К эволюции основных типов жизненных схем у иксодид (Ixodidae)
advertisement
")
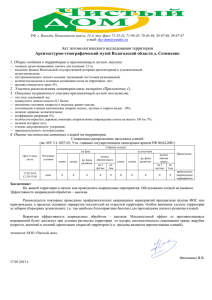
ПАРАЗИТОЛОГИЯ, УДК 576.895.421 XIX, 2, 1985 : 576.1 К ЭВОЛЮЦИИ ОСНОВНЫХ ТИПОВ ЖИЗНЕННЫХ СХЕМ У ИКСОДИД (IXODIDAE) 1 А. Бердыев Использование метода актуализма и сравнительно-исторического метода к экологическим типам жизненных схем иксодовых клещей позволили изложить взгляды на эволюции типов их паразитизма. Отмечается древность гнездово-норовых (начало) и молодость пастбищно-стойловых иксодид (поздний неоген). Пастбищно-подстерегающие и пастбищно-норовые клещи составляют переходные формы и произошли в конце мела и в позднем палеогене соответственно. Вопросы современного взаимоотношения сочленов паразитарной системы «иксодовые клещи—возбудители болезней—позвоночные животные» привлекают внимание многих исследователей. Решение этих сложных вопросов представляется затруднительным, когда недостаточен исторический материал по каждому из сочленов. Сравнительно недавно, после опубликования рецензии Коренберга (1981) на нашу монографию, у нас возникло решение анализировать возможные пути происхождения выделенных типов жизненных схем у иксодид. Эта рецензия наряду с положительным отзывом автора содержит ряд дискуссионных и интересных вопросов по поводу происхождения того или иного типа жизненной схемы у клещей. Наши взгляды на основные направления эволюции паразитизма у иксодовых клещей излагаются в настоящем сообщении. Нами (Бердыев, 1974, 1980) в результате обобщения собственных наблюдений по фенологии и экологии иксодид аридных районов, а также сравнения их с данными литературы пересмотрены типы жизненных схем у иксодид. Кроме известных ранее гнездово-норовых и пастбищно-подстерегающих форм иксодид, были выделены группы клещей, ведущих пастбищно-норовый и пастбищностойловый тип паразитизма. Эти четыре экологических типа иксодид включают в себя все группы иксодид, по меньшей мере фауны Палеарктики. Согласно Померанцеву (1937, 1948), Филипповой (1977), мы (Бердыев, 1980) как исходную форму брали гнездово-норовый тип паразитизма, давший начало в эволюции остальным типам паразитизма у иксодид. Как известно, характеристика исторических изменений типов паразитизма дает возможность истолковать причину и сущность угасания или процветания некоторых групп паразитов, обитающих в экосистемах далекого или позднего геологического прошлого. В связи с этим некоторые направления эволюции групп Ixodidae, в частности их историко-морфологическая дифференцировка, отражены в работах Померанцева (1948), Шульце (Schulze, 1932, цит. по: Померанцев, 1948), Дубинина (1958), Филипповой (1977), Балашова (1982). Из-за отсутствия палеонтологического материала исследователям пришлось воспользоваться сравнительно-морфологическим и функциональным анализом. Вместе с тем паразитологам-экологам приходится судить об эволюции парази1 Материалы этой статьи доложены на заседании IV Всесоюзного акарологического совещания по проблемам изучения паразитических клещей в ноябре 1981 г. в Ленинграде. 434 тизма у отдельных групп паразитов, используя данные, уже полученные в палеозоологии, палеоботанике, палеоклиматологии и др. Реконструкция истории развития позвоночных животных — прокормителей иксодид в экосистемах в прошлых геологических эпохах и сравнение этих данных с результатами, полученными при изучении современных типов жизненных схем (метод «актуализма» по Завадскому и Колчинскому, 1977) позволяют построить научные гипотезы о времени и направлениях эволюции паразитизма у иксодовых клещей. Использование этого подхода в виде экстраполяции на прошлое в единстве со сравнительно-историческим методом проливает свет в познании причин эволюции паразитизма иксодовых клещей, по которым нет палеонтологических данных. Необходимым условием следует считать параллельное изучение клещей и их прокормителей как единой системы паразит—хозяин. Существуют разные взгляды в отношении места происхождения гнездовонорового типа паразитизма у иксодид. Так, Померанцев (1948) писал, что этот тип паразитизма возник гн ПП ПН ПС параллельно и независимо по крайней мере в трех основных центрах формиIV рования: африкано-европейском, центрально-азиатском и североамериканш ском. Дубинин (1958), хотя и допускает вероятность происхождения 1хоdidae в мелу, все же местом происхождения икII содид, по его мнению, Основные направления эволюции паразитизма у иксодид. I I — антропоген, II — неоген, III — палеоген, IV — мел. Типы жизненных схем: Г Н — гнездовоноровый; ПП — пастбищно-подстерегающий; П Н — пастбищноноровый; ПС — пастбищно-стойловый. могли быть тропические страны, а затем уже клещи расселились в более северные и холодные зоны. С этим мнением автора трудно согласиться, когда известно, что даже в палеогене господствовал изотермический климат и существовало только два термических пояса: жаркий тропический и квазитропический (Синицын, 1980) и возникновение арктического и субарктического поясов, четкие границы климатических зон оформились лишь в период четвертичных ледников. Сравнивая морфологический материал по иксодинам — примитивной группе среди иксодид, Филиппова (1977) высказывается в пользу гипотезы о первичности гнездово-норового типа паразитизма. Об этом свидетельствует примитивность опорно-фиксаторных и экологических приспособлений представителей этой группы. Формирование этого типа паразитизма было связано с древними мелкими стационарно-логовищными примитивными животными — однопроходными и сумчатыми, появившимися на земле в меловом периоде. Известно, что по ходу эволюции, как правило, полнота положительных связей паразита с хозяином нарастает (Беклемишев, 1954). Это позволяет предполагать, что прародители современных иксодид, совершенствуясь, устанавливают постепенно закономерные связи с животными, не имеющими постоянных убежищ, т. е. они начинают вести жизненную схему по пастбищно-подстерегающему типу паразитизма. Совершенствование опорно-фиксаторных и экологических приспособлений клещей, связанных с изменениями климата нашей планеты способствовало, очевидно, в дальнейшем закреплению у иксодид (см. рисунок) пастбищно-подстерегающего типа паразитизма. 135 В палеогене (палеоцен, эоцен, ранний олигоцен), когда равномерно изотермический климат преобладал на планете (Синицын, 1965; 1980), вероятно, сформировался и господствовал пастбшцно-подстерегающий тип паразитизма, поскольку при этом климате ни клещам, ни их хозяевам не требовалось убежища. Таким образом, палеоген можно рассматривать как эпоху повсеместного расселения иксодовых клещей. В этот период начинают развиваться плацентарные млекопитающие, которые достигают расцвета в неогене. Ареал пастбищно-подстерегающих клещей, очевидно, занимал обширную территорию суши, включая и современные пустынные территории. Уже в олигоцене в южных областях Азии существовали автохтонные группы землероев, характерные для открытых пространств (Шевырева, 1983). Особенно это процветало в позднем олигоцене. В неогене постепенный переход климата внутренней Азии к аридности способствовал формированию фауны открытых пространств (роющие грызуны, парнокопытные). Азиатская часть Палеарктики считается центром развития и распространения грызунов (Шевырева, 1983). Все это повлекло за собой постепенное формирование паразитизма у иксодид по пастбшцно-норовому типу (см. рисунок). Кроме того, постепенное опустынивание территории способствовало либо вынужденному оттеснению в мезофильные районы клещей с пастбищно-подстерегающим типом паразитизма, обитающим здесь ранее, либо, по мнению Филипповой (1977), — вторичному возврату некоторых видов клещей к гнездово-норовому типу паразитизма. Однако основная часть пастбищно-подстерегающих клещей на таких территориях вынуждена была адаптироваться к условиям и изменить жизненную схему в сторону пастбшцнонорового типа. Есть все основания полагать, что центром возникновения и дальнейшего расселения клещей пастбищно-норового типа явились аридные и открытые территории современной Центральной и Средней Азии. Об этом косвенно свидетельствуют палеонтологические данные: обнаружение в неогеновых отложениях остатков гиппарионовой фауны, парнокопытных, роющих грызунов (Беляева, Трофимов, 1967; Бирман и др., 1971; Горелов, 1972; Шевырева, 1983). В те времена нынешняя территория Туркменистана находилась в давно сложившейся аридной зоне. Аридность территорий сопровождалась возрастанием континентальности климата, сопровождаемой выраженной сезонностью. При этих условиях в неблагоприятных сезонах клещи укрывались в норах, убежищах. Активность неполовозрелых фаз этих клещей приурочена к летнему сезону, когда обилие их прокормителей достаточно высоко. Проявление активности голодных имаго к весеннему и осеннему сезонам также является адаптивным к аридным условиям признаком. Дальнейшая депрессия климата и возрастание континентальности на аридных территориях, очевидно, повлияли на тип жизненной формы прокормителей имаго иксодид — копытных. У предков современных домашних животных, обитающих на открытых пространствах, появились поведенческие приспособительные признаки (стадность, суточный ритм активности и отдыха). В пещерах и естественных понижениях, например под навесами скал, т. е. в местах, защищенных от сырой и холодной погоды зимой и от палящих лучей солнца — летом, стадные копытные устраивали постоянные лежки. В течение длительной эволюции у некоторых форм иксодид, паразитировавших ранее на таких животных, также эволюцинировали жизненные циклы, параллельно приспособленные к новым условиям. Отпадение сытых самок клещей и их дальнейшее размножение происходило уже в лежках — в непосредственной близости от мест отдыха прокормителей; постепенно личинки и нимфы, ранее питавшиеся на норовых млекопитающих, переходили на паразитирование на молодняке копытных. Очевидно, таким образом и шло формирование типа паразитизма современных пастбищно-стойловых клещей. К этой же эпохе можно отнести становление типов развития трех-, двух- и однохозяинных циклов развития у иксодид. Мы считаем, что пастбшцно-стойловый тип паразитизма мог эволюционировать в основном из клещей пастбищно-норового типа паразитизма. Следовательно, центром происхождения этих клещей могли быть открытые пространства аридной территории Центральной и Средней Азии. На остальные современные аридные территории других континентов (например, Африки) паст136 бшцно-стойловые формы иксодид могли расселиться в поздние геологические эпохи. Об этом свидетельствует и то, что в Африке, в том числе на месте современной пустыни Сахары, в послеледниковый период и даже в недавнем прошлом (около 6000 лет назад) существовали сплошные лесные массивы (Гаврилов, 1976; Леме, 1976), что является нехарактерной для пастбищно-стойловых клещей стацией. Вероятность происхождения пастбищно-стойловых клещей от более генерализованного пастбшцно-подстерегающего типа менее убедительна, поскольку, на наш взгляд, тенденция к сокращению числа встреч клещей с хозяином для кровососания могла появиться вслед за аридизацией территории. Учитывая, что в настоящее время пастбищно-стойловые клещи встречаются в основном на аридных территориях, происхождение их от пастбищно-норовых групп клещей вполне очевидно, поскольку ареал последних также совпадает с аридными и семиаридными территориями. Появление человека оказало значительное влияние на эволюцию биосферы в целом. Начало трудовой деятельности представителей рода Homo в четвертичном периоде привело к исчезновению некоторых крупных млекопитающих и птиц. Одомашнивание полезных для человека животных (последние 10— 15 тыс. лет) способствовало еще большему закреплению и прогрессированию пастбищно-стойлового типа паразитизма у иксодовых клещей. Этому во многом способствовала гарантированная обеспеченность клещей источником питания и укрытием: содержание животных в помещениях — стойловое или стойлово-пастбищное содержание скота. Следовательно, современные синантропные стойловые клещи, безусловно, трансформировали из дикой природы ближе к человеку, вместе с одомашненным скотом. При этом трансформация клещей из лежек диких копытных в стойла домашних животных и наоборот, на наш взгляд, не сопровождается изменениями в строении и экологии паразитов. Пастбищно-стойловые клещи, несмотря на кажущуюся некоторым исследователям (Коренберг, 1981) близость по происхождению к гнездово-норовым иксодидам, все же резко отличаются от них прогрессивностью своей экологии: нападение на крупных позвоночных, активность в поисках хозяина, высокая плодовитость и соответственно высокая численность, изменение типа развития в сторону сокращения смены хозяев, агрессивность самцов и копуляция на хозяине. Именно к этой группе отнесены нами однохозяинные клещи, например Hyalomma scupense, Boophilus calcaratus, тип развития которых, очевидно, является завершающей формой в их эволюции (Беклемишев, 1954). Следовательно, однохозяинные иксодовые клещи в антропогене находятся в эволюционном тупике, лишающем этих клещей возможности дальнейшего экологического прогрессирования или возврата к предыдущим жизненным схемам. Все эти факты дают нам основание еще раз утверждать, что пастбищностойловые виды клещей, как самостоятельная жизненная схема у иксодид, произошли вторично, т. е. они произошли от пастбищно-норовых клещей и сложились в течение длительной эволюции задолго до антропогена. Таким образом, исторически более древние и примитивные группы — гнездово-норовые клещи, а пастбищно-стойловые иксодиды представляют собой сравнительно молодую и высокоспециализированную по уровню развития жизненную схему. Литература Б а л а ш о в Ю. С. Паразито-хозяинные отношения членистоногих с наземными позвоночными. Л., Наука, 1982. 318 с. Б е к л е м и ш е в В. Н. Паразитизм членистоногих на наземных позвоночных. Основные направления его развития. — Мед. паразитол. и паразитарные болезни, 1954, т. 29, вып. 1, с. 3—20. Б е л я е в а Е. И., Б. А. Т р о ф и м о в. Основные этапы развития наземных третичных млекопитающих азиатской части СССР. — В кн.: Стратиграфия и палеонтология мезозойских и палеоген-неогеновых континентальных отложений азиатской части СССР. Л., Наука, 1967, с. 209-215. Б е р д. ы е в А. Типы жизненных схем у иксодовых клещей (Parasitiformes, Ixodidae). — Паразитология, 1974, т. 8, вып. 3, с. 227—233. Б е р д ы е в А. Экология иксодовых клещей Туркменистана и их роль в эпизоотологии природно-очаговых болезней. Ашхабад, Ылым, 1980. 281 с. 4 П а р а з и т о л о г и я , № 2, 1985 г. 137 Б п р м а н А. С., В. И. Ж и г а й л о, Jl. М. Р а с ц в е т а е в, Л. И. X о з а ц к и й г Н. С. Ш е в ы р е в а. О находке плиоценовых позвоночных в молассовых отложениях Восточного Копетдага. — Бюл. МОИП, отд. геологии, т. 46, вып. 2, 1971, с.99—106. Г а в р и л о в В. П. Путешествие в прошлое земли. М. Недра, 1976. 144 с. Г о р е л о в Ю. К. К формированию фауны наземных позвоночных. — Бюл. МОИП, отд. биол., 1972, т. 17, вып. 6, с. 27—37. Д у б и н и н В . Б. К вопросу о происхождении и развитии типов жизненных циклов у иксодовых клещей.— Изв. Казан, филиала АН СССР. Сер. биол. наук., 1958, № 6, с. 95— 126. З а в а д с к и й К. М.,Э. И. К о л ч и н с к и й . Эволюция эволюции. Историко-критические очерки проблемы. Л., Наука, 1977. 235 с. К о р е н б е р г Э . И. Рец. на кн.: А. Бердыев. Экология иксодовых клещей Туркменистана и их роль в эпизоотологии природно-очаговых болезней. Ашхабад, 1980. 282 с. — Мед. паразитол. и паразитарные болезни, 1981, № 3, с. 85—87. Л е м е Ш. Основы биогеографии. —М., Прогресс, 1976. 308 с. П о м е р а н ц е в Б . И . О паразитических адаптациях Ixodoidea (Acarina). — Изв. АН СССР Сер. биол. наук, 1937, № 4, с. 1423—1437. П о м е р а н ц е в Б . И . Географическое распространение клещей Ixodoidea и состав их фауны в Палеарктической области. — Тр. ЗИН АН СССР, 1948, т. 7, вып. 3, с. 132—148. С и н и ц и н В . М., Древние климаты Евразии. Часть I. Л., Изд. ЛГУ, 1965, 167 с. С и н и ц и н В . М . Природные условия и климаты территории СССР в раннем и среднем кайнозое. Л., Изд. ЛГУ, 1980. 104 с. Т и м о ф е е в - Р е с о в с к и й Н. В., Н . Н . В о р о н ц о в , А. В. Я б л о к о в. Краткий очерк теории эволюции М., Наука, 1977. 301 с. Ф и л и п п о в а Н. А. Иксодовые клещи подсем. Ixodinae. — Фауна СССР. Паукообразные. Т. 4, вып. 4. Л., Наука, 1977. 396 с. Ш е в ы р е в а Н . С. Грызуны (Rodentia, Mammalia) неогена Евразии и Северной Африки — эволюционная основа плейстоценовой и современной фауны грызунов (неоген-современность). М., Наука, 1983, с. 9—145. Институт зоологии АН ТССР, Ашхабад Поступило 2 XI 1983* ON THE EVOLUTION OF THE MAIN TYPES OF LIFE SCHEMES IN (IXODIDAE) IXODIDS A. Berdyev SUMMARY The burrow-nest life scheme of ixodid ticks, which was formed at the beginning of the Chalk, gave rise to the subsequent types of parasitism: pasture (end of the Chalk), pastureburrow (end of Paleogene) and pasture-stall one (late Neogene). Changes in the climate in different geological epochs and regions together with the hosts' evolution, natural selection in the aggregate of variability and heredity of ixodids in their populations were the cause of the evolution.