilicornis). Большинство видов урботолерантной группы относится к луговым видам (76 %) и характеризуются широкой биотопической приуроченностью (83 %). По категориям численности группа представлена главным образом видами-субдоминантами (12 из 18 видов, 66 %). Большинство из указанных урбофильных и урботолерантных видов пчёл сем. Megachilidae в антропогенных и урбанизированных ландшафтах легко привлечь с помощью искусственных гнездовий [Мариковская Т.П. и др., 2001]. Урбофобные виды. Эта группа характеризуется максимальным видовым обилием и насчитывает 22 видa пчёл сем. Megachilidae (45 % видового обилия мегахилид исследуемой территории) (табл. 6.5). В неё входят виды, избегающие урбанизированные и преобразованные человеком территории; при росте урбанизации среды их численность сокращается [Бейко В.Б., Березин М.В., 1992]. Из всех представителей урбофобных видов только H. acuticornis и M. melanopyga встречаются в антропогенных местообитаниях (слабо изменённых и изменённых (нарушенных) ландшафтах). В состав группы урбофобных видов входит большинство лугово-лесных видов мегахилид (H. leucomelana, H. tuberculata, C. mandibularis), 4 луговостепных вида (A. tridentatus, C. afra, M. analis, M. argentata), один эвритопный (C. quadridentata). Остальные 13 видов мегахилид принадлежат к биотопической группе луговых видов. Подавляющее большинство урбофобных видов (14 из 18, 78 %) характеризуются узкой биотопической приуроченностью. По категориям численного обилия урбофобные виды распределяются следующим образом: к категории очень редких относятся 10 видов мегахилид (45 % видового обилия), к категории редких – 5 видов (23 %). Остальные являются субдоминантами. 203 Сравнение полученных данных с литературными показывает значительное сходство в распределении пчёл сем. Megachilidae антропогенных территориях. Большинство авторов на отмечают благоприятность многих урбанизированных ландшафтов для обитания пчелиных, а наличие подходящих мест для гнездования и обилие кормовых ресурсов считают основным фактором, привлекающим пчёл на урбанизированные территории. О.Л. Конусова (2000 а) отмечает заметную роль пчёл сем. Megachilidae в зоне малоэтажной застройки жилых районов г. Томска. Анализ структуры популяций пчёл, в том числе мегахилид, позволяет судить о степени антропогенного давления на ландшафты. Пчёлы сем. Megachilidae могут быть использованы в качестве индикаторов урбанизированности территории [Бейко В.Б., Березин М.В., 1992; Курмакаев Е.В., 2003]. Таким образом, на территории Кузнецко-Салаирской провинции установлено неравномерное распределение населения пчёл сем. Megachilidae по орографическим регионам. Наибольшее число видов мегахилид отмечено в Кузнецкой котловине (46 видов, 94 % от общего числа видов исследуемой территории); суммарное численное обилие отловленных здесь особей составляет 60 % от общих сборов. В пределах только Кузнецкой котловины встречаются 11 видов мегахилид. На территории Горной Шории встречаются 30 видов пчёл сем. Megachilidae (61 % видового обилия). Численное обилие особей составляет около 22 % от общих сборов. В Кузнецком Алатау отмечено наименьшее число видов пчёл сем. Megachilidae (26 видов, 53 % видового обилия Кузнецко-Салаирской провинции). Численное обилие мегахилид в этом регионе составляет 16 % от общих сборов. Основу населения мегахилид всех орографических регионов КузнецкоСалаирской провинции составляют представители трибы Megachilini (к этой трибе принадлежат 52 % видов мегахилид Кузнецкой котловины, 50 % видов Горной Шории и 61 % видов Кузнецкого Алатау). Численное обилие 204 представителей указанной трибы в орографических регионах составляет 53, 61 и 69 % соответственно. По видовому и численному обилию во всех орографических регионах преобладают виды луговой биотопической группы. Максимальное численное обилие везде демонстрирует эвритопный M. ligniseca (17 % в Кузнецкой котловине, 17,3 % в Горной Шории и 69 % в Кузнецком Алатау). Из 49 видов пчёл сем. Megachilidae 17 видов (35 % от общего числа видов) встречается во всех орографических регионах; 13 видов (26, 5 %) отмечено на территории только одного региона, остальные 19 видов (38 %) встречаются в двух орографических регионах. Наибольшее котловина и фаунистическое Горная Шория сходство демонстрируют (коэффициент сходства Кузнецкая Чекановского- Съёренсена для этих территорий равен 0,74). Наибольшее число типичных биотопов (12) исследовано на территории Кузнецкой котловины. В этом орографическом регионе наибольшее видовое (38 видов, 80,4 %) и численное (73,8 %) обилие пчёл сем. Megachilidae установлено для разнотравно-злаковых суходольных лугов. Из встречающихся в биотопах этого типа видов мегахилид представители 24 видов демонстрируют здесь максимальное численное обилие по сравнению с биотопами других типов. Остепнённые разнотравно-злаковые луга и луговые степи уступают разнотравно-злаковым суходольным лугам по видовому и численному обилию мегахилид (39, 11 и 26, 4,3 % соответственно). Остальные биотопы региона мало привлекательны для пчёл этого семейства. В Кузнецком Алатау исследовано 9 типичных биотопов. Максимальное видовое (24 вида, 92 %) и численное (69 %) обилие мегахилид также установлено для разнотравно-злаковых суходольных лугов. Меньшее видовое и численное обилие пчёл сем. Megachilidae в Кузнецком Алатау демонстрируют разнотравно-злаковые лесные и высокотравные лесные луга (15, 10 и 19, 7 % соответственно). 205 Наименьшее число биотопов исследовано в Горной Шории (7). Максимальное видовое (83 %) и численное (75 %) обилие мегахилид отмечено на суходольных разнотравно-злаковых лугах. Более половины видов этого региона встречается в биотопах только этого типа. Большинство видов демонстрирует здесь максимальное численное обилие. Разнотравнозлаковые лесные луга горной Шории уступают суходольным в отношении видового (30 %) и численного (15,5 %) обилия мегахилид. По численности на разнотравно-злаковых суходольных лугах всех орографических регионов преобладают представители трибы Megachilini. Максимальное численное обилие в биотопах этого типа демонстрирует M. ligniseca (21 % от сборов на разнотравно-злаковых суходольных лугах в Кузнецкой котловине, 29 % – в Кузнецком Алатау, 17 % – в Горной Шории). В антропогенно преобразованных местообитаниях население мегахилид существенно отличается от природных территорий. На преобразованных человеком территориях отмечены 29 видов мегахилид (59 % видового обилия). По степени возрастания антропогенной нагрузки на исследуемой территории выделено 3 группы ландшафтов (слабо изменённые, изменённые (нарушенные) и преобразованные). Из них наибольшее видовое и численное обилие пчёл сем. Megachilidae отмечено в местообитаниях преобразованного типа (обочины дорог, сельская застройка, урбанизированные территории), что объясняется разнообразием экологических и микроклиматических условий, обилием мест для гнездования и отсутствием пестицидного пресса. По степени устойчивости к факторам городской среды выделено 3 группы мегахилид. К группе урбофильных относятся 9 видов мегахилид (18 % видового обилия), увеличивающих свою численность на урбанизированных территориях по сравнению с природными биотопами в 1,7 раз (40 % от сборов в природных биотопах и 68 % численного обилия от сборов в городской среде). Виды этой группы характеризуются высокой экологической пластичностью, широкой биотопической приуроченностью, 206 полилектичностью и способностью заселять разнообразные постройки человека. Численность представителей этой группы в городской среде почти вдвое превышает численность других групп (урботолерантной, включающей 18 видов, и урбофобной – 22 вида). 207 ВЫВОДЫ 1. Видовое разнообразие пчёл семейства Салаирской провинции представлено Кузнецко- Megachilidae 49 видами 12 родов 5 триб подсемейства Megachilinae. Впервые для фауны Западной Сибири указаны 1 род (Bathanthidium Mavromoustakis, 1953) и 6 видов мегахилид, для Кузнецко-Салаирской провинции – 13 видов. 2. Основу населения мегахилид составляют представители трибы Megachilini (25 видов, 51 % видового и 57 % численного обилия). Доминируют два вида – Megachile ligniseca и Coelioxys inermis (18,6 и 6,2 %, соответственно, от общих сборов мегахилид). К субдоминантам относятся 25 видов, редким – 9, очень редким – 13. 3. В Кузнецкой котловине отмечено наибольшее число видов – 46 (94 % – видовое и 60 % – численное обилие). В горных районах видовое разнообразие пчёл семейства Megachilidae значительно снижается и составляет 26–30 видов в различных орографических регионах. 4. Предпочитаемыми биотопами пчёл сем. Megachilidae в Кузнецко- Салаирской провинции являются: суходольные (43 вида, 71,5 % от общих сборов) и остепнённые (18; 10,9) разнотравно-злаковые луга, луговые степи (12; 4,3) Кузнецкой котловины и лесные разнотравно-злаковые луга (9; 15,5) Горной Шории. 5. Пчёлы сем. Megachilidae представлены четырьмя биотопическими группами: луговой, лугово-степной, лугово-лесной, эвритопной. По видовому и численному обилию во всех орографических регионах преобладают луговые виды (69–72 и 56–73 % – видовое и численное обилие в разных регионах); по численному обилию выделяются также эвритопы (17–55 %). Виды мегахилид с широкой биотопической приуроченностью составляют 70 % видового обилия. 6. В Кузнецко-Салаирской провинции преобладают широко распространенные виды мегахилид – транспалеарктические полизональные (31 вид, 63,2 % – видовое и 64,3 % – численное обилие). 208 7. Основу населения пчёл сем. Megachilidae Кузнецко-Салаирской провинции составляют широкие полилекты (18 видов, 52,9 % – видовое и 61,3 % – численное обилие). Трофический спектр мегахилид включает 80 видов растений 58 родов 21 семейства. Наиболее привлекательными являются растения семейств Fabaceae, Asteraceae, Rosaceae. Для широких олиголектов основную составляют Fabaceae, кормовую базу Asteraceae и на исследуемой Lamiaceae, территории узких олиголектов – Campanulaceae (род Campanula). 8. На антропогенных территориях видовое богатство пчёл сем. Megachilidae снижается по сравнению с естественными и составляет 29 видов (59 % от общего числа видов в Кузнецко-Салаирской провинции). По степени устойчивости к факторам городской среды мегахилиды представлены 3 группами – урбофилы (9 видов), урботолеранты (18) и урбофобы (22). Численность урбофильных видов мегахилид в городской среде увеличивается в 1,7 раз по сравнению с природными биотопами. В состав урбофильной группы входят массовые (доминантные и субдоминантные) виды, характеризующиеся полилектичностью и широкой биотопической приуроченностью. 209 ПРАКТИЧЕСКИЕ РЕКОМЕНДАЦИИ 1. Рекомендуется включить в списки редких видов следующих мегахилид: Hoplitis tuberculata, Osmia leaiana, O. uncinata, Anthidium manicatum, A. punctatum, Bathanthidium sibiricum, Megachile argentata, M. centuncularis, M. circumcincta, M. rotundata, M. melanopyga. Сохранение этих видов тесным образом связано с охраной мест их обитания – остепнённых и суходольных разнотравно-злаковых лугов и степных участков Кузнецко-Салаирской провинции. 2. Мероприятия по сохранению видового разнообразия пчёл сем. Megachilidae в антропогенных местообитаниях наиболее целесообразно проводить в городских парках и садах, где в качестве охранных мероприятий рекомендуется регулирование рекреационной нагрузки. На рудеральных участках и деградированных лугах в черте городской и сельской застройки, а также в наиболее уязвимых биотопах – луговых степях Кузнецкой котловины – действенной мерой охраны пчёл может стать организация микрозаповедников. 3. Для привлечения пчёл сем. Megachilidae с целью увеличения видового разнообразия на антропогенно преобразованных и урбанизированных территориях рекомендуется высадка растений, принадлежащих к предпочитаемым кормовым семействам: Asteraceae, Fabaceae, Rosaceae и Campanulaceae (на клумбы, бордюры и т. д.). 4. Ряд показателей можно использовать в биоиндикационных целях. Например, индикаторными показателями усиления антропогенного воздействия на биоценозы является рост численности урбофильных видов пчёл сем. Megachilidae (Anthidium manicatum, A. septemspinosum, Chelostoma rapunculi, Osmia Coelioxys bicolor, inermis, Megachile Hoplitis ligniseca, tridentata, M. Lithurgus fuscipennis, versicolor) по сравнению с урботолерантными и урбофобными видами, а также отсутствие урбофобных видов, что свидетельствует антропогенного воздействия. о наличии даже небольшой степени 210 СПИСОК ЛИТЕРАТУРЫ Амолин, А.В. Гнездостроящие пчёлы трибы Anthidiini (Hymenoptera: Megachilidae) юго-востока Украины / А.В. Амолин // Проблеми екології та охорони природи техногенного регіону: міжвід. зб. наук. пр. – Донецьк: ДонНУ, 2008. – Вип. 8. – С. 96–103. Андреев, А.В. Оценка биоразнообразия, мониторинг и экосети / А.В. Андреев. – Кишинев: BIOTICA, 2002. – 168 с. Балахонова, В.А. Фаунистический обзор насекомых Южного Притоболья / В.А. Балахонова // Вестник Курганского ун-та. – Сер. «Естественные науки». – Курган: Изд-во Курганского гос. ун-та, 2009. – Вып. 2. – № 1 (15). – С. 17–20. Бейко, В.Б. Урботолерантные перепончатокрылые насекомые Москвы и перспективы их дальнейшего сохранения / В.Б. Бейко // Животные в городе Матер. науч.-практ. конф. – М.: ИПЭЭ РАН, 2000. – С. 68–69. Бейко, В.Б. Пчелиные (Apoidea) как индикаторы антропогенных воздействий в городских экосистемах / В.Б. Бейко, М.В. Березин // Экол. нормирование: проблемы и методы. Тез. науч.-координац. совещ. Пущино, 13–17 апр., 1992 г. – М., 1992. – С. 16–17. Бейко, В.Б. Перепончатокрылые насекомые в Красной книге Москвы / В.Б. Бейко, М.В. Березин // Животные в городе. Матер. науч.-практ. конф. – М.: ИПЭЭ РАН, 2000. – С. 69–71. Белова, К.А. Пчёлы (Hymenoptera: Apidae) Курумбельской степи / К.А. Белова // 50-я юбилейная Междунар. науч. студ. конф. «Студент и научно-технический прогресс». Новосибирск, 13–19 апр., 2012 г. Прогр. докл. – Новосибирск, 2012. – С. 25. Белова, К.А. Сравнительный анализ населения пчёл (Hymenoptera, Apidae) равнинных и горных степей Западной Сибири / К.А. Белова, Я.Р. Телегина // Матер. XVIII междунар. экол. студ. конф. «Экология России и сопредельных территорий. – Новосибирск, 2013. – С. 41. 211 Благовещенская, Н.Н. Изучение диких пчелиных в природе и лаборатории / Н.Н. Благовещенская // Уч. пос. к спецкурсу. – Ульяновск: Ульяновский пед. ин-т, 1983. – 79 с. Благовещенская, Н.Н. Ретроспективный анализ фауны пчелиных Среднего Поволжья / Н.Н. Благовещенская. – Ульяновск: Ульяновский гос. пед. ин-т, 1994. – 42 с. Богатырёв, Н.Р. Влияние антропогенной нагрузки на численность и видовой состав шмелей в парках Новосибирска / Н.Р. Богатырёв // Антропогенные воздействия на сообщества насекомых. – Новосибирск: Наука, 1985. – С. 128–134. Бубнова, Т.В. Методические рекомендации по сбору насекомых / Т.В. Бубнова. – Горно-Алтайск, 1988. – 67 с. Бывальцев, А.М. Население шмелей (Hymenoptera: Apidae, Bombini) Новосибирска и его окрестностей / А.М. Бывальцев // Сиб. экол. ж. – 2009. – № 3. – С. 395–404. Бывальцев, А.М. Сообщество шмелей (Hymenoptera: Apidae, Bombini) Новосибирска и его окрестностей / А.М. Бывальцев // Энтомол. исследования в Северной Азии. Матер. VIII межрегион. совещ. энтомологов Сибири и Дальнего Востока с участием заруб. учёных. Новосибирск, 4–7 окт., 2010 г. – Новосибирск: Товар-во науч. изд-й КМК, 2010. – С. 45–46. Вдовин, В.В. Салаирский кряж / В.В. Вдовин, А.М. Малолетко // История развития рельефа Сибири и Дальнего Востока. Алтае-Саянская область. – М.: Наука, 1969. – С. 141–156. Вдовин, В.В. Кузнецко-Салаирская провинция / В.В. Вдовин // Рельеф Алтае-Саянской горной страны. – Новосибирск: Наука, 1988. – С. 40–71. Воронцов, А.Г. Геоботаника. Уч. пос. для ун-тов и пед. ин-тов / А.Г. Воронцов. – М.: Высш. шк., 1973. – 384 с. Внуковский, В.В. Некоторые новые данные по энтомофауне Омского округа / В.В. Внуковский // Изв. Зап.-Сиб. геогр. об-ва. – Омск, 1930. – Т. 7. – С. 181–187. 212 Волошина, Т.А. Перспективы введения в культуру местных видов диких одиночных пчёл – опылителей люцерны / Т.А. Волошина // Тр. Зоол. ин-та АН СССР. – 1984. – Т. 128. – С. 87–93. Воскресенский, С.С. Геоморфология Сибири / С.С. Воскресенский. – М.: Изд-во МГУ, 1962. – С. 214–223. Гвоздецкий, Н.А. Физико-географическое районирование СССР. Характеристика региональных единиц / Н.А. Гвоздецкий. – М.: Изд-во МГУ, 1968. – 575 с. Гвоздецкий, Н.А. Физическая география СССР. Азиатская часть / Н.А. Гвоздецкий, Н.И. Михайлов. – М.: Мысль, 1978. – 512 с. Городков, К.Б. Типы ареалов насекомых тундры и лесных зон европейской части СССР / К.Б. Городков // Ареалы насекомых евр. части СССР. Карты 179–221. – Л.: Наука, 1984. – С. 3–20. Гришина, Е.М. Пчелиные (Hymenoptera, Apoidea) и необходимость их охраны / Е.М. Гришина // Пробл. охраны природы Западной Сибири. – Томск: Изд-во Томск. ун-та, 1980. – С. 93–97. Гуляева, А.Ф. Краткая характеристика растительного покрова Кузнецкой котловины / А.Ф. Гуляева, А.В. Климов // Вестник гос. пед. академии. – 2013. – № 1 (26). – URL: http://vestnik.kuzspa.ru/articles/139/ (дата обращения: 31.03.14). Давыдова, М.И. Физическая география СССР. Т. 1 / М.И. Давыдова, Э.М. Раковская, Г.К. Тушинский. – М.: Просвещение, 1989. – 240 с. Давыдова, Н.Г. Фауна пчёл (Hymenoptera, Apoidea) Якутии. I / Н.Г. Давыдова, Ю.А. Песенко // Энтомол. обозр. – 2002. – Т. 81, вып. 3. – С. 582–599. Давыдова, Н.Г. Фауна пчёл (Hymenoptera, Apoidea) Якутии: Автореф. дис. … канд. биол. наук / Давыдова Нина Григорьевна. – СПб, 2003. – 22 с. Давыдова, Н.Г. Фауна пчёл (Hymenoptera: Apoidea) Лено-Амгинского междуречья / Н.Г. Давыдова // II Симп. стран СНГ по перепончатокрылым 213 насекомым. 8-й Коллоквиум Рос. секции Междунар. союза исследователей общест. насекомых (IUSSI). СПб, 13–19 сент., 2010 г. Прогр. и тез. докл. – СПб: Зоол. ин-т РАН, 2010. – С. 46. Данилов, Ю.Н. Материалы по фауне осообразных (Hymenoptera, Vespiformes) и пчелиных (Hymenoptera, Apoidea) из окрестностей г. Барнаула (Алтайский край) / Ю.Н. Данилов // Энтомол. исследования в Северной Азии. Матер. VII Межрег. совещ. энтомологов Сибири и Дальнего Востока (в рамках Сиб. зоол. конф.). Новосибирск, 20–24 сент., 2006 г. – Новосибирск, 2006. – С. 55–56. Додин, А.Л. Геология и полезные ископаемые Кузнецкого Ала-Тау / А.Л. Додин. – М. – Л., 1948. – 286 с. Дубинкин, С.Ф. Алтае-Саянская область / С.Ф. Дубинкин // Геология СССР. Западная Сибирь. – М.: Недра, 1967. – Т. 14, ч. 1. – С. 585–594. Дунаев, Е.А. Методы эколого-энтомологических исследований / Е.М. Дунаев. – М.: МосгорСЮН, 1997. – 44 с. Еремеева, Н.И. Влияние факторов городской среды на структуру населения шмелей / Н.И. Еремеева //Сиб. экол. ж. – 2002. – № 4. – С. 441–448. Еремеева, Н.И. Фауна и биотопическое распределение шмелей в степном поясе Кузнецко-Салаирской горной области / Н.И. Еремеева, С.Л. Лузянин // Тр. Кемер. отд.-ния РЭО. – Кемерово: Компания Юнити, 2005. – Вып. 3. – С. 18–24. Еремеева, Н. И. Фауна шмелей (Hymenoptera, Apidae) урбоценозов г. Кемерово и его окрестностей / Н.И. Еремеева, С.Л. Лузянин // Живые объекты в условиях антропогенного пресса / Матер. X Междунар. науч.практ. экол. конф. Белгород, 15–18 сент., 2008 г. – Белгород, 2008 а. – С. 70–71. Еремеева, Н.И. Шмели (Hymenoptera, Apidae: Bombus Latreille и Psithyrus Lepeleiter) Кузнецко-Салаирской горной области: фауна, ландшафтнобиотопическое распределение и трофические связи / Н.И. Еремеева, С.Л. Лузянин // Тр. РЭО. – СПб. – 2008 б. – Т. 78, вып. 2. – С. 25–52. 214 Еремеева, Н.И. Материалы по фауне антофорид (Hymenoptera, Anthophoridae) Кузнецко-Салаирской горной области / Н.И. Еремеева, Д.А. Сидоров // Муравьи и защита леса. Матер. XII Всерос. Мирмекологического симп. и сателлитного совещ. «Экология и поведение общест. перепончатокрылых: теор. проблемы и практ. использование» в рамках Сиб. зоол. конф. Новосибирск, 7–14 августа, 2005 г. – Новосибирск: ООО «Талер-Пресс», 2005. – С. 36–39. Еремеева, Н.И. Материалы по фауне и экологии мегахилид (Hymenoptera, Megachilidae) Кузнецко-Салаирской горной области / Н.И. Еремеева, Д.А. Сидоров // Энтомол. исследования в Северной Азии: Матер. VII Межрегион. совещ. энтомологов Сибири и Дальнего Востока. Новосибирск, 20–24 сент., 2006 г. – Новосибирск: Талер-Пресс, 2006. – С. 222–224. Еремеева, Н.И. К фауне шмелей Горной Шории / Н.И. Еремеева, С.Л. Лузянин, Д.А. Сидоров // Тр. Кемер. отд.-ния РЭО. – Кемерово: Компания Юнити, 2003. – Вып. 2. – С. 17–23. Еремеева, Н.И. Структура населения и экологические особенности пчёл (Hymenoptera, Apoidea) горно-таёжных экосистем Кузнецкого Алатау / Н.И. Еремеева, С.Л. Лузянин, Д.А. Сидоров // Изв. Санкт-Петербургской лесотехн. академии. – СПб, 2010. – Вып. 192. – С. 85–92. Еремева, Н.И. Пчёлы (Hymenoptera, Apoidea) как компонент лесных экосистем Горной Шории / Н.И. Еремеева, С.Л. Лузянин, Д.А. Сидоров // Изв. Санкт-Петербургской лесотехн. академии. – 2009. – Вып. 187. – С. 115–123. Еремеева, Н.И. К изучению шмелей и шмелей-кукушек Кузнецкого Алатау / Н.И. Еремеева, Д.А. Сидоров, С.Л. Лузянин // Вестник Томск. гос. ун-та. – 2004. – № 11. – С. 22–25. Еремеева, Н.И. Коллетиды (Hymenoptera, Colletidae) Кузнецко-Салаирской горной области / Н.И. Еремеева, Д.А. Сидоров, С.Н. Яковлева // Тр. Кемер. отд.-ния Рус. энтомол. о-ва – Кемерово: Компания Юнити, 2006. – Вып. 4. – 215 Стр. 24–29. Еремеева, Н.И. Предварительные данные по фауне пчёл семейства Halictidae Кузнецко-Салаирской горной области / Н.И. Еремеева, С.Н. Яковлева, Д.А. Сидоров // Проблемы и перспективы общей энтомологии: тез. докл. XIII съезда Рус. энтомол. о-ва. Краснодар, 9–15 сент., 2007 г. – Краснодар: КубГАУ, 2007. – С. 106–107. Еремеева, Н.И. Урботолерантные насекомые: состав и особенности видов / Н.И. Еремеева, С.В. Блинова, С.Л. Лузянин // Изв. Самарского науч. центра РАН. – 2010. – Т. 12 (33), № 1 (8). – С. 1970–1972. Еремеева, Н.И. Редкие виды перепончатокрылых насекомых КузнецкоСалаирской горной области / Н.И. Еремеева, С.В. Блинова, С.Л. Лузянин, Д.А. Сидоров, С.Н. Яковлева // Изв. Самарского науч. центра РАН. – 2011. – Т. 13 (39), № 1 (5). – С. 1088–1091. Иванов, С.П. coerulescens Особенности (Hymenoptera: гнездования Apoidea: пчёл-опылителей Megachilidae) в Osmia искусственных гнездилищах / С.П. Иванов, А.В. Фатерыга // Экосистемы Крыма, их оптимизация и охрана (Тематич. сб. науч. тр.). – Симферополь: ТНУ, 2006. – Вып. 16. – С. 52–57. Иванов, С.П. Параллелизмы в развитии гнездостроительных инстинктов одиночных пчёл и ос (Hymenoptera: Megachilidae, Megachilinae; Vespidae, Eumenidae) / С.П. Иванов, А.В. Фатерыга // Исследования по перепончатокрылым насекомым. – М., 2007. – С. 205–217. Иванов, С.П. Материалы сравнительного изучения гнездования пчёллисторезов (Hymenoptera, Megachilidae Megachile Latr.). Сообщение I. Разнообразие гнездовых построек / С.П. Иванов // Уч. зап. Таврического нац. ун-та им. В.И. Вернадского. Сер. «Биология, химия». – 2010. – Т. 23 (62), № 3. – С. 68–78. 216 Иванов, С.П. Чеклист пчёл-мегахилид (Hymenoptera: Apoidea: Megachilidae) фауны Крыма / С.П. Иванов, М.А. Филатов, А.В. Фатерыга // Экосистемы Крыма, их оптимизация и охрана (Тематич. сб. науч. тр.). – Симферополь: ТНУ, 2007. – Вып. 17. – С. 3–12. Игнатенко, Е.В. Фауна пчёл (Hymenoptera, Apoidea) Амурской области // Е.В. Игнатенко, М.Ю. Прощалыкин // Евразиат. энтомол. ж. – 2005. – № 4 (3). – С. 243–250. Ильиных, Н.И. Почвы Кузнецкого Алатау / Н.И. Ильиных. – Красноярск, 1970. – 165 с. Ильичёв, А.И., Кузбасс: Ресурсы. Экономика. Рынок / А.И. Ильичёв, М.П. Виткин, Н.В. Калишев. – Кемерово: Кемер. полиграфкомбинат, 1995. – 288 с. Ильичёв, А.И. География Кемеровской области: природные условия и ресурсы: Уч. пос. / А.И. Ильичёв, Л.В. Соловьёв. – Кемерово: Кемер. книжн. изд-во, 1994. – 366 с. Ковалёва, С.Р. Лесные почвы горного окаймления юго-востока Западной Сибири (Восточный Алтай, Горная Шория, Салаир) / С.Р. Ковалёва, В.М. Корсунов, С.А. Таранов. – Новосибирск: Наука, 1974. – С. 75–195. Кокуев, Н.Р. Hymenoptera, собранные В.В. Совинским на берегах озера Байкал в 1902 году / Н.Р. Кокуев // Тр. комиссии по изучению озера Байкал. – 1927. – Т. 2. – С. 63–76. Колобков, М.Н. Кузнецкий бассейн (очерки природы и хозяйства) / М.Н. Колобков. – Кемерово: Кемер. книжн. изд-во, 1956. – С. 13–31. Конусова, О.Л. Пчёлы (Hymenoptera, Apoidea) Томской области и их трофические связи / О.Л. Конусова // Пробл. сохранения биол. разнообразия Юж. Сиб.: 1 Межрегион. науч.-практ. конф. Кемерово, 21–22 мая, 1997 г. – Кемерово, 1997. – С. 81–82. 217 Конусова, О. Л. Пчелиные семейства Andrenidae (Hymenoptera) Южной тайги Западной Сибири / О.Л. Конусова // Пробл. энтомологии в России. Сб. науч. тр. XI Съезда РЭО. СПб, 23–26 сент., 1997 г. – СПб.: ЗИН РАН, 1998. – Т. 1. – С. 207–208. Конусова, О.Л. Сообщества пчелиных (Hymenoptera, Apoidea) в ландшафтах юга Томской области / О.Л. Конусова // Экология и рац. природопользование на рубеже веков. Итоги и перспективы. Матер. междунар. конф. – Томск, 2000 а. – Т. 1. – С. 123. Конусова, О.Л. Структура населения шмелей Томского Приобья и тенденции её изменения за последние 30 лет / О.Л. Конусова // Экология пойм сибирских рек и Арктики. Тр. II совещ. 22–26 нояб. 2000 г. – Томск: «STT», 2000 б. – С. 100–103. Конусова, О.Л. К фауне и экологии шмелей (Hymenoptera, Apidae) Томской области / О.Л. Конусова // XII Съезд РЭО. СПб, 19–24 авг., 2002 г. Тез. докл. – СПб, 2002. – С. 177. Конусова, О.Л. Экологическая характеристика фауны пчёл (Hymenoptera, Apoidea) южной тайги Томского Приобья / О.Л. Конусова, В.В. Янюшкин // Сиб. экол. ж. – Томск, 2000. – № 3. – С. 283–286. Конусова, О.Л. К фауне и экологии пчёл семейства Andrenidae (Hymenoptera, Apoidea) юго-востока Томской области / О.Л. Конусова, Л.В. Гришаев // Энтомол. исследования в Северной Азии. Матер. VII Межрегион. совещ. энтомологов Сибири и Дальнего Востока (в рамках Сиб. зоол. конф.). Новосибирск, 20–24 сент., 2006 г. – Новосибирск, 2006. – С. 239–241. Конусова, О.Л. Материалы по фауне и экологии пчёл-коллетид (Hymenoptera, Colletidae) юго-востока Западной Сибири / О.Л. Конусова, Д.А. Сидоров, Н.И. Еремеева // Вестник Томск. гос. ун-та. – 2007. – № 301. – С. 212–218. Королюк, А.Ю. Луговые степи Алтае-Саянской горной области. Общая характеристика / А. Ю. Королюк, Н.И. Макунина // KRYLOVIA. Сиб. ботан. 218 ж. – 2000.–– Т. 2, № 1. – С. 26–-37. Красная книга города Москвы / под ред. Самойлова Б.Л., Морозовой Г.В. – М.: Департамент природопользования и охраны окружающей среды г. Москвы, 2011. – 930 с. Красная книга Курганской области. Изд-е второе. – Курган: Изд-во Курган. гос. ун-та, 2012. – С. 158–158. Красноборов, И.М. Определитель растений Кемеровской области / И.М. Красноборов, Э.Д. Крапивкина, М.Н. Ломоносова, Г.П. Будникова, Д.Н. Шауло, П.А. Волобаев, А.Т. Мальцева, Г.И. Яковлева, А.А. Красников, Е.И. Вибе, К.С. Байков, Н.Н. Тупицына, Д.А. Герман, Н.Ф. Вылцан, И.И. Гуреева. – Новосибирск: Изд-во СО РАН, 2001. – 477 с. Крылов, Г.В. Леса Западной Сибири / Г.В. Крылов. – М.: Изд-во АН СССР, 1961. – 255 с. Кузьмин, П.М. Фауна пчелиных семейств Megachilidae, Anthophoridae Курганской области / П.М. Кузьмин, А.Е. Молчанов // Фауна беспозвоночных Урала. – Челябинск, 1983. – С. 19–25. Кузьмин, П.М. Проблема охраны колоний пчелиных в Зауралье / П.М. Кузьмин, А.Е. Молчанов // Флора и фауна Курганской области, их охрана, экология и изучение в биол. дисциплинах. – Курган, 1987. – С. 122–123. Куминова, А.В. Растительность Кемеровской области / А.В. Куминова. – Новосибирск, 1950. – 167 с. Курмакаев, Е.В. Динамика распределения пчелиных (Hymenoptera, Apoidea) в зависимости от урбанизированности территории / Е.В. Курмакаев // Биология – наука XXI века: сб. тез. 7-й Пущинской школы – конф. молодых учёных. Пущино, 14–18 апр., 2003 г. – Пущино, 2003 – С.184. Лаврентьев, А.И. К вопросу о строении и развитии долин Кузнецкого нагорья / А.И. Лаврентьев // Геология и геофизика. – 1967. – № 11. – С. 65–73. Лавров, С.Д. Материалы к изучению энтомофауны окрестностей Омска / 219 С.Д. Лавров // Тр. Сиб. ин-та сел. хоз-ва и лесоводства (Сиб. с.-х. академии). – Омск, 1927. – Т. 8, вып. 3. – С. 95–96. Лащинский, Н.Н. Растительность Салаирского кряжа / Н.Н. Лащинский. – Новосибирск: Академ. изд-во «Гео», 2009. – 263 с. Лащинский, Н.Н. Флора Салаирского кряжа / Н.Н. Лащинский, Н.В. Седельникова, Т.А. Сафонова, О.Ю. Писаренко, Н.В. Перова, И.А. Горбунова, Н.В. Лащинская. – Новосибирск: Академ. изд-во «Гео», 2007. – 252 с. Лащинский, Н.Н. Природные условия Салаирского кряжа / Н.Н. Лащинский, М.П. Ревякина // Экология сообществ черневых лесов Салаира. – Новосибирск: Наука, 1991. – С. 6–23. Лебедева, Н.В. География и мониторинг биоразнообразия / Н.В. Лебедева, Д.А. Криволуцкий, Ю.Г. Пузаченко, К.Н. Дьяконов, Г.М. Алещенко, А.В. Смуров, В.Н. Максимов, В.С. Тикунов, Г.Н. Огуреева, Т.В. Котова. – М.: Изд-во Науч. и уч.-метод. центра, 2002. – 432 с. Левченко, Т.В. К фауне пчёл семейства Apidae Московской области / Т.В. Левченко // Симп. стран СНГ по перепончатокрылым насекомым. Прогр. и тез. докл. – М.: МГУ им. М.В. Ломоносова. 2006 а. – С. 54. Левченко, Т.В. К фауне пчёл семейства Megachilidae Московской области / Т.В. Левченко // Симп. стран СНГ по перепончатокрылым насекомым. Прогр. и тез. докл. – М.: МГУ им. М.В. Ломоносова, 2006 б. – С. 55. Левченко, Т.В. К фауне пчёл (Hymenoptera, Apoidea) Московской области: предварительный список видов семейства Megachilidae / Т.В. Левченко // Исследования по перепончатокрылым насекомым. – М.: Товарищество науч. изданий КМК, 2007 а. – С. 153–161. Левченко, Т.В. К фауне пчёл семейства Andrenidae (Hymenoptera, Apoidea) Московской области / Т.В. Левченко // Тез. докл. XIII съезда РЭО. – Краснодар: РЭО, КубГАУ, 2007 б. – С. 82–83. Левченко, Т.В. О фауне пчёл семейства Halictidae (Hymenoptera, Apoidea) Московской области / Т.В. Левченко // Тр. Ставропольского отд.-ния РЭО. 220 Вып. 4: Матер. Междунар. науч.-практ. конф. – Ставрополь: Аргус, 2008. – С. 104–105. Левченко, Т.В. Стациальное распределение пчёл (Hymenoptera: Apoidea) Московской студентов, области / Т.В. Левченко // аспирантов и молодых Тез. докл. XVI междунар. ученых «Ломоносов 2009»; конф. секция «Биология». – М.: МАКС Пресс, 2009. – С. 127–128. Левченко, Т.В. Зоогеографическая характеристика фауны пчёл (Hymenoptera, Apoidea) Московской области / Т.В. Левченко // Бюлл. МОИП. – 2009 б. – Т. 114, вып. 1. – С. 14–21. Левченко, Т.В. Материалы по фауне пчёл (Hymenoptera: Apoidea) Московской области. 1. Семейства Melittidae и Colletidae / Т.В. Левченко // Эверсманния. Энтомол. исследования в России и соседних регионах. – 2009 в. – Вып. 17–18. – С. 27–43. Левченко Т.В. К фауне пчёл (Hymenoptera: Apoidea) ландшафтного заказника «Крылатские холмы» (Москва) / Т.В. Левченко // Вестник Мордовского ун-та, 2009. – № 1. – С. 39–40. Левченко, Т.В. Фауна и экология пчёл (Hymenoptera: Apoidea) Московской области: Автореф. дис. … канд. биол. наук / Левченко Тимофей Викторович. – М., 2010. – 23 с. Левченко Т.В. К изучению лектических связей пчёл (Hymenoptera: Apoidea) Московской области / Т.В. Левченко // Тр. Ставропольского отд.ния Рус. энтомол. о-ва. Вып. 6: Матер. III Междунар. интернет-конф. – Ставропольский гос. аграрный ун-т. – Ставрополь: АГРУС, 2010 б. – С. 67–69. Ломакин, Д.И. К истории энтомологических исследований на территории региона до 1916 г. / Д.И. Ломакин, А.В. Толстиков //Aus Sibirien-2006: науч.информ. сб. – Тюмень, 2006. – С. 63–65. Лузянин, С.Л. Трофический спектр шмелей горных территорий Кемеровской области / С.Л. Лузянин // Тез. докл. XIII Междунар. конф. студентов, аспирантов и молодых ученых «Ломоносов – 2006». М., 12–15 221 апр., 2006 г. – М., 2006 а. – С. 143–144. Лузянин, С.Л. Эколого-фаунистическая характеристика шмелей Горной Шории / С.Л. Лузянин // Студент и научно-технический прогресс: Матер. XLIV Междунар. науч. студ. конф. – Новосибирск, 2006 б. – С. 41. Лузянин, С.Л. Эколого-фаунистическая характеристика шмелей (Hymenoptera, Apidae) восточного склона Салаирского кряжа / С.Л. Лузянин // Тр. Кемер. отд.-ния РЭО. – Кемерово: Компания Юнити, 2006 в. – Вып. 4. – С. 78–81. Лыков, В.А. Сравнительная характеристика фауны пчелиных г. Перми и пригорода / В.А. Лыков // Насекомые в естественных и антропогенных биогеоценозах Урала. Матер. IV Совещ. энтомологов Урала. Пермь, 24–26 марта, 1992 г. – Екатеринбург: Наука. Урал. отд.-ние, 1992. – С. 89–91. Лыков, В.А. Фауна и некоторые вопросы экологии пчелиных (Hymenoptera, Apoidea) в городе Перми / В.А. Лыков // Фауна и экология насекомых Урала. – Пермь: Пермский гос. ун-т. – 1993. – С. 81–100. Лыков, В.А. Пчелиные в условиях г. Перми / В.А. Лыков // Экол. и охрана окр. среды: Тез. докл. 2 Междунар. науч.-практ. конф. Пермь, 12–15 сент., 1995 г. – Пермь, 1995. – Ч. 4. – С. 34–35. Лыков, В.А. Обзор фауны пчелиных (Hymenoptera, Apoidea) Пермского края / В.А. Лыков // Вестник Перм. ун-та. – 2007. – Вып. 5. – С. 61–68 Лыков, В.А. Пчёлы (Hymenoptera, Apoidea) островной Кунгурской лесостепи / В.А. Лыков // Вестник Перм. ун-та. – Пермь, 2008. – Вып 9 (25). – С. 32–36. Малолетко, А.М. Палеогеография предалтайской части Западной Сибири в мезозое и кайнозое / А.М. Малолетко. – Томск: Изд-во Томcк. ун-та, 1972. – 227 с. Мариковская, Т.П. К биологии пчелиных (Apoidea) юго-востока Казахстана / Т.П. Мариковская // Тр. Всесоюзного энтомол. о-ва. – 1972. – Т. 55. – С. 187–216. 222 Мариковская, Т.П. перепончатокрылых Сохранение (Hymenoptera) в разнообразия антропогенных жалящих биотопах Юго- Восточного Казахстана с помощью искусственных приманочных гнездилищ / Т.П. Мариковская, П.А. Есенбекова, В.Л. Казенас // Структура і функціональна роль тваринного населення в природних та трансформованих екосистемах: Тези I міжнародної конференції Дніпропетровськ, 17–20 вересня, 2001 г. – Дніпропетровськ: ДНУ, 2001. – С. 80–82. Матысюк, Н.Т. О каменистых степях Кузнецкой котловины / Н.Т. Матысюк // Природа и экономика Кузбасса. – Новокузнецк, 1980. – С. 99–102. Михайлов, Н.И. Горы Южной Сибири / Н.И. Михайлов. – М.: Географгиз, 1961. – С. 135–141. Молчанов, А.Е. Насекомые / А.Е. Молчанов // Сберечь обязаны: Редкие, исчезающие животные и растения, памятники природы и заказники Курганской области. Справочник. – Челябинск, 1989. – С. 104–135. Молчанов, А.Е. Материалы к типизации и эволюции гнездовий пчелиных (Apoidea) / А.Е. Молчанов, П.М. Кузьмин // Фауна беспозвоночных Урала. – Челябинск: Гос. пед. ин-т, 1985. – Вып. 6. – С. 60–67. Монсявичюс, В.С. Пчелиные (Hymenoptera, Apoidea) Дайнавской песчаной равнины. 1. Фауна и стациальное распределение / В.С. Монсявичюс // Acta Entomologica Lituanica. – Вильнюс, 1988. – Vol. 9. – С. 95–110. Монсявичюс, В.С. Трофические связи пчелиных Дайнавской песчаной равнины / В.С. Монсявичюс // Тр. Академии наук Литовской ССР. Сер. В. – 1989. – Т. 1, № 105. – С. 61–74. Мытарев, А.А. Экономико-географические особенности и природа Кузбасса / А.А. Мытарев // Природа Кузбасса. – Новокузнецк, 1973. – С. 5–19. Никифорук, К.С. Пчёлы Башкирской АССР / К.С. Никифорук // Зап. Башкир. филиал Геогр. о-ва СССР. – 1957. – Вып. 1. – С. 139–162. 223 Осичнюк, А.З. Фауна Украіни. Бджолині. Бджоли – колетиди / Г. З. Осичнюк. – Київ: Наукова думка, 1970. – Т.12, вип. 4. – 156 с. Осичнюк А.З. Фауна Украіни. Бджолині. Бджоли – андреніди / Г. З. Осичнюк. – Київ: Наукова думка, 1977. – Т.12, вип. 5. – С. 5–327. Осычнюк А.З. Надсем. Apoidea /А.З. Осычнюк, Д.В. Панфилов, А.А. Пономарёва // Определитель насекомых европейской части СССР. Т. 3. Перепончатокрылые. Ч. 1. – Л.: Наука, 1978. – С. 279–519. Павлова, Г.Г. Суходольные луга Средней Сибири / Г.Г. Павлова. – Новосибирск: Наука, 1980. – 216 с. Пантова, А.Ю. Разнообразие диких пчёл (Hymenoptera, Apoidae) в условиях г. Киев / А. Ю. Пантова // Біорізноманіття та роль тварин в екосистемах: Матер. VІІ Міжнар. наукової конф. – Дніпропетровськ: Адверта, 2013. – С. 151–153. Панфилов, Д.В. Общий обзор населения пчелиных Евразии / Д.В. Панфилов // Сб. тр. Зоол. музея МГУ. – 1968. – Т. XI. – С. 18–35. Пашина, Л.С. Опылители семенной люцерны в Восточно-Казахстанской области / Л.С. Пашина // Тр. Ин-та зоол. АН КазССР, 1955. – Т. 4. – С. 226–235. Пашина, Л.С. Новое в экологии пчёл мегахил / Л.С. Пашина // Проблемы экологии. – Томск: Изд-во Томск. ун-та, 1967. – Т. 1. – С. 116–121. Песенко, Ю.А. Материалы по фауне и экологии пчелиных (Hymenoptera, Apoidea) Нижнего Дона. Сообщ. I. Сем. Megachilidae / Ю.А. Песенко // Энтомол. обозр. – 1971. – Т. 50, № 1. – С. 60–78. Песенко, Ю.А. Материалы по фауне и экологии пчелиных (Hymenoptera, Apoidea) Нижнего Дона. Сообщ. IV. Обзор трофических связей / Ю.А. Песенко // Энтомол. обозр. – 1975. – Т. 54, № 3. – С. 555–564. Песенко, Ю.А. О формировании населения одиночных пчелиных (Apoidea) на полях посевной люцерны / Ю.А. Песенко // Зоол. ж. – 1976. – Т. 55, вып. 6. – С. 856–859. 224 Песенко, Ю.А. Люцерновая пчела-листорез Megachile rotundata и её разведение для опыления люцерны (обзор и библиография) / Ю.А. Песенко. – Л.: Наука. – 1982 а. – 136 с. Песенко, Ю.А. Принципы и методы количественного анализа в фаунистических исследованиях / Ю.А. Песенко. – М.: Наука, 1982 б. – 287 с. Петров, Б.Ф. Почвы Алтайско-Саянской области / Б.Ф. Петров // Тр. Почв. ин-та им. В.В. Докучаева. – 1952. – Т. 35. – 245 с. Пономарева, А.А. Пчелиные – опылители бобовых растений Западного Копет-Дага: автореф. дис. … канд. биол. наук / Пономарева Ася Анатольевна. – Л., 1960. – 20 с. Пономарева, А.А. О пчелиных (Hymenoptera, Apoidea) – опылителях некоторых степных растений Центрального Казахстана / А.А. Пономарева // Энтомол. обозр. – 1962. – Т. 41, № 1. – С. 76–82. Пономарева, А.А. О кормовых связях некоторых пчёл подсемейства Anthophorinae и основных опылителей (Hymenoptera, Apoidea) бобовых растений в Центральном Казахстане и Средней Азии / А.А. Пономарева // Полезные насекомые – опылители и энтомофаги: Тр. Зоол. ин-та АН СССР. – 1967. – Т. 38. – С.65–83. Попов, В.В. Палеарктические формы трибы Stelidini Roberts. (Hymenoptera, Megachilidae) / В.В. Попов // Тр. Зоол ин-та АН СССР. – 1932. – Т. I. – С. 375–414. Попов, В.В. Семейство Fideliidae и морфологическая конвергенция среди пчелиных / В.В. Попов // Доклады АН СССР. – 1939. – Т. 22, № 9. – С. 645–647. Попов, В.В. Перепончатокрылые – Hymenoptera / В.В. Попов // Животный мир СССР. II. Пустыни. – М. – Л.: Изд-во АН СССР, 1948. – С. 195–222. Попов, В.В. Сбор и изучение опылителей сельскохозяйственных культур и других растений / В.В. Попов // В помощь работающим на полезащитных лесных полосах. – 1950. – Ч. 2. – 36 с. 225 Попов, В.В. О значении пчелиных (Hymenoptera, Apoidea) в опылении люцерны / В.В. Попов // Тр. Всесоюзного энтомол. о-ва. – 1951. – Т. 43. – С. 65–82. Попов, В.В. О географическом распространении и эволюции пчелиных подрода Erythrosmia Schmied. (Hymenoptera, Megachilidae) / В.В. Попов // Зоол. ж. – 1954 б. – Т. 33, вып. 2. – С. 443–451. Попов, В.В. Пчелиные, их связь с цветковой растительностью и вопрос об опылении люцерны / В.В. Попов // Энтомол. обозр. – 1956. – Т. 35, № 3. – С. 582–598. Попова, Л.М. Стациальное распределение пчелиных (Hymenoptera, Apoidea) Среднего Поволжья / Л.М. Попова // Эколого-генетические исследования устойчивости и продуктивности популяций. Межвуз. сб. – Нижний Новгород, 1990. – С. 29–41. Прокопьев, А.С. К изучению репродуктивной биологии и видового состава опылителей очитков (Sedum L.) на юге Томской области / А.С. Прокопьев, О.Л. Конусова, Т.Н. Беляева // Вестник Томск. гос. ун-та. – 2004. – № 11. – С. 124–126. Проскурякова, А.А. Пчёлы рода Andrena Fabricius, 1775 (Hymenoptera, Andrenidae) Среднего Прииртышья /А.А. Проскурякова // Прогр. докл. 50-й юбилейной Междунар. науч. студ. конф. «Студент и научно-технический прогресс». Новосибирск, 13–19 апр., 2012 г. – Новосибирск, 2012. – С. 25. Прощалыкин, М.Ю. Фауна пчёл (Hymenoptera, Apoidea) Среднего и Нижнего Приамурья / М.Ю. Прощалыкин // Евразиат. энтомол. ж. – 2003. – № 2 (1). – С. 25–29. Прощалыкин, М.Ю. Фауна пчёл юга Дальнего Востока России / М.Ю. Прощалыкин. // Чтения памяти А. И. Куренцова. – Владивосток, 2005. – Вып. XVI. – С. 5–38. Прощалыкин, М.Ю. Фауна пчёл (Hymenoptera, Apoidea) Дальнего Востока России / М.Ю. Прощалыкин // Симп. стран СНГ по перепончатокрылым насекомым. – М., 2006 а. – С. 72. 226 Прощалыкин, Дальнего М.Ю. Востока Пчёлы-клептопаразиты России / М.Ю. (Hymenoptera, Прощалыкин // Apoidea) Матер. VII Межрегионального совещания энтомологов Сибири и Дальнего Востока в рамках Сиб. зоол. конф. – Новосибирск, 2006 б. – С. 273–274. Прощалыкин, М.Ю. Пчёлы рода Coelioxys Latreille, 1809 (Hymenoptera, Apoidea, Megachilidae) фауны Дальнего Востока России / М.Ю. Прощалыкин // Евразиат. энтомол. ж. – 2006 в. – № 5 (4). – С. 318–324. Прощалыкин, М.Ю. Пчёлы (Hymenoptera, Apoidea) острова Монерон / М.Ю. Прощалыкин // Растительный и животный мир острова Монерон (Матер. Междунар. сахалинского проекта). – Владивосток, 2006 г. – С. 250–254. Прощалыкин, М.Ю. Фауна пчёл (Hymenoptera, Apoidea) Забайкалья и Дальнего Востока России / М.Ю. Прощалыкин // Экология в современном мире: взгляд научной молодежи. – Улан-Удэ, 2007 а. – С. 88–89. Прощалыкин, М.Ю. Фауна пчёл (Hymenoptera, Apoidea) Дальнего Востока России / М.Ю. Прощалыкин // Исследования по перепончатокрылым насекомым. Сб. науч. работ. – М.: Товарищество науч. изданий КМК, 2007 б. – С. 162–172. Прощалыкин, М.Ю. Сем. Megachilidae – Мегахилиды / М.Ю. Прощалыкин // Определитель насекомых Дальнего Востока России. Том 4. – Ч. 5. – Владивосток, 2007 в. – С. 889–897. Прощалыкин, М.Ю. Фауна пчёл (Hymenoptera, Apoidea) Еврейской автономной области / М.Ю. Прощалыкин // Чтения памяти А.И. Куренцова. – Вып. XVIII. – Владивосток, 2007. – С. 88–93. Прощалыкин, М.Ю. Ареалогический анализ фауны пчёл (Hymenoptera, Apoidea) Дальнего Востока России / М.Ю. Прощалыкин // Проблемы и перспективы общей энтомологии. Тез. докл. XIII съезда Рус. энтомол. о-ва. – Краснодар, 9–15 сент., 2007 г. – Краснодар, 2007 д. – С. 299. 227 Прощалыкин, М.Ю. Пчёлы-эндемики (Hymenoptera, Apoidea) фауны Дальнего Востока России / М.Ю. Прощалыкин // Чтения памяти А.И. Куренцова. – Вып. XIX. – Владивосток, 2008. – С. 42–51. Прощалыкин, М.Ю. Длиннохоботковые пчёлы (Hymenoptera, Apoidea: Megachilidae, Apidae) Забайкалья / М.Ю. Прощалыкин // Вiсник Харькiвського нац. унiверситету iм. В.Н. Каразiна. Сер.: Бiологiя. – 2009 а. – Вип. 9. – № 856. – С. 136–142. Прощалыкин, М.Ю. История изучения пчёл (Hymenoptera, Apoidea) Забайкалья / М.Ю. Прощалыкин. // Чтения памяти А.И. Куренцова. – Вып. XX. – Владивосток, 2009 б.– С. 69–78. Прощалыкин, М.Ю. Ареалогический анализ фауны пчёл (Hymenoptera, Apoidea) Забайкалья / М.Ю. Прощалыкин // Энтомол. исследования в Северной Азии. – Новосибирск, 2010. – С. 177–178. Прощалыкин, М.Ю. Особенности распространения пчёл (Hymenoptera: Apiformes) на Дальнем Востоке России / М.Ю. Прощалыкин // Определитель Насекомых Дальнего Востока России. Дополнительный том. Анализ фауны и общий указатель названий. – Владивосток: Дальнаука, 2011. – С. 101–115. Прощалыкин, М.Ю. Секция Apiformes – пчёлы / М.Ю. Прощалыкин // Аннотированный каталог насекомых Дальнего Востока России. Т.1. – Перепончатокрылые. – Владивосток: Дальнаука, 2012. – С. 461–466. Прощалыкин, М.Ю. Новые находки пчёл (Hymenoptera, Apoidea, Apiformes) в Сибири / М.Ю. Прощалыкин // Чтения памяти А.И. Куренцова. – Вып. XXIV. – Владивосток, 2013 а.– С. 135–148. Прощалыкин, М.Ю. Пчёлы трибы Anthidiini Ashmead, 1899 (Hymenoptera: Apoidea: Megachilidae) Сибири и Дальнего Востока России / М.Ю. Прощалыкин // Кавказский энтомол. бюлл. – 2013 б. – № 9 (1). – С. 147–158. 228 Прощалыкин, М.Ю. К познанию пчёл (Hymenoptera, Apoidea, Apiformes) Тувы / М.Ю. Прощалыкин // Биоразнообразие Алтае-Саянского экорегиона: изучение и сохранение в системе ООПТ. Матер. межрегион. науч.-практ. конф., посвящённой 20-летию основания заповедника «Убсунурская котловина». Кызыл, 27 июня – 1 июля 2013 г. – Кызыл, 2013 в. – С. 150–151. Прощалыкин, М.Ю. Фауна пчёл (Hymenoptera, Apoidea) острова Сахалин / М.Ю. Прощалыкин, А.С. Лелей, А.Н. Купянская // Растительный и животный мир острова Сахалин (Матер. Междунар. сахалинского проекта). – Владивосток: Дальнаука, 2004. – Ч. 1. – С. 154–192. Радченко, В.Г. Биология пчелиных юго-восточной Украины: Автореф. дис. … канд. биол. наук / Радченко Владимир Григорьевич. – Киев, 1982. – 25 с. Радченко, В.Г. Биология пчёл (Hymenoptera, Apoidea) / B.Г. Радченко, Ю.А. Песенко. – СПб.: ЗИН РАН СССР. – 1994. – 350 с. Радченко, В.Г. «Краснокнижные» виды пчёл семейства Мегахилиды (Hymenoptera, Megachilidae) на карте Крыма / В. Г. Радченко, С.П. Иванов, М.А. Филатов, А.В. Фатерыга // Экосистемы Крыма, их оптимизация и охрана (Тематич. сб. науч. тр.). – Симферополь: ТНУ, 2009. – Вып. 20. – С. 165–179. Рахимов, И.И. Классификация ландшафтов по степени нарастания антропогенной трансформации на примере Среднего Поволжья / И.И. Рахимов, Р.А. Уленгов // Филология и культура. – Казань: Казанский (Приволжский) федеральный ун-т, 2004. – № 2. – С. 166–172. Розенберг, Г.С. Несколько слов об индексе разнообразия Симпсона / Г.С. Розенберг // Самарская Лука. – 2007. – Т. 16, № 3 (21). – С. 581–584. Романькова, Т.Г. Пчелиные рода Megachile Latr. (Hymenoptera, Apoidea, Megachilidae) фауны Сибири и Дальнего Востока СССР / Т.Г. Романькова // Систематика и эколого-фаунистический обзор отдельных насекомых Дальнего Востока. – Владивосток, 1983. – С. 141–147. отрядов 229 Романькова, Т.Г. Пчелиные рода Osmia Panz. (Hymenoptera, Megachilidae) фауны Дальнего Востока России / Т.Г. Романькова // Энтомол. обозр. – 1984. – Т. 63, вып. 2. – С. 141–147. Романькова, Т.Г. Использование приманочных гнёзд для изучения жалящих перепончатокрылых (Hymenoptera, Aculeata) в Приморском крае / Романькова Т.Г., Романьков А. В. // Перепончатокрылые Восточной Сибири и Дальнего Востока. – Владивосток, 1986. – С.130–137. Романькова, Т.Г. Бджолинi родини Megachilidae Забайкалья / Т.Г. Романькова // 4-й зiзд Украiнського ентомол. товариства. – Харькiв, 1992. – С. 145–146. Романькова, Т.Г. Новые данные по фауне пчелиных Сибири и Дальнего Востока (Hymenoptera, Apoidea, Megachilidae) / Т.Г. Романькова // Перепончатокрылые Сибири и Дальнего Востока. Сб. науч. тр. заповедника «Даурский». – Киев, 1994. – Вып. 3. – С. 119–128. Романькова, Т.Г. Megachilidae – Мегахилиды / Т.Г. Романькова // Определитель насекомых Дальнего Востока России. Т. 4. Сетчатокрылые, скорпионницы, перепончатокрылые. Ч.1. – СПб: Наука, 1995. – С. 530–547. Романькова, Т.Г. Трофические связи пчелиных семейства Megachilidae в условиях Приморского края / Романькова Т.Г., Романьков А. В. // Изв. Харьковского энтомол. о-ва. – 1993. – № 1. – С.76–81. Ромасенко, Л.П. Предкуколки некоторых видов пчёл-мегахилид (Hynenoptera, Megachilidae) / Л. П. Ромасенко // Вестник зоологии. – 1989. – № 1. – С. 75. Ромасенко, Л.П. Гнездостроящие мегахилиды (Hynenoptera, Apoidea, Megachilidae) Украины и их экологические особенности: Автореф. дис. … кандидата биол. наук / Ромасенко Людмила Петровна. – Киев: Ин-т зоол. АН УССР, 1984. – 23 с. Ромасенко, Л. П. Пчёлы-мегахилиды (Apoidea, Megachilidae) Украинского государственного Степного заповедника (Хомутовская степь, Каменные могилы) / Л. П. Ромасенко. – Киев: Ин-т зоол. АН УССР. – 1990. – 64 с. 230 Ронгинская, А.В. Динамические процессы в луговых фитоценозах (на примере лугов Салаирского кряжа) / А.В. Ронгинская. – Новосибирск: Наука, 1988. – 160 с. Рутковская, Н. В. Климат / Н. В. Рутковская, К. И. Попова // Физикогеографическое описание и экономическая характеристика. Геология СССР. Западная Сибирь. – М.: Недра, 1967. – Т. 14, ч. 1. – С. 42–44. Сапаев, Е.А. К фауне пчелиных нижней части города Казани (Приволжский и Бауманский район) / Е.А. Сапаев // Защита растений и охрана природы в Татарской АССР. – Казань: Татарское книжн. изд-во, 1089. – С. 82–84. Сапаев, Е.А. Сравнение видового состава диких пчелиных агроценозов Татарии и города Казани / Е.А. Сапаев // Матер. экол. науч. конф. студентов и выпускников биолого-почвенного факультета Казанского университета, посвященной 80-летию со дня рождения проф. В.А. Попова. – Казань: Полиграфкомбинат им. К. Якубаю – 1993. – С. 25–30. Сарычев, М.В. Трофические связи представителей пчёл-мегахилид из рода трахуза с бобовыми северо-востока Приобской лесостепи / М.В. Сарычев // Науч.-техн. бюлл. Сиб. науч.-исслед. ин-та кормов. – Новосибирск, 1988. – № 1. – С. 44–50. Сарычева, З.Н. Флороспециализация антидии флорентийской Anthidium florentinum F. (Apoidea, Hymenoptera, Insecta) в условиях Новосибирской области / З.Н. Сарычева, Н.Д. Тарасенко // Ж. общ. биологии. – 1989. – Т. 50, № 5. – С. 655–662. Седельников, В.П. Флора и растительность высокогорий Кузнецкого Алатау / В.П. Седельников. – Новосибирск: Наука, 1979. – 168 с. Сергеев, М.Г. Закономерности распространения прямокрылых насекомых Северной Азии / М.Г. Сергеев. – Новосибирск: Наука, 1986. – 228 с. Сидоров, Д.А. Материалы по фауне рода Megachile Latr. (Hymenoptera, Megachilidae) Кузнецко-Салаирской горной области и сопредельных территорий / Д.А. Сидоров // Сб. тр. студентов и молодых ученых Кемер. гос. ун-та, посвященный 60-летию победы в Великой Отечественной войне. – 231 Кемерово: Полиграф, 2005. – Т. 3, вып. 6. – С. 335–337. Сидоров, Д.А. Видовой состав и биотопическое распределение пчёл рода Andrena F. (Hymenoptera, Andrenidae) в Кузнецко-Салаирской горной области и сопредельных территориях / Д.А. Сидоров // Образование, наука, инновации – вклад молодых исследователей: матер. I (XXXIII) Междунар. науч.-практ. конф. – Кемерово: Полиграф, 2006. – Т. 3, вып. 7. – С. 302–304. Сидоров, Д.А. Материалы по экологии Andrena dentata Smith, 1879 (Hymenoptera, Andrenidae) в Кузнецко-Салаирской горной области / Д.А. Сидоров // Экология Южной Сибири и сопредельных территорий. Вып. 12. – Абакан: Изд-во Хакасского госун-та, 2008. – Т. 1. – С. 113. Сидоров, Д.А. Фауна пчёл отдельных родов семейства Apidae КузнецкоСалаирской горной области / Д.А. Сидоров, Н.И. Еремеева // Тр. Кемер. отд.ния Рус. энтомол. о-ва. – Кемерово: Компания Юнити, 2008. – Вып. 6. – С. 110–116. Сидоров, Д.А. Находки восточно-палеарктических пчёл-андрен (Hymenoptera, Apoidea, Andrenidae) в Кузнецко-Салаирской горной области / Д.А. Сидоров, Н.И. Еремеева // Евразиат. энтомол. ж. – 2010. – № 9 (1). – С. 87–89. Сидоров Д.А., Яковлева С.Н. Фауна и экология пчёл семейства Anthophoridae (Hymenoptera, Anthophoridae) степной зоны Кузнецко- Салаирской горной области // Образование, наука, инновации – вклад молодых исследователей: матер. I (XXXIIII) Междунар. науч.-практ. конф. студентов, аспирантов и молодых учёных. – Кемерово: Полиграф, 2006. – Т. 3, вып. 7. – С. 301–302. Ситдиков, А.А. К фауне диких пчелиных Удмуртии / А.А. Ситдиков. Фауна и экология животных УАССР и прилежащих районов. – Ижевск: Изд-во Удмурт. гос. ун-та, 1981. – С. 118–125. Ситдиков, А.А. Фауна пчёл (Hymenoptera, Apoidea) Удмуртии, с описанием Melitta udmurtica sp. n. / А.А. Ситдиков // Тр. Зоол. ин-та АН СССР. – 1986. – Т. 156. – С. 103–112. 232 Соловьёв, Л.И. Беседы по краеведению Кузбасса: учебное пособие / Л.И. Соловьёв. – Кемерово: КРИПКиПРО, 2010. – 391с. Тахтаджян, А.П. Жизнь растений. Т. 5 (2). Цветковые растения / А.П. Тахтаджян. – М.: Просвещение, 1980. – 576 с. Трофимов, С.С. Экология почв и почвенные ресурсы Кемеровской области / С.С. Трофимов. – Новосибирск: Наука, 1975. – 300 с. Уткин, Н.А. Насекомые / Н.А. Уткин // Что имеем, как храним. – Курган: Курганск. обл. комитет экологии и природных ресурсов, 1993. – С. 126– 135. Файнер, Ю.Б. Кузнецкая котловина / Ю.Б. Файнер // История развития рельефа Сибири и Дальнего Востока. – М.: Наука, 1969. – С. 146–156. Фасулати, К.К. Полевое изучение наземных беспозвоночных / К.К. Фасулати. – М.: Высшая школа, 1971 г. – 424 с. Фомичёв, В.Д. Геоморфологический очерк / В.Д. Фомичёв // Геология СССР. Кузнецкий бассейн. – М. – Л.: Гос. изд-во геолог. лит-ры, 1940. – Т. 16. – С. 35–40. Хлонов, Ю.П. Деревья и кустарники юго-восточной части Западной Сибири / Ю.П. Хлонов. – Новосибирск: Наука, 1979. – С. 7–22. Ченикалова, Е.В. Пчелиные (Hymenoptera, Apoidea) Центрального Прекавказья, их эколого-биоценотическое значение в агроландшафтах: Автореф. дис. … докт. биол. наук / Ченикалова Елена Владимировна. – СПб – Пушкин, 2005. – С. 15–25. Чернышев, В.Б. Охрана природы и роль полевых обочин и лесополос в агроландшафте / В.Б. Чернышев, А.В. Суязов, В.М. Афонина // Видовые популяции и сообщества в антропогенно трансформированных ландшафтах: состояние и методы его диагностики. Матер. XI Междунар. науч.-практ. экол. конф. Белгород, 20–25 сент., 2010 г. – Белгород, 2010. – С. 13–14. Шварёв, Ю.О. Климат / Ю.О. Шварёв // Западная Сибирь. – М.: Изд-во АН СССР, 1963. – С. 70–99. 233 Шелоховская, Л.В. Фауна и экология пчёл (Hymenoptera, Apoidea) тундровой зоны бассейна нижнего течения р. Индигирки (Северо-Восточная Якутия): Автореф. дис. … канд. биол. наук / Шелоховская Лариса Васильевна. – Якутск., 2009. – 20 с. Шелоховская, Л.В. Материалы по фауне и экологии пчёл (Hymenoptera, Apoidea) тундровой зоны Якутии /Л.В. Шелоховская, Н.Г. Давыдова, М.В. Березин // Вестник СВФУ, 2011. – Т. 8, № 4. – с. 45–50. Шеметов, Г.А. Природные комплексы Горной Шории / Г.А. Шеметов, О.С. Андреева // Шорский сб. Вып. II. Этноэкология и туризм Горной Шории. – Кемерово, 1997. – Вып. 2. – С. 111–133. Шереметова, С.А. Проблемы сохранения редких растений на территориях с высокой антропогенной нагрузкой / С.А. Шереметова // Развитие современной экологии. – 2012. – URL: http://ecotext.ru/34.html (дата обращения: 01.04.14). Шитиков, В.К. Оценка биоразнообразия: попытка формального обобщения / В.К. Шитиков, Г.С. Розенберг // Структурный анализ экологических систем. Количественные методы экологии и гидробиологии (Сб. науч. тр., посвященный памяти А.И. Баканова). – Тольятти: СамНЦ РАН, 2005. – С. 91–129. Шульгин, В.Н. Снежный покров в Горной Шории / В.Н. Шульгин // Шорский сб. Вып. II. Этноэкология и туризм Горной Шории. – Кемерово, 1997. – Вып. 2. – С. 37–50. Шумакова, П.С. Пчелиные (Hymenoptera, Apoidea) – опылители бобовых трав в Кулунде / П.С. Шумакова, З.С. Бабенко, Г.С. Золотаренко // Полезные и вредные насекомые Сибири. – Новосибирск: Наука, 1982. – С. 157–174. Шумилова, Л.В. Ботаническая география Сибири / Л.В. Шумилова. – Томск: Изд-во Томск. ун-та, 1962. – 439 с. Яковлева, С.Н. Материалы по фауне и экологии галиктид КузнецкоСалаирской горной области / С.Н. Яковлева // Образование, наука, инновации – вклад молодых исследователей: матер. II (XXXIV) Междунар. науч.-практ. 234 конф. студентов, аспирантов и молодых учёных. – Кемерово: Полиграф, 2006. – Т. 1, вып. 8. – С. 384–385. Яковлева, С.Н. Предварительные данные по фауне пчёл семейства Megachilidae (Hymenoptera, Apoidea) Кузнецко-Салаирской горной области / С.Н. Яковлева // Тр. Ставропольского отд.-ния Рус. энтомол. о-ва. Вып. 7: Матер. IV Междунар. интернет-конф. «Актуальные вопросы энтомологии». Ставрополь, 20 марта, 2011 г. – Ставрополь: Параграф, 2011. – С. 98–101. Яковлева, С.Н. Состав и структура фауны мегахилид (Hymenoptera, Apoidea, Megachilidae) Кузнецкой котловины / С.Н. Яковлева // Вестник Кемер. гос. ун-та. – Кемерово, 2012. – Выпуск 2 (50). – С. 10–14. Яковлева, С.Н. Материалы по фауне пчёл сем. Megachilidae горных территорий Кузнецко-Салаирской провинции / С.Н. Яковлева // Биоразнообразие, проблемы экологии Горного Алтая и сопредельных территорий: настоящее, прошлое, будущее. Матер. III Междунар. конф. Горно-Алтайск, 1–5 окт., 2013 г. – Горно-Алтайск: РИО ГАГУ, 2013. – С. 123–126. Яковлева, С.Н. Трофические связи пчёл сем. Megachilidae (Hymenoptera, Apoidea) Кузнецко-Салаирской провинции / С.Н. Яковлева // Современные проблемы науки и образование. – 2014. – № 3. – URL: http://www.scienceeducation.ru/117-13506 (дата обращения: 11.06.2014). Ban, C.M. Contributions to the knowledge of Apoid Hymenopterans (Hymenoptera: Megachilidae, Anthophoridae, Apidae) from Maramureş (Romania). Part I / C.M. Ban // Travaux du Museum National d’Histoire Naturelle «Grigore Antipa». – 2005. – Vol. XLVIII. – P. 289–301. Banaszak, J. Megachilid Bees of Europe (Hymenoptera, Apoidea, Megachilidae) / J. Banaszak, L. Romasenko. – Bydgoszcz: Pedagogical Univ. of Bydgoszcz. – 1998. – 241 pp. Banaszak, J. Klucze do oznaczania owadów Polski. Część XXIV, Błonkówki – Hymenoptera. Zeszyt 68 f, Pszczołowate – Apidae. Podrodzina: Megachilinae / J. Banaszak, L. Romasenko, T. Cierzniak // Polskie Towarzystwo Entomologiczne. 235 – Toruń, 2001. – N. 161. – 53 pp. Bohart, G.E. How to manage the alfalfa leaf-cutting bee (Megachile rotundata F.) for alfalfa pollination / G.E. Bohart // Utah Agric. Exp. Sta. Circ. – l962. – Vol. 144. – P. 1–7. Bohart, G.E. Managing the leaf-cutting bee for alfalfa seed yields / G.E. Bohart // Utah State Univ. Extention, Serv. Leafleat. – 1964. – Vol. 104. – P. 1–12. Bohart, G.E. Commercial production and management of wild bees – a new entomological industry / G.E. Bohart // Bull. Soc. Entomol. Amer. – 1970. – Vol. 16, N 1. – P. 8–9. Bosch, J. Analisis de los nidos de algunos Megachilidae nidificantes en cavidades preestablecidas (Hymenoptera, Apoidea) / J. Bosch, N. Vicens, M. Blas // Orsis. – 1993. – N 8. – P. 53–63. Cane, J.H. Substrates and materials used for nesting by North American Osmia bees (Hymenoptera: Apiformes: Megachilidae) / J.H. Cane, T. Griswold, F.D. Parker // Annals of the Entomol. Soc. of America. – 2007. – N 100. – P. 350– 358. Cockerell, T.D.A. Bees collected in Siberia in 1927 / T.D.A. Cockerell // Ann. Mag. nat. Hist. – 1928. – Ser. 10, Vol. 1. – P. 345–361. Delaplane, K.S. Crop pollination by bees / K.S. Delaplane, D.F. Mayer. – Cambridge, MA, USA: CABI Publishing. – 2000. – 50 pp. Eickwort, G.C. Two European species of Chelostoma established in New York State / G.C. Eickwort // Psyche – 1980. – Vol. 87. – P. 315–324. Erlandsson, S. Die schwedischen Arten der Gattung Coelioxys Latr. (Hym., Apidae) / S. Erlandsson // Opusc. Ent. – 1955. – Suppl. XX: 2 – 3. – P. 174–191. Hausl-Hofstatter, U. Bemerkenswerte Megachile-Arten aus der Steiermark (Hymenoptera, Apoidea, Megachilidae) / U. Hausl-Hofstatter // Joannea Zool. – 2007. 9: 15–20. Gogala, A. Contribution to the Knowledge of the Bee Fauna of Slovenia (Hymenoptera: Apidae) / A. Gogala // Scopolia. – 1991. – N. 25. – P. 1–33. Gogala, A. Contribution to the Knowledge of the Bee Fauna of Slovenia II (Hymenoptera: Apidae) / A. Gogala // Scopolia. – 1994. – N. 31. – P. 1–40. 236 Gogala, A. Bee Fauna of Slovenia: Checklist of species (Hymenoptera: Apidae) / A. Gogala // Scopolia. – 1999. – N. 42. – P. 1–79. Griswold, T.L. The Classification of the Osmiini of the Eastern Hemisphere (Hymenoptera, Megachilidae) / T.L. Griswold, C. D. Michener // J. of the Kansas Ent. Soc. – 1997. – V. 70, N. 3. – P. 207–253. Guler, Y. Faunistic Study on Megachilini, Osmiini snd Anthidiini Tribes (Hymenoptera: Megachilidae) in Central Anatolia / Y. Guler, N. Çağatay // J. Ent. Rec. Soc. – 2006. – Vol. 8, N 2. – P. 15–34. Kemp, W.P. Development and Emergence of the Alfalfa Pollinator Megachile rotundata (Hymenoptera: Megachilidae) / W.P. Kemp, J. Bosch // Annals of the Ent. Soc. of America. – 2000. – Vol. 93, N 4. – P. 904–911. Kupianskaya, A.N. Contribution to the bee fauna (Hymenoptera: Apoidea: Megachilidae) of Shantar Islands / A.N. Kupianskaya, M.Yu. Proshchalykin, A.S. Lelej // Far Eastern Entomologist. – 2011. – N. 229. – P. 1–6. Ladurner, E. Laboratory Rearing of Osmia cornuta Latreille (Hymenoptera, Megachilidae) on Artificial Diet / E. Ladurner, B. Maccagnani, D. Tesoriero, M. Nepi, A. Felicioli // Boll. Ist. Ent. «G. Grandi» Univ. Bologna. – 1999. Vol. 53. – P. 133–146. Ladurner, E. Osmia cornuta (Hymenoptera, Megachilidae) densities required for apple pollination: / E. Ladurner, L. Recla, M. Wolf, R. Zelger, G. Felicioli // J. of Apicultural Research. – 2004. – Vol. 43, N 3. – P. 118–122. Levchenko, T.V. Contribution to the bee fauna (Hymenoptera: Apiformes) of the Khanty-Mansi Autonomous Region, Western Siberia, Russia / T.V. Levchenko, K.P. Tomkovich // Entomofauna. – 2014. – Bd. 35, H. 5. – S. 85–100. Maccagnani, B. Osmia cornuta (Hymenoptera, Megachilidae) as a pollinator of pear (Pyrus communis): fruit- and seed-set / B. Maccagnani, E. Ladurner, F. Santi, G. Burgio // Apidologie. – 2003. – Vol. 34. – P. 207–216. 237 Matache, I. Family Megachilidae (Hymenoptera: Apoidea) in Dobrogea (Romania) / I. Matache, C. M. Ban // Travaux du Museum National d’Histoire Naturelle «Grigore Antipa». – 2006. – Vol. XLIX. – P. 297–306. Michener, C.D. The Generic Classification of the Anthidiine Bees (Hymenoptera: Megachilidae) / C.D. Michener // Amer. Museum Novitates. – 1948. –N 1381. – P. 1–29. Michener, C.D. The Classification of the Lithurginae (Hymenoptera: Megachilidae) / C.D. Michener // Pan-Pacific entomologist. – 1983. – Vol. 59, N 1. – 4. – P. 176–187. Michener, C.D. The bees of the world / C.D. Michener. – Baltimore; London: J. Hopkins Univ. Press. – 2000. – 913 pp. Michener, C.D. The bees of the world. 2nd edition / C.D. Michener. – Baltimore; London: J. Hopkins Univ. Press. – 2007. – 953 pp. Monsevičius, V.A Check-list of Wild Bee Species (Hymenoptera, Apoidea) of Lithuania with Data to their Distribution and Bionomics / V.A. Monsevićius // New and Rare for Lithuania Insect Species. Records and Descriotions of 1994– 1995. – Vilnius: Institute of Ecology, 1995. – 145 pp. Monsevicius, V.A. First supplement to the fauna list of Lithuanian wild bees (Hymenoptera, Apoidea) / V.A. Monsevićius // Acta Zool. Lituanica, 2004. – Vol. 14., N 3. – P. 3–9. Monzon, V.H. Foraging behavior and pollinating effectiveness of Osmia cornuta (Hymenoptera: Megachilidae) and Apis mellifera (Hymenoptera: Apidae) on «Comice» pear / V.H. Monzon, J. Bosch, J. Retana // Apidologie. – 2004. – Vol. 35. – P. 575–585. Muller, A. Morphological specializations in Central European bees for the uptake of pollen from flowers with anthers hidden in narrow corolla tubes (Hymenoptera: Apoidea) / A. Muller // Entomologia Generalis. – 1995. – N 20. – P. 43–57. Muller, A. Host-plant specialization in western palearctic anthidiine bees (Hymenoptera: Apoidea: Megachilidae) / A. Muller // Ecological Monographs. – 238 1996. – N 66. – P. 235–257. Muller, A. Unusual host plant of Hoplitis pici, a bee with hooked bristles on its mouthparts (Hymenoptera: Megachilidae: Osmiini) / A. Muller // European J. of Entomology. – 2006. – N 103. – P. 497–500. Noskiewicz, J. Beitrage zur Kenntnis der palaarktischen Arten der Gattung Stelis Panz. (Hym., Apidae) / J. Noskiewicz // Pol. Pismo Ent. – 1961. – N 32. – P. 54–68. Ozbek H. A preliminary review of the Megachilidae of Turkey. Part IV. Megachilini and Lithurgini (Hymenoptera: Apoidea) / H. Ozbek, G. Zanden // Turk. entomol. derg. – 1994. – Vol. 18, N. 3. – P. 157–174. Pachinger, B. Uber das Vorkommen der Steinbienen Lithurgus Latr. (Hymenoptera: Apoidea, Megachilidae) in Osterreich - Okologie, Verbreitung und Gefahrdung / B. Pachinger // Linzer biol. Beitr. – 2004. – Bd. 36, H. 1 – S. 559–566. Praz, C.J. Phylogeny and biogeography of bees of the tribe Osmiini (Hymenoptera: Megachilidae) / C.J. Praz, A. Muller, B.N. Danforth, T.L Griswold, A. Widmer, S. Dorn // Molecular Phylogenetics and Evolution. – 2008. – Vol. 49. – P. 185–197. Přidal, A. Commentary to the subgenera of the genus Coelioxys described by Ruszkowski 1986 (Hymenoptera: Apoidea: Megachilidae) / A. Přidal, B.Tkalců // Entomofauna. – 2001. – Bd. 22, H. 14. – S. 357–364. Proshchalykin, M.Yu. The bees (Hymenoptera, Apoidea) of the Kuril Islands / M.Yu. Proshchalykin // Far Eastern Entomologist. – 2003. – N. 132. – P. 1–21. Proshchalykin, M.Yu. A check list of the bees (Hymenoptera, Apoidea) of the southern part of the Russian Far East / M.Yu. Proshchalykin // Far Eastern Entomologist. – 2004. – N. 143. – P. 1–17. Proshchalykin, M.Yu. The bees of family Megachilidae (Hymenoptera, Apoidea) of Transbaikalia / M.Yu. Proshchalykin // Far Eastern Entomologist. – 2007. – N. 175. – P. 1–18. 239 Proshchalykin, M.Yu. Contribution to the fauna of bees (Hymenoptera, Apoidea) of the Northern part of the Russian Far East / M.Yu. Proshchalykin // Far Eastern Entomologist. – 2009. – N. 194. – P. 6–7. Proshchalykin, M.Yu. Bees of the subgenus Allocoelioxys Tkalců of the genus Coelioxys Latreille (Hymenoptera: Apoidea: Megachilidae) from the Russian Far East / M.Yu. Proshchalykin, A.S. Lelej // Zootaxa. – 2004. – N. 517. – P. 1–6. Proshchalykin, M.Yu. The bees (Hymenoptera, Apoidea) of the Northern part of the Russian Far East / M Yu. Proshchalykin, A.N. Kupiaskaya // Far Eastern Entomologist. – 2005. – N. 153. – P. 1–39. Robertson, C. Flowers and Insects / C. Robertson. – Carlinville (Illinois): Sci. Press, 1928. – 221 p. Romankova, T.G. Additional data of the bee fauna (Hymenoptera, Apoidea: Megachilidae, Apidae) of Siberia and the Russian Far East / T.G. Romankova // Far Eastern Entomologist. – 2003. – N 129. – P. 1–6. Shebl, M.A. The impact of using leafcutting bees (Megachilidae, Hymenoptera) with different fertilization treatments on Alfalfa seed production / M.A. Shebl, S.M. Kamel, T.A. Abu Hashesh, M.A. Osman // Rev. Cienc. Suelo Nutr. / J. Soil. Sci. Plant Nutr. – 2009. – Vol. 9, N 2. – P. 134–141. Stoeckhert, F.K. Die Bienen Frankens / F.K. Stoeckhert // der Deutschen Entomologischen Zeitschrift. – 1933. – 294 ss. Strange, J.P. Global invasion by Anthidium manicatum (Linnaeus) (Hymenoptera: Megachilidae): assessing potential distribution in North America and beyond / J.P. Strange, J.B. Koch, V.H. Gonzalez, L. Nemelka, T. Griswold // Biol. Invasions. – 2011. – V. 13, N. 9. – P. 2115–2133. Ungricht, S. A taxonomic catalogue of the Palaearctic bees of the tribe Osmiini (Hymenoptera: Apoidea: Megachilidae) / S. Ungricht , A. Muller, S. Dorn // Zootaxa. – 2008. – Vol. 1865. – P. 1–253. 240 Warncke, K. Die Bienengattung Osmia Panzer 1986, ihre Systematik in der Westpalaarktis und ihre Verbreitung in der Turkey. 10. Die Untergattung Alcidamea Cress. / K. Warncke // Linzer biol. Beitr. – 1991. – Bd. 23, H. 2. – S. 701–751. Warncke, K. Die westpalaarktischen Arten der Bienengattung Coelioxys Latr. (Hymenoptera, Apidae, Megachilidae) / K. Warncke // Ber. naturf. Ges. – 1992 а. – Bd. 53. – S. 31–77. Warncke, K. Die westpalaarktischen Arten der Bienengattung Stelis Panzer, 1806 (Hymenoptera, Apidae, Megachilidae) / K. Warncke // Entomofauna. – 1992 b. – Bd. 13, H. 22. – S. 341. Whitehead, V.B. African Fideliini: Genus Fidelia Friese (Hymenoptera: Apoidea: Megachilidae: Fideliinae) / V.B. Whitehead, C. D. Eardley // J. of the Cansas Ent. Soc. – 2003. – Vol. 76, N 2. – P. 250–276. Wnukowskij, W. Zur Fauna der Dipteren und Hymenopteren des Bezirks Kuznezk (sudwestliches Sibirien, fruheres Gouvernement Tomsk) / W. Wnukowskij // Mitt. munch. ent. Ges., e. V. – 1926. – Bd. 16, H. 8–12. – S. 92–94. Wnukowskij, W. Verzeichnis der Hymenopteren des Bezirkes Kamenj (sudwestliches Sibirien, fruheres Gouvernement Tomsk) / W. Wnukowsky // Konowia. – 1927. – Bd. 6, H. 1. – S. 31–34. Wnukowsky, W. Einige faunistische Angaben uber die Insecten Sibiriens und des Ussuri-Gebietes / W. Wnukowsky // Zool. Anzeiger. – 1929. – Bd. 83, H. 9/10. – S. 212–220. Wnukowsky, W. Beitrage zur Insekten-Fauna des Bezirkes von Tomsk / W. Wnukowsky // Konowia. – 1936. – Bd. 25, H. 1–2. – S. 113–128. Zanden, G. Notes of some palearctic species of the genus Lithurge Latreille (Hymenoptera, Apoidea, Megachilidae) / G. Zanden // Bull. Rech. Agron. Gembloux. – 1977. – Vol. 12, N 4. – P. 357–362. 241 Zanden, G. Die palaarctischen Arten der Gattung Lithurge Latreille, 1825 (Hymenoptera, Apoidea, Megachilidae) / G. Zanden // Mitt. Zool. Mus. Berl. – 1986. – Bd. 62, No 1. – S. 53–59. Zanden, G. Beitrag zur Systematik und Nomenklatur der palaarktischen Osmiini, mit angaben uber ihre Verbreitung / G. Zanden // Zool. Meded. – 1988. – Vol. 62, no 9. – P. 113–133 Zanden, G. Zur Synonymie palaarctischer Arten der Familie Megachilidae (Insecta, Hymenoptera; Apoidea) / G. Zanden // Linzer biol. Beitr. – 1995. – Bd. 27, H. 1. – S. 427–434. Zanden, G. Notes on the distribution of Palaearctic Hymenoptera Aculeata / G. Zanden // Ann. Ent. Fenn. – 1997. – V. 43, N 1. – P. 39–40.