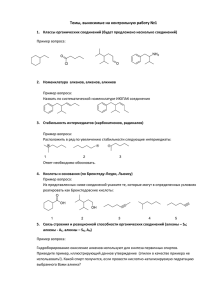
АБИОГЕНЕЗ В АТМОСФЕРЕ: АРХАИЧНЫЙ МЕТАБОЛИЗМ
advertisement

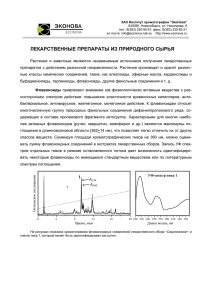
110 ЗАРИЦКИЙ А. Р., ВОРОНЦОВ Ю. П., ПРОНИН В. С. ТЕОРИЯ ЭВОЛЮЦИИ АБИОГЕНЕЗ В АТМОСФЕРЕ: АРХАИЧНЫЙ МЕТАБОЛИЗМ, КОМБИНАТОРНАЯ РЕПЛИКАЦИЯ И ПЕРВЫЕ ВИРУСЫ Зарицкий А. Р., 2Воронцов Ю. П., 3Пронин В. С. 1 Физический институт им. П.Н.Лебедева РАН, http://www.lebedev.ru 119991 Москва, Россия 2 ДГК Больница №13 им. Н.Ф.Филатова Мосгорздрава, http://www.13dgkb.ru 103001 Москва, Россия 3 Первый МГМУ им И.М.Сеченова Минздравсоцразвития РФ, http://www.mma.ru 119991 Москва, Россия Поступила в редакцию 19.03.2014 Представлена чл.-корр. РАЕН В.И. Грачевым 1 Работа посвящена анализу процессов развития нарождающегося живого мира на этапе, непосредственно следующем за «началом начал». Данный период характеризуется дальнейшей интенсификацией выделений свободной энергии локальными источниками в атмосфере, усложнением на этой базе синтезируемых органических соединений, метаболических процессов и архаичных предклеточных систем, что обеспечило формирование предпосылок для появления первых ростков жизни. Появление липидов стало тем системообразующим фактором, который обеспечил реализацию указанных предпосылок в форме архаичных механизмов матричного воспроизводства органических соединений и простейших вирусов. Ключевые слова: абиогенез, метаболизм, комбинаторика, репликация, липиды, вирусы. УДК 576.1+551.7 Содержание 1. Введение (110) 2. Появление разветвленных цепочек реакций метаболизма (111) 3. Предпосылки для появления вирусов (117) 4. Заключение (120) Литература (121) 1. ВВЕДЕНИЕ1 Проблемы абиогенеза – этапа добиологической эволюции Земли – всегда были и остаются предметом неослабевающего интереса и острого столкновения прямо противоположных точек зрения, построений и моделей, имеющих серьёзные основания по причине всеобщей и полной неадекватности понимания (по утверждению известных эволюционистов) как физических условий, так и химических процессов на первичной Земле. В рамках восприятия мира как естественной среды существования реальности, не противоречащей известным законам физики и химии, правдоподобные и логически непротиворечивые гипотезы абиогенеза с возможно меньшим количеством произвольных допущений имеют право на существование в 1 Раздел, за исключением последнего абзаца, написан В.И. Грачевым (ИРЭ им В.А. Котельникова РАН) 1 НОМЕР | ТОМ 6 | 2014 | РЭНСИТ качестве взаимодополнительных концепций. Рассмотрение гипотетических процессов эволюции во временных и пространственных масштабах, лежащих далеко за пределами человеческого опыта, неизбежно принимающих вероятностную форму, с необходимостью отмечает высокую роль случая на разных уровнях эволюционных процессов, не отрицая их глобальных закономерностей. Абиогенез как начало начал происхождения жизни рассматривается в ряду соответствующих гипотез, среди которых общепризнаны такие построения, сочетающие моделирование, эксперимент и натурные наблюдения, как наиболее раннее – гипотеза Опарина-Холдейна [1, 2] коацерватных капель из противоположно заряженных полимеров в первичном бульоне, с восстановительной атмосферой и анаэробными гетеротрофами, частично поддержанная экспериментами C.Миллера-Г.Юри [3] и С.Фокса [4]. Ее ключевые положения – концентрирующая органику ячеистость среды, роль обмена веществ и источников энергии, определяющие первичный органический синтез – остались исходными для последующих подобных гипотез. Г. Вахтершаузер с ТЕОРИЯ ЭВОЛЮЦИИ коллегами [5, 6] предполагает возникновение жизни в зонах вулканической активности на катализаторах – железе, никеле, кобальте и других переходных металлах, фиксировавших СО2 для растущей органики, подкрепив свои построения синтезом пептидов в подобных условиях. М. Рассел с коллегами [7, 8, 9] развивают идею возникновения жизни в теплых щелочных источниках на дне первичного океана – непрерывно-проточных реакторах в насыпях карбонатов, кремния, глины, железоникелевых сульфидов, вскоре открытых в области Срединно-Атлантического хребта. А. Мулкиджанян с коллегами [10, 11] представили инкубаторами самых ранних форм жизни конденсаты испарений наземных геотермальных источников (малые глубины, низкие температуры, катализатор – сульфид цинка, УФ излучение), с соотношением ионного состава (Ka, Na, Mg, Zn, фосфаты), весьма напоминающим современную внутриклеточную среду, не менявшемся, как считают, на протяжении всей истории жизни на Земле. Наконец, в нашей работе [12] абиогенные процессы выведены в газовую фазу – в приземные слои турбулентной, горячей, безкислородной и насыщенной водяными парами, парами неорганических катализаторов и органикой атмосферы, существование которой в первые сотни миллионов лет над расплавленной и постепенно остывающей магмой на поверхности планеты достаточно правдоподобно. Основной акцент сделан на энергетическом аспекте абиогенных процессов с использованием возможных физических факторов. А именно на учете потенциалов на поверхности аморфных сгустков органики, конденсирующейся при остывании окружающей среды – нанокаплях углеводородного аэрозоля, вероятность образования которых в плотных пылевых слоях архаичной атмосферы очевидно достаточно велика. Появление такого относительно устойчивого, неравновесного энерго-сырьевого образования, автономного, напрямую не зависимого от Солнца, тепла Земли и различных излучений, становилось источником свободной энергии за счет анаэробного распада в нем депонированных углеводородов для синтеза органики в окружающей среде. Углеводородная капля с сильным градиентным электрическим полем над ее поверхностью АБИОГЕНЕЗ В АТМОСФЕРЕ... порядка сотен нм может рассматриваться как первичная ячейка примитивного открытого реактора пребиотической химии, в котором синтезировались мономеры – аминокислоты, нуклеотиды, сахара и жирные кислоты. Архаичные механизмы катализа (ускорения реакций) в условиях переизбытка времени только складывались в этой среде, насыщенной парами неорганических катализаторов. При достижении локальных высоких концентраций неорганических полифосфатов становился возможным синтез первых короткоживущих макроэргов, а также полимеров различной длины со случайной последовательностью, в том числе медленно реплицирующихся (лат. replicatio – развертывание; зд.: удвоение, копирование) полинуклеотидов, в частности, фрагментов рибонуклеиновой кислоты (РНК). Спонтанное специфическое спаривание комплементарных (лат. complementum – дополнение) нуклеотидов формировало элементы архаичных механизмов матричного копирования и саморепликации – основы переноса информации. Целью настоящей работы является анализ наиболее вероятного, по нашему мнению, пути эволюционного развития преджизни – архаичных метаболических систем, сформировавшихся непосредственно в газовой среде и на поверхности углеводородных капель, завершившегося появлением первых архаичных вирусов. На этом этапе элементы указанных систем постепенно приобретали черты характерные как для клеточного мира, так и вирусов. 2. ПОЯВЛЕНИЕ РАЗВЕТВЛЕННЫХ ЦЕПОЧЕК РЕАКЦИЙ МЕТАБОЛИЗМА В работе [12] была указана генеральная линия развития живого мира (в том числе предвирусного и предклеточного), которой он следует и по настоящее время. Это – неуклонная интенсификация выделений и утилизации энергии (энергооборота) в его метаболических системах и энергетический выигрыш (более полное использование выделяющейся химической энергии) при совершенствовании живых систем, что позволяют им направлять избыток энергии на создание новых, полезных для их выживания, структур. Следование живого мира этой генеральной линии обеспечивает РЭНСИТ | 2014 | ТОМ 6 | НОМЕР 1 111 112 ЗАРИЦКИЙ А. Р., ВОРОНЦОВ Ю. П., ПРОНИН В. С. его системам удивительную выживаемость при смене условий существования, проявляясь как биологический консерватизм. В свете изложенных в предыдущем абзаце положений причины появления в метаболических системах, ответственных за выделение энергии, длинных и зачастую разветвленных цепочек реакций ясны. В первую очередь это обеспечивает энергетический выигрыш, поскольку позволяет полнее использовать запасенную в исходном сложном веществе химическую энергию. В одном акте (отдельной реакции) превращения исходного энергоемкого субстрата в конечный продукт может выделиться единовременно энергии, в несколько раз превышающее то, что может запасти макроэрг. В этом случае оставшаяся и, возможно, основная часть освободившейся химической энергии перейдет в тепло. Последовательные превращения первичного субстрата могут обеспечить наиболее экономичный путь утилизации энергоемких соединений. Однако, необходимым условием такой экономии является требование, чтобы при каждом шаге превращений выделяемая энергия была достаточна для запаса в конкретном макроэрге, но существенно его не превышала. При этом цепочки реакций могут быть разветвленными, поскольку каждый промежуточный продукт может претерпевать дальнейшие превращения несколькими способами. Иногда выгодно при таких превращениях даже тратить часть макроэргов. Заметим, что группы первых макроэргов как раз и относились к соединениям – участникам таких превращений. При этом основным требованием для действия соединения как макроэрга является обратимость реакций их распада. Огромное многообразие органического материала, в том числе соединений, способных функционировать в таких цепочках, сложившееся к тому времени в атмосфере, являлось предпосылкой для появления сложных путей утилизации энергоемких субстратов. Однако, для реализации этой предпосылки необходимы соответствующие (высокие) концентрации реагентов (катализаторов реакций, макроэргов и субстратов). Системообразующими факторами для реализации указанной предпосылки стали, во-первых, дальнейшее остывание планеты 1 НОМЕР | ТОМ 6 | 2014 | РЭНСИТ ТЕОРИЯ ЭВОЛЮЦИИ и, во-вторых, появление широкого видового спектра архаичных макроэргов. Действие первого фактора привело к усиленной конденсации углеводородов в более крупные капли. Их плотность вблизи поверхности планеты существенно возросла и, соответственно, основная часть органики («преджизнь») сосредоточилась в нижних слоях атмосферы. С появлением первых архаичных макроэргов, способных действовать на поверхности капельных систем и вдали от них – в газово-паровой среде, сама атмосфера по своим свойствам стала похожа на цитоплазму космических масштабов, где реализовались необходимые и достаточные условия для формирования и эффективного функционирования названых цепочек. Энергия распада предшественников современных макроэргов обеспечивала преодоление более высоких активационных барьеров в сопряженных каталитических реакциях синтеза и распада органики. Каталитический синтез соединений мог осуществляться более эффективно. Поэтому темп обогащения атмосферы новыми соединениями, как количественно, так и видами, существенно возрос. В анализе эволюционного развития живых систем и «преджизни» в частности наиболее важной и первоочередной является проблема появления информации о новых соединениях, входящих в их структуры. Э.М. Галимов в своей работе [13] первостепенную роль в рождении новой информации в живых системах отдает комбинаторике их элементов. Правда, он пишет, в основном, о комбинаторике генов в хромосомах. Однако его выводы можно отнести к эволюционному развитию всех сложных органических соединений, и весьма продуктивно. Особенно это касается анализа этапов развития «преджизни», когда крупных полинуклеотидов в природе (в тогдашней атмосфере) еще не было и когда они только начали появляться. Так перестановка части длинной молекулы углеводорода, включающей кроме одинарных также одну или несколько двойных С-Ссвязей, из средины на ее края, может изменить физические свойства этого соединения, но весьма незначительно. А перестановка частей сложных углеродсодержащих соединений, имеющих в своем составе атомы кислорода, азота, ТЕОРИЯ ЭВОЛЮЦИИ металлов и других элементов кроме углерода, напротив, эти свойства может существенно менять. Кроме того, каталитический распад и синтез с участием макроэргов предоставляли неограниченные возможности не только для комбинаторики каких-либо частей конкретного углеродсодержащего соединения, но и комбинаторики его частей с частями соединений других видов и классов. Распад (спонтанный или каталитический) органики предоставлял разного рода части соединений для последующей их сшивки-синтеза в случайно образовывавшихся комплексах с катализаторами и макроэргами. Таким образом, рождалась новая информация. Конечно, проточный характер каталитического метаболизма не был единственным фактором, определяющим ее накопление. В этом направлении действовали и другие факторы. Можно назвать естественную радиоактивность минералов поверхности Земли, солнечную радиацию, мощные электрические разряды в плотной атмосфере и выбросы расплавленной породы и газов предшественниками вулканов на формирующейся коре. Однако, по нашему мнению, доминирующим фактором в рождении новой информации (новых соединений) изза своей массовости была упомянутая выше комбинаторика. Появление первых макроэргов существенно ускорило эволюционное развитие органического мира «преджизни». До этого в основном синтезировались все более усложняющиеся органические соединения: сахара, нейтральные жиры, жирные кислоты, аминокислоты, нуклеиновые кислоты и др. Можно говорить о разрастании их представителей на моноуровне. Участие в синтетических процессах макроэргов обеспечило переход иерархии органического мира на более высокие уровни. Мономеры, имеющие сродство друг к другу, образовывали комплексы. Некоторые из них (комплексов), обладая хотя и малой, но все же каталитической активностью, с участием макроэргов могли катализировать метаболические реакции синтеза и распада новых видов мономерных соединений. Однако эти комплексы были нестойки и быстро разрушались в бурлящей атмосфере тогдашней Земли. Особую роль в образовании более устойчивых комплексов АБИОГЕНЕЗ В АТМОСФЕРЕ... играло (и играет до сих пор) диполь-дипольное взаимодействие. Это касается в первую очередь аминокислот, поскольку многие их виды обладают значительным дипольным моментом. Стабилизации таких комплексов способствовали молекулы воды, создавая гидратную оболочку вокруг взаимодействующих аминокислот. Напомним, что речь идет о времени, когда вода на Земле находилась в ее атмосфере в виде плотного пара. Его парциальное давление на уровне тверди планеты составляло ~200 атмосфер. На поверхности некоторых указанных выше комплексов (агрегатов), которые обладали каталитической способностью2, с участием макроэргов и без них могли происходить реакции установления пептидных C-N связей между аминокислотами, взаимодействующими с такими объектами. Сначала могли синтезироваться пептиды, состоящие из пары аминокислот (в том числе разных видов), потом – из одной кислоты и такой пары и так далее, все более удлиняясь. Короткие пептиды в свою очередь могли образовывать более устойчивые комплексы, чем таковые из мономеров. Они также могли обладать каталитическими свойствами (Возможно подобным феноменом объясняется «кажущаяся» сверхактивность ферментов, катализирующих в клетках некоторые метаболические реакции). На этих комплексах с участием макроэргов могли синтезироваться более длинные пептиды, внося свою долю в процессы накопления в атмосфере более крупных пептидных соединений. Так стартовала эпоха полимерных соединений. Все сказанное для аминокислот и их соединений верно также для всех других видов, семейств и классов органических веществ, появившихся к тому времени в атмосфере. Полимеризация и комбинаторика органики на уровне мономеров и полимерных соединений, задействовавших практически все виды ковалентной связи (C-C, C-O, C-N и др.), 2 Известны работы о наличии ферментативной активности агрегатов органических соединений, например, [14], где речь идет о катализе белковыми агрегатами реакций распада этих же белков в водном растворе. Но такие реакции принципиально обратимы и, при накоплении продуктов прямой реакции и достаточной концентрации макроэргов, обеспечивающих сопряжение гидролиза последних с синтезом исходного продукта, идут в обратном направлении. РЭНСИТ | 2014 | ТОМ 6 | НОМЕР 1 113 114 ЗАРИЦКИЙ А. Р., ВОРОНЦОВ Ю. П., ПРОНИН В. С. определили появление огромного разнообразия сравнительно коротких полимерных органических соединений с соответствующим разнообразием их физико-химических свойств. В этой связи особо отметим сахара, жирные и нуклеиновые кислоты и липиды. Следует более подробно остановиться на некоторых физико-химических свойствах аминокислот. Большинство из них обладают (как указано выше) значительным дипольным моментом, определяющим их диполь-дипольные взаимодействия. Во времена, относящиеся к рассматриваемому периоду развития «преджизни», еще не было естественного отбора по хиральности (типов симметрии L и D) аминокислот, отличающимися, в том числе, направлением результирующего дипольного момента молекул. Поэтому классы и семейства пептидов и их комплексов по своим видам были чрезвычайно богаты, поскольку эта органика синтезировалась из аминокислот обеих хиральностей в любой последовательности. При наличии макроэргов синтез коротких пептидов на более крупных пептидах и их комплексах, несомненно, был широко представлен. В настоящее время такой синтез известен как нерибосомальный. Например, известно, что короткие циклические пептиды-антибиотики могут синтезироваться с участием АТФ на крупных глобулярных белках [15]. Две пептидные их составляющие идентичны (по количеству и последовательности аминокислот) и соединены C-N-связями по принципу голова-хвост. Ясно, что при таком синтезе решающее значение имеет диполь-дипольное взаимодействие аминокислот как первой половины синтезируемого циклического пептида с аминокислотами фермента-глобулы, так и такое же взаимодействие этой половины пептида со второй его частью. При этом основной вклад во взаимодействие аминокислот, непосредственно прилегающих друг к другу, вносит притяжение их противоположно направленных дипольных моментов. Однако вернемся к тем временам и проблемам, которые мы анализируем. Если на агрегатекомплексе из пептидов в месте катализа оказывался пептидный фрагмент с аминокислотами одной хиральности, то на нем могли синтезироваться идентичные по аминокислотам пептиды, но 1 НОМЕР | ТОМ 6 | 2014 | РЭНСИТ ТЕОРИЯ ЭВОЛЮЦИИ другой хиральности. Этот процесс напоминает штамповку деталей с использованием матрицы – пептид одной хиральности и пуассона – другой. В отличие от штамповки такой синтез пептидов мог прекратиться на любой его фазе в результате воздействия тепловых толчков. Поэтому на выходе реакций синтеза могли оказываться пептиды с одинаковой последовательностью аминокислот, но отличающиеся в своем составе по их количеству. Заметим также, что такие процессы воспроизводства простых, коротких пептидов на матрице – пептиде другой хиральности в составе агрегатов допускают, в принципе, большой процент ошибок, так как их основу определяет только диполь-дипольное взаимодействие аминокислот. Это, однако, обеспечивало в процессах синтеза сложных пептидных соединений по методу сшивки богатую базу для комбинаторики, что было важным фактором в формировании новой каталитической базы. Как известно в настоящее время воспроизводство генетического аппарата клетки и информационных частей вирусов, внедрившихся в цитоплазму, является сверхсложным процессом и может протекать только при участии белков-ферментов, объединенных в комплексы [16]. Однако нетрудно усмотреть принципиальное сходство каждого из его отдельных этапов с процессами воспроизводства пептидов, представленные выше в двух предыдущих абзацах. Синтез нуклеозида из нуклеинового основания с участием макроэргов мог происходить также, как и пептидов, на пептидном комплексе, а фосфорилирование синтезированного нового соединения для образования нуклеотида на другом комплексе. Полимеризация нуклеотидов могла, соответственно, происходить на третьем. Конечно, ничто не мешало указанным пептидным комплексам объединиться в более крупные агрегаты. При этом некоторые из них, возможно, сохраняли ферментативные свойства отдельных своих частей. Однако таким способом как крупных полинуклеотидов, так и длинных пептидных цепей в тех условиях получить было невозможно. Прежде чем нарастет цепочка мономеров (будь то пептиды или нуклеотиды) значительной длины, тепловое воздействие, ТЕОРИЯ ЭВОЛЮЦИИ во-первых, разрушит ферментативный пептидный агрегат и/или, во-вторых, оторвет от него растущий полимер. Все же в рассматриваемый период развития «преджизни» средств (каталитической базы и макроэргов) для реализации простейших процессов воспроизводства коротких полинуклеотидов обеих хиральностей хватало и непрерывно наращивалось. Поэтому следует указать только пути, по которым самопроизвольно и эффективно могло происходить их удлинение. Сдерживающим фактором, ограничивающим скорости энергозависимых процессов синтеза все более сложных соединений, в том числе длинных полинуклеотидов, на ранних этапах эволюционного развития «преджизни» являлось отсутствие, во-первых, запасов макроэргов непосредственно в зоне такого синтеза и, во-вторых, механизмов подведения туда всех участников энергозависимых многокомпонентных синтетических реакций. Однако это не означает, что крупных полинуклеотидов и полипептидов, обладающих ферментативной активностью, не было. Малое Рис. 1. Углеводородная капля со слоем липидов и встроенными в этот слой крупными органическими соединениями и их комплексами: 1 – углеводороды; 2 – молекулы липидов; 3 – белковый комплекс, осуществляющий ферментативную разборку липидов; 4 – белково-нуклеотидный комплекс, ассоциированный с липидами; 5 – белковый комплекс, катализирующий реакции распада углеводородов капли и сопряженную с этим распадом реакцию синтеза липидов. Стрелкой указано направление движения комплекса 4 вместе с липидами к комплексу 3. АБИОГЕНЕЗ В АТМОСФЕРЕ... представительство в мире органики пока не могло обеспечить им ведущей роли в эволюционном развитии «преджизни». С появлением липидов положение кардинально изменилось. Синтез липидов осуществлялся преимущественно на поверхности капель. Имея высокое сродство к углеводородам капель, они накапливались на их поверхностях в виде слоя, как показано на рис. 1. Распад липидов происходил (с участием макроэргов и без них) на поверхности капель и непосредственно в газовой среде атмосферы на комплексах органических соединений. В начале активационные барьеры таких реакций (прямой и обратной) преодолевались на поверхности капель за счет их понижения полями, обусловленными наличием Z-потенциала на границе двух сред – конденсированных углеводородов и газовой среды. Однако, по мере охлаждения атмосферы и роста размеров капель, из-за усиления конденсации углеводородов, этот потенциал, достигнув максимальной величины, начал неуклонно уменьшаться. Поэтому доминирующими стали реакции, катализируемые агрегатами соединений (в том числе и из пептидов), адсорбированными непосредственно на углеводородах и липидных слоях капель или в них встроенными и обладавшими соответствующей каталитической активностью. При распаде липидов, наиболее вероятно, отделялась и уходила в газовую среду полярная головка молекулы. Выделяемая при этом химическая энергия могла либо рассеиваться, переходя в тепло, либо использоваться для преодоления активационных барьеров в сопряженных энергозависимых реакциях синтеза сложных соединений. Таким образом, липиды могли выступать в роли макроэргов, но действующих только на поверхности капель. Ацильные цепи распавшихся липидов смешивались с углеводородами капель и утилизировались как их основной энергоемкий материал. Заметим, что при таком каталитическом распаде липидов (отделении их полярных головок или разрыве C-C связей в ацильных цепях) продукты реакций уходят из активной зоны, освобождая место для подхода туда ближайших нативных липидов, немедленно вступающих в процесс разборки. При этом РЭНСИТ | 2014 | ТОМ 6 | НОМЕР 1 115 116 ЗАРИЦКИЙ А. Р., ВОРОНЦОВ Ю. П., ПРОНИН В. С. происходит либо сдвиг всей липидной массы слоя к данному месту разборки липидов вместе с ассоциированными и встроенными в него комплексами и соединениями, либо сам агрегаткатализатор распада липидов сближался с ними. Начали формироваться структуры, способные сопрягать процессы распада липидов с процессами синтеза новых крупных соединений. Появление таких структур с зоной сопряжения синтеза крупных соединений с распадом липидов на поверхности капель сняло указанное выше ограничение скорости синтетических энергозависимых процессов. Представительство крупных органических соединений, в первую очередь, пептидной природы и полинуклеотидов в органическом мире существенно расширилось. Кроме того, полимерные цепочки настолько удлинились, что крупные полипептиды начали сворачиваться в глобулы, причем некоторые из них обладали ферментативной активностью, а нуклеотидный состав информационных молекул в своем развитии оказался достаточным для размещения в них информации о первых пептидных ферментах. В развертывании эволюционных преобразований органического мира «преджизни» отметим еще один важный момент. При движении липидных слоев вместе со всем органическим материалом, с ними ассоциированным, к месту выделения химической энергии распада углеводородов и липидов капель самопроизвольно формировались наиболее благоприятные условия для протекания многокомпонентных реакций синтетических процессов. К мощному источнику энергии, с большим ее запасом (помимо углеводородов в одном месте одновременно можно утилизировать и липиды слоя), подводились все участники таких реакций. Другим важным моментом, в этой связи, являлась деятельность агрегатов соединений и первых ферментов, адсорбированных на поверхности капель, встроенных в липидный слой и способных катализировать реакции процессов синтеза самих липидов. Такая деятельность приводила к сдвигу всей липидной массы от места синтеза липидов, если ферментативные структуры синтеза были надежно закреплены в определенном месте на поверхности 1 НОМЕР | ТОМ 6 | 2014 | РЭНСИТ ТЕОРИЯ ЭВОЛЮЦИИ капель (рис. 1). В противном случае они сами вместе с ассоциированными с ними другими соединениями удалялись по поверхности капли от первоначальной зоны синтеза липидов. Таким образом, появилась основа для формирования механизмов транспорта веществ и их структур в латеральном по отношению к липидным слоям направлении. Устранение препятствий для ускорения синтеза и усложнения органических соединений, в первую очередь полипептидов и полинуклеотидов существенно ускорило их накопление в атмосфере, создав предпосылки для появления, во-первых, информационных молекул (РНК и ДНК), без которых нет ни вирусов, ни клеток, и, во-вторых, ферментов, способных понижать активационные барьеры в простых реакциях метаболизма. Следует напомнить, что в рассматриваемый эволюционный период «преджизни» вся масса воды Земли все еще находилась в виде пара в ее атмосфере. Гидрофобная часть липидов, имея сродство к углеводородам капли, была обращена к ее поверхности, а полярные головки в противоположную сторону (рис. 1). На границе раздела сред – «конденсированная нейтральная липидная (упорядоченные ацильные цепи липидов) и газовая среды» всегда наличествуют поля, обуславливая относительно указанной газовой среды положительный потенциал. Эти поля заставляли полярные головки липидов ориентироваться положительным полюсом к газу, причем положение их продольных осей было близко к нормали к поверхности капли. Подвижные молекулы воды (в данном случае водяного пара атмосферы), имеющие самый высокий удельный дипольный момент из известных веществ, взаимодействуя с дипольными головками липидов, закрепляли такую ориентацию. (Такая геометрия в точности соответствует внутриклеточному слою липидов, обращенных своими полярными головками в цитоплазму.) Водородные связи, устанавливающиеся между ацильными цепями молекул липидов, высокое сродство этих частей к углеводородам капли и диполь-дипольные взаимодействия полярных головок с молекулами паров воды атмосферы обуславливали ТЕОРИЯ ЭВОЛЮЦИИ устойчивость данной системы к разрушающим факторам бурлящей тогдашней атмосферы. С появлением липидов и все большим накоплением их на поверхности капель (вплоть до полного их покрытия) возник новый фактор отбора соединений по их способности взаимодействовать с источником свободной энергии, каковым являлась капля. Для утилизации углеводородов капель ферменты должны были встраиваться в липидный слой. Пептидные соединения в этом отношении имели преимущество, поскольку аминокислоты, из которых они состоят, имея дипольный момент, могли эффективно взаимодействовать с дипольными головками липидов слоя. Полинуклеотиды, наоборот, сродства к липидным слоям не имеют, однако охотно образуют комплексы с пептидными соединениями. Такие комплексы могли встраиваться в липидный слой стороной пептидной части, в то время как полинуклеотидная часть находилась в атмосфере. Это открывало возможность для участия полинуклеотидов в сопряженных реакциях синтеза сложных соединений с использованием одновременно энергии, запасенной липидами, углеводородами и макроэргами капель и атмосферы. Поэтому можно говорить о появлении предпосылок для формирования современных механизмов воспроизводства белков и полинуклеотидов на матричной основе с привлечением информационных молекул РНК и ДНК. Заметим, что полинуклеотиды, адгезированные в пептидных комплексах на поверхности капель, на начальном этапе развития «преджизни» оказались полезными для примитивного метаболизма этих предшественников клеток не как носители информации, а в качестве постоянно действующих депо макроэргов – мононуклеотидов. В данной статье мы не ставили перед собой цель подробно осветить сложнейшую проблему появления механизмов биосинтеза нуклеиновых кислот. Указаны только предпосылки (необходимые условия) и возможные пути эволюционного развития систем «преджизни», которые могли это обеспечить. Огромное видовое разнообразие органических соединений в атмосфере (было из чего выбирать) и наличие для них центров притяжения и отбора на постоянных источниках свободной энергии, каковыми в рассматриваемый период были АБИОГЕНЕЗ В АТМОСФЕРЕ... «активные капли» с липидными слоями на их поверхности, стали решающими факторами в появления первых ростков жизни. Мы имеем в виду предшественников современных вирусов. 3. ПРЕДПОСЫЛКИ ДЛЯ ПОЯВЛЕНИЯ ВИРУСОВ В настоящее время, как известно, вирусы могут воспроизводить свои тела (в том числе информационные части) только в клетках, используя их энергетический метаболизм. Полная сборка вирусов может происходить как в цитоплазме с последующим их выходом из клетки, так и вне клеток после выхода их тел в окружающую среду. Кроме того, популяции вирусов имеют лучшую эволюционную перспективу, если их тела, попадая в цитоплазму, прямо или опосредованно активируют энергетический метаболизм клеток, обеспечивая симбиотическое развитие живого мира. Информационные молекулы вирусов вне клеток часто мутируют и, вернувшись в клетки, вносят в них новую информацию, и, если она полезна для развития клеточного мира, находит свое закрепление в геноме клеток. Таким образом, на сегодняшний день ведущим звеном в отборе новой информации в живом мире является ее полезность для клеток. Однако в тот период зарождения живого мира, который мы рассматриваем, клеток еще не было. Новая информация рождалась в основном в результате комбинаторики соединений, участвующих в качестве субстратов в энергозависимых, простых ферментативных реакциях синтеза крупной органики. Такие реакции в принципе обратимы. Скорости прямых и обратных реакций экспоненциально зависят от высоты их активационных барьеров. Как правило, высота барьера реакций синтеза (установления C-C, C-N, C-O и других связей) много выше, чем обратной реакции (распада соединений). Поэтому зачастую скорости прямых реакций (синтеза) много меньше обратных. Участие макроэргов в реакциях синтеза соединений способно устранять такое несоответствие. Тогда доминирование одного из направлений простейших обратимых реакций может определяться концентрациями в среде, как реагентов, так и макроэргов. Чрезвычайная турбулентность тогдашней РЭНСИТ | 2014 | ТОМ 6 | НОМЕР 1 117 118 ЗАРИЦКИЙ А. Р., ВОРОНЦОВ Ю. П., ПРОНИН В. С. атмосферы определяла неоднородность и быстрое изменение концентраций реагирующих веществ, в том числе и макроэргов в различных регионах газовой среды. Быстрая смена направлений реакций по указанной причине обеспечивала проток реагентов, то есть создавала наиболее благоприятные условия для комбинаторики и, следовательно, для появления новых сложных соединений с новыми свойствами. Среди этих соединений оказались и такие (мы их называем «вирусоподобными телами»), которые обладали по отношению к источникам свободной энергии свойствами, сходными с телами современных вирусов по отношению к клеткам. Такие вещества прямо (катализируя реакции энерговыделений) или опосредовано (образуя комплексы с катализаторами) интенсифицировали процессы энерговыделения, обеспечивая появление более сложных соединений. Что касается полипептидов и полинуклеотидов, то и их цепи последовательно удлинялись. Появились крупные глобулярные полипептиды, в том числе обладающие ферментативными свойствами, и достаточно крупные молекулы РНК и ДНК, способные вместить информацию о таких пептидах. Крупные глобулы полипептидов из-за своего малого представительства не могли пока принять эстафету пептидных комплексов и стать основными участниками каталитических процессов эволюционирующего метаболизма в тогдашней атмосфере. Подчеркнем, что только переход процессов синтеза полипептидов к их матричному воспроизводству с участием полинуклеотидов мог это обеспечить. Рассмотрим один из возможных путей такого перехода. Среди новых сложных соединений находились вещества (в основном соединения пептидной природы), которые, образуя комплексы с информационными молекулами (РНК и ДНК), не утрачивали своего сродства как к углеводородам капель, так и к пептидам и липидам на их поверхности. Понятно, что такие комплексы стали удерживаться на поверхности капель. Появление в реакционной зоне информационных молекул послужило предпосылкой для зарождения простейших механизмов воспроизводства как пептидных соединений на матричной основе с участием 1 НОМЕР | ТОМ 6 | 2014 | РЭНСИТ ТЕОРИЯ ЭВОЛЮЦИИ РНК и ДНК, так и самих информационных молекул. Напомним, что и в настоящее время основой сложных механизмов воспроизводства указанных соединений являются простейшие обратимые ферментативные реакции. Поэтому при избытке в реакционной зоне макроэргов и полного видового представительства реагентов они направлены в сторону синтеза сложных соединений, а, если это условие не выполняется, то в сторону распада вплоть до полной их разборки. (Наглядной иллюстрацией последнего положения является утрата генетического аппарата и существенное обеднение ферментативного пула предшественников эритроцитов крови высших животных и человека после выхода их из костного мозга). По указанной причине в тогдашней атмосфере (в ее нижних слоях) реализовался проток соединений, способных нести закодированную информацию не только о строении пептидов и белков, но и их самих. Проточное равновесие скоростей синтеза и распада органических соединений (причем любое изменение их структуры мы тоже считаем распадом) является необходимым и достаточным условием сохранения их количества и указанной выше способности. Неизбежное по мере остывания планеты сгущение органического материала в нижних слоях атмосферы способствовало ускорению скоростей синтеза информационных молекул (полинуклеотидов) и установлению баланса со скоростями их распада на все боле высоких концентрационных уровнях. По этой же причине усилился синтез макроэргов, и на поверхности капель в липидных слоях сложились условия, при которых неизбежно протекают упомянутые выше простейшие реакции воспроизводства как пептидов, так и РНК и ДНК на матричной основе с использованием указанных информационных молекул. Конечно, кодировались и воспроизводились пока сравнительно короткие полимерные цепи, но по составу и видам они были чрезвычайно разнообразны. Среди новых видов пептидов, воспроизводимых матричным способом, оказались соединения, способные интенсифицировать энерговыделения в реакциях утилизации энергоемких субстратов, протекающих как в газовой среде, так и на поверхности капель. Наметилась петля ТЕОРИЯ ЭВОЛЮЦИИ положительной обратной связи между количеством таких пептидов в природе и скоростью их матричного воспроизводства на РНК. С другой стороны, накопление в атмосфере этих пептидов влекло за собой неуклонную интенсификацию энерговыделений капель. Это, в свою очередь, обуславливало удлинение цепей не только полипептидов, но и информационных молекул вплоть до их способности нести информацию о все более длинных соединениях пептидной природы ферментах, способных катализировать реакции процессов нарождающегося метаболизма, в том числе матричного воспроизводства как белков, так и полинуклеотидов. Для эффективного воспроизводства таким способом тех и других необходимо было только свести их в одно место с мощным источником свободной энергии. Решение этой задачи, как показано выше, обеспечили липиды. Их появление и формирование сплошного липидного слоя на поверхности капель стало решающим системообразующим фактором в появлении механизмов репликации информационных молекул. В результате деятельности встроенных в липидный слой ферментов, катализирующих реакции распада липидов, все соединения, имеющие к ним сродство и адгезированные на поверхности липидного слоя, двигаясь вдоль поверхности капель к указанным ферментам, стали образовывать группы – предшественники современных клеточных органелл, в том числе, ответственных за передачу генетической информации. Если учесть, что в таких группах оказывались как ферменты, способные утилизировать углеводородный материал и липиды капли, так и ферменты, осуществляющие катализ реакций синтеза полимеров на матричной основе, то появление первых простейших упомянутых выше репликационных механизмов стало неизбежным. Именно соединение в одном месте мощного источника свободной энергии и элементов механизмов матричного воспроизводства полинуклеотидов и белков, обеспечило сборку этих механизмов. Получая достаточную энергетическую поддержку, они заработали. Следует подчеркнуть, что появление информационных молекул, способных вмещать АБИОГЕНЕЗ В АТМОСФЕРЕ... информацию о крупных полипептидах, не сузило видовое многообразие органического мира, как и количества веществ каждого из видов. Наоборот, существенно расширило видовой спектр органических соединений, в том числе в сторону их все большего и большего усложнения. За счет более производительного матричного воспроизводства этих веществ их количество в атмосфере непрерывно росло. Это касается также и белков-ферментов. Расширение их видового состава в конечном итоге привело к появлению ферментов, способных катализировать реакции не только энерговыделений и своего матричного воспроизводства, но и репликации как РНК, так и ДНК. Подчеркнем еще раз, что появление и накопление на поверхности капель липидов было тем решающим системообразующим фактором, который обеспечил воспроизводство обеих частей (тел и информационных молекул) первых примитивных вирусов. Отметим, что роль липидов, как энергоемкого архаичного макроэрга в процессах воспроизводства полинуклеотидов и крупных полипептидов до нас не выяснялась. При разрыве С-С-связи при ферментативном распаде такой молекулы выделяется энергия, которой хватает для синтеза из дифосфатов нуклеозидов и неорганического фосфата сразу всех четырех мономеров современных макроэргов в любом наборе: АТФ, ГТФ, ТТФ и УТФ или ЦТФ, необходимых для удлинения полинуклеотидных цепей информационных молекул. Именно эти макроэрги используются в качестве энергоемкого и строительного материала в многокомпонентных ферментативных реакциях процессов воспроизводства информационных и других сложных соединений. В появлении архаичных вирусов следует отметить роль вирусоподобных тел, обеспечивших энергетическую основу для воспроизводства самих себя и информационных частей вирусов, а также защиту последних от агрессивного влияния окружающей среды. Напомним, что под термином «вирусоподобные тела» мы понимаем соединения, как правило, пептидной природы, которые обладали сродством к полинуклеотидам и поэтому были способны образовывать с ними устойчивые комплексы. Эта группа соединений выделилась из ферментативного комплекса капель и при этом РЭНСИТ | 2014 | ТОМ 6 | НОМЕР 1 119 120 ЗАРИЦКИЙ А. Р., ВОРОНЦОВ Ю. П., ПРОНИН В. С. сохранила свою способность катализировать реакции их энергетического метаболизма и/или его активировать. Естественно полагать, что сначала сформировались механизмы репликации РНК, как более простые по сравнению с таковыми для ДНК. Это прямо соответствует генеральной линии эволюционного развития живого мира. Однако напомним о великом разнообразии соединений, что, по нашему мнению, было характерно для органического мира того периода. Наиболее ярко это иллюстрирует тогдашнее одинаковое представительство соединений разной хиральности и их гибридов, в то время как в настоящее время в живом мире присутствуют только соединения одной. Поэтому, весьма вероятно, существовали и были реализованы и другие, не известные на сегодняшний день, более сложные пути формирования механизмов репликации информационных молекул. Однако, по мере обогащения нарождающегося живого мира новыми соединениями с новыми свойствами, эти пути непрерывно изменялись как в сторону упрощения, так и в сторону усложнения в соответствии с непрерывно изменяющимися окружающими условиями. Вопрос первенства появления в природе механизмов репликации РНК или ДНК не столь важен для раскрытия данной темы. Важно только то, что они самопроизвольно появились, и эти соединения стали воспроизводиться матричным способом только на поверхности капель, поскольку в то время эти процессы могли обеспечиваться энергией в полной мере только там. Под воздействием тепловых толчков полинуклеотиды, синтезируемые на поверхности капель in novo, покидали капли. В газовой среде они образовывали комплексы с пептидами, которые защищали их от агрессивного влияния окружающей газовой среды. Среди этих пептидов в силу многообразия их видов нашлись и такие, у которых, во-первых, последовательность аминокислот соответствовала информации, кодируемой последовательностью нуклеотидов в молекулах РНК и ДНК, и, во-вторых, могли активировать процессы выделения свободной энергии при утилизации углеводородов и липидов капли. Это обеспечивало таким пептидам преимущество перед другими 1 НОМЕР | ТОМ 6 | 2014 | РЭНСИТ ТЕОРИЯ ЭВОЛЮЦИИ видами в воспроизводстве на поверхности капель. Соответственно, представительство в атмосфере росло как пептида, так и самой информационной молекулы, поскольку каждая из двух информационных молекул с большей вероятностью могла получать пептид для своей защиты непосредственно после репликации такого полинуклеотида. Сформировавшаяся таким образом петля обратной связи между матричным воспроизводством соединений пептидной природы и репликацией информационных молекул в конечном итоге привели к появлению в атмосфере первых вирусов в количестве, достаточном для поддержания их популяций и дальнейшего эволюционного развития. 4. ЗАКЛЮЧЕНИЕ Рассматриваемый этап абиогенеза представлял собой длинный путь появления все новых и новых, непрерывно усложняющихся, органических соединений. В начале синтезировались мелкие молекулы, далее, все более крупные (мономеры). Выход на новый этап эволюционного развития органического мира ознаменовался появлением полимеров, которые тоже непрерывно усложнялись. Особое место в абиогенезе заняли крупные информационные молекулы – полинуклеотиды. Эти соединения оказались полезными для примитивного метаболизма в качестве постоянно действующих депо макроэргов. Появление первых механизмов матричного воспроизводства сложных органических соединений не только пептидной природы, но и полинуклеотидов ознаменовало завершение данного этапа абиогенеза, обозначаемого нами (вслед за М. Руттеном [17]) как «преджизнь». Архаичные вирусы стали первыми ростками жизни. Их эволюционное развитие в симбиотическом взаимодействии с локальными источниками свободной энергии, утилизирующими запасы углеводородного сырья и липидов в нижних слоях атмосферы, стало впоследствии ведущим фактором в формировании предпосылок для появления первых клеток. Данный этап характеризуется также последовательным усложнением открытого метаболизма капель и процессов с участием ТЕОРИЯ ЭВОЛЮЦИИ органических соединений, протекавших в газовой среде нижних слоев атмосферы. Это выражалось в появлении цепочек реакций (порой разветвленных) превращений исходных энергоемких субстратов в конечные продукты. Самым важным эволюционным достижением этого этапа, помимо появления вирусов, стало формирование на базе протока липидной массы первых механизмов транспорта органики вдоль липидных слоев. Именно этот феномен лежит в основе действия не только указанных выше, но и других молекулярных механизмов, действующих в современных клетках, например, при формировании их органелл, митотической деятельности и других. Поэтому можно утверждать, что первыми предшественниками клеток (протобионтах по А.И.Опарину), наиболее вероятно, были указанные «активные» капли – своеобразные генераторы свободной энергии, которая аккумулировалась в макроэргах и липидах. В отличие от общераспространенных попыток вывести многообразие из единичного наш подход к раскрытию темы противоположный: из многообразия – частное, успешно противостоящее разрушающему действию стихий. Мы указываем также основное свойство этих частных случаев, обеспечивающее такую устойчивость, присущую живой материи – непрерывный проток вещества, информации и энергии. АБИОГЕНЕЗ В АТМОСФЕРЕ... Sites for the Origin and Evolution of Life. Origins of Life, 1985, 15:327-345. 9. Martin W, Baross J, Kelley D, Russel MJ. Hydrothermal Vents and Origin of Life. Nat.Rev.Microbiol., 2008, 6:805-814. 10. Mulkidjanian AY. On the Origin of Life in the Zinc World: 1. Photosynthesizing, Porous Edifices Built of Hydrothermallly Precipitated Zinc Sulfide as Cradles of Life on Earth. Biol.Direct, 2009, 4:26. 11. Mulkidjanian AY, Bychkov AY, Dibrova DV, Galperin MY, Koonin EV. Open Questions on the Origin of Life an Anoxic Geothermal Fields. Orig.Life Evol. Biosph.,2012, 42(5):507-16. 12. Зарицкий АР, Грачев ВИ, Воронцов ЮП, Пронин ВС. Энергетические аспекты абиогенеза в атмосфере на нанокаплях углеводородного аэрозоля. РЭНСИТ, 2013, 5(2):105-125. 13. Галимов ЭМ. Феномен жизни. М., Едиториал УРСС, 2006, 254 с. 14. Maryada Sharma, Manni Luthra – Guptasarma. Degradation of protein upon storage at near-neutral Ph: Indications of a proteolytic/gelatinolytic activity associated with aggregates. Biochimica at Biophysica Acta, 2009, 1790:1282-1294. 15. Березов ТТ, Коровкин БФ. Биологическая химия. М., Медицина, 1990, 416 с. 16. Мари Р, Греннер Д, Мейес П, Родуэлл В. Биохимия человека, т.2. М., Мир, 2009, с.73-81. 17. Руттен М. Происхождение жизни естественным путем. М., Мир, 1973, 411 с. ЛИТЕРАТУРА 1. Опарин АИ. Происхождение жизни. М., Моск. рабочий, 1924. 2. Haldane JBS. The Origin of Life. Rationalist Annual, 1928, 148:3-10. 3. Miller SL, Urey HC. Organic Compound Synthesis on the Primitive Earth. Science, 1959, 130(3370):245251. 4. Fox SW. The Evolutionary Significance of PhaseSeparated Microsystems. Orig.Life, 1976, 7:49-68. 5. Wachtershauser G. The Origin of Life and its Methodological Challenge. J.Theor.Biol., 1997, 187:483-494. 6. Huber C, Eisenreich W, Hecht S, Wachtershauser G. A Possible Primordial Peptide Cycle. Science, 2003, 301:938-940. 7. Russel MJ, Hall AJ. The Emergence of Life from Iron Monosulphide Bubbles at a Submarine Hydrothermal Redox and Ph Front. J.Geol.Soc.London, 1997, 154:377-302. 8. Baross JA, Hoffman SA. Submarine Hydrothermal Vents and Associated Gradient Environments As Зарицкий Александр Романович с.н.с. Физический институт им. П.Н.Лебедева РАН 119991 Москва, Ленинский пр., 53, к. 389. +8 499 132-6309, zaritsky@sci.lebedev.ru Воронцов Юрий Петрович д.м.н., проф., ДГК Больница №13 им. Н.Ф.Филатова Мосгорздрава 103001 Москва, Садовая-Кудринская ул., 15, Yupmkpsp@rambler.ru Пронин Вячеслав Сергеевич д.м.н., проф., Первый Мос. гос. мед. университет им. И.М.Сеченова Минздравсоцразвития РФ 119991 Москва, Трубецкая ул., д.8, стр.2, +8 499 248-3885, vspronin@yandex.ru РЭНСИТ | 2014 | ТОМ 6 | НОМЕР 1 121 122 THEORY OF EVOLUTION ABIOGENESIS IN THE ATMOSPHERE: ARCHAIC METABOLISM, COMBINATORIAL REPLICATION AND PRIMARY VIRUSES Zaritsky Alexander R. Lebedev Physics Institute, Russian Academy of Sciences, http://www.lebedev.ru 119991 Moscow, Russian Federation zaritsky@sci.lebedev.ru Vorontsov Yuri P. Filatov Hospital no. 13, Moscow Department of Healthcare, http://www.13dgkb.ru 103001 Moscow, Russian Federation yupmkpsp@rambler.ru Pronin Vyacheslav S. Sechenov First Moscow State Medical University, Ministry of Healthcare and Social Development of RF, http://www.mmm.ru 119991 Moscow, Russian Federation vspronin@yandex,ru This study represents an analysis of abiogenesis processes at the step immediately succeeding the "beginning of the beginnings". This step exhibits further augmentation of free energy liberation by local atmospheric sources that provides grounds for progressive increase in complexity of organic compounds, metabolic processes, and archaic metabolic pre-cellular systems, and so, forms preconditions for the first life sprouts. The incipience of lipids had become the system-forming factor which provided implementation of the above prerequisites in archaic mechanisms of matrix replication of organic compounds and primary viruses. Keywords: abiogenesis, atmosphere, primary separation, hydrocarbon nanodroplets, autonomous energy source, metabolism, homeostasis, symbiosis. UDC 576.1+551.7 Bibliography – 17 references RENSIT, 2014, 6(1):110-123 REFERENCES 1. Oparin AI. The Origin of Life on Earth. NY, Academic Press, 1938. 2. Haldane JBS. The Origin of Life. Rationalist Annual, 1928, 148:3-10. 3. Miller SL, Urey HC. Organic Compound Synthesis on the Primitive Earth. Science, 1959, 130(3370):245-251. 4. Fox SW. The Evolutionary Significance of PhaseSeparated Microsystems. Orig.Life, 1976, 7:49-68. 5. Wachtershauser G. The Origin of Life and its Methodological Challenge. J.Theor.Biol., 1997, 187:483-494. 6. Huber C, Eisenreich W, Hecht S, Wachtershauser G. A Possible Primordial Peptide Cycle. Science, 2003, 301:938-940. 7. Russel MJ, Hall AJ. The Emergence of Life from Iron Monosulphide Bubbles at a Submarine 1 НОМЕР | ТОМ 6 | 2014 | РЭНСИТ Received 21.01.2013 Hydrothermal Redox and Ph Front. J.Geol.Soc. London, 1997, 154:377-302. 8. Baross JA, Hoffman SA. Submarine Hydrothermal Vents and Associated Gradient Environments As Sites for the Origin and Evolution of Life. Origins of Life, 1985, 15:327-345. 9. Martin W, Baross J, Kelley D, Russel MJ. Hydrothermal Vents and Origin of Life. Nat.Rev. Microbiol., 2008, 6:805-814. 10.Mulkidjanian AY. On the Origin of Life in the Zinc World: 1. Photosynthesizing, Porous Edifices Built of Hydrothermallly Precipitated Zinc Sulfide as Cradles of Life on Earth. Biol. Direct, 2009, 4:26. 11.Mulkidjanian AY, Bychkov AY, Dibrova DV, Galperin MY, Koonin EV. Open Questions on the Origin of Life an Anoxic Geothermal Fields. Orig.Life Evol.Biosph.,2012, 42(5):507-16. THEORY OF EVOLUTION ABIOGENESIS IN THE ATMOSPHERE... 12.Zaritsky AR, Grachev VI, Vorontsov YuP, Pronin VS. Energy Aspects of Abiogenesis in the Atmosphere on Hydrocarbon Aerosol Nanodroplets. RENSIT, 2013, 5(2):105-125 (in Russ.). 13.Galimov EM. Fenomen zhizni [The phenomenon of life]. Moscow, URSS Publ., 2006, 254 p. 14.Maryada Sharma, Manni Luthra – Guptasarma. Degradation of protein upon storage at nearneutral Ph: Indications of a proteolytic/ gelatinolytic activity associated with aggregates. Biochimica at Biophysica Acta, 2009, 1790:1282-1294. 15.Nelson DL, Cox MM. Lehninger Principles of Biochemistry. WH Freeman, 2012, 1100 p. 16.Murray RK, Granner DK, Mayes PA, Rodwell VW. (eds.) Harper’s Biochemistry. NY, Appleton&Lange, 1988. 17.Rutten MG. The Origin of Life by Natural Causes. Amsterdam, Elsevier, 1971. РЭНСИТ | 2014 | ТОМ 6 | НОМЕР 1 123