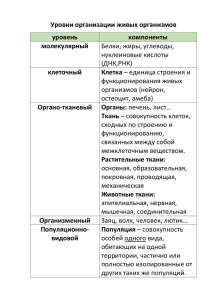
популяционно-онтогенетические исследования
advertisement

РОССИЙСКИЙ ФОНД ФУНДАМЕНТАЛЬНЫХ ИССЛЕДОВАНИЙ МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РЕСПУБЛИКИ МАРИЙ ЭЛ КОМИТЕТ ЭКОЛОГИИ И ПРИРОДОПОЛЬЗОВАНИЯ Г. ЙОШКАР-ОЛЫ ФГБОУ ВПО «МАРИЙСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ» НАЦИОНАЛЬНЫЙ ПАРК «МАРИЙ ЧОДРА» ФГУ ГПЗ «БОЛЬШАЯ КОКШАГА» МАРИЙСКОЕ ОТДЕЛЕНИЕ ОБЩЕСТВА ФИЗИОЛОГОВ РАСТЕНИЙ ПРИНЦИПЫ И СПОСОБЫ СОХРАНЕНИЯ БИОРАЗНООБРАЗИЯ М А ТЕ Р И А Л Ы V Международной научной конференции 9–13 декабря 2013 года Часть I Йошкар-Ола 2013 ББК 28.0:20.1 УДК 57:502.172 П 75 Ответственные редакторы: О. Л. Воскресенская, д-р биол. наук, проф., зав. кафедрой экологии, и.о. декана биолого-химического факультета МарГУ; Л. А. Жукова, д-р биол. наук, проф. МарГУ, заслуженный деятель науки РФ Редакционная коллегия: А. Л. Азин, д-р мед. наук; И. М. Божьеволина, канд. пед. наук; И. А. Гетманец, д-р биол. наук; Н. В. Глотов, д-р биол. наук; Ю. А. Дорогова, канд. биол. наук; В. П. Ившин, д-р хим. наук; В. А. Забиякин, д-р с.-х. наук; Г. О. Османова, д-р биол. наук; Т. А. Полянская, канд. биол. наук: Н. П. Савиных, д-р биол. наук Р е ц е н з е н т ы : Р. И. Винокурова, д-р биол. наук, профессор Поволжского государственного технологического университета; В. М. Пахомова, д-р биол. наук, профессор Казанского государственного аграрного университета. Печатается при поддержке ФГБУ «Российский фонд фундаментальных исследований» (грант № 13-04-06122) Принципы и способы сохранения биоразнообразия: матеП 75 риалы V Международной научной конференции: в 2 ч. / Map. гос. ун-т. – Йошкар-Ола, 2013. – Часть I. – 324 с. ISBN 978-5-94808-795-5 В материалах конференции представлены доклады, посвященные проблемам биоразнообразия на организменном, популяционном и биоценотическом уровнях. Подробно рассмотрены разнообразие жизненных форм, таксономическое, экологическое и структурное биоразнообразие популяций и сообществ на особо охраняемых и нарушенных территориях, физиолого-биохимические адаптации организмов к различным экологическим факторам, медико-биологические аспекты использования биоразнообразия. Показаны математические аспекты анализа биоразнообразия; приведены методы мониторинга абиотических и биотических компонентов экосистем. Особое внимание уделено проблеме формирования экологического мировоззрения школьников и студентов. Для экологов, биологов, специалистов в области охраны природы и рационального природопользования, а также преподавателей и студентов (бакалавров и магистров) биологических и экологических специальностей вузов, учителей и школьников. ББК 28.0:20.1 УДК 57:502.172 © ФГБОУ ВПО «Марийский государственный университет» ISBN 978-5-94808-795-5 2 RUSSIAN FOUNDATION FOR BASIC RESEARCH MINISTRY OF EDUCATION OF THE MARI EL REPUBLIC ECOLOGY AND NATURAL RESOURCES COMMITTEE OF THE CITY OF YOSHKAR-OLA MARI STATE UNIVERSITY MARI CHODRA NATIONAL PARK BOLSHAYA KOKSHAGA STATE NATURE RESERVE MARI EL BRANCH OF THE SOCIETY OF PLANT PHYSIOLOGISTS PRINCIPLES AND METHODS OF BIODIVERSITY CONSERVATION P R O C E E D IN G S of the Fifth International Research Conference 9-13 December 2013 Part I Yoshkar-Ola 2013 3 BBK 28.0:20.1 UDK 57:502.172 P75 Editors-in-Chief: О. L. Voskresenskaya, Dr. Biol. Sciences, Professor, Head of the Department of Ecology, Acting Dean of the Faculty of Biology and Chemistry of Mari State University L. А. Zhukova, Dr. Biol. Sciences, Professor of Mari State University, Honored Scientist of the Russian Federation. Editorial Board: A. L. Azin, Dr. Med. Sciences; I. M. Bozhjevolina, Cand. Ed. Sciences; I. A. Getmanets, Dr. Biol. Sciences; N. V. Glotov, Dr. Biol. Sciences; Y. A. Dorogova, Cand. Biol. Sciences; V. P. Ivshin, Dr. Chem. Sciences; V. A. Zabiyakin, Dr. Agricult. Sciences; G. O. Osmanova, Dr. Biol. Sciences; T. A. Polyanskaya, Cand. Biol. Sciences; N. P. Savinykh, Dr. Biol. Sciences. R e v i e w e r s : R. I. Vinokourova, Dr. Biol. Sciences, Professor of Volga State University of Technology; V. M. Pakhomova, Dr. Biol. Sciences, Professor of Kazan State Agrarian University. Published with financial support of the Russian Foundation for Basic Research (grant № 13-04-06122) P 75 Principles and methods of biodiversity conservation: Proceedings of the Fifth International Research Conference / Mari State University. – Yoshkar-Ola, 2013. – Part I. – 324 p. ISBN 978-5-94808-795-5 The conference proceedings cover the issues of biodiversity at the organismal, population and biocenotic levels. Careful consideration is given to the variety of life forms, taxonomic, ecological and structural biodiversity of populations and communities in protected and disturbed areas, physiological and biochemical adaptation of organisms to different environmental factors, medical and biological aspects of biodiversity. Mathematical aspects of the analysis of biodiversity and methods of the monitoring of abiotic and biotic components of ecosystems are presented. Special attention is given to the shaping of school and university students’ ecological worldviews. Targeted at ecologists, biologists, other professionals in the field of environmental protection and rational nature management; university teachers and students (undergraduate and graduate) of biological and environmental majors, as well as at school teachers and students. BBK 28.0:20.1 UDK 57:502.172 © Mari State University, 2013 ISBN 978-5-94808-795-5 4 ПРЕДИСЛОВИЕ Пятая Международная научная конференция «Принципы и способы сохранения биоразнообразия» посвящена Году охраны окружающей среды в России. На конференции будут заслушаны доклады по актуальным вопросам исследования таксономического, экологического и структурного биоразнообразия наземных и водных экосистем, принципам и методам его изучения и сохранения. В рамках конференции представлено десять секций и одна подсекция. С е к ц и я 1 . Биоразнообразие жизненных форм живых организмов в наземных и водных экосистемах. С е к ц и я 2 . Таксономическое и структурное биоразнообразие на особо охраняемых и нарушенных территориях. С е к ц и я 3 . Мониторинг абиотических и биотических компонентов экосистем. С е к ц и я 4 . Популяционное биоразнообразие и устойчивость популяций. С е к ц и я 5 . Экологические и физиолого-биохимические адаптации организмов для сохранения биоразнообразия. С е к ц и я 6 . Медико-биологические аспекты использования биоразнообразия и здоровье населения. С е к ц и я 7 . Здоровый образ жизни – основа устойчивости ноосферы. С е к ц и я 8 . Рациональное использование биоразнообразия живых организмов человеком; экологическая биотехнология. С е к ц и я 9 . Математические аспекты анализа биоразнообразия. С е к ц и я 1 0 . Формирование экологического сознания – основной путь сохранения биоразнообразия экосистем Земли в XXI веке. Подсекция «Молодежные экологические инициативы». Материалы конференции изданы в 2 книгах: в первую книгу вошли пленарные доклады и материалы секций (с 1 по 4); во вторую книгу – материалы докладов 5–10 секций. Во время проведения конференции организованы выставки: а) книг по биоразнообразию, выпущенных участниками конференции за последние 10 лет; б) онтогенетического гербария, отражающего внутрипопуляционное разнообразие; 5 в) молодежных работ студентов и школьников. Организаторами конференции являются: Российский фонд фундаментальных исследований, ФГБОУ ВПО «Марийский государственный университет», Министерство образования и науки Республики Марий Эл; Комитет экологии и природопользования г. Йошкар-Олы, Национальный парк «Марий Чодра», ФГУ ГПЗ «Большая Кокшага», Марийское отделение Общества физиологов растений. Для публикации в сборнике представлено около 200 докладов более чем от 300 участников конференции, из которых 34 – представители зарубежных вузов и научно-исследовательских центров. Содержание материалов конференции одобрено программным комитетом и редакционной коллегией. По возможности проверена или введена бинарная номенклатура, устранены технические погрешности, не изменяющие сущности текстов. Ответственность за научное содержание статей несут их авторы. Редакционная коллегия выражает признательность за рецензирование материалов конференции доктору биологических наук, профессору Р. И. Винокуровой и доктору биологических наук, профессору В. М. Пахомовой. Оргкомитет конференции благодарит и. о. ректора МарГУ доктора экономических наук М. Н. Швецова за помощь в организации и проведении конференции, доцента МарГУ Е. В. Сарбаеву за техническую подготовку материалов к печати, начальника РИЦ Л. Н. Еремееву, начальника ООП С. А. Филонова, а также литературных редакторов О. С. Березину, О. А. Егошину, оператора компьютерной верстки С. Н. Бастракову, и всех, кто трудился над изданием материалов конференции. 6 ПЛЕНАРНЫЕ ДОКЛАДЫ ПЕРВИЧНОЕ ЗДОРОВЬЕ ЧЕЛОВЕКА И ВЕРОЯТНОСТЬ ФЕНОМЕНА «СТОП-ЭВОЛЮЦИЯ» НА УРОВНЕ HOMO SAPIENS Азин А. Л. Республиканский клинический госпиталь ветеранов войн, г. Йошкар-Ола, azin_rbvv@mail.ru Казанский государственный медицинский университет, г. Казань, kaf_kgmu_rme@mail.ru Марийский государственный университет, г. Йошкар-Ола Эта статья носит дискуссионный характер, так как в ней сопоставляются спорные, т. е. оптимистические и пессимистические концепции современной геронтологии по проблеме эволюционной судьбы человека как биологического вида. История знает немало примеров полного исчезновения видов на Земле... В фундаментальной геронтологии общепринято считать, что видовой биологический предел продолжительности жизни человека должен достигать 120 лет и более. По мнению одного из основателей геронтологии – И. И. Мечникова (1908), для достижения этого предела необходимо каждому индивидууму следовать жизненным принципам ортобиоза (здорового образа жизни). Традиционная демографическая доктрина утверждает, что к 2050 г. численность пожилых людей увеличится в 2–3 раза, поскольку научнотехнический прогресс будет улучшать условия для увеличения продолжительности жизни людей. В научной печати уже появились скептические прогнозы, противоречащие официальным. Предполагается, что люди, родившиеся во второй половине XX века, в основном не будут доживать до пенсионного возраста, и кривая «демографического бума» пойдет вниз, продолжительность жизни людей будет уменьшаться (Один, 2011). В чем причина геронтологического кризиса? Цитируемый автор полагает, что это – катастрофическое ухудшение качества первичного здоровья у лиц, рожденных в конце XX века, вследствие действия факторов «стоп-эволюция человека» и кризиса среды обитания. Первичное здоровье – это совокупность врожденной предрасположенности к заболеваниям с врожденным резервом саногенных процессов. На сегодняшний день отсутствие болезни не отражает истинного здоровья человека, поскольку в результате успехов медицины, сниже7 ния детской смертности и выживания лиц с наследуемыми хроническими заболеваниями и, как следует, отмены законов эволюции, а также изза комплекса факторов: социальных, антропогенных и техногенных, действующих на человека еще до рождения само здоровье первично стало другим (на герминальном, эмбриональном и фетальном уровнях). Можно сказать – это весьма перспективное направление в изучении качества и перспектив жизни каждого человека, пока, к сожалению, мало распространенное в медицинской практике. Оно созвучно известной американской шутке: «хочешь долго прожить, выбирай себе здоровых родителей...» и только. Мне кажется, что общепринятое понятие ВОЗ общественное здоровье носит скорее всего социально-политический, а не медицинский характер. Предполагается, что в индустриальную эпоху, в особенности с развитием «социальной индустрии», появились антропогенные, социальные и техногенные факторы, практически отменившие эволюционные законы существования в природе вида Homo sapiens! Посмотрим, так ли это. К числу основных причин, отменяющих классические законы эволюционного развития человека, предлагается отнести: 1. Коммерциализацию и «индустриализацию» медицины. Действительно, во второй половине ХХ в. медицина, к сожалению, превращается в высокодоходный бизнес. Появилась «неодеонтология», которая для решения рыночных интересов разработала идеологию доказательной медицины (основанной на «уликах»), формально научно-экономической теории – по сути «рыночно-полицейской идеологии» (Один, 2011). Мирное сосуществование «пациент – врач – фармацевт», по мнению автора, стало возможным только под неусыпным оком закона. Канула в Лету «доверительная медицина», когда врач принимает решение, сообразуясь со своим клиническим опытом и его школой, лечит за благодарность, а не за профицит. Далее, начиная с 50-х гг. после открытия антибиотиков, произошло снижение общей и детской смертности. Поэтому в конце века основными причинами смертности стали «нормальные болезни» по В. М. Дильману (1987) – атеросклероз и его последствия, сахарный диабет и метаболический синдром, рак. Но и здесь были приложены значительные медицинские усилия по выживанию лиц с названными хроническими заболеваниями и появлению у них потомства, а это приводит к закономерному накоплению болезней у потомков, причем с опережающим дебютом. Наконец, в современной медицине стал изучаться «эмбриональный стресс», вызванный частыми УЗ-исследованиями в период беременности. Согласно стандартам в России принято 3-кратное скрининговое УЗИ всех беременных на 10–14, 20–24, 32–34 неделях. При этом уста8 новлен шум мощностью до 100 дБ (!) от вторичной вибрации стенки матки, который приводит к усиленным движениям плода во время УЗИ. Неврологические и психические последствия этого воздействия пока мало изучены и противоречивы. Тем не менее, огромное количество женщин стремятся на платной основе проходить дополнительно УЗИ, не задумываясь над возможными неврологическими последствиями для будущего ребенка. 2. Феминизм или эмансипация. Оказалось, что с развитием техники и уменьшением мускульного труда женщина оказалась равной и самостоятельной добытчицей материальных благ. Реализовалась идея Фрейда уровнять секс и любовь в ущерб браку. В итоге вместо «трех К» (Kirche, Kinder, Kuche) в мыслях у современных женщин появилось «два К» (карьера и «клубничка»). Соответственно, уменьшилось число родов и детей, вступающих в эволюцию, и резко увеличилось число генетически скомпрометированных новорожденных. Другим следствием эмансипации стало уклонение от материнства и сдача детей в сиротские приюты. Исследования показали, что распространенное курение женщин как символ равенства полов способствует эстрогендефициту, андрогенизации, интоксикации, ановуляции, снижению репродуктивной способности и увеличению хромосомной патологии плода. 3. К сожалению, в условиях современной либерально-гедонистической морали стал активно пропагандироваться образ гомосексуала. По данным В. В. Кон (1988), явных гомосексуалов в популяции рождается не более 5 %, однако из-за массовой пропаганды однополой любви число латентных гомосексуалов достигает 25 %, естественно, можно согласиться, что это явление не способствует повышению рождаемости (фертильности) в популяции человека. 4. Вот и психологический аспект. В ХХ в. было отменено раздельное обучение мальчиков и девочек. В этих условиях от ребенка стали требовать унифицированного стандарта поведения преимущественно феминного образца, привычного для педагогов-женщин (их большинство). В образовательной системе произошла гендерная дезинтеграция. Так, для мальчиков, к тому же психофизиологически отстающих от девочексверстниц, школа стала источником хронического стресса. Наиболее подробно проблему школьного стресса изучил В. Ф. Базарный (2004), который выделил 7 дидактических стрессов. Юноши и девушки, не прошедшие основополагающие обряды гендерного поведения, гигиены души и тела в семье и школе, превращаются в сообщество инфантильных секс-партнеров. Как следствие, число разводов превышает число браков, резко падает рождаемость, что ведет к феномену «стопэволюции». 9 5. Другой феномен – психоэмоциональный стресс. Более 50 % населения проживает в городах и подвергается стрессорному воздействию урбанизации. Психику современного человека агрессивно фрустрируют СМИ с целью манипуляции общественным сознанием. Используется прием индуцирования страха путем негативации новостей, разжигания чувства недовольства рекламой, громкой ритмичной музыкой в кафе и маркетах. Можно привести и ряд других, в основном, антропогенных факторов воздействия, негативно влияющих как на первичное, так и вторичное здоровье человека. Однако в заключении вернемся к главному вопросу – возможен ли феномен «стоп-эволюция» человека как вида? Ответ на этот вопрос неоднозначен. Если задуматься, то вероятность его формирования высока. Однако, если проанализировать негативное воздействие перечисленных, а также других факторов на первичное здоровье человека, то можно предположить, что эти факторы в принципе контролируемы и управляемы, даже на генетическом уровне. Следовательно, кризис геронтологии невозможен, и каждый человек сможет достигнуть видовой продолжительности жизни. Л и т е р а т ур а Базарный В. Ф. Школьный стресс и демографическая катастрофа в России / В. Ф. Базарный. – Сергиев Посад: Весь, 2004. – 96 с. Дильман В. И. Четыре модели в медицине / В. И. Дильман. – Л.: Медицина, 1987. – 356 с. Кон И. С. Введение в сексологию / И. С. Кон. – М.: Медицина, 1988. – 336 с. Мечников И. И. Этюды оптимизма / И. И. Мечников. – М.: Наука, 1964. – 328 с. Один В. И. Кризис геронтологии: к вопросу о первичном здоровье в XX веке / В. И. Один // Успехи геронтологии. – 2011. – Т. 24, № 1. – С. 11–23. ОХРАНА ВИДОВОГО РАЗНООБРАЗИЯ РАСТЕНИЙ НА ОСОБО ОХРАНЯЕМЫХ ПРИРОДНЫХ ТЕРРИТОРИЯХ В УДМУРТСКОЙ РЕСПУБЛИКЕ Баранова О. Г. Удмуртский государственный университет, г. Ижевск, ob@uni.udm.ru Существуют разные способы сохранения фиторазнообразия, но основным способом его охраны на региональном уровне является заповедование экосистем с последующим слежением за режимом резервации. В связи с этим, необходимо создание в регионах экологически обоснованной и реально действующей системы функционально взаимосвязанных особо охраняемых природных территорий (ООПТ). В связи с этим основными объектами для создания ООПТ разного уровня должны являться, прежде всего, сообщества и ландшафты, имеющие свойства устойчивости и самовосстановления. 10 История формирования сети ООПТ в отдельных регионах России различна, но, наверное, есть и такие, где картина ее формирования сходна с Удмуртской Республикой (УР). Здесь создание единой республиканской сети ООПТ имеет длительную и, к сожалению, не завершенную историю (Особо…, 2002). В 1995 г. была принята схема единой сети ООПТ Удмуртии Постановлением Правительства УР «О схеме ООПТ Удмуртской Республики» № 377 от 18 декабря 1995 г. Схема ООПТ в УР включала 349 объектов различной категории, общей площадью 401,8 тыс. га (около 9 % территории республики). Несмотря на высокий уровень теоретического обоснования схемы, она не обладала правовой основой для придания ей юридического статуса. В схему вошло большое количество достопримечательностей из раннего, стихийно сложившегося списка, не имеющих необходимой научной, организационно-административной и картографической основы (научные критерии выделения, учетные карточки и паспорта природных достопримечательностей, охранные обязательства, картографическая привязка и др.) (Особо…, 2002). С целью сохранения большего числа представителей Красной книги УР в 2005 г. начаты работы по переинвентаризации ООПТ, которые завершились в 2011 г. Данный этап по созданию единой сети ООПТ УР был направлен и на нормативно-правовое обоснование принятой схемы, придание ей официального юридического статуса и изменения ее в соответствии с новыми концепциями, связанными с упором на сохранение биоразнообразия в природных сообществах и разнообразия природных сообществ. Переинвентаризация показала, что ряд ООПТ, ранее входивших в локальные сети административных районов УР, не существует по целому ряду причин, в том числе связанных с хозяйственной деятельностью человека. На основе комплексных научных исследований видового состава биоты территории республики, растительности, водных объектов, ландшафтов и других элементов природных комплексов ООПТ рекомендовано создание новых локальных сетей во всех 25 районах республики. В новый перечень включены как вновь предлагаемые, так и ранее законодательно утвержденные или рекомендованные к созданию ООПТ (Особо…, 2002; Редкие…, 2011). В результате рекомендовано снижение численности ООПТ до 218, которые имеют разное значение, профиль и площади. Для всех предложенных в локальную сеть комплексных ООПТ выявлен видовой состав биоты, распределение видов биоты, занесенных в Красные книги УР и РФ по территории, оценена сохранность флористических комплексов и животного населения различных биотопов (Баранова и др., 2006а, 2006б; Баранова, 2009; Редкие…, 2011 и др.). 11 Кроме памятников природы, на территории Удмуртии имеются ООПТ других категорий. Самыми крупными, ныне существующими, ООПТ на территории УР являются национальный парк «Нечкинский» (20,7 тыс. га), природные парки «Шаркан» (16,5 тыс. га) и «УстьБельск» (1,77 тыс. га). Достаточно большие площади в республике занимают государственные природные заказники, среди них наибольшую площадь имеют охотничьи заказники (294,4 тыс. га, или 6,3 % от общей площади УР), тогда как ботанических заказников только 2 – «Кокманский» (1,82 тыс. га) и «Андреевский сосновый бор» (0,9 тыс. га). Это территории, которые утверждены на правительственном уровне, все остальные ООПТ в настоящее время правового статуса не имеют. За весь период переинвентаризации не было проведено работ по утверждению их на республиканском уровне, для ряда местных ООПТ были приняты постановления на уровне глав администраций районов, и они имеют законодательную основу. Процесс заповедания уникальных природных территорий УР упирается в невозможность решения 2 вопросов: постановку данных территорий на кадастровый учет с их межеванием (отсутствие финансирования на многомиллионные работы) и нежелание землепользователей изменять функциональность территорий с приданием ранга ООПТ (большая часть этих территорий является лесопокрытыми площадями, находящими в активной эксплуатации Министерством лесного хозяйства УР). В настоящее время в Удмуртии из более чем 300 редких видов сосудистых растений законодательно охраняется 145 видов (Красная книга УР, 2012), из них 9 видов относятся к исчезнувшим на территории республики. Кроме того, в УР произрастает 13 видов сосудистых растений, занесенных в Красную книгу РФ (2008). В результате проведенных исследований в 2005–2011 годах было выявлено, что на ООПТ Удмуртии, по схеме 1995 г., произрастает 73 вида растений (54 %), занесенных в Красную книгу УР (2012). В результате планирования новой сети ООПТ в УР планируется охрана 114 видов, что составляет 84 % от общего числа «краснокнижных» растений. Наибольшее число редких видов находится под охраной на территории национального парка «Нечкинский» – 34 вида растений. К природоохранным организациям, призванным решать многие задачи, связанные с охраной природы, относятся ботанические сады. В 2005 году были начаты работы по созданию интродукционных площадок для выращивания представителей местной флоры Удмуртии в Учебном ботаническом саду Удмуртского государственного университета (УБС УдГУ), в том числе редких и исчезающих растений. За период с 2005 по 2012 гг. в интродукционном эксперименте участвовало 12 68 видов редких растений, и 10 видов были реинтродуцированы в естественные сообщества на территории ботанического сада. Кроме этого, УБС УдГУ совместно с кафедрой ботаники и экологии УдГУ проводит работы по клональному микроразмножению редких травянистых растений Удмуртии. В разные годы объектами исследования стали 16 видов, включенных в Красную книгу УР (2012), такие как Aster amellus L., Dianthus borbasii Vandas, Hedysarum alpinum L. и др. (Баранова и др., 2010; Баранова и др., 2011а, 2011б). Для регионов, где вопросы сохранения видового и ландшафтного разнообразия стоят достаточно остро, актуальным является интегрирование наиболее уникальных природных участков в систему Панъевропейской экологической сети. Изумрудная сеть ООПТ международного значения покрывает фактически весь континент: сюда входят страны Евросоюза и Восточной Европы и других регионов. С территории УР в настоящее время включено в Изумрудную сеть 45 объектов, которые отвечают всем ее требованиям (Удмуртская Республика, 2013). Это, возможно, позволит обратить внимание администрации региона на уникальность природных территорий в пределах республики и необходимость разработки мероприятий по их охране. Исследования проведены при финансовой поддержке Министерства природных ресурсов и охраны окружающей среды Удмуртской Республики и финансирования государственного задания на выполнение НИР в ФГБОУ ВПО «УдГУ» № 4.4599.2011. Л и т е р а т ур а Баранова О. Г. Видовой состав растений некоторых особо охраняемых природных территорий Удмуртской Республики / О. Г. Баранова // Принципы и способы сохранения биоразнообразия: сб. материалов II Всерос. науч. конф. – Йошкар-Ола, 2006а. – С. 67–68. Баранова О. Г. Концептуальные основы охраны редких растений в Удмуртской Республике / О. Г. Баранова // Научные исследования как основа охраны природных комплексов заповедников и заказников: материалы Всерос. науч.-практ. конф. – Киров, 2009. – Вып. 1. – С. 15–19. Баранова О. Г. Стратегия создания и сохранения коллекционного фонда редких и исчезающих растений в Ботаническом саду Удмуртского университета / О. Г. Баранова, О. Н. Дедюхина, О. В. Яговкина // Вестн. Удм. ун-та. Сер., Биология. Наука о земле. – 2010. – Вып. 2. – С. 48–54. Баранова О. Г. Создание генетического банка семян редких растений в ботаническом саду Удмуртского университета / О. Г. Баранова, О. Н. Дедюхина, О. В. Яговкина // Науч. ведомости БелГУ. Сер., Естественные науки. – 2011а. – № 9 (104), вып. 15/2. – С. 145–150. Баранова О. Г. Опыт выращивания редких растений в ботаническом саду Удмуртского университета / О. Г. Баранова, О. Н. Дедюхина, О. В. Яговкина // Вестн. Иркут. гос. сельскохоз. акад. – 2011б. – С. 13–19. Красная книга Российской Федерации: Растения и грибы. – М.: Тов. науч. изд-во КМК, 2008. – 854 с. Красная книга Удмуртской Республики / отв. ред. О. Г. Баранова. – Чебоксары: Перфектум, 2012. – 458 с. Особо охраняемые природные территории Удмуртской Республики: сб. / под ред. Н. П. Соловьёвой; сост. О. Г. Баранова, А. Г. Илларионов. – Ижевск, 2002. – 211 с. Редкие и исчезающие виды растений и животных южной половины Удмуртии и их охрана: итоги науч. исслед. (2005–2009 гг.) / О. Г. Баранова, 13 Д. А. Адаховский, А. Г. Борисовский [и др. ]; под общ. ред. О. Г. Барановой. – Ижевск: Удмурт. ун-т, 2011. – 271 c. Удмуртская Республика / О. Г. Баранова, С. В. Дедюхин, Ю. А. Тюлькин и др. // Изумрудная книга Российской Федерации. Территории особого природоохранного значения Европейской России. Предложения по выявлению / ред. тома Н. А. Соболев, Е. А. Белоновская. – М.: Институт географии РАН, 2011–2013. – Ч. 1. – С. 146–162. ПОПУЛЯЦИОННО-ОНТОГЕНЕТИЧЕСКИЕ ИССЛЕДОВАНИЯ И ПРОБЛЕМА СОХРАНЕНИЯ БИОРАЗНООБРАЗИЯ Жукова Л. А.1, Нотов А. А.2 1 Марийский государственный университет, г. Йошкар-Ола, pinus9@mail.ru 2 Тверской государственный университет, г. Тверь, anotov@mail.ru Катастрофический уровень исчезновения видов в разных биомах нашей планеты позволяет считать задачу сохранения биоразнообразия одной из глобальных проблем современности. Для ее решения необходимо проведение разноплановых исследований и реализация специальных программ и проектов. Анализ биоразнообразия и мониторинг состояния природных комплексов должен осуществляться как минимум на четырех уровнях: экосистемном, ценотическом, популяционном и организменном. При этом популяционный уровень является основополагающим для понимания закономерностей организации живых систем ценотического и экосистемного уровней, что неоднократно отмечали разные исследователи (Юрцев, 1987, 1988; Смирнова и др., 1990, 1993; Жукова, 1995, 2008, 2012б; Комаров, 2006; Злобин, 2009; Злобин и др., 2013 и др.). В рамках конвенции ЮНЕСКО о сохранении биологического разнообразия Земли в 2001 г. была принята Национальная стратегия сохранения биоразнообразия России. В ней также отмечена значимость популяционного подхода для решения основных задач стратегии. Среди них: 1) сохранение и восстановление численности и ареалов природных популяций представителей всех царств живых организмов; 2) сохранение разнообразия их популяционной структуры (половой, возрастной, онтогенетической, виталитетной, пространственной и т. д.). Методические основы анализа популяций редких видов растений разработаны достаточно детально (см. Злобин и др., 2013). Однако в практической деятельности по сохранению биоразнообразия популяционно-онтогенетический подход применяется достаточно редко или возможности его используются не полностью. Необходимо осознание того, что только укрепление позиций и дальнейшее развитие популяционной экологии поможет повысить эффективность такой деятельности. Актуально также выяснение основных направлений популяционно-онтогенетических исследований, которые будут способ14 ствовать более глубокому пониманию закономерностей организации фитоценозов и механизмов поддержания их устойчивости. В настоящее время в рамках популяционно-онтогенетического подхода, основоположниками которого были Т. А. Работнов и А. А. Уранов, сформировалось самостоятельное направление научных исследований, решающее различные прикладные и фундаментальные задачи. Описаны онтогенезы более 2000 видов сосудистых растений и некоторых лишайников (см. Онтогенетический атлас…, 1997–2011; Жукова, 2001б; Жукова и др., 2012б; Ведерникова, Жукова, 2005 и др.). Это направление позволяет полно характеризовать структуру популяций, оценивать их жизнеспособность, выявлять их роль в функционировании фитоценозов, прогнозировать основные тенденции развития, разрабатывать рекомендации по контролю численности популяций в составе разных экосистем (Жукова, 1995, 2012б; Злобин, 2009 и др.). Все это дает возможность объективно оценивать состояние популяций редких и исчезающих видов, перспективы их сохранения (Жукова, 1995; Злобин и др., 2013). Подобный подход должен стать основополагающим в природоохранной деятельности. С этих же позиций следует проводить и анализ инвазионных видов, которые представляют реальную угрозу природному биоразноообразию. Актуально проведение школ, совещаний, конференций с более широким участием специалистов в области охраны и рационального природопользования, флористов, геоботаников, которые будут способствовать более активному использованию популяционно-онтогенетического подхода при решении практических задач (Восточноевропейские…, 2004). Необходимо продолжение исследований, связанных с анализом популяционных характеристик и индексов, которые могут быть использованы для оценки уровня стабильности популяций и выяснения направлений ее динамики. Целесообразно более широкое применение индекса замещения – Iз (показатель отношения прегенеративной фракции ко всей взрослой части ценопопуляции (ЦП)) (Жукова, 1995). Этот индекс позволяет определить тип ЦП и прогнозировать дальнейшие перспективы ее развития, выявлять угасающие ЦП (Iз = 0), нестабильные (Iз < 1), перспективные (Iз > 1) ЦП (Полянская, Жукова, 2013). Актуален поиск других индексов и показателей. На наш взгляд, следует также обратить внимание на некоторые направления исследований, представляющие интерес для дальнейшего развития популяционной экологии и биоценологии. Их результаты имееют концептуальное значение для понимания популяционных механизмов функционирования биологических систем ценотического уровня, поэтому оказываются значимыми для решения проблемы сохранения биоразнообразия. Ключ для понимания закономерностей развития 15 биологических систем разного уровня дает концепция поливариантности развития (Жукова, 1986, 2001а, 2010, 2012а; Жукова, Комаров, 1990; Поливариантность…, 2006). В настоящее время систематизирован огромный фактический материал, описаны основные типы поливариантности (Жукова, 2001а; 2008; 2010; 2012а). Поливариантность обеспечивает гетерогенность популяций и адаптационную пластичность, которая является важным механизмом поддержания устойчивости живых систем надвидового уровня. Однако нередко в биоморфологических, экологических, фитоценотических исследованиях поливариантности развития не придают должного значения, хотя это явление в конечном итоге обеспечивает устойчивость фитоценозов в меняющихся условиях среды и при антропогенном стрессе. Актуально изучение поливариантности у представителей разных групп и царств живых организмов и поиск ее новых форм и типов. Теперь мы достаточно информированы о паразитах, сапрофитах, микоризе, понимая их как результат симбиогенеза. Однако четкая картина взаимодействия представителей всех царств и уровней пока не появилась, т. к. почти всегда эти явления рассматриваются в статике. Разработанная концепция дискретного описания онтогенеза растений дает возможность описывать реальные связи организмов на разных этапах их развития. Большая часть предлагаемых в настоящее время моделей организации биоценоза оказывается достаточно примитивными (Гиляров, 2010), поэтому реализация рекомендаций по сохранению уникальных сообществ и представленных в них ценопопуляций редких видов не приносит желаемого результата (Большаков и др., 2009). Значительные темпы деградации растительного покрова приводят к осознанию того, что мы практически ничего не знаем о бесконечном многообразии связей, которые объединяют популяции разных групп организмов в единое целое. Усиливающаяся трансформация растительного покрова создает опасность не успеть изучить и описать все это разнообразие, так как коренные и типичные для определенных биомов ненарушенные фитоценозы на многих территориях уже замещены сильно обедненными вторичными сообществами с упрощенной структурой. Популяционноонтогенетический подход способствовал формированию популяционной парадигмы в биоценологии (Смирнова и др., 1990, 1993; Смирнова, 1998 и др.), которая сделала наши представления о ценозах полнее и глубже, позволила решать природоохранные вопросы более обоснованно (Сохранение…, 2002). Следующим шагом на пути к детализации наших знаний о биоценозе может стать анализ динамики формирования консортивных связей в процессе онтогенеза доминанта консорции. В рамках этой системы 16 сопряжены многочисленные онтогенезы и биологические времена разного уровня и масштаба. В этом отношении особенно интересны древесные растения, на основе организма которых образуется сложная экосистема, включающая разные виды лишайников, мохообразных, грибов, водорослей. В ходе онтогенеза дерева происходит дифференциация различных микрониш, которые являются основой для формирования синузий консортов. Представления о том, что древесное растение является «субстратом», на котором развиваются эпифитные виды, свидетельствуют об упрощенном и примитивном понимании структуры и организации биоценоза. Связи, существующие между его компонентами, значительно сложнее и разнообразнее. Именно такие работы приведут нас к пониманию динамики формирования в биоценозе взаимосвязей на разных структурных уровнях. Важно то, что многие из них оказываются сопряженными с онтоморфогенезом древесных растений, играющих ценозообразующую роль. Какие структурные особенности древесных растений разных онтогенетических состояний обуславливают определенное пространственное и временное распределение разных групп консортов? Нами проведен анализ динамики формирования консорции лишайников и мохообразных в процессе онтогенеза сосны обыкновенной (Pinus sylvestris L.) (Жукова и др., 2013). Этапы развития консорции (табл. 1, 2; рис. 1, 2) обусловлены структурными изменениями, происходящими в ходе онтогенеза сосны. Среди них появление корки, дифференциация ствола и кроны, появление отмирающей древесины. Корка в отличие от перидермы характеризуется меньшей скоростью отделения сбрасываемых элементов, их большей влагоемкостью, неоднородностью поверхности. Наличие ниш в трещинах между чешуйками корки в прикорневой части способствует их постепенному заполнению гумусовыми и торфянистыми субстратами. Уровневая дифференциация ствола обеспечивается разным типом покровных тканей, разным режимом влагообеспеченности. По характеристикам образующихся микрониш существенно отличается комлевая часть ствола. В основании комлевой части может формироваться приствольное возвышение. Оно в месте контакта с основанием ствола способствует появлению в комлевой части видов, растущих на лесной подстилке. Высотная дифференциация в области кроны обеспечивается процессами отмирания нижних ветвей, а впоследствии и появлением отмирающих ветвей в других частях кроны. Мертвая корка и древесина, лишенная корки, представляют специфические микрониши для консортов. На последних этапах онтогенеза объем лишенной корки отмирающей древесины сильно возрастает, что приводит к увеличению доли преимущественно эпиксильных лишайников (табл. 1, рис. 1). 17 Таблица 1 – Динамика изменения в консорции долевого участия мохообразных и лишайников на разных этапах онтогенеза Pinus sylvestris Группы Лишайники g2 g1 g3 – SS 11 (84,6) 40 (75,5) 67 (76,1) Мхи 1 (7,7) 11 (20,9) 14 (15,9) Печеночники 1 (7,7) 2 (3,8) 7 (8,0) 13 53 88 Всего Рисунок 1 – Динамика формирования консорции на разных этапах онтогенеза Pinus sylvestris: 1 – лишайники; 2 – мхи; 3 – печеночники Таблица 2 – Динамика изменения экологических спектров в консорции мохообразных и лишайников на разных этапах онтогенеза Pinus sylvestris Группы Лишайники g1 g2 g3 – SS 11 40 67 10 (90,9) 19 (47,5) 25 (37,3) эпиксилы 0 13 (32,5) 30 ( 44,8) эпигейные 1 (9,1) 7 (17,5) 11 (16,4) 0 1 (2,5) 1 (1,5) эпифиты лихенофильные Мхи 1 11 14 эпифиты 1(100) 6(54,5) 6 (42,9) эпиксилы 0 0 1 (7,1) эпигейные 0 5(45,5) 7 (50,0) Печеночники 1 2 7 эпифиты 1(100) 2(100) 2 (28,7) эпиксилы 0 0 5 (71,3) эпигейные 0 0 0 18 Рисунок 2 – Динамика изменения экологических спектров компонентов консорции в процессе онтогенеза Pinus sylvestris: 1 – эпифиты; 2 – эпиксилы; 3 – эпигейные; 4 – лихенофильные грибы Для всех рассматриваемых групп консорции максимальное видовое богатство наблюдается на старых генеративных и субсенильных особях сосны (табл. 1, рис. 1). В прегенеративном периоде у P. sylvestris отмечено только 2 вида лишайников (Жукова и др., 2013). Характер распределения в генеративном периоде представлен в таблице 1 и на рисунке 1. Анатомо-морфологические преобразования, происходящие в онтогенезе P. sylvestris, усиливают дифференциацию вертикальной структуры дерева, которая способствует увеличению разнообразия микрониш. Эти микрониши разные консорты заселяют в соответствии с особенностями их биологии и экологии, что приводит к существенным различиям в спектрах эколого-субстратных групп на разных этапах формирования консорции (табл. 2, рис. 2). Посредством приствольного возвышения обеспечивается более тесная связь с напочвенным покровом и появление эпигейных видов (табл. 2, рис. 2). Большая часть печеночников представляет эпиксильную группу. Характерное для сосны постоянное образование отмершей древесины обуславливает включение эпиксильных лишайников (табл. 2). Древесина также становится субстратом для ксилотрофных грибов. Очень сложны и интересны связи между грибами и другими элементами экосистем. Мицелий играет роль не только потребителя углеводов, синтезируемых растениями, но и распределителя органических веществ между различными организмами в зависимости от их потребно19 стей. Прекрасная книга С. Э. Смита и Д. Дж. Рида (2012) дает современное представление о ведущей роли грибов в большинстве физиологических процессов, об их роли в сообществах и биомах. Мицелий грибов выполняет функцию интернета, только более информационноемкого и отлаженного. Грибы так же, как и представители других царств, поразному взаимодействуют с растениями разных онтогенетических и виталитетных состояний. Эти сложные взаимодействия и определяют характер развития любой экосистемы и длительность ее существования. Каждый экологический фактор, нарушающий эти взаимодействия, может привести к гибели экосистем и биомов, в которые они входят. Таким образом, успешность реализации стратегий сохранения биоразнообразия во многом зависит от использования результатов популяционного анализа, а дальнейшее развитие популяционно-онтогенетического направления позволит выйти на качественно иной уровень понимания природных процессов и явлений. Л и т е р а т ур а Большаков В. Н. Сохранение биологического разнообразия: от экосистемы к экосистемному подходу / В. Н. Большаков, А. А. Лущекина, В. М. Неронов // Экология. – 2009. – № 2. – С. 83–90. Восточноевропейские леса: история в голоцене и современность: в 2 кн. – М.: Наука, 2004. Гиляров А. М. В поисках универсальных закономерностей организации сообществ: прогресс на пути нейтрализма / А. М. Гиляров // Журн. общ. биологии. – 2010. – Т. 71, № 5. – С. 386–401. Ведерникова О. П. Концепции популяционной биологии / О. П. Ведерникова, Л. А. Жукова // Регионология. – 2005. – № 5. – С. 200–210. Жукова Л. А. Поливариантность онтогенеза луговых растений / Л. А. Жукова // Жизненные формы в экологии и систематике растений: межвуз. сб. науч. тр. – М.: Изд-во МГПИ им. В. И. Ленина, 1986. – С. 104–114. Жукова Л. А. Популяционная жизнь луговых растений / Л. А. Жукова. – Йошкар-Ола: Ланар, 1995. – 225 с. Жукова Л. А. Многообразие путей онтогенеза в популяциях растений / Л. А. Жукова // Экология. – 2001а. – № 3. – С. 169– 176. Жукова Л. А. Популяционно-онтогенетическое направление в России / Л. А. Жукова // Бюл. МОИП. Отд. биол. – 2001б. – Т. 106, вып. 5. – С. 17–24. Жукова Л. А. Поливариантность развития организмов в разных царствах биосферы / Л. А. Жукова // Современные подходы к описанию структуры растения. – Киров: Лобань, 2008. – Разд. 6.3. – С. 240– 260. Жукова Л. А. Новый вариант классификации поливариантности развития организмов и популяций / Л. А. Жукова // Актуальные проблемы экологии, биологии и химии: материалы Всерос. конф. – Йошкар-Ола, 2010. – С. 76–81. Жукова Л. А. Значение биоморфологии для популяционно-онтогенетических исследований / Л. А. Жукова // Актуальные проблемы современной биоморфологии. – Киров: Радуга-ПРЕСС, 2012а. – Разд. 1.2.6. – С. 91–105. Жукова Л. А. Проблема сохранения биоразнообразия и роль популяционноонтогенетического направления / Л. А. Жукова // Биоразнообразие: проблемы изучения и сохранения: материалы Междунар. науч. конф., посвящ. 95-летию кафедры ботаники Тверского гос. ун-та (г. Тверь, 21–24 нояб. 2012 г.). – Тверь: ТвГУ, 2012б. – С. 31–35. Жукова Л. А. Поливариантность онтогенеза и динамика ценопопуляций растений / Л. А. Жукова, А. С. Комаров // Журн. общ. биологии. – 1990. – Т. 51, № 4. – С. 450–461. Жукова Л. А. Онтогенез сосны обыкновенной (Pinus sylvestris L.) / Л. А. Жукова, А. А. Нотов, Н. В. Турмухаметова, И. С. Тетерин // Онтогенетический атлас. – Йошкар-Ола, 2013. – Т. 7 (в печати). Жукова Л. А. Популяционно-онтогенетическая школа Марийского государственного университета и ее роль в изучении биоразнообразия / Л. А. Жукова, 20 Г. О. К. Османова, И. В. Шивцова., О. П. Ведерникова // Вестн. ТвГУ. Сер., Биология и экология. – 2012. – Вып. 26. – С. 107–124. Жукова Л. А. О некоторых подходах к прогнозированию перспектив развития ценопопуляций растений / Л. А. Жукова, Т. А. Полянская // Вестн. ТвГУ. Серия Биология и экология. – 2013. – Вып. 32 (в печати). Злобин Ю. А. Популяционная экология растений: современное состояние, точки роста / Ю. А. Злобин. – Сумы: Унив. кн., 2009. – 263 с. Злобин Ю. А. Популяции редких видов растений: теоретические основы и методика изучения / Ю. А. Злобин, В. Г. Скляр, А. А. Клименко. – Сумы: Унив. кн., 2013. – 439 с. Комаров А. С. Дискретные модели динамики биоразнообразия растительных сообществ при случайных уничтожающих воздействиях // Поливариантность развития организмов, популяций и сообществ / Мар. гос. ун-т. – Йошкар-Ола, 2006. – Разд. 4.2.1. – С. 171–175. Онтогенетический атлас растений / Мар. гос. ун-т. – Йошкар-Ола, 1997–2010. – Т. 1–7. Поливариантность развития организмов, популяций и сообществ / Мар. гос. ун-т. – Йошкар-Ола, 2006. – 326 с. Смирнова О. В. Популяционная организация биоценотического покрова лесных ландшафтов / О. В. Смирнова // Успехи соврем. биол. – 1998. – Т. 118, № 2. – С. 148. Смирнова О. В. Популяционная концепция в биоценологии / О. В. Смирнова, Л. Б. Заугольнова, Р. В. Попадюк // Журн. общ. биологии. – 1993. – Т. 54, № 4. – С. 438. Смирнова О. В. Популяционная организация растительного покрова лесных территорий (на примере широколиственных лесов европейской части СССР) / О. В. Смирнова, А. А. Чистякова, Р. В. Попадюк, О. И. Евстигнеев, В. Н. Коротков, М. В. Митрофанова, Е. В Пономаренко. – Пущино, 1990. Смит С. Э. Микоризный симбиоз / С. Э. Смит, Д. Дж. Рид. – М.: КМК, 2012. – 794 с. Сохранение и восстановление биоразнообразия / В. Е. Флинт, О. В. Смирнова, Л. Б. Заугольнова и др. – М.: Изд-во науч. и учеб.-метод. центра, 2002. – 286 с. Юрцев Б. А. Основные направления современной науки о растительном покрове / Б. А. Юрцев // Ботан. журн. – 1988. – Т. 73, № 10. – С. 1380–1395. Юрцев Б. А. Популяции растений как объект геоботаники, флористики, ботанической географии / Б. А. Юрцев // Ботан. журн. – 1987. – Т. 72, № 5. – С. 581–588. ПЕРСПЕКТИВЫ ПРОИЗВОДСТВА ПЕЛЛЕТ В РЕСПУБЛИКЕ МАРИЙ ЭЛ Ившин В. П.1, Хульт К.2 1 Марийский государственный университет, г. Йошкар-Ола, ivshin1941@mail.ru 2 Университет Йончепинг, г. Йончепинг, Швеция, kenneth.hult@jth.hj.se Топливные гранулы – экологически чистое топливо с содержанием золы не более 3 %. При производстве пеллет в основном используются отходы лесопильных производств и сельского хозяйства, которые ранее в основном вывозились на свалки и гнили, а по прошествии нескольких лет начинали гореть или тлеть. Гранулы отличаются от обычной древесины высокой сухостью (влажность всего 8–12 %, а влажность сырых дров – 30–50 %) и большей – примерно в полтора раза – плотностью, чем дрова. Эти качества обеспечивают высокую теплотворную способность по сравнению со щепой или дровами – при сгорании тонны гранул выделяется приблизительно 3,5 тысяч кВт·ч тепла, что в полтора раза больше, чем у обычных дров. Спрос на древесные брикеты и гранулы, оборудование для их сжигания и производства растет пропорционально ценам на такие традиционные виды топлива как нефть 21 и газ. В некоторых странах Европы, где рынок альтернативных источников энергии наиболее развит, гранулами отапливается до 2/3 жилых помещений. Такое широкое распространение объясняется и экологичностью этого вида топлива – при сгорании выбросы CO2 равны поглощению этого газа во время роста дерева, а выбросы NO2 и летучих органических компонентов значительно снижены благодаря использованию современных технологий сжигания. В 2009 году в Европе насчитывалось приблизительно 650 заводов по выпуску топливных древесных гранул, которые произвели более 10 млн тонн этой продукции. В 2011 году в Европе было произведено около 15 млн тонн пеллет. Согласно оценкам ООН, в России ежегодный объем производства топливных гранул в 2011 г. составлял 750 тыс. тонн, из которых 600 тыс. тонн экспортировалось. Площадь Республики Марий Эл на 60 % занята лесами, и по экономическим и экологическим соображениям производство пеллет представляется одним из перспективных направлений развития экономики Республики. В настоящем докладе обсуждаются два возможных варианта организации производства пеллет: бизнес-план выпускника 2013 г. Президентской программы для народного хозяйства подготовки управленческих кадров Российской Федерации Гурьянова Алексея Александровича, предпринимателя из г. Волжска и проект предпринимателя из Швеции Кеннета Хульта. Отечественный проект основан на перспективе разработки инновационной технологии переработки лесной продукции в Республике Марий Эл, где производство пеллет является высокодоходным элементом в связи с возможностью экспорта продукции в европейские страны. По расчетам автора проекта, рентабельность производства промышленных пеллет составляет 114,05 %, что по меркам бизнеса, основанного на переработке отходов – достаточно высокий результат. Проект предусматривает возможность экспорта продукции в европейские страны с использованием водных путей. Проект зарубежного инвестора основан на аренде лесных площадей в Республике Марий Эл и создании в г. Звенигово высокотехнологичного производства по комплексной переработке лесной продукции с учетом технологических достижений Швеции, которая является лидером производства и практического использования пеллет. Преимуществом второго проекта является возможность на арендованных территориях внедрения современной системы лесопользования, исключающей сплошные вырубки, захламление лесных площадей отходами и организацию свалок. Возможности реализации подобных проектов способствует утверждение Правительством РФ плана мероприятий по созданию благоприятных условий для использования возобновляемых древесных источников в производстве тепловой 22 и электрической энергии и позиция исполнительного директора НП «Национальный Биоэнергетический Союз» Ольги Ракитовой с анализом нынешнего состоянии отрасли и уверенностью в том, как предложенные меры скажутся на развитии биоэнергетики в стране. Л и т е р а т ур а Sikkema R. The European wood pellet markets: current status and prospects for 2020 / R. Sikkema, M. Steiner, M. Junginger, W. Hiegl, M. T. Hansen, A. Faaij // Biofuels, Bioproducts and Biorefining. – 2011. – Vol. 5. – Issue 3. – Р. 250–278. Гурьянов А. А. Разработка бизнесплана деревообрабатывающего цеха / Мар. гос. ун-т; А. А. Гурьянов. – г. Йошкар-Ола 2013. – Режим доступа: http://ru.wikipedia.org/w/index.php?title=Топливные_гранулы& oldid=59231786. ПРОБЛЕМЫ ИЗУЧЕНИЯ БИОРАЗНООБРАЗИЯ В СРЕДНЕЙ ШКОЛЕ Кохов А. В., Русов В. А. ГБОУ Средняя общеобразовательная школа № 1253, г. Москва, rusoval@rambler.ru Знания о биологическом разнообразии организмов является основополагающими в формировании экологического мировоззрения человека, для понимания места человека в биосфере, осознания уникальности каждого индивида в природе. Анализ современного состояния биологического образования в школе показывает резкое сокращение учебного времени на изучение многообразия животного и растительного мира. За последние пятнадцать лет в соответствии с базисными учебными планами количество часов, отведенных на изучение раздела «Растения. Бактерии. Грибы. Лишайники», сократилось со 102 часов до 34 часов, а раздела «Животные» – со 102 до 68 часов. При этом сокращение произошло в основном за счет уменьшения времени, отведенного на изучение видового разнообразия отделов и семейств растений, классов и отрядов животных. Только обзорно изучаются растения, имеющие важное хозяйственное значение. В разделе «Растения. Бактерии. Грибы. Лишайники» на знакомство с основными семействами цветковых растений отводится всего два–три часа. За это время предлагается познакомиться с пятью–семью семействами, их основными характеристиками. Еще хуже обстоит дело с изучением основных отделов растений, а также царств бактерий и грибов. Так, на изучение отделов Моховидные, Папоротникообразные (папоротники, хвощи, плауны), царства Бактерий предлагается по одному часу, а на изучение Грибы – 2 часа. Не лучше дело обстоит и с изучением раздела «Животные». Центральными понятиями этого раздела являются вопросы о внешнем 23 и внутреннем строении организмов, в меньшей степени рассматриваются вопросы о жизнедеятельности организмов. Многообразие организмов рассматривается обзорно, поэтому учащиеся зачастую не знают представителей разных типов животных. Так, на изучение наиболее многочисленных и распространенных животных – насекомых и млекопитающих – планируется не более четырех часов на каждый класс. Знакомство с видовым разнообразием животных ограничивается их перечислением. Сведения об экологии и поведении животных или отсутствуют в учебниках, или даются ограниченно, поэтому учащиеся не знают, что одни из наиболее распространенных в нашей местности животные – муравьи являются хищниками, а поэтому не понимают, почему муравьи являются санитарами леса. Материал школьного учебника насыщен фактами, которые ученик должен просто запомнить. Биологические знания не имеют практической направленности. В результате на школьных олимпиадах вопрос о хромосомном наборе в клетках эндосперма семян голосеменных растений оказывается для школьников более простым, чем вопрос о времени созревания семян сосны, значимости этих растений в биоценозе. Учащиеся не могут отличить лист клена от березы или рябины, хотя эти растения встречаются повсеместно. В контрольных измерительных материалах разного уровня (от внутришкольных и региональных до Единого государственного экзамена) практически отсутствуют задания, контролирующие уровень знаний о биологии отдельных видов, семейств, отрядов, отличий биологических объектов на уровне видовых и родовых критериев. Это связано с тем, что в Федеральном государственном образовательном стандарте определены только требования к знаниям общих характеристик царств, типов (отделов) и классов, что вынуждает учителя сокращать время на изучение видового разнообразия, а уделять больше внимания характеристикам крупных таксонов. При изучении биологических объектов не уделяется внимание рассмотрению их индивидуальных особенностей, экологии организмов и поведению. Виды предстают перед учеником как некие модели, обладающие определенным набором признаков, а не как множество живых организмов со своими индивидуальными особенностями. Компьютеризация современной школы в условиях сокращения учебных часов на изучение биологии усилила эту тенденцию. Все больше учителей поддаются соблазну подмены лабораторных и практических работ презентациями, все чаще взамен демонстраций опытов, постановки эксперимента с живыми объектами на уроках используют виртуальные лаборатории. 24 Аналогичная ситуация складывается и с изучением вопросов онтогенеза организмов в разделе «Общая биология». В учебниках рассматривается исключительно эмбриональное развитие. Поэтому учащиеся могут охарактеризовать стадии бластулы, гаструлы, органогенеза, но стадии постэмбрионального развития описывают с большим трудом и только для одного–двух видов насекомых и лягушки. Развитие растений вообще не рассматривается, а понятие однолетние, двулетние и многолетние связываются только с длительностью жизни, но не с периодами их развития. Подавляющее большинство выпускников школы уверены, что все травянистые растения зацветают в первый год жизни, и лишь немногие учащиеся вспоминают о цветении на второй год жизни растений из семейства Капустные. У выпускника современной школы отсутствуют самые общие представления об онтогенезе растений. Несформированность понятий о биологическом разнообразии в курсах «Ботаника» и «Зоология» создают серьезные проблемы в старших классах при изучении вопросов эволюции и экологии в разделе «Общая биология». Задачей этого раздела является объяснение с позиций генетики, молекулярной биологии, цитологии эволюционного учения, возникновения и существования биологического разнообразия во всех его проявлениях. Отсутствие знаний о видовом разнообразии лишает этот материал практической значимости и становится сугубо теоретическим. Овладев теоретическими основами молекулярной биологии и цитологии в объемах предусмотренных программой, школьники не могут привести примеры, подтверждающие теорию. Например, зная строение и функции пластид, они не могут привести примеры взаимоперехода пластид, например, при созревании плодов. Интерес к предмету снижается. Наиболее привлекательным для старшеклассников остается раздел «Генетика», и понятно почему. Теоретические знания о закономерностях наследственности позволяют объяснять появление признаков у человека, признаков – знакомых, хорошо известных. К сожалению, курс «Биология», по-видимому, ждет судьба, уже постигшая другие предметы естественного цикла, – отсутствие практической значимости. Значимость, ценность изучения биологии в средней школе – это приобретение знаний об объектах живой природы, окружающих нас постоянно и повсеместно, умения различать их, понимать и видеть динамику их развития, механизмы приспособленности к совместному сосуществованию. Именно это и уходит из школьной биологии. 25 Л и т е р а т ур а Константинов В. М. Биология. 7 кл. / В. М. Константинов, В. Г. Бабенко, B. C. Кучменко. – М.: Вентана-Граф, 2010. Никишов А. И. Биология. Растения. Бактерии. Грибы и лишайники: учебник для учащихся 6 кл. / А. И. Никишов, В. П. Викторов. – М.: Владос, 2013. Никишов А. И. Естествознание. Биология. 5–11 кл.: программы для общеобразовательных учреждений / А. И. Никишов. – М.: Мнемозина, 2011. Никишов А. И. Биология. Животные: учебник / А. И. Никишов, И. Х. Шарова. – М.: Владос, 2012. Пономарева И. Н. Природоведение. Биология. Экология. 5–11 кл.: программы / И. Н. Пономарева, Т. С. Сухова, И. М. Швец. – М.: Вентана-Граф, 2011. Серебрякова Т. И. Биология. Растения. 6–7 кл.: учебник / Т. И. Серебрякова. – М.: Просвещение, 2000. Трайтак Д. И. Биология. 5–11 классы: программа / Д. И. Трайтак, Н. Д. Андреева. – М.: Мнемозина, 2011. Трайтак Д. И. Биология. Растения, бактерии, грибы, лишайники. 6 кл.: учебник / Д. И. Трайтак, Н. Д. Трайтак. – М.: Мнемозина, 2012. Хрыпова Р. Н. Биология. Растения, бактерии, грибы, лишайники. 6 кл.: учебник / Р. Н. Хрыпова. – М.: Мнемозина, 2011. ОСОБЕННОСТИ БИОРАЗНООБРАЗИЯ РОССИЙСКОЙ ЧАСТИ КАВКАЗА В УСЛОВИЯХ ИЗМЕНЕНИЯ ОКРУЖАЮЩЕЙ СРЕДЫ Литвинская С. А. Кубанский государственный университет, г. Краснодар, Litvinsky@yandex.ru Важной задачей является сохранение всего многообразия видов в биосфере, ибо она теряет устойчивость и целостность. Причин немало, и практически все они связаны с деятельностью человека. Российская часть Кавказа располагает уникальными экосистемами, имеющими высокую биологическую значимость, обладающими огромным ресурсным потенциалом. Природоохранный статус биологического разнообразия региона для мира чрезвычайно высок. Следует отметить высокий уровень флористического разнообразия и эндемизма отдельных локусов Северного Кавказа. Для региона характерен высокий процент редких и исчезающих видов, подлежащих охране на уровне государства и субъектов федерации, что уже само по себе является индикатором экологической напряженности. Сложность горного рельефа, разнообразие климатических типов, обилие кальцефильных субстратов на столь небольшой территории способствовали интенсивным микроэволюционным процессам. Химизм субстрата, интенсивность солнечной радиации, увеличивающейся с высотой, приводят к появлению устойчивых мутаций и формированию в процессе эволюции локальных эндемичных видов, высокому популяционному и видовому разнообразию и своеобразию флоры. Высокий уровень эндемизма и флористического биоразнообразия связан также со сложным флорогенезом, единством с флорами Крыма, Древнего Средиземноморья, Восточной Европы и Передней Азии (см. табл.). 26 Биоразнообразие региональных флор сосудистых растений Российской части Кавказа Край, республика Биоразнообразие Площадь, тыс. км2 Включено в Красные книги Доля нарушенных земель, % Российская часть Кавказа около 4000 254 184 (РФ) около 44 Краснодарский край 3200 75,5 261 (РФ-119) 61,1 Республика Адыгея 2000 7,8 119 (РФ-52) 60,9 Ставропольский край 2251 66,2 286 (РФ-47) 65,2 Кабардино-Балкария 2985 12,5 72 (РФ-29) 39 Чеченская Республика 2315 15,6 145 (РФ-46) около 30 Республика Дагестан 3600 50,3 165 (РФ-83) 18,1 Карачаево-Черкесия 1903 14,3 42 (РФ-26) 39,2 3,6 81 (РФ-29) более 30 8 101 (РФ-56) 36,5 Республика Ингушетия Северная Осетия-Алания 3054 Это единственный регион России, где представлены субтропики. На Северном Кавказе сохранились уникальные лесные экосистемы (субсредиземноморские, субропические колхидские), не имеющие аналогов в мире, по биоразнообразию регион занимает одно из первых мет в России. Специфическими чертами лесных экосистем региона является высокое флористическое богатство ценотаксонов: биоразнообразие формационных флор Quercus petraea L. ex Liebl. – более 580 видов, Quercus pubescens Willd. – около 540, Fagus orientalis Lipsky – более 400, в можжевеловых редколесьях зарегистрировано более 600 видов сосудистых растений (Литвинская, 2006). Все экосистемы насыщены редкими и исчезающими видами, подлежащими охране на уровне государства. Так, в кубанских широколиственных лесах зарегистрировано произрастание 29 видов, подлежащих охране в РФ, в субсредиземноморских (арчевники, томилляры, горные степи) – 46, в колхидских смешанных лесах – 41 вид, в высокогорьях – 29, в степях Западного Предкавказья – 21 вид. Большую роль в сохранении биологического разнообразия играют заповедные территории. В условиях антропогенного наступления на природные системы заповедники – это единственные уголки нетронутой девственной природы. Каждый заповедник – это уникальная природная коллекция видового и экосистемного разнообразия, аналогов которой в мире нет. Только в заповедниках, где обеспечивается естественная эволюция экосистем и сохраняется их естественное историческое функционирование, возможен экологический мониторинг всех компонентов природной среды и экосистем с целью глубокого изучения 27 и прогнозирования динамики их развития. Именно им отведена определяющая роль в сохранении естественных ландшафтов и популяций видов. Регион обеспечен рядом заповедных территорий, которые сохраняют редкий генофон. В Тебердинском биосферном заповеднике произрастает 1 200 видов сосудистых растений, в Кавказском – 1 700, в Кабардино-Балкарском – 950, в Северо-Осетинском – около 2 000 видов. На территории Сочинского национального парка зарегистрировано 1 658 видов (Тимухин, 2002). Конечно, сложившаяся заповедная сеть не решает проблем сохранения ландшафтного и популяционно-видового разнообразия такого сложного региона, как Российская часть Кавказа. Так, только в северозападной части Большого Кавказа не охраняется 45 % видов, занесенных в региональные Красные книги. Для Северного Кавказа характерна высокая степень антропогенной нарушенности экосистем. По нарушенности земель регион занимает лидирующее положение в России (см. табл.). Причины трансформации аборигенных флор многочисленны и известны и для других регионов России. Следует отметить, что для Российской части Кавказа характерна концентрация многих трансформационных причин в одной экосистеме, проявляющихся на ограниченном пространстве, что делает их чрезвычайно уязвимыми. Северный Кавказ является одним из самых популярных курортов России. Это связано с благоприятным географическим положением, наличием трех теплых морей, уникальными природными минеральными источниками и лечебными грязями, разнообразными ландшафтами и т. д. Основное негативное воздействие оказывают: высокая плотность населения, хозяйственное освоение территорий (мелиорация, террасирование склонов, применение химических средств и использование удобрений в агроландшафтах, берегозащитные мероприятия, создание искусственных водоемов, отказ от традиционных форм ведения хозяйства, эвтрофикация плавневых зон и водоемов, прямое уничтожение декоративных и лекарственных растений, открытая добыча полезных ископаемых, нерегулируемый поток туристов и рекреация, курортное строительство, прокладка трубо- и газопроводов, линий электропередач; изъятие песка с пляжей и гравия из русел рек, пожары, неконтролируемый выпас скота и выход его в субнивальную зону, нефтяное загрязнение, аренда раритетных и лесных территорий, рубки горных водоохранных лесов). Все это неизбежно разрушает жизненную среду вида, популяций, ценозов на огромных площадях, приводит к негативным трансформациям разнообразия и снижению биологической продуктивности естественных биоценозов. Биота горных экосистем оказалась чрезвычайно уязвимой к антропогенным факторам, что привело к различным деформациям естественных сообществ, 28 уменьшению аборигенного биоразнообразия, ослаблению экосистем. В регионе лесные формации теряют специфичность и сближаются по структуре и составу, формационные флоры упрощаются за счет их синантропизации. Анализ спектра биоморф скальнодубовых показал, что в нем преобладают кистекорневые, короткокорневищные (14,3 %) травяные поликарпики. Следует отметить высокий процент однолетних, однодвулетних монокарпиков. По сравнению с показателями по широколиственным лесам Кавказа и бывшего СССР (Жижин, Зеленский, 1975) отмечается явное смещение в сторону увеличения количества стержнекорневых, однолетних и одно-двулетних и дерновинных, что связано с нарушенностью региональных лесов. Экологическая структура флоры отражает количественное распределение видов по группам в зависимости от действия абиотических факторов. По отношению к свету в региональной флоре СЗК преобладают гелиофиты (50 %), на долю сциофитов приходится 9,3 %, а в букняках их – 45 %. Обилие светолюбивых видов свидетельствует о нарушенности фитоценотической структуры лесных ценозов постоянными рубками, которыми систематически подвергались дубняки. В них на долю гелиофитов приходится 30–40 %. Отмечаются и отклонения в структуре жизненных форм по Раункиеру. Так, в черешчатодубовых лесах вследствие нарушенности высок процент терофитов, в гемиксерофильных лесах повышается доля участия хамефитов (до 16,7 %) и гемикриптофитов. Для региона затруднительно установить приуроченность вида к типу фитоценоза. Из-за значительной нарушенности и неоднократной смены лесного ландшафта на луговой и опушечный виды, обладающие широкой эколого-ценотической амплитудой при демутации растительного покрова, занимали близкие экологические ниши, что и привело к обилию видов, произрастающих одновременно на лугах, степях и лесах. В целом отмечается снижение роли сильвантов в дубняках (40–54 %) и увеличение пратантов (до 8–9 %), степантов (6 % – в формации дуба черешчатого). Анализ видов по широте эколого-ценотической амплитуды подтвердил факт деградации. Виды, имеющие стенотопную и гемистенотопную амплитуду, встречаются наравне с эвритопными и гемиэвритопными. На СЗК наибольшую специфичность сохранили буковые леса, где преобладают гемистенотопные и стенотопные виды (260), гемиэвритопные доминируют в дубняках (337–369), что показывает размывание границ между лесными ценофлорами и потерю ими специфичности. Высокая флористическая гетерогенность и видовое богатство лесных ценофлор обусловлено инвазией видов из нарушенных экотопов. Следствием антропогенных воздействий является слабая изоляция и стабильность, низкая степень специфичности лесных формационных 29 флор. Но в целом видовое биоразнообразие увеличилось за счет неоднократных антропогенных сукцессий, аборигенное же биоразнообразие сократилось, что привело к потере устойчивости и упрощению структуры лесных экосистем. Степень общности (коэффициент Жаккара, К) лесных флор высока. Так, К скальнодубовых и черешчатодубовых лесов равен 41, буковых и скальнодубовых – 34. Второй уровень сходства (К = 20–29) имеют флоры Quercus robur L. и Quercus pubescens, хотя экологически они различны. Горные экосистемы Российской части Кавказа, вследствие своей природной уязвимости, чрезвычайно чувствительны к происходящей аридизации климата. Первыми на климатические изменения реагируют экотонные экосистемы верхних горных поясов и популяции эндемичных видов cубнивального пояса. Дальнейшее потепление климата может привести в Кавказском экорегионе к сокращению и потере аборигенного эндемичного локального генофонда. Как показали исследования, в особо опасном состоянии в настоящее время находятся экосистемы субнивального и альпийского поясов Большого Кавказа. Они имеют особо высокое значение интегрального показателя чувствительности к климатическим изменениям (10), в частности к потеплению климата. Особенно чувствительны к аридизации узколокальные эндемичные альпийские виды, тяготеющие к произрастанию на моренах, вблизи ледников и снежников. Это узкие эндемики Psephellus salviifolius Boiss. – Центрального (ЦК) и Восточного Кавказа (ВК), Arabis farinacea Rupr. – ВК, Campanula bellidifolia Adams и Rhinanthus schischkinii Vassilcz. – ЦК, Adams Ranunculus helenae Albov и Oxytropis kubanensis Leskov – эндемики Западного Кавказа, Minuartia brotheriana (Trautv.) Woronow, Scrophularia minima Bieb., Veronica minuta C. A. Mey. и Asperula accrescens Klok. – эндемики Большого Кавказа, Corydalis vittae Kolak. – Западного Закавказья. Редчайший эндемик ЦК и ВК Pseudovesicaria digitata (C. A. Mey.) Rupr. произрастает на щебнистых полуподвижных склонах, сланцевых осыпях, моренах, по краям снежников, где приурочен к путям схода лавин в пределах высот 2500–3600 м над ур. м., причем, коренные популяции приурочены только к субнивальному поясу. К климатическим флуктуациям чувствительные эндемичные виды, произрастающие на скалах и на подвижных осыпях альпийского и субальпийского поясов: Anthriscus ruprechtii Boiss., Ficaria ledebourii Grossh. et Schischk., Thymus pseudopulegioides Klok. et Shost. (кавказские эндемики), Arabis kazbegi Mtskhvet., Ranunculus lojkae Somm. et Levier (эндемики ЦК). Состояние популяций большинства из этих видов угрожаемое. Основные лимитирующие факторы – это климатические флуктуации, хозяйственное освоение высокогорных терри30 торий, ареальная редкость, низкая плотность популяций, узкая экологическая амплитуда, рекреация. В связи с изменением геополитической ситуации в некоторых регионах Российского Кавказа возросла социально-экологическая напряженность. Критическое состояние биоразнообразия горных ценотаксонов прослеживается повсеместно. Существует угроза опустынивания, сопровождающаяся эрозийными процессами, засолением, загрязнением и утратой продуктивности почв, обеднением биологического и экосистемного разнообразия, полной деградацией растительного покрова, загрязнением вод. В последние годы добавились непродуманные экономические реформы и аренда земель. Под влиянием распашки земель и неконтролируемого выпаса исчезли степи Предкавказья. Биоразнообразие на видовом и экосистемном уровнях в Российской части Кавказа невозможно сохранить без экосистемного подхода, без инвентаризации экосистем, учета экотопов, ландшафтов. Сохранение биологического разнообразия и генетических ресурсов Российской части Кавказа должно предусматривать: разработку и внедрение научных основ сохранения биоразнообразия с учетом специфических особенностей экосистем; управление природоохранной деятельностью, расширение сети заповедных территорий и координация их деятельности; осознание значимости сохранения биологического разнообразия и снижение антропогенного прессинга на экосистемы; возрождение рационального национального природопользования; прогнозирование развития биоразнообразия экосистем под воздействием антропогенных факторов. Но все это необходимо проводить на фоне снижения антропогенного прессинга. Экосистемы Российской части Кавказа нуждаются в обеспечении экологической безопасности и сохранении биологического разнообразия на всех уровнях. Они должны выполнять только биосферные функции. Л и т е р а т ур а Жижин Р. П. Изменение грабово-дубовых сосняков Pinetum carpinoso-quercetum Росточья и Малого Полесья под влиянием рекреационных нагрузок / Р. П. Жижин, Н. Н. Зеленский // Укр. ботан. журн. – Киев, 1975. – Т. 32, № 5. – С. 639–644. Литвинская С. А. Биоразнообразие важнейших растительных таксонов Северо-Западного Кавказа / С. А. Литвинская // Экологическое и ландшафтное разнообразие Северного Кавказа и особо охраняемых природных территорий: тр. Тебердинского государственного заповедника. – М., 2006. – Вып. 43. – С. 81–84. Тимухин И. Н. Редкие сосудистые растения Кавказского заповедника и Сочинского национального парка / И. Н. Тимухин // Биоразнообразие и мониторинг природных экосистем в Кавказском государственном природном биосферном заповеднике: труды Кавказского государственного природного биосферного заповедника. – Новочеркасск, 2002. – Вып. 16. – С. 39–65. 31 ЭКОЛОГИЯ ДЕТСТВА И НОВЫЕ ПОДХОДЫ К СОЦИАЛЬНО-ПЕДАГОГИЧЕСКОЙ РЕАБИЛИТАЦИИ ДЕТЕЙ-ИНВАЛИДОВ Морова Н. С. Марийский государственный университет, г. Йошкар-Ола, morovans@gmail.com Научно-техническое развитие общества во многом изменило экологию человека. Несмотря на то, что в человеческом обществе преобладает действие социальных законов, и оно может создавать и поддерживать определенные типы экосистем в соответствии со своими потребностями, биологические свойства организма определяются базовыми законами функционирования систем. Неблагоприятное влияние экологических условий на организм, с одной стороны, могут оказывать отрицательное воздействие на процесс формирования плода, а с другой – способствовать развитию процессов дезадаптации, связанных с ослаблением резистентности новорожденных, что нередко ведет к обострению болезней с преждевременной инвалидизации. На стыке экологии, педиатрии, детской психологии, социальной педагогики и других появилось новое научное направление – экология детства. В Российской Федерации частота детской инвалидности за последние 20 лет увеличилась более чем в 2 раза. Способствует этому множество факторов, среди которых главными являются процесс технизации производства и общества с участившимися авариями, ухудшение экологии из-за стихийных бедствий (яркий пример – наводнение в Амурской области в 2013 г.), распространение СПИДа и наркомании, отсутствие стройной системы профилактики детской инвалидности и т. д. Не может не вызывать тревогу такой факт, что уже в 1993 году более, чем в 40 территориях России показатели смертности превысили цифры рождаемости, а в них на здоровое потомство, составляющее полноценный генофонд, приходилось менее 10 %. Цифры свидетельствуют, что этот процесс продолжается до сих пор. Данные последних лет свидетельствуют о том, что проблема детской инвалидности, замалчиваемая десятилетиями в России, в настоящее время требует качественно иного осмысления. На первый план выдвигается задача предупреждения детской инвалидности, а также создание системы эффективной социальной, медико-психолого-педагогической реабилитации детей с различными ограничениями с рождения до 18 лет, которая бы способствовала интеграции детей-инвалидов в общество, а воспитывающие их семьи обеспечила равными возможностями для активного участия в окружающей жизни. 32 Многоплановость подходов к проблеме социальной реабилитации детей с ограниченными возможностями здоровья свидетельствует о том, что ее решение требует взаимосвязанного рассмотрения медицинского, педагогического, психологического, юридического, экологического и ряда других аспектов. Обобщая все в целом, можно заметить, что проблема социальнопедагогической реабилитации детей с ограниченными возможностями здоровья есть не что иное, как воспаленный нерв социальной экологии детства, поскольку количественный рост детской инвалидности, который наблюдается в ряде регионов, трансформируется в качественное изменение всего общества. Именно поэтому разработка новой концепции социально-педагогигеской реабилитации детей с ограниченными возможностями здоровья, теоретическое обоснование обобщенной интегрированной модели в работе с детьми-инвалидами и воспитывающими их семьями по месту жительства, апробация содержания и технологий социально-педагогической реабилитации с различными категориями детей с ограниченными возможностями здоровья – стали главными приоритетами научной школы по проблеме «Социально-педагогическая реабилитация детей с ограниченными возможностями здоровья», активно действующей в Республике Марий Эл и далеко за ее пределами с 1991 года по настоящее время. Суть данной модели сводится к следующему: «человек, имеющий инвалидность, имеет равные права на участие во всех аспектах жизни общества; равные права должны быть обеспечены системой социальных служб, выравнивающие ограниченные в результате травмы или заболевания возможности. Инвалидность – не есть медицинская проблема. Инвалидность – проблема неравных возможностей». Эти идеи и стали основополагающими для нас при построении теоретической модели социально-педагогической реабилитации детей с ограниченными возможностями здоровья, в основу которой положены следующие концептуальные подходы: П е р в а я – гуманистический характер отношения к ребенку с ограничивающими возможностями и воспитывающей его семье. Это связано с уважением к личности ребенка и его близких, взявших на себя огромную ответственность за воспитание больного ребенка. В т о р а я – личностно-деятельностный подход к ребенку с ограниченными возможностями, который ставит его в обязательную деятельную позицию, мешающую овладеть синдромом «приобретенной беспомощности». Это осуществимо в условиях проявления собственной активности ребенка, которая должна стимулироваться близкими во всех сферах его жизнедеятельности. 33 Т р е т ь я – социальное видение ребенка и всемерное содействие раскрытию его социальной сущности. Отказ от традиционного подхода с господством медицинской модели инвалидности в пользу модели социально-педагогической реабилитации. Таким образом, закрепляется идея, что проблема инвалидности ни в коем случае не ограничивается медицинским аспектом, а выступает как социальная проблема неравных возможностей. Представленные концептуальные идеи дают новый принципиальный подход к триаде «ребенок– общество – государство». Суть этих изменений можно выразить в следующих положениях: 1) основная проблема ребенка, имеющего проблемы в развитии, заключается в изменениях его связи с внешним миром, что выражается в нарушении контактов со сверстниками и взрослыми, ограниченном общении с природой и духовным наследием, лимитированностью в приобретении желаемой специальности и т. д.; 2) ребенок с ограниченными возможностями является равноправным членом общества, в котором живет, а потому он должен и может участвовать в его многогранной жизни. Общество обязано создать для него специальные условия, предоставляющие ему равные возможности для удовлетворения всех гражданских прав; 3) ребенок с особыми нуждами может иметь такие же задатки и способности, как и его сверстники с полноценным здоровьем. Задача демократического общества – помочь ему обнаружить свои дарования и развить их с максимальной пользой как для него самого, так и для общества в целом; 4) ребенок с отклонениями в психофизическом развитии не должен быть пассивным объектом социальной помощи. Она должна быть построена так, чтобы способствовала постепенному превращению ребенка из воспринимающего объекта в активно действующий субъект, развитие и становление которого происходит в семье как лучшей инфраструктуре социальной реабилитации; 5) для успешного осуществления процесса социальной реабилитации ребенка с неполноценным здоровьем необходима мобилизация ресурсов всех трех секторов – государственного, коммерческого (частного) и общественного, которые должны объединиться на платформе оказания максимальной помощи и поддержке семьи; 6) эффективность процесса социальной реабилитации во многом определяется уровнем квалификационного, методически выверенного педагогического руководства со стороны специалистов по работе с семьями, воспитывающими детей с отклонениями в развитии. 34 Таким образом, новые подходы к реабилитации детей с ограниченными возможностями здоровья и инвалидам представляют собой целостную систему взаимосвязанных медико-социальных, психологопедагогических, экологических и других мероприятий, создающих условия для эффективного включения ребенка во взаимодействие с социальной средой. СОХРАНЕНИЕ БИОРАЗНООБРАЗИЯ ПРИРОДНЫХ ЭКОСИСТЕМ ТАТАРСТАНА В УСЛОВИЯХ ИНТЕНСИВНОЙ ХОЗЯЙСТВЕННОЙ ДЕЯТЕЛЬНОСТИ Рахимов И. И., Ибрагимова К. К., Леонова Т. Ш. Казанский (Приволжский) федеральный университет, г. Казань, rakhim56@mail.ru Интенсивная антропогенная трансформация естественных природных ландшафтов, демографический взрыв, глобальное загрязнение окружающей среды оставляют все меньше естественных территорий для живых организмов, приводя к еще большему обострению взаимоотношений человека и объектов биоразнообразия. Татарстан является промышленным регионом с развитой транспортной сетью, большой плотностью населения и множеством населенных пунктов. Основная часть территории – это земли сельскохозяйственного назначения (69,2 % от общей площади Республики Татарстан (РТ). Лесистость республики составляет 16,9 %. В этих условиях серьезная роль возлагается на особо охраняемые природные территории (ООПТ) Республики Татарстан. Для поддержания территориального экологического равновесия площади экстенсивно используемых и охраняемых территорий по отношению к интенсивно эксплуатируемым должны составлять: в лесной зоне – 30–35 %, в зоне лесостепи – 25–30 %. При этом условии сохраняется относительная целостность биотопа, а пространственная мозаичность участков обеспечивает поддержание благоприятных условий для животных с активным образом жизни. Сегодня только 1,3 % территории Татарстана относится к системе особо охраняемых, включая Волжско-Камский государственный природный биосферный заповедник и национальный парк «Нижняя Кама». Создание и расширение сети особо охраняемых природных территорий (ООПТ) с разной степенью хозяйственного, природоохранного и рекреационного использования играет большую роль в охране редких видов растений и животных и способствует сохранению биологического разнообразия Среднего Поволжья. 35 Волжско-Камский край был издавна населен людьми, и большая часть его территории за всю историю освоения претерпела значительную антропогенную трансформацию, которая является причиной изменений в составе флоры и фауны: на громадных площадях естественная древесная растительность полностью замещена агроценозами. Сейчас большая часть региона занята «окультуренной лесостепью» или, точнее, лесопольем, на фоне которого в виде сложной мозаики рассеяны островки лесов. Распаханность большинства районов достигает 70–80 %, поля с монокультурами и частая их смена не допускают формирования устойчивых сообществ. Существенно повлияло на природные комплексы создание каскада водохранилищ (Куйбышевское, Нижнекамское и др.). На протяжении всей истории в Татарстане происходил интенсивный рост численности населения с опережающим ростом числа городских жителей. Так, в начале XVIII в. в городах жило лишь 3 % населения, в 1897 году – 12,0 %, в 1998 г. в Среднем Поволжье горожане составляли 73 %, плотность населения 31,3 чел. на 1 км2. Мощное антропогенное воздействие не только изменило состав фауны, но и привело к ее стабилизации на качественно новом уровне. Основными антропогенными факторами, влияющими на состояние среды обитания животных и растений и определяющими характер антропогенной трансформации природной экосистемы, являются степень развития сельского хозяйства, уровень промышленности и плотность населения. Наиболее характерной чертой растительного покрова РТ является его разнообразие, которое определяется особенностями переходной зоны. На северо-востоке можно увидеть темнохвойные леса таежного типа с участками южной тайги, с запада проникают дубравы, на юге встречаются ковыльные степи. Наряду с хвойными лесами, в республике имеются и широколиственные леса. Изначально это были дубравы Правобережья Волги, пойменные и равнинные дубравы. В связи с интенсивным освоением территории в первую очередь именно эти леса стали сводиться, так как занимали самые богатые почвы, во-первых, а во-вторых, содержали запас ценной древесины. Еще Петр I положил начало интенсивным вырубкам дубрав Правобережья Волги. В настоящее время сохранились небольшие массивы нагорных дубрав на западе и юго-западе республики. В южной и юго-восточной части республики сохранились небольшие по площади степные сообщества, главным образом, по склонам, балкам, неудобным для распашки местам. Когда-то открытые пространства занимали ковыльные степи, в которых господствовали злаковые растения. В сообществах степей и лугов в отличие от лесных ценозов 36 основная конкуренция между растениями идет за влагу и почвенное богатство. Поэтому для них характерна горизонтальная неоднородность в распределении видов по площади. Географическое расположение Татарстана на границе двух природных зон: леса и степи, разнообразие ландшафтов, значительная обводненность территории создают благоприятные условия для обитания многих представителей фауны. По последним данным, в республике встречается более 430 видов только позвоночных животных и несколько тысяч видов беспозвоночных (Попов, Лукин, 1988). Фауна Татарстана претерпела изменения не только в плане сокращения численности животных. По разным причинам численность одних видов животных то сокращается, то увеличивается, одни виды исчезают, а другие появляются. За последние десятилетия в Татарстане интродуцированы виды, ранее здесь не обитавшие и для региона новые: енотовидная собака (Nyctereutes procyonoides (Gray, 1834)), американская норка (Neovison vison (Schreber, 1777)), белка-телеутка (Sciurus vulgaris (Linnaeus, 1758)), ондатра (Ondatra zibethicus (Linnaeus, 1766)). Все виды в новых условиях прижились и расселились по территории всего края. Изменения в составе фауны за последние десятилетия произошли и в связи с созданием водохранилищ. Водный режим водохранилищ создает благоприятные условия для обитания одних видов и приводит к сокращению численности других видов. С образованием каскада водохранилищ были прерваны нерестовые миграции 12 проходных видов рыб: белуга (Huso huso (Linnaeus, 1758)), русский осетр (Acipenser gueldenstaedtii (Brandt, 1833)), севрюга (Acipenser stellatus (Pallas, 1771)), каспийский лосось (Alosa kessleri (Grimm, 1887)) и др. В результате естественной миграции появились бычок-кругляк (Neogobius melanostomus (Pallas, 1814)), игла-рыба (Syngnathus typhle (Linnaeus, 1758)), ротан (Perccottus glenii (Dybowski, 1877)) и др. Из беспозвоночных животных в водохранилище вселены донные рачки – мизиды, призванные в значительной степени повысить его кормовую базу. Все эти процессы требуют постоянного мониторинга. Антропогенное влияние на природные комплексы ощущается всюду, не остаются вне этого даже особо охраняемые природные территории. Татарстан – регион с развитой промышленностью, сельским хозяйством и транспортом. В этих условиях, как отмечалось выше, в республике остро встают вопросы сохранения биологического разнообразия, которые включают разнообразие ландшафтов, биогеоценозов, видов животного и растительного мира. Одной из форм охраны биоразнообразия является создание особо охраняемых природных территорий 37 (ООПТ). Территория природно-заповедного фонда составляет в республике 1,3 % от площади, что крайне мало для сохранения устойчивости природной среды. По мнению экологов, в регионе по разным оценкам от 3 до 10 % площади должно изыматься из хозяйственного использования. Эти территории должны служить резервом для восстановления природы в промышленно развитых регионах. Сохранение естественных природных сообществ – сложная, но крайне актуальная проблема, так как только сохранение биологического разнообразия, которое подразумевает и популяционное, и видовое, и биогеоценотическое, может обеспечить устойчивость экологических систем региона и всей биосферы Земли. Л и т е р а т ур а Красная книга Республики Татарстан. – Казань: Идель-пресс, 2006. – 832 с. Попов В. А. Животный мир Татарии / В. А. Попов, А. В. Лукин. – Казань: Татарское кн. издво, 1988. – 248 с. О НЕКОТОРЫХ ОСОБЕННОСТЯХ СТРУКТУРНОЙ ОРГАНИЗАЦИИ * РАСТЕНИЙ В УСЛОВИЯХ ПЕРЕМЕННОГО УВЛАЖНЕНИЯ Савиных Н. П. Вятский государственный гуманитарный университет, г. Киров, botany@vshu.kirov.ru Изучена биоморфология веха ядовитого (Cicuta virosa L.), поручейника широколистного (Sium latifolium L.), омежника водного (Oenanthe aquaticа (L.) Poir.), частухи подорожниковой (Aliama plantagoaquatica L.), стрелолиста обыкновенного (Sagittaria sagittifolia L.), сусака зонтичного (Butomus umbellatus L.), калужницы болотной (Calrha palustris L.), жерушника земноводного (Rorippa amphibia (L.) Bess.), жерушника болотного (R. palustris (L.) Bess.) и жерушника обоюдоострого (R. × anceps (Wahlenb.) Reichenb.), вероники поручейной (Veronica beccabunga L.), вербейника монетчатого (Lysimachia nummularia L.). Структурная организация особей оценена с использованием 3 категорий модулей на основе системного подхода (Савиных, Мальцева, 2008). При обсуждении результатов использованы данные С. Е. Петровой (2010, 2012) по биоморфологии у Oe. javanica D. C., Oe. аquaticа и S. latifolium. ———— * Данное сообщение выполнено на основе анализа работ Н. П. Савиных (2006), Е. В. Лелековой (2006), Т. А. Мальцевой (2009), совместных работ: Н. П. Савиных, Т. А. Мальцевой (2008), С. В. Шабалкина, Н. П. Савиных (2011а, б). 38 Эти растения обитают в условиях с переменной влажностью и содержанием газов в разные периоды вегетационного сезона. Здесь есть представители всех жизненных форм по степени воздействия особей на среду: явнополицентрические, неявнополицентрические, моноцентрические, ацентрические. Большинству биоморф свойственна тенденция к монокарпичности исходных особей и потомков. Реализуется она поразному. 1. Без образования специализированных органов и их частей путем 1) ранней морфологической дезинтеграции у ацентрических биоморф (L. nummularia, V. beccabunga); 2) «вписывания» в среду на основе имеющихся структур (однолетние соцветия-стрелки при наличии многолетней вегетативной оси у B. umbellatus). 2. Изменение в онтогенезе побега: 1) пролонгация * – своеобразное итеративное ветвление при формировании синфлоресценции из нескольких силлептических побегов последовательных порядков ветвления, значительно повышающее семенную продуктивность у неявнополицентрических и явнополицентрических биоморф, замещающих малолетников (A. plantago-aquatica, S. sagittifolia); 2) пролонгация – полегание побегов после цветения и образование из почек средней зоны торможения монокарпических побегов силлептических боковых розеточных вегетативных побегов, в будущем – вегетативных диаспор у явнополицентрических биоморф (облигатно у R. amphibia) и переходных к ним габитусов (явнополицентрические фенобиоморфы при полегании побегов у C. palustris, Oe. aquaticа); 3) базальная девиация как образование специализированных структур для вегетативного расселения и размножения у явнополицентрических биоморф (столоны и клубни у S. sagittifolia; столоны у Oe. javanica); 4) базальная девиация – запас питательных веществ в розеточной части побега (R. amphibia). 3. Изменения корней: 1) образование корневых отпрысков и развитие из этих диаспор рамет у вегетативных малолетников и однолетников (R. рalustris, R. × anceps); 2) терминальная пролонгация – формирование вторичностержневых корневых систем в виде значительного утолщения и удлинения отдельных придаточных корней с закреплением особи на занятой территории у малолетников (R. рalustris, R. × anceps); образование корневых шишек (S. latifolium). 4. Терминальная аббревиация онтогенеза генеты и рамет: ранняя морфологическая дезинтеграция с образованием компактного (A. planta- ———— * Модусы морфологической эволюции определены в соответствии с представлениями А. Л. Тахтаджяна (1954) и Т. И. Серебряковой (1983). 39 go-aquatica, Oe. aquaticа, C. virosa, S. latifolium) или диффузного (S. sagittifolia, виды рода Rorippa) клонов. Эти изменения определяются, прежде всего, пластичностью элементарного модуля (развертывание побегов из почек без периода покоя, образование и изменение придаточных корней) с последующими изменениями универсального. Кроме того – ранней морфологической дезинтеграцией, что приводит к формированию в течение жизни раметы единственного универсального модуля и совпадению его с основным. Монокарпичность рамет обеспечивает моноцентрическим и неявнополицентрическим особям длительное существование на занятом участке при высокой энергии семенного воспроизведения. У растений с симподиальными длиннопобеговой и полурозеточной моделями, единый центр сохраняется в виде компактного клона; дочерние особи сохраняют кажущееся единство из-за большого числа придаточных корней. Это обеспечивает существование растений в условиях прибрежий. У растений явнополицентрических биоморф преобладает вегетативное образование потомков. Тенденция к монокарпичности рамет у них реализуется в полной мере. Образующиеся диаспоры автономны настолько, что совсем не зависят от материнского организма, способны и цвести, и формировать новые диаспоры на следующий год. При образовании большого числа семян семенное возобновление у этих видов подавлено, преобладает вегетативное воспроизведение. Монокарпичность рамет обеспечивает поликарпичность генеты в целом, значительную независимость особей от меняющихся условий среды. Сформировавшаяся в условиях прибрежий тенденция к монокарпичности рамет, еще в большей степени проявляется у гидрофитов. Исследование поддержано Грантом РФФИ № 13-04-01057 Л и т е р а т ур а Лелекова Е. В. Биоморфология водных и прибрежно-водных семенных растений северо-востока Европейской России: дис. … канд. биол. наук / Е. В. Лелекова. – Пермь, 2006. – 189 с. Мальцева Т. А. Биоморфология некоторых кистекорневых гигрогелофитов: дис. … канд. биол. наук / Т. А. Мальцева. – Сыктывкар, 2009. – 204 с. Петрова С. Е. Биоморфология прибрежно-водных зонтичных (на примере родов Sium L., Berula W. D. J. Roch., Cicuta L., Oenanthe L. / С. Е. Петрова // Леонид Васильевич Кудряшов. Ad me moriam: сб. ст. / ред. А. К. Тимонин. – М.: МАКС Пресс, 2012. – С. 129–140. Петрова С. Е. Пластичность биоморфы Oenanthe aquaticа (L.) Poir.) в связи с прибрежно-водной средой обитания / С. Е. Петрова, Р. П. Барыкина // Бюл. Моск. о-ва испытат. прир. Отд. биол., 2010. – Т. 115, № 5. – С. 11–19. Савиных Н. П. Род вероника: морфология и эволюция жизненных форм / Н. П. Савиных. – Киров: Изд-во ВятГГУ, 2006. – 324 с. Савиных Н. П. Модуль у растений как структура и категория / Н. П. Савиных, Т. А. Мальцева // Вестн. Твер. гос. ун-та. Сер., Биология и экология. – 2008. – Вып. 9. – С. 227–234. Серебрякова Т. И. О некоторых модусах морфологической эволюции цветковых растений / Т. И. Серебрякова // Журн. общ. биологии. – 1983. – № 5. – С. 579–592. Тахтаджян А. Л. Вопросы эволюционной морфологии растений / А. Л. Тахтаджян. – Л.: Изд-во Ленинградского ун-та, 1954. – 40 216 с. Шабалкина С. В. Строение побеговых систем некоторых видов рода Rorippa Scop. с позиции модульной организации / С. В. Шабалкина, Н. П. Савиных // Научные ведомости Белгородского государственного университета. Сер., Естественные науки. – 2011. – № 9 (104), вып. 15/1. – С. 16–22. Шабалкина С. В. О корнеотпрысковой жизненной форме Rorippa × anceps (Wahlenb.) Reichenb. (Brassicaceae) / С. В. Шабалкина, Н. П. Савиных // Вестник Тюменского государственного университета. – 2012. – № 6. – С. 118–122. Шабалкина С. В. Биоморфология Rorippa amphibia (Brassicaceae) / С. В. Шабалкина, Н. П. Савиных // Растительные ресурсы. – 2012. – Т. 48, вып. 3. – С. 315–325. ГЕНЕТИЧЕСКИЕ ОСНОВЫ СОРТОВОГО РАЗНООБРАЗИЯ ЯРОВОЙ ПШЕНИЦЫ ПРИ ДЕЙСТВИИ ПОВЫШЕННЫХ ТЕМПЕРАТУР И ЗАСУХИ Хохлова Л. П. Казанский (Приволжский) федеральный университет, г. Казань, Ludmila.Khokhlova@kpfu.ru Молекулярно-генетические механизмы различных сортов сельскохозяйственных культур при действии экстремальных факторов являются малоизученными. В то же время известно, что эти механизмы, связанные прежде всего с экспрессией генов и синтезом стрессовых белков, во многом определяют ответ растительных клеток на неблагоприятные внешние условия и тем самым на формирование фенотипической устойчивости растений в природных условиях. Почти полностью отсутствуют исследования, в которых была бы прослежена связь между экспрессией генов стрессовых белков и сортоспецифической реакцией разных по устойчивости сортов яровой пшеницы на действие повышенных температур и засухи. Эти знания необходимы для улучшения генетической адаптации и выносливости пшеницы к этим факторам, что представляет одну из главных задач региональных программ селекции в зонах с рискованным земледелием, часто страдающих от засухи. К таким регионам, несущим огромные потери урожая пшеницы вследствие ежегодно повторяющихся засух, относится Татарстан. В природных условиях, как правило, засухи и повышенные температуры действуют сопряженно, и поэтому изучение их совместного и отдельного действия на экспрессию генов позволяет приблизиться к пониманию многофакторной генной регуляции устойчивости растений и возможности поиска потенциальных молекулярных маркеров засухоустойчивости растений. Целью нашей работы являлось обнаружение генотипически детерминированной экспрессии генов трех стрессовых белков – дегидрина DHN14, серин-треонин киназы W55a и липокалина TaTIL, у отличающихся по теплоустойчивости восьми сортов яровой пшеницы при отдельном и комбинированном сочетании гипертермии и водного дефицита. Опыты проводили на листьях 7-суточных проростков пшеницы 41 (Тriticum aestivum L.). Теплоустойчивость растений тестировали по термостабильности мембран, которую характеризовали температурой порового повреждения мембран (ТППМ), величиной угла наклона термограмм и коэффициентом повреждаемости мембран. ТППМ соответствовала резкому (фазному) повышению проницаемости мембран на термограммах после 5-минутного прогрева листьев при нарастающем действии повышенных температур в диапазоне 49–56 °С (Валиуллина и др., 2009). Проведенный таким образом анализ термостабильности мембран позволил ранжировать все сорта на три группы: Омская 33 и МиС – высокоустойчивые (ТППМ – 55 °С), Дебют, Амир, Тризо, Тулайковская 10 – среднеустойчивые (ТППМ – 54 °С), Тимер и Закамская – низкоустойчивые (ТППМ – 52 °С). Растения выращивали в течение 7 суток в сосудах с почвой при 22–23 °С, фотопериод – 10 ч, влажность воздуха – 35 %, освещенность – 50 кЛк (контрольный вариант). Затем растения подвергали тепловому стрессу и засухе при различных режимах (опытные варианты): 1 – тепловой шок (ТШ, 38 °С – 30 мин, 40 °С – 30 мин, 42 °С – 2 ч); 2 – предадаптация (45 °С – 15 мин, 23 °С – 2 ч) и ТШ; 3 – засуха и ТШ (засуху создавали путем прекращения полива 5-суточных проростков в течение двух суток); 4 – засуха, предадаптация и ТШ. Экспрессию трех генов изучали, используя обратно-транскриптазную ПЦР. Выделение РНК проводили методом TPIzol, на основе РНК синтезировали кДНК с помощью специального набора («Roche», Германия) в процессе ОТ-ПЦР. Затем получали амплификаты к ДНК с помощью специфических праймеров, подобранных для DHN14, W55a и TaTIL по програме Vector NTI. Реакционную смесь использовали для электрофоретического разделения амплификатов кДНК в агарозном геле, которые визуализировали в УФ-трансиллюминаторе («Serva», Германия) при длине волны 254 нм и фотографировали ССD камерой «Nikon» (Япония) при той же длине волны. Размер амплификатов кДНК DHN14 составлял 325 п. н., W55a – 1006 п. н. и TaTIL – 688 п. н. В качестве маркера молекулярных фрагментов использовали фрагменты ДНК New England Biolabstm lkb-100 bp DNA Ladder («New England Biolabs», США). Экспрессию генов оценивали полуколичественно по ширине и интенсивности окрашивания полос амплификатов. Дегидрины (LEA-белки позднего эмбриогенеза) являются наиболее многочисленной группой гидрофильных водорастворимых белков. Экспрессия их генов происходит под действием различных стрессоров с обезвоживающим эффектом – засухи, низких температур, засоления, АБК (Сlose, 1996). Корреляцию между выносливостью растений к этим факторам и экспрессией генов дегидриновых белков наблюдали 42 у нескольких видов сельскохозяйственных культур, включая пшеницу. Было высказано предположение о протекторной роли дегидринов в стабилизации мембран и защите цитоплазматических белков от денатурации по типу шаперонов (Campbell and Close, 1997). Полученные нами результаты свидетельствуют о том, что экспрессия гена дегидрина DHN14 сохраняется у всех сортов как в контроле, так и при различных режимах температурного и водного стрессов (рис. 1). Несмотря на то, что синтез дегидринов индуцируется, главным образом, стрессорными факторами окружающей среды, однако, вероятно, определенное количество дегидринов может находиться в клетке также в нормальных условиях, то есть являться конститутивными. Поскольку применяемый нами полуколичественный метод ОТ-ПЦР является высокочувствительным, то нам удалось обнаружить сигналы об экспрессии данного гена у всех исследуемых сортов в контрольном варианте. При этом не было выявлено дифференцированной активности данного гена у исследуемых сортов пшеницы, отличающихся выносливостью к повышенным температурам. Такие результаты указывают на необходимость применения количественного анализа для установления различных уровней экспрессии гена DHN14 при заданных условиях. И, кроме того, возможно проявление специфики генов различных дегидринов, так как в работе (Lopez et al., 2003) была установлена связь между накоплением общих дегидринов и приобретением сортоспецифической устойчивости проростков семи сортов озимой пшеницы в течение нарастающего воздействия засухи. Рисунок 1 – Электрофореграммы транскриптов гена DHN14 в листьях разных сортов яровой пшеницы: К – контроль (23 °С); 1– тепловой шок – ТШ (38 °С – 30 мин, 40 °С – 30 мин, 42 °С – 2 ч); 2 – предадаптация (45 оС – 15 мин, 23 оС – 2 ч) и ТШ; 3 – засуха и ТШ; 4 – засуха, предадаптация и ТШ 43 У эукариотов, включая растения, обратимое фосфорилирование белков с участием протеинкиназ/протеинфосфатаз является центральным звеном в восприятии и трансдукции стрессоров окружающей среды и составляет основной механизм контроля клеточных функций при ответах растений на неблагоприятные условия. Существует более 600 генов, кодирующих протеинкиназы в геноме Arabidopsis (Morris, Walker, 2003). Важная роль принадлежит серин/треонин протеинкиназам (SnRKs), разделенным на три подсемейства – SnRK1, SnRK2, SnRK3 (Hrabak et al., 2003). В течение нескольких лет основное внимание было уделено подсемействам SnRK1 и SnRK3, однако мало известно о специфической для растений подгруппе SnRK2. Было сделано предположение, что данная подгруппа протеинкиназ участвует в восприятии изменений окружающей среды (Shen et al., 2001). Недавно был изолирован из пшеницы новый ген W55a этого семейства протеинкиназ, содержащий 11 стандартных для протеинкиназы каталитических доменов, а также сер/тре активный сайт (Zhao-Shi Xu et. al., 2009). Авторы показали, что транскрипты W55a накапливались в листьях проростков при засухе и солевом стрессе, что указывает на участие W55a в ответе пшеницы на абиотический стресс. Более того, усиленная экспрессия этого гена, введенного в геном Arabidopsis, повышала выживаемость трансгенных растений и вызывала более интенсивное развитие корней в условиях имитации засухи. В связи с этим, авторы сделали вывод, согласно которому ген W55a может повышать уровень засухоустойчивости путем стимуляции роста и развития корней. В другой работе (Xinguo Mao et al., 2010), чтобы выяснить функции протеинкиназ SnRK2, были получены трансгенные растения арабидопсиса, сверхэкспрессирующие TaSnRK 2.4 пшеницы. При этом наблюдали увеличение выносливости трансгенных растений к засухе, засолению и замораживанию, что подтверждалось физиологическими результатами – уменьшением скорости потери воды, повышением относительного содержания воды, стабильности клеточных мембран, увеличением фотосинтетического и осмотического потенциалов, а также отсутствием отрицательного влияния на рост и урожай растений. По мнению авторов, TaSnRK 2.4 – многофункциональный регуляторный белок, который можно использовать в трансгенной селекции для улучшения выносливости сельскохозяйственных культур к абиотическим стрессам. Проведенный в нашей работе анализ экспрессии гена серин/треонин киназы W55a показал, что у всех восьми сортов транскрипты присутствуют в норме (рис. 2, контроль). У сортов Мис, Амир, Тризо и Тулайковская 10 активность гена сохраняется как при контрольных, так и при всех стрессовых условиях, тогда как у сортов Омская 33, Тимер и Закамская экспрессия отсутствует при действии засухи и теплового шока 44 (вариант 3). Выявлено, что у сорта Дебют при предадаптации и ТШ (вариант 2) и при сочетании этих факторов на фоне засухи (вариант 4) экспрессия гена серин/треонин киназы W55a также не наблюдалась. В то же время влияние ТШ как на фоне засухи, так и без нее (варианты 1 и 3, соответственно) вызывало экспрессию этого гена. В связи с этим можно заключить, что именно предадаптация проростков к действию гипертермии приводит к выключению транскрипции гена W55a у данного сорта. Выявленные в экспериментах особенности гена W55a могут указывать на то, что у сортов Омская, Тимер и Закамская тепловой шок на фоне засухи блокирует активность этого гена, а у сорта Дебют сходный эффект отмечен в результате предадаптации. Известно, что температурное воздействие приводит к изменению активности стартовых ферментов сигнальных систем, большинство из которых локализовано в плазмалемме (Тарчевский, 2002). Поскольку исследуемая нами серин/треонин киназа W55a локализуется в мембране (Zhao-Shi Xu et al., 2009), то, вероятно, происходящая при действии абиотических стрессов сортоспецифическая реорганизация белково-липидных комплексов мембран приводит к дифференцированному нарушению экспрессии данного гена, вследствие чего отсутствие активности гена W55a может вызывать изменения в передаче различных сигналов внутри клетки у сортов Омская 33, Дебют, Тимер и Закамская. Рисунок 2 – Электрофореграммы транскриптов гена W55a в листьях разных сортов яровой пшеницы: К – контроль (23 °С); 1– тепловой шок – ТШ (38 °С – 30 мин, 40 °С – 30 мин, 42 °С – 2 ч); 2 – предадаптация (45 оС – 15 мин, 23 оС – 2 ч) и ТШ; 3 – засуха и ТШ; 4 – засуха, предадаптация и ТШ 45 Рисунок 3 – Электрофореграммы транскриптов гена TaTIL в листьях разных сортов яровой пшеницы: К – контроль (23 °С); 1– тепловой шок – ТШ (38 °С – 30 мин, 40 °С – 30 мин, 42 °С – 2 ч); 2 – предадаптация (45 оС – 15 мин, 23 оС – 2 ч) и ТШ; 3 – засуха и ТШ; 4 – засуха, предадаптация и ТШ В литературе практически полностью отсутствуют сведения по обнаружению экспрессии генов еще одной важной группы стрессовых белков – липокалинов среди разных по устойчивости сортов яровой пшеницы. Липокалины широко распространены у животных, насекомых и бактерий. У растений известны два температурозависимых липокалина – у пшеницы – TaTIL и арабидопсиса – AtTIL (Frenette-Charron, Breton, 2002). Первоначально липокалины считались транспортными белками, но затем появились данные о том, что они могут быть вовлечены в выполнение многих других важных функций, например, регуляция клеточного роста, метаболизма и иммунного ответа, связывание с клеточными рецепторами, биогенез и восстановление поврежденных мембран, а также индукция апоптоза. Появление транскриптов TaTIL у пшеницы является тканеспецифичным, так как их накопление при воздействии стрессорных факторов происходило в листьях, в формирующихся семенах и в вегетирующих растениях (Rock, 2000). Поскольку липокалины относятся к стероид-связывающим белкам, то TaTIL может участвовать в транспорте молекул стерола к мембране, что способствует сохранению упорядоченности фосфолипидов во время ответа на действие высоких температур (Demel, De Kruiy, 1976). На рисунке 3 представлены электрофореграммы транскриптов гена липокалина TaTIL разных сортов яровой пшеницы. Сигналы гена появлялись в контроле и сохраня46 лись во всех вариантах. По данным литературы, экспрессия гена липокалина в большей степени обнаруживается при действии теплового шока, чем водного дефицита. Во всех опытных вариантах нашего эксперимента присутствовал тепловой шок, и поэтому в данном случае трудно разделить действие этих двух факторов. Полученные результаты указывают на отсутствие каких-либо заметных различий в экспрессии гена липокалина среди исследуемых сортов пшеницы. Таким образом, при изучении экспрессии генов белков – потенциальных участников стрессовых ответов – дегидрина DHN14, серин/треонин киназы W55a, липокалина TaTIL у восьми сортов яровой пшеницы с разным уровнем теплоустойчивости установлено, что транскрипты этих генов обнаружены у растений всех сортов, выращенных при нормальной температуре, и это указывает на конститутивный характер экспрессии исследуемых генов. Однако по многочисленным данным, имеющимся в литературе, их усиленная экспрессия в трансгенных растениях повышала уровень устойчивости к неблагоприятным факторам, вызывающим обезвоживание – засухе, повышенным и низким температурам, засолению. Как показали результаты наших экспериментов, сортовая специфика отмечена только для гена серин/треонин киназы W55a, выключение транскрипции которого вызывала предадаптация растений к действию гипертермии (теплового шока) только у среднеустойчивого сорта, а в условиях теплового шока на фоне засухи блокировка активности этого гена отмечена как для устойчивых (Омская 33), так и неустойчивых (Тимер, Закамская) сортов. В то время как два других гена дегидрина DHN14 и липокалина TaTIL экспрессировались при разных комбинациях температурного и водного режимов у всех сортов. Исходя из представленных в работе результатов впервые проведенных исследований экспрессии генов дегидрина DHN14, серин/треонин киназы W55a и липокалина TaTIL у восьми сортов яровой пшеницы с разным уровнем устойчивости, можно заключить, что изученные гены являются конститутивными и функционируют как в норме, так и при стрессовых режимах температурного и водного факторов. Однако наибольшей чувствительностью к воздействующим факторам, вплоть до полного выключения из общей скоординированной генной регуляции защитных систем, характеризуется ген серин/треонин киназы W55a, являющийся важным звеном клеточного сигналинга. Л и т е р а т ур а Валиуллина Р. Н. Мембранные механизмы устойчивости разных сортов яровой пшеницы к повышенным температурам / Р. Н. Валиуллина, В. В. Рябовол, Л. П. Хохлова // Актуальные проблемы сельскохозяйственной науки и практики в современных условиях и пути их решения: материалы Всероссийской научно-практической конференции молодых ученых. – Казань, 2009. – С. 69–73. Тарчевский И. А. Сигнальные системы клеток расте47 ний / И. А. Тарчевский. – М.: Наука, 2002. – 294 с. Campbell S. A. Dehydrins: genes, proteins and association with phenotypic traits / S. A. Campbell, T. Y. Close // New Phytol. – 1997. – Vol. 137. – P. 61–74. Close T. J. Dehydrins: Emergence of a biochemical role of a family of plant a biochemical role of a family of plant dehydration proteins / T. J. Close // Physiologia Plantarum. – 1996. – Vol. 97. – P. 795–803. Demel R. A. Consequences of the interaction of calcium with dioleoylphosphatidate-containing model membranes: calcium-membrane and membrane-membrane interactions / R. A. Demel, B. De Kruij // Biochim. Biophys. Acta. – 1976. – Vol. 457. – P. 109–132. Frenette-Charron J.-B. Molecular and structural analyses of a novel temperature stress-induced lipocalin from wheat and Arabidopsis / J.-B. Frenette-Charron, G. Breton // Elsevier Science B. V. – 2002. – Vol. 517. – P. 129–132. Hrabak E. M. The Arabidopsis CDPK-SnRK superfamily of protein kinases / E. M. Hrabak, C. W. Chan, M. Gribskov // Plant Physiol. – 2003. – Vol. 132. – P. 666–680. Lopez Cesar G. et al. Dehydrin Expression and Drought Tolerance in Seven Wheat Cultivars / Cesar G. Lopez, Gary M. Banowetz, C. James Peterson, and Warren E. Kronstad // Crop Sci. – 2003. – Vol. 43. – P. 577–582. Morris E. R. Receptor-like protein kinases: the keys to response / E. R. Morris, J. C. Walker // Curr. Opin. Plant Biol. – 2003. – Vol. 6. – P. 339–342. Rock C. D. Pathways to abscisic acidregulated gene expression / C. D. Rock // New Phytologist. – 2000. – Vol. 148. – P. 357– 396. Shen W. The salt stress-inducible protein kinase gene, Esi47, from the salt-tolerant wheatgrass Lophopyrum elongatum is involved in plant hormone signaling / W. Shen, A. G´omezCadenas, E. L. Routly // Plant Physiol. – 2001. – Vol. 125. – P. 1429–1441. Xinguo Mao. TaSnRK 2.4, an NF1-type serine/threonine protein kinase of wheat (Triticum aestivum L.), confers enhanced multistress tolerance in Arabidopsis / Mao Xinguo, Zhang Hongying, Tian Shanjun, Chang Xiaoping and Jing Ruilian // Journal of Experimental Botany. – 2010. – Vol. 61. – P. 683–696. Zhao-Shi X. W55a Encodes a Novel Protein Kinase That Is Involved in Multiple Stress Responses / X. Zhao-Shi, L. Li, N. Zhi-Yong // Journal of Integrative Plant Biology. – 2009. – Vol. 51. – P. 58–66. СОХРАНЕНИЕ И ИЗУЧЕНИЕ БИОРАЗНООБРАЗИЯ ДИКОРАСТУЩИХ ВИДОВ РОДА HORDEUM L. В КОЛЛЕКЦИИ ВИР Чернов В. Е., Пендинен Г. И. Всероссийский научно-исследовательский институт растениеводства им. Н. И. Вавилова, vechernov@mail.ru Снижение биологического разнообразия среди сортов сельскохозяйственных растений приводит как известно к ускорению процессов деградации признаков сорта, снижению качества урожая и его количественных показателей (Nevo, 1992). Одним из возможных способов получения исходного материала растений для селекции растений является интрогрессия генетического материала дикорастущих видов, сородичей культурных растений (Пендинен, Чернов, 2005). Ячмень является третьей по значимости злаковой культурой, а для северных регионов лидирует в первой тройке после ржи. Однако, в связи стем, что культурный ячмень является в северных регионах строгим самоопылителем, статус биоразнообразия этой культуры значительно снижен. Использование трансформации, или прямой генетической модификаци, для ячменя неэффективно, что связано с низкой регенерационной способно48 стью этой культуры in vitro (Чернобровкина, Сидоров, 2007). Одним из возможных способов повышения биоразнообразия культурного ячменя является использование в гибридизации дикорастущих видов Hordeum. Наиболее эффективным в настоящее время является использование дикорастущего подвида Hordeum vulgare ssp. spontaneum (С. Koch.) Thell., а также остающихся в природе случайных высевов культурных сортов (rewild barley). Для вовлечения в селекционный процесс других видов, более отдаленных систематически, необходимы всесторонние исследования этих видов, генетики их совместимости с культурным ячменем и изучения физиологических параметров дикорастущих видов рода Hordeum. Следует отметить, что физиологические процессы дикорастущих видов яченя изучены мало, в основном это физиологические показатели устойчивости к некоторым неблагоприятным факторам среды. Базовые физиологические параметры дикорастущих видов Hordeum не изучались. Для сохранения и изучения дикорастущих видов рода Hordeum в ВИРе создается и поддерживается коллекция этих видов, состоящая из семенной и клоновой составляющих. Род Hordeum L. относится к трибе Triticeae Dum. cемейства злаков Poaceae (Цвелев, 1976). Наиболее современной является классификация рода Hordeum, разработанная Ботмером с соавторами. Род Hordeum, согласно классификации Ботмера с соавторами, включает 4 секции: Hordeum, Anisolepis, Critesion, Stenostachys (Bothmer, Jacobsen, 1985; Bothmer, Jacobsen, Baden et al., 1991; Jacobsen, Bothmer, 1992). Среди представителей рода Hordeum встречаются виды с однолетним (H. vulgare, H. murinum L., H. marinum Hudson) и многолетним (H. bulbosum L., H. jubatum L., H. chilense Roemer&Schultes, H. secalinum Schreber и др.) образом жизни. Часть видов являются самонесовместимыми перекрестноопыляемыми формами (H. bulbosum L., H. Brevisubulatum (Trinius) Link), большинство видов самосовместимы с преобладанием самоопыления (H. marinum, H. murinum, H. patagonicum (Haumann) Covas и др.). Различные образцы видов характеризуются озимым или яровым типом развития. Не все озимые виды зимостойки. Для ряда видов существует проблема всхожести семян, полученных при культивировании растений в условиях, отличающихся от естественных в ареалах их произрастания. Для эффектвного изучения биологических особенностей дикорастущих ячменей необходимо их стабильное культивирование в различных условиях проводимых экспериментов. Как правило, цель культивирования дикорастущих ячменей ограничивается размножением семенного материала для получения новых семенных репродукций. Однако для проведения репрезентативных физиологических, генетических экспериментов, а также 49 морфометрических измерений необходимо культивирование относительно большого числа растений (из природного популяционного сбора), в связи с чем необходимо изучение и оптимизация параметров культивирования видов рода Hordeum в контролируемых искусственных условиях. В Лаборатории биотехнологии ВИР создана коллекция дикорастущих видов Hordeum, охватывающая видовое разнообразие рода, включающая виды с однолетним и многолетним образом жизни, с различными системами размножения: самосовместимые формы с преобладанием самоопыления, самонесовместимые перекресноопыляемые виды. В Лабораториях биотехнологии и молекулярной и экологической генетики ВИР проводится многолетнее изучение рода Hordeum с целью определения оптимальных методов и условий культивирования дикорастущих видов ячменя в полевых условиях, в почвенной культуре, в условиях лабораторного эксперимента in vivo и in vitro. Наиболее интенсивно изучались особенности культивирования дикорастущих видов в культуре тканей, что связано с разработкой технологий культивирования, размножения и межвидовой гибридизации видов Hordeum с культурным ячменем для интрогрессии полезных признаков дикорастущих видов в геном H. vulgare. В результате исследований установлено влияние соотношения форм азота в питательных средах на интенсивность роста недифференцированных тканей дикорастущих видов in vitro, что, возможно, связано с различной интенсивностью поглощения разных форм азота различными видами Hordeum in vivo. Показано, что виды секции Hordeum рода Hordeum имеют однотипную структуру каллусной ткани и низкую эффективность регенерации по сравнению с видами других секций Anisolepis, Critesion, Stenostachys этого рода. Эти особенности учитывались при получении и микроклональном размножении межвидовых гибридов H. vulgare c дикорастущими видами, а также при микроклонировании образцов с низкой семенной продуктивностью (Суриков, Чернов, Пендинен, 1992). При культивировании дикорастущих видов ячменя необходимо учитывать фотопериодическую активность различных видов и отдельных образцов ячменя. Проведенные нами исследования выявили значительную чувствительность большинства видов Hordeum к длине светового дня. Изучались также почвенные условия в местах природного расселения дикорастущих видов, а также культивирование в условиях моделирующих экстремальные почвенные показатели. Установлено, что природные почвенные условия, в частности кислотность почвы, довольно близки в ряде географически удаленных друг от друга районах обитания дикорастущих видов ячменя. Более того, показано, что культивирование дикорастущих видов на искусст50 венном торфосодержащем субстрате, при поливе раствором Хогланда с рН = 4,6 не вызывает каких-либо летальных изменений у растений дикорастущих видов, таких как H. spontaneum, H. murinum, H. secalinum. В результате исследований разработаны методики сохранения и размножения для видов Hordeum c различными биологическими характеристиками, позволяющие сохранить биологическое разнообразие рода, сочетая культивирование в искусственных условиях экспериментального поля, в условиях закрытого грунта, в лабораторных условиях in vivo и in vitro. Они позволяют стабильно культивировать все дикорастущие виды рода Hordeum, получая либо семенные поколения, либо сохраняя образцы в виде клонов. Л и т е р а т ур а Пендинен Г. И. Цитогенетические механизмы, обеспечивающие интрогрессию селекционно ценных генов у злаков / Г. И. Пендинен, В. Е. Чернов // Идентифицированный генофонд растений и селекция. – СПб., 2005. Суриков И. М. Использование дикорастущих видов Hordeum в межвидовых скрещиваниях с культурным ячменем / И. М. Суриков, В. Е. Чернов, Г. И. Пендинен // Труды по прикладной ботанике, генетике и селекции. – СПб., 1992. – Т. 148. – С. 18–28. Цвелев Н. Н. Злаки СССР / Н. Н. Цвелев. – Л.: Наука, 1976. Чернобровкина М. А. Влияние параметров биолистической трансформации ярового ячменя (Hordeum vulgare L.) на уровень транзиентной экспрессии репортерного гена GFP / М. А. Чернобровкина, Е. А. Сидоров, И. А. Баранов, П. Н. Харченко, С.В. Долгов // Известия РАН, Сер., Биология. – 2007. – № 6. – С. 669–675. Bothmer R. An Ecogeographical Study of the Genus Hordeum / R. Bothmer, N. Jacobsen, C. Baden, R. B. Jørgesnen, I. LindeLaursen. – IBPGR. – Rome, 1991. – 127 p. Jacobsen N. Supraspecific groups in the genus Hordeum / N. Jacobsen, R. Bothmer // Hereditas. – 1992. – Vol. 116, № 1–2. – P. 21–24. Nevo E. Origin, evolution ,population genetics and resources for breeding of wild barley / E. Nevo // Barley: Genetics, Biochemistry, Molecular Biology and Biotechnology. – Bristol, UK. CAB International, 1992. – Р. 19–43. 51 Секция 1 БИОРАЗНООБРАЗИЕ ЖИЗНЕННЫХ ФОРМ ЖИВЫХ ОРГАНИЗМОВ В НАЗЕМНЫХ И ВОДНЫХ ЭКОСИСТЕМАХ КЛАССИФИКАЦИЯ И ЭКОЛОГИЧЕСКОЕ РАЗНООБРАЗИЕ ФИТОЦЕНОЗОВ ПОДЗОНЫ ТЕМНОХВОЙНЫХ ЛЕСОВ ЧЕЛЯБИНСКОЙ ОБЛАСТИ Гетманец И. А.1, Москвина И. В.2 1 Челябинский государственный университет, г. Челябинск, igetmanec@mail.ru 2 МБОУ СОШ № 9, г. Куса, kusa_crbomk@mail.ru Исследования были проведены в Кусинском районе Челябинской области, относящемся по ботанико-географическому районированию к лесной зоне, подзоне хвойно-широколиственных и южно-таежных хвойных лесов западного склона Урала, Катав-Златоустовскому району широколиственно-темнохвойных и сосново-березовых лесов, Юрюзанско-Златоустовскому подрайону сосново-березовых лесов (Колесников, 1961). Подзона хвойно-широколиственных и южно-таежных хвойных лесов западного склона Урала представляет собой горный аналог подзон хвойно-широколиственных лесов и южной тайги Восточно-Европейской равнины и Предуралья. Катав-Златоустовский район широколиственно-темнохвойных и сосново-березовых лесов расположен на западном макросклоне Южного Урала, занимая нижний пояс хребтов и полосу предгорий в интервале высот от 250–300 до 650–700 м н. у. м. Растительный покров района, отличающийся значительным разнообразием и неоднородностью, представлен широким набором лесных, луговых, болотных и степных ценозов. Юрюзанско-Златоустовский подрайон характеризуется хребтовоувалистым рельефом со средними высотами 400–700 м н. у. м., расчлененными глубокими речными долинами. Согласно материалам Л. А. Соколовой (1951) и других авторов, наибольшая роль в сложении растительного покрова подрайона в границах Челябинской области из хвойных лесов принадлежит сосновым, лиственнично-сосновым и смешанным липово-сосновым лесам. 52 В связи с большой расчлененностью рельефа и разнообразием климатических условий наблюдается пестрый по составу почвенный покров. Под хвойными и смешанными лесами преобладают горные серые лесные почвы. Под массивами смешанных лесов с травянистомоховым покровом встречаются горные дерново-подзолистые. При анализе геоботанического материала исследованной территории Кусинского района с использованием сайта http://mfd.cepl.rssi.ru/flora/ princip.htm, согласно доминантной классификации, выделено четыре группы типов леса: ельники мелкотравные (неморально-бореальные), ельники травяные неморальные, сосняки сложные (бореальнонеморальные), широколиственные леса травяные неморальные. Перечисленные фитоценозы по эколого-флористической классификации относятся к ассоциациям, приведенным в таблице. Группы типов леса и ассоциации, характерные для фитоценозов Кусинского района Доминантная классификация (приведены группы типов леса) Эколого-флористическая классификация Ельники мелкотравные (неморально-бореальные) Piceetaparviherbosa (nemoralo-borealia) Quercoroboris – Piceetumabietis Ельники травяные неморальные Piceetanemoraloherbosa Quercoroboris – Tilietumcordatae Сосняки сложные (неморально-бореальные) Pineta sylvestris composite (nemoralo-borealia) Quercoroboris – Pinetum sylvestris Широколиственные леса травяные неморальные QuercoTilietanemoraloherbosa Quercoroboris – Tilietumcordatae Таким образом, в нашей работе мы воспользовались двумя основными традиционными подходами к классификации фитоценозов: доминантным (физиономическим) и эколого-флористическим. Для наших исследований наиболее подходящим является первый подход, т. к. речь идет о лесной растительности бореальных, суббореальных и неморальных лесов, где в составе сообществ присутствуют явные доминанты с сильно выраженными эдификаторными свойствами. Использование второго подхода, основанного на группировке сообществ в соответствие с подобием флористических составов и отражающего экологические условия и стадию сукцессии, также целесообразно вследствие того, что на исследуемой территории в результате хозяйственной деятельности человека коренные сообщества сменились или сменяются производными. 53 Выделение семи ассоциаций, на наш взгляд, связано с большим разнообразием экологических режимов местообитаний лесных фитоценозов данного района и воздействием на них антропогенного фактора. Л и т е р а т ур а Колесников Б. П. Очерк растительности Челябинской области в связи с ее геоботаническим районированием / Б. П. Колесников // Флора и лесная растительность Ильменского государственного заповедника им. В. И. Ленина: тр. Ильменск. гос. заповед. – Свердловск: УФАН СССР, 1961. – Вып. 8. – С. 105–129. Куликов П. В. Конспект флоры Челябинской области (сосудистые растения) / П. В. Куликов. – Екатеринбург; Миасс: Геотур, 2005. – 537 с. Соколова Л. А. Основные черты растительности западного склона (северной части) Южного Урала / Л. А. Соколова // Тр. ботан. ин-та им. В. Л. Комарова АН СССР. – 1951. – Сер. 3, вып. 7. – С. 134–180. – Режим доступа: http://mfd.celp.rssi.ru/flora/princip.htm. ПРЕДВАРИТЕЛЬНЫЕ МАТЕРИАЛЫ О ПАРАЗИТАРНЫХ ИНВАЗИЯХ У ПРЕДСТАВИТЕЛЕЙ ГЕРПЕТОФАУНЫ РЕСПУБЛИКИ МАРИЙ ЭЛ Дробот Г. П., Ведерников А. А., Ремизова О. С., Мальцева Н. Л., Свинин А. О. Марийский государственный университет, г. Йошкар-Ола, droga59@mail.ru Амфибии и рептилии – чуткие биоиндикаторы трансформации экосистем и отдельных их частей. Они реагируют на изменения экосистем сдвигами в структуре популяций, фенологии, численности, морфофункциональных показателей. Поскольку одной из первых на действие неблагоприятных факторов среды реагирует кровь, гематологические параметры пригодны для оценки состояния животных в условиях загрязнения (Жукова, Пескова, 1999; Дробот, Мальцева, Ведерников, 2011). В ходе проведенных нами ранее исследований (Дробот, Ремизова, 2012) по изучению цитологической картины крови ящерицы прыткой (Lacerta agilis (Linnaeus, 1758)), обитающей в биотопах с различной антропогенной нагрузкой, в цитоплазме эритроцитов были выявлены «образования» удлиненной формы, напоминающие кровепаразитов. По данным Д. Б. Васильева (2003, 2005), среди внутриклеточных паразитов крови у рептилий наиболее распространенными являются представители трех семейств – Haemogregarinidae, Hepatozoidae и Karyolisidae. Обнаруженные нами в эритроцитах ящерицы прыткой организмы по косвенным признакам совпадают с кровепаразитами ящериц, описанными другими авторами (Хайрутдинов, Соколина, 2007). Внешне все эти паразиты сходны по форме. Как правило, имеют колбасовидное или слегка изогнутое тело, лишенное пигмента, крупное, центрально или терминально расположенное ядро седловидной формы. Наблюдаемые нами паразиты характеризуются вытянутым полупрозрачным телом, лишен54 ным какой-либо капсулы. При этом оболочка паразита вступает в прямой контакт с ядром эритроцита. Внешние морфологические признаки позволяют предположить, что имеет место инвазия кровяным паразитом Karyolysus lacertae (Danilewsky, 1889), характерным для семейства настоящие ящерицы (Lacertidae). В эритроцитах периферической крови ящериц стадии развития кариолизуса определяются как трофозоиты и гамонты. При этом оболочка паразита вступает в прямой контакт с ядром эритроцита, которое в этом случае смещено в сторону от своего обычного положения. Нередко оно как бы расплющивается телом гамонта вдоль длинной оси эритроцита. Эти изменения, как полагают И. З. Хайрутдинов и Ф. М. Соколина (2007), – следствие токсического, кариолитического и механического воздействия паразита и, возможно, результат интенсификации жизнедеятельности ядра-прокормителя. Обнаружение нами кровяного паразита позволило думать о возможном влиянии фактора наличия паразита в эритроцитах крови ящериц на структуру лейкоцитарной формулы (Дробот, Ремизова, 2012) с учетом обитания животных в различных по антропогенной нагрузке биотопах. Изучали мазки крови прытких ящериц (Lacerta agilis (Linnaeus, 1758)), которых отлавливали в мае 2010 г. в различных биотопах на территории Республики Марий Эл: 1. ООПТ Государственный природный заповедник «Большая Кокшага» (60 км от г. Йошкар-Олы; «условно чистая» зона) (12 особей); 2. Промышленная окраина г. Йошкар-Олы (территория, прилегающая к ЗАО «Мясокомбинат» и заводу «Контакт») (3 особи); 3. Окраина пгт. Куженер (8 особей). Местообитания 2 и 3 зоны с антропогенной нагрузкой. Мазки крови готовили и окрашивали по стандартным методикам (Лабораторные методы исследования в клинике, 1987). Для обработки полученных результатов (анализ структуры лейкоформулы и зараженности ящериц) применяли однофакторный дисперсионный анализ, в ходе которого установлено, что фактор «наличие паразитов» в эритроцитах крови ящерицы прыткой является значимым для показателей: доля нейтрофилов у самок (Р = 0,000136) и самцов (Р = 0,000808) и доля эозинофилов у самок (Р = 0,032098). Установлено, что доля нейтрофилов у самок и самцов, зараженных кровяным паразитом, в 1,4 больше, чем у здоровых особей. Также выявлены различия по доле эозинофилов: у самок, зараженных гемопаразитом, этот показатель в 1,2 раза меньше, чем у особей, в крови которых гемопаразит не обнаружен. Дополнительно для определения статистической связи между фактором наличия паразита в эритроцитах крови ящерицы прыткой и 55 параметрами лейкоцитарной формулы нами был проведен корреляционный анализ, который показал, что данный фактор высоко скоррелирован только с показателем «процентное содержание нейтрофилов» (r = 0,654831; Р = 0,000697). Следовательно, можно предположить, что увеличение доли нейтрофилов и сокращение доли эозинофилов напрямую связано с зараженностью рептилий кровяным паразитом. Высокий процент нейтрофилов по сравнению с эозинофилами можно объяснить тем, что в отличие от млекопитающих, у которых показана высокая антипаразитарная активность как раз эозинофилов, у ящериц нейтрофилы также обладают этой способностью и кроме того высокой противовоспалительной активностью (Горышина, Чага, 1990). Наибольший процент зараженных гемопаразитом особей ящериц был отмечен в выборках из заповедника – 66,6 %. В пгт. Куженер он составил 37,5 %, в то время как на территории г. Йошкар-Олы не было обнаружено ни одной зараженной особи. Промышленная зона г. Йошкар-Олы характеризуется низкой плотностью прыткой ящерицы, это зона с высокой антропогенной нагрузкой, возможно, этот факт неблагоприятно отражается на жизнедеятельности как самих ящериц, так и клещей, способствующих их заражению. В изученных местообитаниях (исключение пгт. Куженер) наряду с ящерицами нами были изучены особи лягушки озерной (Rana (Pelophylax) ridibunda (Pallas, 1771)), в эритроцитах у которых также были обнаружены внутриклеточные гемопаразиты. Они в соответствии с описанной морфологией (Amphibian medicine and captive husbandry, 2001) напоминают гемогрегарин, чью видовую принадлежность еще предстоит определить. Следует отметить, что вышеуказанные случаи зараженности ящериц и амфибий гемопаразитами на территории Республики Марий Эл установлены нами впервые. Также ранее нами были описаны случаи гранулематозного воспаления в печени и кишечнике у особей лягушки озерной, которые первоначально были приняты за проявления саркоидоза из-за схожести морфологической картины (Ведерников, Дробот, 2012). Как показали недавние исследования, гранулематозное воспаление этих органов, возможно, было вызвано паразитарной инвазией, так как на некоторых препаратах в очагах воспаления было отмечено наличие округлых образований, напоминающих яйца или редии трематод (Ведерников, Дробот, 2013). Дальнейшее исследование данной патологии продолжается. При анализе зараженности популяций амфибий в изученных местообитаниях была установлена сходная, как и в случае с ящерицами, кар56 тина – наиболее выраженное паразитарное инвазирование внутриклеточными гемопаразитами характерно для особей, обитающих на «условно чистой» территории заповедника, в то время как процент зараженных особей лягушек из промышленных окраин города Йошкар-Олы существенно меньше. Подобные результаты согласуются с литературными данными, в которых указывается, что увеличение загрязнения местообитаний, может приводить, как правило, к уменьшению численности паразитических организмов (Беэр, 2002). Поскольку представленные выше данные получены сравнительно недавно и являются в некотором роде предварительными, то подробное изучение разнообразия и видовой принадлежности паразитов герпетофауны Республики Марий Эл, а также связанных с ними патологических состояний является предметом дальнейших исследований. Л и т е р а т ур а Беэр С. А. Паразитизм и проблема биоразнообразия / С. А. Беэр // Теоретические и прикладные проблемы паразитологии. – М.: Наука, 2002. – С. 25–36. Васильев Д. Б. Ветеринарная герпетология: ящерицы / Д. Б. Васильев. – М.: Проект-Ф, 2005. – 408 с. Васильев Д. Б. Паразитарные болезни рептилий в условиях многовидовой ассоциации зоопарка / Д. Б. Васильев // Материалы XI Mеждунар. вет. конгресса. – М., 2003. – C. 203–205. Ведерников А. А. Гранулематозное воспаление в печени и кишечнике амфибий, обитающих на территории Республики Марий Эл / А. А. Ведерников, Г. П. Дробот // Симбиоз-Россия 2013: сб. тезисов VI Всерос. с международным участием Конгресса молодых ученыхбиологов. – Иркутск: Аспринт, 2013. – С. 331–332. Ведерников А. А. О случае саркоидоза у бесхвостых амфибий на территории Республики Марий Эл / А. А. Ведерников, Г. П. Дробот // Вопросы герпетологии: материалы Пятого съезда Герпетологического общества им. А. М. Никольского, Минск, 24–27 сентября 2012 г. – Минск: ИООО «Право и экономика», 2012. – С. 46–48. Горышина Е. Н. Сравнительная гистология тканей внутренней среды с основами иммунологии / Е. Н. Горышина, О. Ю. Чага. – Л.: Изд-во ЛГУ, 1990. – 318 с. Дробот Г. П. Ответная реакция некоторых тканей лягушки озерной (Rana Ridibunda Pallas, 1771) на антропогенную нагрузку / Г. П. Дробот, Н. Л. Мальцева, А. А. Ведерников // Вестник Оренбургского государственного университета. – 2011. – № 12 (131). – С. 65–67. Дробот Г. П. Использование гематологических показателей Lacerta agilis L. для оценки антропогенно нарушенных территорий / Г. П. Дробот, О. С. Ремизова // Известия Самарского научного центра Российской академии наук. – 2012. – Т. 14, № 1 (8). – С. 1887–1890. Жукова Т. И. Реакция крови бесхвостых амфибий на пестицидное загрязнение / Т. И. Жукова, Т. Ю. Пескова // Экология. – 1999. – № 4. – С. 288–292. Лабораторные методы исследования в клинике / под ред. В. В. Меньшикова. – М.: Медицина, 1987. – 361 с. Хайрутдинов И. З. Зараженность ящерицы прыткой Lacerta agilis L. гемогрегариной Karyolysus lacerate (Danilewsky, 1889) / И. З. Хайрутдинов, Ф. М. Соколина // Ученые записки Казанского гос. ун-та. – 2007. – Т. 149, кн. 1: Естественные науки. – С. 103–112. Amphibian medicine and captive husbandry / Kevin M. Wright and Brent R. Whitaker, editors // Original ed. Printed and Published by Krieger Publishing Company Krieger Drive Malabar, Florida 32950. – 2001. – Р. 190–221. 57 АНАТОМИЧЕСКОЕ СТРОЕНИЕ НАДЗЕМНЫХ ПОБЕГОВ ЗВЕРОБОЯ ПРОДЫРЯВЛЕННОГО (HYPERICUM PERFORATUM L.) ПОЛУКУСТАРНИЧКОВОЙ ЖИЗНЕННОЙ ФОРМЫ Дубровная С. А., Мавлюдова Л. У. Казанский (Приволжский) Федеральный университет, г. Казань, o_v_i@pochta.ru Изучение изменчивости габитуса растений в различных экологоценотических условиях способствует познанию процессов их адаптации к изменяющимся условиям среды. Изменчивость внешнего облика определяется изменением формы роста и способом нарастания побегов, что в отдельных случаях ведет к формированию новой жизненной формы. Проявление той или иной жизненной формы следует рассматривать как проявление «нормы реакции» вида, формирующейся под воздействием факторов среды. Изучение биологии зверобоя продырявленного в различных типах растительности выявило своеобразие внешнего облика, способствующего выживанию, размножению и удержанию занятой территории. У зверобоя продырявленного формируются длиннокорневищные, стержнекорневые, корневищно-стержневые, стержнекорневые корнеотпрысковые жизненные формы (Гонтарь, Годин, 2002; Пахоменко, 2012). Согласно анализу литературных источников (Шишкин, Бобров, 1949) в роде зверобой из 51 вида 27,4 % не являются травами. Это кустарники, кустарнички. Положив в основу классификации особенность строения надземных и подземных побегов и особенность строения корневой системы, жизненную форму зверобоя продырявленного в условиях остепненных лугов можно трактовать как полукустарничковую стержневую факультативно корнеотпрысковую (Дубровная, Мавлюдова, 2012). При выделении полукустарничковой жизненной формы учитывали длительность жизни и анатомическую структуру надземных осевых побегов. Анатомическая структура надземных побегов H. perforatum у особей разных онтогенетических состояний была изучена (Мазей, 2010). Однако автор не указывал, какая часть побега была использована для анализа анатомической структуры. В ходе работы нами было проанализировано более 100 побегов у 37 растений средневозрастного онтогенетического состояния и сходного габитуса. Срезы проводили на плагиотропных участках анизотропных побегов и в основании ортотропных побегов возобновления (рис. 1). 58 Рисунок 1 – Строение особей H. perforatum полукустарничковой жизненной формы: А – главный корень; Б – плагиотропные участки анизотропных побегов; В – побеги возобновления Проведенные исследования показали, что из всех побегов возобновления ортотропного положения около 90 % были однолетними. У них насчитывался один годичных прирост, величина которого могла существенно варьировать. Двулетние ортотропные побеги встречались крайне редко (рис. 2; табл.). Показатели календарного возраста различных типов надземных побегов Тип побега Календарный возраст побега, год 1 2 3 всего побегов Плагиотропные 9 (18,7 %) 34 (70,9 %) 5 (10,4 %) 48 Ортотропные 52 (91,2 %) 5(8,8 %) 0 57 Проведенные исследования показали, что побеги возобновления, как привило, характеризуются целостной сердцевиной, из всех изученных побегов только 14,3 % характеризовались нарушенной сердцевиной в основании побега. 59 Рисунок 2 – Анатомическое строение двулетнего побега. Стрелкой указаны весенние сосуды Таким образом, можно отметить, что в развитии надземных побегов зверобоя отмечается определенная тенденция, а именно, плагиотропные участки анизотропных побегов были многолетними, на которых развивались однолетние монокарпические побеги с годичным циклом развития. Можно отметить, что для зверобоя продырявленного в условиях суходольных лугов формируется типичная полукустарничковая жизненная форма на основе симподиально-базитонической длиннопобеговой и ортотропной симподиальной длиннопобеговой модели побегообразования. Л и т е р а т ур а Гонтарь Э. М. Онтогенез зверобоя продырявленного / Э. М. Гонтарь, В. Н. Годин // Онтогенетический атлас лекарственных растений / Мар. гос. ун-т. – Йошкар-Ола, 2002. – Т. 3. – С. 206–213. Дубровная С. А. Разнообразие жизненных форм Hypericum perforatum L. на территории Республики Татарстан / С. А. Дубровная, Л. У. Мавлюдова // Ученые записки Казанского университета. Сер., Естественные науки. – 2012. – Т. 153, кн. 2. – С. 170–182. Мазей Н. Г. Анатомические особенности вегетативных органов зверобоя продырявленного в ходе онтогенеза / Н. Г. Мазей // Известия ПГПУ им. В. Г. Белинского. – 2010. – № 21. – С. 9–19. Пархоменко М. В. Биологические особенности и структура ценопопуляций зверобоя продырявленного (Hypericum perforatum L.) в условиях Саратовской области: автореф. дис. … канд. биол. наук / М. В. Пархоменко. – Саратов: СГУ, 2012. – 20 с. Шишкин Б. К. Род Зверобой / Б. К. Шишкин, Е. Г. Бобров // Флора СССР. – Л.; М., 1949. – Т. 15. – С. 201–253. 60 ВЛИЯНИЕ ОТМЕНЫ ВЫПАСА НА БИОРАЗНООБРАЗИЕ РАСТИТЕЛЬНОСТИ МНОГОЛЕТНИХ ПАСТБИЩ В ПОЙМЕ РЕКИ УГРЫ Ермакова И. М., Сугоркина Н. С. Московский государственный педагогический университет, г. Москва, geranium.08@mail.ru Многолетний мониторинг проводится с 1965 г. в Калужской области на Залидовских лугах поймы реки Угры в национальном парке «Угра». Для оценки влияния резкой смены хозяйственного использования на динамику биоразнообразия многолетних пастбищ проанализированы данные наблюдений 3-х пастбищных участков с начала их мониторинга до 2001 г. (был выпас) и с 2002 по 2010–2011 гг., когда использование участка 9 стало эпизодически сенокосным, а участки 10 и 15 были выведены из использования (табл. 1). Предыстория использования участков разная: участок 10 до 1980 г. располагался у выхода из загонов скота, после перевода стада в другой загон оказался на проходе стада к воде и был очень сильно сбит; участок 9 находился на проходе стада, но сбивался меньше; участок 15.1 – нижней части берегового вала, где стадо бывало не ежедневно, повреждался при пробеге скота сильно. Пастьба на всех участках до 2001 г. включительно была испокон веку. Наши наблюдения наиболее длительны на участке 9 (Ермакова, Сугоркина, 2012). Жизненные формы (ЖФ) ценопопуляций встречающихся видов (далее именуемых видами) и подходы к построению спектров ЖФ описаны ранее (Ермакова, Сугоркина, Петросян, 2005; Ермакова, Сугоркина, 2010, 2011). На участке 10 при прекращении выпаса увеличилось биоразнообразие: выросли среднегодовое число видов с 19,0 до 22,9 и число ЖФ с 6,3 до 8,4. Роль растений разных ЖФ изменилась. До 2002 г. больше всего было малолетников – 23,3 %, стержнекорневых – 21,1 %, на 3-м месте оказались – рыхлокустовые и длиннокорневищные – 15,8 %. После 2002 г. стержнекорневых стало 29,4 %, длиннокорневищных – 21,8 %, на 3-м месте рыхлокустовые – 14,7 % (табл. 1). По годам стержнекорневых было почти всегда много, после 2002 г. их всегда больше других ЖФ, только в 1988, 1992, 1997, 2001 гг. они оказались на 2-м месте. Однолетников было много до прекращения выпаса, они занимали 1–2-е места. После прекращения выпаса их стало значительно меньше (в среднем 6,0 % против 23,3 % при выпасе), рыхлокустовые на 3-м месте до и после 2002 г. Длиннокорневищные появились в значительном количестве с 1997 г. (рис., табл. 1). 61 По проективному покрытию доминантов разных ЖФ в пастбищный период наибольшим было покрытие малолетников (29,4 %), после прекращения выпаса – длиннокорневищных (32,3 %), в общем за весь период наблюдений длиннокорневищные доминанты покрывали 21,3 % площади, малолетники – 11,5 % (табл. 2). Таблица 1 – Изменение среднегодового числа видов и участия видов по ЖФ (%) при выпасе и без выпаса Участки Наличие выпаса Годы 10 есть 15 нет есть 9 нет есть нет 1987–2001 2004–2011 2000–2001 2002–2011 1980–2001 2002–2010 Число видов 19,0 ± 1,2 22,9 ± 0,4 39,5 ± 1,5 36,2 ± 1,5 35,2 ± 1,2 34,6 ± 0,8 ЖФ* / Число ЖФ 6,3 ± 0,2 8,4 ± 0,1 9,0 ± 0 8,6 ± 0,2 8,9 ± 0,2 9,9 ± 0,2 1 21,1 29,4 31,6 33,1 27,8 22,6 2 5,3 3,6 6,3 1,7 8,4 5,5 3 3,0 9,1 5,1 8,3 9,8 14,3 4 0 2,0 2,5 2,8 0 2,6 5 15,8 21,8 15,2 28,6 18,8 23,7 6 9,0 2,0 11,4 5,9 8,7 4,0 7 0 3,6 0 0,3 1,4 2,2 8 0 0 0 0 0,3 0 9 15,8 14,7 8,9 6,9 8,1 10,6 10 6,8 6,3 6,3 5,5 9,6 8,1 11 23,3 6,0 12,7 4,5 5,9 4,3 12 0 1,6 0 0,3 1,1 1,9 13 0 0 0 2,1 0 0 * ЖФ (1–13) см. под рисунком. На участке 15 при отмене выпаса число видов сначала увеличилось, затем снизилось, в среднем за год уменьшилось с 39,5 до 36,2. Наибольшее число видов наблюдалось в сухом 2002 г. (55) и в 2004 г. (46). Число ЖФ было наибольшим в годы без выпаса – 2004 и 2007 (11 и 10), среднегодовое число в годы с выпасом – 9, без выпаса – 8,6. 62 Изменение спектров видов по ЖФ на участках мониторинга 9, 10, 15. ЖФ: 1 – стержнекорневые; 2 – кистекорневые; 3 – короткокорневищные; 4 – корнеотпрысковые; 5 – длиннокорневищные; 6 – наземно-ползучие; 7 – луковичные; 8 – плотнодерновинные; 9 – рыхлокустовые; 10 – корневищно-кустовые; 11 – малолетники; 12 – полупаразиты; 13 – кустарники (Ермакова, Сугоркина, 2011). М % – среднегодовое относительное число видов по ЖФ. Спектры ЖФ участка 9 с 1980 по 2002 гг. (см. Ермакова, Сугоркина, Петросян, 2005) Роли ЖФ почти не менялись – на 1-м месте по числу видов все годы были стержнекорневые, на 2-м – длиннокорневищные. 3-е место в разные годы занимали разные ЖФ: короткокорневищные в 2007, 2008, 2010 гг., малолетники – в 2001, 2002 гг. (вместе с рыхлокустовыми и корневищно-кустовыми), наземноползучие в 2000 г. В среднем по годам с выпасом и без на 1-м месте были стержнекорневые, на 2-м – длиннокорневищные, на 3-м в годы с выпасом – малолетники, без выпаса – короткокорневищные (табл. 1). 63 Таблица 2 – Среднегодовое проективное покрытие (П) доминирующих видов по ЖФ (%) при выпасе (ПВ), без выпаса (ПБ) и общее (ПО) Участки 10 (1987–2011 гг.) 15 (2000–2011 гг.) 9 (1980–2011 гг.) ЖФ / П ПВ ПБ ПО ПВ ПБ ПО ПВ ПБ ПО 1 9,7 2,3 5,1 20,0 4,8 7,8 21,7 5,5 13,8 1,0 1,0 1,0 4,5 2,2 26,0 17,8 2 3 4 5 6,0 6 3,5 2,5 1,6 32,3 21,3 8,0 1,3 8,0 1,2 1,0 47,9 39,9 10,0 1,2 1,0 37,0 18,9 9 2,7 5,1 4,2 3,2 4,2 16,3 41,1 28,4 10 4,0 3,9 3,9 1,0 0,8 14,1 21,8 17,8 11 29,4 0,8 11,5 1,0 0,8 Однако проективное покрытие доминантов выявило, при выпасе преобладали стержнекорневые (до 20 % площади), после отмены выпаса – длиннокорневищные – почти 48 % в основном за счет костра безостого (Bromopsis inermis (Leys.) Holub), вид с 2004 г. постепенно наращивал покрытие от 8 до 70 % в 2008 г., затем снизил покрытие до 40–50 %. Разнообразие ЖФ доминирующих видов увеличилось в первые годы без выпаса с 3 до 5, затем снизилось до 1–2 (табл. 2). Особенно много доминантов было в 2004 г. (5), затем число их снизилось, в 2008 и 2010 гг. был 1 доминант Bromopsis inermis. Содоминантами при выпасе были овсяница луговая (Festuca pratensis Huds.) и одуванчик лекарственный (Taraxacum officinale Wigg.) без выпаса – стержнекорневые Taraxacum officinale (2002), лопух паутинистый Arctium tomenotsum (2004, 2005), длиннокорневищные – сныть обыкновенная (Aegopodium podagraria L.) (2007, 2009, 2011), рыхлокустовые – Festuca pratensis (2000–2004), ежа сборная (Dactylis glomerata L.) в 2004 г. и др. Сравнение с участком 10 показало, что роль длиннокорневищных доминантов при отсутствии выпаса становится на участках главной, но на участке 10 до выпаса преобладали малолетники, а на участке 15 стержнекорневые. На участке 9 при прекращении выпаса среднегодовое число видов уменьшились с 35,2 до 34,6, число ЖФ увеличилось с 8,9 до 9,9. Изменилась роль ЖФ: больше всего стало длиннокорневищных – 23,7 %, на 2-м месте – стержнекорневые – 22,6 %; при выпасе было наоборот, преобладали стержнекорневые – 27,8 %, на 2-м месте – длиннокорневищ64 ные – 18,8 %, на 3-м – короткокорневищные (14,3 %), вместо корневищно-кустовых (9,6 %), 4-е место – рыхлокустовые (10,6 %) вместо кистекорневых (8,4 %) и рыхлокустовых (8,1 %). По годам наблюдений происходили флюктуационные изменения числа видов по ЖФ, но при выпасе преобладали стержнекорневые (в 2001 г. с длиннокорневищными), после отмены выпаса почти всегда – длиннокорневищные; на 2-м месте при выпасе чаще были длиннокорневищные (в 1988 г. корневищно-кустовые, в 1992 г. еще и короткокорневищные); после отмены выпаса 1-е место все годы занимали длиннокорневищные; в 2007 г. роли этих ЖФ одинаковые, в 2008 г. ЖФ поменялись местами. В 2004, 2005 и 2010 гг. в части описаний 2-е место было у короткокорневищных, иногда со стержнекорневыми (рис., табл. 1). По проективному покрытию в пастбищный период главную роль среди доминантов играли наземноползучие – 37 % (клевер луговой – Amora repens (L.) C. Presl), на 2-м месте были стержнекорневые (21,7 %), на 3-м – рыхлокустовые (16,3 %). С отменой выпаса наземноползучие исчезли из состава доминантов, на 1-е место вышли рыхлокустовые (41,1 %), на 2-е – длиннокорневищные (26 %), на 3-е – корневищно-кустовые (21,8 %) (табл. 2). Перераспределение ролей растений разных ЖФ происходило постепенно. Усиление позиции рыхлокустовых произошло на 3-й год после смены использования – доминантами стали Dactylis glomerata, полевица бледная (Agrostis diluta Kurcz.). У корневищно-кустовых перестала доминировать овсяница красная (Festuca rubra L.), но стал доминантом мятлик узколистный (Poa angustifolia L.). Через 2 года после отмены выпаса перестал доминировать стержнекорневой Taraxacum officinale, через 2 года почти постоянным доминантом стал длиннокорневищный Bromopsis inermis. При уменьшении влияния выпаса (переводе стада в другой загон в 1990-м году), снизилось очень высокое ранее покрытие (20–60 %) наземноползучего Amora repens, вид не доминировал с 1992 г. Только Festuca pratensis доминировал практически весь период мониторинга с одинаковым покрытием. После отмены выпаса на участке 10 среднегодовая видовая насыщенность увеличилась почти на 4 вида, а на менее сбитых участках 9 и 15 снизилась на 0,6–3 вида соответственно. Число ЖФ возросло на участках 10 и 9 и убавилось на 0,4 на участке 15. С отменой выпаса произошло перераспределение ролей представителей разных ЖФ. На 1-е место по числу видов на участке 10 вышли 65 стержнекорневые вместо малолетников, на участке 9 длиннокорневищные заменили стержнекорневые, на участке 15 смены стержнекорневых не произошло, они остались и после прекращения выпаса. На 2-е место на участке 10 вышли длиннокорневищные вместо стержнекорневых, на участке 9 длиннокорневищные, занявшие 1-е место, сменились стержнекорневыми; на участке 15 смены не произошло – оставались длиннокорневищные. На 3-м месте на участке 10 оставались рыхлокустовые, на участке 9 рыхлокустовые сменились короткокорневищными, на участке 15 малолетники сменились короткокорневищными (табл. 1). Анализ показал, что при самом большом повреждении участка, на нем по числу видов преобладают малолетники, при меньшем – стержнекорневые. При отмене выпаса стержнекорневые заменяются длиннокорневищными, малолетники – стержнекорневыми, а на 2-е место выходят длиннокорневищные. Изменяется суммарное проективное покрытие доминантов: на участке 10 малолетники уступили 1-е место длиннокорневищным, на участке 15 – стержнекорневые – длиннокорневищным, на участке 9 наземноползучие рыхлокустовым (табл. 2). На участке с очень сильным выпасом, на стадии сбоя, выживают лучше всего малолетники, которые могут возобновляться на всегда существующих пустых местах. При уменьшении нагрузки преимущества получают стержнекорневые. При прекращении выпаса быстро разрастаются длиннокорневищные, медленнее короткокорневищные и рыхлокустовые растения. Таким образом, при резкой смене пастбищного использования на эпизодически сенокосное или отсутствие использования, произошло изменение биоразнообразия, числа видов и спектра и числа ЖФ, ЖФ доминантных видов, их суммарного проективного покрытия. Направления изменений на 3-х участках обычно не совпадали. Л и т е р а т ур а Ермакова И. М. Динамика спектров жизненных форм растений пойменных лугов / И. М. Ермакова, Н. С. Сугоркина // Биоморфологические чтения к 150-летию со дня рождения К. Раункиера: материалы конф. – Киров, 2010. – C. 218–223. Ермакова И. М. Жизненные формы растений Залидовских лугов Калужской области / И. М. Ермакова, Н. С. Сугоркина // Ботан. журн. – 2011. – Т. 96, № 3. – С. 316–341. Ермакова И. М. Влияние смены хозяйственного использования на динамику растительности пойменных лугов / И. М. Ермакова, Н. С. Сугоркина // Биоразнообразие: проблемы изучения и сохранения: материалы Междунар. научн. конф. – Тверь, 2012. – С. 107–110. Ермакова И. М. Динамика спектра жизненных форм луговых растений пойменного ландшафта в национальном парке «Угра» / И. М. Ермакова, Н. С. Сугоркина, В. Г. Петросян // Экологические проблемы сохранения исторического и культурного наследия. – М., 2005. – С. 405–425. 66 СРАВНИТЕЛЬНОЕ ИЗУЧЕНИЕ ВИДОВ И СОРТОВ ЛАПЧАТОК Ефремова Л. П., Бирюкова О. А., Денисова Т. А. Поволжский государственный технологический университет, г. Йошкар-Ола, eflukm@rambler.ru Лапчатка (Potenttila L.) – один из крупнейших по числу видов род растений из семейства Розовые (Rosaceae). Род представлен чаще всего многолетними, реже однолетними и двулетними травянистыми растениями и полукустарниками. Среди них много декоративных растений, которые можно использовать в различных видах цветочного оформления. Цветки растений этого рода обладают разнообразной окраской, чаще всего белой, также встречаются красные, желтые, оранжевые, розовые. Среди растений рода лапчатка встречаются виды, цветущие ранней весной и в середине лета. Цель исследования: сравнительная оценка 6 видов и сортов лапчатки, и выявление наиболее перспективных представителей для цветочного оформления средней полосы России. Объектами исследований были лапчатка белая (Potentilla alba L.); лапчатка непальская (Potentilla nepalensis Hook); лапчатка темнокроваво-красная (Potentilla atrosanguinea Lodd. ex D. Don.); лапчатка темно-кроваво-красная 'Мастер Флорис' (Potentilla atrosanguinea 'Master Floris' hort.); лапчатка темно-кроваво-красная 'Мсье Ройлэнд' (Potentilla atrosanguinea 'Mons. Rouilland' hort.); лапчатка гибридная 'Уайт Бьюти' (Potentilla x hybridа 'White Beauty' hort.) из коллекции Ботанического сада ПГТУ. В июне 2011 г. проводили черенкование вышеуказанных видов и сортов лапчаток, укоренение черенков производили в смеси торфа и песка в соотношении 1 : 1 в холодном парнике. В сентябре укоренившиеся растения пересаживали на гряды открытого грунта. В ходе исследования вели фенологические наблюдения в течение двух вегетационных сезонов, а также изучали декоративные и хозяйственно-биологические признаки молодых растений, выращенных из черенков и маточников. При наблюдении за сезонным развитием исследуемых растений отмечали следующие фенофазы: развертывание листьев, появление бутонов, начало цветения, конец цветения, завязывание плодов, созревание семян. К декоративным признакам относили: высоту растений без цветоноса, высоту цветоноса, диаметр цветков, окраску цветков, продолжительность цветения, количество цветоносов. К хозяйственно-биологическим признакам относили: процент укоренившихся черенков, процент прижившихся после пересадки молодых растений, устойчивость к болезням и вредителям, степень разрастания, зимостойкость. Обработка данных по результатам исследования прово67 дилась в программе Microsoft Excel. Различия считались статически достоверными при р ≤ 0,05. На основе совокупности декоративных и хозяйственно-биологических признаков была проведена оценка перспективности видов и сортов лапчаток для использования в цветочном оформлении средней полосы России. При проведении оценки перспективности учитывалась успешность прохождения всех фенофаз, декоративные признаки: окраска, продолжительность цветения, сохранение декоративности после цветения; хозяйственно-биологические признаки: процент укоренившихся черенков и приживаемость молодых растения в открытом грунте, зимостойкость, устойчивость к болезням и вредителям. Процент укореняемости черенков варьировал от 50 до 95. Наибольшие показатели были у сортов лапчатки темно-кроваво-красной (92– 95 %), лапчатки белой (85 %), более низкие – у лапчатки непальской (60 %) и лапчатки гибридной (50 %). Различались исследованные культуры и по показателям приживаемости после высадки в открытый грунт: лапчатка темно-кроваво-красная (92–100 %), лапчатка гибридная (89 %), лапчатка белая (76 %) и лапчатка непальская (57 %). Одним из ведущих методов изучения интродуцентов является сравнительное исследование ритмов сезонного развития. Это позволяет не только установить сроки прохождения различных фаз развития, но и оценить устойчивость к неблагоприятным факторам окружающей среды, продуктивность, декоративность наблюдаемых растений. Фенологические наблюдения позволили установить, что в 2011 г. начало вегетации всех культур приходилось на первую декаду мая. В связи с тем, что растения относятся к разным видам, даты наступления фенофаз существенно отличались. Раньше всех фаза бутонизации начиналась у лапчатки белой (1 декада мая). В третью декаду мая бутонизацию наблюдали у лапчатки гибридной; в третью декаду июня – у лапчаток темно-кроваво-красной и непальской, в первую декаду июля – у лапчатки темно-кроваво-красной. Раньше всех начинала цвести лапчатка белая – во вторую декаду мая, в третью декаду мая зацветала лапчатка гибридная, в первую декаду июля – лапчатки темно-кроваво-красная и непальская, во вторую декаду июля начинали цвести сорта лапчатки темно-кроваво-красной. Завязывание плодов и образование семян происходило у всех изученных культур. Однако, при проверке семян на всхожесть они не прорастали. В 2012 г. лапчатка темно-кроваво-красная и ее сорта зацветали на 2 недели раньше, в то время как у остальных лапчаток разница в датах начала цветения составляла 2–3 дня. Одним из значимых показателей для оценки перспективности декоративных культур является продолжительность цветения. Быстрее всех культур отцветала лапчатка гибридная 'Уайт Бьюти', продолжитель68 ность ее цветения составляла 13 и 10 суток соответственно в 2011 и 2012 годах. Остальные лапчатки цвели на протяжении 3–4 недель. Наибольшая высота молодых растений без цветоноса была у сорта лапчатки темно-кроваво-красной 'Мастер Флорис' (21,26 ± 1,21 см), наименьшая у лапчатки белой (10,67 ± 0,36 см). Максимальное количество цветков на цветоносе наблюдалось у лапчатки белой (7,38 ± 1,61 шт.), минимальное у сорта лапчатки темно-кроваво-красной 'Мсье Ройлэнд' (3,89 ± 0,41 шт.). Диаметр цветков молодых растений колебался в пределах от 1,32 ± 0,04 см (лапчатка белая) до 2,48 ± 0,06 см (лапчатка темно-кроваво-красная). Максимальное количество цветоносов насчитывалось у лапчатки белой (2,53 ± 0,38 шт.). Высота цветоноса варьировала от 8,53 ± 0,47 см у лапчатки белой до 34,51 ± 0,76 см у лапчатки темно-кроваво-красной 'Мсье Ройлэнд'. Растения, развившиеся из черенков, за период исследования не достигали высоты маточников (р ≤ 0,05), исключение составляла лапчатка белая. Количество образовавшихся цветков на цветоносе у маточников большинства видов и сортов было больше, чем у молодых растений, у лапчатки белой эти показатели не отличались. Достоверных различий между молодыми и маточными растениями по диаметру цветков не было. Число цветоносов у молодых растений лапчатки темно-кровавокрасной и ее сортов, лапчатки гибридной 'Уайт Бьюти' было меньше, чем у маточников (р ≤ 0,05). Лапчатка белая по этому показателю не отличалась от маточных растений. В ходе наблюдений за маточными растениями изучаемых видов и сортов была отмечена гибель маточников лапчатки непальской после зимы 2011 г., остальные исследуемые культуры не пострадали. К хозяйственно-биологическим признакам относили поражаемость болезнями и вредителями. У маточных растений лапчатки белой были отмечены признаки поражения патогенными грибами. Проведенные исследования показали, что все изученные виды и сорта лапчатки успешно развиваются в климатических условиях Республики Марий Эл. Исследованные культуры проходили все фазы сезонного развития. На основе изучения сезонного развития, декоративных и хозяйственно-биологических признаков были выявлены наиболее перспективные виды и сорта для выращивания в условиях климата средней полосы России: лапчатка белая, лапчатка темно-кроваво-красная и ее сорта 'Мастер Флорис' и 'Мсье Ройлэнд', лапчатка гибридная 'Уайт Бьюти'. Они могут быть использованы в групповых посадках, миксбордерах, на каменистых горках. 69 ОНТОГЕНЕТИЧЕСКИЙ ГЕРБАРИЙ КАК ОСНОВА ДЛЯ ИЗУЧЕНИЯ ВНУТРИПОПУЛЯЦИОННОГО БИОРАЗНООБРАЗИЯ Козырева С. В., Ведерникова О. П., Жукова Л. А. Марийский государственный университет, г. Йошкар-Ола, ecology@marsu.ru В настоящее время огромное внимание уделяется изучению и сохранению биоразнообразия, для этого существует множество организаций, одной из которых являются гербарии. Гербарий – может быть крупным, самостоятельным, научным учреждением, работающим с гербарными коллекциями или так называют коллекции, высушенных растений, обработанных определенным способом и являющихся основой для изучения внутрипопуляционного биоразнообразия. Такие коллекции могут быть частью фондов музея. В этом качестве и может выступать Онтогенетический гербарий, являясь частью Популяционно-онтогенетического музея, который с 15 июля 2008 года был официально зарегистрирован при кафедре экологии Марийского государственного университета и входит в Евразийскую ассоциацию университетских музеев (Смуров и др., 2012). 16 июля 2013 года была получена информация об официальной регистрации гербария (под кодом MARI) в базе данных НьюЙоркского ботанического сада, где регистрируются гербарии со всего мира (сайт http://sweetgum.nybg.org/ih/herbarium.php?irn=176924). Во многих странах собраны коллекции систематического гербария, насчитывающие миллионы гербарных листов, которые служат основой для изучения флористического биоразнообразия. Крупнейшими и старейшими гербариями являются гербарии в Лондоне (Королевский ботанический сад Кью), в США (Национальный гербарий и гербарий в НьюЙоркском ботаническом саду), а в нашей стране – гербарии: Ботанического института (БИН) им. В. Л. Комарова (Санкт-Петербург), МГУ им. М. В. Ломоносова, Тверского и Томского государственных университетов и т. д. В то же время внутривидовое биоразнообразие изучено крайне слабо. На решение этой проблемы направлено создание Онтогенетического гербария в Марийском государственном университете. Огромная заслуга в этом принадлежит заслуженному деятелю науки Российской Федерации, доктору биологических наук, профессору Л. А. Жуковой. В основу гербария были положены обширные гербарные коллекции, собранные в экспедициях учениками А. А. Уранова, привезенные из различных районов России и ближнего зарубежья. Сейчас фонды Онтогенетического гербария преимущественно состоят из гербарных образцов растений флоры Республики Марий Эл. Онтогенетический гербарий используют как в научных, так и педагогических целях, поэтому он состоит из научной и учебной час70 тей. Научная часть насчитывает более 5300 смонтированных гербарных листов 607 видов растений из 369 родов и 86 семейств; учебная часть включает 286 видов из 198 родов и 60 семейств. Экспонаты в гербарии расположены по алфавиту латинских названий семейств и видов внутри этих семейств. Систематический каталог постоянно обновляется, в нем указаны онтогенетические состояния видов, их количество, места сборов и основные коллекторы. Онтогенетический гербарий представляет собой гербарий растений, находящихся на разных этапах индивидуального развития. Онтогенетические состояния выделяются с использованием концепции дискретного описания онтогенеза (Работнов, 1950; Уранов, 1975) с учетом морфологических признаков-маркеров. Каждый вид в Онтогенетическом гербарии представлен экземплярами в различных онтогенетических состояниях от проростков до сенильных растений (полный онтогенез), а в ряде случаев – неполный, т. к. в природе сразу собрать полный онтогенез достаточно сложно и поэтому такой гербарий постоянно дополняется. Наш Онтогенетический гербарий имеет дубликаты некоторых видов растений, которыми мы готовы обмениваться, а также можем принять гербарии онтогенезов любых видов растений на хранение. Всю информацию можно получить по e-mail: ecology@marsu.ru и в опубликованных работах (Жукова и др., 2002; Zhukova et al., 2003; Онтогенетический гербарий…, 2003; Жукова и др., 2004; Жукова и др., 2007; Структурное разнообразие…, 2009; Жукова и др., 2009; Козырева и др., 2010; Жукова и др., 2010; Популяционно-онтогенетический музей…, 2010; Козырева и др., 2011; Развитие идей…, 2011; Создание и развитие…, 2013; Ведерникова и др., 2013). Работа с образцами Онтогенетического гербария позволяет получить навыки определения онтогенетических состояний растений разных биоморф, может быть использован экологами, ботаниками, ресурсоведами, сотрудниками ботанических садов, национальных парков, заповедников, преподавателями, аспирантами, докторантами, студентами биологических специальностей, школьниками и учителями школ с углубленным изучением биологии и является научной базой для изучения, демонстрации и сохранения внутрипопуляционного биоразнообразия. Результаты работы обобщены в четырех томах периодического издания «Онтогенетический атлас лекарственных растений» и в двух томах «Онтогенетический атлас растений», где описаны онтогенезы 237 видов растений разных жизненных форм из 62 семейств, авторами которых были 195 исследователей, работающих в популяционноонтогенетическом направлении. Пятый том Онтогенетического атласа растений на Всероссийском конкурсе РУДН (г. Москва) получил 71 Диплом III степени – за лучшую книгу 2010 года в области экологии. В ближайшее время выйдет VII том. В настоящее время гербарии востребованы для образования, поскольку хранимые и представляемые ими образцы – это первичные источники информации о растениях. Поскольку наш Онтогенетический гербарий не имеет мировых аналогов, то его образовательное и прикладное значение велико, и способствует интенсивному развитию популяционно-онтогенетического направления и популяционной экологии в целом. Л и т е р а т ур а Ведерникова О. П. Популяционно-онтогенетический музей в Марийском государственном университете / О. П. Ведерникова, С. В. Козырева, Л. А. Жукова // Труды XIII съезда Русского ботанического общества «Современная ботаника в России» и конференции «Научные основы охраны и рационального использования растительного покрова Волжского бассейна» (г. Тольятти, 16–21 сентября 2013 г.). – Тольятти: Кассандра, 2013. – Т. III. – С. 258–260. Жукова Л. А. Онтогенетический гербарий / Л. А. Жукова, О. П. Ведерникова, Г. О. Османова // Фундаментальные и прикладные проблемы популяционной биологии: сб. тезисов докладов VI Всероссийского популяционного семинара (2–6 декабря 2002 г.). – Нижний Тагил, 2002. – С. 62–64. Жукова Л. А. Популяционно-онтогенетический музей как центр изучения и сохранения биоразнообразия / Л. А. Жукова, Г. О. Османова, О. П. Ведерникова // Экология города Йошкар-Олы. – Йошкар-Ола: Типография Правительства Марий Эл, 2004. – С. 174–182. Жукова Л. А. Онтогенетический гербарий / Л. А. Жукова, О. П. Ведерникова, Г. О. Османова // Экология города ЙошкарОлы: научное издание. – Йошкар-Ола, 2007. – С. 234–238. Жукова Л. А. Популяционноонтогенетический музей как центр изучения биоразнообразия / Л. А. Жукова, О. П. Ведерникова, С. В. Козырева // Актуальные проблемы экологии, биологии и химии: материалы конференции по итогам научно-исследовательской работы за 2008 г. – Йошкар-Ола, 2009. – С. 39–41. Жукова Л. А. Онтогенетический гербарий в Марийском государственном университете / Л. А. Жукова, О. П. Ведерникова, С. В. Козырева // Изучение растительных ресурсов Волжско-Камского края: сб. науч. тр. / гл. ред. В. Г. Папченков, сост., отв. за выпуск А. В. Дмитриев. – Чебоксары, 2010. – Вып. 1. – С. 33–44. Козырева С. В. Популяционно-онтогенетический музей и Онтогенетический гербарий Марийского государственного университета как центр изучения и сохранения биоразнообразия / С. В. Козырева, О. П. Ведерникова, Л. А. Жукова // Принципы и способы сохранения биоразнообразия: сб. материалов IV Всерос. науч. конф. с международным участием / Мар. гос. ун-т. – Йошкар-Ола, 2010. – С. 185–197. Козырева С. В. Популяционноонтогенетический музей Марийского государственного университета / С. В. Козырева, О. П. Ведерникова, Л. А. Жукова // Онтогенетический атлас растений / Мар. гос. ун-т. – Йошкар-Ола, 2011. – Т. VI. – С. 270–276. Онтогенетический атлас лекарственных растений / Мар. гос. ун-т; отв. ред. Л. А. Жукова. – Йошкар-Ола, 1997. – Т. 1. – 240 с.; 2000. – Т. 2. – 268 с.; 2002. – Т. 3. – 280 с.; 2004. – Т. 4. – 240 с.; 2007. – Т. 5. – 372 с.; Т. 6. – 336 с. Онтогенетический гербарий / Л. А. Жукова, О. П. Ведерникова, Г. О. Османова, С. В. Козырева // Седьмые Вавиловские чтения. Глобализация и проблемы национальной безопасности России в XXI веке: сб. материалов / под общ. ред. В. П. Шалаева. – ЙошкарОла: МарГТУ, 2003. – Ч. 2. – С. 154–156. Популяционно-онтогенетический музей – основа изучения и сохранения биоразнообразия / Л. А. Жукова, С. В. Козырева, Г. О. Османова, О. П. Ведерникова // Проблемы изучения и сохранения растительного мира Евразии: материалы Всерос. науч. конф. с международным участием, посвященной памяти выдающегося ученого Л. В. Бардунова (Иркутск, 15–19 сентября 2010 г.). – Иркутск: Изд-во Ин-та 72 географии им. В. Б. Сочавы СО РАН, 2010. – С. 582–584. Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах / Т. А. Работнов // Тр. БИН АН СССР. Сер. 3, Геоботаника. – М.: АН СССР, 1950. – Вып. 6. – С. 7–204. Развитие идей А. А. Уранова в работе Популяционно-онтогенетического музея Марийского государственного университета / Л. А. Жукова, Г. О. Османова, О. П. Ведерникова, С. В. Козырева // Современные проблемы популяционной экологии, геоботаники, систематики и флористики: материалы международной научной конференции, посвященной 110-летию А. А. Уранова (Кострома, 31 октября – 3 ноября 2011 г.). – Кострома: КГУ им. Н. А. Некрасова, 2011. – С. 99–103. Смуров А. В. Музеи университетов Евразийской ассоциации: Аннотированный справочник / А. В. Смуров, В. В. Снаткин, С. Ю. Ливанцова; под ред. В. А. Садовничего, В. Н. Семина. – 3-е изд. – М., 2012. – 433 с. Создание и развитие Популяционно-онтогенетического музея и его роль в подготовке профессиональных кадров / С. В. Козырева, Г. О. Османова, О. П. Ведерникова, Л. А. Жукова // Музеи евразийских университетов в поддержании и развитии общего образовательного пространства: материалы Междунар. науч.-метод. конф. (Томск, 26–29 сентября 2012 г.) / под ред. Э. И. Черняка. – Томск: Изд-во Томского ун-та, 2013. – С. 46–53. Структурное разнообразие растений и методы его изучения для вузов и школ / Л. А. Жукова, О. П. Ведерникова, Г. О. Османова, С. В. Козырева // Труды VIII Международной конференции по морфологии растений, посвященной памяти И. Г. и Т. И. Серебряковых. – М.: МПГУ, 2009. – С. 186–188. Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов / А. А. Уранов // Науч. докл. высш. школы. Биол. наука. – 1975. – № 2. – С. 7–33. Zhukova L. A. Ontogenetic herbarium as a method of study of intrapopulation biodiversity / L. A. Zhukova, O. P. Vedernikova, G. O. Osmanova // Modern problems of biondication and biomonitoring. Proc. XI Intern. Symp. on biondicators. – Syktyvkar, 2003. – P. 463–472. РАЗВИТИЕ КУСТАРНИЧКА PENTAPHYLLOIDES FRUTICOSA (L.) O. SCHWARZ В ГОРНОМ АЛТАЕ Комаревцева Е. К. Центральный сибирский ботанический сад СО РАН, г. Новосибирск, elizavetakomarevceva@yandex.ru Pentaphylloides fruticosa (L.) O. Schwarz – летнезеленый геоксильный кустарник из семейства Rosaceae высотой до 150 см с дизъюнктивным ареалом. Основная (азиатская) часть ареала находится в Северной, Центральной и Средней Азии. Фрагменты ареала отмечаются в Северной Америке, Европе, на Урале и Кавказе (Коропачинский, Встовская, 2002). Почти везде вид тяготеет к горным местообитаниям, произрастая от предгорий до альпийского пояса включительно (Горчаковский, 1960; Куминова, 1960). Основная жизненная форма – геоксильный кустарник, в разных условиях формирующий моноцентрический, неявно- и явнополицентрический типы биоморф (Комаревцева, 2010). Исследования проводились в Горном Алтае в августе 1999 г. на Семинском перевале (1700 м н. у. м.), на альпийском деградированном лугу. Онтогенез изучен по методике Т. А. Работнова (1950), 73 А. А. Уранова (1975) и его учеников (Ценопопуляции растений, 1976). Морфогенез описан согласно подходам И. Г. Серебрякова (1962), М. Т. Мазуренко, А. П. Хохрякова (1977). Для особи определяли абсолютный возраст по числу годичных колец на спиле ксилоподия или главного корня растения. В данных условиях в ходе сложного онтогенеза Р. fruticosa развивается как гипоэпигеогенно-геоксильный кустарничек высотой до 35 см, образующий куртины из парциальных побегов и кустов (рис. 1). Рисунок 1 – Сложный онтогенез кустарничка P. fruticosa. Условные обозначения: ПП – первичный побег, ГК – главный корень, ПЗ – побег замещения, ПФ – побег формирования, КС-П – ксилоподий, – вегетативный побег, – генеративный побег, – отмерший побег, – гипогеогенный ксилоризом; p, j, im, v, g, ss – онтогенетические состояния При отмирании материнского куста куртина распадается на системы парциальных побегов и кустов (Комаревцева, 2013). Рассмотрено 122 системы парциальных кустов и побегов, включающих более 850 парциальных образований. Частный онтогенез парциальные образования начинается с формирования побега имматурного типа из верхушечной почки ксилоризома (рис. 2). Имматурные парциальные побеги (imвег) – побеги, развивающиеся из верхушечной почки гипогеогенного ксилоризома (I тип) высотой 2–14 см или образовавшиеся при полегании и последующем укоренении ПФ (II тип) высотой 10–26 см. Преобладают побеги imвег I типа, что свидетельствует о вегетативном разрастании в основном за счет образования гипогеогенных ксилоризом. Возраст побегов I типа 2–6 лет, а побегов II типа 3–11 лет. Группа виргинильных парциальных кустов (vвег) высотой 6–30 см развивается в процессе кущения парциальных побегов. Подземная часть представлена ксилоподием диаметром 0,2–1,3 см, покрытым придаточными корнями и соединенным ксилоризомом с другим парциальным образованием. По наличию или отсутствию отмерших ПФ выделены следующие подгруппы. Молодые виргинильные парциальные образова74 ния (v1вег) в возрасте 3–15 лет состоят только из живых 2–6 ПФ. Если куст не зацветает, то он переходит в подгруппу v2вег или v*вег образований. При зацветании с 8 лет парциальный куст переходит в группу g1вег кустов. Взрослые виргинильные парциальные образования (v2вег) состоят из 2–6 живых и 1–12 отмерших ПФ и находятся в возрасте 7–30 лет. С 10 лет возможен переход в g2вег кусты. Угнетенные виргинильные парциальные образования (v*вег) в возрасте 7–35 лет состоят из 1 живого ПФ и до 20 остатков отмерших ПФ. Они появляются при отмирании ПФ у imвег или у v1вегобразований. В группе виргинильных образований преобладают v2вег и v*вег кусты, находящиеся приблизительно в равном соотношении. Молодые виргинильные образования (v1вег) немногочисленны. Рисунок 2 – Частный онтогенез парциального образования. Условные обозначения: ПП – первичный побег, ГК – главный корень, ПЗ – побег замещения, ПФ – побег формирования, КС-П – ксилоподий, – вегетативный побег, – генеративный побег, – отмерший побег, – гипогеогенный ксилоризом; p, j, im, v, g, ss – онтогенетические состояния 75 Генеративные парциальные кусты высотой 7–35 см состоят из 2–8 ПФ, из которых 1–6 цветущие. Диаметр ксилоподия 0,4–1,8 см. Возможны перерывы в цветении. В ксилоподии отмечаются признаки разрушения, но годичные кольца просматриваются. В группе выделены следующие подгруппы. Генеративные парциальные кусты, состоящие только из живых ПФ (g1вег), развиваются их v1вег кустов, быстро зацветающих в благоприятных условиях. Находятся в возрасте 8–23 лет. Генеративные кусты, имеющие в составе отмершие побеги (g2вег), развиваются из v2вег и g1вег кустов. В кусте кроме живых есть 1–11 отмерших ПФ. Возраст кустов от 10 до 43 лет. Угнетенные генеративные парциальные кусты (g*вег) образуются из v*вег кустов, имеют один цветущий ПФ и 1–8 остатки отмерших ПФ, находятся в возрасте 8–32 лет. Субсенильные парциальные образования (ssвег) состоят из 2–9 живых ПФ и 3–21 отмерших ПФ. По высоте и развитию ПФ не отличаются от виргинильных кустов, но имеют сильно разрушенный ксилоподий диаметром 0,7–1,5 см. Возраст таких образований колеблется от 24 до 40 и более лет. Сенильные парциальные образования не обнаружены. Таким образом, в альпийском поясе Р. fruticosa развивается как гипоэпигеогенно-геоксильный кустарничек. В ходе сложного онтогенеза в середине генеративного периода происходит вегетативное размножение с глубоким омоложением вегетативного потомства. В частном онтогенезе парциального образования наряду с нормальным ходом развития от парциального побега (imвег) до парциального генеративного куста (g2вег) около половины парциальных виргинильных кустов представлены угнетенными однопобеговыми растениями (v*вег), которые зацветая, образуют группу угнетенных генеративных образований (g*вег). Почти равное соотношение vвег и v*вег указывает на важную роль последней группы растений в удержании видом территории. Л и т е р а т ур а Горчаковский П. Л. О распространении и условиях произрастания дазифоры кустарниковой (Dasiphora fruticosa (L.) Rydb) в связи с реликтовой природой ее уральских местонахождений / П. Л. Горчаковский // Записки Свердл. отд. Всесоюзн. ботан. общ-ва. – Свердловск, 1960. – Вып. 1. – С. 3–22. Коропачинский И. Ю. Древесные растения Азиатской России / И. Ю. Коропачинский, Т. Н. Встовская. – Новосибирск: Изд-во СО РАН, филиал «Гео», 2002. – C. 341–342. Куминова А. В. Растительный покров Алтая / А. В. Куминова. – Новосибирск: Изд-во СО АН СССР, 1960. − 450 с. Уранов А. А. Возрастной спектр фитоценопопуляции как функция времени и энергетических волновых процессов / А. А. Уранов // Научн. докл. высш. шк. Биол. науки. – 1975. – № 2. – С. 7–34. Серебряков И. Г. Экологическая морфология растений / И. Г. Серебряков. – М.: Высш. шк., 1962. – 378 с. Ценопопуляции растений (основные понятия и структура) / отв. ред. А. А. Уранов, Т. И. Серебрякова. – М.: Наука, 1976. – 217 с. Мазуренко М. Т. Структура и морфогенез кустарников / М. Т. Мазуренко, А. П. Хохряков. – М.:Наука, 1977. – 160 с. Комаревцева Е. К. Поливариантность онтогенеза Pentaphylloides fruticosa (L.) O. Schwarz в Горном 76 Алтае / Е. К. Комаревцева // Принципы и способы сохранения биоразнообразия: материалы IV Всерос. конф. с международным участием (Йошкар-Ола, 22–26 сентября 2010 г.) / Мар. гос. ун-т. – Йошкар-Ола, 2010. – С. 202–203. Комаревцева Е. К. Онтогенез кустарничка Pentaphylloides fruticosa (L.) O. Schwarz в Горном Алтае / Е. К. Комаревцева // Modern Phytomorphology: 2nd International Scientific Conference on Plant Morphology (14– 16 May 2013, Lviv, Ukraine). – Lviv, 2013. – Vol. 4. – С. 118–122. ВИДОВОЙ СОСТАВ МАКРОЗООБЕНТОСА РЕК АБХАЗИИ Мингазова Д. Ю.1, Мингазова Н. М.1, Набеева Э. Г.1, Иванова В. М.1, Дбар Р. С.2 1 Казанский (Приволжский) федеральный университет, г. Казань, nmingas@mail.ru 2 Институт экологии Республики Абхазия, г. Сухум Сохранение уникального генофонда гидробионтов возможно только в естественных или близких к естественным условиям окружающей среды. Одним из наиболее показательных индикаторов изменений природных водных экосистем являются организмы зообентоса вследствие длительного формирования их сообществ. В настоящей работе изучение зообентоса проводилось на водных объектах Республики Абхазия (бывшей Грузинской Республики). Реки Республики Абхазии во многом сохраняют естественные условия, в силу малой антропогенной нарушенности или отсутствия воздействия, поэтому состояние их биотических сообществ во многом может рассматриваться как эталонное. Исключением являются устьевые участки рек в местах впадения у населенных пунктов и участки рек в местах добычи полезных ископаемых. Большинство водных объектов Абхазии является малоисследованными и неисследованными вследствие труднодоступности горных водных объектов, а также вследствие военных событий 1992–2008 гг. и длительного периода непризнания независимости республики. Таким образом, результаты экспедиций приобретают важное значение для выделения видового состава и состояния биотических сообществ. Экспедиции были организованы Казанским федеральным университетом (Лабораторией оптимизации водных экосистем) в сотрудничестве с Институтом экологии Республики Абхазии, Абхазским государственным университетом (кафедра экологии и зоологии) и Комитетом по экологии Республики Абхазии. В ходе исследований были изучены биотические сообщества 23 крупных и малых рек Республики Абхазии. В исследуемых реках Республики Абхазии было обнаружено 104 вида из числа организмов зообентоса, принадлежащих 4 типам, 7 классам, 18 отрядам, 56 семействам и 78 родам. 77 Виды зообентоса были исследованы на предмет встречаемости в реках Республики Абхазии (см. табл.). Наиболее часто встречались такие виды, как Heptagenia sulphurea (52,17 %), Gammarus crispus и Cheumatopsyche lepida (34,783 %). Встречаемость видов макрозообентоса в реках Абхазии (2011–2012 гг.) Crustacea Встречаемость, % Plecoptera Paramoera udehe 4,4 Leuctra sp. Assellus monticola 4,4 Leuctra fusca Gammarus crispus 34,8 Leuctra lippopus Gammarus duebeni Gammarus lacustris Oligochaeta Eiseniella tetraerda Aulodrilus pluriseta Stylodrilus heringianus Lumbriculidae spp. Hirudinea Batracobdella paludosa Erpobdella octoculata Chloroperla apicalis Perla burmeisteriana Perla sp. 13,0 4,4 Cloeon dipterum 4,4 4,4 Baetis fuscatus 4,4 13,0 Baetis rhodani 13,0 Isoperla obscura 21,7 Baetis vernus 4,4 4,4 Isogenus nubecula 4,4 Baetis buceratus 4,4 4,4 Agnetina sp. 4,4 4,4 4,4 4,4 8,9 Mollusca Gastropoda sp. 4,4 Lymnaea ovata 4,4 Lymnaea intermedia Lymnae auricularia Physa bulla ВстреВстречаемость, Ephemeroptera чаемость, % % Ephemeroptera 4,4 4,4 sp. Baetopus 4,4 4,4 wartensis Procloeon 4,4 8,9 ornatum 4,4 8,9 4,4 Brachyptera braueri Coleoptera Coleoptera sp. Hydraena gracilis Peltodytes caecus Dytiscus marginalis Helichus substriatus 4,4 4,4 4,4 4,4 78 13,0 8,9 13,0 52,2 4,4 8,9 4,4 Heptagenia flava 4,4 4,4 Ecdyonurus sp. 4,4 8,9 Ecdyonurus affinis Ecdyonurus venosus Epeorus rheophilis Diptera Blepharicera fasciata Prosimulium hirtipes Nigrobaetis pumilus Potamanthus luteus Rhitrhrogena sp Heptagenia sulfurea Heptagenia fuscogrisea Heptagenia coerulans 8,9 8,9 17,4 4,4 Physa fontinalis 8,9 Wilhelmia lineata 4,4 Physa tastei 4,4 Ibisia marginata 4,4 4,4 Elliptera omissa 4,4 Caenis macrura 8,9 4,4 4,4 Culex theileri Culex mimeticus Prionocera turcica Hemerodromia sp. Cricotopus baricomalis Cricotopus silvestris Syndiamesa nivosa Prodiamesa bathyphila Psectrocladius dilatatus Clinotanypus nervosus Simmulium caucaicum Zavrelia pentatoma Chironomidae sp. Odonata Onychogomphus forcipatus Ophiogomphus obscurus Gomphus vulgatissimus Cordulegaster annulatus 4,4 4,4 Acentrella sp. 4,4 Bithynia tentaculata Ancylus fluviatilis Valvata cristata Theodoxus sp. 4,4 Pisidium amnicum 4,4 Sphaeriastrum rivicola 4,4 Trichoptera Trichoptera sp. 8,9 Notidobia ciliaris 4,4 Cheumatopsyche lepida Hydropsyche ornatula Hydropsyche angustipennis Holocentropus picicornis Allotrichia pallicornis Apatania sp. 34,8 13,0 13,0 4,4 4,4 4,4 Ecnomus tenellus 8,9 Glossosoma boltoni 17,4 Agapetus sp. 4,4 Rhyacophyla pascoei Polycentropus flavomaculatis Molanna angustata Brachycentrus subnubilis Mystacides azurea 8,9 4,4 4,4 4,4 4,4 4,4 4,4 4,4 8,9 8,9 8,9 4,4 4,4 4,4 4,4 4,4 8,9 4,4 Hemiptera Nepa cinerea 4,4 Planariidae 8,9 Bdellocephala punctata 79 4,4 4,4 Oligoneuriella pallida Siphlonorus aestivalis 4,4 4,4 Для оценки меры сходства видового состава между исследованными реками использовался коэффициент Серенсена. Выявлено, что ни одна из исследуемых рек не обладает уникальным видовым составом. Наиболее схожими с точки зрения видового состава являются малые реки Ряпшь и Цеера (индекс Серенсена – 0,44) и относительно крупные реки Абхазии Галидзга и Кодор (0,42). Наиболее специфичным видовым составом зообентоса обладает р. Мааниквара у Ново-Афонской пещеры, которая имеет схожие виды только с р. Лидзава около пещеры у с. Лидзава (2 вида, индекс Серенсена – 0,27). В заключение отметим, что выявленный видовой состав макрозообентоса вносит существенный вклад в исследование биологического разнообразия малоисследованных и не исследованных водных объектов Абхазии, качество вод которых во многом является эталонным для водоемов и водотоков мира. Данные могут стать базовыми для мониторинга состояния водных объектов Абхазии. ОЦЕНКА БИОЛОГИЧЕСКОГО РАЗНООБРАЗИЯ ВОДНЫХ ОБЪЕКТОВ И ПРИБРЕЖНЫХ ТЕРРИТОРИЙ Г. КАЗАНИ Мингазова Н. М., Деревенская О. Ю., Палагушкина О. В., Замалетдинов Р. И., Набеева Э. Г., Зарипова Н. Р. Казанский (Приволжский) федеральный университет, г. Казань, nmingas@mail.ru Биоразнообразие – один из важнейших показателей, характеризующих устойчивость природных экосистем. При антропогенном воздействии и загрязнении наблюдается снижение видового богатства растений и животных, что особенно характерно для водных объектов урботерриторий. Город Казань считается вторым городом в Российской Федерации (после Санкт-Петербурга) по количеству водных ресурсов и водных объектов. Современный город располагается на берегу крупного Куйбышевского водохранилища, через весь город проходит река Казанка и цепь озер Кабан. В городе располагается также 4 притока р. Казанки (р. Нокса, Киндерка, Сухая и Солонка), более 180 малых озер и водноболотных угодий, имеются искусственные водные объекты – пруды, каналы, дрены. Большая часть водных объектов относятся к очень малым по площади, мелководным, зарастающим, иногда заболачиваемым озерам. Изучение биоразнообразия гидрофлоры и гидрофауны города с учетом разнотипности водных объектов представляет большой научный и практический интерес, служит целям оценки экологического состояния и мониторинга. 80 Исследования проводились Лабораторией водных экосистем (ЛОВЭ) КГУ в 2007–2008 гг. по программе инвентаризации и паспортизации водных объектов г. Казани. Всего было обследовано более 200 разнотипных объектов с разным химическим составом вод. В видовом составе фитопланктона было определено 190 таксонов рангом ниже рода из 8 отделов. По количеству видов преобладали зеленые (39 %) и диатомовые водоросли (31,6 %), что характерно для водоемов умеренной зоны. Наиболее часто отмечались Nitzschia palea (Kiitz) W. Sm. (65 %) из диатомовых водорослей, Chlamydomonas sp. (54 %) из зеленых, Euglena viridis Ehr. (52 %) из эвгленовых. Более 50 видов встречалось лишь 1–2 раза. Фитопланктон представлен в основном космополитными видами (88 %), также было определено 7 % голарктических и 2 % бореальных видов (Мингазова и др., 2007). Систематический анализ водной и водно-болотной растительности выявил 51 вид гидрофитов и гелофитов, входящих в 43 рода, 26 семейств, 4 класса и 3 отдела. Доминирующими видами являются поточник сжатый (Blysmus compressus (L.) Panz. ex Link), касатик сибирский (Iris sibirica L.), осока двудомная (Carex dioica L.), белозор болотный (Parnassia palustris L.), сальвиния плавающая (Salvinia natans (L.) All.). Редкими для водоемов г. Казани можно считать следующие виды – телорез алоевидный (Stratiotes aloides L.), дзанникелия ползучая (Zannichellia repens Boenn.), рдест пронзеннолистный (Potamogeton perfoliatus L.), омежник водный (Oenanthe aquatica (L.) Poir.), ситник черный (Juncus atratus Krock), осока сближенная (Carex appropinquata Schum.), шелковник жестколистный (Batrachium circinatum (Sibth.) Spach.), рогоз Лаксмана (Typha laxmanni Lepech.) и хвощ приречный (Equisetum fluviatile L.). В составе зоопланктона было выявлено 204 вида: коловраток – 99 видов, ветвистоусых ракообразных – 71, веслоногих рачков – 57, принадлежащих к 34 семействам, из 7 отрядов и 2 классов. Наиболее часто встречались из коловраток – Keratella quadrata (Muller) (27 %), Brachionus angularis Gosse (19 %); из ветвистоусых – Chydorus sphaericus (O. F. Muller) (47 %), Bosmina longirostris (O. F. Muller) (26 %), Simocephalus vetulus (O. F. Muller) (24 %), Daphnia longispina O. F. Muller (23 %); из веслоногих ракообразных – Eucyclops serrulatus (Fischer) (29 %), Mesocyclops leuckarti (Claus) (28 %). Редкими являются более 30 видов. В составе зообентоса водных объектов г. Казани выявлено 163 вида из 8 классов: олигохеты – 13 видов, пиявки – 6, нематоды – 1, моллюски 2 классов – 47, ракообразные – 2, паукообразные – 2 и насекомые – 91 вид. Выявлены «краснокнижные» виды – Argironeta aquatica L. (паук81 серебрянка), Ranatra linearis (L.) (ранатра палочковидная), Nepa сinerea (L.) (водный скорпион). Редко встречались хирономиды – Potthastia gaedii Meigen, Stictochironomus rosenscholdi Zetterstodt, Monopelopia tenuicalar Kieffer, Prodiamesa olivacea Meigen, Orhtocladius clarci Soponis, стрекозы – Coenagrion armatum Char, Ischnura elegans L., Coenagrion pulchellum L., Somatochlora metallica Vander Linden, двукрылые – Mistacides niger L., бабочки – Nymphula stagnata Donavan, Elophila nymphaeta L., Paraponyx stratiotata L., поденки – Baetis rodani Pictet, полужесткокрылые – Notonecta glauca L. Также редкими являлись представители групп Ptychopteridae, Megaloptera, Empedidae, Poridae (Bradysia sp., Megaselia sp.), Drosophilidae (Scaptomyza phalerata puppe Meigen). В ходе исследований водных объектов и околоводных территорий в г. Казани было зарегистрировано также 30 видов рыб, 11 видов земноводных, 4 вида пресмыкающихся, 104 вида птиц и 14 видов млекопитающих. Наибольшее число видов, внесенных в Красную книгу РТ, приуроченных к водоемам г. Казани, приходится на долю птиц (10 видов): болотная сова (Аsio flammeus), длиннохвостая неясыть (Strix uralensis), европейский козодой (Caprimulgus europaeus), зеленый дятел (Picus viridis), камышница (Gallinula chloropus), серая неясыть (Strix aluco), ушастая сова (Asio otus), большая выпь (Botaurus stellaris), малая выпь (Ixobrychus minutus), пастушок (Rallus aquaticus). В прибрежных биотопах обитают 3 вида амфибий – тритон гребенчатый (Triturus cristatus), жерлянка краснобрюхая (Bombina bombina), жаба серая (Bufo bufo), 1 вид рептилий – веретеница ломкая (Anguis fragilis), 1 вид млекопитающих – горностай (Mustela erminea), 1 вид рыб – обыкновенный подкаменщик (Cottus gobio). Результаты проведенных исследований, несомненно, указывают на значительное разнообразие растительного и животного мира водоемов г. Казани как в видовом, так и в таксономическом отношении. Так, в исследованных малых озерах выявлено до 50–80 % от числа видов, известных для озер Среднего Поволжья (Биоразнообразие и типология…, 2009). Но количество видов в водных объектах различается. С точки зрения видового разнообразия наиболее ценными являются озера и водно-болотные угодья, расположенные на первой надпойменной террасе р. Казанки и Волги, а также заливы водохранилища. Водные объекты представляют собой существенное звено в сохранении биоразнообразия урботерриторий, обеспечивают сохранение малочисленных видов животных в условиях города, нуждаются в охране и благоустройстве. 82 Л и т е р а т ур а Биоразнообразие и типология карстовых озер Среднего Поволжья / под ред. Н. М. Мингазовой. – Казань: Казанский гос. ун-т, 2009. – 222 с. Мингазова Н. М. Биоразнообразие водных объектов г. Казани / Н. М. Мингазова, О. Ю. Деревенская, О. В. Палагушкина, Л. Р. Павлова, Э. Г. Набеева, Н. Р. Зарипова, Р. И. Замалетдинов, Т. А. Кондратьева, Ю. И. Павлов, Е. Н. Унковская, М. Г. Борисович, Л. Ю. Халиуллина // Ученые записки Казанского университета. Сер., Естественные науки. – 2008. – Т. 150, кн. 4 – С. 252–260. РАЗНООБРАЗИЕ СИСТЕМ ГОДИЧНЫХ ПОБЕГОВ QUERCUS ROBUR L. В ЦЕНТРЕ ЕВРОПЕЙСКОЙ ЧАСТИ РОССИИ Стаменов М. Н. Институт физико-химических и биологических проблем почвоведения РАН, г. Пущино, mslv-eiksb@inbox.ru Годичный и элементарный побеги являются основными структурными единицами побегового тела растения (Гетманец, 2011), их разнообразие и изменение в ходе онтогенеза зависят как от действия эндогенных факторов, так и от экологических условий. У вида-идентификатора широколиственных лесов дуба черешчатого Quercus robur L. разнообразие путей индивидуального развития описано преимущественно на уровне крупных скелетных осей (Серебряков, 1962); особенности собственно побегов рассмотрены в работе Т. Н. Астаповой (1954). В то же время модульный подход к изучению побеговой системы Quercus robur L., анализ элементарных структурных единиц применялся гораздо реже. Например, можно отметить работу О. Б. Михалевской (1987). Поэтому целью нашей работы была типизация систем годичных побегов и характеристика их количественных признаков у прегенеративных особей, произрастающих в сообществах с различным уровнем освещенности и различным сукцессионным статусом. Исследования проводились в 2012 и 2013 годах в Южном Подмосковье и в заповеднике «Калужские Засеки». В Подмосковье изучались сформированные 50–60-летние березняки и сосняки с выраженной ярусностью и густым подлеском, разреженные сосняки, сложные липняки (разреженный древостой и ветровальное «окно»), луговая поляна с затенением отдельными деревьями и суходольный луг. В Калужской области изучались заросшие 20–25-летним березняком пашни с различной плотностью интенсивно растущих широколиственных видов и суходольный луг. Всего описано 182 особи. Описывались системы годичных побегов в дистальных частях скелетных ветвей и лидерных осей по 83 измененной методике (Бобровский, Бобровская, 1998). У материнских осей (элементарных побегов) и боковых побегов ветвящихся метамеров измерены значения количественных признаков: длины побегов и число междоузлий. Системы годичных побегов классифицированы по сочетанию следующих признаков: число элементарных побегов, наличие или отсутствие у них ветвления, наличие пролепсиса, характер дополнительного роста боковых побегов за вегетационный сезон (второй сезонный прирост из апикальной почки и/или пролепсис). Наибольшее разнообразие типов побеговых систем выявлено при максимальной освещенности – на лугах. На лугах в Московской и Калужской областях преобладают годичные побеги с ветвлением из зимовавших почек без дополнительного сезонного роста боковых побегов, при этом в луговом сообществе «Калужских Засек» значительна и доля годичных побегов с дополнительным сезонным ростом боковых побегов. В луговом сообществе в «Калужских Засеках» значительна доля годичных побегов с двумя элементарными, каждый из которых несет боковые побеги из зимовавших почек. Кроме того, на суходольном лугу в «Калужских Засеках» выявлены годичные побеги, у которых в пределах одного элементарного побега присутствует как пролептическое ветвление, так и ветвление из перезимовавших почек (на следующий год). Годичные побеги с пролептически ветвящимися элементарными побегами выявлены, в основном, в луговых сообществах Московской и Калужской областей. На луговой поляне в Московской области спектр систем годичных побегов и преобладающие типы побегов в целом такие же, как и на суходольном лугу. В раннесукцессионных лесных сообществах «Калужских Засек» разнообразие типов побеговых систем выше, чем в лесных сообществах Московской области, вероятно, это связано с отсутствием на зарастающих пашнях выраженного второго яруса древостоя и мозаичностью расселения подроста других древесных видов. Кроме того, приживание сеянцев дуба происходило в еще слабо сформированных и сомкнутых древостоях, в отличие от лесов Московской области. В Московской области в сомкнутых сосновых и березовых древостоях с подлеском преобладают неветвящиеся годичные побеги с одним элементарным побегом, в разреженных древостоях неветвящиеся и ветвящиеся годичные побеги с одним элементарным выявлены в примерно равном соотношении. В луговых сообществах Московской и Калужской областей наиболее полный побеговый спектр выявлен у особей старшего имматурного состояния, в лесных сообществах – у особей виргинильного состояния. 84 Значения количественных признаков ветвящихся материнских осей во всех сообществах достоверно выше значений у неветвящихся осей, начиная со старшего имматурного состояния. В лесных сообществах динамика значений количественных признаков при переходе от младших возрастных состояний к старшим индивидуальна для каждого сообщества, в луговых сообществах значения последовательно возрастают с изменением возрастного состояния. Наибольшие значения длин элементарных побегов и числа их междоузлий выявлены у неветвящихся побегов летней стадии роста в луговых сообществах – в среднем, 25– 37 см в зависимости от возрастного состояния. Наиболее существенные различия между скелетными ветвями и лидерной осью по значениям количественных признаков обнаружены в луговых сообществах также у летних («Ивановых») неветвящихся элементарных побегов. Модальное число боковых побегов на материнских осях в лесных сообществах – 1–4, в луговых – 5–8. Число боковых побегов увеличивается с изменением возрастного состояния и яруса кроны. В лесных сообществах преобладают материнские оси с укороченными боковыми побегами, в луговых – с чередованием укороченных и удлиненных. С изменением возрастного состояния положение первого бокового побега смещается ближе к основанию материнской оси. Положение наиболее длинного междоузлия на элементарном побеге является наиболее вариабельной из всех описанных характеристик. Таким образом, различия в экологических условиях между сообществами (прежде всего – различия в уровне освещения) сказываются на вариабельности строения и значений количественных признаков у систем годичных побегов подроста Quercus robur L. Работа выполнена № 12-04-01734. при финансовой поддержке проекта РФФИ Л и т е р а т ур а Астапова Т. Н. Рост и формирование побегов дуба в лесах Подмосковья / Т. Н. Астапова // Уч. зап. МГПИ. – 1954. – Т. XXXVII. – С. 135–155. Бобровский М. В. Структурно-функциональные элементы кроны и их пространственные отношения у Acer platanoides (Aceraceae) и Populus tremula (Salicaceae) / М. В. Бобровский, Н. Е. Бобровская // Ботан. журн. – 1998. – № 4. – С. 114–124. Гетманец И. А. Экологическое разнообразие и биоморфология рода Salix L. Южного Урала: автореф. дис. … д-ра биол. наук / И. А. Гетманец. – Омск, 2011. Михалевская О. Б. Ритмичность процессов роста и морфогенеза побегов в роде Quercus L. / О. Б. Михалевская // Морфогенез и ритм развития высших растений: межвуз. сб. науч. тр. / под ред. Т. И. Серебряковой. – М.: Изд-во Моск. гос. пед. ин-та им. В. И. Ленина, 1987. – С. 33–38. Серебряков И. Г. Экологическая морфология растений / И. Г. Серебряков. – М.: Высш. шк., 1962. 85 К ЭКОЛОГИИ НЕМАТОД РОДА APHELENCHOIDES Хусаинов Р. В. Центр паразитологии Института проблем экологии и эволюции им. А. Н. Северцова РАН, г. Москва, ren_khusainov@yahoo.com Представители рода Aphelenchoides являются обширной группой нематод, которые населяют различные биоценозы и экологические ниши. Они обнаружены в почве, растительных остатках, древесине, на мхах и лишайниках, в пресноводном детрите и иле, а также внутри растений (Барановская, 1981; Hunt, 1993). Афеленхоидесы широко распространены по всему земному шару, то есть являются космополитами. К группе фитопаразитов принадлежат лишь несколько представителей данного рода, большинство же питается содержимым мицелия грибов-микромицетов. Тем не менее, приуроченность некоторых видов к определенному местообитанию и их трофическая специализация до сих пор остается неясной. В 2011–2013 гг. были проведены сборы ряда субстратов в различных ценозах Европейской части России на предмет обнаружения нематодафеленхид. Результаты подтвердили широкую встречаемость афеленхоидесов в различных биотопах. Они встречались в гниющих растительных остатках, на погибших растениях, на поверхности камней и деревьев, на мхах и лишайниках, на поверхности шляпочных грибов. Афеленхоидесы также обнаруживались на некротизированных листьях и в береговом детрите, где их численность колебалась от 3 до 12 особей/100 см3 субстрата. Наличие афеленхоидесов на том или ином субстрате еще не означает, что это их основная среда обитания, – они могли быть смыты потоками дождевой воды или были занесены иным способом. Наиболее часто и в высокой численности (60–300 особей/ 100 см3 субстрата) свободноживущие афеленхоидесы присутствовали в растительных остатках и на поверхности поврежденной или погибшей древесины. Таким образом, они приурочены к обитанию в условиях сапробиотической среды, где в достаточной мере содержится пищевой ресурс в виде плесневых грибов. Результаты исследований также показали, что некоторые виды имеют строгую привязанность к определенной среде обитания и вертикальной структуре биоценоза (так, виды A. macromucrons, A. rhytium, A. eximius – типичные ксилобионты), другие встречаются в различных биотопах без привязки к ярусности (A. composticola, A. saprophilus). Афеленхоидесы отличаются большим разнообразием своих биологических и экологических особенностей. Паразитами растений являются A. besseyi (Christie, 1942), A. fragariae (Ritzema Bos, 1890) Christie, 1932, 86 A. ritzemabosi (Schwartz, 1911) Steiner & Buhrer, 1932 и, возможно, A. panaxofolia Liu et al., 1999. С водной средой ассоциированы только 4 вида: A. fluviatilis Andrassy, 1960, A. gynotylurus Timm & Franklin, 1969, A. marinus Timm & Franklin, 1969, A. robustus Gagarin, 1997. Один вид (A. xylocopae Kanzaki, 2006) обнаружен на пчеле (Xylocopa appendiculata circumvolans). Остальные представители рода, как правило, являются ризо-, сапро- или ксилобионтами. Несмотря на то, что многие авторы отмечают наличие афеленхоидесов в стеблях, листьях и корнях растений, наличие у них способности к фитопаразитизму требует отдельных экспериментальных подтверждений. Обнаружение афеленхоидесов на насекомых, скорее всего, является форезией, а не паразитизмом. Вопрос питания афеленхоидесов микроводорослями также пока остается открытым. Исследования нематод рода Aphelenchoides будут продолжены по направлениям выяснения их трофической специализации, жизненного цикла и экологической роли. Л и т е р а т ур а Барановская И. А. Нематоды растений и почв: афеленхоидиды и сейнуриды / И. А. Барановская. – М.: Наука, 1981. – 235 с. Gagarin V. G. Aphelenchoides robustus sp. n., a new freshwater nematode from archipelago Novaya Zemlya (Nematoda: Aphelenchoididae) / V. G. Gagarin // Zoosystematica Rossica. – 1997. – Vol. 5 (2). – P. 209–211. Hunt D. J. Aphelenchida, Longidoridae and Trichodoridae: their systematics and bionomics / D. J. Hunt. – Wallingford, UK: CABI Publishing, 1993. – 352 pp. Kanzaki N. Description of Aphelenchoides xylocopae n. sp. (Nematoda: Aphelenchoididae), the first observed association between nematodes and carpenter bees / N. Kanzaki // Nematology. – 2006. – Vol. 8 (4). – P. 555–562. Liu W. A new species of genus Aphelenchoides, leaf nematode of American ginseng, Aphelenchoides panaxofolia n. sp. (Nematoda: Aphelenchoidae) / W. Liu, X. Wu, Y. Duan, Y. Liu // Acta Phytopathologica Sinica. – 1999. – Vol. 29 (4). – P. 360–363. 87 Секция 2 ТАКСОНОМИЧЕСКОЕ И СТРУКТУРНОЕ БИОРАЗНООБРАЗИЕ НА ОСОБО ОХРАНЯЕМЫХ И НАРУШЕННЫХ ТЕРРИТОРИЯХ О ФИТОРАЗНООБРАЗИИ ЛУГОВ МАРИЙСКОГО ПРИСУРЬЯ Бекмансуров М. В., Мустаева Н. В. Марийский государственный университет, г. Йошкар-Ола, ecology@marsu.ru Создание Чебоксарского водохранилища привело к существенному преобразованию биогеоценотического покрова долины р. Волга и ее притоков, в том числе и р. Суры – вырублен лес на значительных площадях, исчезли пойменные дубравы, произошло затопление и подтопление высокопродуктивных сенокосных и пастбищных угодий. В 2012– 2013 гг. нами проведены исследования луговых фитоценозов, в правобережной части долины р. Суры в пределах границы Республики Марий Эл. На данной территории водоразделы и крутые склоны второй надпойменной террасы заняты главным образом широколиственными лесами и их производными. Травяные фитоценозы расположены на участках первой и пологих склонах второй надпойменной террасы, свободных от древесной растительности. Пойма Суры затоплена водохранилищем. Небольшие по площади травяные фитоценозы сохранились и на водораздельной территории в окрестностях населенных пунктов. В последнем случае происходит их зарастание кустарниками и подростом деревьев. Долинные луговые фитоценозы пока еще используются местным населением как сенокосные угодья и пастбища, их деградация отмечена лишь на опушках и избыточно увлажненных участках. В ходе исследований нами выполнено 27 геоботанических описаний на пробных площадях размером 100 м2. Пространственная структура луговых фитоценозов весьма сложная – хорошо дренированные участки здесь чередуются с переувлажненными, регулярно выкашиваемые угодья с заброшенными или нерегулярно используемыми сенокосами. Усложняют пространственную структуру лесные колки и ручьи. В силу указанных причин использование методов кластеризации и ординации для выделения сходных групп геоботанических описаний оказалось не эффективным. Поэтому для дифференциации пула описаний мы использовали фитоиндикационный метод 88 Шкала Tr (Экологические…, 2010), проведя обработку флористических списков по шкалам увлажнения (Hd) и трофности почв (Tr) (Цыганова, 1983). При этом использована компьютерная программа EcoscaleWin (Компьютерная…, 2008). Экологическое пространство местообитаний луговых фитоценозов, представленное как результат ординации балльных оценок по шкалам увлажнения и трофности почв показано на рисунке 1, из которого видно, что массив описаний весьма четко разделяется на 3 группы по фактору увлажнения. Первая группа представляет собой совокупность 17 геоботанических описаний, выполненных на хорошо дренированных участках первой и склонах второй надпойменной террасы Суры, а также на водораздельных территориях с свеже-лесолуговым увлажнение. 10 9,5 9 8,5 8 7,5 7 6,5 6 1 2 3 10 12 14 16 18 Шкала Hd Рисунок 1 – Экологическое пространство местообитаний луговых фитоценозов по увлажнению и солевому режиму почв Эту группу дифференцируют мезофиты: тимофеевка луговая (Phleum pratense L.) (класс константности V, среднее обилие 1), ежа сборная (Dactylis glomerata L.) (V, 1), нивяник обыкновенный (Leucanthemum vulgare Lam) (V, 2), подмаренник мягкий (Galium mollugo L.) (IV, 2), цикорий обыкновенный (Cichorium intybus L.) (IV, +), подорожник ланцетный (Plantago lanceolata L.) (IV, 1). Среди прочих видов с высоким постоянством, но меньшим обилием (± 1) здесь встречаются лютик едкий (Ranunculus acris L.), зверобой пятнистый (Hypericum maculatum Crantz), горлюха ястребинковая (Picris hieracioides L.), первоцвет весенний (Primula veris L.), черноголовка обыкновенная (Prunella vulgaris L.), щавель кислый (Rumex acetosa L.), клевер луговой (Trifolium pratense L.), горошек четырехсемянный (Vicia tetrasperma (L.) Schreb). 89 Вторая группа из 7 описаний характеризуется бόльшим увлажнением местообитаний (сыровато-лесолуговым). Ее дифференцируют гигрофиты: таволга вязолистная (Filipendula ulmaria (L.) Maxim.) (V, 1), осока лисья (Carex vulpina L.) (III, 1), девясил иволистный (Inula salicina L.) (II, 1), осока стройная (Carex acuta L.) (III, +), луговик дернистый (Deschampsia cespitosa (L.) P. Beauv.), гравилат речной (Geum rivale L.), ива пепельная (Salix cinerea L.), лапчатка гусина (Potentilla anserina L.), чистец болотный (Stachys palustris L.), осока мохнатая (Carex hirta L.), повой заборный (Calystegia sepium (L.) R. Br.) (III, +). Реже в описаниях этой группы встречаются зюзник европейский (Lycopus europaeus L.), зюзник высокий (L. exaltatus L. f.), вербейник обыкновенный (Lysimachia vulgaris L.), мята полевая (Mentha arvensis L.), щавель водяной (Rumex aquaticus L.). Наличие в описаниях этой группы ивы пепельной свидетельствует о нерегулярном сенокошении и начальных этапах зарастания лугов. Третью группу описаний характеризуют виды, произрастающие в местах избыточного болотно-лесолугового увлажнения, где временно или постоянно имеются небольшие стоячие или слабопроточные водоемы. К ним относятся: сусак зонтичный (Butomus umbellatus L.), частуха подорожниковая (Alisma plantago-aquatica L.), щавель приморский (Rumex maritimus L.), стрелолист обыкновенный (Sagittaria sagittifolia L.), почечуйник почечуйный (Persicaria maculata (Raf.) A. Love & D. Love), рогоз узколистный (Typha angustifolia L.). Спектр семейств, лидирующих по числу видов, представлен на рисунке 2. Рисунок 2 – Спектр ведущих семейств в луговых сообществах 90 Структурное разнообразие травяных сообществ отражают 14 эколого-ценотических групп сосудистых растений (Биологическое…, 2003). Среди них доминируют виды свежих лугов (56), травяноболотные (30), нитрофильные (20), опушечные (19). Следует отметить высокий уровень фиторазнообразия луговых фитоценозов исследованной территории, в особенности долинных. Их альфа-разнообразие характеризуют 166 видов сосудистых растений 40 семейств. Средняя видовая насыщенность составляет для каждой из групп описаний 23, 19 и 7 видов на 100 м2. Высокий уровень биоразнообразия луговых фитоценозов обуславливает необходимость их включения в состав территории будущего заказника республиканского значения, создание которого планируется в ближайшее время. При этом следует поощрять сенокошение и умеренный выпас на лугах долины Суры, что будет препятствовать их дальнейшей деградации и зарастанию древесной растительностью. Работа выполнена при финансовой поддержке Департамента экологической безопасности, природопользования и защиты населения Республики Марий Эл Л и т е р а т ур а Биологическое разнообразие растительного покрова национального парка «Марий Чодра»: научное издание / Л. Б. Заугольнова, Л. А. Жукова, М. В. Бекмансуров и др. – Йошкар-Ола, 2003. – Ч. 1. – 136 с. Экологические шкалы и методы анализа экологического разнообразия растений: монография / Мар. гос. ун-т; Л. А. Жукова, Ю. А. Дорогова, Н. В. Турмухаметова [ и др. ]; под общ. ред. проф. Л. А. Жуковой. – Йошкар-Ола, 2010. – 368 с. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoscaleWin: учеб. пособие / Е. В. Зубкова, Л. Г. Хакина, Т. И. Гроклина, Ю. А. Дорогова. – Йошкар-Ола, 2008. – 96 с. ХАРАКТЕРИСТИКА ЛЕСНОГО ПОКРОВА МАРИЙСКОГО ПРИСУРЬЯ Бекмансуров М. В., Рыбакова М. А. Марийский государственный университет, г. Йошкар-Ола, ecology@marsu.ru В 2012–2013 гг. нами проведены исследования лесных фитоценозов в правобережье р. Суры в пределах границ Республики Марий Эл. Район почти целиком сложен мощной толщей пермских карбонатных пород татарского яруса, расчленен глубокими и узкими оврагами и неширокими долинами рек. Плато и верхние части склонов водоразделов прикрыты лессовидными суглинками. На данной территории произрастают главным образом широколиственные леса и их производные. Все они относятся к категории защитных. Пойма Суры здесь в настоящее время полностью залита Чебоксарским водохранилищем, первая надпоймен91 ная терраса простирается от нескольких десятков метров до 1 км. Склон коренной террасы крутой, местами осыпающийся. В ходе исследования нами по стандартной методике выполнено 51 геоботаническое описание на пробных площадях размером 20 × 20 м. Для выделения групп описаний, сходных по флористическому составу и встречаемости видов, использованы методы многомерной статистики – кластерный анализ на основе алгоритма Варда и метод непрямой ординации описаний в абстрактных осях варьирования с использованием соответствий с удаленным трендом (Джонгман и др., 1999). В первый кластер объединились большинство описаний (39), выполненных главным образом в липняках, дубняках, кленовниках, березняках, осинниках и ясеневниках, а также посадках сосны сибирской и сосны обыкновенной. В подлеске, подросте и травяно-кустарничковом ярусе всех этих фитоценозов преобладают одни и те же виды растений, главным образом неморальной эколого-ценотической группы. Следовательно, большинство из них являются производными от дубняков. Доминантами древесного яруса в порядке убывания являются липа сердцевидная (Tilia cordata Mill.), клен платанолистный (Acer platanoides L.), ясень обыкновенный (Fraxinus excelsior L.), дуб черешчатый (Quercus robur L.), береза повислая (Betula pendula Roth). В подросте преобладают клен платанолистный, вяз шершавый (Ulmus glabra Huds.). И в древостое, и в подросте примерно 20 % фитоценозов этой группы произрастает ясень высокий, включенный в Красную книгу Республики Марий Эл. В районе нашего исследования проходит северная граница ареала сплошного распространения этого вида. В левобережной части Марий Эл он встречается крайне редко. В подлеске преобладают лещина (Corylus avellana L.), рябина обыкновенная (Sorbus aucuparia L.), черемуха птичья (Padus avium Mill.). Менее распространены бересклет бородавчатый (Euonymus verrucosa Scop.) и жимолость обыкновенная (Lonicera xylosteum L.). В травяно-кустарничковом ярусе обычными видами являются сныть обыкновенная (Aegopodium podagraria L.), пролесник многолетний (Mercurialis perennis L.), медуница неясная (Pulmonaria obscura Dumort.). Практически повсеместно распространены дубравные эфемероиды, также дифференцирующих данный кластер – зубянка пятилистная (Dentaria quinquefolia M. Bieb.), ветреничник лютичный (Anemonoides ranunculoides (L.) Holub), хохлатка плотная (Corydalis solida (L.) Clairv.) и хохлатка Маршалла (Corydalis marschalliana (Pall. ex Willd.) Pers.). Зубянка пятилистная и хохлатка Маршалла включены в Красную книгу Республики Марий Эл (2013). По доминантной классификации большинство сообществ данной группы относятся к группе типов широколиственных неморальных лесов. 92 Видовое богатство сообществ этого кластера образуют 125 видов сосудистых растений при средней видовой насыщенности 20 видов на 400 м2. Второй кластер включает в себя 3 описания дубняков, являющихся искусственными насаждениями и располагающихся на водораздельных территориях. Дуб черешчатый является абсолютным доминантом древостоя. Ярус подлеска сообществ этого кластера образуют лещина обыкновенная, черемуха птичья, рябина обыкновенная, калина обыкновенная (Viburnum opulus L.), яблоня лесная (Malus sylvestris Mill.). В подросте незначительным обилием, присутствуют клен платанолистный и липа сердцелистная. В травяно-кустарничковом ярусе доминирует сныть обыкновенная. Также здесь произрастают чистяк весенний (Ficaria verna Huds.), земляника обыкновенная (Fragaria vesca L.), лютик ползучий (Ranunculus repens L.), горошек заборный (Vicia sepium L.), норичник узловатый (Scrophularia nodosa L.), осока лесная (Carex sylvatica Huds.), горицвет кукушкин (Coccyganthe flos-cuculi (L.) Fourr.), ежа сборная (Dactylis glomerata L.), луговик дернистый (Deschampsia cespitosa (L.) P. Beauv.), душистый колосок обыкновенный (Anthoxanthum odoratum L.). Обращает на себя внимание большое количество луговых видов. По-видимому посадки были созданы на заброшенных сельхозугодиях. Фиторазнообразие сообществ данного кластера характеризуют 68 вида сосудистых растений. Средняя насыщенность составляет 33 вида на 400 м². Третий кластер включает в себя 9 описаний, выполненных в серо- и черноольшаниках и ивняках долины Суры и глубоких оврагах. В древесном ярусе наряду с ольхой встречаются вяз гладкий (Ulmus laevis Pall.), береза пушистая (Betula pubescens Ehrh.). Черноольшаники произрастают в условиях застойного увлажнения, а сероольшаники в достаточно хорошо дренированных оврагах. В подлеске доминирует черемуха птичья. Реже встречаются лещина обыкновенная и яблоня лесная. В подросте преобладают вяз гладкий, клен платанолистный. В травяно-кустарничковом ярусе наиболее распространена крапива двудомная (Urtica dioica L.). Дифференцирующими видами кластера являются гигрофиты: хмель обыкновенный (Humulus lupulus L.), зюзник европейский (Lycopus europaeus L.), чистец болотный (Stachys palustris L.) и подмаренник болотный (Galium palustre L.). Фиторазнообразие сообществ данного кластера характеризуют 130 видов сосудистых растений. Средняя видовая насыщенность низкая, всего 16 видов на 400 м². 93 Анализ структурного разнообразия исследованных фитоценозов с использованием классификации эколого-ценотических групп сосудистых растений Европейской России (Восточноевропейские…, 2004) показал, что в сообществах первых двух кластеров преобладают виды неморальной группы, за ними следуют нитрофильные и луговые виды (см. табл.). В сообществах третьего кластера доля неморальных и нитрофильных видов практически одинакова. Доля видов бореальной эколого-ценотической группы здесь существенно ниже, чем в широколиственных лесах левобережной части Республики Марий Эл (Биологическое…, 2003), где соотношение бореальных и неморальных видов примерно одинаковое. Спектры эколого-ценотических групп в лесных сообществах Число видов разных эколого-ценотических групп Класстеры неморальная нитрофильная (черноольшаниковая) бореальная луго вая I 50* 20 7 6 II 29 14 2 9 III 28 25 4 11 аллювиальная низинных болот 1 1 2 * из расчетов исключены виды встречаемостью менее 5 %. В целом исследованная территория характеризуется высоким уровнем фиторазнообразия. Нами обнаружено 202 вида сосудистых растений 59 семейств, из них 6 видов включены в Красную книгу Республики Марий Эл. Работа выполнена при финансовой поддержке Департамента экологической безопасности, природопользования и защиты населения Республики Марий Эл Л и т е р а т ур а Биологическое разнообразие растительного покрова национального парка «Марий Чодра»: научное издание / Л. Б. Заугольнова, Л. А. Жукова, М. В. Бекмансуров и др. – Йошкар-Ола, 2003. – Ч. 1. – 136 с. Восточноевропейские леса: история в голоцене и современность. – М.: Наука, 2004. – Кн. 1. – 479 с. Джонгман Р. Г. Г. Анализ данных в экологии сообществ и ландшафтов / Р. Г. Г. Джонгман, С. Дж. Ф. Тер Брак, О. Ф. Р. Ван Тонгерен. – М.: РАСХН, 1999. – 306 с. Красная книга Республики Марий Эл. Растения. Грибы / Мар. гос. ун-т. – Йошкар-Ола, 2013. – 324 с. 94 ХОЗЯЙСТВЕННАЯ ДЕЯТЕЛЬНОСТЬ НА ПРУДУ ТРЕТИЙ (ГПЗ «САРАТОВСКИЙ») И ЕЕ ВЛИЯНИЕ НА СОСТАВ ГНЕЗДОВОГО НАСЕЛЕНИЯ ПТИЦ Беляченко А. А. СГТУ имени Ю. А. Гагарина, г. Саратов, belyachenkoaa@mail.ru Пруды на территории государственного природного заказника (ГПЗ) «Саратовский» являются своеобразным местообитанием с уникальным сочетанием экологических факторов. Существенный градиент увлажнения при движении от уреза воды обусловливает высокую степень пространственной неоднородности данного природно-территориального комплекса. Следствием этого является ярко выраженная поясность прибрежно-водной растительности. В силу существенных различий характеристик растительных сообществ этих поясов (высоты травостоя, проективного покрытия, биоморфологического спектра и т. д.) они оказываются пригодными для гнездования разных видов птиц. Таким образом, в каждом растительном поясе формируется своеобразное население птиц. Кроме того, гнездовые участки отдельных видов птиц неоднородны и включают участки из нескольких зон. Пруды на территории ГПЗ «Саратовский» используются населением для различных целей. Среди них разведение рыбы, выпас и водопой скота, отдых. Это не может не сказываться на структуре населения птиц прилегающей к пруду территории. На некоторых прудах эти изменения незначительны и растянуты во времени, на других – хозяйственная деятельность ведется очень активно, вследствие чего изменения резки и хорошо заметны. Исследования проводились на пр. Третий, находящемся между с. Красавка и с. Никольское (координаты центра пруда 51º13,816´с. ш., 47º47,577´в. д.). Учеты птиц проводились на фиксированных маршрутах, заложенных вдоль берегов пруда таким образом, чтобы учетчик мог фиксировать птиц не только на зеркале водоема, но и на прилегающей территории (Ларина, 1981; Buckland at al., 2005). Население птиц исследовалось с 15 мая по 15 июля. Антропогенная нагрузка оценивалась исходя из частоты использования пруда и прилегающей территории в результате хозяйственной деятельности населения (табл.). Самой низкой антропогенная нагрузка оказалась в 2011 году. Пруд только начал использоваться для разведения рыбы. Арендаторы проводили подкормку только один раз, а серьезных колебаний уровня воды не наблюдалось. В 2012 г. подкормка проводилась также только однажды, однако часть акватории пруда была реорганизована для разведения молоди рыб. Осенью пруд был спущен, что сказалось на уровне воды в 2013 г. В этом же году подкормка рыбы стала проводиться регулярно, 95 а ее объемы возросли. Кроме того, местные жители стали использовать пруд для отдыха и рыбалки. На пруду в последний год исследований регулярно отмечались рыбаки, находящиеся здесь продолжительное время. Влияние выпаса и водопоя скота также усиливалось с 2011 по 2013 гг. Это связано с увеличением поголовья стада с 75–80 до 100– 120 голов (по глазомерной оценке) и появлением в его составе овец. Как следствие этого происходит более интенсивное вытаптывание растительности вблизи уреза воды и стравливание луговых (пырейных) участков и участков сорной растительности в пределах неудобий и сенокосов вблизи пруда. К иным видам использования акватории пруда и прилегающей территории относятся выявленные в 2012 г. факты браконьерства, а также вырубка сухостойных деревьев на плотине пруда. В 2013 г. была проведена сплошная рубка некрупных кустов ив (Salix) вдоль всей береговой линии пруда. Кроме этого, стал активно использоваться участок дороги, находящийся вблизи берега пруда. Характер использования акватории и прилегающей территории пруда Третий в 2011–2013 гг. Год Использование пруда и прилегающей территории Неделя наблюдений (с 15 мая по 15 июля) 1 2 3 4 5 6 7 8 Выпас и водопой скота 2011 Разведение рыбы Отдых людей Иные виды использования Выпас и водопой скота 2012 Разведение рыбы Отдых людей Иные виды использования Выпас и водопой скота 2013 Разведение рыбы Отдых людей Иные виды использования – влияния фактора не отмечено. – в ходе каждого второго учета птиц фиксируются результаты использования пруда и прилегающей территории в данном виде хозяйственной деятельности. – в ходе каждого учета птиц фиксируются результаты использования акватории пруда и прилегающей территории в данном виде хозяйственной деятельности. 96 Описанные выше изменения структуры хозяйственного использования акватории пруда привели к существенным изменениям в составе и структуре гнездового населения птиц. С интенсификацией хозяйственной деятельности наблюдается снижение числа видов (с 24 до 17) и суммарной численности (с 82 до 57 пар). Наибольшее обеднение видового разнообразия наблюдается среди группировки водных и околоводных птиц. Единовременная вырубка сухостойных деревьев приводит к исчезновению из состава орнитофауны скворцов (Sturnus vulgaris) и полевых воробьев (Passer montanus). При этом обилие обыкновенного скворца, чье гнездование связано с использованием дупел и морозобойных трещин таких деревьев, снизилось до нуля, в то время как несколько пар полевых воробьев перешли к гнездованию в старых гнездах сорок (Pica pica). Интенсивное вытаптывание и стравливание прилегающих к пруду луговых участков в результате выпаса сказывается на численности желтой трясогузки (Motacilla flava). Наиболее стабильным оказывается обилие птиц кустарниковой группировки. Их гнездовые участки расположены в зарослях полыни высокой (Artemisia procera) (серая славка (Sylvia communis), северная бормотушка (Hippolais calligata), варакушка (Luscinia svecica) и лоха узколистного (обыкновенный жулан (Lanius collurio), славка-завирушка (S. curruca). Также относительно стабильным оказывается обилие обыкновенной (Emberiza citrinella) и садовой (E. hortulana) овсянок, чьи гнезда чаще всего расположены под защитой кустарников или невысоко над землей (чаще всего это куртины лоха (Elaeagnus sp.), окруженного зарослями полыни высокой). Обилие двух видов каменок (обыкновенной (Oenanthe oenanthe) и плясуньи (O. isabellina) за период исследования несколько увеличивается. Возможно, это связано с увеличением числа старых брошенных нор рыжеватого суслика, пригодных для гнездования. Некоторые птицы в указанный период гнездятся в пределах других участков, однако пруд является частью этих участков или используется птицами для добывания корма. Изменения в составе и структуре растительности пруда сильно сказываются на поведении этих птиц. В частности, пруд используется для охоты орланом-белохвостом (Heliaeetus albicilla). При этом присадой для птиц служили высокие сухие деревья на плотине. После того, как деревья были спилены, птица реже стала посещать пруд. Другие виды, такие как серая (Ardea cinerea) и белая цапли (Egretta alba), несмотря на постоянное беспокойство со стороны человека, продолжают посещать пруд, питаясь молодью рыб на мелководье. Одним из неблагоприятных факторов, воздействующим на все гнездовое население птиц пруда и его окрестностей являются врановые. 97 Подкормку рыб арендаторы осуществляют большим количеством жмыха подсолнечника и иногда комбикорма. Этот пищевой ресурс привлекает большое количество грачей (Corvus frugileus) и галок (C. monedula), прилетающих сюда из находящихся поблизости колоний в лесополосах и с. Красавка. Одновременно у береговой линии пруда может скапливаться до 300–350 грачей. Они являются существенным фактором беспокойства для гнездового населения пруда, а нередко уничтожают кладки других птиц. Это прежде всего сказывается на численности открыто гнездящихся водных и околоводных птиц. На пруду также отмечен ряд фактов браконьерства. Многие водоплавающие птицы, такие как кряква (Anas platyrhynchos), пеганка (Tadorna tadorna), лысуха (Fulica atra), чомга (Podiceps cristatus) и чирок-трескунок (A. querquedula) отмечены в пределах акватории, однако из-за беспокойства не гнездились на пруду, несмотря на наличие подходящих для гнездования мест. Таким образом, антропогенное воздействие приводит к снижению видового разнообразия орнитофауны природно-территориального комплекса пруда Третий. Наиболее сильно сказываются на обилии птиц выпас скота вблизи пруда и разведение рыбы, сопровождающееся подкормкой. Л и т е р а т ур а Ларина Н. И. Учебное пособие по методике полевых исследований экологии наземных позвоночных / Н. И. Ларина, В. Л. Голикова, Л. А. Лебедева. – Саратов: Изд-во Сарат. ун-та, 1981. – 98 с. Buckland S. T. Introduction to distance sampling. Estimating abundance of biological populations / S. T. Buckland, D. R. Anderson, K. P. Burnham at al. – Oxford: Oxford University Press, 2005. – 432 p. ЭКОЛОГИЧЕСКАЯ ОЦЕНКА НЕКОТОРЫХ РАСТИТЕЛЬНЫХ СООБЩЕСТВ ЧЕЛЯБИНСКОГО (ГОРОДСКОГО) БОРА Гетманец И. А.1, Калашников Н. В.2 1 2 Челябинский государственный университет, г. Челябинск, igetmanec@mail.ru Центр детский экологический г. Челябинска, г. Челябинск, school85nk@mail.ru Рекреационные леса имеют особую ценность для поддержания здоровья и сохранения работоспособности населения. Особенно их роль возрастает в условиях напряженной экологической обстановки, характерной для большинства крупных промышленных городов, к числу которых относится г. Челябинск. Сохранение состояния Челябинского городского бора является актуальной задачей вследствие того, что он выполняет рекреационное, санитарно-гигиеничное, водорегулирующее значение, а также представляет, собой реликтовое сообщество перигляциальной лесостепи. 98 Проведенные немногочисленные исследования Челябинского (городского) бора были посвящены определению критериев к его устойчивости, рекреационной нагрузке, разработке интегрального индекса, оценке суммарного эффекта сочетанного действия рекреационных нагрузок и техногенного загрязнения (Мейлах, 2006). Работ, посвященных эколого-биологической характеристике доминатов древесного яруса, не было проведено. Настоящие исследования проведены в период с 2011 по 2013 гг. в пределах территории Экологической тропы «Загадки соснового бора» Челябинского (городского) бора – памятника природы, на стационарных площадках № 1 и № 2, выбор которых основан на степени антропогенного воздействия. Причем площадка № 2 принята нами как эталонная, вследствие минимальной рекреационной нагрузки. Для оценки экологического разнообразия видов древесных синузий, слагающих растительные сообщества Челябинского (городского) бора целесообразно использовать характеристику экологической толерантности (Экологические…, 2010). Анализ индексов толерантности показал, что к гемистенобионтной группе можно отнести клен платанолистный, дуб черешчатый, лиственницу архангельскую, что свидетельствует об ограниченных возможностях распространения. Широкими адаптационными возможностями к климатопу (высокая потенциальная возможность использовать разнообразные местообитания) обладают сосна обыкновенная и береза повислая, относящиеся к гемиэвривалентной группе. Что касается фактора освещенности/затенения, то все виды отнесены к эвривалентной и эврибионтной фракциям, т. е. могут существовать в различных режимах освещенности. Что касается климатопических факторов, то только лиственница архангельская относится к мезобионтной группе, причем проявляет гемистеновалентные позиции по факторам увлажнения и трофности почв. Эти факторы, для данного вида, являются лимитирующими. Все остальные виды по совокупному отношению к эдафотопическим факторам отнесены к гемиэврибионтной группе, а сосна обыкновенная к эврибионтной группе. Можно заключить, что все древесные виды обладают большими адаптационными возможностями к вариациям климатопа и эдафотопа. Фитоиндикационные методы основаны на использовании в качестве фитометров растительных сообществ или ЦП видов. Для оценки параметров среды нами составлены видовые списки растений в рамках стандартных геоботанических описаний с указанием обилия. После обработки в компьютерном комплексе EcoScaleWin с использованием 99 фитоиндикационных шкал были получены совокупные параметры среды. Из представленной таблицы можно констатировать следующее: – изученные площадки отличаются по климатопическим параметрам, несмотря на то, что они находятся в одном климатическом режиме. Особенно это касается континентальности, аридности/гумидности и криоклиматическому фактору. Это объясняется разным видовым составом и соответственно разными требованиями к среде тех или иных видов; – что касается эдафотопических факторов, то отличия прослеживаются по увлажнению почв (от сухо-лесолугового до влажнолесолугового), солевому режиму (от небогатых к довольно богатым почвам). Для осуществления прогнозов на развитие того или иного сообщества используется критерий – популяционная стратегия или популяционное поведение, определяющий способность вида господствовать или занимать подчиненное положение в его структурно-функциональных подразделениях, т. е. ярусах. Остановимся на анализе популяционных стратегий видов древесных растений слагающих площадку № 2. В своей работе мы использовали классификацию типов стратеги Л. Г. Раменского (1938) и исследованные древесные растения распределены следующим образом: Вид Тип стратеги Береза повислая эксплерент Клен платанолистный патиент, патиент-виолент Дуб черешчатый патиент-виолент, виолент-патиент, патиент Лиственница архангельская патиент Сосна обыкновенная патиент, виолент, виолент с чертами эксплерента, эксплерент Анализ таблицы позволил выявить виды с широкими фитоценотическими позициями – это клен, дуб, сосна. Но учитывая характер ЦП дуба черешчатого и клена платанолистного можно предположить смену растительных сообществ в пользу этих видов. Береза не представляет конкуренции для сосны, т. к. этот вид по своей стратегии реактивный, т. е. способен захватывать лишь освобождающиеся территории. Проведение экологической оценки растительных сообществ на исследуемых участках бора по критериям возраста, онтогенетической структуры популяции, бонитировочным таблицам и типам стратегий 100 показало, что восстановительных сукцессий вряд ли стоит ожидать и что данные сообщества очень далеки от климаксовых и находятся на разных этапах дигрессии вследствие внедрения лиственных пород, сорно-рудеральных комплексов травянистых растений, а также отсутствия возобновления. Л и т е р а т ур а Восточноевропейские леса: история в голоцене и современность / отв. ред. О. В. Смирнова. – М.: Наука, 2004. – Кн. 1. – 479 с. Мейлах Э. В. Рекреационная устойчивость островных боров Южного Урала: дис. … канд. биол. наук: 03.00.16 / Э. В. Мейлах; [Место защиты: Урал. гос. ун-т им. А. М. Горького]. – Екатеринбург, 2003. – 129 с. Природа Челябинской области / под ред. М. А. Андреевой. – Челябинск: Изд-во ЧГПУ, 2000. – 269 с. Экологические шкалы и методы анализа экологического разнообразия: монография / Мар. гос. ун-т; Л. А. Жукова, Ю. А. Дорогова, Н. В. Турмухаметова [и др. ]; под общ. ред. Л. А. Жуковой. – Йошкар-Ола, 2010 – 368 с. ТАКСОНОМИЧЕСКОЕ РАЗНООБРАЗИЕ И ЭКОТИПЫ ГИГРОГЕЛИОФИТОВ ОЗЕРА УВИЛЬДЫ Гетманец И. А.1, Байкова И. В.2 1 Челябинский государственный университет, г. Челябинск, igetmanec@mail.ru 2 МБОУ СОШ № 75 (филиал), г. Челябинск, irishenchik@mail.ru Озеро Увильды располагается в предгорной части восточного склона Уральских гор и относится к группе горно-предгорных водоемов. Оно является памятником природы регионального значения. К настоящему времени специальных гидроботанических исследований не было проведено, что и послужило отправной точкой данной работы. Полевые исследования проведены в июне–августе 2013 г. в период максимального развития растительности. При описании растительных сообществ применена стандартная методика экологических профилей (Катанская, 1981; Катанская, Распопов, 1983) и определено видовое разнообразие (Вейсберг, 1999; 2007). Далее приводим аннотированный список растений. ОТДЕЛ MAGNOLIOPHYTA (ANGIOSPERMAE) – ПОКРЫТОСЕМЕННЫЕ Класс Magnoliopsida (Dicotyledones) – Двудольные Подкласс Magnoliidae – Магнолииды Nymphaeaceae Salisb. – Кувшинковые Nupha rlutea (L.) Smith, 1809 – Nympha ealutea L. – Кубышка желтая. В воде на глубине 2–3,5 м. Гидрофит. Водный. Ceratophyllaceae S. F. Gray – Роголистниковые Ceratophyllum demersum L., 1753 – Роголистник погруженный. В воде на глубине 1–3 м. Гидрофит. Водный. 101 Brassicaceae Burnett (Cruciferae Juss.) – Капустные, или Крестоцветные Rorippa amphibia (L.) Bess., 1822 – Sisymbrium amphibium L. var. (β.) aquaticum L. – Nasturtium amphibium (L.) R. Br. – Жерушник земноводный. Берег, пойменный луг, мелководье. Гидрогигрофит. Прибрежноводный. R. palustris (L.) Bess., 1822 – Sisymbrium amphibium L. var. (α.) palustre L. – S. palustre (L.) Leyss. – Nasturtium palustre (L.) DC. – Rorippa islandica auct., non Borb. – Ж. болотный. Влажный луг, берег. Гигрофит. Прибрежно-сорный. Класс Liliopsida (Monocotyledones) – Однодольные Подкласс Alismatidae – Алисматиды Butomaceae Rich. – Сусаковые Butomus umbellatus L., 1753 – Сусак зонтичный. Берег и мелководье, заболоченный луг. Гидрогигрофит. Прибрежный. Hydrocharitaceae Juss. – Водокрасовые Elodea canadensis Michx., 1803 – Элодея канадская. В воде на мелководье. Гидрофит. Водный. Stratiotes aloides L., 1753 – Телорез обыкновенный. В воде на глубине 2–5 м. Гидрофит. Водный. Alismataceae Vent. – Частуховые Alisma plantago-aquatica L., 1753 – A. plantago L. – A. plantago var. latifolium Kunth – A. michaletii Aschers. et Graebn. var. (α.) latifolium Kunth – Частуха подорожниковая. Берег и мелководье, заболоченный луг. Гигрофит. Прибрежный. Sagittaria natans Pall., 1776 – S. alpina Willd. – Стрелолист плавающий. В прибрежной зоне. Гидрогигрофит. Прибрежно-водный. S. sagittifolia L. 1753 – С. обыкновенный. Берег и мелководье. Гигрофит. Прибрежный. Cyperaceae Juss. – Осоковые Carex aquatilis Wahlenb., 1803 – Осока водная. Заболоченный луг, берег. Гигрофит. Прибрежно-болотный. Poaceae Barnhart (Gramineae Juss.) – Мятликовые, или Злаки Alopecurus aequalis Sobol., 1799 – A. fulvus Smith – A. aristulatus Michx. – Лисохвост равный. Берег, сырой луг. Гигрофит. Прибрежно-болотный. 102 Phragmites australis (Cav.) Trin. ex Steud., 1841 – P. communis Trin. – Arundo phragmites L. – A. australis Cav. – Тростник южный (т. обыкновенный). Берег, заболоченный луг. Гигрофит. Прибрежно-болотный. Подкласс Arecidae – Арециды Lemnaceae S. F. Gray – Рясковые Lemna minor L., 1753 – Ряска малая. В воде на тихом мелководье. Гидрофит. Водный. L. trisulca L., 1753 – Р. трехдольная. В воде на тихом мелководье. Гидрофит. Водный. Typhaceae Juss. – Рогозовые Typha angustifolia L., 1753 – Рогоз узколистный. Берег, мелководье. Гидрогигрофит. Прибрежно-болотный. T. latifoliaL., 1753 – Р. широколистный. Берег, мелководье. Гидрогигрофит. Прибрежно-болотный. Таким образом, таксономический состав оз. Увильды представлен 17 видами относящимися к 1 отделу, 2 классам, 10 семействам, 13 родам. Видовое разнообразие распределено неравномерно по профилю водоема, оно зависит от глубины водоема и удаленности от берега. В связи с этим В. Г. Папченков в структуре флоры водоемов выделяет три группы экотипов: − настоящие водные растения – гидрофиты; − прибрежно-водные растения, включающие гелофиты и гигрогелофиты; − заходящие в воду береговые (околоводные) растения, включающие гигрофиты, гигромезо- и мезофиты (Папченков, 2003). Анализ экотопической структуры показал, что значительная часть видов относится к гидрогигрофитам (5 видов, или 29,4 %) и гигрофитам (5 видов, или 29,4 %), собственно гидрофиты представлены 6 видами, или 35,3 %. В данном экотопе не представлены гелофиты. Л и т е р а т ур а Катанская В. М. Высшая водная растительность континентальных водоемов СССР. Методы изучения / В. М. Катанская. – Л.: Наука, 1981. Кокин К. А. Экология высших водных растений / К. А. Кокин. – М.: Изд-во МГУ, 1982. Катанская В. М. Методы изучения высшей водной растительности / В. М. Катанская, И. М. Распопов // Руководство по методам гидробиологического анализа поверхностных вод и донных отложений. – Л.: Гидрометеоиздат, 1983. Папченков В. Г. Продукция макрофитов вод и методы ее изучения / В. Г. Папченков // Гидроботаника: методология, методы: материалы шк. по гидроботанике / ИБВВ им. И. Д. Папанина РАН. – Рыбинск: ОАО Рыбин. дом печати, 2003. 103 ФЛОРА ГОРОДА МЕЛЕУЗА (РЕСПУБЛИКА БАШКОРТОСТАН) Голованов Я. М. Ботанический сад-институт Уфимского научного центра РАН, г. Мелеуз, jaro1986@mail.ru Исследования флоры города Мелеуза, относящегося к южной промышленной зоне Республики Башкортостан, были проведены в течение 2012 г. с помощью широко используемого в урбанофлористике маршрутного метода. Город Мелеуз основан во второй половине XVIII века как торговое село на Оренбургском тракте. В 1958 году рабочий поселок Мелеуз стал городом районного подчинения. Численность населения в г. Мелеузе равняется 61,4 тыс. чел. Площадь города – 35,5 км2. Согласно природному районированию Республики Башкортостан город расположен в Предуральском степном районе, по преимуществу на левобережье р. Белой. По результатам исследований, флора г. Мелеуза насчитывает 640 видов, 351 род и 80 семейств высших растений. При этом отмечено подавляющее господство двудольных растений класса Magnoliopsida. На долю сосудистых споровых приходится лишь 3 вида. Наиболее крупными по числу видов семействами на территории г. Мелеуза являются: Asteraceae (99 видов, 15,5 % от общего числа видов), Роасеае (63 вида, 9,8 %), Fabaceae (40 видов, 6,3 %), Brassicaceae (37 видов, 5,8 %), Rosaceae (31 вид, 4,8 %). Ранжировка десяти важнейших семейств флоры, представляющих собой «головную часть флористического спектра» (Малышев, 1973), приведена в таблице. В целом спектр ведущих семейств флоры г. Мелеуза согласуется со спектром флоры Голарктики. Высокое положение семейств Brassicaceae, Fabaceae, Lamiaceae, Apiaceae отражает общий процесс ослабления в урбанофлоре ее зональных черт, в городах умеренной зоны проявляющийся в смещении основных параметров в термоксерическом направлении и в сторону флор, сформировавшихся в экстремальных условиях. Флора г. Мелеуза также характеризуется высоким процентом видов десяти ведущих семейств, которые в изученной флоре занимают 60,3 % от общего видового состава. Число видов в семействах варьирует от 1 до 99. Ведущими родами во флоре г. Мелеуза являются: Carex (15 видов), Artemisia (11 видов), Galium, Veronica, Potamogeton (по 10 видов), Poa (9 видов), Rumex, Chenopodium, Viola (по 8 видов). 104 Спектр ведущих семейств флоры г. Мелеуза Семейство Число видов г. Мелеуз Процент от общего числа видов Asteraceae 99 15,5 Poaceae 63 9,8 Fabaceae 40 6,3 Brassicaceae 37 5,8 Rosaceae 31 4,8 Caryophyllaceae 25 3,9 Lamiaceae 23 3,6 Apiaceae 24 3,8 Cyperaceae 23 3,6 Scrophulariaceae 21 3,3 Всего 386 60,3 Важной характеристикой флоры является биологический спектр ее жизненных форм. Флора города характеризуется как гемикриптофитнотерофитная. Преобладающей биоморфой во флоре города являются гемикриптофиты (352 вида, 50,3 % от общего видового состава). Повышение доли гемикриптофитов характерно для флор умеренной зоны Евразии. Вторую позицию занимают терофиты (120 видов, 18,8 %), широко представленные на различных нарушенных городских местообитаниях. Сокращение в городских условиях влажных местообитаний объясняет снижение в спектре доли гидрофитов (22 вида, 2,4 % от общего видового состава), гелофитов (11 видов, 1,7 %), гелофитовгемикриптофитов (11 видов, 1,7 %), гелофитов-гидрофитов (2 вида, 0,3 %). Снижение доли хамефитов (20 видов, 3,1 %) связано с их повышенной механической уязвимостью представителей этой жизненной формы (Березуцкий, Кашин, 2008). Низкую адаптационную активность в урбанизированной среде показывают и криптофиты (38 видов, 5,8 %). Отмечено повышение роли фанерофитов (54 вида, 8,4 %), связанное как с хорошей адаптацией древесных видов к антропогенным местообитаниям, так и с дичанием древесных интродуцентов из культурных насаждений (Березуцкий, 1999). Согласно классификации жизненных форм флоры г. Мелеуза, по И. Г. Серебрякову, больше половины видов флор составляют поликарпические растения (382 вида, 59,7 % от общего видового состава), среди которых преобладают стержнекорневые растения (107 видов, 16,7 %). Основную часть поликарпических трав составляют аборигенные виды растений. Вторую по величине группу составляют монокарпические 105 растения (190 видов, 29,7 %), среди которых преобладают однолетники (120 видов, 18,8 %) и двулетники (37 видов, 5,8 %), большую часть которых составляют адвентивные растения. Согласно структуре флоры г. Мелеуза, по составу долготных групп видов ведущие роли играют евразиатская (404 вида, 63,1 %, от общего видового состава) и мультиконтинентальная группа элементов флоры (118 видов, 18,4 %). Третье место занимают виды с широким культигенным ареалом (76 видов, 7,9 %), высокое положение данной группы характерно для флор городов, где происходят активные процессы дичания культивируемых видов. Согласно структуре флоры г. Мелеуза, по составу широтных групп видов ведущее положение во флорах городов занимают плюризональные виды (223 вида, 34,8 % от общего числа видов), значительная часть которых – сорные виды с широким распространением. Второе место в рассматриваемом спектре занимают виды, характерные для лесостепной и степной зон (135 видов, 21,1 %). Помимо вышеназванных групп видов, также значимы и виды с широким культигенным ареалом (76 видов, 11,9 %). Ведущее положение во флоре г. Мелеуза играют виды мезофитной ориентации (мезофиты, гигромезофиты, ксеромезофиты) (447 видов, 69,8 % от общего видового состава), второе место занимают виды ксерофитной ориентации (ксерофиты, мезоксерофиты) (81 вид, 12,6 %). На долю видов переувлажненных местообитаний (гидрофиты, гидрогигрофиты, гигрофиты, мезогигрофиты) приходится 112 видов, 17,5 %. Согласно фитосоциологическому анализу ведущую роль во флоре г. Мелеуза играют виды классов Festuco-Brometea (120 видов, 18,9 % от общего видового состава) и Molinio-Arrhenatheretea (88 видов, 13,7 %). Также высока доля видов синантропных классов Stellarieteamediae (71 вид, 11,1 %) и Artemisieteavulgaris (36 видов, 5,6 %). В связи с большим наличием прибрежно-водных местообитаний высока доля видов классов Phragmito-Magnocaricetea (51 вид, 8,0 %) и Potametea (18 видов, 2,8 %). Во флоре г. Мелеуза отмечено 173 адвентивных видов растений, из них 39 археофитов и 134 неофита. Основную роль в формировании адвентивной фракции флоры играет непреднамеренный занос (95 видов, 54,9 % от общего видового состава адвентивной фракции флор). Доля преднамеренно занесенных и ушедших из культуры растений также велика (78 видов, 45,1 %). Среди групп видов по степени натурализации ведущую позицию занимают эпекофиты (86 вида, 53,2 %). Меньшую долю имеют эфемерофиты (30 видов, 17,3 %). Среди эфемерофитов преобладает группа преднамеренно занесенных растений. Также высока доля колонофитов (31 вид, 22,5 %). Группа агриофитов насчитывает 106 11 видов, что составляет 6,3 от общего видового состава адвентивной фракции флор. Л и т е р а т ур а Березуцкий М. А. Антропогенная трансформация флоры / М. А. Березуцкий // Ботан. журн. – 1999. – Т. 84, № 6. – С. 8–19. Березуцкий М. А. Антропогенная трансформация флоры и растительности / М. А. Березуцкий, А. С. Кашин. – Саратов: Изд. центр «Наука», 2008. – 100 с. Малышев Л. И. Флористическое районирование на основе количественных признаков / Л. И. Малышев // Ботан. журн. – 1973. – Т. 58, № 11. – С. 1581–1588. К ОЦЕНКЕ ФИТОЦЕНОТИЧЕСКОЙ ПРИУРОЧЕННОСТИ РЕДКОГО ВИДА IRIS PUMILA L. В БАШКИРСКОМ ПРЕДУРАЛЬЕ Голованов Я. М., Крюкова А. В., Абрамова Л. М. Ботанический сад-институт УНЦ РАН, г. Уфа, jaro1986@mail.ru, abramova.lm@mail.ru В деле охраны биоразнообразия редких и исчезающих растений важную роль играет изучение сообществ с их участием, поскольку сохранение конкретных популяций невозможно, если не охраняются ценозы, в которых они произрастают. Кроме того, сами сообщества с редкими видами зачастую являются объектами охраны на ООПТ различного уровня, т. к. представляют редкие типы растительности. В этой связи в ходе проведения популяционных исследований в Башкирском Предуралье, нами проводилась также оценка фитоценотической приуроченности редкого вида флоры Республики Башкортостан Iris pumila L. Ирис карликовый включен в Красную книгу РБ со статусом редкий вид (III категория) (Красная книга…, 2011). Ксерофит. Произрастает в каменистых и луговых степях, на остепненных лугах; по щебнистым и каменистым склонам степных холмов, на обнажениях, предпочитает участки с разреженной растительностью. На выгонах сохраняется в зарослях кустарников (можжевельника казацкого и др.). Основная часть популяций сохранилась в предгорьях Южного Урала в южной лесостепной зоне РБ. В рамках фитоценотических исследований было выполнено 15 геоботанических описаний растительных сообществ с участием Iris pumila на пробных площадках площадью 64–100 м² в зависимости от границ фитоценоза. Эколого-флористическая классификация проведена методом классического синтаксономического анализа (Braun-Blanquet, 1964). Геоботанические описания были введены в базу данных TURBOVEG (Hennekens, 1995). Для принятия синтаксономических решений, полученных в результате обработки, синтаксоны сравнивались 107 с единицами, ранее описанными в РБ и других регионах (Ямалов и др., 2012). В таблице приведена краткая характеристика изученных ценопопуляций Iris pumila в Башкирском Предуралье, название ценопопуляций дано по близлежащим населенным пунктам. Наиболее часто ирис карликовый произрастает в богатовидовых красивейшековыльных степях ассоциации Astragalo austriacae-Stipetum pulcherrimae, являющихся зональным типом степей Предуралья Республики Башкортостан. Описанные площадки занимали при этом, как правило, степные склоны различной экспозиции. Флористический состав сообществ насчитывает от 38 до 50 видов на площадке, в среднем 43 вида. Общее проективное покрытие варьирует от 65 до 90 % на площади описания 64–100 м2. В составе ценозов велика доля петрофитностепных видов. Ирис карликовый был отмечен на всех площадках с обилием 1. Подобные сообщества были описаны в Зианчуринском и Федоровском районах РБ. Краткая характеристика сообществ с участием Iris pumila Местообита- ОПП травония стоя, % Ценопопуляции Характерные виды Ассоциация Astragalo austriacae-Stipetum pulcherrimae г. Высокая, ур. КучТопкан, Юлдашево, Тазларово настоящая степь 65–90 Artemisia campestris, Echinops ruthenicus, Galatella villosa, Koeleria sclerophylla, Stipa pulcherrima Сообщество Stipa capillata [Helictotricho-Stipetalia] г. Караултау, петрофитная Якшимбетово, Кужастепь нак, Холодный Ключ 70–85 Agropyron pectinatum, Astragalus macropus, A. testiculatus, Festuca pseudovina, Potentilla arenaria, Stipa capillata Ассоциация Scorzonerо austriacaе-Stipetum lessingianae Кужанак, Арсенево, Лена настоящая степь 75 Centaurea carbonata, Galatella villosa, Echinops ruthenicus, Stipa capillata, S. lessingiana Ассоциация Spiraeo hypericifoliaе-Amygdaletum nanae Верх. Муйнак кустарниковая степь 50–65 Elytrigia lolioides, Festuca pseudovina, Spiraea hypericifolia, Stipa capillata Помимо сообществ вышеназванной ассоциации часто ирис карликовый нередко отмечался и в сообществах с участием ковыля волосатика или тырсы – сообщество Stipa capillata [Helictotricho-Stipetalia]. Это 108 были, как правило, выпасаемые участки настоящих и петрофитных степей на плакорных и склоновых местообитаниях. Описанные площадки занимали при этом, как правило, степные склоны южной экспозиции. Флористический состав данных сообществ насчитывает от 29 до 45 видов на площадке, в среднем 37 видов. Общее проективное покрытие варьирует от 70 до 85 % на площади описания 64 м2. В составе ценозов также велика доля петрофитно-степных видов. Ирис карликовый был отмечен на всех площадках с обилием ± 2. Такие сообщества были описаны в Куюргазинском, Мелеузовском и Зианчуринском районах РБ. Относительно редко изучаемый вид отмечался в сообществах ассоциации Scorzonerо austriacaе-Stipetum lessingianae, представляющей собой лессингоковыльные степи южных районов Башкирии. Флористический состав ценозов насчитывает от 35 до 40 видов на площадке, в среднем 38 видов. Общее проективное покрытие равняется 75 % на площади описания 64–100 м2. Для ценофлоры описанных сообществ также характерна высокая роль петрофитных видов союза HelictotrichoStipion порядка Helictotricho-Stipetalia. Ирис карликовый был отмечен на всех площадках с обилием ± 1. Наиболее редким типом сообществ с участием ириса карликового являются сообщества ассоциации Spiraeo hypericifoliaе-Amygdaletum nanae. Так, в популяции, расположенной по восточному склону горы у д. Верх. Муйнак, ирис карликовый отмечался среди разреженных зарослей спиреи зверобоелистной на верхней границе склона, по известняковым осыпям. Флористический состав сообществ насчитывает 25–26 видов на площадке, в среднем 25 видов. Общее проективное покрытие варьирует от 50 до 65 % на площади описания 100 м2. Ирис карликовый был отмечен на всех площадках с обилием ± 1. Таким образом, в ходе проведенных исследований выявлено, что сообщества с участием Iris pumila относятся к классу Festuco-Brometea Br. -Bl. et R. Tx. in Br. Bl. 1949, объединяющему сообщества степной растительности и, в частности, к порядку настоящих степей – Helictotricho-Stipetalia Toman 1969. Исследованные ценозы с участием изучаемого вида относятся к трем ассоциациям: Astragalo austriacaeStipetum pulcherrimae Yamalov 2012., Scorzonerо austriacaе-Stipetum lessingianae Yamalov 2012., Spiraeo hypericifoliaе-Amygdaletum nanae Solomeshch et al. 1994 и сообществу Stipa capillata [HelictotrichoStipetalia]. Л и т е р а т ур а Красная книга Республики Башкортостан: в 2 т. Т. 1: Растения и грибы / под ред. д-ра биол. наук, проф. Б. М. Миркина. – 2-е изд., доп. и перераб. – Уфа: МедиаПринт, 2011. – 384 с. Ямалов С. М. Продромус растительных сообществ Республики Башкортостан / С. М. Ямалов, В. Б. Мартыненко, Л. М. Абрамова, В. Б. Голуб, Э. З. Баишева, 109 А. В. Баянов. – Уфа: Гилем, 2012. – 83 с. Braun-Blanquet J. Pflanzensociologie. Grundzuge der Vegetationskunde. 3 Aufl. / J. Braun-Blanquet. – Wien-New-York, 1964. – 865 S. Hennekens S. M. TURBO (VEG). Software package for input processing and presentation of plantsociological data. User’s guide / S. M. Hennekens // IBN-DLO Wageningen et University of Lancaster. – 70 p. РАЗНООБРАЗИЕ И ЧИСЛЕННОСТЬ БАКТЕРИЙ ЦИКЛА АЗОТА В ПОЧВАХ ТЕХНОГЕННО НАРУШЕННЫХ ТЕРРИТОРИЙ Гордеева В. А.1, Жуйкова Т. В.1, Ившина И. Б.2, Костина Л. В.2, Безель В. С.3 1 ФГБОУ ВПО Нижнетагильская государственная социально-педагогическая академия, г. Нижний Тагил 2 ФГБОУ ВПО Пермский государственный национальный исследовательский университет, г. Пермь 3 Институт экологии растений и животных УрО РАН, г. Екатеринбург Азот является одним из важнейших биофильных элементов, во многом определяющий характер и направление микробиологических процессов в различных экосистемах. В настоящее время на функционирование почвенной микробиоты значительное влияние оказывает поступление в почву химических веществ техногенной природы, в частности, тяжелых металлов. Следствием этого, как правило, выступает снижение общей численности микроорганизмов (Тяжелые металлы…, 1989), в том числе аммонификаторов и нитрификаторов. Однако, в микробном сообществе загрязненной почвы также могут появиться нехарактерные для нормальных условий, устойчивые к тяжелым металлам виды микроорганизмов (Кобзев, 1980), либо возможно увеличение численности отдельных устойчивых к токсикантам групп бактерий. Последнее было показано для денитрификаторов, количество которых в загрязненной тяжелыми металлами почве возрастало (Наплекова, 1982; Сорокин, 1990). Следовательно, толерантность микроорганизмов к загрязнению почвы зависит от их принадлежности к различным систематическим группам. Эти структурные изменения микробиологического комплекса приводят к трансформации одного из наиболее важных биогеохимических циклов – круговорота азота. В связи с этим, целью данного исследования было изучение почвенной микробиоты, участвующей в превращениях цикла азота в условиях химического загрязнения среды. Исследования проведены в одном из промышленных центров – г. Нижний Тагил (Свердловская область, 58°42′ с. ш. и 61°20′ в. д.). Район исследования относится к таежной географической зоне, подзона южной тайги. Промышленный комплекс района исследования пред- 110 ставлен предприятиями черной и цветной металлургии, горнодобывающей и химической промышленности, машиностроения и рядом других. Исследование почвенного микробиологического комплекса проведено на агроземах и техноземах в разной степени загрязненных тяжелыми металлами. Максимальное содержание подвижных форм тяжелых металлов в почвах в мкг/г следующее: Cu – 569, Pb – 227, Cd – 3, Zn – 5640, Fe – 50654, что от 16 (для железа) до 493 (для цинка) раз выше минимальных региональных фоновых значений. По остальным элементам превышение фоновых показателей варьирует в пределах 64–84 раза. В соответствие с интегральным уровнем загрязнения участки были отнесены к фоновой (Si = 1,00 отн. ед.), буферной (Si = 3,33–6,19 отн. ед.) и импактной (Si = 22,78–30,00 отн. ед.) зонам загрязнения. Названия зон даны в соответствие с номенклатурой ЮНЕП (Global…, 1973). Отбор и подготовка почвенных образцов для микробиологического анализа проведены в вегетационный сезон 2012 г. в соответствии с методическими рекомендациями (Инструментальные методы…, 1982). Для выявления и учета представителей бактерий цикла азота использовали чашечный метод и метод предельных разведений. При этом подсчет наиболее вероятного числа микроорганизмов определяли по статистической таблице Мак-Креди. Посев почвенных образцов проводили на элективные культуральные среды (Парактикум…, 2002): аммонификаторы – в мясопептонный бульон; денитрификаторы – среду Гильтея; нитрификаторы I и II фазы – среду Виноградского. Численность свободноживущих аэробных азотфиксаторов рода Azotobacter учитывали методом комочков обрастания на среде Эшби. Культивирование микроорганизмов проводили при 28–30 оС в течение 7–21 суток. Все определения проводили в 3–5-кратной повторности. Для более эффективной десорбции микроорганизмов с поверхности почвенных частиц осуществляли предварительную подготовку образцов почвы, включающую ультразвуковую обработку почвенной суспензии (1 : 10) в течение 2– 5 мин с помощью низкочастотного диспергатора Soniprep 150 (MSE) (Sanyo, Япония). Микробиологические исследования были проведены на базе лаборатории алканотрофных микроорганизмов Института экологии и генетики микроорганизмов УрО РАН (г. Пермь). Статистический анализ выполнен на основе методов описательной статистики в ПСП Statistica-6.0 (StatSoft, Inc. 1984–2001). Множественные сравнения проведены S-методом Шеффе. Эколого-трофические группы почвенного микробного сообщества техногенно нарушенных территорий характеризовали по показателям 111 численности бактерий физиологических групп, участвующих в циклах азота (табл.). В ходе микробиологических исследований почвы в 2012 году были выделены аммонифицирующие, нитрифицирующие (II фаза), денитрифицирующие, азотфиксирующие бактерии. Отметим, что нитрифицирующие бактерии первой фазы в 2012 г. были не выделены в почве исследуемых территорий, хотя в предыдущий год их численность варьировала от 4,0–6,5 × 104 на участках буферной зоны до 2,5 × 107 на фоновой и максимально загрязненных территориях. Численность бактерий цикла азота в исследуемых биотопах (M ± m) Физиологические группы бактерий Токсическая нагрузка, отн. ед. 1,00 3,33 6,19 22,78 30,0 1,0·108 0,2·108 0,9·108 0,2·109 0,2·109 Нитрифицирующие (I фаза), клеток / г почвы 0,0 0,0 0,0 0,0 0,0 Нитрифицирующие (II фаза), клеток / г почвы 8,0·105 3,8·106 1,40·106 2,0·106 1,8·106 Денитрифицирующие, клеток / г почвы 0,2·103 0,9·105 0,13·104 4,4·107 2,3·107 Азотфиксирующие (анаэробы), % 23,56 ± 19,48 0,67 ± 0,58 9,78 ± 5,58 97,78 ± 1,87 100,0 ± 0,00 Аммонифицирующие, клеток / г почвы Из таблицы видно, что с увеличением химического загрязнения почвы тяжелыми металлами общая численность микроорганизмов всех исследуемых групп, участвующих в цикле азота, возрастает. Наиболее представленной группой на всех участках являются аммонификаторы. Численность данной группы имеет тенденцию к росту с увеличением общего уровня загрязнения почвы (табл.). Однако статистически значимого влияния уровня загрязнения на данную группу не установлено (S-методом Шеффе (1,00–6,19 и 22,78–30,00 отн. ед.): F = 1,38; df = 4; 11; p > 0,05). Второе место по численности занимают нитрифицирующие (105–106) и денитрифицирующие бактерии (103–107). Методом множественных сравнений Шеффе подтверждено статистически значимое различие между численностью нитрифицирующих и денитрофицирующих бактерий в почве фоновой и буферной зон и участков импактной зоны (нитрифицирующие бактерии: F = 11,58; df = 4; 11; p < 0,01; денитрофицирующие бактерии: F = 3,44; df = 4; 11; p < 0,001). Высокая численность бактерий данных групп на максимально загрязненных тяжелыми металлами участках свидетельствует о их высокой устойчивости к ток112 сикантам. Полученные результаты согласуются с данными Д. Г. Звягинцева с соавт. (1997), которые изучали микробиоту на дерновоподзолистых почвах обогащенных свинцом. Высокую устойчивость к загрязнению также проявляют азотфиксирующие бактерии (табл.). Доля комочков обрастания на среде Эшби, по которой судили о численности свободноживущих аэробных азотфиксаторов рода Azotobacter, на фоновом участке – 24 % (табл.). В почве буферных зон доля бактерий данной группы уменьшается в 2,4–35 раз. Дальнейший рост загрязнения приводит к увеличению процента комочков обрастания до 98–100 %, что статистически значимо отличается от остальных территорий (S-метод: F = 10,76; df = 4; 45; p < 0,01). Таким образом, азотфиксирующие бактерии рода Azotobacter проявляют повышенную чувствительность к средним уровням загрязнения, что согласуется с данными В. Ф. Валькова с соавт. (1997). Однако, высокие дозы тяжелых металлов оказывают стимулирующее действие на процессы азотфиксации. Аналогичные результаты были получены в модельных экспериментах с загрязненной тяжелыми металлами почвой другими авторами (Умарова, Азиева, 1980; Звягинцев и др., 1997). Таким образом, в условиях повышенного загрязнения почвы тяжелыми металлами наблюдается увеличение общей численности микроорганизмов участвующих в цикле азота. Отмеченное увеличение численности почвенных микроорганизмов и плотности микробных популяций, может быть связано с повышенным содержанием гумуса (от 3,63 до 4,12 %) и уровня рН почв исследуемых территорий. Ранее нами было установлено, что в градиенте увеличивающегося загрязнения почвы тяжелыми металлами кислотность почвенного раствора изменяется от 6,32 до 8,57 единиц, содержание гумуса возрастает от 3,6 до 4 % (Кайгородова и др., 2012). Согласно исследованиям Г. В. Ковалевой с соавт. (2007), увеличение численности микроорганизмов в антропогенно нарушенных биотопах связано именно с подщелачиванием почвы. Кроме того, с точки зрения К. Е. Гиллера с соавт. (Giller et al., 1998), длительное техногенное воздействие полиметаллического загрязнения на почвенные комплексы способствует формированию механизмов специфической устойчивости и адаптации микробного сообщества к токсическому влиянию тяжелых металлов. Работа выполнена при финансовой поддержке Правительства Свердловской обл. и РФФИ (проект № 13-04-96056 р_урал_а); Программы Президиума УрО РАН (проект 12-И-4-2051) Л и т е р а т ур а Вальков В. Ф. Влияние загрязнения тяжелыми металлами на микроскопические грибы и Azotobacter чернозема обыкновенного / В. Ф. Вальков, С. И. Колесников, К. Ш. Казеев, 113 С. С. Тащиев // Экология. – 1997. – № 5. – С. 388−390. Звягинцев Д. Г. Микробиологические и биохимические показатели загрязнения свинцом дерново-подзолистой почвы / Д. Г. Звягинцев, А. В. Кураков, М. М. Умаров, З. Филипп // Биология почв. – 1997. – № 9. – С. 1124−1131. Инструментальные методы в почвенной микробиологии / под общ. ред. Е. А. Андреюк. – Киев: Наук. думка, 1982. – 176 с. Кайгородова С. Ю. Характеристика антропогенно деградированных почв и микробного комплекса территорий Нижнетагильского промузла / С. Ю. Кайгородова, Т. В. Жуйкова, В. А. Жуйкова, В. С. Безель, И. Б. Ившина // Современные проблемы загрязнения почв: материалы IV Международной научной конференции. 27–30 мая 2013 г. Москва. – М., 2013. – С. 226–321. Кобзев В. А. Взаимодействие загрязняющих почву тяжелых металлов и почвенных микроорганизмов (обзор) // Загрязнение атмосферы, почвы и растительного покрова. – 1980. – № 10(86). – С. 51–68. Ковалева Г. В. Структура бактериальных сообществ в естественных и антропогенно нарушенных бурых лесных почвах ботанического сада п-ва Муравьева-Амурского / Г. В. Ковалева, Т. Г. Добровольская, А. В. Головченко // Почвоведение. – 2007. – № 5. – С. 1−6. Наплекова Н. Н. Влияние солей некоторых тяжелых металлов на физиологическую активность целлюлозоразрушающих микроорганизмов / Н. Н. Наплекова // Изв. Сиб. отд. АН СССР. Сер., Биол. наук. – 1982. – Вып. 2, № 10. – С. 79–85. Практикум по биологии почв: учеб. пособие / Г. М. Зенова, А. Л. Степанов, А. А. Лихачева, Н. А. Манучарова. – М.: Изд-во МГУ, 2002. – 120 с. Сорокин Д. Ю. Окисление соединений азота гетеротрофными организмами / Д. Ю. Сорокин // Успехи микробиологии. – М.: Наука, 1990. – Т. 24. – С. 100–127. Тяжелые металлы как фактор антропогенного воздействия на почвенную микробиоту / С. В. Левин и др. // Микроорганизмы и охрана почв. – М., 1989. – С. 5–46. Умарова М. М. Некоторые биохимические показатели загрязнения почв тяжелыми металлами / М. М. Умарова, Е. Е. Азиева // Тяжелые металлы в окружающей среде. – М., 1980. – № 9. – С. 16−20. Giller K. E. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: a review / K. E. Giller, E. Witter, S. P. McGrath // Soil Biol. Biochem. – 1998. – Vol. 30. – P. 1389−1414. Global environmental monitoring system (GEMS) SCOPE Rep. 3. – Canada, 1973. БИОМАССА ПРОСТРАНСТВЕННЫХ МИКРОГРУППИРОВОК ВИДОВ РАСТЕНИЙ НАПОЧВЕННОГО ПОКРОВА В ТЕМНОХВОЙНЫХ ЛЕСАХ КОСТРОМСКОЙ ОБЛАСТИ Грозовская И. С.1, Ханина Л. Г.2, Бобровский М. В.1, Смирнов В. Э.2, Романов М. С.2 1 Институт физико-химических и биологических проблем почвоведения Российской академии наук, г. Пущино 2 Институт математических проблем биологии Российской академии наук, г. Пущино, irinagrozovskaya@gmail.com Напочвенный покров (травяно-кустарничковый и мохово-лишайниковый ярусы) является важным компонентом лесных экосистем. Известно, что основной ролью напочвенного покрова в функционировании лесных экосистем являются образование и отчуждение органического вещества (Ремезова, 1964; Тужилкина, 2012). В этой связи для моделирования динамики напочвенного покрова необходимо выделить группы 114 видов растений, сходных по биомассе, химическому составу видов и др. (Комаров и др., 2007). Для обобщения существующей в литературе информации по характеристикам биомассы и продуктивности лесного напочвенного покрова нами была разработана база данных «Биомасса». База данных включает в себя информацию по биомассе видов и групп видов недревесных сосудистых растений, мохообразных и лишайников в лесных сообществах (Ханина и др., 2013). Анализ базы данных показал, что недостаточно информации по биомассе старовозрастных высокотравных и крупнопапоротниковых темнохвойных лесов. Поэтому целью нашего исследования является сбор дополнительного материала по биомассе напочвенного покрова данных типов леса. В качестве объектов исследования выбраны еловые и пихто-еловые леса, расположенные на северо-востоке Костромской области. Сбор данных проводили в 2011–2012 гг. в шести типах леса: пихто-ельнике бореально-мелкотравно-чернично-крупнопапоротниковом, пихто-ельнике разнотравном, ельнике черничном, липо-ельнике крупнопапоротниковом, ельнике бореально-мелкотравном и ельнике высокотравном (Грозовская, 2012). Для сбора полевого материала по биомассе напочвенного покрова в каждом из типов леса заложены учетные площади размером 40 × 40 м2. В напочвенном покрове выделяли микрогруппировки видов по сходству видового состава и доминирующей экологоценотической группе (ЭЦГ) видов. В микрогруппировках закладывали площадки 25 × 25 см2, для которых составляли список видов сосудистых растений. На площадках срезали надземную часть растений с разделением на виды. Методом монолитов на тех же площадках отбирали подземные части растений с разделением их по видовой принадлежности (Андреева, 2002). В напочвенном покрове исследованных типов леса выделены пять основных типов микрогруппировок (табл.). Среднее число видов сосудистых растений, попадавших в пределы закладываемых площадок 25 × 25 см2, в разных типах микрогруппировок варьировало от 4 до 6. В образовании разных типов микрогруппировок участвует определенный набор видов сосудистых растений, из которых 1–3 вида являются доминантными и/или имеют высокий балл константности. Для микрогруппировок мы составили список видов сосудистых растений, являющихся доминантными (3–5 балл обилия по шкале Браун-Бланке) и имеющих высокий балл константности 4–5 (4 – 60–80 %, 5 – 80–100 % площадок, на которых был встречен вид) (Смирнова и др., 2006) (табл.). Oxalis acetosella L. встречается с высокой константностью во всех микрогруппировках, кроме высокотравной, но является доминан115 том только для Br микрогруппировки, где имеет балл обилия 4–5. В Vm, Lf и TH микрогруппировках доминантные виды образуют более 90 % всей надземной биомассы микрогруппировки. В Br микрогруппировке доля биомассы доминантных видов составляет более 70 %. Только в BN микрогруппировке доля биомассы доминантных видов составила меньше половины от всей надземной биомассы (44 %) (рис. 1). Доминантные и высококонстантные виды растений в микрогруппировках Тип микрогруппировки Бореальная (Br) Виды-доминанты Oxalis acetosella, Maianthemum bifolium (L.) F. W. Schmidt, Gymnocarpium dryopteris (L.) Newman, Phegopteris connectilis (Michx.) Watt Бореально-неморальная (BN) Stellaria holostea L., Oxalis acetosella Черничная (Vm) Vaccinium myrtillus L., Oxalis acetosella Крупнопапоротниковая (Lf) Dryopteris dilatata (Hoffm.) A. Gray, Oxalis acetosella Высокотравная (TH) Aconitum septentrionale Koelle, Athyrium filix-femina (L.) Roth, Cirsium oleraceum (L.) Scop., Diplazium sibiricum (Turcz. ex Kunze) Sa. Kurata, Filipendula ulmaria (L.) Maxim. Примечание. В таблице представлены виды растений, имеющие в микрогруппировках балл обилия 5. Рисунок 1 – Долевое участие доминантных видов сосудистых растений в образовании биомассы выделенных типов микрогруппировок 116 Для микрогруппировок была рассчитана суммарная надземная, подземная биомасса сосудистых растений. Надземная биомасса растений возрастает в ряду Br – BN – Vm – Lf – TH, подземная биомасса – Br – BN – Vm – TH – Lf (рис. 2). Самые низкие средние значения по надземной и подземной биомассе имеет Br и BN микрогруппировки, промежуточные – Vm, самые высокие – Lf и TH микрогруппировки. В целом, по нашим данным, надземная биомасса Lf и TH микрогруппировок сходна между собой, в то время как по подземной биомассе Dryopteris dilatata, абсолютно доминирующий в Lf микрогруппировке, явно лидирует. Полученные экспериментальные результаты хорошо согласуются с литературными данными, где наблюдается увеличение значений медиан по надземной и подземной биомассам сосудистых растений в ряду Br – BN – Vm – TH – Lf (рис. 2). Рисунок 2 – Надземная и подземная биомасса сосудистых растений по авторским и литературным данным Белые прямоугольники построены по экспериментальным данным; серые – по литературным данным. Числа под осью означают величину выборки. Типы микрогруппировок (см. табл.) 117 Количественный анализ подтвердил наличие существенных различий между пространственными микрогруппировками растений, выделенными в напочвенном покрове по доминирующей ЭЦГ видов. Наиболее хорошо эти различия отмечаются по значениям надземной и подземной биомассы сосудистых растений (рис. 2). Выявленные различия по биомассе подтверждают обоснованность высказанного ранее предположения (Комаров и др., 2007; Ханина и др., 2013) о перспективности использования пространственных микрогруппировок в качестве элементарной единицы в задаче генерализации напочвенного покрова при включении его в круговоротные модели лесных экосистем. Л и т е р а т ур а Андреева Е. Н. Методы изучения лесных сообществ / Е. Н. Андреева, И. Ю. Баккал, В. В. Горшков и др. – СПб.: НИИХимии СПбГУ, 2002. – 240 с. Грозовская И. С. Биопродукционные характеристики живого напочвенного покрова старовозрастных пихтоельников северо-востока Костромской области / И. С. Грозовская // Известия Самарского научного центра Российской академии наук. – 2012. – Т. 14, № 1(6). – С. 1445–1448. Комаров А. С. Моделирование динамики органического вещества в лесных экосистемах / А. С. Комаров, О. Г. Чертов, А. В. Михайлов и др.; отв. ред. В. Н. Кудеяров. – М.: Наука, 2007. – 380 с. Ремезова Г. Л. Накопление органического вещества в процессе развития травяного покрова дубравы / Г. Л. Ремезова // Ботан. журн. – 1964. – Т. 49, № 6. – С. 894– 900. Смирнова О. В. Биоразнообразие и сукцессионный статус старовозрастных темнохвойных лесов / О. В. Смирнова, М. В. Бобровский, Л. Г. Ханина, В. Э. Смирнов // Успехи современной биологии. – 2006. – Т. 126, № 1. – С. 27–49. Тужилкина В. В. Структура фитомассы и запасы углерода в растениях напочвенного покрова лесов на северо-востоке европейской России / В. В. Тужилкина // Растительные ресурсы. – 2012. – Т. 48, № 1. – С. 44–50. Ханина Л. Г. Анализ базы по биомассе лесного напочвенного покрова для моделирования его динамики в круговоротных моделях лесных экосистем / Л. Г. Ханина, И. С. Грозовская, В. Э. Смирнов, М. С. Романов, М. В. Бобровский // Хвойные бореальные зоны. – 2013. – Т. 31, № 1–2. – С. 22–29. ФАУНА ПОЗВОНОЧНЫХ ЖИВОТНЫХ ПАМЯТНИКОВ ПРИРОДЫ РЕСПУБЛИКИ МАРИЙ ЭЛ «БОЛОТО ЖЕЛЕЗНОЕ» И «БОЛОТО БЕРЕЗОВОЕ» Дробот В. И. Марийский государственный университет, г. Йошкар-Ола, drobot58@gmail.com Объектами наших исследований были орнитоценозы особо охраняемых природных территорий (ООПТ) «Болото Железное» и «Болото Березовое». ООПТ расположены в Волжском районе Республики Марий Эл, находятся на расстоянии 900 м друг от друга. Объекты имеют статус памятников природы регионального (республиканского) значения с 1987 года. Причиной создания данных ООПТ явилось наличие в их границах большого разнообразия гидрофильных птиц, в том числе ред118 ких и охраняемых. Несмотря на официально установленный природоохранный статус, мониторинг состояния экосистем указанных ООПТ на протяжении всего срока их существования не проводился. Имевшие место единичные исследования носили случайный характер и осуществлялись в инициативном порядке. Последние исследования предметного характера датируются 70–80-ми годами ХХ века. Достоверная оперативная информация о состоянии популяций охраняемой на ООПТ фауны отсутствует. В то же время во многих регионах не только Российской Федерации, но и мира отмечается тенденция негативных изменений в структурах ценозов гидрофильных групп птиц, особенно колониальных, включая «краснокнижные» виды. В связи с необходимостью оценить экологическую ситуацию на охраняемых природных объектах и подтвердить их природоохранный статус, целью наших исследований было изучение современного состояния популяций позвоночных животных на ООПТ. В данной работе отражены результаты изучения орнитофауны природоохранных территорий: видовой состав, численность, биотопическое распределение птиц, в том числе редкой фауны. Материалом для работы послужили результаты учетов, проведенных в полевой сезон 2013 года. На протяжении июня–сентября на каждой ООПТ было проведено по 6 учетов. Полевые исследования осуществлялись в соответствии со стандартными методиками (Романов, Мальцев, 2005). Современная фауна птиц ООПТ «Болото Железное» включает 38 видов из 8 отрядов, 19 семейств. В прежних исследованиях на этой ООПТ по одним данным указывалось обитание 11 видов (Ефремов, Балдаев, 1985), по другим – 29 видов (Балдаев, 1989). В целом за все годы наблюдений встречен 51 вид. Фауна птиц ООПТ «Болото Березовое» в настоящее время представлена 32 видами из 6 отрядов, 18 семейств. Максимальное видовое разнообразие по прежним данным оценивалось в 30 видов (Балдаев, 1989). Общее число видов птиц, зарегистрированных за все годы наблюдений на этой ООПТ, составляет 47. Изменения в видовом составе произошли как в результате элиминирования одних видов, так и выявления новых. По сравнению с данными последних исследований в настоящее время на обеих ООПТ в целом не было встречено 14 видов: поганка красношейная (Podiceps auritus (Linnaeus, 1758)), выпь большая (Botaurus stellaris (Linnaeus, 1758)), утка серая (Anas strepera (Linnaeus, 1758)), шилохвость (Anas acuta (Linnaeus, 1758)); погоныш (Porzana porzana (Linnaeus, 1766)), перевозчик (Actitis hypoleucos (Linnaeus, 1758)), бекас (Gallinago gallinago (Linnaeus, 1758)), веретенник большой (Limosa limosa (Linnaeus, 1758)), 119 чайка сизая (Larus canus (Linnaeus, 1758)), чайка малая (Larus minutus (Pallas, 1776)); свиязь (Anas penelope (Linnaeus, 1758)), травник (Tringa totanus (Linnaeus, 1758)), улит большой (Tringa nebularia (Gunnerus, 1767)), дупель (Gallinago media (Latham, 1787)). В то же время в наших учетах в границах ООПТ и их окрестностей было обнаружено 19 видов, ранее не указанных в списках их обитателей: огарь (Tadorna ferruginea (Pallas, 1764)), лунь полевой (Circus cyaneus (Linnaeus, 1766)), коршун черный (Milvus migrans (Boddaert, 1783)), канюк обыкновенный (Buteo buteo (Linnaeus, 1758)), цапля серая (Ardea cinerea Linnaeus, 1758), журавль серый (Grus grus (Linnaeus, 1758)), крачка речная (Sterna hirundo Linnaeus, 1758), голубь сизый (Columba livia Gmelin, 1789), ласточка деревенская (Hirundo rustica Linnaeus, 1758), жаворонок полевой (Alauda arvensis Linnaeus, 1758), трясогузка желтая (Motacilla flava Linnaeus, 1758), трясогузка желтолобая (Motacilla lutea (S. G. Gmelin, 1774)), трясогузка белая (Motacilla alba (Linnaeus, 1758)), скворец обыкновенный (Sturnus vulgaris (Linnaeus, 1758)), сорока (Pica pica (Linnaeus, 1758)), ворона серая (Corvus cornix (Linnaeus, 1758)), чекан луговой (Saxicola rubetra (Linnaeus, 1758)), синица большая (Parus major (Linnaeus, 1758)), воробей полевой (Passer montanus (Linnaeus, 1758)). Большинство из указанных видов непосредственно не обитают на ООПТ, но, несомненно, связаны с ними биоценотически, посещая объекты на водопое, кормежке, во время отдыха. Вероятно, что некоторые из них в прежние годы не учитывались в связи с приоритетами исследований гидрофильных птиц. Особо следует отметить два вида птиц, у которых северные границы ареалов проходят южней границ Республики Марий Эл: трясогузка желтолобая (Motacilla lutea (S. G. Gmelin, 1774)) и огарь (Tadorna ferruginea (Pallas, 1764)). Эти виды ранее на территории республики не встречались. Наши наблюдения являются первыми свидетельствами их обитания на территории РМЭ. Максимальная численность всех видов птиц, обитавших в сезон 2013 года на обеих ООПТ, составила 2071 особь. Из них 1283 особи отмечены в начале гнездового периода. Остальные 788 особей составили прирост популяций за гнездовой период. Ведущую роль в функционировании зооценозов ООПТ играют колониальные птицы, которые создают надежную защиту не только членам колонии, но и прочим видам, обитающим в пределах ООПТ. Согласно литературным данным на ООПТ «Болото Железное» с 1926 года существует колония чайки озерной (Larus ridibundus (Linnaeus, 1766)). В 70–90-х годах ХХ века численность чаек составляла от 1500 до 3000 гнездящихся особей (750–1 500 пар) (Балдаев, 1989; Ефремов, Балдаев, 1985). В 2013 году на ООПТ «Болото Железное» в начале гнездо120 вого периода нами было учтено 400–450 взрослых птиц (200–225 пар). Перед началом кочевок максимальная численность чаек в колонии ООПТ «Болото Железное» уже составляла около 600 особей. По сравнению с прежними годами численность гнездящихся в колонии чаек сократилась в 6,7 раза. Современная площадь колонии составляет 3,5– 4 га. Дата появления колонии чаек на ООПТ «Болото Березовое» не определена. В 70–90-х годах прошлого века их численность оценивалась в 60–100 особей (30–50 пар) (Балдаев, 1989). В 2013 году в начале гнездового периода учтено 450–500 особей (225–250 пар), что в 4,5–8,3 раз больше по сравнению с данными других исследований. К концу гнездового периода численность чаек в колонии ООПТ «Болото Березовое» составляла 600–650 особей. Современная площадь колонии составляет 6–6,5 га. Суммарная численность гнездящихся чаек в колониях обеих ООПТ в настоящее время составляет 850–900 особей. Таким образом, к 2013 году, несмотря на рост численности чаек в колонии ООПТ «Болото Березовое», произошло общее снижение численности вида в 1,8– 3,4 раза. Колонии крачек располагаются только на ООПТ «Болото Березовое». По разным сведениям в прежние годы здесь обитало от 40 до 300 особей крачки черной (Chlidonias niger (Linnaeus, 1758)) и 50–600 особей крачки белокрылой (Chlidonias leucopterus (Temminck, 1815)) (Балдаев, 1989; Красная книга…, 2002). В 2013 году в предгнездовой период в учетах присутствовало 15–20 особей крачек черных и около 80 особей крачек белокрылых. Кроме того, в 2013 году впервые отмечено гнездование крачки речной (Sterna hirundo (Linnaeus, 1758)) в количестве 15– 20 пар. В конце гнездового периода численность популяций указанных видов крачек с учетом молодых птиц составляла соответственно 30, 100 и 55 особей. В настоящее время колония крачек на ООПТ «Болото Березовое» занимает площадь 2–2,5 га и состоит из 70 пар: 10 пар – крачка черная, 20 пар – крачка речная и 40 пар – крачка белокрылая. Из редких птиц, включенных в Красную книгу Республики Марий Эл, на обеих ООПТ в прежние годы обитало 8 видов. Из них в 2013 году не было обнаружено 4 вида: поганка красношейная (Podiceps auritus (Linnaeus, 1758)), выпь большая (Botaurus stellaris (Linnaeus, 1758)), пастушок (Rallus aquaticus (Linnaeus, 1758)), веретенник большой (Limosa limosa (Linnaeus, 1758)). На момент проведения исследований на обеих ООПТ достоверно установлено обитание 5 «краснокнижных» видов: выпь малая (Ixobrychus minutus (Linnaeus, 1766)), камышница (Gallinula chloropus (Linnaeus, 1758)), крачка черная (Chlidonias niger (Linnaeus, 1758)), крачка белокрылая (Chlidonias leucopterus (Temminck, 1815)), журавль серый (Grus grus (Linnaeus, 1758)). Причем журавли были 121 отмечены здесь впервые. Численность редких видов птиц, обитавших на обеих ООПТ к началу гнездования, составляла 149 особей, к концу гнездового периода с учетом выросших птенцов – 183 особи. Для колониальных птиц в пределах ООПТ определены границы колоний, для редких видов зафиксированы координаты встреч. Все данные занесены на топографическую основу. По результатам проведенных исследований можно сделать следующие выводы: 1. ООПТ «Болото Железное» и ООПТ «Болото Березовое» характеризуются высоким разнообразием видов и высокой численностью представителей орнитофауны, в первую очередь колониальных и редких охраняемых видов. Это свидетельствует о высоком природоохранном значении этих ООПТ. 2. За последние 40–50 лет произошли заметные, в том числе негативные изменения в структуре орнитокомплексов ООПТ. Это требует продолжения изучения орнитоценозов для выяснения вызвавших их причин и предупреждения вероятных негативных последствий. 3. В настоящее время на обеих ООПТ обитает 5 редких видов птиц, включенных в Красную книгу РМЭ. Их суммарная численность в сезон 2013 года составила 183 особи. 4. Обнаружены новые для фауны Республики Марий Эл виды птиц: трясогузка желтолобая (Motacilla lutea (S. G. Gmelin, 1774)) и огарь (Tadorna ferruginea (Pallas, 1764)). Для оптимизации функционирования обследованных ООПТ, организации охраны популяций редких видов фауны РМЭ рекомендуется: 1. Включить в Красную книгу РМЭ новые для фауны республики виды. 2. Расширить границы существующих ООПТ, включив в их состав биотопы обитания трясогузки желтолобой (Motacilla lutea (S. G. Gmelin, 1774)), с целью обеспечения расселения и освоения видом новых территорий. Исследования проведены при поддержке Департамента экологической безопасности, природопользования и защиты населения Республики Марий Эл. Л и т е р а т ур а Балдаев Х. Ф. Железное и Березовое болота – царство пернатых / Х. Ф. Балдаев // Памятники природы Марийской республики. – Йошкар-Ола: Марийское кн. изд-во, 1989. – С. 60–64. Ефремов П. Г. Животный мир Марийской АССР. Птицы / П. Г. Ефремов, Х. Ф. Балдаев. – Йошкар-Ола: Марийское кн. изд-во, 1985. – 156 с. Красная книга Республики Марий Эл. Редкие и исчезающие виды животных. – Йошкар-Ола: Изд-во Марийского полиграфкомбината, 2002. – 164 с. Романов В. В. Методы исследований экологии наземных позвоночных животных: количественные учеты: учеб. пособие / В. В. Романов, И. В. Мальцев. – Владимир: Изд-во Владим. гос. ун-та., 2005. – 79 с. 122 РОМАШКА ДУШИСТАЯ (CHAMOMILLA SUAVEOLENS (PURSH) RYDB.) В НЕКОТОРЫХ МЕСТООБИТАНИЯХ НА ТЕРРИТОРИИ РЕСПУБЛИКИ МАРИЙ ЭЛ Жукова О. В. Марийский государственный университет, г. Йошкар-Ола, olga-v-zhukova@mail.ru Одной из причин сокращения биологического разнообразия является распространение инвазионных видов (Виноградова и др., 2009). В связи с этим инвазионные виды растений не могут оставаться без внимания специалистов. Из 52 инвазионных видов, описанных в «Черной книге флоры Средней России» (2009), на территории Республики Марий Эл произрастает 41 вид (Жукова, 2013). К их числу принадлежит ромашка душистая (Chamomilla suaveolens (Pursh) Rydb.), завезенная из Северной Америки, распространившаяся в Европе в 70-х годах позапрошлого столетия. В 1884 г. ромашка была посеяна в Казанском Ботаническом саду, в 1896 г. она упоминается там уже как одичавшая, а 1933 г. «в Маробласти встречается на каждом шагу» (Васильков, 1934). Этот вид распространен очень часто по всей территории Республики Марий Эл, произрастает в населенных пунктах, выгонах, пустырях, обочинах дорог (Абрамов, 1995). В НИЛ «Гербарий (YOLA) им. Н. В. Абрамова» хранится 19 гербарных листов ромашки душистой из 7 административных районов республики (Звениговский, Килемарский, Куженерский, Медведевский, Моркинский, Новоторъяльский, Советский). На территории Куженерского района в рудеральном местообитании вблизи сельскохозяйственных построек плотность ромашки душистой около 7350 раст./м2, на зарастающем асфальтовом покрытии у сельскохозяйственных складов – 358 раст./м2. В результате обхода территории г. Йошкар-Олы (маршрут протяженностью около 37,5 км) выявили места массового распространения ромашки в северо-западной, центральной и юго-восточной части города и места произрастания единичных растений в центральной части города. В одном из мест массового распространения ромашки, на газоне у автомобильной дороги на перекрестке улиц, плотность составляет 14 раст./м2. Это свидетельствует о том, что ромашка душистая распространена в разнообразных нарушенных местообитаниях Республики Марий Эл. Автор выражает благодарность д-ру биол. наук, проф. Н. В. Глотову за обсуждение материалов; канд. биол. наук, доц. Ю. Г. Суетиной за консультации; зав. НИЛ «Гербарий YOLA им. Н. В. Абрамова» Л. А. Абрамовой за предоставленные гербарные листы растений; студ. А. С. Локшину за помощь в сборе материала. 123 Работа выполнена при поддержке Гранта РФФИ (проект № 12-04-01251). Л и т е р а т ур а Абрамов Н. В. Конспект флоры Республики Марий Эл / Н. В. Абрамов. – ЙошкарОла: РФФИ; МарГУ, 1995. – 192 с. Васильков Б. К истории флоры Марийской области (из работ геоботанической экспедиции /Марийского научно-исследовательского института): Отдельный оттиск из журнала «МАО». 1933. № 11–12 / Б. Васильков. – Йошкар-Ола: Типография МАРГИЗА, 1934. – 18 с. Виноградова Ю. К. Черная книга флоры Средней России / Ю. К. Виноградова, С. Р. Майоров, Л. В. Хорун. – М.: ГЕОС, 2009. – 494 с. Жукова О. В. Инвазионные виды растений на территории Республики Марий Эл / О. В. Жукова // Актуальные проблемы экологии, биологии и химии: материалы Республиканской научно-практической конференции / Мар. гос. ун-т. – Йошкар-Ола, 2013. – Вып. 4. – С. 22–23. ИЗМЕНЕНИЕ БИОРАЗНООБРАЗИЯ РАСТЕНИЙ В ХОДЕ СУКЦЕССИОННЫХ ПРОЦЕССОВ НА ЗАЛЕЖАХ Закамская Е. С., Зотина С. Г. Марийский государственный университет, г. Йошкар-Ола, zakamskay@mail.ru В настоящее время интенсивное ведение хозяйственной деятельности, нарушение агротехники, влияние экономических факторов и целого ряда других причин приводят к выведению сельскохозяйственных земель из севооборота. В связи с этим особую актуальность приобретает решение вопросов, связанных с перспективами использования залежей и оценки интенсивности процессов их восстановления. Целью работы является изучение видового и структурного биоразнообразия растений в ходе сукцессионных процессов на залежных землях. Исследования проводились в июле 2007, 2008 и 2013 гг. в Медведевском районе Республики Марий Эл на территории садового товарищества «Маяк». Ранее на территории, которая в дальнейшем была отведена под садоводческое товарищество, произрастал елово-березовый с липой мелколистной лес. После вырубки и раскорчевки деревьев почва возделывалась под посадку картофеля в течение 10–15 лет. Для изучения демутационных процессов, происходящих на залежных землях были выбраны участки размером 400 м2, которые не возделывались в течение 1 года, 2, 3, 5, 6, 7 лет. Под залежными землями (залежами) мы понимаем распаханные и неиспользуемые в течение нескольких лет, заброшенные земли. На каждом участке были проведены стандартные геоботанические описания (Программа и методика…, 1974). В ходе работы выявляли флористический состав изучаемых фитоценозов, определяли проективное покрытие (ПП) ценопопуляции (Цп) каждого вида растений. Принадлежность видов травянистых растений к эколого-ценотической 124 группе (ЭЦГ) растений определяли согласно баз данных ценофонда лесов европейской России (http://mfd.cepl.rssi.ru/flora/ecogroup.html) и института математических проблем биологии РАН (http://www.impb.ru/ index.php?id=div/lce/ecg). Видовые названия растений приводятся по сводке К. С. Черепанова (1995). В ходе зарастания залежных территорий увеличивается флористическое разнообразие. На залежи первого года произрастало 16 ценопопуляций разных видов сосудистых растений, относящихся к 9 семействам. Через 6–7 лет флористическое разнообразие увеличилось до 39 видов и 19 семейств (рис. 1). Из сообществ исчезает семейство Подорожниковые (Plantaginaсеае), появляются семейства Бурачниковые (Boraginaceae), Гвоздичные (Caryophyllaceae), Гераниевые (Geraniaceae), Зверобойные (Hypericaceae), Лютиковые (Ranunculaceae), Осоковые (Cyperaceae), Розовые (Rosaceae), Фиалковые (Violaceae). Однако, несмотря на то, что, в ходе сукцессионного процесса таксономическое разнообразие изменилось количественно и качественно, появились новые таксоны, ведущими по числу видов семействами являются Сложноцветные (Compositae) и Бобовые (Fabaceae) (рис. 2). Происходит смена доминант: в первый год преобладает марь белая (Chenopodium album L) – проективное покрытие составляет 50 %; во второй – иван-чай узколистный (Chamaenerion angustifolium (L.) Scop.) (ПП 60 %), и начиная с третьего года – вейник тростниковидный (Calamagrostis arundinacea (L.) Roth) (ПП на 3 год – 30 %, в последующие – 100 %). 45 Число ЦП растений 40 35 30 25 20 15 10 5 0 1 год 2 года 3 года 5 лет 6 лет 7 лет Возраст залежи Рисунок 1 – Динамика числа ЦП растений при зарастании залежей 125 25,0 50 45 20,0 40 35 15,0 30 % % 25 10,0 20 15 10 5,0 5 0 0,0 1 год сложн. 2 года бобовые 3 года гречишн. 5 лет мятликов. сложн. 7 лет 6 лет бобовые мятликов. норичн. Рисунок 2 – Динамика ведущих семейств при зарастании залежей Для оценки биоразнообразия и сукцессионной динамики растительности используется состав эколого-ценотических групп растений (Биоразнообразие..., 2006). В первый год на залежах преобладают эксплерентные сорные растения. В дальнейшем, в ходе сукцессионного процесса их доля снижается, увеличивается биоразнообразие луговых видов. Так как исследованные участки в прошлом были заняты смешенным елово-березовым с липой мелколистной лесом и находятся в непосредственной близости от лесного ценоза, начиная с 5 года в растительных сообществах начинают преобладать ценопопуляций, относящихся к лесной группе (рис. 2., табл.). В состав лесной фракции входят растения, относящиеся к бореальной, неморальной, боровой и черноольшанниковой группам, с преобладанием неморальных видов. Среди луговых растений первенство принадлежит свеже-луговой ЭЦГ (табл.). % 60 50 40 30 20 10 0 1 год 2 года 3 года 5 лет 6 лет 7 лет ЭЦГ сорная луговая лесная Рисунок 3 – Динамика эколого-ценотической структуры на залежи 126 Состав эколого-ценотических групп растения Br Nm Pn Nt Wt MFr MDr Rd 1 год 0,0 5,9 0,0 17,6 5,9 23,5 5,9 41,2 2 года 4,0 8,0 8,0 16,0 4,0 28,0 8,0 24,0 3 года 6,3 6,3 6,3 15,6 6,3 31,3 12,5 15,6 5 лет 8,6 25,7 2,9 14,3 0,0 37,1 5,7 5,7 6 лет 12,8 25,6 5,1 10,3 2,6 33,3 7,7 2,6 7 лет 10,5 23,7 2,6 7,9 2,6 39,5 7,9 5,3 Таким образом, в ходе сукцессионного процесса на залежных землях происходит увеличение таксономического разнообразия, на смену сорным растениям приходят лесные виды Л и т е р а т ур а Биоразнообразие и сукцессионный статус старовозрастных темнохвойных лесов Европейской России / О. В. Смирнова, М. В. Бобровский, Л. Г. Ханина, В. Э. Смирнов // Успехи совр. биол. – 2006. – Т. 126, № 1. – С. 27–49. Программа и методика биогеоценологических исследований. – М.: Наука, 1974. – С. 14–23. Расширенная система экологоценотических групп видов сосудистых растений для бореальной, гемибореальной и умеренной лесных зон Европейской России [Электронный ресурс] / Институт математических проблем биологии РАН – Режим доступа: http://www.impb.ru/index.php?id=div/lce/ecg, свободный. Черепанов С. К. Сосудистые растения России и сопредельных государств (в пределах бывшего СССР) / С. К. Черепанов. – СПб.: Мир и семья, 1995. – 992 с. Эколого-ценотические группы растений / Ценофонд лесов Европейской России. – Режим доступа: http://mfd.cepl.rssi.ru/flora/ecogroup.html, свободный. ФРАГМЕНТАЦИЯ ПОПУЛЯЦИЙ РЕДКИХ ВИДОВ РАСТЕНИЙ КАК ФОРМА ИХ ДЕГРАДАЦИИ Клименко А. А. Сумский национальный аграрный университет, г. Сумы, Украина, hgrip@rambler.ru Сокращение биоразнообразия – основная проблема, которая стоит перед людьми ХХІ века (Злобин, Клименко, 2010). Тенденции, наметившиеся в прошлом столетии, усугубляются и прогрессируют. Потеря биологического вида, как составляющей части биоразнообразия, – это процесс, который проходит несколько этапов: – снижение численности особей в крупной локальной популяции; – расчленение популяции на отдельные фрагменты; 127 – формирование метапопуляций под влиянием антропогенных факторов (например, разрушение местообитаний вида) или реже природных факторов так, что обмен генетическим материалом становится практически невозможным; – постепенная деградация популяций, которая проявляется на организменном и популяционном уровнях; – полное вымирание локальных популяций, что, в свою очередь, приводит к исчезновению вида в данном регионе. Комплексные исследования популяций редких видов растений в их динамике и выявление этапов, в которых они находятся в конкретный промежуток времени, это проблемы, требующие постоянного контроля и проведения стационарных популяционных исследований (Злобин, Скляр, Клименко, 2013). В ином случае, для сохранения биоразнообразия мы сможем сделать лишь одно – поставить напротив вида природоохранный статус «Исчезнувший». Основной задачей, которую на данное время ставит перед собой фитосозология, является определение количества популяций каждого редкого вида с дальнейшим их исследованием (Дідух, 2010). Относительно количества и качества популяций редких видов растений возможны три основных варианта: а) небольшое количество популяций одного вида с высокой численностью особей; б) много популяций одного вида с низкой численностью особей; в) небольшое количество популяций одного вида (иногда единичные популяции) с низкой численностью особей. Отсюда следует, что охарактеризовать природоохранный статус того или иного вида можно при наличии сведений относительно количества популяций вида и качества особей в этих популяциях. Целью нашей работы было на основе модельного объекта проанализировать количество и качество популяций с дальнейшей характеристикой прогноза стабильности данного вида. В качестве модельного вида изучались популяции Liliummartagon L. Вид занесен в Красную книгу Украины (2009), имеет природоохранный статус «неоцененный». L. martagon произрастает во многих областях Украины, потому для исследований, в качестве модельной территории, был выбран Национальный природный парк «Деснянско-Старогутский» (Украина, Сумская область), где охраняется данный вид. Сбор полевого материала проводился на протяжении 2009–2013 годов. Устанавливали количество популяций, численность особей в их динамике, измеряли морфометрические параметры с использованием неразрушающих методов морфометрии (Панченко, 2007). В процессе статистической обработки материала при помощи некоммерческих специализированных компьютерных программ ANONS и VITAL (разра128 ботчик – Злобин Ю. А.) определяли онтогенетическую и виталитетную структуру популяций. На территории Национального природного парка «ДеснянскоСтарогутский» исследовали четыре локальных популяции L. martagon (Клименко, 2011), разделенных между собой территориально. Условно три популяции можно отнести к так называемым лесным, поскольку они находятся в фитоценозах с высокой сомкнутостью деревостоя и хорошо развитым подлеском: популяция из экотона между Betuletacorylosa Pinetacorylosa-convallariosum (П1), популяция из фитоценоза Pinetacorylosa-convallariosum (П2) и Pinetacorylosa-sparsiherbosum (П3). Четвертая популяция находится на лесной опушке в фитоценозе Fraxinetacorylosa-convallariosum (П4). Было установлено, что под влиянием различных эколого-ценотических условий, которые действовали достаточно продолжительный период времени, данные популяции формировались как полностью изолированные. Сходство между особями L. martagon в разных популяциях прослеживаются только в ювенильном состоянии, тогда как уже виргинильные особи значительно отличаются между собой даже в тех случаях, когда фитоценотические условия несколько схожие (например П2 и П3) (Клименко, 2012). В связи с этим имеются значительные различия в популяционных структурах – онтогенетической и виталитетной. Онтогенетический анализ показал, что у лесных популяций (П1, П2 и П3) преобладали ювенильные особи с полным отсутствием генеративных растений. У популяции П4, произрастающей на опушке, были представлены все онтогенетические группы особей с примерно равным количеством как ювенильных, так и генеративных растений (25,5 % и 21,6 % соответственно). Ввиду отсутствия у трех популяций из четырех исследуемых генеративных растений, виталитетный анализ был проведен для особей каждого онтогенетического состояния в отдельности, а впоследствии выведен интегральный индекс качества популяции. В результате выяснили, что у популяции из фитоценоза Fraxinetacorylosa-convallariosumиндекс качества популяции самый высокий – Q = 0,33. Лесные популяции оценивались как депрессивные или равновесные, однако, при полном отсутствии цветущих и плодоносящих растений. Еще одним важным показателем является общая численность особей в популяциях L. martagon. Исследования на протяжении пяти лет показали, что только в популяции № 4 преобладали позитивные тенденции, во всех остальных популяциях отмечали постепенное уменьшение числа особей L. martagon, порой даже до полного отсутствия надземной части растений на протяжении всего вегетативного периода (П3). 129 Такой комплексный подход к изучению популяций показал, что только у опушечной популяции П4 отмечено увеличение числа особей, полночленная онтогенетическая структура и высокий уровень жизненности. Фрагментация популяций приводит к образованию неустойчивых, угнетенных и малочисленных групп растений одного вида. Неоптимальные фитоценотические условия произрастания являются причиной формирования популяций с неполной онтогенетической структурой и низким состоянием виталитета. Такие популяции обречены или на постепенное вымирание, или на существование в угнетенном вегетативном состоянии особей без возможности образования генетов и, таким образом, эволюционного прогресса. В. М. Ивонин с соавторами (2012), рассматривавшие проблему фрагментации лесов, пришли к выводу, что крупные фрагменты (площадью от 43 га) характеризуются слабым уровнем деградации лесной среды. Именно под полог таких крупных фрагментов авторы и рекомендуют пересаживать редкие и находящиеся под угрозой исчезновения растения. Все технологические аспекты по данному вопросу широко высветлены в монографии «Экология и охрана дикой природы» (Yahner, 2000). Л и т е р а т ур а Дідух Я. П. Підсумкищо до третього видання «Червоної книги України. Рослиннийсвіт» / Я. П. Дідух // Зб. «Рослин. світ у Червоній книзі України: впровадження глоб. стратегії збереження рослин». – К.: Альтерпрес, 2010. – С. 12–13. Злобін Ю. А. Що ми знаємо і що не знааємо при рідкісні рослини / Ю. А. Злобін, Г. О. Клименко // Чорномор. ботан. журн. – 2010. – Т. 6, № 2. – С. 150–161. Злобин Ю. А. Популяции редких видов растений: теоретические основы и методика изучения: монография / Ю. А. Злобин, В. Г. Скляр, А. А. Клименко. – Сумы: Университетская книга, 2013. – 439 с. Ивонин В. М. Фрагментация горных лесов при размещении объектов Олимпиады – 2014 / В. М. Ивонин, М. Д. Пиньковский, А. В. Егошин // Лесное хозяйство. – 2012. – № 1. – С. 31–34. Клименко Г. О. Онтогенетична структура ценопопуляцій рідкісних видів рослин на території Національного природного парку «Деснянсько-Старогутський» / Г. О. Клименко // Укр. ботан. журн., 2011. – Т. 68, № 5. – С. 663–671. Клименко Г. О. Фенотипічна мінливість деяких видів рідкісних рослин / Г. О. Клименко // Актуальні проблеми ботаніки та екології: Міжнародна конференція молодих учених, 19–23 вересня 2012 р. (Ужгородський національний університет): матеріали конф. – Ужгород, 2012. – С. 143– 144. Панченко С. М. Неразрушающие методы морфометрического анализа рідких растений и их применение на примере Huperziaselago (Huperziaceae) / С. М. Панченко // Заповідна справа в Україні. – 2007. – Т. 13, вип. 1–2. – С. 106–110. Червона книга України. Рослинний світ. / ред. Я. П. Дідух. – К.: Глобал-консалтинг, 2009. – 900 с. Yahner, Richard H. Eastern DeciduousForest: Ecologyand Wildlife Conservation / Richard H. Yahner. – Minneapolis: University of Minnesota Press, 2000. – 279 p. 130 ВИДОВОЕ РАЗНООБРАЗИЕ ЗООПЛАНКТОНА КАК ПОКАЗАТЕЛЬ ЭКОЛОГИЧЕСКОГО СОСТОЯНИЯ МАЛЫХ ВОДОТОКОВ НИЖНЕГО НОВГОРОДА Макеев И. С. 1, Гаврилко Д. Е. 2 1 Нижегородский государственный университет им. Н. И. Лобачевского 2 Национальный исследовательский университет, г. Н. Новгород igmakeyev@mail.ru Малые водоемы и водотоки являются резерватами биоразнообразия водных, полуводных и околоводных сообществ и для городской среды имеют важное рекреационное значение. Экосистемы малых водотоков (малых рек и каналов) из-за низкой буферной емкости и очень большой площади контакта с береговой зоной очень чувствительны к загрязнению и хозяйственному преобразованию водосборной территории. В условиях урбаногенной среды эти водные объекты испытывают интенсивную антропогенную нагрузку, обусловленную промышленным, транспортным и хозяйственно-бытовым загрязнением. Экологическая группа зоопланктона активно участвует в саморегуляции водных экосистем. Благодаря фильтрации и седиментации взвешенных в воде минеральных и детритных частиц организмы зоопланктона способствуют самоочищению воды, повышению ее прозрачности. Видовой состав и видовое разнообразие зоопланктона служат показателями скорости течения, экологической гетерогенности и качества водной среды. Зоопланктон малых рек формируется благодаря наличию рефугиумов – периферических и придаточных отделов водной экосистемы, в которых развиваются автохтонные сообщества зоопланктона (Богатов, 1995; Крылов, 2005). Поэтому изучение зоопланктоценозов малых рек требует проведение облова основных типов прибрежно-водных биотопов, а также притоков и пойменных придаточных водоемов, накапливающих планктонофонд. Зоопланктон рипальной зоны характеризует транзитный обменный фонд реки. Исследования зоопланктона 5 малых рек Н. Новгорода проведены в ходе отбора серии количественных и качественных проб 9–10.07.2013 на разных участках и в разных биотопах. Малые реки Старка и Кова протекают по возвышенной территории окско-волжского водораздела, остальные водотоки расположены в низменной части левобережной долины р. Оки. По результатам исследований комитетом охраны окружающей среды и природных ресурсов г. Н. Новгорода водотоки характеризуются как «грязные» и «очень грязные». Преобладают нефтепродукты, железо, марганец, тяжелые металлы, аммоний, легко окисляемые 131 органические вещества. Наименее загрязненной является р. Гниличка, а наиболее – р. Ржавка. Наименьшее видовое богатство и количественное развитие зоопланктона (табл.) отмечается в реках Старке и Кове. Это обусловлено относительно быстрым течением и высокой степенью загрязнения берегов и вод. Доминирует по численности и биомассе эврибионтный вид Bosmina longirostris (O. F. Müller, 1785). В р. Кове субдоминантом по численности является тепловодная прудовая коловратка Hexarthra mira (Hudson, 1871), выносимая из придаточных луж. Высокое видовое богатство и количественное развитие зоопланктона наблюдается в р. Левинке с медленным течением и искусственно расширенным и подпруженным руслом. В среднем участке доминантом по численности выступает коловратка Kellicottia bostoniensis (Rousselet, 1908) – интродуцент из Северной Америки, недавно обнаруженный в 13 разнотипных озерах Северо-западного и Центрального регионов России, включая Владимирскую область (Жданова, Добрынин, 2011); западных и югозападных районов Нижегородской области (Баянов, 2013). Численность вида в р. Левинке достигает 67 тыс. экз./м3. Интересно отметить, что интродуцент сосуществует с аборигенным видом Kellicottia longispina (Kellicott, 1879) (до 500 экз./м3), постепенно вытесняя последний. Река Ржавка, несмотря на сильное загрязнение, имеет хорошо развитый комплекс зоопланктона, ее истоком является Шуваловский мелиоративный канал, где количественное и качественное развитие зоопланктона невысокие. По численности и биомассе наибольшего развития планктон достигает в прудообразном расширении русла в среднем течении. Абсолютным доминантом здесь выступает тепловодный, α-мезосапробный вид ветвистоусого рачка Daphnia pulex (Leydig, 1860). Видовое богатство максимально в нижнем течении. Р. Гниличка имеет каскад прудов, болотное питание и хорошо развитую погруженную и плавающую растительность, поэтому здесь отмечается большое число видов и, в основном, высокая численность зоопланктона. Исключением является нижний пруд, сильно заросший телорезом, где размножение многих видов зоопланктона подавлено. Индекс Шеннона не вполне адекватно характеризует видовое разнообразие вследствие сильной зависимости от выравненности видов. В случаях высокой численности или биомассы планктона, что обычно обусловлено массовым развитием одного, реже двух видов, значение индекса меньше 1,0. Некоторые авторы (Свирская, 1992; Андроникова, 1996) интерпретируют такое состояние влиянием экстремальных условий среды. 132 Показатели видового богатства (S), численности (N, тыс. экз./м3), биомассы (B, мг/м3), индекса видового разнообразия Шеннона (HN, HB, бит) и состава доминирующих видов Кова Старка Левинка Гниличка Ржавка Водоток Место отбора проб HN Доминант + субдоминанты S N B исток 8 2,00 12,4 2,835 2,571 Nauplii+ Copepoditi Eudiaptomus gracilis+ Eurycercus lamellatus вход в коллектор 4 6,41 38,9 1,558 2,275 Nauplii+ Copepoditi Ceriodaphnia pulchella + Daphnia pulex выход коллектора 7 6,33 27,0 2,574 2,738 Copepoditi + Nauplii Copepoditi + + Keratella quadrata D. pulex прудовое расширение 7 устье 16 пруд № 1 17 23,52 332,4 0,703 1,262 B. longirostris B. longirostris речной уч. 15 49,89 68,1 0,540 1,083 B. longirostris B. longirostris пруд № 2 12 26,2 2,965 1,691 A. priodonta A. priodonta+ Copepoditi верхний участок 17 84,14 4229 3,230 2,410 Copepoditi +Nauplii A. priodonta +E. gracilis+ Copepoditi средний участок 16 66,59 11,79 0,766 2,884 K. bostoniensis+ Keratella quadrata B. longirostris + Copepoditi устье 3 0,141 B. longirostris +Nauplii A. priodonta+ Copepoditi выше устья р. Старки 5 0,548 0,775 1,470 1,450 B. longirostris + H. mira B. longirostris+ H. mira ниже устья р. Старки 3 0,074 H. mira+ B. longirostris B. longirostris+ Copepoditi HB по N 55,66 2482 0,658 0,268 Daphnia pulex 3,26 4,07 26,4 2,734 2,123 0,35 1,983 2,087 0,10 1,753 1,551 133 по B D. pulex+ Cyclops strenuus Nauplii+Copepoditi. D. pulex+ + D. pulex Copepoditi Таким образом, несмотря на значительное загрязнение, зоопланктон малых рек г. Н. Новгорода в середине лета в целом имеет естественный характер развития и уровень видового разнообразия обычный для водотоков такого типа. Наибольшее видовое богатство и количественное развитие отмечается обычно на участках замедленного течения при наличии развитой водной растительности. Внедрение интродуцента K. bostoniensis в планктон водотоков замедленного течения облегчается лабильностью их экосистем. Зоопланктон рек Ржавки и Левинки имеет лучшие показатели экологического состояния по сравнению с исследованиями 2000–2004 гг. (Шурганова, Макеев, 2005). Л и т е р а т ур а Андроникова И. Н. Структурно-функциональная организация зоопланктона озерных экосистем / И. Н. Андроникова – СПб.: Наука, 1996. – 208 с. Баянов Н. Г. Встречаемость и уровень развития Kellicottia bostoniensis (Rousselet, 1908) в озерах Нижегородской области / Н. Г. Баянов // Чужеродные виды в Голарктике: Тез. докл IV междунар. симпоз. (ИБВВ РАН, Борок, 22–28 сент. 2013). – С. 48. Богатов В. В. Комбинированная концепция функционирования речных экосистем / В. В. Богатов // Вестник ДВО РАН. 1995. – № 3. – С. 51–61. Жданова С. М. Kellicottia bostoniensis (Rousselet, 1908) (Rotifera: Brachionidae) в водоемах Европейской России / С. М. Жданова, А. Э. Добрынин // Биология внутренних вод. – 2011. – № 1. – С. 45–52. Крылов А. В. Зоопланктон равнинных малых рек / А. В. Крылов. – М.: Наука, 2005. –263 с. Свирская Н. Л. Мониторинг зоопланктона / Н. Л. Свирская // Руководство по гидробиологическому мониторингу пресноводных экосистем. – СПб.: Гидрометеоиздат, 1992. – С. 123–134. Шурганова Г. В. Зоопланктон / Г. В. Шурганова, И. С. Макеев // Экологическое состояние водных объектов Нижнего Новгорода. – Н. Новгород: Изд-во ННГУ, 2005. – С. 89–98. ОСОБЕННОСТИ РАСПРОСТРАНЕНИЯ И УЧЕТ ПО НОРАМ БАРСУКА ОБЫКНОВЕННОГО (MELES МELES BRISSON, 1762) НА ТЕРРИТОРИИ ЖИГУЛЕВСКОГО ГОСУДАРСТВЕННОГО ЗАПОВЕДНИКА Мартынов А. В., Фокина М. Е. Самарский государственный университет, г. Самара, mariyafok@mail.ru Барсук (Meles мeles (Brisson, 1762)) относится к отряду Хищные – Carnivora (Fissipedia), семейству Куньи – Mustelidae. Он является одним из самых крупных представителей этого семейства. Это широко распространенный вид: по всей Европе, до северной Карелии, а в Сибири – до Сургута, встречается также на Кавказе, в Закавказье, в Крыму, в Средней, Центральной и Восточной Азии, на Дальнем Востоке доходит на север до Николаевска-на-Амуре. Наиболее многочислен барсук в средне-лесной полосе, в лесостепи и в горах (Новиков, 1956). В Жигулевском заповеднике обитает Meles meles L., 1758 – барсук обыкновенный или европейский. Объектом исследования являлась популяция барсука 134 на территории Жигулевского заповедника имени И. И. Спрыгина в районе Морквашинского и Бахиловского лесничеств. Жигулевский заповедник имени И. И. Спрыгина входит в центральный массив Жигулей, ограниченный с запада Морквашинской, а с востока – Ширяевской долинами. В пределах заповедника хребет рассекается двумя долинами: Бахиловой Поляной и Молебным оврагом. Северные склоны Жигулей круто опускаются к Волге и разделены большим количеством ущельеобразных «оврагов» или «буераков» на отдельные отроги, покрытыми лиственными лесами. Окончания отрогов увенчиваются скальными обнажениями – утесами, «шишками» или «шиханами». Ширина Жигулей с севера на юг колеблется от 2 до 3 км. В составе древостоев преобладает осина, клен остролистый и береза повислая («Зеленая книга…», 1995). Местом проведения исследования выбраны Морквашинское и Бахиловское лесничества, так как в этих районах, по данным прошлого века, барсук был широко распространен, а подобные исследования не проводились на территории Жигулевского заповедника более 15 лет. Для выявления особенностей распространения барсука был выбран метод картирования. Городки наносились на карту масштабом 1 : 100 м с использованием системы GPS. Обход маршрутов производился весной – с марта по конец мая, летом – в июле, и осенью – с сентября по конец ноября в 2011–2012 гг. Исследования начинались, когда средняя температура днем становилась выше 5 ºC, и заканчивались, когда она понижалась до –2ºС. Время проведения исследований – с 7 до 15 часов. Маршруты исследования прокладывались в наиболее вероятных местах обитания барсука, в основном в горных районах, оврагах. Всего было проложено 35 маршрутов. В описания городков вносились дата обнаружения городка, время, ставилась метка на карте, отмечалась также удаленность от населенных пунктов, количество входов, наличие обитателей (жилая или брошенная), наличие уборной, общая характеристика местности. В ходе исследований было обнаружено 6 городков барсука. Все они находились в малопроходимых удаленных местах. В основном это были густо заросшие в летнее время овраги. Городок № 1 оказался брошенным, вероятно, по причине частого посещения этой зоны людьми. Вокруг было обнаружено много бытового мусора, оставленного дачниками близлежащего дачного массива. Городок № 2 находится на минимальном удалении от населенного пункта. Он расположен на крутом спуске в овраге, норы его расположены в 2 яруса. В летнее время этот овраг является труднопроходимым изза обильного зарастания крапивой и различными кустарниками. Выхо135 дов в этом городке пять, что может свидетельствовать о том, что городок довольно старый и постоянно используется. Городки № 5 и 6 находятся в зоне, где достаточно много поваленных деревьев. Они являются не постоянным местом обитания, так как уборная рядом не была обнаружена. Таким образом, на территории Жигулевского заповедника в 2012 г. обнаружено 6 городков барсука, 5 из них являются жилыми, один заброшен из-за частого посещения этого места людьми. Городки № 2–6 – менее подвержены влиянию антропогенного фактора, так как находятся на склонах оврагов в валежнике и удалены от населенных пунктов и дачных массивов. Рисунок 1 – Расположение городков барсука на территории Жигулевского заповедника Особенности барсучьих нор проведены в таблице. Особенности обнаруженных барсучьих нор на территории Жигулевского заповедника № городка Заселенность Количество входов Наличие уборных Удаленность норы от населенного пункта (км) 1 – 3 – 1,9 2 + 5 + 0,8 3 + 4 + 3,8 4 + 5 + 2,1 5 + 2 – 3,4 6 + 4 – 1,2 136 По данным 30–60-х годов прошлого столетия барсук обыкновенный – широко распространенный вид, которого можно было встретить во всех районах заповедника (Белянин, 1981). По данным П. Б. Юргенсона (1968), в 30-е годы на территории Жигулевского заповедника барсук был многочисленным. В это время было зафиксировано 55 городков. В 1980 г. В. Н. Белянин отметил лишь 24 городка. В 2000 году барсук был зарегистрирован лишь в Зольненском лесничестве (Вехник, 2000). Мы сравнили данные о численности барсука на территории Жигулевского заповедника, результаты приведены на рисунке 2. Рисунок 2 – Количество городков барсука обыкновенного на территории Жигулевского заповедника в разные годы (данные 1933 г. – по Юргенсону (1968), данные 80-х – по Белянину (1981), 2012 г – собственные данные). За промежуток времени с 30-х годов прошлого века по нынешний период произошло значительное уменьшение численности городков барсука (с 55 до 6) на территории Жигулевского заповедника. Это, вероятно, связано с охотой на него в период закрытия заповедника, а также с изменением ландшафта в связи с геолого-добывающей деятельностью и расширением зон рекреации. Л и т е р а т ур а Белянин В. Н. Млекопитающие Жигулевского заповедника / В. Н. Белянин // Сб. науч. тр. ЦНИЛ Главохоты РСФСР. – М., 1981. – С. 89–103. Вехник В. П. Критические замечания к фауно-таксономическому составу млекопитающих Самарской Луки / В. П. Вехник // Биологическое разнообразие заповедных территорий: оценка, охрана, мониторинг: сб. науч. тр. / Жигулев. гос. заповед. им. И. И. Спрыгина. – Самара, 2000. – С. 310–317. «Зеленая книга» Поволжья: Охраняемые природные территории Самарской области / сост. А. С. Захаров, М. С. Горелов. – Самара: Книжное изд-во, 1995. – 352 с. Новиков Г. А. Хищные млекопитающие фауны СССР / Г. А. Новиков. – М.; Л., 1956. – 294 с. Юргенсон П. Б. Охотничьи звери и птицы (прикладная экология) / П. Б. Юргенсон. – М.: Лесная пром-ть, 1968. 137 СОСТОЯНИЕ ЗЕЛЕНЫХ НАСАЖДЕНИЙ САДОВО-ПАРКОВЫХ ТЕРРИТОРИЙ г. КАЗАНИ Мударисов Р. Г. Казанский (Приволжский) федеральный университет, г. Казань, raffikk@mail.ru Антропогенное давление на зеленую зону территории городов являются актуальной темой для научного изучения. Зеленые зоны являются своеобразным аналогом лесной экосистемы, но в значительной степени преобразованной человеком. Наиболее изменены природные ландшафты на территориях современных городов и промышленно-городских агломераций. Из года в год города становятся средой жизни всевозрастающего числа людей. Возросшее внимание к изучению проблем городской среды и накопление сведений, касающихся приспособлений живых объектов к существованию в тесном контакте с человеком, способствовали выбору настоящей темы. Цель исследований – учет древесно-кустарниковых насаждений, выяснение их видового состава и состояния, учет повреждений деревьев и выяснение их причин, а также оценка ухода за древесными насаждениями. Работа по инвентаризации уличных насаждений проводилось в течение 2 лет, вегетативный период 2011–2012 гг. Всего обследовано 3 сада (Лядской, им. Маяковского и Ленинский), 11 парков (Победы, ДК Химиков, Урицкого, Крылья Советов, Петрова, Шурале (Кырлай), Молодоженов, Горького, Черное озеро, Миллениум) и 6 скверов (по ул. Тимирязева, Кирова, Чехова, по ул. Тинчурина, Гоголя, Тукая). Проводилась работа по уточнению их площадей, измерению площади под газонами и аллеями, выяснению площади вытоптанных участков, определению видового состава растений и их состояния, проводился учет повреждений деревьев и выяснение их причин, включая пораженность грибными заболеваниями и вредными насекомыми (Бакин, 2000). Отмечены следующие виды древесных растений, произрастающих в скверах, садах и парках города: 1) береза бородавчатая; 2) дуб черешчатый; 3) вяз обыкновенный; 4) ива белая; 5) осина; 6) тополь бальзамический; 7) тополь серебристый; 8) клен остролистный; 9) клен татарский; 10) клен ясенелистный; 138 11) каштан конский; 12) туя западная; 13) липа мелколистная; 14) ясень обыкновенный; 15) рябина обыкновенная; 16) черемуха Маака; 17) черемуха обыкновенная ; 18) вишня пенсильванская; 19) яблоня лесная; 20) ель голубая; 21) ель обыкновенная; 22) лиственница сибирская; 23) лиственница европейская; 24) сосна обыкновенная. Самыми распространенными деревьями парков и скверов являются клен ясенелистный (50 %), липа мелколистная (20 %). Очень слабо представлены хвойные породы: ели, лиственницы, сосны – тремячетырьмя деревьями в отдельных скверах (Мударисов, Валиев, 2013). В заключение можно дать несколько предварительных выводоврекомендаций: 1. В уличных насаждениях следует разнообразить видовой состав, широко внедряя не только клен ясенелистный и липу мелколистную, но и более декоративные породы – ясень пушистый, тополь берлинский, лиственницу сибирскую, черемуху Маакка и др. 2. Наиболее благоприятной для роста древесных насаждений является посадка их на газонах; даже при частичном огораживании в этом случае почти полностью исключаются повреждения древесных насаждений. При высаживании деревьев не на газонах надо применять приствольные решетки. Необходимо широко внедрять в озеленение улиц кустарники. 3. При любых условиях парки и скверы должны быть защищены от улиц плотными и высокими защитными зелеными посадками. 4. Являясь хорошим шумовым экраном, эффективным озонатором и озеленительным агентом, клен ясенелистный может быть рекомендован в качестве основного посадочного материала на производственных площадках, для создания естественных зеленых фильтров-экранов, ограждающих промышленные предприятия (Экология города Казани, 2005). Л и т е р а т ур а Бакин О. В. Сосудистые растения Татарстана / О. В. Бакин, Т. В. Рогова, А. П. Ситников. – Казань: Изд-во Казанского ун-та, 2000. – 496 с. Экология города Казани. – Казань: Изд-во «Фэн» Академии наук РТ, 2005. – 576 с. Мударисов Р. Г. Привлекательность и экологическая оценка зеленых насаждений города Казани в связи с проведением Универсиады 2013 / Р. Г. Мударисов, М. Р. Валиев // Экология России: на пути к инновациям: межвузовский сб. науч. тр. / сост. Т. В. Дымова. – Астрахань: Изд-во Нижневолжского экоцентра, 2013. – Вып. 7. – 214 с. 139 ОСОБЕННОСТЬ ЭКОЛОГИИ PASSER DOMESTICUS L., 1758 И PASSER MONTANUS L., 1758 САДОВО-ПАРКОВЫХ ТЕРРИТОРИЙ ГОРОДА КАЗАНИ Мударисов Р. Г., Леонова Т. Ш., Мирсаитов Н. Г. Казанский (Приволжский) федеральный университет, г. Казань, raffikk@mail.ru Изучение синантропизации животных имеет важное общебиологическое значение. Деятельность человека – наиболее динамичный фактор, действующий в биосфере. Поэтому проблема приспособления животных к новым условиям, постоянно меняющимся антропогенным условиям затрагивает закономерности макроэволюции, а проблема формирования урбоценозов – проблемы эволюции сообществ. На примере домового и полевого воробьев можно проследить все стадии процессов синатропизации и урбанизации, что придает данным видам свойства уникальной экологической и эволюционной модели (Леонова, 2012). Работы по обследованию живых объектов в различных природных комплексах г. Казани и ее зеленой зоны проводились с ноября 2005 по 2013 гг. Обследовано 3 сада, 11 парков и 6 скверов. 1. «Победы». 2. «ДК Химиков». 3. «Урицкого». 4. «Крылья советов». 5. Сад по улице Тимирязева. 6. «Петрова». 7. Сквер им. Кирова. 8. «Кырлай». 9. Сквер им. Чехова. 10. Сквер по улице К. Тинчурина. 11. «Молодоженов». 12. «Буревестник». 13. ЦПКиО им М. Горького. 14. Сквер у Чеховского рынка. 15. Лядской сад. 16. Ленинский сад. 17. Парк «Черное озеро». 18. Сквер им. Тукая. 19. Парк «Миллениум». Домовый воробей (Passer domesticus) Многочисленный оседлый гнездящийся вид. Один из ведущих доминантных видов. Плотность населения максимальных значений достигает осенью и равняется 290 ос/10 га, а минимальная – 6 ос/10 га – летом. Встречается практически повсеместно во всех садах и парках города. Относительная численность в парках – 30–40 % , в городских скверах – до 60 %. Для гнездования активно используются различные укрытия в строениях, искусственные гнездовья (парки «Урицкого» и «Молодоженов»), дупла деревьев (сквера по ул. Тимирязева, «ДК Химиков» и др.). Также найдено гнездо в разбитом плафоне паркового освещения (Кырлай) (Мударисов, 2011). Это типичная зерноядная птица, любит питаться в мусорных контейнерах, на земле, в период гнездования переходит на питание беспозвоночными. Большей частью это такие беспозвоночные, как насекомые, кузнечики, жужелицы, хрущи и др. Особи данного вида единично не встречаются, встречаются в основном большими скоплениями. 140 Колониальность воробьев наблюдается и в период гнездования. Антропогенное кормовое разнообразие в кормушках не является препятствием, не говоря уже о толерантности к отпугивающим средствам. Диапазон корма расширен. На территории отдельных парков на долю домового воробья приходится до 30–40 % численности птиц (Ленинский сад, сквер им. Кирова). В зимний период воробьи вместе с другими синантропными видами птиц концентрируются вблизи различных источников пищи. Полезная роль воробьев в городе общеизвестна, в Казани наблюдается массовое поедание семян сорных растений многочисленными стаями воробьев. Полевой воробей (Passer montanus) Обычный оседлый гнездящийся, синантропный вид. Максимальная плотность в парках города была отмечена осенью и составляла 372 ос/ 10 га. Летом плотность населения уменьшается, т. к. большая часть птиц покидает город и в поисках более кормных мест перемещается в пригород. При поиске корма в основном перемещаются по газонам, обочинам дорог, реже кормятся на асфальте или на открытой земле. Отмечено, что по сравнению с домовыми воробьями предпочитают участки с более густой и высокой травой. Основным источником питания являются семена маревых, цикория, подорожника, одуванчика, птичьей гречихи, составляющие до 93 % всех потребляемых семян. Корма антропогенного происхождения потребляются значительно реже. Среди поедаемых полевым воробьем насекомых преобладают гусеницы, мелкие бабочки, двукрылые и перепончатокрылые. В зимнее и весеннее время используют городские кормушки, потребляя мелкие фрагменты оставленных там продуктов (Экология, 2005). Полевой воробей играет значительную роль в защите городских зеленых насаждений от вредителей и в большом количестве поедает семена сорных растений (Рахимов, 2012). Полученные количественные данные позволили сделать следующие выводы: 1. Численность полевого воробья превышает на 15–30 % от общего числа домового. 2. Домовый воробей выбирает места для гнездования в условиях городской застройки, а полевой – в естественных условиях парков, скверов, садов, домов частного сектора и лесопосадок. 3. Плотность домового воробья больше в центральных парках («Ленинский сад», сад им. «Лядова» и др.) с наибольшей антропогенной нагрузкой, и наименьшая доля с низкой антропогенной нагрузкой и с наибольшей лесистостью. Для полевого воробья характерны условия лесоопушочнего комплекса. Наибольшая плотность регистрируется в парках ЦПКиО им. М. Горького, парк «Черное озеро» и др. 141 160 140 Общее количество особей 120 100 80 60 40 20 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Садово-парковые территории Зима Весна Лето Осень Рисунок 1 – Сезонное распределение Passer domesticus по садово-парковым территориям г. Казани с 2005–2013 г., ос/ 10 га 800 700 Общее количество особей 600 500 400 300 200 100 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Садово-парковые территории Зима Весна Лето Осень Рисунок 2 – Сезонное распределение Passer montanus по садово-парковым территориям г. Казани с 2005–2013 г., ос/ 10 га 142 1200 1000 Особей/га 800 600 400 200 0 Январь Февраль Март Апрель Май Июнь Июль Август Сентябрь Октябрь Ноябрь Декабрь Месяцы Домовый воробей (Passer domesticus) Полевой воробей (Passer montanus) Рисунок 3 – Сезонное распределение плотности воробьев за 2005–2013гг. Л и т е р а т ур а Леонова Т. Ш. Взаимоотношения синантропных видов птиц в городе Казани (на примере домового и полевого воробьев, серой вороны, грача, галки) / Т. Ш. Леонова, И. И. Рахимов, Р. Г. Мударисов // Врановые птицы в антропогенных и естественных ландшафтах Северной Евразии: 10-ая Международная конференция, 17–21 сентября. 2012. – С. 45–48. Мударисов Р. Г. Формирование видового состава птиц на территории парка «Миллениум» г. Казани / Р. Г. Мударисов // Филология и культура. – 2011. – № 25. – С. 78–81. Рахимов И. И. Систематический обзор птиц парков, садов и скверов города Казани / И. И. Рахимов, Р. Г. Мударисов, А. А. Закиров // Биодиверситиология: Современные проблемы сохранения и изучения биологического разнообразия: сб. материалов II Международной науч.-практ. конф. / под ред. А. В. Дмитриева, Е. А. Синичкина. – Чебоксары: Типография «Новое время» 2010. – 164 с. Рахимов И. И. Экологоповеденческая адаптация воробьев к условиям урбанизированной среды / И. И. Рахимов, Т. Ш. Леонова // Вестник ЧГПУ им. И. Я. Яковлева. – 2012. – № 2(74). – С. 124– 130. Экология города Казани. – Казань: Изд-во «Фэн» Академии наук РТ, 2005. – 576 с. 143 СОСТОЯНИЕ ИНДИКАТОРНОГО КОМПОНЕНТА КОРЕННЫХ ЛЕСНЫХ ФИТОЦЕНОЗОВ КАК ПОКАЗАТЕЛЬ ПРИРОДООХРАННОЙ ЦЕННОСТИ ЭКОСИСТЕМ Нотов А. А. Тверской государственный университет, г. Тверь, anotov@mail.ru В последние годы возрастает интерес к изучению индикаторов биологически ценных лесных фитоценозов (см. Выявление…, 2009). Информация о состоянии такого индикаторного компонента позволяет оценить уровень антропогенной трансформации экосистем и степень их функциональной устойчивости (Выявление…, 2009). В лесных фитоценозах, испытавших разные варианты антропогенного воздействия, существенно уменьшается активность индикаторных видов, а многие виды вообще исчезают. Однако перспективы использования получаемых при этом данных выяснены еще не в полной мере. Необходимы специальные исследования на модельных территориях с разным уровнем активности индикаторных видов биологически ценных лесных сообществ. В качестве таких территорий можно рассматривать ЦентральноЛесной государственный природный биосферный заповедник (ЦЛГПБЗ) и национальный парк «Завидово» (Нотов и др., 2012б). Среди охраняемых территорий Центра Европейской России они представляют особый интерес. Заповедный режим в них установлен на начальных этапах становления нашего государства. Территории их уникальны по многим характеристикам природной среды. ЦЛГПБЗ приурочен к центральной части Каспийско-Балтийского водораздела, имеет площадь 24,4 тыс. га (Истомин, 2008). В заповеднике сохранились фрагменты старовозрастных коренных сообществ южной тайги с характерной структурой и видовым составом (Миняев, Конечная, 1976; Шапошников, 1988; Нотов и др., 2012а). Землепользование на территории водораздела всегда имело экстенсивный характер, который не препятствовал возобновлению зональных лесных сообществ (Носова, 2009). Национальный парк «Завидово» имеет более значительную площадь (125,4 тыс. га) и включает крупные лесные и болотные массивы (Нотов, 2010). На сильно заболоченных водоразделах сохранились участки старовозрастных черноольшаников и интересные переходные болота, окружающие массивы верховых болот. В ЦЛГПБЗ и в национальном парке выявлен высокий уровень видового богатства каждого компонента флоры (Миняев, Конечная, 1976; Ignatov et al., 1998; Нотов, 2010; Нотов и др., 2011, 2012а, 2012б; Потемкин, Нотов, 2012; Конечная, 2012). 144 Антропогенно более изменен и фрагментирован растительный покров национального парка «Завидово» (Нотов, 2010). В этой связи даже в сообществах с участием старовозрастных древесных пород и фрагментов коренных фитоценозов национальный парк характеризуются меньшим числом индикаторных видов, снижением широты их распространения и фитоценотической роли (Нотов и др., 2012б). Анализ индикаторных видов в ЦЛГПБЗ и национальном парке «Завидово» выполнен в 2011–2013 гг. Испльзован подход и методики, разработанные для Северо-Западной России (Выявление…, 2009). Нами определены общий объем и состав индикаторного компонента, роль индикаторных видов мохообразных и лишайников в синузиях и сообществах. Выделены основные комплексы индикаторных видов, связанные с разными флороценотическими группами и типами фитоценозов (Нотов и др., 2012а; 2012б). Созданы ГИС-базы данных на основе системы «Заповедник» (Добриденев, 2005; Нотов и др., 2012а). Оценка встречаемости и сопряженности индикаторных видов в разных фитоценозах позволила выявить характер приуроченности основных комплексов индикаторных видов к определенным древесным породам и микронишам. Часть видов четко связана с фитоценозами, в которых представлены широколиственные породы. Выделяется также группа видов, более обычных в приручьевых и пойменных сероольшаниках и черноольшаниках. Особое внимание уделено видам, связанным с коренными старовозрастными ельниками. Часть индикаторных видов встречается в заболоченных фитоценозах разного состава с высоким уровнем атмосферной влажности (Нотов и др., 2012а). Сопряженный анализ данных по заповеднику и национальному парку позволил выявить существенные различия по характеристикам индикаторного компонента. В сообществах сходного состава, при наличии характерных древесных пород сопоставимого возраста, сходном наборе субстратов и микрониш встречаемость и активность индикаторных видов различны. В национальном парке меньше общий объем индикаторного компонента, ниже средний уровень видового богатства основных синузий, отсутствуют некоторые редкие индикаторные виды лишайников и мохообразных (Нотов и др., 2012б). Более высок по сравнению с национальным парком уровень специфичности состава индикаторных лишайников ЦЛГПБЗ. Они приурочены к фрагментам старовозрастных широколиственно-еловых лесов, особенно к приручьевым и пойменным экотопам с постоянно высоким уровнем атмосферной влажности. Эти виды крайне чувствительны к уровню содержания влаги, сохранности микрониш, характерных для 145 старовозрастных коренных сообществ. Находки многих из них в национальном парке «Завидово» маловероятны. Основные комплексы индикаторных видов, обнаруженные в ЦЛГПБЗ (Нотов и др., 2012б), представлены в пределах национального парка единичными видами, имеющими очень низкое обилие. Не выявлен в национальном парке богатый эпифитный комплекс с Lecanactis abietina (Ach.) Körb, характерный для старовозрастных коренных ельников. Синузии с представителями этого комплекса формируются в ЦЛГПБЗ на комлевых участках стволов старых елей в условиях повышенного затенения и высокой атмосферной влажности (Нотов и др., 2012 а,б). В их составе могут встречаться Arthonia leucopellaea (Ach.) Almq., Cliostomum lepadinum (Ach.) Ach., Thelotrema lepadinum (Ach.) Ach., Schismatomma pericleum (Ach.) Branth et Roster (рис. 1). В национальном парке «Завидово» в эпифитных синузиях в основании стволов старых елей, доминируют широко распространенные виды с высокой активностью. Среди них Cladonia digitata (L.) Hoffm., виды рода Lepraria, Chaenotheca ferruginea (Turner ex Sm.) Mig. Они занимают практически все ниши, которые в старовозрастных коренных сообществах заселяют индикаторные виды. Отсутствуют в национальном парке эпифитные синузии, распространенные в приручьевых и пойменных сообществах ЦЛГПБЗ, включающие Menegazzia terebrata (Hoffm.) A. Massal., Heterodermia spp., Frullania oakesiana Austin, Ulota crispa (Hedw.) Brid. В национальном парке Menegazzia terebrata встречается очень редко в старовозрастных топких черноольшаниках по краю крупных болотных массивов, а сам отмеченный комплекс не выражен (Нотов и др., 2011; 2012б). В фитоценозах с участием широколиственных пород в ЦГПБЗ отмечен высокий уровень богатства индикаторных видов эпифитных базифильных лишайников и мохообразных. Многие из них приурочены к союзу Lobarion. Синузии с Lobaria pulmonaria (L.) Hoffm., встречаются не только на стволах широколиственных пород, но и на старых осинах, реже рябинах и серой ольхе. В верховьях р. Межи в состав синузий c Lobaria pulmonaria входят, как правило, Collema subnigrescens Degel., Leptogium saturninum (Dicks.) Nyl., Nephroma bellum (Spreng.) Tuck., N. laevigatum Ach., N. parile (Ach.) Ach., N. resupinatum (L.) Ach. (рис. 2). Полнота состава синузий зависит от степени участия широколиственных пород или осины в структуре древостоя, преобладающего возраста древесных пород, уровня атмосферной влажности экотопа. 146 Рисунок 1 – Характер распространения Lecanactis abietina (кружки) и некоторых других индикаторных видов лишайников (треугольники) в верховьях реки Межи (ЦЛГПБЗ): Al – Arthonia leucopellaea, Tl – Thelotrema lepadinum, Cl – Cliostomum leprosum, Sp – Schismatomma pericleum 147 Рисунок 2 – Характер распространения Lobaria pulmonaria (кружки) и некоторых других индикаторных видов лишайников (треугольники): Cs – Collema subnigrescens, Ls – Leptogium saturninum, Nb – Nephroma bellum, Nl – N. laevigatum, Np – N. parile, Nr – N. resupinatum Таким образом, большое разнообразие приручьевых экотопов с фрагментами старовозрастных коренных сообществ наряду с большой протяженностью и относительно низким уровнем фрагментации лесных и болотных массивов определяет более значительную фитоценотическую роль индикаторного компонента биологически ценных лесных 148 сообществ в ЦЛГПБЗ по сравнению с национальным парком «Завидово». Даже при наличии фрагментов леса со старовозрастными экземплярами разных древесных пород основные комплексы индикаторных видов в национальном парке сильно трансформированы или утрачены и замещены синузиями более активных эвритопных видов. Сохранение разнообразия комплексов индикаторных видов, которые в данный момент являются еще относительно устойчивыми, определяет особую эталонную ценность лесных массивов ЦЛГПБЗ. Л и т е р а т ур а Выявление и обследование биологически ценных лесов на Северо-Западе Европейской части России: учеб. пособие: в 2 т. – СПб., 2009. – Т. 1. – 238 с.; Т. 2. – 258 с. Добриденев А. И. ГИС – «Заповедник»: справочно-методическое издание / А. И. Добриденев. – М., 2005. – 92 с. Истомин А. В. Мелкие млекопитающие в региональном экологическом мониторинге (на примере Каспийско-Балтийского водораздела) / А. В. Истомин. – Псков: Изд. ПГПУ, 2008. – 278 с. Конечная Г. Ю. Сосудистые растения Центрально-Лесного заповедника (Аннотированный список видов) / Г. Ю. Конечная. – М.: Изд. Комиссии РАН по сохранению биологического разнообразия, 2012. – 75 с. (Флора и фауна заповедников; Вып. 118). Миняев Н. А. Флора Центрально-Лесного государственного заповедника / Н. А. Миняев, Г. Ю. Конечная. – Л.: Наука, 1976. – 104 с. Носова М. Б. Споровопыльцевые диаграммы голоценовых отложений как источник информации об антропогенном воздействии на растительность в доисторический период (на примере ЦентральноЛесного заповедника) / М. Б. Носова // Бюл. МОИП. Отд. биол. – 2009. – Т. 114, вып. 3. – С. 30–36. Нотов А. А. Аннотированный список лихенофлоры Тверской области / А. А. Нотов, Д. Е. Гимельбрант, Г. П. Урбанавичюс. – Тверь: Твер. гос. ун-т, 2011. – 124 с. Нотов А. А. Возможности использования ГИС-технологий для выяснения характера распространения индикаторных видов лишайников и мохообразных / А. А. Нотов, А. Д. Потемкин, Д. Е. Гимельбрант, В. П. Волков, А. В. Павлов // Динамика многолетних процессов в экосистемах ЦЛГПБЗ. – Великие Луки, 2012а. – С. 328–356. (Тр. ЦЛГПБЗ; Вып. 6). Нотов А. А. Индикаторные виды лишайников и мохообразных старовозрастных коренных лесных сообществ как элемент мониторинга экосистем заповедников и национальных парков / А. А. Нотов, А. Д. Потемкин, Д. Е. Гимельбрант, В. П. Волков, А. В. Павлов, В. А. Нотов // Многолетние процессы в природных комплексах заповедников России: материалы Всерос. науч. конф., посвящ. 80-летию Центрально-Лесного государственного природного биосферного заповедника (20–24 авг. 2012 г., пос. Заповедный, Тверская обл.). – Великие Луки, 2012б. – С. 132–139. Нотов А. А. Национальный парк «Завидово»: Сосудистые растения, мохообразные, лишайники / А. А. Нотов. – М.: Деловой мир, 2010. – 432 c. Потемкин А. Д. Некоторые итоги изучения печеночников и антоцеротовых Центрально-Лесного государственного природного биосферного заповедника / А. Д. Потемкин, А. А. Нотов // Динамика многолетних процессов в экосистемах Центрально-Лесного заповедника: материалы Юбилей. конф., посвящ. 80-летию заповедника. – Великие Луки, 2012. – С. 336–344. (Тр. ЦЛГПБЗ; Вып. 6). Шапошников Е. С. Ассоциации еловых лесов ЦЛГЗ / Е. С. Шапошников: дис. … канд. биол. наук – Л., 1988. – 216 с. Ignatov M. S. Bryophyte flora of Zentral'no-Lesnoj Biosphere Nature Reserve (European Russia, Tver Province) / M. S. Ignatov, E. A. Ignatova, E. N. Kurayeva, T. Yu. Minayeva, A. D. Potemkin // Arctoa. – 1998. – T. 7. – P. 45–58. 149 ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ РАСТЕНИЙ БОРЕАЛЬНОЙ ЭКОЛОГО-ЦЕНОТИЧЕСКОЙ ГРУППЫ Полянская Т. А. Национальный парк «Марий Чодра», пос. Красногорский, zamnayki@mail.ru Бореальная эколого-ценотическая группа (ЭЦГ) является основным компонентом растительного покрова лесной зоны на значительной по площади территории и играет особую роль в сохранении типичной структуры коренных лесных сообществ. Современный уровень трансформации бореальных фитоценозов отражается на структуре ЦП видов, входящих в его состав, изменяется строение и функционирование. Поэтому оценка состояния ЦП возможна только в рамках популяционноонтогенетического подхода, наиболее полно отражающего представление о ЦП как надорганизменной структуре. Цель настоящей работы – выявить специфику онтогенетической структуры ЦП модельных бореальных видов разных жизненных форм в различных частях их ареалов в лесной зоне Европейской России. В работе продолжено изучение бореальной эколого-ценотической группы (ЭЦГ) растений, выделенной О. В. Смирновой совместно с Л. Б. Заугольновой (Оценка и сохранение…, 2000). Объектами исследования являются ЦП 30 видов, представляющих разные жизненные формы: 3 вида кустарничков, 3 вида малолетников, 8 видов короткокорневищных растений, 12 – длиннокорневищных, 1 – рыхлодерновинный, 3 – подземно-столоноклубнеобразующих многолетника (табл. 1). Большинство из них имеют обширные циркумбореальные ареалы, произрастают в самых разнообразных экологических условиях. Сбор полевого материала проводился в 1995–2011 гг. в различных частях ареалов бореальных видов растений: в Мурманской, Архангельской, Нижегородской, Московской, Самарской и Челябинской областях, в Республиках: Марий Эл, Чувашия и Башкортостан (табл. 1). Для классификации ЦП применили методы, разработанные Т. А. Работновым (Работнов, 1950), Л. А. Жуковой (Жукова, 1995), А. А. Урановым и О. В. Смирновой (Уранов, Смирнова, 1969), В ходе работы вычислены общепринятые популяционные характеристики: плотность особей на 1 м2, коэффициент возрастности (Уранов, 1975), индексы восстановления и замещения (Жукова, 1985). Для вегетативно-подвижных видов были рассчитаны: специальные индексы восстановления/ и коэффициент возрастности. В результате анализа большого материала (288 ЦП) нами показано, что большинство исследованных ЦП бореальных растений имеют левосторонний базовый спектр. Значительная часть (62,8 %) – молодые, 150 нормальные, неполночленные с максимумами на виргинильной или имматурной группах. 13,8 % всех ЦП – зрелые, нормальные, большую их часть составляют ЦП длиннокорневищных и короткокорневищных растений. Доля участия других типов ЦП незначительна. Регрессивные ЦП отсутствуют (табл. 1). № п/п ЦП ЦП вида инвазионные перспективныне неустойчивые угасающие Всего Таблица 1 – Некоторые характеристики ценопопуляций бореальных видов Кустарнички и п/кустарнички 1. Linnaea borealis L. 1 9 10 2. Vaccinium myrtillus L. 7 17 24 3. Chamaepericlymenum suecicum (L.) Aschers. & Graebn. 1 1 4. Melampyrum pratense L. 1 3 54 5. Melampyrum sylvaticum L. 2 2 4 6. Moehringia trinervia (L.) Clarv. 7. Cicerbita alpina (L.) Wailr. 8. Crepis sibirica L. 9. Geranium sylvaticum L. 10. Omalotheca norvegica (Gunn.) Sch. Bip. & F. Schultz Малолетники 1 5 4 9 Короткокорневищные 3 3 1 1 3 4 5 2 3 6 2 7 3 13 11. Oxalis acetosella L. 9 11 20 12. Solidago virgaurea L. 7 10 17 13. Viola biflora L. 4 14. V. selkirkii Pursh ex Goldie 4 15. Galium triflorum Michx. 4 4 16. Lamium album L. 3 1 17. Maianthemum bifolium (L.) F. W. Shmidt 20 20 18. Moehringia lateriflora (L.) Fenzl. 9 9 4 1 2 7 6 3 17 Длиннокорневищные 151 4 ЦП № п/п ЦП вида инвазионные перспективныне неустойчивые 3 2 3 угасающие Всего Продолжение табл. 1 19. Moneses uniflora (L.) A. Gray 20. Myosotis sylvatica Ehrh. ex Hoffm. 21. Orthilia secunda (L.) House 4 8 12 22. Pyrola media Sw. 8 2 10 5 8 5 10 23. Pyrola minor L. 2 1 6 24. Pyrola rotundifolia L. 9 5 14 25. Saussurea alpina (L.) DC 1 4 26. Viloa epipsila Ledeb. 3 2 2 7 2 Рыхлодерновинные 27. Luzula pilosa (L.) Willd 8 1 9 Подземно-столонообразующие многолетники 28. Adoxa moschatellina L. 29. Circaea alpina L. 1 3 4 30. Trientalis europaea L. 10 8 18 90 157 Всего 9 22 9 19 288 Это определяется, прежде всего, жизненной формой вида, экологическими особенностями, различной историей фитоценозов, занимающих разнообразные экотопы, формирующиеся в разное время и подвергавшихся неодинаковым антропогенным воздействиям. Незначительные колебания демографических параметров связаны с особенностями прохождения онтогенеза в различных экологических условиях Нами предложена новая классификация ЦП, позволяющая выделить угасающие, неустойчивые и перспективные ЦП на основе соотношения подроста и взрослой части ЦП. Критерием их выделения является сравнение доли прегенеративной фракции и взрослой части ЦП, что показывает рассчитываемый индекс замещения (Жукова, 1985; 1995). Анализ популяционной структуры ЦП бореальных видов показал, что их можно подразделить на угасающие, неустойчивые и перспективные по соотношению подроста и взрослой части ЦП, т. е. по индексу замещения. Если индекс замещения равен 0, то такие ЦП следует относить к угасающим. Они составляют 6,6 % от всех ЦП бореальной ЭЦГ. 62,2 % – неустойчивые ЦП, у которых индекс замещения меньше 1, и 31,2 % ЦП – перспективные с индексом замещения более 1 (табл. 2). 152 Таблица 2 – Классификация ценопопуляций бореальных растений по индексу замещения (I з) № п/п Типы ЦП Количество ЦП (шт.) % от общего числа ЦП 1. Угасающие (I з. . = 0) 19 6,6 2. Неустойчивые (Iз. < 1) 179 62,2 3. Перспективные (Iз. . >1) Всего 90 31,2 288 100,0 Угасающие популяции выделены у Сicerbita alpina (3), Omalotheca norvegica (3), Viola selkirkii (2), Galium triflorum (3), Myosotis sylvatica (5), 3 ЦП Saussurea alpina (1) и ЦП Luzula pilosa (1). Это реально доказывает, что лесные сообщества, подвергаясь в настоящее время интенсивному антропогенному воздействию, не могут обеспечить оптимальными условиями обитания ЦП входящих в них бореальных видов. Данное заключение подтверждает и отсутствие регрессивных ЦП, которые должны завершать большую волну популяционных потоков (Уранов, 1975). Для сохранения ЦП многих бореальных видов в природных ЦП требуется широкомасштабный мониторинг, осуществление которого возможно на основе длительного мониторинга ЦП. Л и т е р а т ур а Жукова Л. А. Популяционная жизнь луговых растений / Л. А. Жукова. – Йошкар-Ола: РИИК «Ланар», 1995. Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России / отв. ред. Л. Б. Заугольнова. – М.: Научный мир, 2000. – 196 с. Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах / Т. А. Работнов // Тр. БИН АН СССР. – М.; Л., 1950. – Сер. 3, вып. 6. – С. 77–204. Уранов А. А. Возрастной состав фитоценопопуляций как функции времени и энергетических волновых процессов / А. А. Уранов // Биологические науки. – 1975. – № 2. – С. 17–29. – 224 с. Уранов А. А. Классификация и основные черты развития популяций многолетних растений / А. А. Уранов, О. В. Смирнова // Бюл. МОИП. Отд. биол. – 1969. – Т. 74, вып. 1. – С. 119–134. РАСПРОСТРАНЕНИЕ И ЧИСЛЕННОСТЬ АМФИБИЙ И РЕПТИЛИЙ АНТРОПОГЕННЫХ ЛАНДШАФТОВ ГОРОДА ЙОШКАР-ОЛЫ Свинин А. О., Гаранин В. И. Казанский (Приволжский) федеральный университет, г. Казань, ranaesc@gmail.com Глобальное сокращение биоразнообразия и численности амфибий и рептилий, наблюдаемое в настоящее время, вызывает большие опасения 153 среди исследователей (Blaustein, Wake, 1990; Gibbons et al., 2000; Stuart et al., 2004). Представляется важным изучение и сохранение популяций не только редких, но и массовых видов амфибий и рептилий, обитающих как в естественных, так и в подверженных воздействию антропогенных факторов ландшафтах. Особое внимание уделяется видам, населяющим городскую черту. В Среднем Поволжье были проведены комплексные исследования всех видов амфибий и рептилий, а также отдельных представителей герпетофауны, обитающих в условиях города (Лебединский, Ушаков, 1980; Замалетдинов, Хайрутдинов, 2005; Файзулин, 2012). Тем не менее, данные, относящиеся к городу Йошкар-Оле (северная часть Среднего Поволжья), немногочисленны и отрывочны (Ефремов и др., 1984; Garanin, 2000 и др.), тогда как комплексное исследование герпетофауны до сих пор отсутствует. В связи с этим целью исследования было выявить распространение и численность популяций амфибий и рептилий в черте города ЙошкарОлы. Материал был собран в период с 2007 по 2013 гг. Всего было изучено 10 локалитетов в черте города: 1) микрорайон «Чихайдарово»; 2) микрорайон «Чигашево»; 3) лесопарк «Сосновая роща»; 4) лесопарк «Дубовая роща»; 5) микрорайон «Тарханово»; 6) микрорайон «Заречный»; 7) окрестности МУП «Водоканал», р. Шоя; 8) поселок Знаменский; 9) ул. Березово; 10) водоем по ул. Йывана Кырли. Для оценки численности амфибий был использован метод маршрутных учетов (Динесман, Калецкая, 1952). Ранее в окрестностях и на территории города Йошкар-Олы были отмечены краснобрюхая жерлянка (Garanin, 2000, р. 94), серая жаба (Ефремов и др., 1984, с. 25), озерная (Ведерников, Дробот, 2011, с. 159; Свинин, Забиякин, 2012, с. 28), прудовая (Garanin, 2000, с. 114) и травяная (Ефремов и др., 1984, с. 28) лягушки, остромордая лягушка, прыткая и живородящая ящерицы (Дробот и др., 2008, с. 414). В настоящее время на территории города Йошкар-Олы было отмечено 6 видов амфибий и 3 вида рептилий, что составляет 50 % от региональной герпетофауны. Из земноводных отмечены гребенчатый тритон (Triturus cristatus (Laurenti, 1768)), зеленая жаба (Bufo viridis (Laurenti, 1768)), остромордая лягушка (Rana arvalis (Nilsson, 1842)), травяная лягушка (R. temporaria (Linnaeus, 1758)), озерная лягушка (Pelophylax ridibundus (Pallas, 1771)) и прудовая лягушка (P. lessonae (Camerano, 1882)). Из рептилий были встречены прыткая ящерица (Lacerta agilis (L., 1758)), живородящая ящерица (Zootoca vivipara (Jacquin, 1787)) и обыкновенный уж (Natrix natrix (Linnaeus, 1758)). В окрестностях 154 города помимо перечисленных выше видов также отмечены серая жаба (Bufo bufo (L., 1758)) и съедобная лягушка (Pelophylax esculentus (L., 1758)). Таким образом, из видов герпетофауны, указанных ранее для черты города и его окрестностей, не была найдена лишь краснобрюхая жерлянка (Bombina bombina (L., 1761)), что, возможно, связано с сокращением северо-восточной границы ареала данного вида. Гребенчатый тритон был отмечен нами в небольшом водоеме (165 × 40 м) микрорайона «Чихайдарово» (точка № 1). Отловлен один взрослый самец весной 2010 года, а также зарегистрировано наличие личинок в августе 2011 и 2012 гг. Зеленая жаба в брачный период встречается в небольших постоянных и временных водоемах. Так, в небольших временных водоемах, расположенных за микрорайоном «Тарханово» (точка № 5), в репродуктивный период вид образует большие скопления, состоящие из 50 вокализирующих самцов. Отдельные вокализирующие самцы зарегистрированы нами по ул. Березово (точка № 9). Ранее (в 2003–2004 гг.) в последнем местообитании находился небольшой водоем (около 20 м2), в котором ежегодно размножались зеленые жабы, достигая высокой плотности (до 20 особей) на местах нереста, однако, в настоящее время данное местообитание разрушено. Озерная лягушка заселяет русло реки Малая Кокшага (точки № 3, 4) и расположенные вблизи крупные водоемы («Чихайдарово», «Чигашево», точки № 1, 2, а также Нолькинские пруды по личному сообщению Х. Ф. Балдаева). Прудовая лягушка найдена в двух местообитаниях – небольшие пруды в микрорайоне «Заречный» (точка № 6) и крупный пруд-водохранилище в поселке Знаменский (точка № 8). Гибриды P. esculentus на территории города найдены не были, однако, были встречены в окрестностях поселка Медведево в популяционной системе REL-типа. Озерная лягушка была повсюду многочисленна (от 30 до 100 экземпляров на 100 м береговой линии). Прудовая лягушка в микрорайоне «Заречный» была многочисленна (30 особей в небольшом водоеме 225 площадью м2), тогда как в поселке «Знаменский» было встречено всего 3 особи. Травяная лягушка встретилась нами в лесопарке «Сосновая роща» и в лесопарке «Дубовая роща» по берегу реки Малая Кокшага (точки № 3, 4). Остромордая лягушка отмечена в 4 точках (№ 3–5, 7). Снижение численности бурых лягушек (1–2 экземпляра на 100 м маршрута в 2011 г. против 10–15 особей в 2007 г. в пострепродуктивный период), по всей видимости, связано с засушливым летом и малоснежной зимой 2010 года. Тем не менее, было зарегистрировано восстановление чис155 ленности остромордой лягушки, наблюдавшееся в последний год (около 10 пар особей и 20 кладок в небольшом временном водоеме, локалитет № 5). Прыткая ящерица наиболее многочисленный вид рептилий, составляющий 10–50 особей на 100 м маршрутной линии. Живородящая ящерица предпочитает более влажные биотопы и селится по берегам водоемов. В подходящих биотопах ее численность достигает 10 экземпляров на 100 м маршрута. Обыкновенный уж встречается по берегам крупных рек, прудов, водохранилищ и достигает численности 2–10 особей на 100 м маршрутного учета. Таким образом, наиболее многочисленными амфибиями в городской черте были озерная лягушка и зеленая жаба, наиболее редкими – гребенчатый тритон, прудовая и травяная лягушки. Среди рептилий все три вида характеризовались высокой численностью в подходящих биотопах. Л и т е р а т ур а Ведерников А. А. Некоторые гистологические особенности кишечника и печени лягушки озерной (Rana ridibunda Pallas, 1771), обитающей в условиях урбанизированной среды / А. А. Ведерников, Г. П. Дробот // Актуальные проблемы экологии, биологии и химии: сб. материалов конф. по итогам НИР БХФ за 2010 год /Мар. гос. ун-т. – ЙошкарОла, 2011. – Вып. 2. – С. 159–162. Динесман Л. Г. Методы количественного учета амфибий и рептилий / Л. Г. Динесман, М. Л. Калецкая // Методы учета численности и географического распределения наземных позвоночных. – М.: Высш. шк., 1952. – С. 329–341. Дробот Г. П. Особенности гематологических показателей некоторых видов амфибий и рептилий, обитающих на урбанизированной территории / Г. П. Дробот, Л. И. Губайдуллина, А. Л. Жгулева, А. Ф. Насибуддина, Н. В. Летунова // Принципы и способы сохранения биоразнообразия: материалы III Всерос. науч. конф. – Йошкар-Ола; Пущино, 2008. – C. 414–415. Ефремов П. Г. Животный мир Марийской АССР. Наземные позвоночные. Земноводные, пресмыкающиеся, млекопитающие / П. Г. Ефремов, В. А. Корнеев, Ю. Н. Русов. – Йошкар-Ола: Маркнигиздат, 1984. – 128 с. Замалетдинов Р. И. Земноводные и пресмыкающиеся / Р. И. Замалетдинов, И. З. Хайрутдинов // Экология города Казани. – Казань: Изд-во «Фэн» Академии наук РТ, 2005. – С. 191–203. Лебединский А. А. К изучению амфибий урбанизированной территории в связи с проблемой их охраны / А. А. Лебединский, В. А. Ушаков // Комплексное изучение и рациональное использование природных ресурсов. – Калинин, 1980. – С. 180–181. Свинин А. О. Морфологическая характеристика популяций озерной лягушки (Pelophylax ridibundus (Pallas, 1771)), обитающей в черте г. Йошкар-Олы / А. О. Свинин, В. А. Забиякин // Актуальные проблемы экологии, биологии и химии: материалы конф. по итогам НИР БХФ за 2011 г. / Мар. гос. унт. – Йошкар-Ола, 2012. – Вып. 3. – С. 28–31. Файзулин А. И. Земноводные как биоиндикаторы состояния окружающей среды в условиях Среднего Поволжья: половозрастная структура популяций / А. И. Файзулин // Известия Самарского научного центра Российской академии наук // Спецвыпуск «Безопасность. Технологии. Управление». – 2008. – Вып. 9. – С. 271–274. Blaustein A. R. Declining amphibian populations – a global phenomenon / A. R. Blaustein, D. B. Wake // Trends in Ecology and Evolution. – 1990. – Vol. 5. – P. 203– 204. Gibbons J. W. The Global Decline of Reptiles, Déjà Vu Amphibians / W. J. Gibbons, D. E. Scott, T. J. Ryan, K. A. Buhlmann, T. D. Turberville, B. S. Metts, J. L. Greene, T. Mills, Y. Leiden, S. Poppy and C. T. Winne // BioScience. – 2000. – Vol. 50. – P. 653–666. 156 Stuart S. Status and trends of amphibian declines and extinctions worldwide / S. Stuart, J. S. Chanson, N. A. Cox, B. E. Young, A. S. L. Rodrigues, D. L. Fishman, R. W. Waller // Science. – 2004. – Vol. 306. – P. 1783–1786. СТРУКТУРНОЕ РАЗНООБРАЗИЕ ЭКОСИСТЕМ ПРОЕКТИРУЕМОЙ ЗОНЫ ЗАТОПЛЕНИЯ ЧЕБОКСАРСКОЙ ГЭС (ДО 68 ОТМЕТКИ) Сидоренко М. В., Юнина В. П. Нижегородский государственный университет, г. Нижний Новгород, eco_smv@mail.ru В течение вегетационного периода 2012 г. авторами были проведены исследования растительности проектируемой зоны затопления Чебоксарского водохранилища (ЗЗЧВ) с НПУ 68,0 м. По общепринятым методам физико-географических исследований (Жучкова, 2004) определялись природные условия в ЗЗЧВ, включая рельеф, почвенный покров (степень преобразования и загрязнения). Оценивалось состояние растительного покрова обследуемых участков на пробных площадях и на маршрутах. Описание лесных растительных сообществ проводилось с использованием методов лесной таксации и геоботанических исследований (Анучин 1981; Ипатов, Мирин, 2008). Определялись условия местопроизрастания, характеризовались все ярусы лесного сообщества: древостой, подрост и подлесок, травяно-кустарничковый ярус, живой напочвенный покров. На основании результатов проведенных исследований была составлена легенда карты растительности, в которую вошли все типы растительных ассоциаций, включая коренные, условно-коренные и производные растительные сообщества ЗЗЧВ. С использованием разработанной для ЗЗЧВ легенды, материалов полевых исследований, имеющейся планово-картографической основы, космических- и аэрофотоснимков составлена карта растительности масштаба 1 : 100 000. Это позволило в полной мере охарактеризовать наземную и прибрежно-водную растительность ЗЗЧВ. В основе легенды карты отражено типологическое разнообразие растительных сообществ в районе исследований и связь с ландшафтной структурой территории, учтены пространственно-ботанические закономерности распределения растительности и ее антропогенная динамика. В этих целях в легенде отражена типологическая дифференциация растительного покрова. На карте показаны основные типологические категории (лесные сообщества – формации и типы), луговые, луговоболотные и кустарниковые сообщества (ивняки). Лесные сообщества приведены с указанием коренных и условно-коренных (сосняки, ельни157 ки, дубравы, ольшаники и др.), а также производных (березняки, осинники и др.). В классификации растительности отражены основные зональные особенности растительного покрова, отражающие специфику района исследований, включающего в себя самые разнообразные лесорастительные зоны и подзоны, а также ботанические районы и подрайоны: от степных участков, широколиственных и смешанных лесов до южнотаежных растительных сообществ. Следует отметить, что условия произрастания пойменной растительности существенно сглаживают зональные особенности района исследований. Таким образом, составленная карта растительности зоны влияния Чебоксарского водохранилища с НПУ 68,0 м отражает как динамические характеристики и смены растительных сообществ (производная растительность), так и природные условия произрастания, т. е. коренные типы растительности. Наземные экосистемы в ЗЗЧВ характеризуются значительным структурным разнообразием. Среди лесных экосистем широко представлены хвойные бореальные леса (с участием бореальных элементов) – еловые (преобладающая порода ель – Picea abies (L.) Karst.) и сосново-еловые (Pinus sylvestris L., Picea abies). К ним относятся коренные типы – еловые и сосново-еловые зеленомошные, елово сфагноводолгомошные, еловые травяно-болотные, а также их производные с преобладанием осины (Populus tremula L.), березы повислой (Betula pendula L.) и ольхи черной (Alnus glutinosa (L.) Gaertn.) (всего 4 коренных и 4 производных сообщества). Сосновые леса представлены сосновыми кустраничково-зеленомошными, сфагново-долгомошными и сфагновыми, а также прозводными насаждениями из осины и березы повислой (всего 3 типа коренных и 3 типа производных лесов). Елово-сосновые леса встречаются, по сравнению с предыдущими группами лесов, на более бедных по трофности местообитаниях, занимают небольшие возвышенности и склоны водоразделов, надпойменных террас и участков высокой поймы. Среди них отмечено 4 типа коренных и 6 типов производных лесов. Хвойные субнеморальные леса характеризуются участием как бореальных (кислица обыкновенная (Oxalis acetosella L.), майник двулистный (Majanthemum bifolium (L.) F. M. Schmidt), черника (Vaccinium myrtillus L.), брусника (Vaccinium vitis-idaea L.) и др.), так и неморальных видов: сныть обыкновенная (Aegopodium podagraria L.), копытень европейский (Asarum europaeum L.), звездчатка ланцетолистная (Stellaria holostea L.) и др. Еловые леса здесь представлены еловыми кислично-широкотравными (1 группа коренных и 2 – производных типа). Сосновые леса включают сосново кислично-широкотравные (1 группа коренных и 2 – производных типа). 158 Очень широко представлены в ЗЗЧВ широколиственно-хвойные и хвойно-широколиственные леса. В данных типах лесных экосистем наряду с хвойными и мелколиственными породами в состав древостоя могут входить или доминировать широколиственные породы (Quercus robur L., Acer platanoides L., Ulmus laevis Pall.). В зависимости от условий местопроизрастания данные лесные сообщества могут сильно различаться по составу древесного яруса, подлеска и травянокустарничкого яруса. Так же широко представлены и производные модификации коренных типов леса, с преобладанием березы, осины, реже других пород. Данные группы лесов занимают повышенные местообитания (небольшие повышения и пологие склоны водоразделов и надпойменных террас, редко участки высокой поймы) и произрастают на достаточно плодородных подзолистых почвах, легко- и среднесуглинистых, иногда супесчаных или связнопесчаных, подстилаемых суглинками. Данная группа лесов включает: еловые с дубом (дубово-еловые широкотравные – 1 коренной и 2 производных типа), еловые с липой (липово-еловые широкотравные – 2 коренных и 4 производных типа), сосновые с дубом (дубово-сосновые широкотравные – 1 коренной и 3 производных типа), сосновые с липой (липово-сосновые широкотравные – 2 коренных и 5 производных типов), дубовые с елью (еловодубовые широкотравные – 1 коренной и 2 производных типа). Широколиственные леса занимают в ЗЗЧВ повышенные местообитания (небольшие повышения и пологие склоны водоразделов и надпойменных террас, участки высокой поймы). Почвы достаточно плодородные, серые лесные, легко- и среднесуглинистые, иногда супесчаные, подстилаемые суглинками. В ЗЗЧВ распространены дубовые, липовые, липово-дубовые с кленом и вязом леса: липово-дубовые с кленом, ясенем и вязом кустарниковые широкотравные (1 – коренной и 9 – производных типов), липово-дубовые широкотравные (приручьевые) (1 – коренной и 5 – производных типов), дубовые широкотравновлажнотравные (пойменные) (1 – коренной и 9 – производных типов) Мелколиственные леса представлены в ЗЗЧВ черноольховыми (1 – коренной и 1 – производный типы) – занимают заболоченные участки пойм и окраины болот со слабым стоком воды, а также пушистоберезовыми (Betula pubescens Ehrh.) – занимают заболоченные участки пойм и окраины болот, западины (1 – коренной тип, 1 – производный, 1 – антропогенная модификация). Ивовые леса (Salix fragilis L., S. alba L., S. caprea L.) широко представлены в ЗЗЧВ (1 коренной и 2 – производных типа), занимают поймы рек и речек. Почвы аллювиальные, разного механического состава, чаще песчаные. Подлесок слагают: шиповник, кустарниковые виды ив 159 (Salix cinerea L., S. triandra L., S. viminalis L., S. myrsinifolia Salisb, S. acutifolia Willd. и др.). Часто ивняки представлены только кустарниковыми сообществами (пионерные сообщества на песчаных отмелях и наносах рек). Луга и травяные болота являются в основном производной антропогенной модификацией растительных сообществ поймы, образованной на месте пойменной древесно-кустарниковой растительности, преобразованной в сенокосы или пастбища, реже – пашни. По типу местообитаний выделяются свежие и влажные (преобладают ксеромезофиты и мезофиты) злаково-разнотравные пойменные луга (сенокосы и пастбища), а также сырые пойменные луга и травяные болота (камышовые, манниковые, остроосоковые, щучково-разнотравные, двукисточниковые). Таким образом, при составлении карты растительности ЗЗЧВ было выделено 31 типологическая единица коренных растительных сообществ и антропогенных модификаций, а также 60 производных. Наибольшую долю по площади ЗЗЧВ имеют пойменные луга разных типов и лесные экосистемы. В случае реализации проекта подъема уровня Чебоксарского водохранилища до проектной отметки 68,0 м будет оказано негативное воздействие на природные комплексы ЗЗЧВ и прилегающие к ней территории: прогнозируется резкое снижение биологического разнообразия (видового, ландшафтного и наземных экосистем) пойменных сообществ Нижегородского Поволжья, ожидается почти полное исчезновение пойменных дубрав р. Волги (в ЗЗЧВ находятся последние участки этих типов растительности волжской поймы), трансформация наземных и прибрежно-водных экосистем (от полного затопления до заболачивания прилегающих территорий), исчезнет ряд пунктов произрастания охраняемых видов растений, в т. ч. единственного места произрастания в Нижегородской области для одного вида – Jovibarba globifera (L.) J. Parnell, произойдет негативное воздействие на ряд ООПТ Нижегородской области (от полного и частичного затопления до заболачивания) – более 20-ти ООПТ. Л и т е р а т ур а Анучин Н. П. Лесная таксация / Н. П. Анучин. – М., 1981. – 552 с. Жучкова В. К. Методы комплексных физико-географических исследований: учеб. пособие для студ. вузов / В. К. Жучкова. – М.: Изд. центр «Академия», 2004. – 368 с. Ипатов В. С. Описание фитоценоза. Методические рекомендации: учеб.-метод. пособие / В. С. Ипатов, Д. М. Мирин. – СПб., 2008. – 71 с. 160 ТАКСОНОМИЧЕСКОЕ РАЗНООБРАЗИЕ ОРХИДНЫХ НА ОСОБО ОХРАНЯЕМЫХ ТЕРРИТОРИЯХ И В ВОДООХРАННЫХ ЗОНАХ НИЖЕГОРОДСКОЙ ОБЛАСТИ Сидоренко М. В. Нижегородский государственный университет, г. Нижний Новгород, eco_smv@mail.ru В результате проведенных автором в течение 10 лет исследований состояния популяций охраняемых видов семейства Орхидных (Orchidaceae Juss.) на особо охраняемых природных территориях (ООПТ) и в водоохранных зонах Нижегородской области, а также в проектируемой зоне затопления Чебоксарской ГЭС (повышение уровня с 63 до 68 отметки) выявлены новые местообитания редких видов, установлено их внутривидовое разнообразие. Полученные данные могут быть использованы для организации новых ООПТ, разработке мер по сохранению ценопопуляций орхидных в природе, а также для интродукции видов в культуру с целью сохранения генофонда. В наибольшей степени исследованиям подверглись ООПТ регионального значения, расположенные в г. Н. Новгороде: памятники природы «Малиновая гряда», «Щелоковский хутор», «Слуда». Кроме того, обследовались следующие ООПТ: «Оленина гора» в Лысковском районе, «Балахонинская пещера» в Ардатовском районе, «Территория Затон «Окский» – д. Оленино» в Богородском районе, а также заказник «Пустынский» в Арзамасском районе. Впервые выявлено произрастание ряда охраняемых видов – любки зеленоцветковой (Platanthera chlorantha (Cust.) Reichenb.) и ятрышника шлемоносного (Orchis militaris L.) – на территории памятника природы «Малиновая гряда» в г. Н. Новгороде, ятрышника шлемоносного – на территории памятника природы «Территория Затон «Окский» – д. Оленино» в Богородском районе, башмачка настоящего (Cypripedium calceolus L.) и ятрышника шлемоносного – в окрестностях с. Хабарское. Обследованы популяции (несколько участков) редкого охраняемого вида, занесенного в Красную книгу РФ и Красную книгу Нижегородской области – ятрышника шлемоносного по коренным склонам р. Оки в окрестностях г. Павлово и кокушника длиннорогого (Gymnadenia conopsea (L.) R. Br.). За период исследований было установлено несколько новых мест произрастания пальчатокоренника Траунштейнера (Dactylorhiza traunsteineri (Saut.) Soo): в окрестностях пос. Желнино, в водоохраной зоне р. Пыра (Балахнинский район), в водоохранных лесах р. Ваи (Уренский район). На территории ООПТ «Семиречье» (Балахнинский район) был выявлен новый для области вид – пальчатокорен161 ник Мейера (Dactylorhiza meyeri (Reichenb. fil.) Aver., или D. hebridensis (Wilmott) Aver.), отличающийся по некоторым морфологическим признакам от пальчатокоренника Фукса, к которому его часто относят (Dactylorhiza fuchsii var. meyeri (Reichenb. fil.) Soo) (Маевский, 2006). В некоторых обследованных местообитаниях выявлены белоцветковые формы следующих видов орхидных: пальчатокоренников Фукса (Dactylorhiza fuchsii (Druce) Soo), пятнистого (Dactylorhiza maculata (L.) Soo.) и Траунштейнера; ятрышника шлемоносного; кокушника длиннорогого. В 2005 году автором было установлено произрастание на территории ООПТ «Щелоковский хутор» (г. Нижний Новгород) нового для Нижегородской области вида – пальчатокоренника балтийского (Dactylorhiza baltica (Klinge) Orlova). Впоследствии данный вид был выявлен автором также на территории ООПТ «Слуда» и «Малиновая гряда» (здесь, возможно, произрастает гибрид пальчатокоренников Фукса и балтийского), а также в окрестностях г. Павлово. Для всех обнаруженных на территории Нижегородской области ценопопуляций пальчатокоренника балтийского характерно небольшое число растений, а также приуроченность к выходам грунтовых вод, произрастание в открытых (луга) или полузакрытых местообитаниях (опушки леса). Учитывая небольшое число выявленных ценопопуляций и их малочисленность, пальчатокоренник балтийский необходимо включить в Красную книгу Нижегородской области и разработать меры по его охране. Важной мерой по сохранению генофонда редких и охраняемых видов орхидных является введение в культуру, культивирование растений для получения семенного материала. Имеются успешные попытки интродукции и получение посадочного материала на базе ботанических садов в г. Нижнем Новгороде, Москве и других регионах. Накоплен опыт (как отечественный, так и зарубежный) по содействию процессов опыления дикорастущих растений и посеву семян в подготовленную почву, по другим агротехническим приемам, улучшающим процессы возобновления популяций. Данные по характеристике ценопопуляций орхидных Нижегородской области, полученные автором, могут быть использованы для интродукции в культуру наиболее декоративных форм, в т. ч. белоцветковых и гибридных растений. Л и т е р а т ур а Маевский П. Ф. Флора средней полосы европейской части России / П. Ф. Маевский. – 10-е изд. – М.: Товарищество научных изданий КМК, 2006. – 600 с. 162 ВАЛЕЖНЫЙ КОМПЛЕКС КАК ФАКТОР СОХРАНЕНИЯ БИОРАЗНООБРАЗИЯ НА ГОРОДСКИХ ООПТ НИЖНЕГО НОВГОРОДА Темнухин В. Б. Нижегородский государственный архитектурно-строительный университет, г. Нижний Новгород, temnuhin@rambler.ru Экологическая значимость валежа для лесных экосистем несомненна, причем во многом она обусловлена сохранением биоразнообразия флоры и фауны, необходимой и достаточной устойчивости экосистемы (Стороженко, 1991). Казалось бы, этот давно доказанный факт должен быть непременно использован при проектировании и эксплуатации ООПТ, особенно городских. Однако, анализ официальных сведений, приведенных об ООПТ Нижнего Новгорода (Бакка, Киселёва, 2008) указывает на прямо противоположное. Бесспорно, это приводит к существенным потерям в биоразнообразии, особенно тех видов, которые трофически либо топически связаны с древесными остатками, что в перспективе не может не сказываться на состоянии ООПТ. В частности, значительная часть редких видов грибов-ксилотрофов обитает на крупномерных древесных остатках разной степени разложения, образующихся при гибели деревьев предельного возраста для данных условий роста. Так, например, все установленные нами в нынешнем году местообитания чешуйницы древесинной Lepiota lignicola Karst., занесенной в Красную книгу СССР (Фёдоров, 1990), связаны с крупномерными старовозрастными деревьями березы, липы и лиственницы, произрастающими в Нижнем Новгороде, а также их древесными остатками. Таким образом, количество и качество древесных остатков должно учитываться при проектировании ООПТ в городских насаждениях, а для рациональной эксплуатации таких ООПТ необходимо ввести нормы оставления и/или сохранения древесных остатков разного качества. В противном случае неизбежны деградация и распад экосистемы. Л и т е р а т ур а Бакка С. В. Особо охраняемые природные территории Нижегородской области: аннотированный перечень / С. В. Бакка, Н. Ю. Киселёва. – Н. Новгород, 2008. – 560 с. Стороженко В. Г. Структура грибных сообществ лесных биогеоценозов / В. Г. Стороженко // Научные основы устойчивости лесов к дереворазрушающим грибам / под ред. С. Э. Вомперского. – М.: Наука, 1992. – С. 21–31. Фёдоров Ф. В. Грибы / Ф. В. Фёдоров. –3-е изд., перераб. и доп. – М.: Росагропромиздат, 1990. – 366 с. 163 РАЗНООБРАЗИЕ ЧЛЕНИСТОНОГИХ КОНСОРТОВ НЕКОТОРЫХ ЛИСТВЕННЫХ ДЕРЕВЬЕВ В УСЛОВИЯХ УРБАНИЗИРОВАННОЙ СРЕДЫ Турмухаметова Н. В. Марийский государственный университет, г. Йошкар-Ола, bonid@mail.ru С ценопопуляцией автотрофного растения постоянно связаны прямыми и обратными биоценотическими отношениями разнообразные организмы, образующие совместно с ним консорцию (Хмелев и др., 2000). Одними из древесных видов, широко распространенными в Республике Марий Эл и часто используемыми в озеленении г. ЙошкарОлы, являются береза повислая (Betula pendula Roth) и липа сердцевидная (Tilia cordata Mill.). Консортивные связи B. pendula и T. cordata с различными организмами достаточно разнообразны. Учеными постоянно проводится изучение вредителей древесных растений в условиях лесов (Гусев, 1984). Однако городская среда является по различным параметрам достаточно специфической как для растений, так и для беспозвоночных животных. Известно, что загрязняющие вещества влияют на растительноядных насекомых как непосредственно, так и опосредованно – через кормовые растения. Воздействие промышленных и иных выбросов на растения изменяет в свою очередь видовой состав насекомых-фитофагов (Тарасова и др., 2004). Цель данной работы – описание разнообразия насекомых и паукообразных, обитающих в кронах березы повислой (Betula pendula Roth) и липы сердцевидной (Tilia cordata Mill.) в различных экологических условиях г. Йошкар-Олы. Исследование проводили в 2009–2012 годах в районах города, различающихся степенью загрязнения среды промышленно-транспортными выбросами: зоны наименьшего (ООПТ «Сосновая роща»), слабого (селитебная зона) и среднего загрязнения (промышленно-транспортная зона). Изучались деревья средневозрастного онтогенетического состояния. Насекомых и паукообразных собирали в нижней части кроны деревьев и на стволах вручную, методами встряхивания, использования стволовых ловушек (Хмелев и др., 2000; Цуриков, 2004); по характерным повреждениям вегетативных и генеративных структур деревьев определяли некоторых насекомых или клещей-фитофагов (Гусев, 1984); рассчитывали относительное обилие дендробионтов (Песенко, 1982). В изученных насаждениях березы повислой обнаружены особи 84 видов насекомых и паукообразных, в насаждениях липы сердцевидной – 100 видов. Большее разнообразие насекомых-дендробионтов в консорции T. cordata обусловлено энтомофильностью особей вида. Структура 164 консорции изученных лиственных деревьев включает три концентра: первый концентр представлен фитофагами – растительноядными насекомыми и клещами, второй – энтомозоофагами. В смешанные 1, 2 концентры входят пантофаги, во 2, 3 – пауки, являющиеся зоофагами первого и второго порядков. Выявлены трофические (биотрофические, эккрисотрофические, сапротрофические), топические, форические, фензивные и фабрические связи членистоногих животных с детерминантами консорций. Среди фитофагов B. pendula и T. cordata доминируют Coleoptera. В кронах B. pendula обнаружены жуки семейств Curculionidae, Apionidae, Attelabidae, Chrysomellidae, Elateridae, Throscidae; к этому перечню для T. cordata добавляются Mordellidae, Staphylinidae. Виды семейств Dascillidae, Nitidulidae, Byturidae ранее не были описаны как фитофаги T. cordata. Сосущими листья детерминантов консорции являются представители отряда Homoptera – Aphidoidae, Aphrophoridae, Cicadellidae, Psyllidae, Membracidae, Fulgoridae; представители Hemiptera – Acanthosomatidae, Lygaeidae, Pentatomidae, Miridae. Среди энтомозоофагов преобладают Coccinellidae (Coleoptera) и Aranei. К консортам смешанного порядка относятся Pyrrhocoris apterus (Linnaeus, 1758), Pentatoma rufipes (Linnaeus, 1758), Panorpa communis (Linnaeus, 1758), Formica rufa L., Formica truncorum Fabr., Vespula vulgaris (Linnaeus, 1758) и другие. В кронах B. pendula и T. cordata встречаются и сходные виды; индекс Чекановского-Съеренсена составляет 0,35. Так, например, типичные вредители B. pendula – Betulapion simile (Kirby, 1811), Kleidocerys resedae (Panzer, 1797) и Byctiscus betulae (Linnaeus, 1758), обнаружены и на T. cordata. Большинство обнаруженных насекомых не входят в списки основных вредителей B. pendula и T. cordata (Гусев, 1984), что возможно, связано с недостаточной изученностью консорции данных детерминантов в нашем регионе. В изученных насаждениях B. pendula преобладающими трофическими группами являются филлофаги – листогрызущие и сосущие листья насекомые, а также карпофаги, в насаждениях T. cordata – еще и антофилы. В условиях среднего загрязнения среды в многочисленных разрывах корки B. pendula увеличивается обилие видов-эккрисотрофов и отмечены повреждения опасного стволового вредителя Scolytus ratzeburgi (Janson, 1856), что может свидетельствовать об ослаблении деревьев вследствие ухудшения качества среды обитания. В консорциях B. pendula и T. cordata преобладают полифаги. Доля монофагов Aphidoidae (Homoptera), Cecidomyiidae (Diptera) и Eriophyidae (Acarina) 165 увеличивается в условиях среднего загрязнения среды промышленнотранспортными выбросами. При разной степени антропогенной нагрузки изменяются консортивные связи с видом-детерминантом. В условиях среднего загрязнения среды промышленно-транспортными выбросами отмечается уменьшение видового разнообразие консортов 1-го и 2-го концентров B. pendula и T. cordata, при этом увеличивается относительное обилие наиболее массовых вредителей листьев березы повислой и липы сердцевидной: тлей, клещей и галлиц до 4–5 баллов (P < 0,05). Однако одни зоофаги (божьи коровки и златоглазки) здесь же представлены с высоким обилием, другие (клопы и паукообразные) – с низким. Хищные членистоногие контролируют численность филло- и карпофагов деревьев. Изменчивость консортивных связей с детерминантом популяционной консорции можно рассматривать как функциональную поливариантность (Жукова, 2006) средневозрастных генеративных особей B. pendula и T. cordata. Разнообразие консортивных связей с древесным растением может свидетельствовать о степени устойчивости его популяции к антропогенной нагрузке. Полученные результаты согласуются с данными фитоиндикационного исследования, проведенного в этих же пунктах (Турмухаметова, 2005). Так, по различным морфометрическим и фенологическим признакам береза повислая является более газоустойчивой к промышленнотранспортным выбросам среды, чем липа сердцевидная. Причем, у B. pendula в условиях среднего загрязнения среды отмечается стимулирование ростовых процессов побегов и степени плодоношения, что, скорее всего, привлекает наиболее устойчивых филло- и карпофагов. У T. cordata в неблагоприятных городских условиях также отмечается активация функционирования репродуктивной сферы при подавлении вегетативной, что привлекает большее разнообразие антофилов и эккрисотрофов. В насаждениях изученных лиственных деревьев в селитебной и промышленной зонах, по сравнению с насаждениями ООПТ, увеличивается разнообразие и обилие членистоногих (клещей, тлей, личинок галлиц и молей), трофически не связанных с поверхностными тканями растений и формирующих защитные приспособления и уменьшается разнообразие открытоживущих фитофагов. Таким образом, исследуя видовой состав и относительное обилие насекомых и паукообразных в естественных условиях и городских насаждениях деревьев, можно судить о качестве среды обитания, степени загрязнения их теми или иными вредными веществами. 166 Л и т е р а т ур а Гусев В. И. Определитель повреждений лесных, декоративных и плодовых деревьев и кустарников / В. И. Гусев. – М.: Лесная пром-ть, 1984. – 472 с. Жукова Л. А. История развития популяционно-онтогенетического направления в России и его перспективы / Л. А. Жукова // Поливариантность развития организмов, популяций и сообществ: научное издание / Мар. гос. ун-т. – Йошкар-Ола, 2006. – С. 7–32. Песенко Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях / Ю. А. Песенко. – М.: Наука, 1982. – 287 с. Тарасова О. В. Насекомые-филлофаги зеленых насаждений городов: Видовой состав и особенности динамики численности / О. В. Тарасова, А. В. Ковалев, В. Г. Суховольский, Р. Г. Хлебопрос. – Новосибирск: Наука, 2004. – 180 с. Турмухаметова Н. В. Особенности морфогенеза побегов и феноритмов Betula pendula Roth. и Tilia cordata Mill. в условиях городской среды: автореф. дис. … канд. биол. наук / Н. В. Турмухаметова. – Новосибирск, 2005. – 19 с. Хмелев К. Ф. Методы изучения популяций и их консортивных связей в природных и антропогенно-трансформированных экосистемах / К. Ф. Хмелев, А. А. Афанасьев, А. И. Кирик, В. В. Негробов, В. В. Онищенко // Экологический мониторинг. Методы биологического и физико-химического мониторинга: учеб. пособие / под ред. Д. Б. Гелашвили. – Н. Новгород: Изд-во ННГУ, 2000. – Ч. 4. – С. 220–258. Цуриков М. Н. Гуманные методы исследования беспозвоночных / М. Н. Цуриков // Запов. справа в Украiнi. – 2004. – Т. 9, вып. 2. – С. 52–57. К ВОПРОСУ О ФЛОРИСТИЧЕСКОМ РАЗНООБРАЗИИ РАЙОНА ПЯТИГОРЬЯ, КАК ЧАСТИ ОСОБО ОХРАНЯЕМОГО ЭКОЛОГО-КУРОРТНОГО РЕГИОНА РФ «КАВКАЗСКИЕ МИНЕРАЛЬНЫЕ ВОДЫ» Утенкова С. Н. Государственный университет управления, г. Москва, lana_utenkova@mail.ru Район Пятигорья охватывает возвышенно-горно-островную часть региона Кавказских Минеральных Вод, находящегося на юге Ставропольского края. Территории Кавказский Минеральных Вод в 1992 году Указом Президента РФ (от 27.03.1992 г. № 309) и решением Правительства РФ (6.06.1992 г. № 462) придан статус «Особо охраняемого эколого-курортного региона РФ». В геоморфологическом отношении территория представлена системой предгорных наклонных террасированных равнин и весьма своеобразным проявлением в интрузивной форме новейшего вулканизма, получившего в литературе название лакколитов, криптолакколиты и дайков (Чупахин, 1974). Довольно сложная архитектоника, а также положение района на стыке равнинной и горной территории способствовало тому, что на настоящий момент, на сравнительно незначительной по площади территории (≈1630 км2), образовалась весьма оригинальная и своеобразная флора, вопрос об изучении которой имеет особую значимость, т. к. регион является уникальным и одним из богатейших во флористическом отношении на территории Предкав167 казья. Выявление видов, нуждающихся в охране, определение генофонда полезных видов с проведением всестороннего флористического анализа, частью которого является экологический и географический анализы) и установлением специфических особенностей флоры является важной основой для прогнозирования развития региона и тенденции изменения экологической обстановки. В исследуемой флоре подавляющее большинство видов составляют покрытосеменные растения (Magnoliophyta) – 1567 видов (98,18 % от общего числа видов флоры), среди которых преобладают двудольные (Magnoliopsida) – 1247 вида (78,13 %). В целом, в систематическом отношении флора Пятигорья характеризуется следующими чертами: достаточно высокой степенью гетерогенности; высокой видовой насыщенностью; большим процентом участия крупнейших и крупных семейств; высоким процентом участия семейств, насчитывающих один вид; значительным участием крупных (полиморфных), средних и бедных родов; большим количеством родов, представленных одним видом. Все это, прежде всего, говорит о специфичности флоры и, как следствие этого, ее богатстве и оригинальности. На исследуемой территории нами выделяются семь флороценотипов: лесной, луговой, степной, полупустынный, пустынный, водный и сорный. Флороценотипы образованы флороценоэлементами, объединяемыми в ряде случаев во флороценосвиты. Таких флороценоэлементов выделяется 16: лесной, равнинно-луговой, субальпийский, альпийский, степной, полупустынный, кальцепетрофильный, псаммофильный, галофильный, аргиллофильный, гигрофильный, гидрофильный, гидатофильный, сегетальный и рудеральный. Помимо ценотипно верных флороценоэлементов имеется множество переходных, которые образуются, во-первых, при взаимном контактировании различных сообществ, а вовторых, из-за широкой экологической амплитуды элементов флоры. Наибольшее разнообразие видов наблюдается среди луговой растительности; на втором месте по числу видов стоит степной комплекс, а за ним – лесной, что еще раз указывает на особое положение исследуемой территории, как места где происходит смешение кавказской (лесной и луговой флоры) и южнорусской (степной). В изучаемой флоре насчитывается 1120 (70,1 %) ценотипно верных видов, обладающих строгой приуроченностью к определенному фитоценозу. Процент перекрытия составляет 29,8 %, т. е. около трети видов флоры экологически пластичны, не обладают строгой приуроченностью к определенному ценозу, одной фитоценоэкологической нише, а могут встречаться в двух, трех, а иногда и четырех различных местообитаниях. В систематическом плане в большинстве флороценотипов по числу флороценоэлементов первое 168 место принадлежит семейству Asteraceae, только в полупустынном на первое место выходит семейство Caryophyllaceae и в водном – Cyperaceae. Географическая структура флоры также заслуживает к себе внимания. Главенствующее место занимают бореальные геоэлементы (система географических элементов флоры Н. Н. Портениера (1993), дополненная А. Л. Ивановым (1998)) – 627 видов (39,3 %), среди которых, в свою очередь, преобладают кавказские (12,6 % – от общего числа видов флоры и 32,1 % – от количества видов в группе бореальных элементов). Значительным количеством (111 видов, 7,0 и 17,7 %) представлен европейский геоэлемент, а также евро-сибирский (71 вид, 4,5 и 11,3 %) и евро-кавказский (71 вид, 4,5 и 11,3 %). Понтические и понтическоюжносибирские виды также имеют достаточно большой процент участия: понтические – 64 вида (4,0 и 10,2 %), понтическо-южносибирские – 61 вид (3,8 и 9,7 %). Доля же панбореального и эвксинского геоэлементов невелика – всего 3,7 и 3,5 %. На втором месте по численности видов находится группа общеголарктических геоэлементов (403 вида, 25,2 % флоры). Основная часть (80,6 %) этой группы – палеарктические геоэлементы (325 видов, 20,4 % флоры). На третьем месте стоит группа связующих геоэлементов – 249 видов (15,6 % флоры). Среди них значительно выделяется, составляя 50,6 % от общего количества видов в группе связующих элементов, субкавказский геоэлемент – 126 (7,9 % флоры). Незначительно, по сравнению с предыдущей группой, обеднена видами группа древнесредиземноморских геоэлементов – 243 вида (15,2 % флоры). Среди них ведущую роль играют общедревнесредиземноморский (90 видов, 5,6 % флоры и 37,0 % группы) и западнодревнесредиземноморский (45 видов, 2,8 % флоры и 18,5 % группы). Другие геоэлементы распределяются в следующей последовательности: восточнодревнесредиземноморский (23 вида, 1,4 и 9,5 %); средиземноморский и ирано-туранский геоэлементы представлены одинаковым количеством видов (21 вид, 1,3 и 8,6 %); туранский (16 видов, 1,0 и 6,6 %); армено-иранский (14 видов, 0,9 и 5,8 %); крымсконовороссийский (13 видов, 0,8 и 5,4 %). Плюрирегиональный и адвентивный географические элементы представлены незначительным количеством видов (39 – 2,4 % и 36 – 2,3 % соответственно). Таким образом, бореальный характер флоры района не вызывает сомнений – в целом, ее можно определить как бореально-общеголарктическо-древнесредиземноморскую. При определении типа флоры в данном случае необходимо учитывать большой процент участия кавказских, палеарктических и европейских видов, т. к. именно они играют существенную роль в общей картине флоры района. 169 Для флоры территории Пятигорья характерно явление эндемизма условно трех типов: стеноэндемизма (настоящего эндемизма), эвриэндемизма и субэндемизма. Флора Пятигорья содержит 5 стеноэндемичных видов; 4 эвриэндемика и 21 субэндемика Предкавказья. Новейшее расо- и видообразование особенно ярко выражено в родах Papaver, Astragalus, Centaurea, обособление апомиктических рас – в роде Hieracium. Исследуемый район можно определить как единый Пятигорский рефугиум, большинство реликтов которого имеют доплейстоценовый возраст. Реликтовость флоры района имеет смешанный характер. Она представлена 18 третичными реликтами, 41 гляциальным и 42 ксеротермическими. Таким образом, по всем приведенным выше данным видно, что район является уникальным с точки зрения флористического разнообразия, а, следовательно, требует дальнейшего изучения и, самое важное, охраны растительных комплексов. Л и т е р а т ур а Иванов А. Л. Флора Предкавказья и ее генезис / А. Л. Иванов. – Ставрополь: Изд-во СГУ, 1998. – 204 с. Портениер Н. Н. Географический анализ флоры бассейна реки Черек Безенгийский (Центральный Кавказ). II. Географические элементы / Н. Н. Портениер // Ботан. журн. – 1993. – Т. 78. – № 11. – С. 1–18. Чупахин В. М. Физическая география Северного Кавказа / В. М. Чупахин. – Ростов: Изд-во Ростовского ун-та, 1974. – 196 с. ДИНАМИКА ИЗМЕНЕНИЯ СТРУКТУРЫ ЛЕСНЫХ НАСАЖДЕНИЙ И ФИТОМАССЫ ЛУБЯНСКОГО УЧАСТКОВОГО ЛЕСНИЧЕСТВА ЗА 1940–2007 ГОДЫ Фархуллин Р. Ш. Казанский федеральный университет, г. Казань, rusfara@yandex.ru Лубянское участковое лесничество, расположено в северной части Республики Татарстан, на юго-востоке Кукморского муниципального района. Протяженность лесничества с севера на юг составляет 14 км, с запада на восток 36 км. Общая площадь лесничества по состоянию на 01.01.2007 год составляла 12955 га. В лесничестве преобладают бурые лесные супесчаные (левобережье Вятки) и коричнево-бурые лесные типичные (правобережье Вятки) почвы (Газизуллин, 2005). В лесорастительном отношении лесничество относится к подзоне южной тайги, которую профессор Марков в пределах ТАССР называет подзоной елово-пихтовых смешанных лесов (Марков, 1948). 170 Особенностью лесничества является его водоохранное значение, как лесного массива, лежащего в береговой защитной зоне ВолжскоКамского водохранилища. В настоящее время исходные коренные растительные формации еловых и сосновых лесов на большей площади лесничества утратили свой первоначальный характер и сменились лиственными насаждениями. Концентрированные сплошные рубки за короткий срок коренным образом изменили внешний облик лесничества, сменив на преобладающей площади ландшафт спелых хвойных древостоев вырубками, возобновившимися порослевыми лиственными молодняками. Сосновые и елово-пихтовые насаждения, занимавшие в 1926 году – 83 % площади лесничества, а в 2007 году составили – 52 % (рис.). Особенно резко сократилась площадь ельников. В 1926 г. они занимали 42 %, а 2007 г. – 13 % покрытой лесом площади (Сводные материалы Министерства лесного хозяйства Республики Татарстан, 2007). Хозяйственно ценные хвойные насаждения сменились мягколиственными. Этот процесс смены явился результатом хозяйственной деятельности, при которой заботы и мероприятия по восстановлению хвойных пород были явно недостаточны и если состав лесов пополнялся культурами сосны, то в отношении восстановления елово-пихтовых лесов никаких мероприятий не проводилось. В возрастной структуре в 1940 г. спелые насаждения занимали 67 % от всей лесопокрытой площади (рис.). В конце пятидесятых годов преобладали уже молодняки – 46 %, спелые насаждения составляли 32 % (сосна, ель). Это явилось следствием расширенного отпуска леса за счет использования эксплуатационных запасов в закрепленной за Лубянским леспромхозом сырьевой базе, по которой за отдельные годы отпускалось до 150 тыс. м3, т. е. более чем в два раза против установленного отпуска (Фархуллин Р. Ш., 2012). Общее пользование лесом с 1 га лесной площади составило 12 м3 при ежегодном приросте 3,1 м3, тогда как обычное пользование лесом в других лесхозах 2 группы лесов ТАССР с 1 га лесной площади составляло 4–5 м3. В 2007 г. в породном составе преобладали сосна (38,2 %), береза (26 %), ель (14 %). Доля хвойных насаждений была увеличена до 52,2 %. В возрастной структуре преобладали молодняки – 29 % (сосна, ель), средневозрастные – 27 % (сосна, береза), спелые – 21,8 % (береза). При этом среди хвойных пород доля спелых составляла всего 7,3 %. 171 2007 год 1940 го д 4% 21% 20% 29% 4% 8% 20% 67% 27% молодняки средневозрастные приспевающие спелые и перестойные молодняки среднев озрастные приспев ающие спелые перестойные Рисунок 1 – Динамика покрытой лесом площади Лубянского участкового лесничества по группам возраста с 1940 по 2007 годы Используя таблицу средних значений плотности древесины при нормальной влажности, характерные для Умеренной зоны, мы вычислили фитомассу лесных насаждений Лубянского участкового лесничества за 1940 и 2007 годы (Уголев, 2001). В результате анализа мы получили, что в 1940 году общая сумма фитомассы равнялась 1170,3 Мт. Из них на долю хвойных приходилось – 88,2 %, твердолиственных пород – 4,1 %, мягколиственных – 7,7 %. В 2007 году общая фитомасса в лесничестве увеличилась и была равна 1423,4 Мт. Из них на долю хвойных приходилось – 52,5 %, твердолиственных пород – 2,1 %, мягколиственных – 45,4 %. Таким образом, за более чем 60 лет фитомасса хвойных и твердолиственных насаждений уменьшилась в 1,5 раза, а фитомасса мягколиственных насаждений увеличилась в 7 раз. В перспективном периоде перед лесничеством стоят серьезные задачи по восстановлению хозяйственно-ценных пород на больших площадях и по восстановлению существующих лиственных молодняков в направлении улучшения состава древесных пород и перевода их в хвойные хозяйства. В связи с истощением эксплуатационных запасов леса и возрастающей разнообразной потребностью в древесине для удовлетворения местных нужд, экономическое значение и ценность лесов Лубянского участкового лесничества еще более возрастает в дальнейшем периоде хозяйства. Таким образом, рекомендации будут заключаться в сохранении ресурсно-экологического потенциала лесов лесничества, повышение их 172 качественного состояния и биоразнообразия. При искусственном лесовосстановлении и лесоразведении необходимо стремиться воссоздавать возможную местную (естественную для данной зоны) полночленность биоценозов, свойственных данным экотопам. Л и т е р а т ур а Газизуллин А. Х. Почвенно-экологические условия формирования лесов Среднего Поволжья. Т. 1: Почвы лесов Среднего Поволжья, их генезис, систематика и лесорастительные свойства: научное издание / А. Х. Газизуллин. – Казань: РИЦ «Школа», 2005. – 496 с. Марков М. В. Растительность Татарии / М. В. Марков. – Казань: Татгосиздат, 1948. – 128 с. Сводные материалы Министерства лесного хозяйства по учету лесного фонда Лубянского лесхоз-техникума на 01.01.2007 г. Уголев Б. Н. Древесиноведение с основами лесного товароведения. Третье переработанное и дополненное издание: учеб. для лесотехнических вузов / Б. Н. Уголев. – М.: МГУЛ, 2001. – С. 110–111. Фархуллин Р. Ш. Динамика лесного потенциала Лубянского участкового лесничества Республики Татарстан (1926– 2007) / Р. Ш. Фархуллин, К. К. Ибрагимова // Общество, современная наука и образование: проблемы и перспективы: сборник научных трудов по материалам Международной научно-практической конференции. – Тамбов: ТРОО «Бизнес-Наука-Общество», 2012. – Ч. 8. – С. 139–140. ЭДЕМИКИ НАЦИОНАЛЬНОГО ПАРКА «ПРИЭЛЬБРУСЬЕ» (ЦЕНТРАЛЬНЫЙ КАВКАЗ) Цепкова Н. Л., Тубаева М. А. Институт экологии горных территорий КБНЦ РАН, г. Нальчик, cenelli@yandex.ru Эндемики являются наиболее показательными элементами флоры, по которым можно судить о ее оригинальности. По данным А. И. Галушко (1976), флора западной части Центрального Кавказа насчитывает в своем составе около 124 эндемичных таксонов: видов, разновидностей и гибридов. В задачи нашей работы входило выявление эндемиков на территории национального парка «Приэльбрусье», о приуроченности их к высотным поясам. Национальный парк «Приэльбрусье» (НП), образованный в 1986 г. на площади 101,2 тыс. га, охватывает западные части северных макросклонов Главного и Бокового Кавказских хребтов, включая районы Приэльбрусья с высочайшей вершиной Европы вулканом Эльбрус (5642 м н. ур. м.) и верховья рек Баксан (южное Приэльбрусье) и Малка (северное Приэльбрусье). В соответствии с системой высотно-поясной структуры Кавказа (Соколов, Темботов, 1989) территория НП относится к эльбрусскому варианту восточно-северокавказского типа поясности, характерными особенностями которого являются отсутствие лесного пояса и доминирование субальпийского пояса над другими поясами (альпийским, субнивальным). 173 Субнивальный пояс в НП «Приэльбрусье» (западная часть северного макросклона Центрального Кавказа) начинается примерно на высоте 2900–3000 м н. у. м. Верхней его границей считается «уровень 365» (абсолютная высота, выше которой снежный покров держится 365 дней в году). Альпийский пояс ограничен высотами 2500–2600 – 2800– 2900 м н. у. м. Оба пояса отличаются своеобразными экологическими условиями, обусловленными экстремальными режимами радиации, тепла и влаги, выходами коренных пород, массивами скальных обнажений, близостью ледников и снежников. Пестрота условий обитания наряду с изолированностью территории (островной эффект) считаются факторами формирования эндемизма (Галушко, 1976). По данным, полученным нами во время маршрутно-экспедиционных обследований высокогорных ландшафтов нацпарка, общими для альпийского и субнивального поясов являются 22 кавказских эндемика (в группу кавказских эндемиков мы включили собственно кавказские, условно кавказские и узкорегиональные). Большинство из эндемиков встречаются на моренах, щебнистых склонах, скалах, реже – на лугах. К ним относятся Androsace raddeana Somm. et Levier, проломник Радде (территория НП, рассеянно), Anemone speciosa Adams. ex E. Pritz., ветреница видная (в НП повсеместно, обычно), Asperula cristata (Somm. et. Levier) V. Krecz., ясменник гребенчатый (повсеместно, обычно), Astragalus levieri Freyn ex Somm. et Levier, астрагал Левье (повсеместно, обычно), Campanula biebersteiniana Schult., колокольчик Биберштейна (повсеместно, обычно), C. ciliatа Stev., колокольчик реснитчатый (повсеместно, обычно), Cerastium undulatifolium Somm. et Levier, ясколка волосистолистная (повсеместно, обычно), Cerastium polymorphum Rupr., ясколка полиморфная (повсеместно, обычно), Delphinium caucasicum C. A. Mey., живокость кавказская (урочище Джилысу, ущелья Адылсу, Адырсу), Draba bryoides DC., крупка моховидная (территория НП, обычно), D. scabra C. A. Mey., крупка шероховатая (ущелья Адылсу, Шхельда, редко), D. supranivalis Rupr. крупка приледниковая (территория НП, рассеянно), Minuartia buschiana Schischk., минуарция Буша (каменистые склоны в ущельях НП; обычно), Pedicularis crassirostris Bunge, мытник толстоклювый (в ущельях и урочищах НП, рассеянно), P. subrostrata C. A. Mey., мытник почтиносатый (в ущельях и урочищах НП, рассеянно), Primula bayernii Rupr., первоцвет Байерна (ущелье Адылсу, редко; вид занесен в Красную книгу Кабардино-Балкарии (2000)), Pseudovesicaria digitata (C. A. Mey.) Rupr., лжепузырник пальчатый (урочище Джилысу, редко), Ranunculus trisectilis Oves., лютик трехрассеченный (повсеместно на территории НП, обычно), Trifolium polyphyllum C. A. Mey., клевер многолистный (повсеместно на территории НП, рассеянно; занесен в Красную книгу 174 КБР), Trygonocaryum involucratum (Stev.) Kusn., тригонокариум оберточный (верховья Баксанского ущелья, редко), Taraxacum porphyranthum Boiss., одуванчик пурпурноцветковый (в ущельях и урочищах НП, рассеянно), Valeriana saxatilis C. A. Mey., валериана скальная (скалы, каменистые места; рассеянно). В пределах только альпийского пояса обнаружено 5 эндемиков: Crocus scharojanii Rupr., шафран Шарояна (альпийские луга на склонах горы Чегет), Euphrasia amblyodonta Juz., очанка тупозубая (альпийские луга НП; рассеянно), Gentiana angulosa Bieb., горечавка угловатая (альпийские луга в ущельях НП; обычно), G. oschtenica (Kusn.) Woronow, горечавка оштенская (на щебнистых склонах в урочище Джилысу; редко), Salix apoda Trautv., ива безногая (луга, каменистые склоны, заросли рододендрона; рассеянно на территории НП). Субальпийский пояс в НП «Приэльбрусье» занимает большие пространства на высотах от 1500–1800 до 2400–2500 м и отличается благоприятными биоклиматическими факторами для развития растительности, что объясняет флористическое и фитоценотическое разнообразие. Интересным является нахождение в этом поясе двух узкорегиональных эндемиков (эндемиков Баксанского ущелья в пределах НП), включенных в Красную Книгу КБР (2000) – Cicer balcaricum Galuschko, нут балкарский (на осыпных склонах в районе пос. Эльбрус; очень редко) и Daphne baksanica Pobed., волчеягодник баксанский (на сухих склонах в Баксанском ущелье; очень редко). В субальпах в пределах НП «Приэльбрусье» встречается 7 эндемиков, общих с альпийским поясом. В Красную книгу КБР из них включены два вида – Betula raddeana Trautv., береза Радде (в ущельях НП; очень редко) и Rhododendron caucasicum Pall., рододендрон кавказский (в ущельях НП на каменистых открытых местах, в лесных сообществах; обычно). Десять эндемиков имеют широкую амплитуду высотного распространения, встречаясь как в субальпийском поясе, так и в альпийском и субнивальном. В Красную книгу КБР занесен один вид – Potentilla divina Albov, лапчатка удивительная (в трещинах скал; изредка на склонах Главного и Бокового хребтов в пределах парка). Всего на территории национального парка «Приэльбрусье» по предварительным данным обнаружено 49 эндемиков. Наибольшим богатством кавказскими эндемиками отличаются альпийский и субнивальный пояса, что подтверждает вывод А. И. Галушко (1976) о тесной связи между числом эндемиков и высотным положением территории, об оригинальности по количеству эндемичных видов верхних поясов. Р. М. Середин (1979) также говорит о том, что число эндемичных видов Северного Кавказа, характерных для высокогорий, в 2,5 раза превышает 175 число эндемиков, произрастающих в среднем поясе, предгорьях и на низменности. Исследования по выявлению эндемичных видов в НП «Приэльбрусье» планируется продолжить. Л и т е р а т ур а Галушко А. И. Флора Северного Кавказа и вопросы ее истории / А. И. Галушко. – Ставрополь: Изд-во Ставропольского госпединститута, 1976. – 202 с. Красная книга Кабардино-Балкарской республики. Редкие, находящиеся под угрозой исчезновения виды животных и растений / отв. ред. И. В. Иванов – Нальчик: Эль-Фа, 2000. – 308 с. Середин Р. М. Флора и растительность Северного Кавказа: учеб. пособие / Р. М. Середин. – Краснодар: КубГУ, 1979. – 88 с. Соколов В. Е. Млекопитающие. Насекомоядные / В. Е. Соколов, А. К. Темботов. – М.: Наука, 1989. – С. 3–27. РЕДКИЕ И ОХРАНЯЕМЫЕ ВИДЫ БЕСПОЗВОНОЧНЫХ В БАССЕЙНЕ Р. ЦИВИЛЬ (ЧУВАШСКАЯ РЕСПУБЛИКА) Яковлев В. А.1, 2, Яковлева А. В.1 1 Институт проблем экологии и недропользования АН Республики Татарстан, 2 Казанский (Приволжский) федеральный университет, г. Казань В ходе исследования зообентоса бассейна реки Цивиль в летнеосенний период 2013 г. были обнаружены виды беспозвоночных из следующих систематических групп: губки, гидры, нематоды, олигохеты, пиявки, моллюски, ракообразные (равноногие), паукообразные (водяные клещи), насекомые (поденки, стрекозы, полужесткокрылые, жесткокрылые, большекрылые, ручейники, двукрылые). Основу состава беспозвоночных формируют широко распространенные в Палеарктике виды, обычные для водотоков и стоячих водоемов Среднего Поволжья. Однако среди них было выявлено четыре вида, представляющие значительный интерес в связи с их редкостью, исчезновением и включением в Красные книги некоторых субъектов Российской Федерации и даже ряда стран Европы. В первую очередь следует отметить обнаружение личинок (нимф) относительно редкого в Палеарктике вида – поденки белой Polymitarcis (=Ephoron) virgo Oliver, 1791 (семейство Polymitarcidae, Ephemeroptera). Всего 11 экз. личинок были обнаружены в реке Большой Цивиль. Форма тела личинок несколько похожа на тело медведки – обитателя почвы. Они, как этот вид, роют в грунте ходы с помощью верхних челюстей и сильных ног. Двигая жабрами, они создают там приток воды, содержащей кислород и пищу (преимущественно детрит и мелкие организмы) (Elliot et al., 1988; Монаков, 1998). Распространен этот, вид в Голарктике (США, Европа, включая территорию Российской Федерации, а также в Южной Сибири и Китае). Встречался ранее по всей Волге 176 (Бенинг, 1924). Он указан для рек Республики Татарстан и включен там в Красную книгу (Красная Книга Республики Татарстан, 2006; Яковлев, 2009). Второй, также крайне редкий вид поденок, Ephemera lineata Eaton, 1870 (семейство Ephemeridae, Ephemeroptera), был обнаружен 11.09.2013 г. в устье р. Средний Цивиль. Глубина реки составляла 0,1−0,2 м (грунт − песок с остатками растений). Численность личинок этого вида составляла 10 экз./м2, а биомасса − 0,37 г/м2. Известен как обитатель быстро текущих рек с илисто-песчаным грунтом. Личинки ведут скрытный образ жизни. Питаются детритом и водорослями (Монаков, 1998). Вид распространен в Европе. Отмечено повсеместное сокращение численности, включен в Красную книгу Республики Татарстан (2006) (Яковлев, 2009). Следующий вид также представляет интерес. Четыре особи ранатры палочковидной Ranatra linearis Linne, 1758 (семейство Nepidae, Heteroptera) были выловлены 11.09.2013 г. в р. Бол. Цивиль в районе впадения в нее р. Унга. Вид включен в Красные книги многих субъектов Российской Федерации. Ранее он был указан для ряда водных объектов Республики Чувашия, где и внесен в Красную книгу (Егоров и др. 2006, Корнилов и др., 2007; по: Красная…, 2010). Распространение: западная часть Палеарктики. Представляет большой интерес обнаружение типичного реофильного вида плавта летнего Aphelocheirus aestivalis (Fabricius, 1803) (Aphelocheiridae, Heteroptera). Клоп относительно многочислен в верховьях р. Цивиль, на участках, представляющих собой перекаты, сложенных гравием, галькой, реже крупнозернистым песком. Скорость течения воды была повсеместно выше 0,3 м/с. Глубина воды – от 0,1 до 1,0 м. Плавт получает кислород из воды с помощью жабр и, соответственно, чувствителен к его недостатку. Ведет придонный образ жизни, почти не всплывает, сидит неподвижно на поверхности камней и на песчаном грунте или двигается по дну в основном в темное время суток. Он отмечен для водоемов Турции, Грузии и Египта (Канюкова, 1971; 1989; 1998). В составе полиморфного вида выделяют различное количество подвидов, форм и даже видов. В Европейской части России пока отмечен лишь A. aestivalis. Он указан для малых рек междуречья Суры и Мокши (Каменев, 2002), рек Казанка, Меша, Свияга, Лесной Зай и ряда других рек Закамья Республики Татарстан (Экологические…, 2003; Красная…, 2006). Он обнаружен в реках Удмуртской Республики (Каргапольцева и др., 2012), Пермской и Оренбургской областей. Плавт A. aestivalis включен в Красную книгу Норвегии как почти находящийся под угрозой исчезновения (Coullianas et al., 2008). На данный момент его статус в Чувашской Республике можно оценивать как вид, принад177 лежащий к IV категории – «Вид с неопределенным статусом», поскольку достаточных сведений о состоянии его популяции пока нет. Таким образом, из четырех относительно редких беспозвоночных, обнаруженных в 2013 г. в бассейне р. Цивиль, лишь ранатра палочковидная включена в Красную книгу Чувашской Республики. Остальные три вида также нуждаются в охране. Они могли бы рассматриваться как потенциальные кандидаты в следующее издание Красной книги. За исключением фитофильной ранатры, остальные виды – типичные реофилы, чувствительные к загрязнению и, особенно, заиливанию ложа реки. Соответственно, поступление тонких взвесей с водосборной площади, активно используемой сельскохозяйственной деятельностью, представляет угрозу для них. Следующей причиной их исчезновения может стать зарегулирование течения плотинами. В верхней части бассейна угрозу для них также представляет выпас крупного рогатого скота. Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований и Министерства образования и молодежной политики ЧР (проект № 13-04-97158 р_поволжье_а). Л и т е р а т ур а Бенинг А. Л. К изучению придонной жизни реки Волги / А. Л. Бенинг. – Саратов: Б. и., 1924. – 398 с. Канюкова Е. В. Водные клопы северного Присурья / Е. В. Канюкова // Материалы 1-й научн. конф. по проблеме фауны, экологии, биоценологии и охраны животных Присурья. – Саранск. 1971. – С. 23–24. Канюкова Е. В. Полужесткокрылые рода Ranatra (Heteroptera, Nepidae) фауны СССР / Е. В. Канюкова // Энтомол. обозрен. – 1989. – Т. 68, вып. 1. – С. 121–124. Канюкова Е. В. Список видов водных полужесткокрылых (Hetroptera) бассейна р. Волги / Е. В. Канюкова // Энтомологические исследования в Чувашии: материалы энтомол. конф. Чебоксары, 24–25.10.1997 г. – Чебоксары, 1998. – С. 37– 41. Каменев А. Г. Биопродуктивность и биоиндикация малых водотоков междуречья сообществ Суры и Мокши. Макрозообентос / А. Г. Каменев. – Саранск: Изд-во Мордов. унта, 2002. – 120 с. Каргапольцева И. А. Материалы к фауне водных полужесткокрылых (Heteroptera) Удмуртской Республики / И. А. Каргапольцева, Н. В. Холмогорова, М. А. Грандова // Вестник Удмуртск. ун-та. Сер., Биология. Наука о Земле. – 2012. – Вып. 2. – С. 38–46. Красная книга Республики Татарстан. – Казань: Идель-Пресс, 2006. – 832 с. Красная книга Чувашской Республики. Т. 1. Ч 2: Редкие и исчезающие виды животных. – Чебоксары: ГУП «ИПК «Чувашия», 2010. – 372 с. Монаков А. В. Питание пресноводных беспозвоночных / А. В. Монаков. – М.: Б. и., 1998. – 320 с. Яковлев В. А. Охраняемые водные беспозвоночные Республики Татарстан / В. А. Яковлев. – Казань: Изд-во Казанск. ун-та, 2009. – 140 с. Экологические проблемы малых рек Республики Татарстан (на примере Меши, Казанки и Свияги) / под ред. В. А. Яковлева. – Казань: Фэн, 2003. – 289 с. Coullianos C. C. Norwegian water bugs. Distribution and ecology (Hemiptera– Heteroptera: Gerromorpha and Nepomorpha) / C. C. Coullianos, J. Økland, K. A. Økland // Norw. J. Entomol. – 2008. – Vol. 55. – P. 179–222. Elliot J. M. Larvae of the British Ephemeroptera / J. M. Elliot, U. H. Humpesch, A. N. Macan // Freshwater biol. Ass. Sci. Publ. – 1988. – No 49. – 145 p. 178 Секция 3 МОНИТОРИНГ АБИОТИЧЕСКИХ И БИОТИЧЕСКИХ КОМПОНЕНТОВ ЭКОСИСТЕМ ДИНАМИКА СОДЕРЖАНИЯ АСКОРБИНОВОЙ КИСЛОТЫ В ЛИСТЬЯХ ДРЕВЕСНЫХ ВИДОВ РАСТЕНИЙ В НАСАЖДЕНИЯХ С РАЗЛИЧНОЙ ТЕХНОГЕННОЙ НАГРУЗКОЙ Бухарина И. Л.1, Кузьмин П. А.2 1 Удмуртский государственный университет, г. Ижевск 2 Казанский федеральный университет, г. Елабуга В адаптивных реакциях участвуют различные метаболиты, в том числе низкомолекулярной природы – аскорбиновая кислота. Аскорбиновая кислота рассматривается учеными как важный элемент в формировании ответных адаптивных реакций древесных растений на повышающийся уровень загрязнения окружающей среды (Кулагин, 2006). Исходя из этого, мы поставили перед собой цель изучить особенности динамики содержания в листьях аскорбиновой кислоты, как элемента антиоксидантной системы защиты, в период активной вегетации аборигенных видов древесных растений. Исследования проведены в г. Набережные Челны. Комплексный индекс загрязнения атмосферы (ИЗА) показывает очень высокое загрязнение (ИЗА = 15,3) и превышение уровня предельно допустимой концентрации по бенз(а)пирену, формальдегиду, фенолам и оксидам углерода и азота (Атлас…, 2005; Государственный…, 2011). Объект исследования древесные растения: аборигенные виды – клен остролистный (Acer platanoides L.), липа мелколистная (Tilia cordata Mill.) и береза повислая (Betula pendula Roth). Изучаемые виды произрастали в городе в составе насаждений различных экологических категорий: магистральные посадки и санитарнозащитные зоны (СЗЗ) промышленных предприятий ОАО «КамАЗ», являющихся основными загрязнителями города. В качестве зоны условного контроля (ЗУК) выбрана территория Челнинского лесничества (лесостепная зона, общая площадь лесничества 9539 га). Пробные площади закладывали регулярным способом (по 5 шт. в каждом районе, размером не менее 0,25 га). В пределах пробной площади проведен отбор (по 10 растений каждого вида) и нумерация учетных 179 древесных растений. Учетные особи имели хорошее жизненное состояние и средневозрастное онтогенетическое состояние (g2). Для проведения лабораторных физиолого-биохимических анализов были взяты пробы верхушечных вегетативных годичных побегов и срединных (ассимилирующих) листьев. Отбор проводился с помощью секатора на шесте со средней и нижней части (исключая нижние ветви) кроны древесных растений южной экспозиции. Смешанную пробу не проводили (каждая особь выступала как повторность), для каждой особи анализы проводили в трех повторениях. Количественное содержание аскорбиновой кислоты определяли по ГОСТ 24556-89. Анализы проводили в лаборатории «Экологии и физиологии растений» биологического факультета Елабужского института (филиала) ФГАОУ ВПО «Казанский (Приволжский) федеральный университет». Математическую обработку материалов провели с применением статистического пакета «Statistica 5.5». Для интерпретации полученных материалов использовали методы описательной статистики и дисперсионный многофакторный анализ (по перекрестно-иерархической схеме, при последующей оценке различий методом множественного сравнения LSD-test). Дисперсионный многофакторный анализ результатов исследований показал, что на содержание аскорбиновой кислоты в листьях древесных растений достоверное влияние оказали видовые особенности (Р = 5,68·10–5), комплекс условий места произрастания (Р < 10–5), фазы вегетации (Р < 10–5) а также взаимодействия этих факторов (Р < 10–5) (табл. 1). У всех изучаемых видов древесных растений, не зависимо от условий произрастания, количество данного метаболита снижалось в течение вегетации, что свидетельствует о снижении активности окислительно-восстановительных процессов у растений. У липы мелколистной, березы повислой и клена остролистного, произрастающих в насаждениях СЗЗ промышленных предприятий, содержание аскорбиновой кислоты в листьях по сравнению с контролем было выше: в июне – на 110,5; 84,8 и 150,5; в августе – на 56,9; 67,5 и 90,6, соответственно. В магистральных насаждениях тенденция была иной: у липы мелколистной и у березы повислой содержание данного метаболита, по сравнению с ЗУК, было значительно ниже в июне – на 155,4 и 230, в июле – на 80,2 и 103,7, в августе – на 40,9 и 110,3, соответственно. У клена остролистного, наоборот, оно было значительно выше, причем в июне – на 178,5, в июле – на 28,4, в августе – на 2,6 мг %. 180 Таблица 1 – Динамика содержания аскорбиновой кислоты в листьях древесных растений, произрастающих в различных категориях насаждений г. Набережные Челны, мг % Месяц Вид древесного растения Tilia cordata Mill. Betula pendula Roth. Acer platanoides L. Зона условного контроля (НСР05 = 2,4) июнь 329,6 372,8 176,3 июль 186,3 222,5 120,7 133,4 188,7 104,7 август Санитарно-защитные зоны промышленных предприятий июнь 440,1 457,6 326,8 июль 175,0 218,6 154,5 август 190,3 256,2 195,3 Магистральные посадки июнь 174,2 142,8 июль 106,1 118,8 354,8 149,1 август 92,5 78,6 107,3 Сходная динамика получена ранее в результате научных исследований, проведенных в крупном промышленном центре Уральского региона г. Ижевске (Бухарина, 2007; 2011). Таким образом, повышение степени техногенной нагрузки приводит к возрастанию содержания аскорбиновой кислоты в листьях у изучаемых видов древесных растений в насаждениях СЗЗ промышленных предприятий. Однако у особей в магистральных насаждениях имеются отличия в динамике содержания аскорбиновой кислоты в листьях. Схожие с растениями в насаждениях санзон реакции отмечены лишь у клена остролистного, а у остальных изучаемых видов наблюдается достоверное снижение содержания концентрации данного метаболита. На наш взгляд, это свидетельствует о наличии иных физиологических процессов, компенсирующих негативное воздействие техногенной нагрузки в урбаносреде. Л и т е р а т ур а Атлас Республики Татарстан / гл. ред. Г. В. Поздняк. – М., 2005. – 700 с. Бухарина И. Л. Эколого-биологические особенности древесных растений в урбанизированной среде / И. Л. Бухарина, Т. М. Поварницина, К. Е. Ведерников. – Ижевск: ФГОУ ВПО Ижевская ГСХА, 2007. – 216 с. Бухарина И. Л. Особенности динамики содержания аскорбиновой кислоты и танинов в побегах древесных растений в условиях г. Ижевска / И. Л. Бухарина // Растительные ресурсы. – М., 2011. – № 2. – С. 109–117. Кулагин А. А. Реализация адаптивного потенциала древесных растений в экстремальных лесорастительных условиях: автореф. дис. … д-ра биол. наук / А. А. Кулагин. – Уфа-Тольятти, 2006. – 181 32 с. Государственный доклад «О состоянии природных ресурсов и об охране окружающей среды Республики Татарстан в 2011 году» (29.06.2012 г.). – URL: http://www.eco.tatarstan.ru/rus/info.php?id=424234 (дата обращения: 15.07.2012). ОСОБЕННОСТИ ЛЕСОРАСТИТЕЛЬНОГО РАЙОНИРОВАНИЯ СРЕДНЕГО ПОВОЛЖЬЯ Глушко С. Г. Казанский государственный аграрный университет, г. Казань, glushkosg@mail.ru В ходе лесорастительного районирования РФ в 2007, 2009 гг. (Перечень, 2009) лесорастительные условия Среднего Поволжья «приведены в известность». Постоянный набор новых фактов, анализ поступающей информации позволяют выявлять современные тенденции лесообразовательного процесса, существенно воздействующие на лесорастительные условия (Галиуллин, 2011; Прохоренко, 2012, и др.). Целью наших исследований стал учет современных тенденций динамики лесов, определяющих формирование лесорастительных условий. В задачи входило: оценить устойчивость, выявить основные тенденции динамики лесов, дать рекомендации по результатам лесорастительного районирования. Объектом исследования стали леса Среднего Поволжья. Работа выполнялась на кафедре таксации и экономики лесной отрасли Казанского ГАУ при помощи Казанского филиала ФГУП «Рослесинфорг», с привлечением студентов, специалистов других организаций (лесной научно-образовательный кластер РТ). Проводились маршрутно-рекогносцировочные и стационарные работы с использованием материалов лесоустройства, научной и справочной литературы, с применением общепринятых методик. По итогам лесорастительного районирования 2009 г в Среднем Поволжье установлена зона хвойно-широколиственных лесов (ХШЛ) и лесостепная зона, кроме того присутствуют фрагменты таежной (южно-таежной) и степной лесорастительных зон. В соответствии с данными лесной науки, на севере региона распространен таежный тип растительности (бореальный), в центральной части преобладает широколиственный тип (неморальный), и на юге господствует степной тип растительности (в т. ч. лугово-степной). В настоящее время леса в зоне ХШЛ подверглись значительной фрагментации, остатки условно-коренных лесов чередуются с лесами вторичными, разной степени производности, в том числе с дериватами, олуговелыми и т. п. участками. Процессы распространения липы на се- 182 вер, массового усыхания ели, дуба и березы, лесные смены, заслуживают внимания исследователей. ХШЛ региона в своей южной полосе, в Приволжском и ЮжноПредуральском районах ХШЛ по С. Ф. Курнаеву (Курнаев, 1958), фактически деградировали. Из состава лесов выпадают хвойные породы и дуб, распространяются по богатым условиям – липа, по бедным – береза. Сохранение лесов способствует сохранению биоразнообразия. На месте деградирующих ХШЛ в регионе активно формируется зона широколиственных лесов (ШЛ). Вопрос автохтонности или наоборот вторичности региональной зоны ШЛ требует решения, но наличие данной зоны в настоящее время не вызывает сомнений. Формируемая зона ШЛ Среднего Поволжья крайне фрагментирована, лесные сообщества неустойчивы, находятся на разных стадиях своего восстановительного развития, господствуют леса производные. Зона ШЛ имеет тенденцию к продвижению на север региона, за счет распространения липы по относительно богатым условиям, вместо массово усыхающих сложных ельников. Обособление зоны ШЛ можно представить как последствие процессов антропогенной фрагментации лесов. Современные тенденции лесообразовательного процесса следует зафиксировать в итогах лесорастительного районирования. Так как современная зона ШЛ формируется в значительной степени за счет вытеснения хвойных пород, то она может быть подразделена на две полосы: Северная полоса ШЛ-С, кроме собственно широколиственных и иных лиственных насаждений, содержит фрагменты хвойных лесов естественного происхождения. В южной полосе ШЛ-Ю хвойные леса естественного происхождения имеют реликтовый характер, распространены участки олуговелые, остепненные и степные. Мы предлагаем вместо принятого действующими нормативами разделения ШЛ между зонами ХШЛ и лесостепей, выделить фактически существующую зону ШЛ. Ниже нами схематично сопоставлены итоги нормативного (Перечень, 2009) лесорастительного районирования региона с фактически сложившимися лесорастительными зонами, устанавливаемыми по типам растительности и иным «нормативам» лесной науки. Предлагаемое нами выделение зоны ШЛ, осуществляемое преимущественно за счет зоны ХШЛ тем более оправдано, что в ШЛ есть зональные лесные формации (дубовая, липовая), а в ХШЛ нет. Более того, в рассматриваемых ХШЛ по факту деградируют все зональные лесные субформации (хвойно-широколиственные). 183 Схема лесорастительного районирования Среднего Поволжья по действующим в лесном хозяйстве нормативам (2009 г.) и фактически сложившееся Лесорастительные зоны – 2009 г. Лесорастительные зоны – по факту Таежная Таежная Хвойно-широколиственная Хвойно-широколиственная Широколиственная (ШЛ-С,Ю) Лесостепная Лесостепная Степная Степная Полоса исследуемых подтаежных лесов включает в себя зону ХШЛ. Наличие зон ХШЛ и ШЛ в регионе свидетельствует о разнообразии лесов. Подтаежные ХШЛ играют роль экотона между таежным и широколиственным типами растительности. В свою очередь ХШЛ вместе с ШЛ есть экотон – буфер между лесами и лесостепью. Буфер в виде ХШЛ и ШЛ с соответствующими лесорастительными зонами, вероятно, следует отнести к формам приокеанической растительности (Глушко, 1994). Прослойка широколиственной растительности, расположенная между тайгой и степью, участвующая в формировании южной тайги, ХШЛ, ШЛ, лесостепи, есть отличительный признак лесов, формирующихся в особых природно-климатических (муссонных) условиях. Отсутствие широколиственной растительности в экотоне между лесом и степью, есть признак континентальных, центрально-евразийских лесостепей Сибири. Определенное сходство равнинного Зауралья и равнинного Предуралья обусловило отсутствие ХШЛ и ШЛ в Зауралье и их присутствие в Среднем Поволжье в качестве узкой и крайне неустойчивой полосы, требующей должной охраны и воспроизводства. Выделение зоны широколиственных лесов и сохранение лесов региона, будут способствовать сохранению условий местообитания многих видов флоры и фауны. Лесорастительное районирование в регионе Среднего Поволжья необходимо провести на научной основе, в целях осознанного воспроизводства лесов (Глушко, 2010), приведенных в необходимую известность. Л и т е р а т ур а Галиуллин И. Р. Деградация лесов в регионе Среднего Поволжья и ее исследование / И. Р. Галиуллин, С. Г. Глушко, И. Р. Саитов // Современные аспекты сохранения биоразнообразия и пользования природными ресурсами: материалы Всерос. науч.-практ. конф. – Казань: Изд-во Казанского ГАУ, 2011. – Вып. 1. – С. 6–10. Глушко С. Г. Проблемы реконструкции лесов Среднего Поволжья / С. Г. Глушко // Инновационное развитие агропромышленного комплекса: материалы Всерос. науч.-практ. конф. – Казань: Изд-во Казанско184 го ГАУ, 2010. – Т. 77, ч. 2. – С. 325–328. Курнаев, С. Ф. Лесорастительное районирование подзоны южной тайги и хвойно-широколиственных лесов европейской части СССР / С. Ф. Курнаев. – М.: Типография МЛТИ, 1958. – 22 с. Перечень лесорастительных зон и лесных районов: приказ Минсельхоза России № 37, от 04.02.2009 г. – 23 с. Глушко С. Г. Приокеаническая растительность Российского Дальнего Востока / С. Г. Глушко, Н. Б. Прохоренко // Чтения, посвященные 110-летию Общества изучения Амурского края: тез. докл. Владивосток: Приморский объединенный музей, 1994. – С. 255–257. Прохоренко Н. Б. Перспективы сохранения хвойно-широколиственных, подтаежных лесов в Среднем Поволжье / Н. Б. Прохоренко, С. Г. Глушко // Лес, лесной сектор и экология: материалы Всерос. науч.-практ. конф. – Казань: Изд-во Казанского ГАУ, 2012. – С. 60–63. ОСНОВНЫЕ КОМПОНЕНТЫ РЕГИОНАЛЬНЫХ МОДЕЛЕЙ КОМПЛЕКСНОГО БИОМОНИТОРИНГА Дементьева С. М., Нотов А. А., Мейсурова А. Ф., Иванова С. А., Зуева Л. В. Тверской государственный университет, г. Тверь, dementeva1948@mail.ru Единая государственная система экологического мониторинга в России предусматритвает деятельность по оценке уровня загрязнения окружающей среды (Постановление…, 2013). Однако получаемые в ходе такого мониторинга данные будут односторонними без информации о состоянии биологических систем разного уровня. В этой связи необходима разработка моделей комплексного биомониторинга, которые так же, как и модели мониторинга состояния окружающей среды (Постановление…, 2013), должны функционировать на национальном, региональном и локальном уровнях. Актуально выявление универсальных компонентов региональных моделей и поиск дополнительных, которые отражают специфику ресурсно-технического потенциала региона. К числу обязательных направлений в рамках биомониторинга следует отнести исследования по инвентаризации региональной биоты, изучение биологических инвазий. В пределах лесной зоны приобретает актуальность анализ состояния индикаторного компонента биологически ценных лесных сообществ (Выявление…, 2009; Нотов и др., 2012). При наличии соответствующей материально-технической базы возможно дополнение модели физико-химическими методами исследования биологических объектов. На примере Тверской области, включающей центральную часть Каспийско-Балтийского водораздела, разработана региональная модель биомониторинга, которая может представлять интерес для других регионов Центрального Федерального округа (Нотов и др., 2013). Реализация идеи биомониторинга была начата с инвентаризации региональной флоры. С 90-х гг. XX в. она приобрела комплексный харак185 тер. Проведенные исследования позволили оценить уровень видового богатства и степень уникальности разных компонентов флоры. Более детально изучены особо охраняемые природные территории. Среди них Центрально-Лесной государственный природный биосферный заповедник (ЦЛГПБЗ) и национальный парк «Завидово» (Нотов, 2010; Нотов и др., 2011 и др.). На этапе функционирования модели биомониторинга начато специальное изучение индикаторного компонента старовозрастных коренных лесных сообществ по методике, разработанной в СевероЗападной России в рамках совместного шведско-российского проекта (Выявление…, 2009). Нами проведен анализ индикаторных видов в ЦЛГПБЗ и национальном парке «Завидово», в некоторых районах Тверской области. Использованы возможности ГИС-баз и технологий (Нотов и др., 2012 и др.). Характер количественного распределения поллютантов в ЦЛГПБЗ по данным ИК с пектроскопического анализа образцов Hypogymnia physodes: 72–80 номера пунктов сбора образцов; ___ – области распространения поллютантов 186 В 2010 г. обобщены данные об инвазионных видах Тверского региона (Виноградова и др., 2011). Организован мониторинг инвазионной и адвентивной фракций флоры. В рамках программы биомониторинга Верхневолжья осуществлена разработка Черной книги Тверской области. В ней предложены также методы оценки уровня воздействия инвазионной фракций на региональные экосистемы (Виноградова и др., 2011). В качестве исходной количественной характеристики нами предложено использовать суммарную активность инвазионной фракции, которую можно определить для каждого района и природного комплекса. Дальнейший мониторинг инвазионной фракции должен осуществляться в рамках программы ведения Черной книги (Виноградова и др., 2011). Он предполагает следующие направления исследований: 1) изучение структуры и динамики популяций инвазионных видов, анализ адвентивного компонента флоры; 2) выявление путей и способов заноса инвазионных видов, изучение экологии и биологии адвентивных растений; 3) оценка экономического ущерба, наносимого наиболее агрессивными инвазионными видами; 4) разработка конкретных мер, препятствующих внедрению чужеродных видов в природные растительные сообщества региона (Виноградова и др., 2011). Объектом мониторинга стали не только инвазионные виды, но и адвентивный компонент флоры в целом. Начат специальный анализ адвентивной фракции в наиболее оригинальных с точки зрения состава флоры природных комплексах (Ржевско-Старицкое Поволжье, Вышневолоцко-Новоторжский вал). К сожалению, активность инвазионных видов на этих территориях постоянно увеличивается. Использование специальных подходов и методов биоиндикации начато в 1998–2002 гг. В этот период в промышленных районах Тверской области проведено исследование рекреационных зон с использованием лихеноиндикационного подхода (Уразбахтина и др., 2005). В 2002 г. в модель биомониторинга включены физико-химические методы анализа биологических объектов. Использован новый подход с применением Фурье-ИК спектрального анализа слоевищ лишайников (Уразбахтина и др., 2005; Meisurova et al., 2011 и др.). В настоящее время реализуется программа комплексного биомониторинга Верхневолжья с применением разных подходов и методов (Дементьева и др., 2012; Нотолв и др., 2013). Традиционные подходы оценки состояния компонентов природной среды сочетаются с современными физико-химическими методами исследования. Имеющаяся в настоящее время в Тверском государственном университете материально-техническая база позволяет существенно расширить программу мониторинговых исследований. Широко применяется в мониторинговых 187 исследованиях Фурье-ИК спектральный анализ слоевищ лишайников (Meisurova et al., 2011). В отличие от других методов он позволяет выявлять широкий спектр поллютантов, дает интегральную оценку состояния атмосферы, включая данные об экотоксикантах, поступавших в атмосферу в прежние годы. Он дает возможность также не только проводить анализ уровня загрязнения атмосферы, но и оценивать характер воздействия поллютантов на живые объекты (Meisurova et al., 2011). Полученные результаты позволили оценить уровень и характер загрязнения атмосферы наиболее распространенными поллютантами (аммиак, аэрозоли серной и азотной кислот) в разных промышленных центрах и их окрестностях. Фурье-ИК спектральный анализ слоевищ лишайников оказался также эффективным методом оценки влияния лесных пожаров на фитоценозы охраняемых природных территорий (Мейсурова, Нотов, 2012). В момент пожара увеличиваются концентрации оксида углерода, диоксида азота и взвешенных частиц, которые являются первичными экотоксикантами. При высокой температуре воздуха во время лесных пожаров образуются вторичные поллютанты (например, формальдегид). Площадь территории, испытавшей воздействие дыма, зависит от типа пожара, особенностей его локализации, направления и силы ветра. Содержащиеся в дыме химические вещества оказывают определенное воздействие на разные компоненты лесного фитоценоза. С помощью спектрального анализа слоевищ Hypogymnia physodes (L.) Nyl. в 2011 г. был выявлен характер распространения области задымления, которая образовалась в результате пожара, возникшего на территории ЦЛГПБЗ в верховьях реки Тюзьмы в 1999 г. (рис.). В сочетании с Фурье-ИК спектральным анализом лишайников в мониторинговых исследованиях мы используем дополнительную приборную базу. Разовые концентрации экотоксикантов, способствующих кислотному загрязнению атмосферы, определяются нами с помощью газоанализатора Miran (Нотов и др., 2013). Он позволяет обнаруживать в воздухе более 20 компонентов органических и неорганических поллютантов. Для идентификации тяжелых металлов в почве, воде, растительных объектах и лишайниках нами используется атомно-адсорбционный спектрометр с индуктивно-связанной плазмой. Такой анализ актуален в связи с расположением в пределах региона Калининской атомной электростанции, в окрестностях которой реализуется специальная программа мониторинга наземных и водных экосистем. Таким образом, использование приоритетных и дополнительных подходов в региональной модели биомониторинга позволяет осуществлять в настоящее время достаточно точную оценку состояния биосистем 188 разного уровня. Сочетание исследований, позволяющих получить достаточно полную характеристику состояния регионального биоразнообразия, а также анализа экосистем с помощью физико-химических методов дают возможность своевременно выявлять динамику и факторы, способствующие трансформации биосистем. Разработанная модель может быть успешно внедрена в других регионах. Л и т е р а т ур а Виноградова Ю. К. Черная книга флоры Тверской области: чужеродные виды растений в экосистемах Тверского региона / Ю. К. Виноградова, С. Р. Майоров, А. А. Нотов. – М.: КМК, 2011. – 279 с. (Сер. «Чужеродные виды России»). Выявление и обследование биологически ценных лесов на Северо-Западе Европейской части России: учеб. пособие: в 2 т. – СПб., 2009. – Т. 1. – 238 с.; Т. 2. – 258 с. Дементьева С. М. Комплексный мониторинг экосистем как элемент региональной стратегии сохранения биоразнообразия / С. М. Дементьева, А. А. Нотов, А. Ф. Мейсурова, С. А. Иванова, А. В. Павлов, Е. А. Андреева, Л. В. Зуева // Биоразнообразие: проблемы изучения и сохранения: материалы Междунар. науч. конф., посвящ. 95-летию кафедры ботаники Тверского гос. ун-та (г. Тверь, 21–24 нояб. 2012 г.). – Тверь: ТвГУ, 2012. – С. 26–28. Мейсурова А. Ф. Фурье-ИК спектроскопический анализ слоевищ лишайников как метод оценки характера воздействия лесных пожаров на фитоценозы заповедных территорий / А. Ф. Мейсурова, А. А. Нотов // Многолетние процессы в природных комплексах заповедников России: материалы Всерос. науч. конф., посвящ. 80-летию Центрально-Лесного государственного природного биосферного заповедника (20–24 авг. 2012 г., пос. Заповедный, Тверская обл.). – Великие Луки, 2012. – С. 140–145. Нотов А. А. Аннотированный список лихенофлоры Тверской области / А. А. Нотов, Д. Е. Гимельбрант, Г. П. Урбанавичюс. – Тверь: ТвГУ, 2011. – 124 с. Нотов А. А. Возможности использования ГИС-технологий для выяснения характера распространения индикаторных видов лишайников и мохообразных / А. А. Нотов, А. Д. Потемкин, Д. Е. Гимельбрант, В. П. Волков, А. В. Павлов // Динамика многолетних процессов в экосистемах Центрально-Лесного заповедника. – Великие Луки, 2012. – С. 328–356. (Тр. ЦЛГПБЗ; Вып. 6). Нотов А. А. Комплексный биомониторинг природных экосистем центральной части Каспийско-Балтийского водораздела / А. А. Нотов, А. Ф. Мейсурова, С. М. Дементьева // Фундаментальные исследования. – 2013. – № 10 (5). – С. 1090–1094. Нотов А. А. Национальный парк «Завидово»: Сосудистые растения, мохообразные, лишайники / А. А. Нотов. – М.: Деловой мир, 2010. – 432 c. Постановление Правительства РФ от 6 июня 2013 г. № 477 «Об осуществлении государственного мониторинга состояния и загрязнения окружающей среды» (вместе с «Положением о государственном мониторинге состояния и загрязнения окружающей среды») // КонсультантПлюс [Электронный ресурс]. – 2013. – Режим доступа: http://www.consultant.ru/document/ cons_doc_LAW_147245/#p30. Уразбахтина (Мейсурова) А. Ф. Применение метода ФурьеИК спектроскопии для лихеноиндикации атмосферного загрязнения в городских районах / А. Ф. Уразбахтина (Мейсурова), С. Д. Хижняк, С. М. Дементьева, А. А. Нотов, П. М. Пахомов // Растительные ресурсы. – 2005. – Т. 41, вып. 2. – С. 139–147. Meisurova A. F. IR spectroscopic study of the chemical composition of epiphytic lichens / A. F. Meisurova, S. D. Khizhnyak, P. M. Pakhomov // Journal of Applied Spectroscopy. – 2011. – Vol. 78, iss. 5. – P. 711–718. 189 ИСПОЛЬЗОВАНИЕ ГИСТО-МОРФОМЕТРИЧЕСКИХ ПОКАЗАТЕЛЕЙ ПЕЧЕНИ И МИОКАРДА РЕПТИЛИИ ДЛЯ ОЦЕНКИ КАЧЕСТВА НАЗЕМНЫХ ЭКОСИСТЕМ Дробот Г. П., Арсланова А. А., Ведерников А. А. Марийский государственный университет, г. Йошкар-Ола, droga59@mail.ru Вырубка лесов, мелиорация, развитие сельского хозяйства приводят к трансформации ландшафтов, изменениям гидрологического режима и климата региона. Урбанизация, промышленное загрязнение, рекреация оказывают дополнительное влияние на экосистемы, что, в свою очередь, отражается на состоянии популяций амфибий и рептилий (Мильто, 2007). Исследование живых организмов позволяет оценить совокупное физиологическое воздействие антропогенных загрязнений в данном биоценозе (Акуленко, Гассо, Клименко, 2012). В настоящее время анализ структурно-функциональных изменений в печени животных приобретает определенное значение для оценки здоровья их среды обитания. К сожалению, морфологические показатели печени для биоиндикационных исследований используются относительно редко (Дробот, Соловьева, Васина и др., 2007; Дробот, Трубачева, Малюта и др., 2011, Акуленко, Гассо, Клименко, 2012). Работы же, посвященные изучению ответной реакции сердечной мышечной ткани на антропогенное воздействие, – единичны (Дробот, Глотов, Емельянова, 2001; Дробот, Мальцева, Ведерников, 2011). В связи с вышеизложенным цель настоящей работы заключалась в проведении гистологического и морфометрического исследования печени и миокарда ящерицы прыткой (Lacerta agilis Linnaeus, 1758) и оценке возможного применения выбранных морфологических параметров для биоиндикационного исследования антропогенно нарушенных территорий. Для достижения цели были поставлены следующие задачи: 1. Изучить гистологическое строение печени и миокарда, и динамику некоторых морфометрических параметров этих тканей у ящерицы прыткой из разных местообитаний. 2. Определить биоиндикаторную ценность выбранных морфологических параметров печени и миокарда ящерицы прыткой. Объектом исследования служили печень и сердце ящерицы прыткой (Lacerta agilis Linnaeus, 1758). Половозрелых особей отлавливали в мае 2010–2011 гг. на территории Республики Марий Эл в трех местообитаниях: 1. ООПТ государственный природный заповедник ГПЗ «Большая Кокшага» (♂ – 8 особей и ♀ – 3 особи) – «условно чистая зона». 2. пгт. Куженер (♂– 4 особи и ♀– 6 особей). 3. Район Мясокомби- 190 ната г. Йошкар-Ола (только ♂ – 3 особи). Местообитания 2 и 3 – «антропогенно трансформированные зоны». Гистологическую обработку материала проводили по стандартным методикам (Трубачёва, Дробот, Абдрушевич, 2003). При помощи сетки Автандилова на световом микроскопе Micros MC – 40 M (Австрия) количественно оценивали площади функциональных зон в печени (гепатоциты) и миокарде (мышечные волокна), которые обозначали как паренхима. Для исследования использовали поле, содержащее 25 точек (Автандилов, 1990). Для определения толщины мышечных волокон и толщины стенки миокарда применяли винтовой окуляр-микрометр МОВ-1-15х ГОСТ 7865-56. Замеряли по 20 волокон и по 20 участков сердечной стенки (миокард) у каждой особи. Доли митотически делящихся, клеток с микроядрами, двуядерных, некротических и дистрофически измененных клеток определяли из расчета на 1000 клеток у одного животного. При статистической обработке материала оценивали однородность выборок, проводили двухфакторный дисперсионный анализ (Лакин, 1990). В печени ящерицы прыткой, обитающей во всех изученных местообитаниях гистологическая картина характеризовалась относительной нормой, хотя на препаратах встречались участки органа с неравномерным кровенаполнением. В печени ящериц всех групп выявлены клетки с вакуольной дистрофией, вплоть до баллонной формы. Данный вид дистрофии встречается в печени животных, обитающих на неблагополучных территориях (Акуленко, Гассо, Клименко, 2012). У ящерицы прыткой из района мясокомбината г. Йошкар-Олы в печени обнаружены очаги хронического гранулематозного воспаления. При морфометрии печени изучали показатели: доли митотических, двуядерных, некротических и дистрофических клеток, доли клеток с микроядрами, определяли площади паренхимы и стромы, рассчитывая коэффициент их соотношения. Предварительная оценка дисперсий с помощью тестов Кохрена, Хартли и Бартлетта показала их однородность (Р > 0,08). Из всех параметров печени существенные различия установлены только для долей двуядерных и некротических гепатоцитов, которые отличны у ящериц разных полов. Так у самок доли двуядерных клеток в паренхиме печени меньше (Р = 0,045), а доли некротических клеток больше (Р = 0,04), чем таковые у самцов. Возможно, это свидетельствует о большей чувствительности печени самок рептилий к повреждающим воздействиям либо о недостаточной выраженности у них компенсаторно-восстановительных процессов. При изучении срезов сердца вне зависимости от места отлова ящериц установлено типичное гистологическое строение органа, грубых 191 нарушений в его структуре не выявлено. Наряду с обычными мышечными волокнами в миокарде обнаружены гипертрофированные волокна. В данном случае проявляется мозаичность структуры миокарда, характерная для сердца позвоночных животных (Целлариус, Семенова, Непомнящих, 1980). Морфометрический анализ сердца ящерицы прыткой включал определение толщины мышечных волокон, толщины стенки желудочков сердца, определение площади паренхимы и стромы и коэффициента их соотношения в левых желудочке и предсердии. Предварительная проверка дисперсий по критериям Кохрена, Хартли и Бартлетта установила их однородность (Р > 0,09). Двухфакторный дисперсионный анализ коэффициента соотношения паренхимы к строме в желудочке сердца ящерицы прыткой показал, что оказывается значим фактор Местообитание (Р = 0,04), второй фактор – Пол и взаимодействие факторов не значимо (Р > 0,78). Величина данного коэффициента у животных из ГПЗ «Большая Кокшага» превышает таковую у ящериц из пгт. Куженер (Р = 0,04). Выводы: 1. В печени ящерицы прыткой, обитающей на антропогенно трансформированных территориях (пгт. Куженер и район Мясокомбината г. Йошкар-Ола), наиболее характерные морфологические изменения отмечены в сосудистом русле органа (неравномерное кровенаполнение крупных сосудов; эритростазирование капилляров) и его паренхиме (развитие очагов дистрофии, некрозов и гранулематозного воспаления). Из всех изученных морфометрических параметров печени ящерицы прыткой наиболее значимыми являются доли двуядерных и некротически измененных клеток, имеющие половые различия. 2. В тканевой структуре миокарда ящерицы прыткой всех изученных групп не установлено грубых нарушений. У ящериц, обитающих в пгт. Куженер, по сравнению с особями из ООПТ ГПЗ «Большая Кокшага», установлено сокращение площади функциональной паренхимы миокарда, что выражается в значимом уменьшении величины коэффициента соотношения паренхимы к строме (Р = 0,04). 3. Для биоиндикационного мониторинга качества среды наряду с гистологическим описанием структуры печени и миокарда рептилий можно использовать такие их морфометрические показатели как доли дистрофически и некротически измененных клеток (печень); коэффициент соотношения паренхимы к строме (миокард). Л и т е р а т ур а Автандилов Г. Г. Медицинская морфометрия / Г. Г. Автандилов. – М.: Медицина, 1990. – 218 с. Акуленко Н. М. Исследование печени земноводных и пресмыкающихся в целях определения степени антропогенного загрязнения биоценоза: возможности метода / Н. М. Акуленко, В. Я. Гассо, Е. Ю. Клименко // Вопросы герпетологии: материалы Пятого 192 съезда Герпетологического общества им. А. М. Никольского. 24–27 сентября 2012 г., Минск, Беларусь. – Минск: ИООО «Право и экономика», 2012. – С. 11–14. Акуленко Н. М. Амфибии и рептилии как индикаторы антропогенного загрязнения / Н. М. Акуленко, О. Д. Некрасова // Мониторинг природных и техногенных сред: материалы Всеукраинской науч. конф. – Симферополь: ДИАЙПИ, 2008. – С. 306–309. Дробот Г. П. Морфологические особенности мышечной ткани амфибий, обитающих на урбанизированных территориях / Г. П. Дробот, Н. В. Глотов, Л. В. Емельянова // Популяция, сообщество, эволюция: V Всерос. популяционный семинар 26–30 ноября 2001 г., Казань, – Казань: ЗАО «Новое знание», 2001. – Ч. 1. – С. 137–140. Дробот Г. П., Мальцева Н. Л., Ведерников А. А. Ответная реакция некоторых тканей лягушки озерной (Rana Ridibunda Pallas, 1771) на антропогенную нагрузку / Г. П. Дробот, Н. Л. Мальцева, А. А. Ведерников // Вестник Оренбургского государственного университета. – 2011. – № 12 (131). – С. 65–67. Дробот Г. П. Изучение ряда морфофункциональных особенностей ящериц, обитающих в промышленной зоне г. Йошкар-Олы / Г. П. Дробот, Т. В. Соловьева, Е. М. Васина, Н. В. Летунова, А. Р. Насибуллина // Экология города Йошкар-Олы: научное издание / Мар. гос. ун-т; отв. ред. О. Л. Воскресенская. – Йошкар-Ола, 2007. – С. 209–216. Дробот Г. П. Гистоморфометрические характеристики печени рыб как биоиндикаторы качества водной среды / Г. П. Дробот, В. С. Трубачева, О. В. Малюта, С. Г. Васин, О. И. Павловская // Вестник Марийского государственного технического университета. Сер., Лес, экология, природопользование. – 2011. – № 3 (13). – с. 101–107. Лакин Г. Ф. Биометрия / Г. Ф. Лакин. – М.: Высш. шк.. – 1990. – 350 с. Мильто К. Д. Земноводные и пресмыкающиеся Северо-Запада России: Оценка биоразнообразия: автореф. дис. … канд. биол. наук / К. Д. Мильто. – СПб., 2007. – С. 3–21. Трубачева В. С. Большой практикум по гистологической технике / Мар. гос. ун-т; В. С. Трубачева, Г. П. Дот, Л. И. Абдрушевич. – Йошкар-Ола, 2003. – 47 с. Целлариус Ю. Г. Морфологические типы изменений миофибрилл мышечных клеток сердца / Ю. Г. Целлариус, Л. А. Семенова, Л. М. Непомнящих // Архив патологии. – 1980. – № 12. – С. 3–14. МОНИТОРИНГ СОДЕРЖАНИЯ НЕФТЕПРОДУКТОВ В ПОЧВЕ РАЗНЫХ ЗОН Г. ЙОШКАР-ОЛЫ Иванова Р. Р., Малкова Т., Куфтина Н. И., Глаголев Д. Поволжский государственный технологический университет, г. Йошкар-Ола, ivanova.rufina@yandex.ru В последние десятилетия происходит быстрое развитие автотранспорта, увеличивается число автомобилей в личном пользовании и транспортных системах городов. В основном в качестве топлива для автомобилей используются нефтепродукты (бензин, керосин, мазут). Возрастание использования нефтепродуктов ведет к их распространению в окружающей среде и увеличению их содержания в почвах городских территорий. Рост содержания нефтепродуктов в почве урбанизированных территорий отмечают многие исследователи. Данная проблема актуальна и для г. Йошкар-Олы. Автомобилями заполнены дворы в жилых районах, где автомобили часто стоят вплотную к детским площадкам и на газонах, машины припаркованы к обочине улиц в центре и промышленной зоне. Автомобильный транспорт 193 в настоящее время является основным загрязнителем атмосферного воздуха города, его доля в общей массе выбросов составляет 85 %. При этом сами выбросы автотранспорта составляют 65 %, а 20 % – продукты возгонки и терморазложения масел, 9 % – испарения бензина и 6 % – продукты износа резины и металла (Воскресенская и др., 2007). Автотранспорт – также главный антропогенный источник загрязнения почвы урбанизированной территории нефтепродуктами. Исследование содержания нефтепродуктов в почве функционально различных территорий не проводилось, поэтому целью работы было изучение содержания нефтепродуктов в почве разных зон г. Йошкар-Олы. Объектами исследования была территория жилых районов, детских площадок в детских садах и дворах, промышленной зоны и зоны отдыха г. Йошкар-Олы. Фоновое содержание нефтепродуктов определяли в Центральном парке культуры и отдыха (ЦПКиО). На объектах были отобраны пробы почвы согласно ГОСТ 17.4.4.02-84 (1984). Определение содержания нефтепродуктов в почве проводили согласно Методике измерений массовой доли нефтепродуктов в пробах почв, грунтов, донных отложений, илов, осадков сточных вод, отходов производства и потребления гравиметрическим методом (Методика…, 2010). Результаты исследования содержания нефтепродуктов в почве представлены в таблице. Как видно из таблицы, фоновое содержание нефтепродуктов в ЦПКиО составило 18,09 мг/кг, в зоне отдыха лесопарке «Дубовая роща» – 189,60 мг/кг, лесопарке «Сосновая роща» – 50,10 мг/кг. Содержание нефтепродуктов в пробах почвы, взятых с функционально разных территорий, варьирует в пределах от 18,09 до 813,71 мг/кг. Содержание нефтепродуктов в почве разных зон г. Йошкар-Олы № пробы Объект исследования Содержание нефтепродуктов, мг/ кг Селитебная зона 1 ул. Машиностроителей, д. 8 813,71 2 ул. Анциферова, д. 15 460,90 3 ул. К. Либкнехта, д. 66 121,20 4 ул. Строителей, д. 32 540,40 5 ул. Эшкинина, д. 6 99,4 х±m 387,12 ± 152,36 Детские площадки во дворах жилых массивов 1 ул. Комсомольская, д. 92А 74,03 194 № пробы Объект исследования Содержание нефтепродуктов, мг/ кг 2 ул. Петрова, д. 13А 52,42 3 ул. З. Космодемьянской, д. 128 88,91 4 ул. Панфилова, д. 30 200,0 5 ул. Пролетарская, д. 71 225,00 6 ул. К. Маркса, д. 121 537,50 7 ул. Кирпичная, д. 5 37,50 х±m 173,66 ± 75,61 Детские площадки на территории детских садов 1 д/с № 76, ул. Петрова 46,70 2 д/с № 58, ул. Чехова 84,28 3 д/с № 7, ул. К. Либкнехта 76,32 4 д/с № 49, ул. Целинная 25,92 5 д/с № 44, ул. Лазо 45,03 6 д/с № 64, ул. Пролетарская 15,00 7 д/с № 13, переулок Заводской 280,00 х±m 81,88 ± 39,32 Промышленная зона 464,6 1 ЗАО «Йошкар-Олинский мясокомбинат» 2 ЗАО «Инструментальный завод» 402,5 3 ООО «Объединение Родина» 238,2 4 ОАО «Гардиан» 197,5 5 ОАО «Контакт» 190,2 6 ОАО «ММЗ» 277,7 7 ОКТБ «Кристалл» 146,8 8 ООО «Еврофуд» 93,2 х±m 251,30 ± 55,95 Зона отдыха 1 Дубовая роща 189,60 2 Сосновая роща 50,10 3 Центральный парк культуры и отдыха 18,09 Норматив для РМЭ 1000,00 195 Установлено, что содержание нефтепродуктов в почве всех обследованных территорий не превышает норматив, установленный для РМЭ на уровне 1000 мг/кг. В тоже время отмечаются различия в содержании нефтепродуктов в разных зонах города. Наиболее высокий уровень содержания нефтепродуктов в почве отмечен в селитебной зоне – 387,12 ± 152,36 мг/кг, особенно на ул. Машиностроителей (813,71 мг/кг). На втором месте по содержанию нефтепродуктов в почве – промышленная зона – 251,30 ± 55,95 мг/кг. В почве детских площадок во дворах жилых домов содержание нефтепродуктов варьировало от 37,50 до 537,50 мг/кг, а в почве детских площадок в детских садах – от 15,00 до 289,00 мг/кг. Наименее загрязненной нефтепродуктами зоной является территория лесопарка Сосновая роща (50,10 мг/кг) и ЦПКиО (18,09 мг/кг). При этом содержание нефтепродуктов в почве жилой зоны в 20 раз превышает уровень фона, и даже в почве детских садов отмечается превышение фонового содержания в 4 раза. При оценке степени загрязнения почвы нефтепродуктами мы руководствовались рекомендациями Пиковского (1993), который предлагает считать концентрации нефтепродуктов в почвах до 100 мг/кг фоновыми, экологической опасности для среды они не представляют. Концентрации от 100 до 500 мг/кг считать повышенным фоном. Нефтепродукты в таких количествах активно утилизируются микроорганизмами или вымываются дождевыми потоками без вмешательства человека. Загрязненными почвами можно считать почвы, содержащие более 500 мг/кг нефтепродуктов. При этом содержания от 500 до 1000 мг/кг относятся к умеренному загрязнению, от 1000 до 2000 – к умеренно опасному загрязнению, от 2000 до5000 мг/кг к сильному, опасному загрязнению, и свыше 5000 мг/кг к очень сильному загрязнению, подлежащему санации (Пиковский, 1993). Анализ показал, что к умеренно загрязненным территориям г. Йошкар-Олы среди обследованных объектов следует отнести дворы по ул. Машиностроителей (813,71 мг/кг) и ул. Строителей, д. 32 (540,40 мг/кг); детскую площадку во дворе по ул. К. Маркса, д. 121 (537,50 мг/кг). Промышленная зона и большинство детских площадок во дворах жилой зоны характеризуются как повышенный фон. А детские площадки на территории детских садов и зона отдыха соответствуют фону. Таким образом, проведенные исследования не выявили существенного загрязнения почвы города г. Йошкар-Олы нефтепродуктами, но в то же время отмечены различия в их содержании в разных городских зонах. Наиболее чистыми являются зоны отдыха и территории детских площадок, более загрязнены дворы селитебной зоны. 196 Л и т е р а т ур а Воскресенская О. Л. Экология города Йошкар-Олы / Мар. гос. ун-т; О. Л. Воскресенская и др. – Йошкар–Ола, 2007. – 230 с. ГОСТ 17.4.4.02-84. Охрана природы. Почвы. Методы отбора и подготовки почвы для химического, бактериологического, гельминтологического анализа. – М.: Изд-во стандартов, 1985. –11 с. Методика измерений массовой доли нефтепродуктов в пробах почв, грунтов, донных отложений, илов, осадков сточных вод, отходов производства и потребления гравиметрическим методом. – М., 2010. – 41 с. ОЦЕНКА ВОЗДЕЙСТВИЯ АЗС НА СОСТОЯНИЕ ОКРУЖАЮЩЕЙ СРЕДЫ В Г. ЙОШКАР-ОЛЕ Иванова Р. Р., Токарева Н. А. Поволжский государственный технологический университет, г. Йошкар-Ола, ivanova.rufina@yandex.ru Автомобильный транспорт в настоящее время бурно развивается, значительно повышая качество жизни человека, но рост количества автомобилей сопровождается негативными последствиями как для самого человека, так и для окружающей среды. Автотранспорт с одной стороны, потребляет из атмосферы кислород, а с другой – выбрасывает в нее отработавшие газы, картерные газы, оксиды углерода, оксиды свинца и другие вещества, тем самым влияя на окружающую среду, животный и растительный мир, в том числе и на человека. Выбросы автомобильного транспорта содержат около 300 вредных веществ, обладающих токсическим и нередко канцерогенным действием (Воскресенская, 2007 и др.). На сегодняшний день ни один автомобиль не может обойтись без топлива, которое обеспечивается автозаправочной станцией. Автозаправочные станции – это многофункциональный комплекс, оснащенный всем необходимым для качественного обслуживания, заправки и ремонта автотранспорта. Опасности, исходящие от автозаправочных станций, недооценить невозможно: повышенное загрязнение воздуха выхлопными газами и углеводородами, возникновение чрезвычайных ситуаций, сопровождающихся взрывами, пожарами и разливом топлива (Аксенов, 1986 и др.). Данная проблема актуальна и для города Йошкар-Олы, где количество автотранспорта и автозаправок в последние два десятилетия резко увеличилось. Для обслуживания автотранспортного потока в городе функционирует 36 АЗС, расположенных как в его центре, так и на окраинах. Сегодня автотранспорт – главный загрязнитель атмосферного воздуха города Йошкар-Олы, доля выбросов которого в общей массе выбросов составляет более 85 %. В городе существенно увеличилась 197 антропогенная нагрузка как на окружающую среду в целом, так и на ее отдельные компоненты. Целью работы была оценка воздействия автозаправочных станций на экологическое состояние территории АЗС г. Йошкар-Олы. Объектом исследования была территория АЗС, расположенных в разных районах города Йошкар-Олы: АЗС Лукойл на ул. Й. Кырля; АЗС Транс – Ойл, на ул. Прохорова; АЗС № 11 на Сернурском тракте; АЗС Лукойл – МарийЭл № 7 на Оршанском тракте; АЗС № 1 на ул. К. Маркса; АЗС Лукойл Волгонефтепродукт на ул. Красноармейская слобода; АЗС Лукойл № 9 на Кокшайском проезде. Контрольным объектом служил лесопарк «Дубовая роща», фоновое содержание нефтепродуктов определяли в Центральном парке культуры и отдыха (ЦПКиО). Для оценки состояния окружающей среды проводилось визуальное обследование территории АЗС с целью выявления соответствия требованиям СНиП 2.07.01-89 (1999), исследование возможной токсичности почвы методом биотестирования с использованием такого тест-объекта, как дафнии Daphna magna Straus (Методика…, 2006), определение содержания нефтепродуктов в почве территории АЗС (Методика…, 2010). Отбор проб почвы проводился согласно ГОСТ 17.4.4.02-84 (1984). Оценка воздействия АЗС на окружающую среду проводилась по состоянию почвы в санитарно-защитной зоне АЗС, т. к. почва в конечном итоге является накопителем веществ, поступающих в атмосферу и непосредственно на поверхность территории. Почва отражает предыдущие воздействия и может служить источником информации для интегральной оценки экологического состояния окружающей среды (Давыдова , 2002; Воскресенская и др., 2007). Результаты исследования показали, что все обследованные АЗС располагаются согласно СНиП 2.07.01-89* «Градостроительство. Планировка и застройка городских и сельских поселений» пункта Сооружения и установка для хранения и обслуживания транспортных средств, на допустимом расстоянии от жилых помещений 50 метров и более 100 метров от таких объектов, как больницы, поликлиники, детские сады, школы и др., что не создает угрозу безопасности населения. Непосредственно возле автозаправочных станций территория благоустроена, созданы клумбы с цветами, газоны, высажены деревья. На территории каждой АЗС размещены контейнеры для ТБО, вокруг объектов исследования замусоренности не выявлено. Отмечено также наличие на каждой заправке предупреждающих плакатов: «Не мусорить», «Не курить» и другие ограничительные знаки. 198 Исследование проб почвы с территории АЗС на возможную токсичность с помощью тест-объекта дафнии (показатели БКР – безвредная кратность разбавления и ЛКР – летальная кратность разбавления) показало, что на АЗС № 1 и АЗС № 9 почва соответствует IV классу опасности и оценивается, как возможное воздействие исследуемого фактора на окружающую среду. Все остальные пробы почвы V класса опасности – т. е. исследуемый фактор не создает угрозы для живых организмов. В контроле – Дубовой роще и ЦПКиО почва также соответствует V классу опасности. Результаты исследования содержания нефтепродуктов в почве АЗС представлены в таблице. Как видно из таблицы, фоновое содержание нефтепродуктов в ЦПКиО составило 18,09 мг/кг, на контрольном объекте лесопарке «Дубовая роща» – 189,60 мг/кг. При этом содержание нефтепродуктов в пробах почвы, взятых с территории АЗС, варьируется в пределах от 381,53 до 1077,11 мг/кг. Содержание нефтепродуктов в почве территории АЗС № пробы Объект исследования Содержание нефтепродуктов, мг/ кг 1 АЗС Лукойл 381,53 2 ЗАО Транс – Ойл 424,97 3 АЗС № 11 823,70 4 АЗС ЛУКОЙЛ – МАРИЙ ЭЛ № 7 736,94 5 АЗС № 1 881,80 6 ЛУКОЙЛ ВОЛГОНЕФТЕПРОДУКТ 811,00 7 АЗС № 9 1077,11 8 Дубовая роща (контроль) 189,60 9 Центральный парк культуры и отдыха 18,09 10 Норматив для РМЭ 1000,00 Наибольшее количество нефтепродуктов выявлено в пробе почвы, взятой на территории АЗС № 9 – 1 077,11 мг/кг, что превышает рекомендуемый уровень содержания нефтепродуктов в почве для РМЭ (1 000 мг/кг) в 1,1 раза. В пробах почвы с территории остальных обследованных АЗС содержание нефтепродуктов не превышает норматив, но в десятки раз превышает фоновое содержание в почве ЦПКиО и 2– 5 раз – содержание в почве контрольного объекта Дубовой рощи. Аналогичные уровни содержания нефтепродуктов в почве городов получены другими исследователями. Так в городах Самарской и Мос199 ковской областей в среднем по территории содержание нефтепродуктов варьируется от 5 до 5100 мг/кг. Таким образом, проведенные исследования не выявили резких нарушений состояния окружающей среды в зоне территории исследованных АЗС г. Йошкар-Олы, но в то же время отмечается тенденция к увеличению загрязнения почвы нефтепродуктами и повышению ее токсичности. Л и т е р а т ур а Аксенов И. Я. Транспорт и охрана окружающей среды / И. Я. Аксенов, В. И. Аксенов – М.: Транспорт, 1986. – 378 с. Воскресенская О. Л. Экология города Йошкар-Олы / Мар. гос. ун-т; О. Л. Воскресенская и др. – Йошкар-Ола, 2007. – 230 с. ГОСТ 17.4.4.02-84. Охрана природы. Почвы. Методы отбора и подготовки почвы для химического, бактериологического, гельминтологического анализа. – М.: Изд-во стандартов, 1985. – 11 с. Давыдова С. Л. Тяжелые металлы как суперэкотоксиканты ХХI века / С. Л. Давыдова – М., 2002. – 140 с. Методика определения токсичности водных вытяжек из почв, осадков, сточных вод и отходов, питьевой, сточной и природной воды по смертности тест-объекта Дафния – М.: Фед. служба по экологическому, технологическому и атомному надзору, 2006. – 32 с. Методика измерений массовой доли нефтепродуктов в пробах почв, грунтов, донных отложений, илов, осадков сточных вод, отходов производства и потребления гравиметрическим методом. – М., 2010. – 41 с. Строительные нормы и правила СНиП 2.07.01-89 «Градостроительство. Планировка и застройка городских и сельских поселений» (Утв. Постановлением Госстроя России от 25.08.1999). СРАВНИТЕЛЬНЫЙ ГЕНЕТИЧЕСКИЙ АНАЛИЗ РОДА ПНЕВМОЦИСТОВ С ТОЧКИ ЗРЕНИЯ МЕТАБОЛИЧЕСКИХ ПУТЕЙ Поролло А. А.1,2, Кушион М. Т.2,3 1 Научно-исследовательский центр при детском госпитале г. Цинциннати, США, alexey.porollo@cchmc.org 2 Университет Цинциннати, г. Цинциннати, США, melanie.cushion@uc.edu 3 Медицинский центр для ветеранов, г. Цинциннати, США Пневмоцисты (Pneumocystis genus) представляют собой род малоизученных грибов-аскомицетов. Все члены этого рода – паразитические грибы, живущие в легких млекопитающих, в том числе и человека. У людей с ослабленной иммунной системой (больных СПИДом, перенесших трансплантацию органов или проходящих терапии, понижающие иммунитет) пневмоцисты начинают активно размножаться, заполняют альвеолярное пространство и вызывают пневмоцистую пневмонию (пневмоцистоз), которая при неадекватном лечении может привести к легочной недостаточности и, как следствие, летальному исходу. Пневмоцисты нечувствительны к широко используемым антигрибковым препаратам на основе производных азола. Несмотря на несколь200 ко десятилетий исследований, репертуар препаратов для лечения пневмоцистоза остается очень ограниченным и состоит из триметопримсульфаметоксазола и пентамидина. Первый бинарный препарат может иметь ограниченное применение у людей с выраженной реакцией на сульфамидные препараты. Кроме того, наблюдается возрастающая устойчивость пневмоцистов к этому препарату (Nahimana et al., 2004). Второй препарат обладает повышенной токсичностью и не рекомендуется к применению больным СПИДом. Отсутствие новых лекарств для лечения пневмоцистоза связано, в первую очередь, с недостаточным пониманием биологии этого организма. Более того, до сих пор не удается вырастить культуру этого гриба: по извлечении патогена из легких организма-хозяина грибная культура живет максимум 7–10 дней. В результате невозможно проводить полноценные скрининги на новые лекарства и выполнять какие-либо генетические манипуляции с этим организмом для изучения его биологии и разработки новой таргетной терапии. Появление технологий нового поколения для секвенирования геномов позволило получить 3 генома представителей пневмоцистов: Pneumocystis jirovecii (вид, инфицирующий людей), Pneumocystis carinii (вид, инфицирующий крыс) и Pneumocystis murina (вид, инфицирующий мышей). Ранее мы провели анализ строения генов в геноме P. carinii на предмет распределения количества и длины интронов (Slaven et al., 2006), охарактеризовали экспрессирующие гены (Cushion et al., 2007), и провели поиск генов аналогичных известным таргетам антигрибковых препаратов для других грибковых инфекций (Porollo et al., 2012). В данной работе нами представлен сравнительный анализ указанных выше геномов для выявления метаболических путей, обуславливающих паразитический характер грибов рода пневмоцистов, а также для идентификации общих для всех и специфичных для каждого вида энзимов для последующего определения круга потенциальных таргетов при разработке новых лекарств. Геномы трех видов пневмоцистов были получены из следующих источников: P. jirovecii – NCBI Genome Database http://www. ncbi. nlm. nih. gov/genome?LinkName=pubmed_genome&fr om_uid=23269827 P. carinii – Pneumocystis Genome Project http://pgp. cchmc. org/ P. murina – Pneumocystis murina Database http://www. broadinstitute. org/annotation/genome/Pneumocystis_group/ MultiHome. html 201 Для поиска гомологии между ДНК секвенированных пневмоцистов и известными последовательностями энзимов использовалась программа SHARKhunt (Pinney et al., 2005) с 3412 профилями различных ферментов, полученных из базы данных PRIAM (Claudel-Renard et al., 2003). Уровень гомологии при поиске генов был установлен по критерию E < 10–6. Соотнесение энзимов к метаболическим путям было выполнено в соответствии с описанием биологических путей в базе данных KEGG Pathway. Поиск по гомологии генов, кодирующих ферменты, дал следующие результаты: 317, 329 и 334 вероятных генов в P. jirovecii, P. carinii и P. murina, соответственно. Диаграмма Венна на рисунке 1 иллюстрирует общее распределение обнаруженных ферментов между данными тремя видами пневмоцистов. Рисунок 1 – Суммарное распределение идентифицированных ферментов в секвенированных геномах пневмоцистов Подавляющее большинство генов (275) оказалось общим между тремя видами пневмоцистов, что является дополнительным подтверждением, что три генома, полученных в независимых исследовательских лабораториях с применением разных технологий секвенирования и алгоритмов последующей сборки фрагментов ДНК, были собраны в целом правильно. Кроме того, виды, инфицирующие грызунов (P. carinii и P. murina), имеют больше общих генов, чем с видом, инфицирующим людей (P. jirovecii). В то же время, каждый из проанализированных видов имеет уникальные энзимы: 21 в P. jirovecii и по 25 в P. carinii и P. murina, соответственно. Вероятно, эти индивидуальные для каждого вида гены участвуют в предопределении видового разнообразия в роде пневмоцистов и адаптации каждого вида паразита к особенностям вида своего хозяина, и потому представляют отдельный интерес для будущих исследований биологии этих организмов. В таблице приведены выборочно (в виду ограничений по размеру текста) метаболические пути для иллюстрации суммарного распределе202 ния обнаруженных ферментов в геномах пневмоцистов. Для сравнения там же приведены ферменты из генома таксономически близких, но свободно живущих делящихся дрожжей (Schizosaccharomyces pombe). Суммарное распределение ферментов в секвенированных пневмоцистах в сравнении с делящимися дрожжами (S. pombe). Обозначения: Pc = P. carinii, Pm = P. murina, Pj = P. jirovecii, Sp = S. pombe Категория / Биологический путь (KEGG Pathway) Sp Pc Pm Pj Метаболизм аминокислот Alanine, aspartate and glutamate metabolism Glycine, serine and threonine metabolism Cysteine and methionine metabolism Valine, leucine and isoleucine degradation Valine, leucine and isoleucine biosynthesis Lysine biosynthesis Lysine degradation Arginine and proline metabolism Histidine metabolism Tyrosine metabolism Phenylalanine metabolism Tryptophan metabolism Phenylalanine, tyrosine and tryptophan biosynthesis 21 22 27 8 7 13 8 29 12 13 8 9 20 11 11 9 5 0 1 4 8 1 1 2 8 10 12 10 8 8 0 0 4 8 1 1 3 7 8 12 10 7 7 1 0 3 9 1 1 2 2 10 Метаболизм углеводов Glycolysis / Gluconeogenesis Citrate cycle (TCA cycle) Pentose phosphate pathway Pentose and glucuronate interconversions Fructose and mannose metabolism Galactose metabolism Starch and sucrose metabolism Amino sugar and nucleotide sugar metabolism Inositol phosphate metabolism Pyruvate metabolism Glyoxylate and dicarboxylate metabolism Propanoate metabolism Butanoate metabolism 24 16 15 4 13 11 14 17 10 18 10 12 8 17 16 11 3 10 8 16 15 12 11 9 6 3 16 15 10 3 10 5 16 12 12 14 9 5 4 15 14 11 3 11 6 14 14 11 11 8 5 4 Энергетический метаболизм Oxidative phosphorylation Methane metabolism Nitrogen metabolism Sulfur metabolism 9 13 11 14 9 10 11 3 10 11 12 2 10 9 11 3 203 Категория / Биологический путь (KEGG Pathway) Метаболизм липидов Fatty acid biosynthesis Fatty acid elongation Fatty acid degradation Synthesis and degradation of ketone bodies Steroid biosynthesis Glycerolipid metabolism Glycerophospholipid metabolism Ether lipid metabolism Arachidonic acid metabolism Linoleic acid metabolism Sphingolipid metabolism Biosynthesis of unsaturated fatty acids Sp Pc Pm Pj 5 6 5 2 16 15 29 10 7 5 8 6 4 2 2 2 8 6 11 4 2 1 6 3 4 3 2 2 8 7 13 5 1 2 5 4 3 2 2 2 10 6 11 4 0 1 7 3 Как видно из таблицы, все пневмоцисты имеют значительно меньшее число ферментов в биологических путях метаболизма аминокислот по сравнению с делящимися дрожжами, что наверняка является одним из факторов определения их паразитической природы, то есть необходимостью заимствования аминокислот у хозяина. Эта недостаточность была впервые отмечена в работе группы Хаузера, которая первая секвенировала полный геном P. jirovecii (Hauser et al., 2010), и теперь подтверждена в других пневмоцистах нашим анализом. Интересным представляется тот факт, что у видов, инфицирующих грызунов, имеются 5 общих дополнительных энзимов по сравнению с P. jirovecii. Метаболизм углеводов представлен значительно лучше и почти полностью соответствует составу ферментов этой категории в делящихся дрожжах. Однако, особо следует отметить два биологических пути, метаболизм крахмала и сахарозы (Starch and sucrose metabolism) и метаболизм фосфоинозитола (Inositol phosphate metabolism), где пневмоцисты обладают дополнительными ферментами по сравнению с S. pombe, что может указывать на дополнительную зависимость паразитических организмов от доступа поли-, ди- и моносахаридов. Энергетический метаболизм практически идентичен делящимся дрожжам, за исключением метаболизма серы (Sulfur metabolism), который почти отсутствует в пневмоцистах. Большинство биологических путей в категории метаболизма липидов имеют меньшее число ферментов по сравнению с делящимися дрожжами. В этом контексте, следует особо выделить биосинтез стероидов (Steroid biosynthesis). Рисунок 2 показывает схему этого пути с ферментами, выделенными серыми квадратами для S. pombe и указан204 ными стрелками для проанализированных видов пневмоцистов. Пневмоцисты не имеют полной цепочки синтеза эргостерина, что может объяснить их нечувствительность к ингибиторам этого пути (основная цель антигрибковых препаратов на основе производных азола). Ранее эспериментально было показано, что, в отличие от других грибов, пневмоцисты не имеют эргостерин, а используют холестерин, который вероятно заимствуется у организмов-хозяинов (Kaneshiro, 2004). Рисунок 2 – Схема биосинтеза стероидов с помеченными ферментами, которые были обнаружены в пневмоцистах и делящихся дрожжах. Схема пути представляет модификацию оригинальной версии в KEGG Pathway: убрана цепочка синтеза фитостерола как неотносящегося к рассматриваемой биологической системе 205 Таким образом, данная работа представляет детальный сравнительный анализ недавно секвенированных геномов рода пневмоцистов с точки зрения метаболических путей. Показано, что большинство ферментов являются общими среди проанализированных видов, но имеется небольшое количество уникальных ферментов для каждого вида. Проведенная работа служит основанием для проверки гипотез о причинах паразитического характера рассмотренных грибов и выявления целей для разработки новых препаратов против пневмоцистоза. Л и т е р а т ур а Claudel-Renard C., Chevalet C., Faraut T., Kahn D., 2003. Enzyme-specific profiles for genome annotation: PRIAM. Nucleic Acids Res 31, 6633–6639. Cushion M. T., Smulian A. G., Slaven B. E., Sesterhenn T., Arnold J., Staben C., Porollo A., Adamczak R., Meller J., 2007. Transcriptome of Pneumocystis carinii during fulminate infection: carbohydrate metabolism and the concept of a compatible parasite. PLoS One 2, e423. Hauser P. M., Burdet F. X., Cisse O. H., Keller L., Taffe P., Sanglard D., Pagni M., 2010. Comparative genomics suggests that the fungal pathogen pneumocystis is an obligate parasite scavenging amino acids from its host's lungs. PLoS One 5, e15152. Kaneshiro E. S., 2004. Sterol metabolism in the opportunistic pathogen Pneumocystis: advances and new insights. Lipids 39, 753–761. Nahimana A., Rabodonirina M., Bille J., Francioli P., Hauser P. M., 2004. Mutations of Pneumocystis jirovecii dihydrofolate reductase associated with failure of prophylaxis. Antimicrob Agents Chemother 48, 4301–4305. Pinney J. W., Shirley M. W., McConkey G. A., Westhead D. R., 2005. metaSHARK: software for automated metabolic network prediction from DNA sequence and its application to the genomes of Plasmodium falciparum and Eimeria tenella. Nucleic Acids Res 33, 1399–1409. Porollo A., Meller J., Joshi Y., Jaiswal V., Smulian A. G., Cushion M. T., 2012. Analysis of Current Antifungal Agents and Their Targets within the Pneumocystis carinii Genome. Curr Drug Targets 13, 1575–1585. Slaven B. E., Porollo A., Sesterhenn T., Smulian A. G., Cushion M. T., Meller J., 2006. Large-scale characterization of introns in the Pneumocystis carinii genome. J Eukaryot Microbiol 53 Suppl 1, S. 151–153. ИСПОЛЬЗОВАНИЕ ДРЕВЕСНЫХ РАСТЕНИЙ В ЛОКАЛЬНОМ МОНИТОРИНГЕ СОСТОЯНИЯ ОКРУЖАЮЩЕЙ СРЕДЫ г. ЙОШКАР-ОЛЫ Сарбаева Е. В., Воскресенский В. С. Марийский государственный университет, г. Йошкар-Ола, sarbaevaev@mail.ru Высокий уровень урбанизации городов, рост количества автотранспорта и интенсификация промышленного производства с каждым годом усиливают прессинг на природную среду, резко снижают качество городской среды. Система озеленения городов является обязательным условием создания комфортных условий для населения. Древесные растения – самые крупные и долговечные элементы данной системы, они наиболее эффективны как источники кислорода, фитонцидов, агенты 206 оптимизации светового и температурного режимов, уловители техногенных газов и аэрозолей. Изменение состояния растений в городской среде и различные интегральные показатели, характеризующие древесные растения в условиях города, могут быть использованы для оценки и контроля качества городской среды, осуществляемых в рамках локального мониторинга. Биологический мониторинг окружающей среды возможен по функциональным (прирост биомассы в единицу времени, изменение окраски листьев, аккумуляция продуктов техногенеза живыми организмами, ход биохимических и физиологических процессов) и структурным (численность и состав видов) признакам. Информация лишь о концентрации загрязняющих веществ в воздухе, почве и органах растений не отражает реальную нагрузку на биоценоз и не позволяет прогнозировать возможные эффекты их воздействия без изучения ответной реакции всех структурных элементов растительного сообщества (Павлов, 2006). Многие древесные растения являются довольно чувствительными к действию различных атмосферных токсикантов, поэтому некоторые из них могут применяться в качестве тест-объектов в экологическом мониторинге. Чувствительные фитоиндикаторы указывают на присутствие загрязняющего вещества в воздухе или почве ранними морфологическими реакциями – изменением окраски листьев (появление хлорозов; желтая, бурая или бронзовая окраска), различной формы некрозами, преждевременным увяданием и опаданием листвы. У многолетних растений загрязняющие вещества вызывают изменение размеров, формы, количества органов, направления роста побегов или изменение плодовитости. Подобные реакции обычно неспецифичны (Биологический контроль…, 2007). Также для осуществления мониторинга городской среды целесообразно применение следующих показателей: индекса жизненности, содержания серы, оводненности тканей, активности пероксидазы, каталазы и других признаков, наиболее полно отражающих состояние растений. В течение ряда лет нами проводился локальный мониторинг состояния окружающей среды (на территории г. Йошкар-Олы) с помощью древесных растений (туя западная – Thuja occidentalis L., ель обыкновенная – Picea abies (L.) Karst., ель колючая – Picea pungens Engelm., сосна обыкновенная – Pinus silvestris L., липа сердцевидная – Tilia cordata Mill., береза повислая – Betulla pendula Roth.). Древесные растения в условиях урбанизированной среды испытывают значительные колебания физиологических параметров в зависимости от условий произрастания. Растения промышленных районов г. Йошкар-Олы характеризуются меньшей оводненностью тканей по 207 сравнению с растениями селитебных районов и пригородной зоны. Так, у особей T. cordata общее содержание воды в листьях снижалось с 45,6 до 36,4 %, у P. silvestris – с 47,2 до 38,6 %. Остальные виды характеризовались меньшей вариабельностью данного признака. При оценке водоудерживающей способности тканей изучаемых растений было выявлено, что максимальные потери воды из тканей наблюдались у растений промышленной зоны: у P. silvestris – до 15,33 %, у T. occidentalis – до 14,00 %, у T. cordata – 10,32 %. У всех изученных видов отмечалось повышение активности пероксидазы при увеличении загрязнения окружающей среды. Так, у T. occidentalis отмечен рост активности указанного фермента с 6,6 в селитебной зоне до 8,9 ΔД 670 г -1·с-1 в промышленной зоне г. ЙошкарОлы; аналогичная тенденция обнаружена у особей P. pungens и P. silvestris: активность пероксидазы у данных растений в более загрязненном районе исследования возрастала почти в 2,5 раза, а у B. pendula и T. cordata – в 1,7–1,9 раз. Максимальное увеличение активности пероксидазы (более чем в 3 раза) отмечено у особей P. abies, произраставшей в промышленном районе г. Йошкар-Олы. Активность каталазы у всех изученных растений по мере увеличения антропогенной нагрузки имела тенденцию к снижению: у B. pendula и T. cordata каталазная активность была максимальной и у растений промышленной зоны г. Йошкар-Олы снижалась лишь на 30–40 %. В то же время у хвойных растений (T. occidentalis, P. pungens P. abies и P. silvestris), исследованных в районе с высоким уровнем автотранспортного потока и функционирования промышленных предприятий, активность каталазы снижалась практически в 2 раза. Известно, что комплекс городских антропогенных факторов (трансформация природной среды, ее загрязнение, нарушение правил создания и содержания городских насаждений и др.) и влияние ряда природных факторов на древесные растения (вредители, болезни, экстремальные погодные условия и др.) снижают устойчивость и нарушают нормальную жизнедеятельность растений в городах. Вначале ослабление растений проявляется на физиолого-биохимическом уровне, затем распространяется на ультраструктурный и клеточный уровни и лишь после этого развиваются видимые признаки ослабления – хлорозы и некрозы тканей листа, уменьшение их размеров и преждевременное опадание, торможение роста растений, усыхание ветвей и побегов и др. (Мальхотра, Хан, 1988). В связи с этим особое значение приобретает диагностика состояния древесных насаждений с помощью физиологобиохимических показателей, которые не только дают возможность выявить признаки угнетения растений на ранних стадиях воздействия фи208 тотоксикантов, но и позволяют судить об уровне загрязнения окружающей среды. Наши исследования позволили предположить, что среди изученных видов наиболее чувствительными являлись хвойные растения местной флоры – P. abies и P. silvestris, у которых рассмотренные физиологические показатели оказались более вариабельными, чем у иных видов. В то же время лиственные растения (B. pendula и T. cordata), а также хвойные растения-интродуценты (T. occidentalis, P. pungens) в условиях городской среды проявляли большую стабильность физиологических параметров и оказались более устойчивыми видами. Тем не менее они также обладают определенным диапазоном изменчивости показателей, обладающих высокой информативностью при экологическом мониторинге и могут служить биоиндикаторами загрязнения окружающей среды в урбанизированных экосистемах. Исследование выполнено при поддержке ФЦП, соглашение № 14.B37.21.1111 «Экологические аспекты функционального состояния растений в условиях городской среды» Л и т е р а т ур а Павлов И. Н. Биологический мониторинг техногенного загрязнения по морфометрическим показателям древесных растений / И. Н. Павлов // Фундаментальные исследования. Физико-математические науки. – 2006. – № 8. – С. 34–37. Биологический контроль окружающей среды: биоиндикация и биотестирование / О. П. Мелехова, Е. И. Егорова, Т. И. Евсеева и др.; под ред. О. П. Мелеховой и Е. И. Егоровой. – М.: Академия, 2007. – 288 с. Мальхотра С. С. Биохимическое и физиологическое действие приоритетных загрязняющих веществ / С. С. Мальхотра, А. А. Хан // Загрязнение воздуха и жизнь растений. – Л., 1988. – С. 141–190. СОСТОЯНИЕ НАСАЖДЕНИЙ НА ТЕРРИТОРИЯХ ОБЩЕГО ПОЛЬЗОВАНИЯ ГОРОДА ЙОШКАР-ОЛЫ Соловьева О. С., Кудрявцева А. Е., Васильева В. К. Поволжский государственный технологический университет, г. Йошкар-Ола, sol-oxana@yandex.ru, anjutka90@inbox.ru, monsun-91@mail.ru Зеленые насаждения являются важным элементом городской среды. При озеленении городских территорий зеленые насаждения осуществляют многие функции, улучшающие среду проживания людей: − санитарно-гигиенические; − защитные, являясь барьером между проезжей частью и пешеходными тротуарами, обеспечивают безопасность людей, защищают от действия ветра, перегрева, снижают уровень шума; 209 − психологического воздействия, влияя на органы чувств человека окраской листьев, цветков, плодов древесных растений, их ароматом; − эстетические, украшая улицы, бульвары, скверы, парки, подчеркивая архитектурный стиль застройки (Зарубин, 1986). В настоящее время многие древесные растения, используемые в озеленении города Йошкар-Олы, достигли возраста физиологического старения. Изменяется видовой и возрастной состав городских насаждений в результате антропогенного изменения внешних условий. Площадь, занятая зелеными насаждениями в городах и вокруг них, постоянно сокращается за счет бесконтрольных вырубок, пожаров, постоянного воздействия вредных химических веществ и погодных условий. Мониторинг состояния древесно-кустарниковых растений на объектах общего пользования г. Йошкар-Олы показал, что большая часть (78 %) находится в удовлетворительном и неудовлетворительном состоянии, т. е. это растения с неправильно или слаборазвитой кроной, с различной степенью повреждениями, ранениями, дуплами и т. д. Для определения состояния древесных растений, произрастающих на территориях общего пользования города Йошкар-Олы, проведена инвентаризация зеленых насаждений. Сводные данные по количеству усыхающих и сухостойных древесных растений представлены в таблице. Сводный анализ состояния насаждений, произрастающих на территории г. Йошкар-Олы Количество усыхающих Количество усохших Наиболее распространенные виды деревьев и кустарников шт. % шт. % шт. % 1 2 3 4 5 6 7 Количество Деревья Ель колючая 632 2,24 0 0,00 3 0,47 Лиственница сибирская 426 1,51 1 0,23 17 3,99 Ель европейская 340 1,20 2 0,59 24 7,06 Липа мелколистная 9944 35,23 16 0,16 41 0,41 Береза повислая 5333 18,89 31 0,58 129 2,42 Рябина обыкновенная 2109 7,47 35 1,66 134 6,35 Клен американский 1666 5,90 4 0,24 3 0,18 Клен остролистный 1246 4,41 4 0,32 2 0,16 Клен ясенелистный 1050 3,72 1 0,10 0 0,00 Яблоня ягодная 869 3,08 4 0,46 21 2,42 210 Количество усыхающих Количество усохших Наиболее распространенные виды деревьев и кустарников шт. % шт. % шт. % 1 2 3 4 5 6 7 Тополь бальзамический Количество 855 3,03 3 0,35 28 3,27 Всего деревьев: 28229 82,32 106 0,38 539 1,91 Пузыреплодник калинолистный 1123 18,53 10 0,89 9 0,80 Карагана древовидная 651 10,74 1 0,15 0 0,00 Сирень обыкновенная 583 9,62 0 0,00 1 0,17 Боярышник кровавокрасный 580 9,57 1 0,17 13 2,24 Акация желтая 460 7,59 0 0,00 3 0,65 Вишня обыкновенная 393 6,48 0 0,00 2 0,51 Можжевельник казацкий 208 3,43 0 0,00 8 3,85 Всего кустарников: 6061 17,68 9 0,15 99 1,63 Всего деревьев и кустарников: 34290 100 115 0,34 638 1,86 Кустарники Основываясь на данных проведенной инвентаризации зеленых насаждений, произрастающих на улицах города Йошкар-Олы, можно сделать нижеизложенные выводы. По жизнеспособности наблюдается среди проанализированных 34 290 деревьев и кустарников 115 шт. в усыхающем состоянии – 0,34 %, 638 шт. в усохшем состоянии – 1,86 %, среди которых большую часть составляют: береза повислая, рябина обыкновенная, липа мелколистная, ель европейская, туя западная, боярышник кроваво-красный, свидина белая. Наибольшее количество усохших и усыхающих деревьев наблюдается в следующих районах: − территория бульвара Чавайна – 82 дерева в усохшем состоянии, 26 деревьев в усыхающем; − ул. Красноармейская Слобода – 75 деревьев при достаточно узком видовом разнообразии, из них 32 березы повислой и 43 дерева туи западной; − ул. Машиностроителей – 13 деревьев в усохшем состоянии, большинство из которых – липа мелколистная (6 шт.); 211 − ул. Мира – 13 деревьев в усыхающем состоянии, большинство из которых – липа мелколистная (10 шт.); − ул. Осипенко – 6 деревьев и 4 кустарника пузыреплодника в усохшем состоянии; − пр. Гагарина – 13 деревьев в усохшем состоянии, большая доля из которых приходится на рябину обыкновенную; − ул. Павленко – 5 деревьев в усохшем состоянии; − ул. Первомайская – 12 деревьев и 1 кустарник в усохшем состоянии; − ул. Пролетарская – 1 дерево и 8 кустарников в усохшем состоянии, среди которых – боярышник кроваво-красный и акация желтая; − ул. Петрова – 21 дерево в усохшем состоянии; − ул. Прохорова – 26 деревьев в усохшем состоянии, из которых значительная доля приходится на березу повислую, 3 дерева в усыхающем состоянии; − ул. Советская – 41 дерево в усохшем состоянии, из них значительная доля приходится на рябину обыкновенную; − ул. Соловьева – 5 деревьев в усохшем состоянии; − ул. Строителей – 8 деревьев в усыхающем состоянии и лишь одно дерево в усохшем; − ул. Суворова – 4 дерева в усохшем состоянии и 3 дерева в усыхающем; − ул. Эшкинина – 12 деревьев березы повислой и 1 кустарник калины обыкновенной в усохшем состоянии; − ул. Эшпая – 7 деревьев и 9 кустарников в усохшем состоянии; − по ул. Авиации 1 дерево липы мелколистной и 7 кустарников можжевельника обыкновенного в усохшем состоянии; − ул. Баумана – 15 деревьев в усохшем состоянии; − бульвар Победы – 21 дерево в усохшем состоянии; − ул. Воинов-Интернационалистов – 12 деревьев и 35 кустарников усохшие; − ул. Героев Сталинградской битвы – 4 дерева и 2 кустарника в усохшем состоянии; − ул. Карла Либкнехта – 6 деревьев в усохшем состоянии и 7 в усыхающем; − ул. Карла Маркса – 5 деревьев в усохшем состоянии; − ул. Кирова – 4 дерева в усохшем состоянии; − ул. Кирпичная – 17 деревьев тополя бальзамического в усохшем состоянии; 212 − ул. по Козьмодемьянскому тракту – 7 деревьев в усохшем состоянии; − Ленинский проспект – 12 деревьев и 7 кустарников в усохшем состоянии. На фоне всего многообразия древесно-кустарниковой растительности Парка культуры и отдыха ХХХ-летия ВЛКСМ 18 деревьев находится в усохшем состоянии, 9 деревьев в усыхающем, 9 кустарников в усохшем состоянии. На территории сквера Наты Бабушкиной в озеленении использованы липа мелколистная, рябина обыкновенная, ель европейская, береза повислая, яблоня лесная, барбарис обыкновенный, бузина черная, среди которых 24 дерева в усохшем состоянии, большая доля из которых приходится на рябину обыкновенную. На территории Юбилейной площади распространены береза повислая, черемуха обыкновенная, яблоня лесная, ель колючая, сирень обыкновенная, вишня обыкновенная. Среди всего многообразия используемых видов 13 деревьев и один кустарник находятся в усыхающем состоянии, где значительную долю составляет рябина обыкновенная. Сквер по ул. Пушкина отличается также большим разнообразием видов используемой древесно-кустарниковой растительности, среди которых доля усохших деревьев незначительная, что составляет 4 дерева. На территории сквера на Кокшайском проезде среди всего многообразия 17 деревьев и 8 кустарников в усохшем состоянии, большую долю среди которых составляют ель европейская и боярышник кровавокрасный. По данным отчета по работе Комитета экологии и природопользования только за 1 год рассмотрены с проведением экологической оценки заявки на проведение сноса и реконструкции 2116 деревьев, 568 кустарников, живой изгороди 434,5 п. м, 46865,98 м2 естественного травяного покрова и 4163 м2 газона по причине строительства объектов, ремонта подземных коммуникаций, также процесс усыхания после засухи все еще продолжается. С каждым годом площадь зеленых насаждений нашего города уменьшается в связи с активной застройкой и тем, что многие насаждения физиологически старые и не выдерживают постоянного воздействия антропогенной нагрузки, поэтому необходим регулярный контроль и соблюдение основных агротехнических мероприятий по созданию и уходу за зелеными насаждениями. Для этого необходима разработка и принятие соответствующих нормативных правовых актов. В городском округе «Город Йошкар213 Ола» в настоящее время отсутствуют правила создания и содержания зеленых насаждений. Контроль в области охраны окружающей среды должен осуществляться в целях обеспечения выполнения в процессе хозяйственной и иной деятельности мероприятий по охране окружающей среды, рациональному использованию и восстановлению природных ресурсов, а также в целях соблюдения требований в области охраны окружающей среды, установленных законодательством в области охраны окружающей среды. Л и т е р а т ур а Зарубин Г. П. Гигиена города / Г. П. Зарубин, Ю. В. Новиков. – М.: Медицина, 1986. – 272 с. ВЛИЯНИЕ КОЛЕБАНИЯ УРОВНЯ СЕВЕРНОГО МАЛОГО АРАЛА НА ЭКОЛОГИЧЕСКУЮ ОБСТАНОВКУ В РЕГИОНЕ Шынбергенов Е. А., Сиханова Н. С.1, Нургизаринов А. М.2 1 Казанский (Приволжский) федеральный университет, г. Казань, muhtasar_08@mail.ru 2 Кызылординский государственный университет имени Коркыт Ата, г. Кызылорда Приаралье, пережив непоправимые потери, утратило свое природнохозяйственное значение, и ради сохранения природы региона было решено спасти хотя бы небольшую часть Аральского моря. В природоохранных целях в августе 2005 года была сооружена Кокаральская плотина, отделяющая Северное Аральское море от Большого моря. Экологический и социально-экономический эффекты наблюдали в том же году, с момента окончания строительства. Огромный объем пресной воды Сырдарьи, который уходил в Большое море, стал собираться в Северном Малом море. Как и предполагалось, результаты, достигнутые путем разделения Северного Аральского моря, сохранились до конца года. Это – уровень зеркала моря (Балтийская система-БС), достигший +4,0 метров, это – увеличение зеркала моря на +874,0 км2. Плотина принесла и экономическую выгоду: в 2006 году в Северном море было выловлено 2,3 тыс. т рыбы, что в 10 раз больше, чем в 2005 году. Некоторые заброшенные приморские населенные пункты ожили, стало развиваться рыболовство и животноводческое хозяйство. Уменьшились соляные ветры, пылесброс в атмосферу, восстановился микроклимат. Рабочий уровень Кокаральской плотины рассчитан на 40,0 м, если эта тенденция сохранится, обьем воды, собирающейся в Северном Малом море, не будет превышать 21,0 км3. Для Малого моря этого недостаточ214 но, потому что из четырех частей Северного Арала полноводие будет досигнуто только в Центральной части и в заливе Шевченко, а югозападные окраины заливов Бутакова и Сарышыганак, лежащие выше указанной отметки, будут маловодными. Останутся вдали от моря густонаселенный город Аральск и приморские населенные пункты (Карашалан, Бугень, Каратуп, Тастубек, Акеспе, Акбасты). Следовательно, Кокаральская плотина не может полностью решить экологические проблемы. После сдачи в эксплуатацию Кокаральской плотины в 2005 году ожидалось, что вся вода реки Сырдарьи, впадающей в Северный Арал, останется в самом море. Минеральность морской воды должна была уменьшиться. Так как с завершением проекта масса воды Малого моря увеличится, уровень соли уменьшится с прежних 23,0 г/л до 17,0 г/л, следовательно, вода в Северном море станет более опресненной. Наши наблюдения подтвердили это предположение (табл.). Минеральный состав воды Аральского моря, 2009 год Моря ррН Жесткость, мг-экв НСО3 Мг/литр Cl SO4 Ca Mg Na+К твердый остаток Северное Аральское море 7,45 90,0 183,0 7090,0 3789,09 600,0 729,6 4797,5 16090,0 Большое Аральское море 7,45 93,0 256,2 13612,8 3622,8 580,0 772,2 9267,5 26512,0 В таблице можно видеть, что минеральные показатели вод двух морей разные. Соленость (твердый остаток) Большого Аральского моря 26,5 г/л соответствует уровню 2000 года. Соленость Северного Аральского моря – 16,1 г/л, на 10,4 г/л меньше показателей Большого моря. Поскольку основную массу воды Малого моря составляет речная вода, то здесь отмечена и меньшая концентрация соли. Воды обоих морей имеют слабую щелочную реакцию – рН = 7,45. Жесткость морской воды соответственно 90 и 93 мг-экв., такая жесткость свойственна только морской воде. Жесткость воды определяется концентрациями солей кальция (Са) и магния (Mg). В водах Малого и Большого морей концентрации солей щелочных металлов близки: в водах Малого моря Са – 600 мг/л и Mg – 729,6 мг/л, в Большом море соответственно 580,0 и 772,2 мг/л, это очень высокие показатели. Если жесткость пресной воды превышает 12–15 мг-экв., то ее запрещают использовать в хозяй215 стве, например, для орошения, животноводства, в бытовых целях. Общая щелочность воды Малого моря – 183,0 мг/л, это на 73,2 мг/л меньше, чем в Большом море. Концентрация ионов хлора в Большом море на 6 522,8 г/л больше, чем в Малом море. Растворимость в воде и продвижение этого элемента не зависит от температуры среды, т. е. ионы хлора в водах Большого моря накопились с давних времен. Концентрации ионов сульфата (SO4) в водах двух морей близки: в Малом море – 3 789,1 мг/л сульфата, а в водах Большого моря – 3 622,8 г/л ионов сульфата, концентрация этих ионов в воде зависит от температурного режима. Ионов натрия и калия в воде Малого моря – 4 797,5 мг/л, а Большого моря – 9 267,5 г/л, ионы натрия в основном соединяются с ионами сульфата и оседают в виде мирабилитных солей (Na2SO4 × 10Н2О). Эти соли и собираются на обезвоженном грунте морского дна. В летние месяцы температура воздуха поднимается, и мирабилит на поверхности грунта испаряет воду, появляется тенардит – сухой сульфат натрия (Na2SO4), порошок белого цвета, который сдувается с поверхности грунта, попадает на поля, пашни и нарушает плодородие почвы. Восточная маловодная часть Аральского моря в 70-х годах прошлого века быстро осушилась, оставшуюся на поверхности грунта солевую массу уносили пылевые бури. Этот процесс в первые 10–15 лет шел очень динамично. Потом восточный берег моря отодвинулся далеко на запад, уровень соленой воды в пластах понизился, вследствие чего остановился процесс капиллярного выхода воды по профилю грунта. По данным на сентябрь 2011 года, в высохшие озера Карашалан, Баян, Жыланды, Домалак, Картма, Кызылжарма поступило 82,0 млн. м3 воды. Это в свою очередь способствовало появлению растительности в приморских и прибрежных зонах. В регионе сформировался мягкий микроклимат, пригодный для домашнего скота, хищных зверей и птиц. Местные жители восстанавливают рыболовное хозяйство и используют влажные земли в качестве пашни. Л и т е р а т ур а Нургизаринов А. Н. Арал өңірінің экологиялық күйіне байланысты биологиялық қорлардың қайтадан қалпына келуі / А. Н. Нургизаринов, Р. Х. Курманбаев // Қазіргі заманғы Арал өңірінің экологиялық жағдайы, проблемаларды шешу перспективалары: Халықаралық ғылыми-практикалық конференция. – Қызылорда, 2011. – 59–61 б.б. Нургизаринов А. Аралдың экологиялық тынысы / А. Нургизаринов. – Алматы: Ғылым, 2006. – 225 б. Саданов А. К. Арал өңірінде орнықты дамудың ғылыми негізгі / А. К. Саданов, А. Н. Нургизаринов. – Астана: Ақарман, 2008. – 170 с. 216 РАЗНООБРАЗИЕ И ОЦЕНКА СОСТОЯНИЯ ЗЕЛЕНЫХ НАСАЖДЕНИЙ Г. КАЗАНИ ПО ДРЕВЕСНОЙ РАСТИТЕЛЬНОСТИ Юпина Г. А., Никитин А. В., Мингазова Н. М. Казанский (Приволжский) федеральный университет, г. Казань, lucky033@yandex.ru. Озеленение городов требует постоянного мониторинга состояния зеленых насаждений. В г. Казани по результатам инвентаризации зеленых насаждений, проведенной Казанским университетом по муниципальному контракту, на 1.01.2008 г. к насаждениям общего пользования было отнесено 190 объектов, из них 16 парков, 5 садов, 109 скверов, 58 зеленых зон и городской лес «Лебяжье», расположенный в черте города в Кировском, Московском, Приволжском и Советском районах (табл. 1). Таблица 1 – Количество объектов зеленых насаждений общего пользования Кол-во парков Кол-во садов Авиастроительный 2 – 4 4 Вахитовский 5 3 37 9 Кировский 2 1 14 6 Московский 2 – 9 6 Ново-Савиновский 2 – 8 7 Приволжский 2 1 22 13 Советский 1 – 15 13 Итого по Казани: 17 5 109 58 Название района Кол-во скверов Кол-во зеленых зон В зеленых насаждениях общего пользования произрастает 274 895 деревьев, принадлежащих к 71 виду. Из 71 видов деревьев 22 вида принадлежат к природной флоре средней полосы России. Наряду с растениями природной флоры республики Татарстан в г. Казани произрастают деревья экзоты (33 вида). Ведущими древесными породами в городе являются клен ясенелистный, липа сердцевидная, береза повислая, тополь секции бальзамической и сосна обыкновенная. Анализ распространения видов деревьев по районам города показал, что в Авиастроительном и Советском районах доминируют клен ясенелистный и липа сердцевидная; в Вахитовском районе – липа сердцевидная и клен ясенелистный; в Кировском районе – клен ясенелистный и тополь секции бальзамической; в Московском и Ново-Савиновском районах – липа сердцевидная и береза повислая; в Приволжском – береза повислая и клен ясенелистный. 217 Клен ясенелистный является наиболее распространенной древесной породой городских зеленых насаждений. В озеленении Казани с середины ХХ в. этот вид занимал ведущее место из древесных пород. В настоящее время прослеживается тенденция на снижение в посадках количества клена ясенелистного, как и тополя секции бальзамической. В условиях загрязнения воздушной среды, нарушения и загрязнения почвенной структуры, заражения деревьев и кустарников паразитическими организмами происходит большое разнообразие как повреждений, так и поражений, что приводит к появлению различных заболеваний. У древесно-кустарниковой растительности происходит нарушение роста, усыхание ветвей, дифолиация, изменения в облиствлении, появление некрозов и хлорозов, суховершинность, фитопатологические болезни, различные механические повреждения. При анализе данных по инвентаризации зеленых насаждений г. Казани были установлены количественные и процентные показатели жизненного состояния деревьев. Анализ проведенных исследований показал, что наибольшее количество деревьев – 160 251 экз. – относится к III классу жизнеустойчивости. Это ослабленные посадки, со средним облиствлением и усыханием кроны, различными механическими повреждениями, грибными болезнями. К данному классу относятся 58 % всех древесных насаждений. Наибольшее число таких деревьев было выявлено в Советском районе г. Казани – 39 127 экз., в Вахитовском районе – 35 688 экз. и в Приволжском районе – 33 558 экз. Таблица 2 – Оценка жизненного состояния деревьев в насаждениях общего пользования Район г. Казани Кол-во живых деревьев I II III IV V Кол-во погибших деревьев Авиастроительный 22724 629 8990 9295 2849 961 457 Вахитовский 50767 775 8381 35688 5187 736 1072 Кировский 23647 1100 4449 9779 5222 3097 320 Московский 22803 181 3339 14196 3805 1282 599 Ново-Савиновский 31658 206 5657 18608 5803 1384 564 Приволжский 57144 637 14698 33558 7076 1175 1097 Советский 66152 472 19163 39127 6570 820 1112 Итого по Казани 274895 4000 64677 160251 36512 9455 5221 100 1,5 23,5 58,3 13,3 3,4 % Класс жизнеустойчивости деревьев 218 Незначительно ослабленные деревья II класса жизнеустойчивости отмечены в количестве 64 677 экз. (23,5 %). Наибольшее их число отмечалось в Советском районе (19 163 экз.), а наименьшее в Московском (3 339 экз.) и Кировском (4 449 экз.) районах. Такие декоративные деревья произрастают на улицах города, в парках и скверах – это и молодые, и средневозрастные посадки. Высокодекоративные, здоровые деревья I класса жизнеустойчивости отмечены в количестве 4 000 экз., что составляло 1,5 % от всех посадок. Наименьшее их число произрастает в Московском районе (181 экз.), а наибольшее – в Кировском районе города (1 100 экз.). Учитывая долю сильно ослабленных и усыхающих деревьев (IV–V классов жизнеустойчивости) от общего числа живых деревьев, можно констатировать, что наиболее ослабленные древесные насаждения произрастают в Кировском районе г. Казани (35 % деревьев в районе сильно ослабленные и усыхающие). Высокий процент ослабления отмечается в Московском и Ново-Савиновском районах г. Казани, сильно ослабленных и усыхающих деревьев в них составляет 22 %. Таким образом, основу зеленых насаждений города Казани составили ослабленные (58,3 %) деревья. Это указывает на недостаточный уход, многие насаждения заброшены и нуждаются в благоустройстве. Наряду с жизненным состоянием деревьев был проведен анализ возрастного состава зеленых насаждений. Основное количество деревьев, произрастающих на территории города, – это молодые деревья I класса возраста в количестве 106 585 экз., что составляет 38,8 % от всех древесных посадок. Молодняки преобладают в Советском, НовоСавиновском, Авиастроительном районах города. В основном это насаждения реконструированных или вновь сформированных парков, скверов и молодая поросль зеленых зон. Несколько в меньшем числе произрастают средневозрастные деревья III класса возраста – 90 504 экз. (32,9 %). Наибольшее количество средневозрастных деревьев в Вахитовском, Кировском, Московском, Приволжском районах. На долю жердянок (II класс возраста) приходится 48 495 деревьев, что составляет 17,6 % всех древесных насаждений города. Количество таких деревьев варьирует от 3 489–4 372 до 6 788–9 978 экз. и насчитывает 12 774 экз. на территории Советского района. На долю приспевающих деревьев (IV класс возраста) приходится 17 366 экз., что составляет 6,3 % от общего количества деревьев в городе. Численность приспевающих деревьев изменяется от 1 192–1 518 экз. (Ново-Савиновский, Московский районы) до 2 354 экз. (Авиастроительный район) и достигает 4 013–5 203 экз. в Вахитовском и Приволжском районах. В результате проведенных исследований следует заключить, что наибольшее количество скверов, садов, парков сосредоточено в Вахи219 товском районе (при небольшой площади), а наибольшее количество зеленых зон располагается в Приволжском, Советском районах г. Казани. Низкое количество зеленых насаждений в Авиастроительном районе объясняется плотной застройкой, большим количеством производственных и промышленных зон. В озеленении использовалось около 275 тысяч деревьев 71 вида. Основу зеленых насаждений города на 2007–2008 гг. составили ослабленные деревья, что указывает на недостаточный уход за древесной растительностью. Данные исследований можно использовать для целей мониторинга состояния зеленых насаждений, т. к. за 2009–2013 гг. в связи со строительством объектов Универсиады и осуществлением муниципальных программ по озеленению строительных площадок ситуация с зелеными насаждениями города изменяется. 220 Секция 4 ПОПУЛЯЦИОННОЕ БИОРАЗНООБРАЗИЕ И УСТОЙЧИВОСТЬ ПОПУЛЯЦИЙ ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ ПОСКОННИКА КОНОПЛЯНОГО (EUPATORIA CANNABINUM L.) Агаева П. Н. 1, Османова Г. О. 2 1 Институт ботаники Национальной академии наук Азербайджана, г. Баку 2 Марийский государственный университет, г. Йошкар-Ола, gyosmanova@yandex.ru Посконник конопляный (Eupatoria cannabinum L.) – многолетнее, короткокорневищное травянистое растение из семейства астровые (Asteraceae). Растение с высоким, до 2,0 м, ветвистым стеблем. Листья супротивные, глубокорассеченные на 3–5 ланцетных крупнопильчатых заостренных долей, снизу железистых. Цветки грязно-розовые, обоеполые, трубчатые, с двумя длинными нитевидными долями столбика пестика, собраны по 4–6 в корзинки. Обертки корзинок продолговатые. Корзинки собраны в густую щитовидную метелку. Время цветения – июнь–август. Плоды – цилиндрические ребристые семянки с хохолком из одного ряда волосков (Скворцов, 2000). Ареал распространения охватывает Скандинавию, Среднюю, Атлантическую и Южную Европу, Балканский полуостров, Малую Азию, Турцию, Армению и Иран; встречается в Австралии и Северной Африке. Произрастает в европейской части России (кроме Севера), на Украине (кроме Крыма), в Беларуси, на Кавказе, на юге Западной Сибири, в юго-западных районах Средней Азии. E. cannabinum предпочитает берега рек и озер, встречается по обочинам канав, на болотистых лугах и в сырых зарослях кустарников (Определитель сосудистых…, 1995). Посконник конопляный – лекарственное растение. Надземная часть растения содержит сесквитерпеновый лактон, зупаторнопикрин, зупирин, альфа-лакгуцерол, 1-инозит, эфирное масло (до 0,3 %), рутин, гиперозид, дубильные вещества, сапонины, аскорбиновую кислоту, смолу, холин, инулин, астрагалин, изокверцитрин, кумаровую и феруловую кислоты, ароматические оксикислоты (кофейная, хлорогеновая, изохлорогеновая). В корнях обнаружен инулин (Биохимия…, 1968). В народной медицине настой травы используют для регулирования деятельности желудочно-кишечного тракта, при заболеваниях печени 221 и селезенки, водянке и отеках, т. к. он обладает слабительным, мочегонным, потогонным и желчегонным действием. Настойку из E. cannabinum применяют при гриппе и простудных заболеваниях, а наружно – при ушибах, для заживления язв и ран. Растение ядовито, поэтому его внутренне употребление допустимо только по назначению и под наблюдением врача. Посконник конопляный нуждается в охране и включен в Красную книгу Карелии (1995) со статусом 3 (LC) как редкое (вызывающее наименьшие опасения) растение, поэтому изучение структуры его ценопопуляций (ЦП) актуально. Исследования ЦП E. cannabinum проводили в августе 2013 г. на территории Губинского района Республики Азербайджан в окрестностях села Минара по дороге в село Хыналыг. Губинский район относится к области Большого Кавказа (в пределах Азербайджана) и обозначается в литературе как БК (кубинск.) и БК (вост.) ботанико-географический район. Данные GPS: N 41º 14.396'; E 048º 18.556', высота над уровнем моря составляет 1098 м. E. cannabinum встречается в Губинском районе на сухих каменистых, щебнистых и глинистых склонах, а также на равнинных территориях в зарослях кустарников. Размещение растений в исследуемых местообитаниях носило диффузный характер. Для характеристики растений любого вида важно знать его устойчивость в ценозе, способность возобновляться и поддерживать определенную численность. Для характеристики процессов самоподдержания E. cannabinum были рассчитаны индексы восстановления и замещения (Жукова, 1995). Тип ценопопуляций определяли по классификации «дельта-омега» Л. А. Животовского (2001), основанной на совместном использовании индексов возрастности (∆) А. А. Уранова (1975) и эффективности (ω) (Животовский, 2001). Результаты изучения онтогенетической структуры ЦП E. cannabinum в исследованных местообитаниях нормальные неполночленные и представлены особями прегенеративного, генеративного и постгенеративного периодов. Спектр онтогенетических состояний в ЦП 1 одновершинный центрированный (рис. А), с макимумом на группе особей средневозрастного генеративного состояния (24,4 %). В ЦП 1 E. cannabinum, произрастающей близ родника Назлы-булаг, преобладали особи генеративного периода (51,3 %). Доля особей прегенеративного периода составила 39,0 %. Растения в этой ЦП составили всего 4,9 %. Немногочисленной была группа особей постгенеративного периода (9,7 %). В ЦП 2 (каменистые склоны оврага в окрестностях села Хыналыг) онтогенетический спектр E. cannabinum двухвершинный (рис. Б) с максимумом на группе особей виргинильного и средневозрастного генера222 тивного онтогенетических состояний (по 24,4 %). Минимум в спектре (1,4 %) составили растения ювенильной группы E. cannabinum. Сенильные особи E. cannabinum в ЦП 2 отсутствовали. Особи E. cannabinum произрастали в условиях высоких горных склонов и не испытывали влияния антропогенного фактора. Невысокая доля особей ювенильного онтогенетического состояния, по-видимому, связана с поздними сроками сбора материала. Поскольку в обеих ЦП высока доля особей генеративного периода, особенно фракции средневозрастных генеративных растений, то и нагрузка на среду будет выше, поэтому индекс эффективности (Животовский, 2001) имеет высокие значения: в ЦП 1 – ω = 0,602, а в ЦП 2 – ω = 0,653. А относительно низкие (Δ = 0,335 и Δ = 0,319 соответственно) значения индекса возрастности (Уранов, 1975) можно объяснить наличием в обеих ЦП достаточно высокой доли особей прегенеративного периода (39,0 и 38,9 % соответственно). А Б Спектр онтогенетических состояний ценопопуляций E. cannabinum: А – ЦП 1, Б – ЦП 2 Процессы самоподдержания в исследуемых ЦП E. cannabinum идут одинаковыми темпами. Значения индексов восстановления и замещения для обеих ЦП E. cannabinum практически одинаковы и составляют: в ЦП 1 (Iв = 0,41; Iз = 0,37), а в ЦП 2 (Iв = 0,41; Iз = 0,39). В заключение можно отметить, для E. cannabinum основным способом самоподдержания является семенное. Засчет формирующихся на корневище многочисленных почек возобновления происходит образование системы надземных побегов и разрастание особей E. cannabinum, поэтому возможно и вегетативное размножение (черенкованием и делением куста). 223 Л и т е р а т ур а Биохимия растений. – М.: Мир, 1968. – 624 с. Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений / Л. А. Животовский // Экология. – 2001. – № 1. – С. 3–7. Жукова Л. А. Популяционная жизнь луговых растений / Л. А. Жукова. – Йошкар-Ола: РИИК «Ланар», 1995. – 223 с. Красная книга Карелии / науч. ред. Э. В. Ивантер, О. Л. Кузнецов. – Петрозаводск: Карелия, 1995. – 286 с. Определитель сосудистых растений Центра европейской части России / И. А. Губанов, К. Н. Киселева, В. С. Новиков, В. Н. Тихомиров. – М.: Аргус, 1995. – 560 с. Скворцов В. Э. Атлас – определитель сосудистых растений таежной зоны Европейской России: определитель по генеративным и вегетативным признакам, региональные списки редких и охраняемых видов / В. Э. Скворцов. – М.: Гринпис России, 2000. – 587 с. Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов / А. А. Уранов // Биол. науки. – 1975. – № 2. – С. 7–34. ФАУНИСТИЧЕСКАЯ СТРУКТУРА ПОЧВООБИТАЮЩИХ ЖЕСТКОКРЫЛЫХ (COLEOPTERA) ПОЙМЫ МАЛЫХ РЕК КОСТРОМСКОЙ ОБЛАСТИ Анциферов А. Л. ¹, Полежаева А. Ю. ² ¹ ОГБУК «Музей природы Костромской области», г. Кострома, ancifer.ost@yandex.ru ² ФГБОУ ВПО «Костромской государственный университет им. Н. А. Некрасова», г. Кострома, zhuzhefilka@mail.ru Почвообитающие жесткокрылые, как наиболее характерные представители группы герпетобионтов, в нашей работе представлены преимущественно видами семейств жужелиц (Carabidae), стафилинов (Staphylinidae), мертвоедов (Silphidae), а также, в незначительной степени, карапузиков (Histeridae) и нарывников (Meloidae). Благодаря большому видовому разнообразию, численности и широкому распространению они характеризуются как чувствительные биоиндикаторы, энтомофаги и сапрофаги и являются важным звеном в биоценозах. Поэтому неслучайно данная группа насекомых в целом либо ее отдельные представители широко используются в наше время как модельные объекты для изучения экологических закономерностей. Изучение фаунистического состава жуков-герпетобионтов проводили в летние периоды с 2007 по 2012 г. в пойменных зонах малых рек волжского бассейна, притоков I, II и III порядка: Ветлуга, Кострома, Покша, Сендега, Корба на разных участках русла. Территориально исследованиями были охвачены Костромской, Судиславский, Солигаличский, Шарьинский и Пыщугский административные районы Костромской области. Для отлова и учета жесткокрылых применяли стандартный метод почвенных ловушек Барбера (Тихомирова, 1975) в сочетании с фикси224 рующей жидкостью (крепкий раствор поваренной соли либо уксусной кислоты). Ловушки располагали стационарно на участках каждого отдельного элемента рельефа поймы порциями по 5–10 шт. в направлении от уреза воды к коренному берегу. При выборе контрольных площадок ориентировались на характерные различия в структуре рельефа поймы, выделяя такие элементы, как бечевник, береговой (прирусловый) вал, тыловой шов и надпойменная терраса (Пармузин, Карпов, 1994). В работе приведены данные по всем указанным территориальным объектам в обобщенном виде. Всего за период исследований было учтено 4854 особи жуковгерпетобионтов, принадлежащих к 105 видам из 5 семейств, представленных в следующем соотношении (по суммарному обилию от общей совокупности): жужелицы – 65,7 %, стафилины – 21 %, мертвоеды – 9,5 %, карапузики – 2,8 % , нарывники – 1 % (рис.). В пределах каждого отдельного элемента рельефа поймы характер данного соотношения (по сравнению с общим) практически не изменяется. Исключением является надпойменная терраса, где процентная доля жужелиц по-прежнему высока, но все же снижается до 56,7 % при возросшем обилии стафилинов до 28,3 % (рис.). Предположительно это вызвано резко обособленной формой и характером фитоценоза надпойменной террасы. Как правило, это возвышенные части речной долины, естественные горизонтальные или слабо наклонные площадки на склонах речных долин, ограниченные уступами (Важнов, 1976). По типу растительности исследуемые участки надпойменных террас отличаются лесным характером видового состава с наличием древесного яруса, подлеска, подстилки и соответствующего типа почвы. Соотношение семейств почвообитающих жесткокрылых (общее и в отдельных структурных элементах) речных пойм 225 В данном случае можно судить о большей преферентности представителей семейства стафилинов в отношении участка надпойменной террасы, нежели остальных элементов поймы. К примеру, такие виды, как Staphylinus caesareus Cederh. и Quedius nitidipennis Steph. обнаруживаются только в пределах этой структуры рельефа. А Philontus splendens Fabricius, Domene stilicina Erichson, Xantolinus tricolor Fabricius и Medon melanocephalus Fabricius резко прибавляют в численности по сравнению с другими участками и заметно преобладают в зоне надпойменной террасы. Это так называемые эуценные (характерные) виды – свойственные исключительно одному биоценозу или, что случается чаще, гораздо обильнее в них встречающиеся, чем в других биоценозах (Дажо, 1975). В целом по доминантному составу наиболее массовыми видами пойменных жуков-герпетобионтов отмечены Trechus secalis Paykull (11,5 %), Pterostichus oblongopunctatus Fabricius (9,8 %), Domene stilicina (7,8 %), Carabus nemoralis O. Müller (7,7 %), Philontus splendens (7,1 %), Pterostichus melanarius Illiger (6,1 %). По системе О. Ренконена (1938) данная группа видов (с долей 5 % и более от общей массы) отнесена к доминантам. Однако для каждого отдельного элемента поймы состав и соотношение доминантов индивидуальны. Эврибионтный вид, убиквист (по Дажо) Trechus secalis (Carabidae) с тем или иным изменением обилия доминирует во всех выделенных участках поймы, в то время как Astenus uniformis Duv. (Staphylinidae), Platynus assimilis Paykull и Carabus granulatus Paykull (Carabidae) в массе встречаются только в пределах зоны бечевника; Poecilus versicolor Sturm. (Carabidae) – в зоне берегового вала; Oiceoptoma thoracica Herbst. (Silphidae), Patrobus assimilis Chaudoir и Pterostichus strenuus Panzer (Carabidae) доминируют только на участке тылового шва; Carabus hortensis L. (Carabidae) – типичный доминирующий вид надпойменной террасы с лесным фитоценозом. Для данных биотопов – это характерные эуценные виды (табл.). Доминантный состав и биотопическое распределение жуков-герпетобионтов речной поймы* Доминантный состав Бечевник Patrobus assimilis 12,3 Carabus granulatus 6,2 Astenus uniformis 8,9 Trechus secalis 7,3 Береговой вал Тыловой шов Надпойм. терраса 20,1 5,3 11,4 Domene stilicina 12,4 8,9 8 Carabus nemoralis 6,8 7,9 10,8 226 Доминантный состав Бечевник Philontus splendens Pterostichus melanarius 7 Береговой вал Тыловой шов Надпойм. терраса 5,3 6,2 10,5 6,6 Poecilus versicolor 6,1 6 Pterostichus oblongopunctatus 8,9 Oiceoptoma thoracica 9,9 Patrobus assimilis 19,1 6 Pterostichus strenuus 5,1 Carabus hortensis 5,7 * Числовые значения в ячейках таблицы – процентная доля от общей массы жуков в данном элементе поймы. В итоге проведенного анализа можно сделать вывод о наличии характерного видового состава фауны жуков-герпетобионтов в экосистеме речной поймы, формирующегося в определенной зависимости от ее рельефной структуры и степени привязанности того или иного вида к конкретному биоценозу. Л и т е р а т ур а Важнов А. Н. Гидрология рек / А. Н. Важнов. – М.: МГУ, 1976. – 239 с. Дажо Р. Основы экологии / Р. Дажо. – М.: Прогресс, 1975. – 415 с. Пармузин Ю. П. Словарь по физической географии / Ю. П. Пармузин, Г. В. Карпов. – М.: Просвещение, 1994. – 367 с. Тихомирова А. Л. Учет напочвенных беспозвоночных / А. Л. Тихомирова // Методы почвенно-зоологических исследований. – М.: Наука, 1975. – С. 73–85. Renkonen O. Statisch-Okologische Untersuchungen Über die ferrestrische Käferwelt der finnischen Bruchmoore / O. Renkonen // Ann. Zool. Soc. Zool. – Bot. Fenn. Vanamo, 1938. – Bd. 6. – 231 ss. РАЗМНОЖЕНИЕ ВИДОВ РОДА HEDYSARUM L. IN VITRO Ахметова А. Ш., Зарипова А. А. Учреждение Российской академии наук Ботанический сад-институт Уфимского научного центра РАН, г. Уфа, al_sham@mail.ru В данной работе объектом изучения являлись представители рода Hedysarum: копеечник серебристолистный (Hedysarum argyrophyllum Ledeb.) и копеечник крупноцветковый (Hedysarum grandiflorum Pall.). Это редкие, находящиеся под угрозой исчезновения декоративные виды, занесенные в Красную книгу РФ (2008) и в ряд региональных Красных книг Российской Федерации (Красная книга..., 1998; 2001), эндемичны для республики Башкортостан, известны для 23 изолированных, по преимуществу скальных, местонахождений. Разработка альтернатив227 ных способов расширения сырьевой базы исследуемых видов дает уникальную возможность сохранить их природные популяции, что способствует сохранению многообразия растительного мира. Проблема сохранения H. argyrophyllum и H. grandiflorum in situ может быть решена путем введения этого растения в культуру тканей, создания генных банков и репатриацией, полученного в ходе микроклонального размножения материала, в естественные сообщества. Целью настоящего исследования было изучение особенностей культивирования на каждом этапе размножения in vitro редких и исчезающих видов H. argyrophyllum и H. grandiflorum. Работу в асептических условиях, стерилизацию питательных сред и посадочного материала проводили согласно имеющимся рекомендациям (Бутенко, 1964; Калинин и др., 1980). Поверхностную стерилизацию проводили с использованием в качестве стерилизующих агентов ртутьсодержащего соединения диацида в концентрации 0,1 % и лизоформина в концентрации 1,0 %. Семена предварительно обрабатывали 70 %-м этанолом в течение 0,5 мин, а затем одним из вышеперечисленных стерилизующих растворов. Экспозиция воздействия стерилизующих растворов составляла от 4 до 12 мин (табл.). Использованные нами стерилизующие растворы по-разному влияли на жизнеспособность семян и последующее развитие проростков. Применение лизоформина для дезинфекции семян не являлось эффективным, так как в этом случае получение стерильной культуры составляло 12,2–30,6 %. Причем при 12-минутном выдерживании семян в лизоформине число инфицированных и нежизнеспособных было максимально и составляло 69,4–75 %. При стерилизации семян H. аrgyrophyllum и H. grandiflorum в 0,1 % растворе диацида в течение 8 мин было получено максимальное количество стерильной культуры (83,3–87,4 %), которая характеризовалась интенсивным ростом. Влияние стерилизующего раствора и его экспозиции на получение стерильных культур Hedysarum argyrophyllum и Hedysarum grandiflorum (%) Стерилизующий раствор Генотип диацид 0,1 % лизоформин 1,0 % экспозиция стерилизации, мин. 4 6 8 экспозиция стерилизации, мин. 6 10 12 H. аrgyrophyllum 12,5 ± 4,77 42,6 ± 6,73 83,3 ± 5,38 12,2 ± 4,67 20,4 ± 5,76 30,6 ± 6,58 H. grandiflorum 16,7 ± 5,07 41,7 ± 7,12 87,4 ± 4,58 13,7 ± 4,73 17,3 ± 5,25 25,0 ± 6,00 228 Интенсивность морфогенеза зависит от наличия регуляторов роста, использованных в питательной среде. Одним из наиболее мощных индукторов морфогенеза, который принято называть стимулирующим фактором или сигналом морфогенеза, является изменение соотношения между цитокининами и ауксинами, входящими в состав питательных сред. Для увеличения коэффициента размножения и получения микропобегов, способных к ризогенезу, было проведено исследование по подбору оптимальных концентраций регуляторов роста. Для H. grandiflorum оптимальной питательной средой для размножения является среда с добавлением 1,0 мг/л БАП, коэффициент размножения составил 4,9, в то время как у H. аrgyrophyllum на данной среде коэффициент размножения составил 2,0, и наблюдалась значительная витрификация побегов – 75 %. Сравнительный анализ изучения влияния регуляторов роста показал, что под воздействием 0,5 мг/л БАП и 0,2 мг/л ИУК коэффициент размножения H. аrgyrophyllum, а также средняя длина побегов существенно превышали значение этих показателей на среде с 1,0 мг/л БАП. Коэффициент размножения составил 3,8. Содержание в среде 0,5 мг/л кинетина и 0,2 мг/л ИУК уменьшало коэффициент размножения у исследуемых видов, одновременно стимулируя образование каллуса. Коэффициент размножения на среде с 2,0 мг/л БАП и 0,1 мг/л ИУК различался в 2 раза (от 4,2 до 8,0). Если у H. аrgyrophyllum наблюдалось разрастание тканей побега и образование большого числа адвентивных почек (коэффициент размножения 8,0), то почти все побеги H. grandiflorum были витрифицированные. Одновременно на данной среде индуцировался и рост побегов. У H. grandiflorum она составила14,8 мм, у H. Аrgyrophyllum – 12,3. Установлено также, что коэффициент размножения представителей рода Hedysarum в процессе культивирования существенно изменяется. Отмечена общая для всех изучаемых видов тенденция: на первых пассажах коэффициент размножения постепенно возрастал, достигая наибольших значений на 3–5 субкультивированиях, снова снижаясь на 6–7 пассажах. Максимальный коэффициент размножения H. аrgyrophyllum на 4 субкультивировании, H. Grandiflorum – на 5. Для стимуляции процесса ризогенеза на полученных микропобегах из питательной среды исключили цитокинины. Результаты экспериментов по подбору питательных сред на этапе укоренения показывают значительные отличия скорости укоренения в зависимости от концентрации ауксинов, применяемых для индукции ризогенеза. Микропобеги укореняли на среде 1/2 MS, дополненной ауксинами: 1) ИМК 0,5 мг/л; 2) ИМК 0,2 мг/л + НУК 0,2 мг/л. Результаты эксперимента указывают 229 на целесообразность использования в качестве индуктора ризогенеза для микропобегов H. аrgyrophyllum и H. grandiflorum ИМК. Оптимальной оказалась концентрация 0,5 мг/л, обеспечивающая высокий процент укорененных побегов (73,8 и 88,6 %). Интенсивность ризогенеза составила 4,7 и 6,4 шт. на эксплант, а длина корней достигала 58,7 и 71,4 мм. Следовательно, питательную среду ½ MS, дополненную ИМК 0,5 мг/л, можно рекомендовать как относительно универсальную для укоренения растений-регенерантов H. argyrophyllum и H. grandiflorum. Таким образом, проведена оценка эффективности различных подходов стерилизации семян Hedysarum argyrophyllum Ledeb и Hedysarum grandiflorum L. при введении в культуру in vitro, разработаны условия обеспечения оптимального роста и развития растений за счет подбора питательной среды и физических условий культивирования. В результате исследований было установлено, что реализация морфогенетического потенциала определяется видовыми особенностями. Л и т е р а т ур а Бутенко Р. Г. Культура изолированных тканей и физиология морфогенеза растений / Р. Г. Бутенко. – М., 1964. – 272 с. Калинин В. Ф. Методы культуры тканей в физиологии и биохимии растений / В. Ф. Калинин, В. В. Сарнацкая, В. Е. Полищук. – Киев: Наукова думка, 1980. – 488 с. Красная книга Республики Башкортостан. Редкие исчезающие виды высших сосудистых растений. – Уфа: Китап, 2001. – Т. 1. – С. 75–76. Красная книга Российской Федерации. Растения и грибы. – М.: Товарищество научных изданий, 2008. – 885 с. Красная книга Оренбургской области. – Оренбург: Оренбургское кн. изд-во, 1998. – 176 с. ОНТОГЕНЕТИЧЕСКАЯ И ВИТАЛИТЕТНАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ ЯСЕНЯ ОБЫКНОВЕННОГО НА ГРАНИЦЕ АРЕАЛА Бекмансуров М. В., Соловьева М. Н. Марийский государственный университет, г. Йошкар-Ола, ecology@marsu.ru В Республике Марий Эл природные популяции ясеня (Fraxinus excelsior L.) встречаются только в Горномарийском районе, где располагается северо-восточная граница ареала его сплошного распространения (Алимбек, 1967; Заугольнова, 1974). Ясень здесь растет в широколиственных лесах с участием дуба, липы, клена. Чистых насаждений практически не образует. Очень редко встречаются ясеневники площадью менее гектара с незначительным участием других широколиственных видов в древостое (Красная книга..., 2013). Сбор материала проводили в широколиственных лесах и их производных на водораздельных территориях, а также в долине р. Суры. 230 В геоморфлогическом отношении район исследования существенно отличается от низменного левобережья. Изученная территория расположена на Приволжской возвышенности и сложена в основном породами татарского яруса, имеет наибольшее вертикальное расчленение в пределах республики – 118 м, большой коэффициент овражной (0,27 км/км2) и балочной эрозии (1,31 км/км2) (Энциклопедия…, 2009). Поверхностные отложения представлены в основном покровными лессовидными суглинками. Цель работы: охарактеризовать онтогенетическую структуру ценопопуляций (ЦП) ясеня обыкновенного. На исследованной территории ясень встречается в древостое и подросте липняков, березняков, осинниках, а также в серо- и черноольшаниках и в искусственных насаждениях сосны сибирской и сосны обыкновенной. В сообществах с участием и доминированием ясеня заложена 21 пробная площадь размером 500 м², проведено их геоботаническое описание. На каждой пробной площади подсчитывали численность особей каждого онтогенетического состояния с учетом уровня жизненности и в дальнейшем строили виталитетно-онтогенетические спектры. Выделение онтогенетических состояний осуществляли по диагнозам, предложенным Л. Б. Заугольновой (1974). Жизненность особей оценивали по 3-бальной шкале: 1 – нормальная, 2 – пониженная, 3 – низкая. В изученных ценопопуляциях ясеня онтогенетический спектр левосторонний, преобладают проростки и особи j-состояния, что свидетельствует о благоприятных условиях для семенного размножения. Нормальные полночленные ценопопуляции ясеня отмечены в липняках: лунниково-снытевом, крапивном и крапивно-пролесниковом. Однако онтогенетический спектр здесь резко левосторонний за счет большого числа ювенильных особей (несколько сотен на пробной площади) (рис., А). В то же время доля имматурных, виргинильных и генеративных растений невелика – не более десятка. Известно, что всходы ясеня устойчивы к затенению, но не переносят конкуренции с видами травянистого яруса. Сходный тип спектра отмечен в зрелых древостоях сосны обыкновенной и сибирской, имеющих искусственное происхождение, однако генеративные растения здесь практически не встречаются. Лишь за пределами пробной площади обнаружено несколько растений g1 и g2-состояния. Возможно, их отсутствие объясняется рубками ухода, которые проводятся в посадках. В березняках и осинниках снытевых ценопопуляции нормальные неполночленные, отсутствуют ювенильные особи (рис., Б). Спектр левосторонний с максимумом на виргинильных растениях первой подгруппы. При этом и среди генеративных растений и в подросте преоб231 ладают особи нормального уровня жизненности. Ясень, являясь светолюбивым видом, успешно растет и в березняках, и в осинниках, где сомкнутость крон древостоя и развитие подлеска существенно ниже, чем в липняках. Виталитетно-онтогенетический спектр ценопопуляций ясеня обыкновенного в различных лесных сообществах: А – липняки, Б – березняки и осинники; В – ясеневники; Г – серо- и черноольшаники. Жизненность: 1 – нормальная, 2 – пониженная, 3 – низкая В ясеневниках спектр правосторонний и фрагментарный. Ценопопуляции нормальные неполночленные. Преобладают деревья g1 состояния (рис., В). Плотность составляет 12,4 экз./100 м2. Такой спектр, в котором отсутствуют ювенильные особи и малочисленны иматурные, говорит о том, что возобновление ясеня под своим пологом затруднено. В серо- и черноольшаниках спектр левосторонний (рис., Г). Ценопопуляции нормальные, неполночленные. Немногочисленные генератив232 ные особи характеризуются пониженной жизненностью и не обеспечивают восстановления ценопопуляции, плотность которой очень низкая (2 экз./100 м2). Пониженная жизненность связана с тем, что в условиях близкого залегания грунтовых вод в ольшаниках взрослые особи медленно растут и погибают из-за гибели молодых корней (Заугольнова, 1974). Таким образом, несмотря на то, что изученная популяция ясеня обыкновенного располагается на северо-восточном пределе ареала, ее состояние можно оценить как удовлетворительное. По-видимому, распространение F. excelsior на восток ограничено антропогенной деятельностью – здесь располагаются аграрные районы республик Марий Эл, Чувашии и Татарстана, поэтому уровень лесистости очень низкий. К северу от исследованной территории располагается Чебоксарское водохранилище и низменное левобережье Волги с условиями, не подходящими для ясеня: зандровым ландшафтом, мощной толщей песчаных древнеаллювиальных отложений, бедными оподзоленными почвами. Работа выполнена при финансовой поддержке Департамента экологической безопасности, природопользования и защиты населения Республики Марий Эл Л и т е р а т ур а Алимбек Б. М. Васильсурские дубравы / Б. М. Алимбек // Охрана и обогащение природы Марийской АССР. – Йошкар-Ола: Марийское кн. изд-во, 1967. – С. 80–89. Заугольнова Л. Б. Ясень обыкновенный / Л. Б. Заугольнова // Флора московской области. – М.: МГУ, 1974. – Вып. 1. – С. 142–159. Красная книга Республики Марий Эл. Растения. Грибы / Мар. гос. ун-т. – Йошкар-Ола, 2013. – 324 с. Энциклопедия Республики Марий Эл. – Йошкар-Ола, 2009. – 872 с. ЭКОЛОГО-МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ОСОБЕЙ И ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ КРОВОХЛЕБКИ ЛЕКАРСТВЕННОЙ (SANGUISORBA OFFICINALIS L.) Ведерникова О. П., Халиуллина Л. Р. Марийский государственный университет, г. Йошкар-Ола, ecology@marsu.ru Детальное изучение морфологических особенностей особей и онтогенетической структуры ценопопуляций (ЦП) позволяет оценить состояние и прогнозировать все изменения их развития, что имеет большое значение для рационального использования и продуктивного долголетия фитоценозов. Цель работы – охарактеризовать эколого-биологические особенности особей и онтогенетическую структуру ЦП кровохлебки лекарствен233 ной (Sanguisorba officinalis L.) – многолетнего травянистого лекарственного растения в национальном парке «Нижняя Кама». В работе использовали общепринятые популяционно-онтогенетические и геоботанические методы, а также методику определения экологической валентности, стеноэврибионтности видов (Работнов, 1950; Уранов, 1975; Жукова, 1995, 2004; Животовский, 2001). В 1989 г кровохлебка включена в «Перечень дикорастущих растений, подлежащих полной охране на всей территории ТАССР» (цит. по: Иванова, 1988). В связи с хозяйственной деятельностью человека (затоплением пойм Волги и Камы) вид S. officinalis сократил свои местообитания, становится редким. Сбор материала проводился в 1997 году (ЦП 1 и ЦП 2) на Елабужских лугах и в 2010 году (ЦП 3 и ЦП 4) в Тукаевском районе. На территории национального парка «Нижняя Кама» изучено 4 луговых фитоценоза с ЦП S. officinalis. Результаты обработки флористических списков геоботанических описаний исследованных фитоценозов с ЦП S. officinalis по шкалам Л. Г. Раменского с соавторами (Экологическая оценка…, 1956) с использованием программы EcoScaleWin (Компьютерная обработка…, 2008) показали, что изученные ЦП имеют сходные экологические условия и характеризуются умеренно-переменным увлажнением (10–12), средним почвенным богатством (10–13), слабой аллювиальностью (1–2). Следовательно, по всем полученным данным модельный вид укладывается в диапазоны экологического ареала этого вида, предложенные Л. Г. Раменским с соавторами (Экологическая оценка…, 1956). Для получения достоверной картины по экологической приуроченности данного вида, произрастающего в исследуемых сообществах, нами были использованы результаты обработки геоботанических описаний и рассчитана потенциальная и реализованная экологическая валентность, индекс толерантности и коэффициент экологической эффективности, предложенные Л. А. Жуковой (2004). Анализ экологического разнообразия S. officinalis в ЦП 1–4 позволяет отнести изучаемый нами вид к мезовалентной фракции валентности по переменности увлажнения, аллювиальности, почвенного богатства и засоленности почвы, а по степени увлажнения – к стеновалентной (табл.). Следовательно, фактор увлажнения является лимитирующим для кровохлебки лекарственной. Показатели коэффициента экологической эффективности исследуемого вида показали, что экологические возможности реализованы от 0,19 (по шкале богатства и засоленности (NS) до 0,65 по шкале увлажнения (FE). 234 Экологическая характеристика S. officinalis Диапазон экологического ареала S. officinalis 1 2 3 4 FE (1–120) 58–86 66,0 82,0 64,0 VF (1–20) 7–16 12,0 10,5 NS (1–30) 5–21 13,0 А (1–10) 1–6 1,0 Экологические шкалы Ценопопуляции PEV REV kek. ef. 67,5 0,23 0,15 0,65 10,0 10,0 0,45 0,10 0,22 10,0 10,2 10,1 0,53 0,10 0,19 2,0 2,0 1,5 0,5 0,10 0,20 Примечание: FE – шкала увлажнения, VF – шкала переменности увлажнения, NS – шкала почвенного богатства и засоленности, А – шкала аллювиальности; PEV – потенциальная экологическая валентность, REV – реальная экологическая валентность, kec. ef. – коэффициент экологической валентности. При анализе строения надземной и подземной сфер у особей S. officinalis выявлено 4 жизненные формы: стержнекорневая, стержнекорневая-короткокорневищная, короткокорневищная и клубнеобразующая (рис. 1) и морфологическое разнообразие листьев S. officinalis, проявляющееся в расположении листочков в сложном листе, крае и форме листовой пластинки (рис. 2). Проведенное изучение морфологической поливариантности S. officinalis позволило установить, что на морфологические структуры как подземных, так и надземных побегов могут влиять экзогенные факторы (условия произрастания). Результаты исследования онтогенетической структуры показали, что ЦП 1 и ЦП 2 S. officinalis были зрелыми нормальными неполночленными, так как отсутствовали особи в субсенильном и сенильном состояниях. Онтогенетические спектры – одновершинные с максимумом на группе особей в средневозрастном генеративном состоянии. О зрелости ЦП 1 и 2 S. officinalis свидетельствовали высокие индексы возрастности (0,375 и 0,341 соответственно) и эффективности (0,676 и 0,664 соответственно). В связи с преобладанием взрослых над молодыми растений индексы восстановления были невысокими (0,47 и 0,55 соответственно). ЦП 3 – зрелая нормальная неполночленная, из-за отсутствия проростков и растений в ювенильном, имматурном, субсенильном и сенильном состояниях. Онтогенетический спектр – одновершинный с максимумом на группе особей в средневозрастном генеративном состоянии. О зрелости ЦП 1 свидетельствуют и высокие индексы возрастности (0,405) и эффективности (0,802), но процессы самоподдержания затруднены, так как индекс восстановления невысокий (0,33). 235 v v g1 g3 стержнекорневая стержнекорневая короткокорневищная короткокорневищная клубнеобразующая Рисунок 1 – Морфологическая поливариантность подземных органов особей S. officinalis v g1 g2 Рисунок 2 – Морфологическая поливариантность надземных органов особей S. officinalis g2 ЦП 4 S. officinalis – стареющая нормальная, но неполночленная, так как отсутствовали проростки и растения в ювенильном, имматурном и сенильном состояниях. Онтогенетический спектр – одновершинный с максимумом на группе особей в старом генеративном состоянии. В связи с преобладанием взрослых растений индексы возрастности (Δ = 0,546) и эффективности (ω = 0,639) достаточно высоки. Небольшой индекс восстановления (0,33) свидетельствует о трудностях самоподдержания в ЦП 2 S. officinalis. Таким образом, изученные ЦП 1 и ЦП 2 были зрелыми нормальными, с подростом, что говорит об экологических условиях, благоприятных для самоподдержания S. officinalis в 1997 г., а ЦП 3 и ЦП 4 S. officinalis были нормальными (соответственно зрелой и стареющей), но неполночленными. Отсутствие подроста свидетельствует о том, что процессы самоподдержания затруднены. Возможно, это связано с жарким и сухим вегетационным периодом 2010 г. Л и т е р а т ур а Глотов Н. В. Об оценке параметров возрастной структуры ценопопуляций растений / Н. В. Глотов // Жизнь растений в гетерогенной среде. – Йошкар-Ола: Периодика Марий Эл, 1998. – Ч. 1. – С. 146–149. Жукова Л. А. Популяционная жизнь луговых растений / Л. А. Жукова. – Йошкар-Ола: РИИК «Ланар», 1995. – 223 с. Жукова Л. А. Методология и методика определения экологической валентности, стеноэврибионтности видов растений / Л. А. Жукова // Методы популяционной биологии: сб. материалов VII Всероссийского популяционного семинара. – Сыктывкар, 2004. – Ч. 1. – С. 75–76. Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений / Л. А. Животовский // Экология. – 2001. – № 1. – С. 3–7. Иванова Р. Г. Словарь-справочник по флоре Татарии / Р. Г. Иванова – Казань: Татарское кн. изд-во, 1988. – 304 c. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoScaleWin / Е. В. Зубкова, Л. Г. Ханина, Т. И. Грохлина, Ю. А. Дорогова. – Йошкар-Ола, 2008. – 96 с. Работнов Т. А. Вопросы изучения состава популяций для целей фитоценологии / Т. А. Работнов // Проблемы ботаники. – М.; Л., 1950. – Вып. 1. – С. 465– 483. Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов / А. А. Уранов // Биол. науки. – 1975 – № 2. – С. 7–34. Экологическая оценка кормовых угодий по растительному покрову / Л. Г. Раменский, И. А. Цаценкин, О. Н. Чижиков и др. – М.: Сельхозгиз, 1956. – 472 с. ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ TANACETUM VULGARЕ L. Ведерникова О. П., Ягодарова О. С. Марийский государственный университет, г. Йошкар-Ола, ecology@marsu.ru В настоящее время особый интерес представляют популяции лекарственных растений как объекты всестороннего исследования. Чрезвычайно актуальна оценка их современного состояния, требующая деталь237 ных популяционных исследований, что невозможно без использования популяционно-онтогенетического подхода, а именно без описания онтогенеза и структуры ценопопуляций (ЦП). Целью данной работы было изучение онтогенетической структуры ЦП многолетнего короткокорневищного или длиннокорневищного травянистого растения Tanacetum vulgarе L, имеющего лекарственное значение. Сбор материал проводили на территории Сернурского района Республики Марий Эл в трех фитоценозах (ЦП 1 и ЦП 3 – в рудеральных сообществах, ЦП 2 – в луговом сообществе). В работе использовали общепринятые популяционно-онтогенетические и геоботанические методы (Работнов, 1950; Уранов, 1975; Жукова, 1995; Глотов, 1998; Животовский, 2001). Результаты обработки флористических списков геоботанических описаний исследованных фитоценозов с ЦП T. vulgarе по шкалам Л. Г. Раменского с соавторами (Экологическая оценка…, 1956) с использованием программы EcoScaleWin (Компьютерная обработка…, 2008) показали, что изученные ЦП имели сходные экологические условия местообитаний: довольно богатые почвы (11,0–12,0), сухолуговое и свежелуговое увлажнение (62,0–63,0), умеренно переменное увлажнение (10,0–12,0), слабое влияние выпаса, сенокосная стадия (2,5–3,0), очень слабо аллювиальные почвы (0,1–0,3 см наилка). Таким образом, на территории Сернурского района Республики Марий Эл экологическая характеристика местообитаний ЦП T. vulgarе по анализируемым факторам соответствует диапазонным оценкам, предложенным Л. Г. Раменским с соавторами (Экологическая оценка…, 1956) для данного вида. Анализ онтогенетической структуры ЦП T. vulgarе показал, что все ЦП зрелые нормальные, но неполночленные, из-за отсутствия в ЦП 1 – особей прегенеративного периода и сенильных растений, в ЦП 2 – особей в прегенеративном и постгенеративном периодах, а в ЦП 3 – проростков, особей в ювенильном и имматурном состояниях. Результаты исследования онтогенетической структуры ЦП T. vulgarе в изученных сообществах сведены в таблицу. ЦП 1 T. vulgaris – зрелая, нормальная, с максимумом на группе особей в средневозрастном генеративном состоянии (39,7 %). Индекс эффективности самый низкий (0,529), что свидетельствует о наименьшем воздействии на среду из всех изученных ценопопуляций. Коэффициент возрастности составляет 0,327. Индекс старения небольшой и составлял 0,012. Экологические условия в данном рудеральном сообществе оказались более благоприятными для особей T. vulgaris, о чем свидетельствует самая высокая плотность (табл.). 238 Некоторые популяционные параметры ценопопуляций Tanacetum vulgarе L. № ЦП Тип ЦП 1. Индексы эффективности возрастности старения Плотность, шт./м2 Зрелая нормальная неполночленная 0,529 0,327 0,012 31,2 2. Зрелая нормальная неполночленная 0,772 0,454 – 26,8 3. Зрелая нормальная неполночленная 0,715 0,477 0,057 21,2 ЦП 2 T. vulgaris также можно отнести к зрелой нормальной. Она полностью состоит из особей генеративного периода, что составляет 100 %. Генеративный период – наиболее длительный в жизни многолетних растений. Длительное нахождение особей в генеративном периоде способствует увеличению численности групп, составляющих этот период. Максимум приходился на группу особей в средневозрастном генеративном состоянии (37,3 %). Таким образом, максимальное число составляют наиболее жизнедеятельные особи пижмы обыкновенной, прошедшие половину онтогенеза. Это является показателем зрелых популяций. О зрелости свидетельствуют высокие коэффициент возрастности (0,454) и индекс эффективности (0,772). Индекс эффективности максимальный, поэтому воздействие на среду ЦП 2 T. vulgaris максимальное. Экологические условия на суходольном лугу оказались наиболее благоприятными для особей T. vulgaris. (табл.). В онтогенетическом спектре ЦП 3 T. vulgaris максимум приходился на группу особей в средневозрастном генеративном состоянии (34 %). Это является показателем зрелых нормальных популяций. Постгенеративный период представляют особи в субсенильном и сенильном состояниях. На эти онтогенетические группы приходилось 3,8 и 1,9 % соответственно. Максимальную долю составили особи генеративного периода (94,3 %). Экологические условия обитания в данном рудеральном сообществе оказались менее благоприятными для особей T. vulgaris, чем на суходольном лугу (ЦП 2) и в рудеральном сообществе (ЦП 1), об этом свидетельствует самая низкая плотность из всех изученных ЦП (табл.). Для ЦП T. vulgaris на территории Сернурского района Республики Марий Эл характерен одновершинный базовый спектр. Колебания демографических параметров связаны с особенностями прохождения онтогенеза в разных экологических условиях. Во всех изученных ЦП пре- 239 обладают особи генеративного периода, индексы эффективности и коэффициенты возрастности достаточно высокие. Следовательно, полученные данные дают возможность для практических рекомендаций по оценке состояния ЦП, определения ресурсов и их рационального использования для сбора лекарственного сырья на исследованной территории. Л и т е р а т ур а Глотов Н. В. Об оценке параметров возрастной структуры ценопопуляций растений / Н. В. Глотов // Жизнь растений в гетерогенной среде. – Йошкар-Ола: Периодика Марий Эл, 1998. – Ч. 1. – С. 146–149. Жукова Л. А. Популяционная жизнь луговых растений / Л. А. Жукова. – Йошкар-Ола: РИИК «Ланар», 1995. – 223 с. Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений / Л. А. Животовский // Экология. – 2001. – № 1. – С. 3–7. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoScaleWin / Е. В. Зубкова, Л. Г. Ханина, Т. И. Грохлина, Ю. А. Дорогова. – Йошкар-Ола, 2008. – 96 с. Работнов Т. А. Вопросы изучения состава популяций для целей фитоценологии / Т. А. Работнов // Проблемы ботаники. – М.; Л., 1950. – Вып. 1. – С. 465–483. Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов / А. А. Уранов // Биол. науки. – 1975 – № 2. – С. 7–34. Экологическая оценка кормовых угодий по растительному покрову / Л. Г. Раменский, И. А. Цаценкин, О. Н. Чижиков и др. – М.: Сельхозгиз, 1956. – 472 с. БИОЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ, РАЗМНОЖЕНИЕ И ЗНАЧЕНИЕ КСАНТОЦЕРАСА РЯБИНОЛИСТНОГО (ЧЕКАЛКИН ОРЕХ) – XANTHOCERAS SORBIFOLIA BUNGE НА АПШЕРОНСКОМ ПОЛУОСТРОВЕ АЗЕРБАЙДЖАНА Гасанова М. Ю., Алиева С. А., Гулиев Ф. Б. Мардакянский дендрарий НАН Азербайджана, г. Баку, dendrary@mail.az, 2013@gmail.com, offis.dendrari@hotmail.com С 1926 г. в коллекции Мардакянского дендрария Апшеронского полуострова собраны коллекции древесно-кустарниковых растений, относящихся к различным дендрофлорам. Были проведены научно-исследовательские работы над видами, относящимися к различным дендрофлорам, всесторонне изучены биоэкологические особенности этих растений. Было выявлено, что виды растений из различной флоры обладают широкой экологической амплитудой. У видов растений из различных дендрофлор в условиях Апшерона, в Мардакянском дендрарии все фенологические фазы прошли нормально, образовались цветки, плоды и семена. Многие годы проводятся научно-исследовательские работы с целью интродукции, акклиматизации и размножения с помощью семян древесно-кустарниковых видов. В Мардакянском дендрарии Апше240 ронского полуострова были проведены научно-исследовательские работы над ксантоцерасом рябинолистным (Xanthoceras sorbifolia Bge.) из семейства Sapindaceae Juss. – Сапиндовые. X. sorbifolia – представитель Восточно-Азиатской флоры. Этот вид был впервые обнаружен в конце ХIX века, в 1868 г. со стороны Аббе Девида в Северном Китае. А на Апшеронском полуострове Азербайджана в Мардакянском дендрарии этот вид выращивается только с 1936 года. Этот вид полностью адаптирован к сухому, субтропическому почвенно-климатическому условию Апшерона, всесторонне изучены его биоэкологические особенности. X. sorbifolia – морозоустойчивое растение. Причиной зимостойкости этого вида, как показали исследования в Мардакянском дендрарии, ли изменения физиологических функций, характеризующих предков древесных растений холодного, умеренного и субтропического климатов (Аббасов и др., 1978; Мамедов, 2010). Зимостойкость вырабатывалась в результате исторического развития растений в зависимости от условий внешней среды. В повышении зимостойкости важную роль сыграло изменение функциональных особенностей запасных веществ, что выражается в превращении этих веществ в защитные, в зависимости от погодных условий времени года. По своим биологическим особенностям вступает в нормальную фазу цветения весной. X. sorbifolia – листопадное дерево или кустарник высотой 6–8 метров. Листорасположение очередное. Листья сложные, без прилистников, непарноперистые, с 9–15 листочками, до 25 см длиной. Листочки сидячие, обычно супротивные, иногда очередные, ланцетные, длиной 3– 5 см, шириной 0,7–1,2 см, остропильчатые, блестящие, кожистые, сверху темно-, снизу светло-зеленые с курчавыми волосками, которые после формирования листьев опадают. Соцветие – густая, многоцветковая кисть, длиной до 25 см, расположены на концах побегов и укороченных боковых веточках. Цветки крупные, белые, правильные, обоеполые и тычиночные, диаметром до 4 см, на цветоножках длиной около 1,5 см; чашелистиков 5, продолговатые, длиной 6–7 мм; лепестков 5, обратнояйцевидные, длиной 2 см, шириной 8 мм; тычинок 8; завязь 3-гнездная; рыльце головчато-трехраздельное. X. sorbifolia – однодомное растение. Цветение начинается со второй декады апреля по первую декаду мая и продолжается до полного распускания листьев. Плод – треугольно-яйцевидная или эллиптическая коробочка, диаметром до 5 см. Созревание плодов начинается в августе, а в сентябре они полностью созревают. 241 В трехгнездной коробочке жится до 15–17 семян (в каждом де по 5–6 семян). Семена длиной до 1,8 см, диаметром 1,5 см похожи на мелкие орехи, съедобные (рис. 1). По мере созревания семян коробочка становятся темно-коричневого цвета, верхушка ее растрескивается, и зрелые семена высыпаются. Взрослое растение не очень требовательно к поливу, поскольку этот вид засухоустойчив. Растения на ранних этапах онтогенеза также нуждаются в умеренном поливе. Была отмечена гибель 1–3-летних растений, поливаеРисунок 1 – Внешний вид семян X. sorbifolia мых через день. Такие растения поливают один раз в 8–10 дней. После каждого орошения проводятся агротехнические мероприятия. Рекомендуется посадка светолюбивого растения сразу на постоянный участок, поскольку плохо переносит пересадку. Годовой прирост составляет 22 см. Развитие идет очень быстро. Интенсификация развития продолжается с апреля до конца августа. Хотя X. sorbifolia малотребователен к почвенным условиям, оно лучше развивается на плодородных почвах, на открытых, солнечных участках. Размножается X. sorbifolia семенами и корневыми черенками, возможно и естественное возобновление. Семена не нуждаются в стратификации и в благоприятных условиях способны к прорастанию независимо от времени года. В суровую зиму проростки, полученные от семян, высеянных осенью, погибают. По этой причине семена должны быть высеяны во второй декаде марта (14.03). Глубина заделки семян составила 3–4 см, расстояние между высеянными семенами – 4–5 см. Прорастание семян подземное Рисунок 2 – Молодое вегетативное (рис. 2). Первые всходы были полурастение X. sorbifolia чены 3 апреля, а массово они появи70–75-дневного возраста 242 лись 7 апреля. Первый ассимилирующий лист появился 10 апреля. Всхожесть семян составила 85–90 %. Растение обладает очень мощной корневой системой. После 70– 75 дней длина корневой системы молодых растений достигает 10– 12 см. Растение, выросшее из семян на третий год, цветет и плодоносит. Семенной материал хранят в мешочках, в сухом месте. В таких условиях семена сохраняют всхожесть до 22–24 месяцев. В Мардакянском дендрарии X. sorbifolia используется в озеленении, т. к. обладает высокими декоративными качествами. Красивые соцветия с белыми цветками, многочисленные крупные листья с блестящими листовыми пластинками, крупные, зеленые плоды и семена в раскрывшихся темно-каштановых коробочках придают растению особую эстетическую красоту. Во время фазы цветения, X. sorbifolia высотой 6–8 м полностью покрывается белыми цветами, похожими на мелкие, белые звездочки. Этот необыкновенный вид растения некоторые дендрологи рассматривают как «цветовой» куст и называют «красивым цветовым кустом». В фазе массового цветения декоративность растения проявляется еще больше. Развитие ствола дерева в курчавом виде делает растение привлекательным. X. sorbifolia используется в озеленении как одиночно, так и в групповых посадках. Таким образом, X. sorbifolia является очень ценным декоративным растением, которое используется в озеленении на основе ландшафтной архитектуры. Л и т е р а т ур а Аббасов Р. М. Мардакянский дендрарий / Р. М. Аббасов, У. М. Агамиров, Ф. М. Мамедов, А. М. Садыхов. – Баку: Элм, 1978. – 19 с. Мамедов Т. С. Деревья и кустарники Апшерона. – Баку: Елм вя тящсил, 2010. – 194 с. ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА SCUTELLARIA GALERICULATA L. Гусева А. А. ЦСБС СО РАН, г. Новосибирск, guseva. sc@list.ru Scutellaria galericulata L. – шлемник обыкновенный – многолетние симподиально нарастающее длиннокорневищное растение. Вид широко распространен в России и за ее пределами, заходит в страны Скандинавии, Европы, Средиземноморья, Балканский полуостров, Монголию, Китай и Японию. Произрастает по берегам рек, на сырых и заболоченных лугах, березовых лесах (Зуев, 1997). В настоящее время изучается как перспективное лекарственное растение с выраженным гипотензивным и седативным свойствами (Дудецкая, 2011). 243 Изучена онтогенетическая структура двух ценопопуляций (ЦП) S. galericulata по общепринятым методикам (Серебряков, 1962; Уранов, 1967; Ценопопуляции…, 1976). При изучении онтогенетической структуры за счетную единицу принимался парциальный побег. Материал собран в Хакасии и Красноярском крае. ЦП № 1 была описана по берегу реки Большой Монок в окрестности поселка Большой Монок в Хакасии (h = 412 м, N 52o51’31.3’’, E 090o17’12.1’’). Общее проективное покрытие (ОПП) – 70 %. ЦП № 2 изучена в осоковых зарослях по берегу реки Хабык в окрестностях поселка Малый Хабык в Красноярском крае (h = 273 м, N 54o26’30.0’’, E 092o04’32.7’’). ОПП – 100 %. Изученные ценопопуляции нормальные, неполночленные. Плотность особей высокая (13,5–23,6 особи на м2). Онтогенетические спектры в обеих ЦП центрированные с преобладанием средневозрастных генеративных особей. Отсутствие семенных особей указывает на то, что поддержание ЦП осуществляется только вегетативным путем. Большое число виргинильных особей в ЦП № 2, незначительная доля имматурных и отсутствие ювенильных особей в ЦП № 1, 2, говорит о том, что вегетативное размножение в основном происходит с омоложением рамет до виргинильного состояния. Онтогенетическая структура изученных ценопопуляций По классификации «дельта-омега» (Животовский, 2001) ЦП № 1 является зрелой (∆ = 0,38, ω = 0,84). Максимум на средневозрастной генеративной фракции связан с продолжительным нахождением особей в g2 онтогенетическом состоянии, увеличение численности от имматурных до молодых генеративных определяется вегетативным размножением с омоложением особей. ЦП № 2 является зреющей (∆ = 0,33, ω = 0,76). Накопление особей в виргинильном онтогенетическом состоянии определяется интенсивным вегетативным размножением в условиях повы244 шенной плотности травостоя (ОПП – 100 %). Отсутствие постгенеративного периода в обеих ЦП связано с быстрым отмиранием старых особей и определяется биологией вида. Л и т е р а т ур а Дудецкая Н. А. Состав и содержание фенольных соединений в надземной части Scutellaria galericulata (Lamiaceae) / Н. А. Дудецкая // Растительные ресурсы. – 2011. – Т. 47, вып. 4. – С. 95–105. Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений // Экология, 2001. – № 1. – С. 3–7. Зуев В. В. 3. Scutellaria L. – Шлемник / В. В. Зуев // Флора Сибири. Pyrolaceae-Lamiaceae (Labiatae) / под ред. Л. И. Малышева. – Новосибирск, 1997. – Т. 11. – С. 161–165. Серебряков И. Г. Экологическая морфология растений. Жизненные формы покрытосеменных и хвойных / И. Г. Серебряков. – М.: Высш. шк., 1962. – С. 378. Уранов А. А. Онтогенез и возрастной состав популяций (вместо предисловия) А. А. Уранов // Онтогенез и возрастной состав ценопопуляций цветковых растений. – М.: Наука, 1967. – С. 3–8. Ценопопуляции растений: (основные понятия и структура). – М.: Наука, 1976. – С. 215. ВЛИЯНИЕ ЭКОЛОГИЧЕСКИХ ФАКТОРОВ НА БИОРАЗНООБРАЗИЕ ФЛОРЫ ИЛИСУИНСКОГО ГОСУДАРСТВЕННОГО ПРИРОДНОГО ЗАПОВЕДНИКА АЗЕРБАЙДЖАНА Дадашева А. Г., Абдыева Р. Т. Институт ботаники Национальной академии наук Азербайджана, г. Баку, aida_dadashova@mail.ru Одной из актуальных и важнейших задач на сегодняшний день является защита окружающей среды, охрана и восстановление естественно-природных ресурсов Земли. На территории Азербайджана организованы десятки государственных заповедников, национальных парков и заказников, целью которых является сохранение и увеличение численности редких и исчезающих видов флоры и фауны, восстановление уникальных биогеоценозов, ценных древесных лесных пород. Одной из таких заповедных зон является Илисуинский государственный природный заповедник площадью около 17 381 га, основанный в 1987 г. (рис. 1). Илисуинский государственный природный заповедник – один из живописнейших уголков республики, находится в северо-западной части Азербайджана, на территории Гахского административного района и составляет 11,63 % его общей площади. Территория, занимаемая заповедником, относится к области Большого Кавказа. Рельеф отличается весьма сложной структурой. Почвы горно-луговые дерновые, рыхлодерновые, плотно-дерновые, горно-лугово-лесные, бурые лесные. В высотном отношении расположен в пределах 600–2300 м н. у. м. Климат изменяется от умеренно-теплого влажного до умеренно-холодного 245 влажного. Среднегодовая температура воздуха составляет 11,1– 13,2 °С. Среднегодовое количество осадков 500–1600 мм (Мусеибов, 2001). Территория заповедника покрыта речной сетью. Основными водными бассейнами являются реки Кюрмюкчай, Хамалчай, Агчай, Гумчай и Лекитчай. Рисунок 1 – Географическое расположение Илисуинского государственного природного заповедника Флора Илисуинского государственный природного заповедника изучалась Т. А. Абдуллаевой еще в 1996 году (Абдуллаева, 1996). В связи с динамично протекающими экологическими процессами территория заповедника за два с лишним десятилетия претерпела ряд изменений: действующие здесь различные физические и биотические факторы оказывают влияние на его флору и растительность. Учитывая вышеизложенное, было решено провести мониторинг территории этой заповедной зоны. Рекогносцировочное обследование проводилось в период 2012–2013 гг. В растительном покрове заповедника наблюдается сочетание лесной, луговой, кустарниковой растительности и редколесий. Характерным элементом его фитоландшафта являются высокогорные леса, образованные такими лесными породами, как Fagus orientalis Lipsky (бук восточный), Carpinus orientalis Mill. (граб восточный), Quercus iberica 246 Stev. (дуб иберийский), Q. macranthera Fisch. & C. A. Mey. ex Hohen. (дуб восточный) и т. д. В нижнем ярусе лесов встречаются кустарники (Mespilus germanica L. (мушмула кавказская), Prunus spinosa L. (слива колючая), Corylus avellana L. (лещина обыкновенная), Cornus mas L. (кизил обыкновенный) и виды рода Rosa L. (шиповник) и т. д.). Для лугов типичны разнотравные фитоценозы с участием видов родов Campanula L., Inula L., Heracleum L., Veratrum L., Alchemilla L. и др. В злаковых фитоценозах обычны представители родов Phleum L., Bromus L., Festuca L., Calamagrostis Adans. и др. Из числа реликтовых, редких и исчезающих видов, внесенных в «Красную книгу Азербайджана» (2013), в заповеднике охраняются Castanea sativa Mill. (каштан съедобный), Taxus baccata L. (тисс ягодный), Juniperus foetidissima Willd. (можжевельник тяжелопахучий), Hedera pastuchowii Woron. (плющ пастухова), Rosa azerbajdzhanica Novopokr. (шиповник азербайджанский) и т. д. (Красная Книга Азербайджанской Республики, 2013). В регионе в последние годы наблюдается увеличение нормы выпадающих осадков, особенно в весенний период, когда растительность начинает вегетацию. Ливневый характер дождей вызывает мощные наводнения и сели, которые наносят ущерб не только населенным пунктам, народному хозяйству, но и растительному покрову заповедника. Возникающие эрозии почвы воздействуют на нормальный ход развития растительности. В частности, наблюдается изменение экологии местообитания растений, их смыв и слабое возобновление (рис. 2, 3). Рисунок 2 – Размыв почвы под действием селей 247 Рисунок 3 – Эрозия почвы Вымывая минеральные вещества из верхних горизонтов почвы и нарушая механическую структуру, сели вызывают разрушение ее верхних слоев. В результате снижается продуктивность и качественный состав почв. Отрицательные экзогенные процессы, протекающие на территории заповедника, снижают биоразнообразие его флоры, вызывают деградацию растительности и почвы. Участки, подвергающиеся этим воздействиям, требуют повышенного внимания как с научной, так и с практической стороны. В связи с этим необходимы более длительные и глубокие исследования, конечной целью которых является оптимизация окружающей среды заповедника. Л и т е р а т ур а Абдуллаева Т. А. Флористический анализ и флора Илисуинского Государственного Заповедника: дис. … канд. биол. наук / Т. А. Абдуллаева. – Баку, 1996. – 164 с. (на азерб. яз.). Мусеибов М. А. Физическая география Азербайджана / М. А. Мусеибов. – Баку: Элм, 2001. – 200 с. (на азерб. яз.). Красная Книга Азербайджанской Республики. Редкие и исчезающие виды растений и грибов. – Баку, 2013. – 673 с. (на азерб. яз.). 248 ВЛИЯНИЕ ЖИЗНЕННОСТИ НА МОРФОМЕТРИЧЕСКИЕ И ПРОДУКЦИОННЫЕ ХАРАКТЕРИСТИКИ ПОДРОСТА PICEA OBOVATA LEDEB. Ефименко А. С., Алейников А. А. Центр по проблемам экологии и продуктивности лесов РАН, г. Москва, aseforests@gmail.com Для оценки успешности естественного возобновления и прогнозирования дальнейшего развития лесных сообществ необходимо исследовать подрост лесообразующих видов. В природных лесах со сложной вертикальной структурой для подроста характерна разная жизненность, обусловленная фитоценотическим угнетением. Жизненное состояние подроста проявляется, прежде всего, в вегоразмерных и продукционных характеристиках. Цель исследования – изучить биометрические и продукционные показатели подроста P. obovata с учетом жизненного состояния в темнохвойных лесах Северного Предуралья. Изучение биоморфологических особенностей подроста ели сибирской проводилось в верховьях реки Печоры (Печоро-Илычский заповедник), где уже несколько лет ведутся стационарные исследования малонарушенных лесных экосистем (Смирнова и др., 2011; Алейников, Ефименко, 2012; Луговая и др., 2013). В приведенных выше работах дана подробная характеристика района исследования. Модельные особи подроста ели сибирской были отобраны в бореально-мелкотравных и крупнопапоротниковых сообществах. К подросту отнесены особи начальных этапов онтогенеза: ранние имматурные (im1), поздние имматурные (im2) и ранние виргинильные (v1) растения (Диагнозы…, 1989; Методические…, 2010). Жизненность оценивали по трехбалльной шкале (нормальная, пониженная и низкая) с учетом внешних признаков растения (Злобин, 2009). У каждого растения определяли: календарный возраст, высоту, диаметр стволика у основания, приросты по высоте за последние 5 лет, протяженность кроны по стволику, а также фитомассу хвои, ветвей и стволика. Календарный возраст im1-растений устанавливали путем подсчета линейных приростов по высоте. Возраст im2- и v1-особей определяли по годичным кольцам на спилах у основания стволика. Для исследования надземной фитомассы ели сибирской с помощью секатора и складной пилы Fiskars каждую модельную особь подроста разделяли на отдельные части: ствол, живые и сухие ветви, хвою. В представленной работе фитомасса ветвей рассматривается как сумма живых и сухих ветвей. Для удобства сушки крупные фракции (стволик и ветви) измельчали. Далее каждую фракцию древесного растения помещали в бумажные пакеты, на каждом 249 пакете отмечали номер и вид особи, а также наименование фракции. Затем модельные образцы сушили до достижения воздушно-сухого веса. После этого каждая фракция была взвешена на весах, при этом хвою ранних имматурных растений взвешивали с точностью до 0,0001 г, поздних имматурных и ранних виргинильных – до 0,01 г. Крупные фракции (ветви и стволики) поздних имматурных и ранних виргинильных состояний взвешивали с точностью до 0,1 и 1,0 г соответственно, а ранних имматурных до 0,005 г. Для описания im1-растений проанализированы морфологические показатели у 34 модельных особей, для im2 и v1 – 16 и 15 растений соответственно. Продукционные показатели растений каждого возрастного состояния исследовали на 15 модельных особях. Оценку влияния жизненности на морфологические и продукционные показатели проводили с помощью однофакторного дисперсионного анализа в программе STATISTICA9. Ранний имматурный этап развития у ели сибирской начинается с ветвления стволика и образования первого бокового побега (Давыдычев, Кулагин, 2010; Методические…, 2010). Это состояние характеризуется постепенным увеличением порядка ветвления и формированием рыхлой кроны с недолговечными боковыми побегами. Как правило, im1-растения небольших размеров, с редкой несформированной кроной, состоящей из 1–2 ветвей. Средний возраст особей в этом возрастном состоянии – 10,0 ± 0,55 лет, высота – 22,9 ± 1,37 см. Средний прирост по высоте за последние 5 лет составляет 2,5 ± 0,15 см. Среди морфометрических показателей однофакторный дисперсионный анализ подтвердил достоверное влияние жизненного состояния на календарный возраст и средний прирост по высоте за последние 5 лет. Анализ продукционных характеристик показал, что фитомасса основных фракций у растений разной жизненности достоверно не отличается (табл. 1). Поздние имматурные растения по сравнению с особями предыдущего возрастного состояния характеризуются наибольшей высотой и наибольшими приростами по высоте, а также более сложной, зонтикообразной, сформированной кроной. Календарный возраст im2-растений – 50,7 ± 5,05 лет. Средняя высота особей в этом возрастном состоянии составляет – 150,0 ± 6,94 см, а средний прирост по высоте за последние 5 лет – 4,6 ± 0,72 см. Основные биометрические и продукционные показатели представлены в таблице 2. В результате исследования выявлено, что im-2-растения разной жизненности достоверно отличаются по календарному возрасту, среднему приросту по высоте за последние 5 лет и протяженности кроны. По фитомассе основных фракций растения разной жизненности не отличаются (табл. 2). 250 Таблица 1 – Биометрические и продукционные показатели im1-растений разной жизненности Показатели Возраст, лет Высота, см Диаметр стволика, мм Средний годовой прирост, см/год Протяженность кроны, см Фитомасса хвои, г Фитомасса ветвей, г Фитомасса стволиков, г Жизненность Статистические показатели n М ±m 1 12 8,8 0,35 1,22 σ CV, % 13,9 2 16 10,0 0,97 3,86 38,6 3 6 12,7 1,20 2,94 23,2 1 12 23,5 1,87 6,48 27,6 2 16 22,8 2,48 9,91 43,5 3 6 22,0 2,30 5,64 25,6 1 12 3,6 0,28 0,98 27,0 2 16 3,5 0,40 1,59 45,1 3 6 3,2 0,48 1,17 36,5 1 12 3,2 0,26 0,91 28,6 2 16 2,1 0,15 0,58 27,1 3 6 1,9 0,14 0,34 17,9 1 12 19,7 1,90 6,57 33,3 2 16 16,1 1,68 6,71 41,6 3 6 14,6 2,02 4,95 34,0 1 5 1,01 0,27 0,60 59,2 2 5 0,97 0,26 0,58 60,0 3 5 0,57 0,16 0,35 62,1 1 5 0,43 0,12 0,27 63,4 2 5 0,55 0,16 0,36 65,3 3 5 0,41 0,14 0,30 74,6 1 5 0,98 0,22 0,48 49,1 2 5 1,78 0,57 1,28 72,0 3 4 1,12 0,26 0,51 45,6 р 0,047 0,935 0,815 0,0004 0,201 0,362 0,74 0,342 Примечания: n – число измерений; М – среднее значение; ±m – ошибка среднего значения; σ – стандартное отклонение; СV – коэффициент вариации, %; р – уровень достоверности. Курсивом выделены статистически значимые уровни достоверности. Уровни жизненности: 1 – нормальная, 2 – пониженная, 3 – низкая. 251 Таблица 2 – Биометрические и продукционные показатели im2-растений разной жизненности Показатели Возраст, лет Высота, см Диаметр стволика, мм Средний годовой прирост, см/год Протяженность кроны, см Фитомасса хвои, г Фитомасса ветвей, г Фитомасса стволиков, г Жизненность Статистические показатели n М ±m σ CV, % 1 5 33,8 3,14 7,01 20,8 2 6 45,2 3,09 7,57 16,8 3 5 74,2 7,95 17,77 23,9 1 5 152,3 17,12 38,27 25,1 2 6 140,3 6,52 15,97 11,4 3 5 159,4 13,03 29,13 18,3 1 5 25,8 2,68 6,00 23,2 2 6 23,3 3,47 8,49 36,5 3 5 26,4 2,95 6,59 24,9 1 5 7,2 1,27 3,11 43,2 2 6 3,2 0,33 1,10 34,5 3 5 2,1 0,17 0,46 22,3 1 5 114,0 15,40 34,50 30,1 2 6 84,0 5,20 12,90 15,3 3 5 62,0 7,20 16,10 25,8 1 5 100,2 26,31 58,83 58,7 2 6 60,5 14,43 35,35 58,4 3 5 31,0 7,61 17,02 54,8 1 5 179,9 47,50 106,21 59,0 2 5 192,0 75,27 168,30 87,7 3 5 96,9 26,61 59,51 61,4 1 5 104,5 30,44 68,08 65,1 2 5 91,5 13,27 29,68 32,4 3 5 153,6 35,21 78,73 51,2 p 0,0002 0,545 0,507 0,00007 0,009 0,054 0,55 0,29 Примечания: n – число измерений; М – среднее значение; ±m – ошибка среднего значения; σ – стандартное отклонение; СV – коэффициент вариации, %; р – уровень достоверности. Курсивом выделены статистически значимые уровни достоверности. Уровни жизненности: 1 – нормальная, 2 – пониженная, 3 – низкая. Раннее виргинильное состояние характеризуется началом фазы активного роста древесных растений. У них хорошо развит ствол и крона; прирост в высоту приближается к максимальным значениям. Диаметр ствола превышает диаметр скелетных ветвей в 3 раза и более. В этом онтогенетическом состоянии у деревьев всех видов потребности в свете максимальные (Методические…, 2010). Календарный возраст v1-рас252 тений – 76,0 ± 7,91 лет. Средняя высота растений в этом возрастном состоянии – 270,3 ± 7,59 см, средний прирост по высоте за последние 5 лет – 8,7 ± 1,71 см. Таблица 3 – Биометрические и продукционные показатели v1-растений разной жизненности Показатели Возраст, лет Высота, см Диаметр стволика, мм Средний годовой прирост, см/год Протяженность кроны, см Фитомасса хвои, г Фитомасса ветвей, г Фитомасса стволиков, г Жизненность Статистические показатели n М ±m σ CV, % 1 5 45,2 6,73 15,10 33,3 2 4 67,0 8,05 16,10 24,0 3 6 107,7 3,50 8,60 8,0 1 5 273,1 10,66 23,83 8,7 2 4 262,0 12,23 24,45 9,3 3 6 273,6 15,98 39,15 14,3 1 5 41,1 1,74 3,90 9,5 2 4 51,8 4,83 9,66 18,6 3 6 51,1 3,90 9,55 18,7 1 5 15,6 2,01 4,49 28,8 2 4 7,9 1,25 2,50 31,5 3 5 2,5 0,19 0,43 16,8 1 5 223,7 13,4 29,96 13,4 2 4 184 10,89 21,77 11,8 3 6 139,6 16,2 39,69 28,4 1 5 220,6 52,03 116,34 52,7 2 4 567,2 133,04 266,07 46,9 3 6 127,3 36,42 89,2 70,1 1 4 267,3 89,68 179,36 67,1 2 4 995,5 187,61 375,22 37,7 3 6 586,4 197,89 484,72 82,7 1 4 1173 248,72 497,45 42,4 2 4 3302,6 601,99 1204,00 36,5 3 6 2259,3 464,39 1137,50 50,3 p 0,00001 0,826 0,112 0,0001 0,004 0,0003 0,068 0,342 Примечания: n – число измерений; М – среднее значение; ±m – ошибка среднего значения; σ – стандартное отклонение; СV – коэффициент вариации, %; р – уровень достоверности. Курсивом выделены статистически значимые уровни достоверности. Уровни жизненности: 1 – нормальная, 2 – пониженная, 3 – низкая. 253 Однофакторный дисперсионный анализ показал, что v1-растений разной жизненности достоверно отличаются по календарному возрасту, среднему приросту по высоте за последние 5 лет и протяженности кроны. Среди продукционных характеристик достоверные отличия выявлены только по фитомассе хвои. В результате исследования получены данные о зависимости основных морфологических показателей и продукционных характеристик от жизненного состояния растений. Установлено, что на протяжении трех онтогенетических этапов развития растения разной жизненности достоверно отличаются по календарному возрасту и приростам по высоте, а поздние имматурные и ранние виргинильные растения отличаются еще и по протяженности кроны. Из продукционных характеристик достоверные отличия были отмечены только у v1-растений по фитомассе хвои. Полученные результаты подтверждают, что растения разной жизненности характеризуются различной скоростью прохождения возрастных состояний. Кроме того, достоверные отличия прироста по высоте у растений разной жизненности позволяют использовать этот показатель для определения жизненного состояния в полевых условиях. Работа выполнена при поддержке фонда РФФИ (проекты № 13-04-01491а и № 13-04-01521а) и гранта Президента РФ (проект МК-2102.2012.4) Л и т е р а т ур а Алейников А. А. Характеристика естественного возобновления кедра сибирского (Pinus sibirica) в крупнопапоротниковых и высокотравных лесах Северного Предуралья / А. А. Алейников, А. С. Ефименко // Вестник МГУЛ. Лесной вестник. – 2012. – T. 92, № 9. – С. 52–57. Диагнозы и ключи возрастных состояний лесных растений. Деревья и кустарники. – М., 1989. – 106 с. Давыдычев А. Н. Характеристика предгенеративного периода онтогенеза ели сибирской (Picea obovata Ledeb.) в подзоне широколиственнохвойных лесов Уфимского плато / А. Н. Давыдычев, А. Ю. Кулагин // Бюл. МОИП. Отд. биол. – 2010. – Т. 115, вып. 2. – С. 59–66. Злобин Ю. А. Популяционная экология растений: современное состояние, точки роста: монография / Ю. А. Злобин. – Сумы: Университетская книга, 2009. – 263 с. Луговая Д. Л. Микромозаичная организация и фитомасса напочвенного покрова в основных типах темнохвойных лесов Печоро-Илычского заповедника / Д. Л. Луговая, О. В. Смирнова, М. В. Запрудина, А. А. Алейников и др. // Экология. – 2013. – № 1. – С. 3–8. Методические подходы к экологической оценке лесного покрова в бассейне малой реки. – М., 2010. – 383 с. Смирнова О. В. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике / О. В. Смирнова, А. А. Алейников, А. А. Семиколенных, А. Д. Бовкунов, М. В. Запрудина, Н. С. Смирнов // Лесоведение. – 2011. – № 6. – С. 67–78. 254 СОСТОЯНИЕ СМЕШАННЫХ ЛЕСНЫХ ПОСАДОК СОСНЫ ОБЫКНОВЕННОЙ (PINUS SYLVESTRIS L.) В НАЦИОНАЛЬНОМ ПАРКЕ «МАРИЙ ЧОДРА» Забродин И. В. Марийский государственный университет, г. Йошкар-Ола, dom_356@mail.ru Рост и формирования культур древесных растений изучаются длительное время, однако, оптимальные режимы выращивания древостоев окончательно не установлены даже для такого лесного сообщества, как одновозрастные посадки сосны обыкновенной (Pinus silvestris L). В. И. Рубцов (1996) считал вопрос о посадке P. sylvestris главным вопросом, от правильности решения которого зависит успех выращивания ценных насаждений. Познание особенностей формирования посадок с разными типами совместного произрастания позволяет изучить особенности каждого из них. Повышение устойчивости P. sylvestris имеет большое народнохозяйственное значение. Изучение посадок заключалось в наблюдениях за развитием растений и их сохранностью. Число сохранившихся посадок P. sylvestris определялось на основе проведения сплошного перечета деревьев во второй половине вегетационного периода (август–сентябрь). Учет проводился в рядах, которые были замаркированы в виде небольших углублений в почве с обеих сторон каждого учетного ряда. На учетных рядах сплошь считали особи P. sylvestris по сантиметровым ступеням толщины. Проводили подразделение P. sylvestris по классам Г. Крафта (I – господствующие, II – согосподствующие, III – подчиненные, IV – угнетенные), с определением причин усыхания и повреждения. Для трех центральных ступеней толщины по каждой породе измеряли ширину кроны (при помощи мерной ленты), массу сучков, массу хвои и массу ствола (методом взвешивания), прирост в высоту по годам с 1980 по 2004 год. Живой напочвенный покров учитывали на всех площадях: глазомерно определяли общую степень покрытия и преобладающие виды по биомассе. Вычисляли плотность сложения опада по измеренной толщине и массе подстилки. На пробной площади № 5 где росли четыре ряда P. sylvestris с чередованием с двумя рядами березы повислой (Betula pendula Roth) и наибольшее число стволов P. sylvestris (24 шт.) относились к первым двум классам Крафта, тогда как особи B. pendula – к 4 классу. 255 Распределение деревьев по классам Г. Крафта при разных типах смешения, % Классы Крафта №П П Виды деревьев Состав посадки 1 2 3 4 5 Итого производства 1978 г. 1 P. sylvestris 6P. sylvestris+ 2S. acutifolia 2 P. sylvestris 5P. sylvestris+ 2S. acutifolia 23,8 35,2 33 5,7 2,3 100 3 P. sylvestris 1P. sylvestris +1S. acutifolia 7,6 18,2 42,4 15,2 16,6 100 18,9 29,4 25,9 18,9 6,9 100 18,5 18,5 22,3 29,6 11,1 100 10 17,5 25 22,5 25 100 – – – 100 – 100 22,9 28,6 31,4 14,2 2,9 100 4 5 P. sylvestris B. pendula B. pendula B. pendula 6P. sylvestris +2B. pendula 4P. sylvestris +2B. pendula 18,9 26,2 28,9 21,7 4,3 100 производства 1979 г. 5 P. sylvestris B. pendula 2P. sylvestris+ 1B. pendula 6 P. sylvestris 6P. sylvestris +2S. acutifolia 7 P. sylvestris 4P. sylvestris +2S. acutifolia 8 4 2 P. sylvestris B. pendula P. sylvestris B. pendula P. sylvestris B. pendula P. sylvestris 1 3 B. pendula P. sylvestris B. pendula 5 – 15 30 50 100 21,1 21,1 23,7 28,9 5,2 100 24 18 28 12 18 100 1P. sylvestris+ 1B. pendula 18 22 26 22 12 100 – 14,8 18,5 22,3 44,4 100 10P. sylvestris +2B. pendula 5,4 9,5 27 28,4 29,7 100 4,2 8,3 4,2 29,2 54,1 100 10P. sylvestris+ 3B. pendula 15,2 11,9 25,4 6,8 40,7 100 7,1 21,4 – 28,6 42,9 100 10P. sylvestris +1S. acutifolia 1B. pendula +1S. acutifolia 4,5 5,9 14,9 34,3 40,3 100 – 6,3 – 43,7 50 100 5P. sylvestris+ 2B. pendula 8,8 25 22,1 29,4 14,7 100 – 3,7 14,8 37,1 44,4 100 В результате исследования отмечена зависимость увеличения количества опада в сосняках лишайниково-мшистых с повышением их пол256 ноты. Так, при наименьшей полноте (0,64) и совместном произрастании P. sylvestris и B. pendula в посадках 1978 г. было отмечено 15,02 т/га опада, а при большей полноте (0,76) – 15,8 т/га. Чем больше запас хвороста в напочвенном покрове, тем скорость пожара выше, однако нередко достаточно небольшого количества сильно высохших листьев и веток, чтобы выделяемая ими тепловая энергия создала условия, благоприятствующие образованию типа конвекционной колонки. Пожары сильной интенсивности наблюдаются в тех случаях, когда располагаются толстым и рыхлым слоем и тем более в густых хвойных насаждениях, где в горении могут участвовать и кроны деревьев. Л и т е р а т ур а Анучин Н. П. Лесная таксация / Н. П. Анучин. – М.: Наука, 1982. – 552 с. Антонова Г. Ф. Влияние условий произрастаний на структуру годичного слоя древесины и продуктивность сосны обыкновенной / Г. Ф. Антонова, В. Д. Перевозникова, В. В. Стасова // Лесоведение. – 1999. – № 6. – С. 45–53. Демаков Ю. П. Рост и изреживание древостоев сосны / Ю. П. Демаков. – Йошкар-Ола: МарГТУ, 1998. – С. 36–38. Дитрих В. И. Теоретические основы геометрии режущих устройств сучкорезных машин / В. И. Дитрих // Лесная промышленность. – 2005. – № 3. – С. 28–29. Кроткевич П. Г. Выращивание высококачественной древесины / П. Г. Кроткевич. – М.: Гослесбумиздат, 1955. – 179 с. Успенский Е. И. Лесная пирология / Е. И. Успенский. – Йошкар-Ола: МарГТУ, 1992. – 7 с. Митрофанов С. В. Рост и продуктивность молодняков сосны обыкновенной / С. В. Митрофанов, В. Л. Кузнецов // Лесное хозяйство. – 2004. – № 4. – С. 39–40. Писаренко А. И. Искусственные леса: в 2 ч. / А. И. Писаренко, Г. И. Редько, М. Д. Мерзленко. – М.: ВНИИЦлесресурс, 1992. – Ч. 2. – 240 с. Репринцев Д. Д. Механизация обрезки крон деревьев и кустарников / Д. Д. Репринцев, М. В. Драналюк, В. П. Попиков // Лесное хозяйство. – 2006. – № 1. – 45 с. БИОРАЗНООБРАЗИЕ: КОЛИЧЕСТВЕННЫЙ И КАЧЕСТВЕННЫЙ ПОДХОД Злобин Ю. А. Сумский национальный аграрный университет, г. Сумы, yuz-32@mail.ru Проблема изучения и сохранения биоразнообразия имеет длительную историю. Еще в 1988 г. по инициативе Международного союза биологических наук (IUBS) была принята резолюция о необходимости изучения роли и значения биоты в регуляции многих природных процессов. В 1991 г. Генеральная ассамблея Международного союза биологических наук принимает программу изучения биоразнообразия (Diversitas), а в 1992 году международным сообществом была принята Конвенция о биоразнообразии. Эти международные соглашения активизировали исследовательские работы по проблеме биоразнообразия: за 257 прошедшие три десятилетия ей посвящены многие десятки монографий, сотни и сотни статей. При мониторинге биологического разнообразия обычно используют три его категории (α-, β- и γ-разнообразие), которые были выделены Р. Уиттекером (1980) и ориентированы на установление видового богатства. Помимо этого многие исследователи выделяли в качестве единиц для учета биоразнообразия не только виды растений или животных, но и различные природные структуры от генов и макромолекул до ландшафтов и природных зон (Rands et al., 2010). В итоге учение о биоразнообразии становилось все более аморфным и неопределенным: оно стало сводиться либо только к таксономическому разнообразию, либо охватывать весь набор структур и явлений природной среды планеты, а не только живые организмы (Raven, 2011). Это привело к тому, что пока не установлена вся палитра разнообразия жизни, не выявлены ключевые процессы, определяющие устойчивость биоразнообразия на планете, не раскрыты критические для сохранения живых организмов и биоценозов звенья в иерархии биоразнообразия, поэтому качественные и количественные аспекты биоразнообразия и методы его регистрации требуют дальнейшего переосмысливания и критического анализа. Если рассматривать биоразнообразие именно как разнообразие живых организмов и их сочетаний (а это достаточно логичный подход), то базовыми единицами биоразнообразия должны быть приняты особи, индивидуумы и их закономерные устойчивые сочетания – биоценозы. Такой подход был сформулирован Б. А. Юрцевым еще в 1992 г. словами «биологическое разнообразие – разнообразие организмов и их природных сочетаний», но он не получил дальнейшего развития. Качественные и количественные отличия между особями, таким образом, являются исходной формой биоразнообразия – i-биоразнообразием, а соответствующие отличия между биоценозами должны рассматриваться как b-биоразнообразие. На основе этих двух базовых единиц биоразнообразия строится его иерархия. В ходе анализа разнообразия особей (i-биоразнообразие) по определенному набору признаков идентифицируется их видовая принадлежность, т. е. определяется таксономиический статус, что ведет к новой категории биоразнообразия: видовому, т. е. s-биоразнообразию, которое составляет первое звено в иерархии таксономического разнообразия: вид – род – семейство – порядок – класс – отдел. В настоящее время изучение видового биоразнообразия как части таксономического разнообразия является наиболее «модным» направлением. 258 Видовое разнообразие принято оценивать с помощью двух показателей. Собственно видовое богатство выражают числом видов растений, приходящимся на единицу площади. Равномерность распределения видов по обилию называют выравненностью, для вычисления которой предложено более 40 различных индексов (Лебедева, Криволуцкий, 2002). Один из наиболее эффективных индексов был предложен еще , где Р. МакИнтошем (McIntosh, 1967) и имеет вид S – число видов, ni – число особей i-го вида. Обширная литература по изучению видового биоразнообразия позволяет сделать заключение о ряде общих закономерностей его проявления, в частности, видовое разнообразие выше в среде, содержащей больше природных ресурсов, и в экотонах, существует некоторый оптимальный уровень биоразнообразия, при котором устойчивость биоценозов максимальна, во всех биомах земного шара в настоящее время наблюдается тенденция к снижению уровня биоразнообразия. Одновременно сопоставительный анализ литературных данных выявляет большие нестыковки в выводах разных исследователей. Во многом это связано с субъективностью таксономических решений. При классификации растений могут использоваться линнеоны – виды в их «крупном» понимании, а в других случаях – жорданоны, т. е. мелкие виды, на которые подразделен более крупный линнеевский вид. Субъективность таксономических решений охватывает и семейства, которые понимаются либо в крупном объеме, либо подразделяют некогда крупное семейство на несколько мелких. Качество исходного материала, конечно, зависит и от флористической подготовки исследователя. А. М. Федотов и др. (1998, с. 74) справедливо отмечали, что «в настоящее время квалифицированное определение таксономической принадлежности для большинства видов является почти искусством, доступным, как правило, специалистам по сравнительно узким таксономическим группам». Анализ геоботанических описаний показывает, что они далеко не всегда полные. Часто пропускаются редкие виды, виды, представленные в фитоценозе всходами или ювенильными формами, а также виды, развитие которых имеет сезонный характер. Биоразнообразие не сводится к таксономическому. Для понимания его генезиса необходим анализ так называемого типологического разнообразия, иерархия которого составляет ряд: жизненная форма – тип флорогенеза – гемеробность – эколого-фитоценотическая стратегия – функциональный тип. Типологическое разнообразие в мире растений пока остается изученным гораздо хуже, чем таксономическое, хотя оно в большей степени раскрывает механизмы утраты общего биоразнообразия, снижение которого характерно для современной эпохи. Количе259 ственные критерии для оценки этой категории биоразнообразия мало разработаны. Виды растений в растительном покрове существуют в форме их популяций – совокупностей особей одного вида, занимающих определенную территорию и объединенных системообразующими связями. Популяционная структура таксономического вида является особой формой биоразнообразия. Предметом исследования здесь является количество локальных популяций, которыми представлен тот или иной вид растения, численность особей в них, популяционная плотность, конфигурация и размеры популяционных полей. Важную информацию, особенно при изучении редких видов растений, дает анализ динамики популяций – их устойчивость, прогрессивное расширение или, напротив, деградация (Злобин и др., 2013). Самостоятельный типологический ряд в иерархии биоразнообразия составляет внутрипопуляционное разнообразие (Злобин, 2010). Основными звеньями в иерархии этой категории являются: гендерная структура популяции – возрастная структура (по календарному возрасту) – онтогенетическая структура – виталитетная структура. Эти типы внутрипопуляционной структуры оценивают по соотношению в популяции особей разных категорий соответственно пола, возраста, онтогенетического или виталитетного состояния, выражая результаты либо в форме диаграмм-спектров, либо в виде интегральной оценки специальными индексами (Жукова, 1995; Злобин, 2009; Hayek, 2010). Смещение интересов исследователей на анализ таксономического биоразнообразия с констатацией фактов его снижения в определенной степени бьет мимо цели: оно не раскрывает механизмов утраты биоразнообразия. Высказываются лишь общие соображения о факторах, провоцирующих такую утрату. Это в основном усиливающийся антропогенез и глобальное потепление климата (Heller et al., 2009). С точки зрения сохранения биоразнообразия на планете на первый план должны быть выдвинуто изучение популяционной структуры видов и внутрипопуляционной структуры локальных популяций, которое позволит устанавливать генезис популяций растений и разрабатывать меры по управлению им. Именно популяционный уровень в организации жизни является критическим и наиболее уязвимым при изменении условий существования популяций как растений, так и животных. Как было отмечено выше, самостоятельной формой биоразнообразия является биоразнообразие биоценозов. На сегодня b-биоразнообразие изучено гораздо меньше, чем таксономическое биоразнообразие. Основное внимание здесь пока уделяется иерархии синтаксонов, которая в разных системах классификации растительности неодинакова. 260 Редкие синтаксоны характеризуются в Зеленых книгах, обычные – в текущих геоботанических публикациях. Сопоставительный анализ литературных данных здесь иногда обескураживающий. Например, для национального природного парка ДеснянскоСтарогутский С. М. Панченко (2013) выделено по системе Браун-Бланке 9 классов, 11 порядков, 13 союзов, 22 ассоциации растительности. Кроме того, в рамках двух ассоциаций выделено по 3 субассоциации. Для этой же территории по доминантной классификации установлено наличие 3 типов растительности, 7 классов формаций, 11 групп формаций, 23 формации, 35 субформаций, 104 групп ассоциаций, 242 ассоциации. В обеих системах классификации первичной синтаксономической единицей является ассоциация. Очевидно, что второй способ классификации создает впечатление о более выраженном фитоценотическом биоразнообразии территории по сравнению с первым. Еще сложнее интерпретировать литературные данные по структурно-функциональному биоразнообразию биоценозов. Понимая биологическое разнообразие как разнообразие живых организмов и их устойчивых сочетаний – биоценозов, следует учитывать органическую связь этого типа разнообразия с экологическим разнообразием, базовой единицей которого являются экосистемы. На основе экосистемного разнообразия складывается своя достаточно сложная иерархия экологического разнообразия. В целом, иерархия категорий биологического и экологического разнообразия континуальна (Злобин, 1996), но этот континуум не аморфен, он структурирован. Исследования биологического и экологического разнообразия могут быть продуктивными и сопоставимыми только при четкой приуроченности работ к определенным категориям природного разнообразия и выполняться по унифицированным методикам. Л и т е р а т ур а Жукова Л. А. Популяционная жизнь луговых растений / Л. А. Жукова. – Йошкар-Ола: Ланар, 1995. – 224 с. Злобин Ю. А. Континуум категорий биологического разнообразия / Ю. А. Злобин // Доповiдi Нац. АН Украïни, 1996. – № 6. – С. 152–154. Злобин Ю. А. Популяционная экология растений: современное состояние, точки роста / Ю. А. Злобин. – Сумы: Университетская книга, 2009. – 263 с. Злобин Ю. А. Биоразнообразие фитопопуляций – основа устойчивого существования видов растений и растительных сообществ / Ю. А. Злобин // Вiсн. СНАУ. – 2010. – Вип. 4. – С. 3–10. Злобин Ю. А. Популяции редких видов растений: теоретические основы и методика изучения / Ю. А. Злобин, В. Г. Скляр, А. А. Клименко. – Сумы: Универитетская книга, 2013. – 439 с. Лебедева Д. А. Биологическое разнообразие и методы его оценки / Д. А. Лебедева, Д. А. Криволуцкий // География и мониторинг биоразнообразия: сб. – М.: НУМЦ, 2002. – С. 9–75. Панченко С. М. Лесная растительность Национального природного парка «Деснянско-Старогутский» / С. М. Панченко. – Сумы: Университетская книга, 2013. – 213 с. Уиттекер Р. Сообщества и экосистемы / Р. Уиттекер. – М.: Прогресс, 1980. – 328 с. Федотов А. М. Электронный атлас «Биоразнообразие растительного мира Сибири» / А. М. Федотов, Б. Я. Рябко, 261 А. А. Федотов и др. // Вычисл. техника, 1998. – Т. 3, № 5. – С. 68–78. Юрцев Б. А. Экологогеографическая структура биологического разнообразия и стратегия его учета и охраны / Б. А. Юрцев // Биол. разнообразие: подходы к изучению и сохранению: сб. – СПб.: ЗИН РАН, 1992. – С. 7–21. Hayek L. A. Surveying natural population. Quantitative tools for assessing biodiversity / L. A. Hayek, M. A. Bazzas. – N. Y.: Columbia Univ. Press, 2010. – 592 p. Heller N. E. Biodiversity management in the face of climate change: a review of 22 years of recommendations / N. E. Heller, E. S. Zavaleta // Biol. Conserv. – 2009. –Vol. 142. – P. 14– 32. McIntosh R. An index of diversity and the relation of certain concepts to diversity / R. McIntosh // Ecology. – 1967. – Vol. 48, № 3. – P. 392–404. Rands M. R. Biodiversity conservations: challenges beyond 2010 / M. R. Rands, W. Adams, L. Bennun et al. // Science. – 2010. – Vol. 329. – P. 1298–1303. Raven P. H. Introduction to special issue on biodiversity / P. H. Raven, J. Chase, J. C. Pires // Amer. J. Botany. – 2011. – Vol. 98, № 3. – P. 333–335. ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА СУБПОПУЛЯЦИЙ ЛИШАЙНИКА (LOBARIA PULMONARIA) В ЛЕСАХ СЕВЕРО-ВОСТОКА КОСТРОМСКОЙ ОБЛАСТИ Иванова Н. В. Институт математических проблем биологии Российской академии наук, г. Пущино, Natalya.dryomys@gmail.com Наши исследования посвящены изучению популяций редкого лесного лишайника (Lobaria pulmonaria (L.) Hoffm.). Согласно литературным данным, этот вид встречается в различных по экологическим условиям сообществах, но длительное существование его популяций возможно только в старовозрастных малонарушенных лесах. Это связано с требовательностью L. pulmonaria к микроклиматическим условиям среды, а также с низкой способностью к колонизации соседних территорий (Пыстина, Семенова, 2004, Ockinger et al., 2005). Известно, что зачатки, способные перемещаться на большие расстояния (аскоспоры), образуются редко и только на старых талломах лобарии (Scheidegger et al., 1998). Основываясь на этих данных, мы предположили, что в лесах, образовавшихся после нарушений (сплошных рубок и пожаров), и в лесах, длительное время не подвергавшихся антропогенным воздействиям, структура популяций L. pulmonaria будет различной. Для подтверждения этой гипотезы на северо-востоке Костромской области проведены полевые исследования субпопуляций L. pulmonaria в различных типах южнотаежных лесов. В данной работе под субпопуляцией понимается совокупность талломов лобарии, населяющих один ствол дерева (Михайлова, 2008). Онтогенетическую структуру субпопуляций описывали по методике, изложенной ранее (Иванова, Терентьева, 2012), при этом фиксировалось наличие стадий развития в субпопуляциях лишайника без указания числа талломов. Для описания онтогене262 тической структуры использовалась семибалльная шкала (табл. 1). Типы леса определяли на основе геоботанических описаний по методике, предложенной Л. Г. Ханиной с соавторами (2002). Таблица 1 – Стадии развития L. pulmonaria 1 стадия Таллом цельный в виде чешуйки 2 стадия Таллом с лопастями, легочная складчатость не выражена 3 стадия Таллом имеет взрослый облик, сформированы лопасти и доли, выражена характерная «легочная» складчатость 4 стадия Наличие соредиев (органов вегетативного размножения) 5 стадия Наличие апотециев (органов полового размножения) 6 стадия Наличие некрозов, разрушение центральной части талломов, с сохранением связи между лопастями 7 стадия Отдельные отмирающие лопасти Для полевых исследований было выбрано два участка: первый – на территории Кологривского района (ГПЗ «Кологривский лес») в бассейне р. Сеха. На первом участке описана онтогенетическая структура 30 субпопуляций L. pulmonaria, на втором участке – 218. По лесотаксационным данным, лесной покров на обоих исследованных участках представлен в основном средневозрастными смешанными лесами из Betula sp., Populus tremula L., Picea abies (L.) Karst., Pinus sylvestris L., чаще всего с преобладанием березы в древостое. Эти сообщества образовались на месте сплошных рубок середины XX века, редко с последующим лесовосстановлением. Встречаются осиновые леса в виде небольших фрагментов на водораздельных участках; как правило, они располагаются на месте старых гарей. Фрагменты еловых лесов сохранились в основном в водоохранных зонах вдоль рек, а также как недорубы на труднодоступных водораздельных участках. В результате проведенного исследования выяснено, что в березовых лесах, которые наиболее распространены на данной территории, L. pulmonaria встречается редко и отмечена только в 6 % от всех случаев находок. Чаще же всего лобария встречалась в осиновых (59 % находок) и еловых (34 % находок) лесах. Однократно L. pulmonaria была встречена в сосняке и ивняке (из Salix caprea L.). В целом в исследованной нами выборке описаний субпопуляций отмечено наличие всех онтогенетических состояний L. pulmonaria. Спектр представлен на рисунке. 263 Число деревьев 200 150 100 50 0 1 2 3 4 5 6 7 Стадии онтогенеза L. pulmonaria Онтогенетический спектр L. pulmonaria на исследованной территории Максимумы приходятся на 3-ю и 4-ю стадии онтогенеза, что соответствует еще не приступившим к вегетативному размножению талломам со сформированным взрослым обликом и хорошо выраженной «легочной» складчатостью (3 стадия) и вегетативно размножающимся (с помощью соредиев) талломам L. pulmonaria (4 стадия). Наличие апотециев у лобарии зафиксировано на всех исследованных участках. В спектрах субпопуляций отмечен «провал» на стадии 5 (наличие апотециев). Такой результат согласуется с литературными данными (Scheidegger et al., 1998). Обращает на себя внимание низкая встречаемость субпопуляций со старыми талломами (стадия 7). Видимо, это связано с тем, что L. pulmonaria относительно недавно заселила исследованные нами лесные сообщества. Это предположение подтверждается данными, полученными Н. А. Семеновой при исследовании онтогенетической структуры субпопуляций L. pulmonaria в старовозрастых еловых лесах Печоро-Илычского заповедника (Семенова, 2005). В полученных ею результатах доля старых талломов достигала 20 %, что свидетельствует о длительном существовании популяции на территории. Проведено сравнение онтогенетических спектров субпопуляций L. pulmonaria, обитающих в лесах, относящихся к разных формациям. Было выявлено, что, несмотря на существенно низкую встречаемость лобарии в березняках, чем в осинниках и ельниках, во всех лесах 264 структура субпопуляций лобарии была одинаковой (табл. 1). При этом онтогенетические спектры субпопуляций в лесах разных формаций совпадали по форме со спектром, построенным на основе всех анализируемых данных. Таблица 2 – Частота встречаемости онтогенетических состояний L. pulmonaria в лесах разных формаций Формация Онтогенетические состояния L. pulmonaria Число описаний 1 2 3 4 5 6 7 ельники 10 18 25 22 4 18 2 84 осинники 8 18 25 25 3 19 1 146 березняки 10 22 29 20 4 16 0 16 Важно отметить, что субпопуляции с апотециями встречались во всех лесах с одинаковой частотой 3–4 %. При этом в субпопуляциях, исследованных в малонарушенных лесах Печоро-Илычского заповедника, по данным Н. А. Семеновой (2005), доля талломов с апотециями также была низкой и не превышала 3 %. Таким образом, в ходе проведенного исследования мы выяснили, что доля субпопуляций L. pulmonaria с апотециями одинакова для малонарушенных лесов и средневозрастных лесов, образовавшихся после рубок и пожаров. Однако в средневозрастных лесах доля старых талломов меньше, чем в старовозрастных. Автор благодарит сотрудников ИФХиБПП РАН М. П. Шашкова и В. Н. Шанина, магистранта ПущГЕНИ Л. К. Рочеву за помощь в сборе полевого материала, а также сотрудника ГПЗ «Кологривский лес» Е. В. Терентьеву за помощь в организации полевых исследований. Работа выполнена при финансовой поддержке РФФИ (грант № 13-04-02181 А) Л и т е р а т ур а Иванова Н. В. Онтогенетическая структура субпопуляций редкого лишайника лобария легочная (Lobaria pulmonaria (L.) Hoffm.) В ядре заповедника «Kологривский лес» / Н. В. Иванова, Е. В. Терентьева // Естествознание в регионах: проблемы, поиски, решения: материалы межд. науч. конф. «Регионы в условиях неустойчивого развития» (КостромаШарья, 1–3 ноября 2012 г.): в 2 т. / сост. и отв. ред.: Ю. А. Дорогова, И. Г. Криницын, С. А. Кусманов, А. П. Липаев, К. С. Ситников. – Кострома: КГУ им. Н. А. Некрасова, 2012. – Т. 1. – С. 139–143. Михайлова И. Н. Анализ субпопуляционных структур эпифитных лишайников (на примере Lobaria pulmonaria (L.) Hoffm. / И. Н. Михайлова // Вестник Нижегородского университета им. Н. И. Лобачевского. Сер., Биология. – 2005. – № 1. – С. 124–134. Пыстина Т. Н. Некоторые аспекты изучения экологических особенностей лишайника Lobaria pulmonaria (L.) Hoffm. на европейском северо-востоке России / 265 Т. Н. Пыстина, Н. А. Семенова // Вестник Института биологии Коми НЦ УрО РАН. – 2004. – № 9 (83). – С. 4–9. Семенова Н. А. Структура и численность популяции Lobaria pulmonaria в девственных лесах Печоро-Илычского заповедника / Н. А. Семенова // Грибы в природных и антропогенных экосистемах: тр. междунар. конф., посв. 100-летию начала работы проф. А. С. Бондарцева в бот. ин-те им. В. Л. Комарова РАН (Санкт-Петербург, 24–28 апреля 2005 г.). – СПб., 2005. – Т. 2. – С. 190–192. Ханина Л. Г. Новый метод анализа лесной растительности с использованием многомерной статистики (на примере заповедника «Калужские засеки») / Л. Г. Ханина, В. Э. Смирнов, М. В. Бобровский // Бюл. МОИП. Отд. биол. – 2002. – Т. 107, № 1. – С. 40–48. Ockinger E. Is local distribution of the epiphytic lichen Lobaria pulmonaria limited by dispersal capacity or habitat quality / E. Ockinger, M. Niklasson, S. G. Nilsson // Biodiversity and conservation. – 2005. – Vol. 14, № 3. – Р. 759–773. Scheidegger Ch. Reintroduction and augmentation of populations of the endangered Lobaria pulmonaria: methods and concepts / Ch. Scheidegger, B. Frey, J-Cl. Walser // Lobarion lichens as indicators of the primeval forests of the Eastern Carpathians (Darwin International Workshop, 25–30 May 1998, Kostrino, Ukraine). – P. 33–52. СТРУКТУРА И ДИНАМИКА ПОПУЛЯЦИЙ НЕКОТОРЫХ КАЛЬЦЕФИТОВ СРЕДНЕЙ ВОЛГИ Ильина В. Н. Поволжская государственная социально-гуманитарная академия, г. Самара, 5iva@mail.ru Популяционно-онтогенетическое направление ботанических исследований завоевывает все большую популярность, играя весомую роль среди методов биоиндикации состояния природных комплексов и урбанизированной среды, выявления и мониторинга охраняемых территорий, в создании и ведении Красных книг, определении биоэкологических особенностей видов растений и др. Нами уже 15 летосуществляются исследования популяционной структуры и ее динамики большого числа раритетных представителей средневолжской флоры. Пристальное внимание нами уделяется растениям-кальцефитам – Hedysarum grandiflorum Pall., H. razoumovianum Fisch. еt Helm, H. gmelinii Ledeb., Oxytropis hippolyti Boriss., O. floribunda (Pall.) DC., O. spicata (Pall.) O. et B. Fedtsch., Laser trilobum (L.) Borkh., Linum flavum L., Onosma simplicissima L., Centaurea sumensis Kalen. и C. ruthenica Lam., Adonis vernalis L. и A. wolgensis Stev., Astera lpinus L., Jurinea arachnoidea Bunge и J. ledebourii Bunge и т. д. (Ильина, 2006– 2008, 2009а, 2009б; Ильина, Саксонов, 2010; Родионова, Ильина, 2013 и др.). Структура определена у 2350 популяций (46 представителей), у 215 популяций изучена многолетняя динамика онтогенетической, пространственной и виталитетной структуры. Модельные виды в основном имеют следующие общие особенности биологии и экологии: всходы появляются из семян весной и в начале 266 лета после прохождения естественной зимней стратификации в условиях наибольшей влажности почвы; проростки подвержены катастрофической элиминации; выжившие молодые растения быстро проходят начальные стадии развития, активно наращивая биомассу; при наступлении длительной засухи генеративные растения могут переходить в стадию покоя, переживая неблагоприятные условия среды; воздействие высоких температур приводит к переходу растений в состояние субсенильное, ложное субсенильное или отмирающее; ядро популяции составляют генеративные, наиболее устойчивые и длительно существующие, особи; в основном проявляют патиентные черты; имеют весомый разрыв реальной и потенциальной семенной продуктивности; обладают гетерогенностью онтогенетической и вителитетной структуры популяций; чувствительны к антропогенной нагрузке на местообитания (Ильина, 2011, 2012; Ильина, Горлов, 2011; Ильина, Дорогова, 2012; Родионова, Ильина, 2013 и др.). В популяциях Hedysarum grandiflorum, H. razoumovianum, H. gmelinii, Oxytropis hippolyti, Onosma simplicissima, Centaurea sumensis и C. ruthenica преобладают особи средневозрастного генеративного состояния; Oxytropis floribunda, O. spicata, Linum flavum, Onosma simplicissima, Adonis vernalis, A. wolgensis, Asteralpinus – старые генеративные особи, Laser trilobum, Jurinea arachnoidea и J. Ledebourii – виргинильные растения. В пространственной структуре отмечаются крупные агрегации особей. В скопления входит от 30 % особей (Centaurea sumensis, Linum flavum, Astera lpinus) до 80 % экземпляров (Oxytropis floribunda, O. spicata, H. gmelinii). Распределение по фракциям внутри скоплений в оптимальных условиях местообитаний выглядит следующим образом – в них преобладают растения зрелого генеративного состоянии, остальные фракции хоть и уступают, но незначительно; можно сказать, что основные онтогенетические группы представлены равномерно (примерно в 25 % изученных популяций). При ухудшении условий существования соотношение особей в скоплениях, как и во всей популяции, свидетельствует о ее старении. Виталитетная структура популяций большинства модельных видов гетерогенна. В оптимальных условиях закономерно преобладают особи высокого (первого) уровня жизненности. При возрастающем антропогенном прессе характерны депрессивный и равновесный виталитетные типы ценопопуляций, что также служит подтверждением патиентного типа жизненной стратегии видов. Полученные нами обширные оригинальные данные по биологии и экологии модельных представителей, структуре и динамике их попу267 ляций свидетельствуют о негативном влиянии разных типов хозяйственной эксплуатации на растительный покров в бассейне Средней Волги (Ильина, 2006; 2007; 2009 а,б и др.). Большинство обследованных популяций видов, занесенных в Красную книгу Самарской области и РФ, находятся в угнетенном состоянии, численность особей в них неуклонно снижается. Л и т е р а т ур а Ильина В. Н. Эколого-биологические особенности и структура ценопопуляций редких видов рода Hedysarum L. в условиях бассейна Средней Волги: дис. … канд. биол. наук / В. Н. Ильина. – Самара, 2006. – 253 с. Ильина В. Н. Структура популяций Hedysarum gmelinii Ledeb. на западной границе и в центральной части ареала / В. Н. Ильина // Известия Самарского НЦ РАН. – 2007. – Т. 9, № 1. – С. 153–157. Ильина В. Н. Мониторинг ценотических популяций растений: учеб. пособие / В. Н. Ильина. – Самара: Изд-во СГПУ, 2008. – 92 с. Ильина В. Н. Некоторые результаты исследований ценопопуляций Anemone sylvestries L. (Ranunculaceae) в Самарском Заволжье / В. Н. Ильина // Самарская Лука: проблемы региональной и глобальной экологии. Самарская Лука. – 2009а. – Т. 18, № 4. – С. 159–170. Ильина В. Н. О сохранности фиторазнообразия степей Самарского Высокого Заволжья (на примере Кондурчинских яров) / В. Н. Ильина // Бюл. МОИП. Отд. биол. – 2009б. – Т. 114, вып. 3. – Приложение 1. Часть 1. Экология. Природные ресурсы. Рациональное природопользование. Охрана окружающей среды. – М., 2009. – С. 361–366. Ильина В. Н. Некоторые результаты ординации сообществ с участием Hedysarum gmelinii Ledeb. с использованием экологических шкал Д. Н. Цыганова / В. Н. Ильина // Проблемы современной биологии: Материалы II Международ. научно-практ. конф. (19.10.11). – М.: Спутник+, 2011. – С. 47–51. Ильина В. Н. К вопросу о систематическом положении копеечников Средней Волги / В. Н. Ильина // Международный сборник научных трудов, посв. году Германии в России «Естественные и гуманитарные науки – устойчивому развитию общества». – М., 2012. С. 63–65. Ильина В. Н. Экологическая пластичность флоры урочища «Лысая гора» (Студеный овраг, Красноглинский район г. о. Самара) / В. Н. Ильина // Научный диалог. – 2013. – № 3(15): Естествознание. Экология. Науки о земле. – С. 43–56. Ильина В. Н. К вопросу об онтогенезе и онтогенетической структуре ценопопуляций Jurinea arachnoidea Bunge / В. Н. Ильина, С. Е. Горлов // Известия Самарского НЦ РАН. – 2011. – Т. 13, № 5. – С. 71–74. Ильина В. Н. О положении ценопопуляций копеечника Гмелина (Hedysarum gmelinii Ledeb.) в экологическом пространстве (в условиях бассейна Средней Волги) / В. Н. Ильина, Ю. А. Дорогова // Известия Самарского научного центра Российской академии наук. – 2012. – Т. 14, № 1(7). – С. 1745–1749. Ильина В. Н. Некоторые итоги изучения ценопопуляций адонисов весеннего и волжского (Adonis vernalis L. и A. wolgensis Stev.) в бассейне Средней Волги / В. Н. Ильина, С. В. Саксонов // Бюлл. Главного ботанического сада. – 2010. – Вып. 196. – С. 107–116. Родионова Г. Н. Популяционные стратегии жизни избранных полукустарничков сем. Бобовые (Fabaceae) в условиях антропогенного пресса / Г. Н. Родионова, В. Н. Ильина // Известия Самарского научного центра РАН. – 2013. – Т. 15, № 3 (2). – С. 776–778. 268 ПОПУЛЯЦИОННОЕ БИОРАЗНООБРАЗИЕ И ОЦЕНКА СОСТОЯНИЯ ПОПУЛЯЦИЙ ЮЖНОСИБИРСКИХ ВИДОВ РОДА HEDYSARUM L. (FABACEAE) Карнаухова Н. А. Центральный сибирский ботанический сад СО РАН, г. Новосибирск, karnaukhova-nina@rambler.ru В настоящее время для разработки теоретических основ экологического мониторинга, оценки состояния возобновляемых биологических ресурсов необходимы более глубокие знания о популяционной жизни растений. Это невозможно без изучения популяционно-онтогенетического аспекта неоднородности популяций растений. Поэтому на первый план выдвигается выделение в популяциях онтогенетических групп как совокупности особей (или рамет вегетативного происхождения) одного биологического возраста. Это в свою очередь требует детального изучения онтогенеза растений разных биоморф различной систематической принадлежности (Жукова, 2001). Объекты наших исследований – виды рода Hedysarum L. В составе флоры Южной Сибири мы изучали 16 видов, относящихся к разным жизненным формам. 1. Ксерогенные кустарники (секция Fruticosa B. Fedtsch.) – факультативно корневищно-стержнекорневой ксерогенный кустарник Hedysarum fruticosum Pallas. 2. Травянистые многолетники. 2.1. Стержнекорневые поликарпики. 2.1.1. Длиннопобеговые (секция Gamotion Basin.): а) моноцентрические длинностержнекорневые каудексовые многоглавые базисимподиальные травянистые поликарпики с монокарпическими побегами удлиненного типа – Hedysarum alpinum L., H. austrosibiricum B. Fetsch., H. consanguineum DC., H. neglectum Ledeb., H. theinum Krasnob.; б) полицентрические длиннокорневищно-стержнекорневые базисимподиальные травянистые поликарпики с монокарпическими побегами удлиненного типа -– H. consanguineum, H. inundatum Turcz. 2.1.2. Полурозеточные: а) моноцентрические стержнекорневые каудексовые многоглавые базисимподиальные травянистые поликарпики с монокарпическими побегами полурозеточного типа – Hedysarum dahuricum Turcz. ex B. Fedtsch., H. minussinense B. Fedtsch., H. sangilense Krasnob. et Timoch.; б) факультативно корневищно-стержнекорневые травянистые поликарпики с монокарпическими побегами полурозеточного типа – Hedysarum gmelinii Ledeb., H. tschuense A. I. Pjak et A. L. Ebel. 269 2.1.3. Розеточные: а) моноцентрические стержнекорневые каудексовые многоглавые травянистые многолетники с поликарпическими побегами розеточного типа – Hedysarum chaiyrakanicum Kurbatsky, H. setigerum Turcz. ex Fischer et Meyer, H. turczaninovii Peschkova; б) розеточные подушковидные стержнекорневые многоглавые каудексовые поликарпики с каудикулами – Hedysarum zundukii Peschkova. Изменение жизненной формы и сочетание разных проявлений поливариантности на протяжении онтогенеза особей в естественных условиях произрастания обеспечивают формирование разных путей индивидуального развития, от чего зависит жизненное состояние, проявляющееся в мощности особей и различных перспективах их развития. Одной из главнейших диагностических характеристик популяционного уровня в оценке общего состояния популяций и их критического состояния (или близкого к критическому) является жизненное состояние популяций. При определении реального оптимума организма и популяций видов рода Hedysarum в различных условиях Южной Сибири мы используем метод Л. Б. Заугольновой (1994), основанный на определении как популяционных, так и организменных параметров (Карнаухова, Селютина, 2013). Ценопопуляции ксерогенного кустарника H. fruticosum образованы корневищно-стержнекорневыми растениями за счет корневищ, образующихся из погребенных пазушных почек. Первичный куст и главный корень у таких особей сохраняются, дальнейшее засыпание субстратом или понижение уровня субстрата в конце концов приводит к гибели сначала первичного куста, а затем и неукоренившихся парциальных кустов. Изучение онтогенетической структуры показало, что в ЦП на подвижном песчаном субстрате молодая фракция более многочисленная (53,8 %), чем на более закрепленном субстрате (42,9 % на железнодорожной насыпи), где к тому же отсутствуют ювенильные особи. Онтогенетический спектр в любом случае бимодальный – характерный для этого вида. Характерный спектр моноцентрических поликарпиков из секции Gamotion – центрированный. Но в зависимости от эколого-фитоценотических условий произрастания и антропогенного воздействия тип онтогенетических спектров изменяется. В благоприятных условиях для прорастания семян при хорошем семенном возобновлении происходит накопление молодых особей. Резкие колебания числа всходов и их массовая гибель в высокогорных условиях приводят к возникновению в популяциях волны возобновления: перемещению максимума с ювенильных на другие группы молодых особей. Кроме колебаний в поступлении ювенильных растений, в ЦП часто происходит смещение основ270 ного максимума и формирование бимодальных спектров. Наибольшее число вариантов онтогенетических спектров, обладающих сходством в положении подъемов (g1 и g3), определяют модальный (базовый) спектр этих видов. В отличие от моноцентрических видов, все исследованные ценопопуляции видов секции Gamotion c корневищно-стержнекорневой жизненной формой относятся к молодым. Характерный и базовый спектры полицентрических растений однотипны, так как основной пик образован виргинильными особями вегетативного происхождения. Для онтогенетических спектров моноцентрических полурозеточных и розеточных травянистых многолетников сохраняются те же тенденции, что и для моноцентрических поликарпиков секции Gamotion. Тенденция к уменьшению общей длины «главной оси» моноподиальных и годичных побегов в одном случае способствовала расширению экологической амплитуды и широкому распространению полиморфного вида секции Multicaulia H. gmelinii; в другом – изменение побега в сторону уменьшения укороченной части «главной оси» годичного побега дало ряд преимуществ в приспособлении к крайним условиям обитания в каменистых степях или высокогорных условиях, в более аридных восточных районах. Для ценопопуляций моноцентрических видов в типичных условиях произрастания с достаточным увлажнением характерен центрированный онтогенетический спектр. Достаточная численность молодых растений в этих ценопопуляциях обусловлена хорошим семенным возобновлением. Пик на группе генеративных растений в спектрах определяется значительной длительностью жизни особей в генеративном состоянии. Левосторонность спектров вторичных ложномолодых ценопопуляций (Уранов, Смирнова, 1969) в условиях длительного выпаса определяется хорошим семенным возобновлением копеечников, уменьшением плотности генеративных растений (ускоренное старение) и быстрым выпадением старых особей. Бимодальность и неполночленность онтогенетических спектров видов секций Multicaulia и Subacaulia Boiss. связана с засушливыми условиями, которые препятствуют осуществлению полного онтогенеза. В неполночленных бимодальных правосторонних спектрах ЦП на пастбищах часто отсутствуют особи прегенеративного периода, а основной максимум приходится на старые особи. Сравнительный анализ популяционных и организменных параметров в оценке общего состояния популяций моноцентрических видов свидетельствует об их устойчивом состоянии в природных ценозах Южной Сибири и успешном восстановлении после прекращения антропогенного воздействия. 271 Л и т е р а т ур а Заугольнова Л. Б. Структура популяций семенных растений и проблемы их мониторинга: дис. ... д-ра биол. наук / Л. Б. Заугольнова. – СПб., 1994. – 70 с. Жукова Л. А. Многообразие путей онтогенеза в популяциях растений / Л. А. Жукова // Экология. – 2001. – № 3. – С. 169–176. Карнаухова Н. А. Оценка состояния популяций Hedysarum theinum Krasnob. (Fabaceae) на Алтае / Н. А. Карнаухова, И. Ю. Селютина // Сибирский экологический журнал. – 2013. –№ 4 (6). – С. 543–550. Уранов А. А. Классификация и основные черты развития популяций многолетних растений / А. А. Уранов, О. В. Смирнова // Бюл. МОИП. Отд. биол. – 1969. – Т. 74 (2). – С. 119–134. ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ THYMUS MARSCHALLIANUS WILLD. Колегова Е. Б. Центральный сибирский ботанический сад Сибирского отделения Российской академии наук, г. Новосибирск, kolegova_e@mail.ru Исследование современного состояния ценотических популяций видов растений в настоящее время является одной из основных задач популяционной экологии растений. Особую значимость в данном направлении приобретают исследования хозяйственно-ценных и лекарственных видов растений. Одним из таких видов является Thymus marschallianus Willd. (тимьян Маршалла) – лекарственное растение, широко применяемое в официнальной и народной медицинах. Ареал вида простирается от Западной Европы до Алтая на востоке (Гогина, 1975). Типичным местообитанием являются богаторазнотравнодерновиннозлаковые растительные сообщества степной и лесостепной зон (Растительность…, 1980). Также для вида характерно произрастание в сообществах антропогенно нарушенных степей (насыпь, пашни, обочины дорог, свалки) и в экотонных сообществах. T. marschallianus – неподвижный полукустарничек с ветвящимися моно- и дициклическими генеративными побегами. Взрослое растение представляет собой первичный куст (или клон, состоящий из первичного куста и близко расположенных парциальных структур). Структура куста в основном представлена одно-, двулетними ортотропными побегами, комплексами на их основе и восходящими побегами формирования. В условиях горных степей на вершине холмов, а также в сообществах луговых степей по опушкам березовых и сосновых колков с высокой фитоценотической конкуренцией, восходящие побеги формирования в структуре куста T. marschallianus могут отсутствовать. Онтогенез T. marschallianus полный, сложный. Длительность онтогенеза может достигать 30 лет и зависит от длительности пребывания особей в средневозрастном генеративном состоянии (от 4 до 20 272 лет). Вегетативное размножение наступает в средневозрастном генеративном состоянии. Самоподдержание ценопопуляций осуществляется семенным путем (Колегова, Черемушкина, 2011). Изучение онтогенетической структуры ценопопуляций T. marschallianus проводили в 2010–2013 гг., на Южном Урале и в Западной Сибири. Всего исследовано 9 ценопопуляций (ЦП) по общепринятым методикам (Работнов, 1950; Ценопопуляции растений, 1976; 1988). Выявлено, что все ЦП T. marschallianus нормальные, неполночленные. Тип спектра, совпадающий с характерным центрированным, установлен для ЦП в разных растительных сообществах. Действие антропогенных факторов в луговых и настоящих степях приводит к изменению типа онтогенетического спектра (на левосторонний и бимодальный) и формированию молодых, переходных или старых ценопопуляций. Подобные изменения обратимы. Оценка состояния ЦП T. marschallianus показала, что реальный оптимум достигается в зрелых ЦП в условиях луговых степей на равнинных участках. Условия горных настоящих степей в целом для вида неблагоприятны. Здесь отмечается нерегулярное семенное возобновление, снижение жизнеспособности подроста, длительность средневозрастного генеративного состояния редко достигает 10 лет. Часто реальные оптимумы организма и популяции в таких условиях не совпадают, а значения соответствующих показателей низкие. Но в настоящей степи на равнинных участках между лесных колков, в затенении, где условия произрастания близкие к условиям в сообществах луговой степи, состояние ЦП может приближаться к оптимальному. Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта 12-04-100104-а. Л и т е р а т ур а Гогина Е. Е. Род чабрец (тимьян) – Thymus L. / Е. Е. Гогина // Биологическая флора Московской области. – М.: Изд-во Моск. ун-та, 1975. – Вып. 2. – С. 137–168. Колегова Е. Б. Онтогенез видов рода Thymus L. (Lamiaceae) и структура их ценопопуляций в Хакасии / Е. Б. Колегова, В. А. Черемушкина // Современные проблемы популяционной экологии, геоботаники, систематики и флористики: материалы международной научной конференции, посвященной 110-летию А. А. Уранова (г. Кострома, 31 октября – 3 ноября 2011 г.): в 2 т. / отв. ред.: Ю. А. Дорогова, Л. А Жукова, В. П. Лебедев. – Кострома: КГУ им. Н. А. Некрасова, 2011. – Т. 1. – С. 129–133. Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах / Т. А. Работнов // Тр. БИН АН СССР. Сер. 3, Геоботаника. – 1950. – Вып. 6. – С. 179–196. Растительность европейской части СССР / под ред. С. А Грибовой, Т. И. Исаченко, Е. М. Лавренко. – Л.: Наука, 1980. – 429 с. Ценопопуляция растений: Основные понятия и структура / О. В. Смирнова и др. – М.: Наука, 1976. – 217 с. Ценопопуляция растений: Очерки популяционной биологии / Л. Б. Заугольнова и др. – М: Наука, 1988. – 184 с. 273 ПОПУЛЯЦИОННАЯ МОРФОЛОГИЯ СПОРОФИТА BOTRYCHIACEAE NAKAI И ЕЕ ВЛИЯНИЕ НА СТУКТУРУ ПОПУЛЯЦИЙ Криницын И. Г.1, Зонтиков Д. Н., Лебедев В. П., Латышева Д. Ю., Разгуляева Н. А., Мастерова А. В. Костромской государственный университет им. Н. А. Некрасова, г. Кострома, 1 hek@rambler.ru В популяционно-онтогенетическом направлении, базирующемся на достижениях биоморфологии, особо выделяется популяционная биология папоротников. Н. И. Шориной и др. авторами изучены типичные папоротники. Взаимосвязь популяционной морфологии и структуры видов Botrychiaceae Nakai исследованы пока недостаточно. В связи с этим объектом нашего исследования по-прежнему остаются представители этого семейства. К настоящему времени нами описано 5 видов: Botrychium lunaria, B. multifidum, B. matricariifolium, B. lanceolatum и B. virginianum; в различных географических зонах и эколого-фитоценотических условиях При периодизации онтогенеза использовались традиционные для архегониат маркеры онтогенетических состояний: связь с гаметофитом; наличие спорофора; форма трофофора и спорофора; число и порядок ветвления придаточных корней. Выделено 3 периода и 10 онтогенетических состояний, выявлена поливариантность онтогенеза, влияющая на структуру популяций. П р е р е п р о д у к т и в н ы й п е р и о д включает проросток (P), ювенильные микотрофный (J1) и миксотрофный (J2), имматурный (Im), виргинильный (V) спорофиты. Р е п р о д у к т и в н ы й п е р и о д – молодой (Sp1), зрелый (Sp2) и стареющий (Sp3) спороносящие спорофиты. С е н и л ь н ы й п е р и о д – субсенильный (Ss) и сенильный (S) спорофиты. По классификации, разработанной Л. А. Жуковой, среди модусов преобразований, проявляющихся в поливариантности вегетативной и репродуктивной сфер всем изученным представителям семейсива характерны следующие: увеличение числа метамеров и их элементов, их редукция либо разрастание. Первый надтип (Жукова, 1995) включает в себя 3 типа. I тип – размерная поливариантность (жизненность). При понижении жизненности спорофор не развит и представлен рудиментом между филломофором и трофофором (Sp1-3V состояние). Подобные особи отмечены во всех популяциях, при этом все иные маркеры онтогенетического состояния сохраняются. В процессе онтогенеза особи способны изменять свою жизненность, что является важнейшим механизмом адаптации, функционирующим на организменном и популяционном уровнях. Особи низкой жизненности – резерв, сохраняющий территорию и переходя- 274 щий при изменении условий на более высокий уровень и играющий важную роль в поддержании генетического разнообразия популяций. II тип – собственно морфологическая поливариантность. Проявляется в основном в изменении трофофора. Дистальная часть перьев B. lunaria – цельнокрайняя или тупозубчатая. По этому признаку выделяются вариации – B. lunaria var. normale Roep. и B. lunaria var. subiniscum Roep., первая характерна слабосомкнутым сообществам, вторая – сообществам с более мощной растительностью. Встречаются особи, сочетающие эти признаки. Верхний непарный сегмент бывает квадратным, трапециевидным, треугольным, округлым, яйцевидным, надрезанным и рассеченным до середины и более, почти парным. Трофофоры B. lunaria, B. matricariifolium и B. lanceolatum дихотомически ветвятся, из их перьев могут формироваться трофофоры II порядка. У B. lunaria, B. matricariifolium и B. lanceolatum на нижних перьях трофофора иногда образуются спорангии, либо все перо превращается в подобие спорофора, из перьев материнской вайи формируется подобие дочерней, со спорофором и трофофором. У B. multifidum могут формироваться 2 спорофора на 1 вайе. Данная особенность подтверждает то, что стерилизация при возникновения вегетативных листьев играла важную роль. Подобные соотношения между категориями листьев у растений подтверждают превращение спорофиллов в трофофиллы. Корневища всех 5 видов могут развивать 2–3 листа, что приводит к ветвлению побега и ложному учету численности и описанию биоморфы. При этом корневища в почве располагаются горизонтально, т. е. перемещены в пространстве механическим путем. У B. lunaria, B. matricariifolium и B. lanceolatum ветвление придаточных корней как дихотомическое, так и боковое; у B. multifidum и B. virginianum исключительно боковое. III тип – поливариантность способов размножения. У всех 5 видов, в условиях центральной России, корневых отпрысков или особей вегетативного происхождения нами не обнаружено. Второй надтип. IV тип – ритмологическая поливариантность. Проявляется в сдвигах фенофаз у особей одной и соседних популяций. Сроки начала развития вайи и спороношения зависят от экологических факторов, жизненности и онтогенетического состояния спорофита. V тип – по темпам развития. Выражается в скорости индивидуального развития особей. Вжнейший параметр – продолжительность онтогенетических состояний. Большинство особей имеет нормальные темпы развития, но при высокой плотности, густом травостое и значительном задернении, наблюдается задержка в im, v и sp1-состояниях до 15–20 лет. Иногда наблюдаются случаи неотении, у B. lunaria, B. matricariifo275 lium, B. lanceolatum спорофоры и спорангии на перьях трофофора, формируются в пререпродуктивном периоде, отмечены Sр1-(j), Sр1-(im) и Sр1-(v) спорофиты. Наиболее характерный динамический процесс – временный покой. Обычно в нем пребывает 15–20% особей популяции, но в некоторые годы может перейти и вся популяция. Наиболее стабильная характеристика популяций – возрастные спектры. Почти все изученные популяции зрелые, полночленные с максимумом на Sp2-группе, исключение – 2 популяци B. lunaria, на известковых склонах (респ. Марий Эл), максимум спектра на Sp1-группе. Это явление вызвано следующими причинами: 1) неотения; 2) задержка развития. Анализ геоботанических описаний показал, что они обитают на карбонатных, бедных азотом почвах. Характерный возрастной спектр популяций определяется типом онтогенеза, а признаками, влияющими на реальные спектры, выступают поливариантность и особенности спорового размножения. Наибольшее количество особей репродуктивного периода приходится на Sp2-спорофиты. В годы с нормальной влажностью на их долю приходится до 50%. Несколько меньшее количество приходится на долю Sp1-растений, самая низкая численность – у Sp3. Численность в популяциях Botrychiaceae – Sp1-3V – спорофитов обратно пропорциональна Sp-группе. Она преимущественно пополняется Sp2-особями. В климатически благоприятные годы их численность составляет от 0 до нескольких процентов, в сухие – до 80%, их отсутствие скорее исключение, чем норма в жизни популяций, и свидетельствует о благоприятных условиях среды для развития спорофитов. Изучение динамики популяций показало, что главной чертой возрастного состава являются флюктуационные изменения: переход спорофитов в Sp1-3V-состояние и временный покой. Эти явления вызывают сложности при определении возрастного состава и численности популяций. В засушливые и следующие за ними годы максимум спектра смещается на Sp1-3V-группу, значительное количество особей переходит в состояние покоя. Анабиоз – одно из главнейших средств адаптации растений, спасающее от неблагоприятных воздействий. Продолжительность его – 1–2 года, но зафиксирован и 10-летний период. Он иногда совпадает с явлением реверсии. Наиболее малочисленная группа – спорофиты пререпродуктивного периода. Отсутствие их не говорит о неблагополучии вида в сообществе, т. к. они более подвержены элиминации и в отдельные годы могут, не формируя вайю, продолжать развиваться под землей. Соотношение встречающихся при раскопке J1 и учитываемых J2 показало, что основ- 276 ная их часть переходит в более взрослые состояния (Im и V), не развивая надземный лист или развивая его не каждый год. Таким образом, морфологическая неустойчивость, лабильность побеговых структур и неустойчивый характер ветвления корневой системы, расположение спорангиев как на спороносной, так и на вегетативной частях вайи свидетельствуют о своеобразии и древности таксона. Высокая морфологическая пластичность, лежащая в основе лабильности популяционных систем, позволила этой реликтовой группе сохраниться до наших дней. Погодичные изменения климатических факторов вызывают резкие колебания возрастного спектра: переход в состояние вторичного покоя, временного неспороношения, неотению. Популяции функционируют как «пульсирующие». Растения пререпродуктивного периода составляют незначительную часть популяции, т. к. ранние этапы онтогенеза проходят под землей и молодые спорофиты чаще переходят в анабиоз. Отсутствие их не свидетельствует о критическом состоянии популяций. На численность и положение максимума в возрастном спектре оказывают влияние экологические условия. Состав популяций Гроздовников, который можно описать, условен, т. к. основная часть особей обитает под землей (криптический характер), периодически появляясь на поверхности. АУТЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ПЛАСТИЧНОСТИ МОРФОМЕТРИЧЕСКИХ ПАРАМЕТРОВ В ПОПУЛЯЦИЯХ DACTYLORHIZA ROMANA (SEB. ET MAURI) SOO (ORCHIDACEAE) Кучер Е. Н., Чмелева С. И., Решетник Г. В. Таврический национальный университет им. В. И. Вернадского, г. Симферополь, evgenia.kucher@gmail.com, chmeleva@ukr.net, gvresh@ukr.net Крым является одним из наиболее крупных центров видовой насыщенности орхидных в Украине. Все виды семейства природной флоры полуострова занесены в Красную книгу Украины (Бордзiловський, 1950; Собко, Гапоненко, 1996). Редкость орхидей во многом обусловлена сложностью процесса репродукции. Обобщающим показателем процесса воспроизводства популяции служит репродуктивное усилие. Сравнительная оценка репродуктивного усилия при межпопуляционных сравнениях позволяет охарактеризовать самобытность той или иной фитопопуляции, которая может проявляться в особенностях изменчивости особей, степени скоррелированности параметров морфогенеза и пластичности (Злобин, 1996). Если изменчивость отображает неравноценность особей и разнообразие условий обитания внутри популяции, то пластичность характеризует их обратимые изменения при воздейст277 вии новых условий среды. Методом обнаружения пластичности служит сопоставление средних арифметических значений для растений из разных популяций (Злобин, 1989). Нами было проведено изучение особенностей пластичности метрических и аллометрических параметров у пальчатокоренника римского (Dactylorhiza romana (Seb. et Mauri) Soo). Исследования проводили в двух популяциях Dactylorhiza romana на территории Горного Крыма, расположенных на различной высоте над уровнем моря (растения, входящие в состав популяции 1, произрастают на высоте 800–850 метров над уровнем моря, популяции 2 – 350–400 м), а главное, склоны здесь разных экспозиций (популяция 1 – на югозападном, популяция 2 – на северо-восточном). Особи из популяций изымали в фазах цветения и плодоношения методом полной откопки и отмывки (Тарановская, 1957). Исследованы 17 метрических параметров. Параметры фитомассы: общая, запасающих органов, надземных органов, фотосинтезирующих органов, листьев, цветоноса, репродуктивных структур, брактей, всех цветков или плодов, одного цветка или плода. А также такие параметры, как высота цветоноса, длина соцветия, число листьев, цветков, плодов, семязачатков, семян, площадь фотосинтезирующей поверхности, листьев. В качестве аллометрических параметров оценивали: репродуктивное усилие, фотосинтетическое усилие и площадь фотосинтезирующей поверхности на единицу фитомассы. С целью сравнительной оценки скоррелированности морфологических структур использовали индекс морфологичеcкой интеграции (Злобин, 1989), который был рассчитан нами с учетом корреляционных матриц, включающих все исследованные метрические параметры. Сравнение средних значений всех изученных морфометрических параметров в двух популяциях сопровождалось определением достоверности разницы средних. Средние значения метрических параметров особи в популяции 2 превышают соответствующие значения в популяции 1 (в фазе цветения на 7,9–78,4 %, а в фазе плодоношения на 16,0– 194,8 %). Разница средних арифметических для всех параметров в высшей степени достоверна. Наименьшей пластичностью обладают такие показатели, как число листьев, длина цветоноса, фитомасса одного цветка и одного плода, эти параметры характеризуются наименьшими значениями коэффициента вариации. Резко повышается разница между популяциями в значениях линейных параметров надземной сферы в фазе плодоношения. Связано это с достижением растениями максимума своего размера и лучшей опыляемостью особей в популяции 2 (в среднем у одной особи из популяции 2 завязывается больше плодов, чем в популяции 1). Слабая изменчивость весовых параметров, отмеченная 278 для фитомассы одного цветка и одного плода как внутри отдельной популяции, так и между популяциями, позволяет утверждать, что у исследуемого вида орхидных весовые параметры цветка и плода достаточно жестко стабилизированы. Средние значения аллометрических параметров различаются не столь существенно: в фазе цветения – от 1,5 до 22,6 %, в фазе плодоношения – от 4,7 до 72,7 %. Причем, в отличие от метрических, во многих случаях средние значения аллометрических параметров в популяции 1 были выше, чем в популяции 2. Таким образом, при сравнении двух популяций, несмотря на большую общую мощность особей из популяции 2, относительные вклады в репродукцию и органы фотосинтеза изменяются в фазе цветения незначительно. Тогда как фотосинтетическое усилие в фазе цветения является одним из самых пластичных аллометрических параметров, вариабельность и среднее значение отношения площади фотосинтезирующей поверхности к общей фитомассе особи в двух популяциях почти не отличаются. Другими словами, площадь фотосинтезирующей поверхности, приходящаяся на единицу общей фитомассы, по сравнению с относительным вкладом фитомассы в органы фотосинтеза, довольно консервативна. Исследование аллокации фитомассы особей двух популяций в фазе цветения показало уменьшение вклада фитомассы в листья у особей из популяции 2 (относительный вес листьев особей из популяции 1 – 0,196 ± 0,006, из популяции 2 – 0,165 ± 0,009). Относительный вес цветоноса не изменяется (в популяции 1 – 0,116 ± 0,006, в популяции 2 – 0,116 ± 0,007), слабо изменяется и относительный вес подземных органов (в популяции 1 – 0,597 ± 0,011, в популяции 2 – 0,614 ± 0,019), а вклад в цветки и брактеи у особей из популяции 2 увеличивается (в популяции 1 – 0,090 ± 0,004, в популяции 2 – 0,105 ± 0,008). Данные об аллокации свидетельствуют о «перекачивании» в популяции 2 пластических веществ из листьев в генеративные органы. Наряду с этим, отношение числа семязачатков к площади фотосинтезирующей поверхности, обладающее наибольшей пластичностью среди аллометрических параметров, в популяции 2 снижается. В фазе плодоношения средние значения всех показателей репродуктивного усилия в популяции 2 превосходят соответствующие значения в популяции 1. Большее количество цветков, большие размеры цветоноса, соцветия и особей в целом повышают эффективность аттрактации опылителей, а следовательно, увеличивают значение репродуктивного усилия. Отличия между популяциями в средних значениях репродуктивного усилия, рассчитанных с использованием всех генеративных структур, меньше, 279 чем в средних значениях этих параметров, вычисленных с учетом только фитомассы цветков. Таким образом, сравнительный анализ популяций позволил обнаружить отличия по размеру особей и (в меньшей степени) относительному вкладу фитомассы в генеративные органы. Наименьшей пластичностью обладает в фазе цветения площадь фотосинтезирующей поверхности, приходящаяся на единицу фитомассы. Анализ пластичности репродуктивного усилия подтвердил буферную роль вспомогательных структур соцветия. Самобытность популяции 2 проявляется в наличии индивидуальной репродуктивной тактики. Повышенная репродуктивная способность обусловливается не увеличением количества цветков на единицу фитомассы, а повышением эффективности опыления. Последнему способствует перераспределение пластических веществ внутри растения в пользу возрастания фитомассы репродуктивных структур. Л и т е р а т ур а Злобин Ю. А. Принципы и методы изучения ценотических популяций растений: учеб.метод. пособие / Ю. А. Злобин. – Казань: Изд-во Казанского ун-та, 1989. – 146 с. Злобин Ю. А. Структура фитопопуляций / Ю. А. Злобин // Успехи совр. биол. – 1996. – Т. 116, вып. 2. – С. 133–146. Собко В. Г. Iнтродукцiя рiдксних i зникаючих рослин флори Украiни / В. Г. Собко, М. Б. Гапоненко – К.: Наук. думка, 1996. – 283 с. Тарановская М. П. Методы изучения корневых систем / М. П. Тарановская. – М.: Сельхозгиз, 1957. – 215 с. Бордзiловський Э. I. Родина Зозуленцевi / Э. I. Бордзiловський // Флора УРСР. – Киев: Вид-во АН УРСР, 1950. – Т. 3. – С. 312–405. АНТИМИКОТИЧЕСКИЕ СВОЙСТВА НЕКОТОРЫХ ВИДОВ ANTHEMIS L. ФЛОРЫ АЗЕРБАЙДЖАНА Мустафаева С. Д.1, Бахшалиева К. Ф.2 1 2 Отдел растительных ресурсов Института ботаники НАН Азербайджана, г. Баку Лаборатория биоконверсии растительных субстратов Института микробиологии НАН Азербайджана, г. Баку, Ин-т ботаники НАН Азербайджана, msitara@mail.ru Территория Азербайджанской республики обладает богатой флорой. Здесь распространено почти 5000 высших цветковых растений (Аскеров, 2011). Богатство флоры и разнообразие растительного покрова в Азербайджанской республике связаны с ее сложной историей, сформировавшейся под влиянием разнообразия ее физико-географических, природно-климатических и исторических условий. В мировой флоре семейство Астровых (Asteraceae Bercht. et J. Presl) – одно из самых крупных среди цветковых растений, включающее около 24000–30000 видов, объединенных в 1600–1700 родах (Funk V. A. и др., 2005). Виды этого семейства широко распространены на всех континентах и климатических зонах земного шара. 280 Семейство Asteraceae во флоре Азербайджана насчитывает 584 вида, объединенных в 125 родов (Аскеров, 2008; 2011), которые, в основном, широко используются в быту как пищевые, овощные, кормовые, эфирномасличные, лекарственные, технические, декоративные и пряноароматические растения. Одним из широко распространенных родов этого семейства является род Anthemis L. – пупавка, входящий в подсемейство Asteroideae Lindley и трибу Anthemideae Cass. Во флоре Азербайджана род Anthemis представлен 14 видами (Мустафаева, 2013). Среди полезных растений этого рода есть виды, известные как лекарственные, эфирномасличные, красильные и декоративные (Растительные ресурсы, 1995). В статье представлены результаты исследования антимикотических свойств двух видов рода Anthemis – A. cotula L. – пупавка вонючая и A. candidissima Willd. ex Spreng. – пупавка блестящая по отношению к микроскопическим грибам Trichoderma lignorum (Tode) Harz., Fusarium oxysporum Schlecht. и Aspergillus niger. A. cotula – однолетнее растение высотой 10–45(60) см. Распространена во всех ботанико-географических районах Азербайджана от низменности до среднего горного пояса как сорное растение на полях, в посевах, садах, огородах, лугах, лесных полянах, опушках, мусорных местах, по галечникам рек (Флора Азербайджана, 1961). Ксеромезофит. Относится к средиземноморскому классу ксерофильного географического типа. Согласно литературным источникам в составе A. cotula обнаружены эфирные масла, флавоноиды, полиацетиленовые и серосодержащие соединения, сесквитерпеноиды, стероиды, гетероциклические кислородсодержащие соединения (Растительные ресурсы, 1995). В народной медицине отвар, приготовленный из надземной части A. cotula, применяется при неврозах, гастралгии, проявляет гемостатическое, желчегонное, спазмолитическое, мочегонное, антигельминтное и противоопухолевое, а также инсектицидное действие (Мехтиева, 2011; Растительные ресурсы, 1995). A. candidissima – однолетнее растение высотой от 5 до 30 см. Встречается во всех ботанико-географичесиких районах Азербайджана от низменности до среднего горного пояса на сухих глинистых и щебнистых склонах, песчаных местах, по галечникам рек, иногда как сорное растение – в посевах (Флора Азербайджана, 1961). Ксерофит. Относится переднеазиатскому классу ксерофильного географического типа. По литературным данным (Растительные ресурсы, 1995), в составе A. candidissima обнаружены сесквитерпеноиды. В народной медицине этот вид применяется как гемостатическое и ранозаживляющее средство. 281 Для определения антифунгальной активности испытывались высушенные (воздушно-сухие) надземные части и водные экстракты исследуемых растений. В качестве штамм-культуры использованы Trichoderma lignorum, Fusarium oxysporum и Aspergillus niger, хранящиеся в музее Института микробиологии НАН Азербайджана. Определение антифунгальной активности (Методы…, 1982) проводили в 2 этапа: 1. Для определения роста грибов на твердой питательной среде воздушно-сухое сырье исследуемых растений измельчали до 0,5–1 см, затем увлажняли водопроводной водой до 55–60 %, при pH 6,5– 7. Увлажненный субстрат помещали в чашки Петри и стерилизовали в автоклаве при 1 атм. в течение 45 минут. После стерилизации, биомассу грибов в одинаковом количестве высевали в чашки Петри и выдерживали в термостате при температуре 25–27 оС. На 3, 5 и 7-й дни посева (в зависимости от роста) проводили учет выросших колоний грибов. В качестве контроля использовали шелуху пшеницы. Полученные результаты представлены на рисунках 1 и 2. Рисунок 1 – Рост колоний грибов на растениях Anthemis cotula Рисунок 2 – Рост колоний грибов на растениях Anthemis candidissima 282 Как видно из рисунков 1 и 2, максимальный рост всех грибов на растениях обоих видов наблюдался на 7-й день (5,0–8,0 см). Таким образом, в результате проведенных исследований установлено, что исследуемые виды растений являются питательным субстратом для Trichoderma lignorum, Fusarium oxysporum и Aspergillus niger. 2. Для выявления антифунгальной активности водных экстрактов исследуемых видов проводили экстрагирование надземных частей растений водопроводной водой в соотношении 1 : 10 на водяной бане. Полученный водный экстракт после охлаждения процеживали и разливали в колбы вместимостью 200 мл по 100 мл в каждую, pH доводили до 6,5–7,0 затем стерилизовали в течение 45 минут при 0,5 атм. Далее высевали культуры грибов в колбы с водным экстрактом исследуемых растений. Затем колбы помещали в термостат на 7 суток при температуре 25–27 ºС. В качестве контроля использовали среду Чапека. После фильтрации культурной жидкости определяли вес биомассы. Фильтрат высушивали до постоянного веса при температуре 98 оС. Водные экстракты A. сotula и A. candidissima в отличие от самих исследованных растений во всех разведениях (сухая биомасса составила от 0,16 до 1,43 г/л) проявляют фунгистатическое действие на грибы Trichoderma lignorum, Fusarium oxysporum и Aspergillus niger. Так, водные экстракты A. cotula в отношении всех 3-х грибов проявляют почти одинаковое фунгистатическое действие (сухая биомасса составила 0,28– 0,78 г/л). Тогда как 5 % водный экстракт Anthemis candidissima обладает наибольшей ингибирующей активностью в отношении Fusarium oxysporium (сухая биомасса составила 0,16 г/л), а наименьшей – 15 % водный экстракт того же растения в отношении Aspergillus niger (сухая биомасса составила 1,43 г/л) (рис. 3, 4). Рисунок 3 – Влияние водных экстрактов Anthemis cotula на рост грибов 283 Рисунок 4 – Влияние водных экстрактов Anthemis candidissima на рост грибов Таким образом, результаты исследования позволяют считать водные экстракты Anthemis cotula и A. сandidissima перспективными при создании новых противогрибковых препаратов. Л и т е р а т ур а Аскеров А. М. Высшие растения (Lamiaceae, Asteraceae) Азербайджана. Конспект флоры Азербайджана / А. М. Аскеров. – Баку: Элм, 2008. – Т. III – С. 75–77. Аскеров А. М. Конспект флоры Азербайджана / А. М. Аскеров. – Баку: Элм, 2011. – 202 с. Методы экспериментальной микологии / под ред. В. И. Билай. – Киев: Наукова Думка, 1982. – 500 с. Meхтиева Н. П. Биоразнообразие лекарственной флоры Азербайджана / Н. П. Meхтиева. – Баку: Letterpress, 2011. – 186 с. Мустафаева С. Д. Об изученности семейства Asteraceae Bercht. et J. Presl / С. Д. Мустафаева // Известия НАНА (серия биологии и медицины), 2013. – т. 68, № 1 – С. 17–23. Растительные ресурсы СССР: Цветковые растения, их химический состав, использования. Cемейство Asteraceae (Compositae). – СПб., 1993. – C. 23–25. Флора Азербайджана. – Баку, 1961. – Т. VIII – C. 248–264. Funk V. A. et al. Everywhere but Antarctica: using a supertree to understand the diversity and distribution of the Compositae / V. A. Funk, R. J. Bayer, S. Keeley, R. Chan, L. Watson, B. Gemeinholzer, E. Schilling, J. L. Panero, B. G. Baldwin, N. Garcia-Jacas, A. Susanna & R. K. Jansen // Biol. Skr. – 2005. – № 55. – С. 343–374. К ВОПРОСУ ОЦЕНКИ ФУНКЦИОНАЛЬНОЙ УСТОЙЧИВОСТИ СООБЩЕСТВ С УЧАСТИЕМ РЕДКИХ ВИДОВ РАСТЕНИЙ Невский С. А., Давиденко О. Н. Саратовский государственный университет имени Н. Г. Чернышевского, г. Саратов, biosovet@sgu.ru Оценка степени сформированности и функциональной устойчивости сообществ с участием редких видов растений – одно из перспективных направлений экологических исследований, в комплексе с изучением других факторов позволяющее прогнозировать динамику ценопопуля284 ций охраняемых видов. В литературе имеются данные об успешном использовании индекса Шеннона, рассчитанного через распределение относительных проективных покрытий видов (Мэгаран, 1992), в качестве меры энтропии сообщества (Зырянова, Бугаенко, Бугаенко, 2002; Тараканова, 2002). Построение стандартных распределений геометрических рядов и Мак-Артура, соответствующих экспериментальному, и их последующее сравнение с помощью нормированного индекса Шеннона дает возможность оценить степень сформированности растительного сообщества по близости экспериментального индекса Шеннона конкретного фитоценоза к той или иной теоретической модели распределения. Для оценки функциональной структуры ряда сообществ с участием редких видов растений Саратовской области нами предложено использование индекса сформированности сообщества (Невский, Давиденко, 2011), представляющего собой отношение разности нормированных индексов Шеннона, рассчитанных для распределения Мак-Артура и экспериментальных данных, к разности таковых для распределения Мак-Артура и геометрических рядов. , , – нормированные индексы Шеннона, рассчитанные для распределений Мак-Артура, экспериментальных данных и геометрических рядов соответственно. Значение индекса, стремящееся к единице, свидетельствует о высокой степени сформированности и функциональной устойчивости сообщества. Уменьшение значения индекса свидетельствует о дестабилизации видового состава, структуры и степени устойчивости. Индекс сформированности, выраженный в процентах от максимально возможного, показывает степень сформированности видовой структуры сообщества. В таблице приведены значения индекса сформированности некоторых растительных сообществ Саратовской области, в составе которых были отмечены редкие виды растений. Все изученные сообщества можно разделить на две группы. В первую входят сообщества Stipa capillata + Festuca valesiaca и Festuca valesiaca, характеризующиеся самыми высокими значениями индекса сформированности видовой структуры и средним видовым богатством. Эти фитоценозы расположены вдали от населенных пунктов и не несут на себе видимых следов антропогенного воздействия (табл.). 285 Характеристика фитоценозов с участием редких видов растений Фитоценоз Stipa lessingiana Festuca valesiaca + Stipa lessingiana Охраняемые виды Isf Glycyrrhiza glabra, Stipa pennata 0,56 Iris pumilla, Adonis vernalis 0,52 Glycyrrhiza glabra 0,48 Iris pumilla, Adonis vernalis, Stipa pennata 0,72 Adonis vernalis 0,89 Festuca valesiaca – Artemisia lerchiana Stipa capillata + Festuca valesiaca Festuca valesiaca Вторую группу образуют сообщества с более низкими значениями индекса сформированности – Stipa lessingiana и Festuca valesiaca + Stipa lessingiana, также не испытывающие на себе влияния негативных антропогенных факторов. Невысокие значения индекса сформированности связаны в данном случае с проникновением в состав фитоценозов солодки голой, которая за счет вегетативного размножения значительно увеличивает свою численность и плотность, дестабилизируя функциональную структурированность сообщества. Отмеченные за период исследования тенденции расселения солодки позволяют говорить о положительной динамике численности ее ценопопуляций в исследованных сообществах. В эту же группу входят сообщества, подвергающиеся умеренному воздействию выпаса скота. Однако при полученных значениях энтропии можно говорить лишь об умеренном нарушении функциональной устойчивости данных сообществ, поскольку в их состав входит значительное число типично степных видов, способных при снижении или полном исключении антропогенного воздействия достаточно быстро вернуть биоценоз к исходному состоянию. Л и т е р а т ур а Зырянова О. А. К вопросу изучения видового разнообразия коренных лесов криолитозоны Сибири / О. А. Зырянова, Т. Н. Бугаенко, Н. Н. Бугаенко // Исследовано в России (электронный журнал). – 2002. – С. 2194–2203. Мэгарран Э. Экологическое разнообразие и его измерение / Э. Мэгарран. – М., 1992. – 181 с. Невский С. А. Функциональная структура и устойчивость cтепных растительных сообществ c участием редких видов растений в саратовском Заволжье / С. А. Невский, О. Н. Давиденко // Изв. Сарат. гос. ун-та. Новая серия. Сер., Химия. Биология. Экология. – 2011. – Вып. 2. – С. 116–121. Тараканова Ю. В. Применение индекса Шеннона-Уивера для индикации структуры растительных сообществ / Ю. В. Тараканова // Тез. 6-й Пущинской школы-конф. молодых ученых. – 2002. – С. 189. 286 ВЛИЯНИЕ ГЕНЕТИЧЕСКОЙ ТРАНСФОРМАЦИИ НА ЭФФЕКТИВНОСТЬ РАБОТЫ МЕХАНИЗМОВ АДАПТАЦИИ У РАСТЕНИЙ ТАБАКА Нурминская Ю. В., Максимова Л. А., Копытина Т. В., Еникеев А. Г. Сибирский институт физиологии и биохимии растений СО РАН, г. Иркутск, nurminskaya@sifibr.irk.ru Целью каждого вида живых организмов является сохранить себя неизменным. Неизменность для вида означает неизменность генома, поскольку распространение у всех особей вида какой-либо мутации (пусть даже полезной в данных условиях среды) будет обозначать гибель этого вида-предшественника и появление нового вида (Щербаков, 2012). Поэтому стремление к поддержанию видового гомеостаза является естественным свойством живых организмов. Особенностью генома высших эукариот является его высокая интегрированность, наличие коадаптированных комплексов генов (Жученко, 2004). Генные комплексы – основа адаптационных механизмов в онтогенезе, обеспечивающих популяционный гомеостаз. С другой стороны, окружающая среда нестатична, поэтому если генотип популяции будет единообразен у всех особей, достижение популяционного гомеостаза и стабильности популяции не будет возможно – вид не сохранит себя в изменяющихся условиях среды. Следовательно, высокий популяционный или видовой гомеостаз невозможен без генетического разнообразия. Обеспечение генетической гибкости в филогенезе уравновешивает стремление живого сохранить свой генотип неизменным. Живые организмы обладают механизмами как поддержки популяционного гомеостаза, так и механизмами сохранения биоразнообразия популяции. Таким образом, генотип популяции можно представить как итог равновесия между необходимостью сохранить постоянство генома и необходимостью в поддержке биоразнообразия. С этой точки зрения становится понятным, что внедрение в геном чужеродной генетической конструкции при трансформации не может не сказаться на этом равновесии. Агробактериальную трансформацию можно рассматривать как комплексный стрессирующий фактор, включающий не только факт инсерционной мутации, но и стресс вследствие поранения, инфицирования патогеном и т. д. (Еникеев и др., 2008; Maximova et al., 2012). Как эндогенные, так и экзогенные стрессовые факторы способны влиять на функционирование рекомбинантной системы, увеличивая частоту кроссоверных обменов (Жученко, 2004). Соответственно, стресс вследствие трансформации может вызвать увеличение уровня рекомбинационной изменчивости у последующего поколения (а возможно, и поколений) 287 трансгенных растений. Считается, что генетическая трансформация может приводить к разбалансировке коадаптированных генных комплексов (Животовский, 2001; Жученко, 2004). Это может быть следствием как увеличения числа рекомбинаций, так и непосредственно внедрения чужеродной генетической конструкции в область активных генов. Нарушение слаженной работы генных комплексов может приводить к увеличению онтогенетического шума – случайных, ненаправленных нарушений в работе генома. В настоящее время имеется больше вопросов, чем ответов о том, насколько сильно у трансгенных растений нарушается равновесие между сохранением постоянства генома и поддержкой генетического разнообразия; происходит ли у них (и насколько значительно) разбалансировка коадаптированных генных комплексов. Снижение устойчивости к неблагоприятным факторам у трансгенных растений может служить косвенным признаком о такой разбалансировке, поскольку, как уже отмечалось выше, комплексы генов играют ключевую роль в адаптации организмов. Другим показателем этого может служить увеличение онтогенетического шума. Онтогенетический шум может проявлять себя в виде незначительных отклонений от идеальной симметрии (флуктуирующая асимметрия – ФА) (Захаров, 2001). В данной работе исследовали морфометрические признаки и величину ФА между площадями половин листа у шести поколений Nicotiana tabacum L., трансформированных агробактериальным штаммом A699 со встроенным геном канамицинустойчивости nptII в нормальных условиях (+25o) и при пониженной (+15o) температуре. Обнаружено, что у трансгенных растений в нормальных условиях параметры общей площади листовой поверхности, длины стебля, сухого веса надземной части менялись в поколениях, сначала несколько увеличиваясь к Т4, а затем уменьшаясь в Т5. В Т6 снова наблюдали увеличение вегетативных параметров. В условиях низкой температуры параметры трансформантов были понижены и имели тенденцию к еще большему снижению в поколениях. Самыми низкими значениями вегетативных параметров при +15o имели растения Т5 и Т6, у этих же поколений наблюдали повышенные значения ФА. Растения этих поколений имели повышенные значения ФА и в условиях роста при +25o. Ранее установлено, что трансформированные данной генетической конструкцией растения табака в условиях повышенной освещенности в целом имели более высокий уровень ФА по сравнению с контрольными растениями, одновременно с пониженной длинной стебля (Нурминская и др., 2013). 288 Растения табака, трансформированные генетической конструкцией, не имевшей целевого гена, демонстрировали пониженную устойчивость к таким неблагоприятным факторам, как низкая положительная температура и экстремальная освещенность. Факт трансформации негативно отразился на адаптивном потенциале растений, что свидетельствует о разбалансировке коадаптированных комплексов генов. Тенденция уменьшения ростовых параметров у Т5 и увеличения онтогенетического шума у Т5, Т6 может быть признаком снижения габитуса у трансформантов через определенное число поколений. Таким образом, у данных трансгенных растений даже у Т6 не наблюдается восстановления стабильной работы генома. Это может быть связано с тем, что стресс, вызванный трансформацией, привел к увеличению уровня рекомбинаций, что могло стать одной из причин нарушения работы генных комплексов. Очевидно, что стабильность популяции трансгенных растений может быть достигнута только с достижением нового равновесия между генетическим разнообразием и стабильностью работы комплексов генов. То, что это возможно, доказывает существование в природе видов растений (в том числе и табака), имеющих в геноме гены Т-ДНК Agrobacterium rhisogenes (Matveeva et al., 2012). Л и т е р а т ур а Агробактериальная трансформация как биотический стрессирующий фактор / А. Г. Еникеев [и др. ] // Журнал стресс-физиологии и биохимии растений. – 2008. – Т. 4, № 1. – С. 11–15. – Режим доступа: http://www.jspb.ru/issues/2008/N1/JSPB_2008_1_11–19. Животовский Л. А. Стабилизирующий отбор и приспособленность популяций ГМО / Л. А. Животовский // ГМО – скрытая угроза России. – М.: ОАГБ, ЦЭПР, 2004. – С. 93– 104. Захаров В. М. Онтогенез и популяция: оценка стабильности развития в природных популяциях / В. М. Захаров // Онтогенез. – 2001. – Т. 32, № 6. – С. 404–421. Жученко А. А. Экологическая генетика культурных растений и проблемы агросферы (теория и практика). /. А. А. Жученко. – М.: Агрорус, 2004. – Т. I – 690 с. Процедура трансформации вызывает изменения стабильности развития и интенсивности фотосинтеза в ряду поколений у растений табака / Ю. В. Нурминская [и др. ] // Известия ИГУ. – 2013 (в печати). Щербаков В. П. Эволюция как сопротивление энтропии. I. Механизмы видового гомеостаза / В. П. Щербаков // Журн. общ. биол. – 2005. – Т. 66, № 3. – С. 195–211. Agrobacteriummediated transformation of Nicotiana tabacum by disarmed strain At 699 resulted in considerable raising of growth and development of transgenic plants / L. A. Maximova [et al. ] // Journal of Stress Physiol. and Biochem. – 2012. – Vol. 8, № 1. – P. 138–148. Horisontal gene transfer from genus Agrobacterium to plant Linaria in nature / T. V. Matveeva [et al. ] // MPMI. – 2012. – Vol. 25, № 12. – Р. 1542–1551. 289 ОНТОГЕНЕТИЧЕСКАЯ СТРУКТУРА ЦЕНОПОПУЛЯЦИЙ ПЫЛЬЦЕГОЛОВНИКА КРАСНОГО (CEPHALANTHERA RUBRA (L.) RICH.) Османова Г. О., Чернова С. Ю. Марийский государственный университет, г. Йошкар-Ола, gyosmanova@yandex.ru Биологическое разнообразие, сформированное длительным процессом эволюции, составляет основу целостности экосистем и биосферы в целом. В настоящее время основными причинами сокращения видового разнообразия являются (Стратегия…, 2004): – уничтожение, разрушение и загрязнение местообитаний; – чрезмерное изъятие и истребление природных популяций растений; – интродукция чужеродных видов, (при этом список инвазийных чужеродных видов может пополняться за счет генетически модифицированных сортов растений, последствия и масштабы воздействия которых на природные экосистемы и популяции коренных видов непредсказуемы); – распространение болезней растений. Приоритетными мерами, направленными на сохранение редких и находящихся под угрозой исчезновения видов, являются: – сохранение популяций в естественной среде обитания; – сохранение и восстановление природной среды обитания, реконструкция биотопов; – восстановление утраченных популяций. Сохранение и восстановление редких и нуждающихся в охране видов возможно лишь при знании стратегии их выживания, так как для большинства из них не в полной мере выявлены распространение, особенности биологии и экологии, а у многих видов не изучена их популяционная жизнь. К настоящему времени накоплен значительный материал по онтогенезу, структуре и динамике популяций орхидных в разных частях ареала (Татаренко, 1996; Перебора, 2002; Блинова, 2003; Ишмуратова и др., 2003; Быченко, 2004; Быченко и др., 2006; Фардеева, 1997; Фардеева, Исламова, 2004; Вахрамеева, 2006; Железная, 2008 и др.). К сожалению, орхидные, произрастающие на территории Республики Марий Эл, практически не изучены. Объектом исследования выбран пыльцеголовник красный (Cephalanthera rubra (L.) Rich.) – многолетнее, короткокорневищное травянистое растение из семейства орхидные (Orchidaceae Juss.), криптофит (геофит). Растение имеет европейско-средиземноморскопереднеазиатский тип ареала, а по сезонному ритму развития побегов относится к весенне-летне-осенне-зеленым с периодом зимнего покоя растениям (Перебора, 2002). 290 Пыльцеголовник красный – редкий вид, который, к сожалению, страдает из-за высокой декоративности, поэтому занесен в Красные книги ряда областей и республик России (Республики Марий Эл, Удмуртии, Татарстана и др.) (Абрамов, 1989; Папченков, 1990; Красная книга Республики Марий Эл, 2013; Красная книга Удмуртской Республики, 2001; Красная книга Республики Татарстан, 2006). Изучение особей и ценопопуляций (ЦП) C. rubra проводили в 2011 г. в Моркинском районе Республики Марий Эл в окрестностях деревни Коркатово на каменном карьере близ горы Коркан-Курык на двух участках. Первый участок был расположен на искусственной терассе под горой Коркан-Курык западной экспозиции, на отвалах каменного карьера из мелкого щебня, мергеля, извести, зарастающего молодым березняком с елью обыкновенной и сосной обыкновенной, с 75 % проективным покрытием, с умеренным увлажнением. Второй участок находился на склоне горы Коркан-Курык западной экспозиции, с 35 % проективным покрытием, испытывало недостаток влаги. На каждом участке были заложены трансекты, в пределах которых – примыкающие друг к другу площадки. Общая площадь каждой исследуемой ЦП составила 144 м2. Закладку трансект проводили таким образом, чтобы они пересекали участки, характеризующиеся как относительно высоким, так и низким обилием вида. Особи C. rubra на площадках не выкапывали, а производили их подкопку. Общий объем выборки составил 470 растений. Результаты обработки списков геоботанических описаний (Компьютерная обработка…, 2008) по экологическим шкалам Д. Н. Цыганова (1983), показали, что обе ЦП C. rubra по 3 почвенным факторам (переменности увлажнения, солевого режима почв богатства почв азотом) не отличаются друг от друга. По фактору шкалы солевого режима почв, исследуемые местообитания характеризуются как небогатые / довольно богатые, по шкале богатства почв азотом – от очень бедных азотом почв до бедных азотом почв, по шкале переменности увлажнения – от слабопеременного увлажнения до умеренно переменного увлажнения. По фактору увлажнения почв ЦП 1 C. rubra произрастала в местообитании, характеризующемся как сухолесолугое так и влажно-лесолуговое, а второе местообитание (склон, ЦП 2) – сухо-лесолуговое. Обе ЦП C. rubra произрастали на полуоткрытых пространствах и в светлых лесах. Полученные нами для C. rubra результаты позволяют расширить шкалу кислотности почв и шкалу освещенности-затенения влево на 0,14 и 0,74 ступеней соответственно. Экологическая позиция C. rubra по фактору шкалы увлажнения почв находится в пределах от 11 до 13 ступеней. Нами найдена крайняя точка – 11. По факторам солевого режима почв и богатства почв азотом экологическая позиция имеет срединные 291 значения. Нами рассчитана экологическая валентность вида (потенциальная и реализованная) и коэффициент экологической эффективности (Жукова, 2004). Полученные результаты представлены в таблице. Экологические характеристики ценопопуляций C. rubra по шкалам Д. Н. Цыганова (1983) Коэффициент экологической эффективности (Кес. еf) Диапазон шкалы* Экологическая позиция вида по шкале фактора Hd (1–23) 11–13 0,13 11,00–11,51 0,07 0,54 Tr (1–19) 4–7 0,21 5,69–6,68 0,10 0,48 Потенциальная экологическая валентность (PEV) Экологическая позиция изученных ЦП Реализованная экологическая валентность (REV) Nt (1–11) 1–7 0,64 4,34–4,79 0,13 0,20 Rc (1–13) 7–11 0,38 6,86–7,74 0,14 0,37 Fh (1–11) – – 5,12–6,20 0,19 – Lc (1–9) 4–9 0,67 3,26–3,84 0,18 0,27 * Нd – шкала увлажнения почв, Тr – шкала солевого режима почв, Nt – шкала богатства почв азотом, Rс – шкала кислотности почв, Fh – шкала переменности увлажнения, Lс – шкала освещенности-затенения. Анализ экологического разнообразия изученного вида показал, что C. rubra по шкале кислотности почв гемистеновалентен, по шкале богатства почв азотом гемиэвривалентен, по шкале освещенностизатенения эвривалентен. По факторам шкал увлажнения и солевого режима почв C. rubra стеновалентен, следовательно, данные факторы можно считать лимитирующими для этого вида. Индекс толерантности, рассчитанный по четырем почвенным шкалам, позволил отнести C. rubra к гемистенобионтным растениям. Коэффициент экологической эффективности по разным шкалам находится в диапазоне от 0,13 (шкала богатства почв азотом) до 0,54 (шкала увлажнения почв). Это свидетельствует о том, что C. rubra не реализует свой потенциал полностью. Результаты изучения онтогенетической структуры ЦП показали, что в онтогенетическом спектре C. rubra (рис.) наблюдается невысокая доля ювенильных и имматурных особей по сравнению со взрослыми вегетативными и генеративными. В ЦП 2 растения C. rubra ювенильного онтогенетического состояния отсутствуют. Средняя плотность особей C. rubra в ЦП 1 составила 2,85 на 1 м2, а в ЦП 2 – 0,41 на 1 м2. Изученные ЦП C. rubra являются нормальными, неполночленными. Отсутствующими группами в обеих ЦП являются старые генера292 тивные растения и особи постгенеративного периода. Для орхидных, как отмечают некоторые авторы (Татаренко, 1996; Перебора, 2002; Быченко, 2006 и др.), и в частности у C. rubra, онтогенез может завершиться в конце генеративного периода. Доля особей, % 40 30 20 ЦП1 ЦП2 10 0 j im v1 v2 g1 g2 Онтогенетические группы Спектр онтогенетических состояний C. Rubra Среди особей прегенеративного периода в обеих ЦП доминирующими группами являются особи виргинильного состояния. В ЦП 2 соотношение особей C. rubra в имматурном и виргинильном (v2) состоянии одинаково и составило более 15 %. На долю особей прегенеративного периода в ЦП 1 приходится 38,1 %, а в ЦП 2 – 55,8 %. Особей генеративного периода в ЦП 1 больше, чем в ЦП 2 (61,9 % и 44,2 % соответственно). Высокие значения индекса восстановления – 0,62 (ЦП 1) и 1,27 (ЦП 2) свидетельствуют об активном самоподдержании ЦП C. rubra. Значения индекса возрастности (Уранов, 1975) в обеих ЦП невысоки: в ЦП 1 – ∆ = 0,264, а в ЦП 2 – ∆ = 0,330. Это свидетельствует о молодости этих ценопопуляций. В то же время высоки значения индекса эффективности (Животовский, 2001): в ЦП 1 – ω = 0,670, а в ЦП 2 – ω = 0,666. Поэтому, согласно классификации «дельта-омега» Л. А. Животовского (2001), обе ЦП C. rubra относятся к зреющим. Следовательно, полученные данные дают возможность для практических рекомендаций по оценке состояния ЦП C. rubra. Согласно данным Красной книги Республики Марий Эл (2013), C. rubra имеет статус редкого, уязвимого вида с сокращающейся численностью популяции (V), поэтому в качестве рекомендации можно предложить создание в окрестностях д. Коркатово комплексного заказника для сохранения природных ландшафтов и популяций редких видов растений. 293 А популяционный подход должен составлять основу стратегии сохранения редких и находящихся под угрозой исчезновения видов, так как только сохранение отдельных природных популяций может обеспечить полноценное сохранение вида. Исследование выполнено при поддержке НИР № 5.8479.2013 «Экологический мониторинг и прогнозирование состояния урбанизированных и природных популяций растений». Л и т е р а т ур а Абрамов Н. В. Сосудистые растения флоры Марийской АССР / Н. В. Абрамов. – Йошкар-Ола, 1989. – 147 с. Блинова И. В. Онтогенетическая структура и динамика популяций Cypripedium calceolus (Orchidaceae) в разных частях ареала вида / И. В. Блинова // Ботан. журн. – 2003. – Т. 73, № 6. – С. 36–47. Быченко Т. М. Структурное и популяционное биоразнообразие орхидных Южного Прибайкалья / Т. М. Быченко // Принципы и способы сохранения биоразнообразия: сб. материалов Всерос. науч. конф. / Мар. гос. ун-т. – Йошкар-Ола, 2004. – С. 71–74. Быченко Т. М. Биоразнообразие и мониторинг редких и исчезающих видов растений березового острова (Иркутская область) в связи с вопросами их охраны / Т. М. Быченко, Н. В. Громакова, Н. А. Дулепова // Принципы и способы сохранения биоразнообразия: сб. материалов II Всерос. науч. конф. / Мар. гос. ун-т. – ЙошкарОла, 2006. – С. 74–75. Вахрамеева М. Г. Онтогенез и динамика популяций Dactylorhiza fuchsia (Orchidaceae) / М. Г. Вахрамеева // Ботан. журн. – 2006. – Т. 67, № 11. – С. 1683– 1694. Железная Е. Л. Особенности популяционной биологии некоторых видов орхидных в разных типах фитоценозов (на примере Московской и Брянской областей): автореф. дис. … канд. биол. наук / Е. Л. Железная. – М., 2008. – 21 с. Животовский Л. А. Онтогенетические состояния, эффективная плотность и классификация популяций растений / Л. А. Животовский // Экология. – 2001. – № 1. – С. 3–7. Жукова Л. А. Оценка экологической валентности видов основных эколого-ценотических групп / Л. А. Жукова // Восточно-Европейские леса: история в голоцене и современность. – М.: Наука, 2004. – Кн. 1. – С. 256–270. Ишмуратова М. М. Состояние ценопопуляций некоторых видов сем. Orchidaceae на Южном Урале. Корневищные виды / М. М. Ишмуратова, И. В. Суюндуков, А. Р. Ишбирдин, Т. В. Жирнова, М. И. Набиуллин // Раст. ресурсы. – 2003. – Т. 39. – С. 18– 36. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoScaleWin / Е. В. Зубкова, Л. Г. Ханина, Т. И. Грохлина, Ю. А. Дорогова. – Йошкар-Ола, 2008. – 96 с. Красная книга Удмуртской Республики: Сосудистые растения, лишайники и грибы / под ред. В. В. Туганаева. – Ижевск: Изд. дом «Удмуртский университет», 2001. – 290 с. Красная книга Республики Татарстан: Животные, растения, грибы. – Казань: Идел-Пресс, 2006. – 832 с. Красная книга Республики Марий Эл. Растения. Грибы / Мар. гос. ун-т. – Йошкар-Ола, 2013. – 324 с. Папченков В. Г. О флористических находках в Марийской АССР / В. Г. Папченков // Ботан. журн. – 1990. – № 12. – С. 1773–1778. Перебора Е. А. Орхидные Северо-Западного Кавказа / Е. А. Перебора; отв. ред. И. С. Белюченко. – М.: Наука, 2002. – 253 с. Стратегия сохранения редких видов в России: теория и практика. – М., 2004. – 376 с. Татаренко И. В. Орхидные России: жизненные формы, биология, вопросы охраны / И. В. Татаренко. – М.: Аргус, 1996. – 207 с. Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов / А. А. Уранов // Биол. науки. – 1975 – № 2. – С. 7–34. Фардеева М. Б. Орхидные Республики Татарстан: биология, экология, вопросы охраны / М. Б. Фардеева. – М.: Наука, 1997. – 138 с. Фардеева М. Б. Онтогенез пыльцеголовника красного / М. Б. Фардеева, Г. Р. Исламова // Онтогенетический атлас лекарственных растений: науч. изд. / Мар. гос. ун-т; отв. ред. Л. А. Жукова. – Йошкар-Ола, 2004. – Т. 4. – С. 186–190. 294 ОЦЕНКА УСТОЙЧИВОСТИ К ЭНТОМОВРЕДИТЕЛЯМ НАЗЕМНЫХ ОРГАНОВ ГИБРИДНОГО ПОТОМСТВА СОСНЫ ОБЫКНОВЕННОЙ (PINUS SYLVESTRIS L.) В ИСПЫТАТЕЛЬНЫХ КУЛЬТУРАХ Ребко С. В., Гаврилик О. П., Гарбар Ю. И., Горошко А. И. Белорусский государственный технологический университет, г. Минск, rebkosergey@tut. by Важнейшим фактором стабильности экосистемы служит разнообразие составляющих ее элементов. Это позволяет быстро реагировать на изменение условий среды и за счет гибели слабых, неустойчивых элементов поддерживать устойчивость системы. Именно с точки зрения устойчивости биологическая система не может существовать, если все составляющие ее элементы одинаковы, поскольку они одинаково реагируют на изменение условий среды, ослабляют друг друга, что в дальнейшем ведет к их массовой гибели. Следовательно, разнообразие элементов в экосистемах повышает их устойчивость. Наоборот, малое разнообразие, особенно основного элемента экосистемы, приводит к ее разрушению. На различных этапах или стадиях развития лесные биогеоценозы и составляющие их элементы имеют неодинаковую устойчивость. Это связано с особенностями роста и развития деревьев и лесного биогеоценоза в целом. Так, у организмов низка устойчивость в начале и конце развития, а наибольшая в период размножения. Для древостоя устойчивость зависит, кроме того, от его структуры и взаимного влияния составляющих ее элементов. Воздействие этих показателей на разных стадиях проявляется по-разному, что приводит к неодинаковой устойчивости леса. Устойчивость насаждений – это способность сохранять параметры роста и развития составляющих элементов при изменении внешней среды. Устойчива та система, которая соответствует условиям среды и находится с ней в динамическом равновесии. Основная элементарная частица любой экосистемы – организм. Его жизнеспособность, жизнестойкость и приспособляемость к условиям среды определяют устойчивость биологических систем. Под жизнестойкостью понимают способность деревьев противостоять неблагоприятным условиям среды, под жизнеспособностью – продолжительность их жизни. Устойчивость лесного биогеоценоза в значительной степени зависит от его состава, условий среды и их соответствия друг другу. Чем больше состав леса соответствует условиям среды, тем он более устойчив и продуктивен. Такой лес менее подвержен воздействию неблагоприятных условий среды. В текущем моменте устойчивость насаждений определяется числом и характером 295 отмирания деревьев. В древостое отмирание деревьев – это непрерывный процесс уменьшения их числа по мере увеличения их размеров, причем в процессе естественного изреживания отмирают, как правило, отставшие в росте деревья. О снижении устойчивости древостоя можно судить по массовому отмиранию не только отставших в росте деревьев, но и более высоких ступеней толщины. Взаимосвязь устойчивости и продуктивности леса – одна из важнейших. Более продуктивные биогеоценозы, как правило, более устойчивы, поскольку они разнообразнее по составу, в них более сложны и многообразны связи. Устойчивые леса, как правило, высокопродуктивны, поскольку они высокополнотны, имеют оптимальную густоту древостоя и структуру насаждения. В пределах территории, занимаемой насаждением, условия среды неодинаковы вследствие неоднородности рельефа, почвенно-гидрологических условий, изменения полноты древостоя и других факторов. В результате естественного отбора из многообразия семенного потомства растительных видов формируется генотипический состав насаждений. Приведение насаждений в генетически устойчивое состояние осуществляется путем естественного отбора через конкурентные взаимоотношения растений между собой за эффективное использование ресурсов среды. Исследования за состоянием культур сосны обыкновенной и изучение степени их повреждения вредителями проводили в испытательных культурах различного возраста (табл. 1), созданных на землях, вышедших из сельхозпользования (2002, 2004, 2009, 2012 гг. создания) и на свежих вырубках (2008, 20113 гг. создания). Наиболее опасными вредителями, поражающими надземные органы сосны обыкновенной, являются сосновый подкорный клоп (Aradus cinnamomeus Panz.), большой сосновый долгоносик (Hylobius abietis L.) и побеговьюн летний (Rhyacionia duplana L.). Рекогносцировочный надзор за сосновым подкорным клопом проводили на участках испытательных культур 2002, 2004 и 2008 гг. создания. В культурах производился визуальный осмотр деревьев с определением их состояния и роста путем закладки пробной площади с количеством деревьев не менее 200 шт. с подразделением на поврежденные и неповрежденные. При рекогносцировочном надзоре за большим сосновым долгоносиком и побеговьюном летним долевое участие поврежденных деревьев устанавливали посредством закладки пробной площади с перечетом также не менее 200 деревьев. Для каждого участка сосны обыкновенной рассчитывали средневзвешенный балл категории состояния деревьев. 296 Таблица 1 – Характеристика испытательных культур сосны обыкновенной Год создания (площадь, га) Место расположения Схема посадки, м Густота посадки, шт./га (сохранность, %) Характеристика почвы 2002 (1,6) Негорельское лесничество НУОЛХ (кв. 70, в. 1) 2,0×0,75 6667 (96,7) дерново-подзолистая, слабооподзоленная, контактно-оглеенная, песчаная, на песке связном, сменяемом песком рыхлым, подстилаемой супесью рыхлой моренной 2004 (0,7) Негорельское лесничество НУОЛХ (кв. 72, в. 1) 3,0×1,0 3333 (93,5) дерновая, контактно-оглеенная, песчаная, на песке связном, сменяемом песком рыхлым моренным 2008 (0,4) Негорельское лесничество НУОЛХ (кв. 145, в. 4) 2,5×0,8 5000 (92,1) дерново-подзолистая, слабооподзоленная, оглеенная внизу, песчаная, на песке связном, сменяемом песком рыхлым моренным 2009 (0,3) Негорельское лесничество НУОЛХ (кв. 94, в. 14) 3,0×1,0 3333 (72,0) дерновая, контактно-оглеенная, супесчаная, на супеси рыхлой, подстилаемой супесью связной моренной 2012 (0,5) Ивьевское опытное лесничество ГЛХУ «Ивьевский лесхоз» (кв. 94, в. 4) 2,0×1,0 5000 (81,7) дерновая, контактно-оглеенная, песчаная, на песках рыхлых водноледниковых 2013 (0,2) Негорельское лесничество НУОЛХ (кв. 148, в. 1) 2,5×1,0 4000 (92,9) дерново-подзолистая, слабооподзоленная, супесчаная, на супеси рыхлой, древнеаллювиальной, сменяемой песком связным 2013 (0,5) Засимовское лесничество ГЛХУ «Кобринский лесхоз» (кв. 111, в. 17) 2,5×1,0 4000 (94,1) дерново-подзолистая, слабооподзоленная, временно избыточно увлажняемая, песчаная, на песке связном, сменяемом песком рыхлым 2013 (1,0) Негорельское лесничество НУОЛХ (кв. 162, в. 2) 3,0×1,0 3333 (93,7) дерново-подзолистая, слабооподзоленная, супесчаная, на супеси рыхлой, водно-ледниковой, сменяемой песком связным, подстилаемом супесью рыхлой моренной В результате рекогносцировочного надзора за вредителями надземных органов сосны обыкновенной в испытательных культурах установлено, что на участке 2002 г. создания выявлены растения, поврежденные сосновым подкорным клопом, большим сосновым долгоносиком и побеговьюном летним, однако их количество незначительно – соответственно 2, 9 и 3 деревца (табл. 2). Не обнаружено указанных вредителей на участках 2008, 2012 и 2013 гг. создания, а на участке 2009 г. создания не оказалось поврежденных деревьев сосны большим сосновым долгоносиком. На участке 2004 г. создания не зафиксировано повреждений побегов и почек побеговьюном летним, однако выявлено 3 растения, поврежденные сосновым подкорным клопом (категория состояния деревьев – I,1). Также выявлено 5 деревьев, поврежденных большим сосновым долгоносиком. На участке 2009 г. создания отмечено только 4 дерева, поврежденные побеговьюном летним (категория состояния деревьев – I,1). В результате детального надзора за сосновым подкорным клопом установлено, что на участке 2002 г. создания среднее количество особей вредителя достигает 6 шт./дм2, а на участке 2004 г. создания число особей клопа – всего лишь 2 шт./дм2. Степень заселенности деревьев сосновым подкорным клопом на обоих участках является низкой, следовательно, проведение определенных защитных мероприятий в испытательных культурах не требуется. Таблица 2 – Результаты рекогносцировочного надзора за вредителями наземных органов сосны обыкновенной в испытательных культурах Год создания 2002 2004 2009 Количество растений, шт. неповрежденные поврежденные Категория состояния деревьев сосновый подкорный клоп 218 2 I – здоровые большой сосновый долгоносик 211 9 I,3 – здоровые побеговьюн летний 217 3 I,2 – здоровые сосновый подкорный клоп 216 3 I,1 – здоровые большой сосновый долгоносик 212 5 I,1 – здоровые побеговьюн летний 216 4 I,1 – здоровые Вид вредителя 298 В результате проведенного лесопатологического мониторинга за вредителями наземных органов сосны обыкновенной в испытательных культурах установлено, что состояние лесных культур, созданных гибридным посадочным материалом, характеризуется как хорошее и отличное – средневзвешенный балл категории состояния деревьев колеблется от I до I,3. Исходя из этого можно заключить, что гибридное потомство сосны обыкновенной, произрастающее в испытательных культурах, отличается высокой резистентностью к основным вредителям наземных органов – сосновому подкорному клопу, большому сосновому долгоносику и побеговьюну летнему. ОЦЕНКА УСТОЙЧИВОСТИ ГИБРИДНОГО ПОТОМСТВА СОСНЫ ОБЫКНОВЕННОЙ (PINUS SYLVESTRIS L.) К ПЛАСТИНЧАТОУСЫМ-РИЗОФАГАМ НА ЗЕМЛЯХ, ВЫВЕДЕННЫХ ИЗ СЕЛЬСКОХОЗЯЙСТВЕННОГО ОБОРОТА Ребко С. В., Гаврилик О. П., Гарбар Ю. И., Горошко А. И. Белорусский государственный технологический университет, г. Минск, rebkosergey@tut.by Лес имеет важное глобальное значение не только как источник возобновляемых ресурсов, но и как главный компонент биосферы, играющий природоохранную роль. В полной мере основные функции способны выполнять только здоровые, продуктивные и устойчивые насаждения. Устойчивость – главное свойство и условие существования биологических систем. Лес как наиболее устойчивый комплекс живой природы выступает не только как стабилизатор биосферы, но и как важнейший фактор ее улучшения. Ввиду недостаточной изученности вопросы устойчивости лесных насаждений пока не нашли отражения в лесохозяйственной практике. В связи с длительностью роста леса последствия негативного влияния на его состояние имеют тенденцию к накоплению, что требует прогнозирования устойчивости лесов на перспективу. Кроме того, с усилением масштабов хозяйственной деятельности и ее воздействия на окружающую среду эта проблема приобретает все большее значение. Изучение устойчивости лесных экосистем является сложной проблемой. Для выявления причин снижения устойчивости лесов необходима система методов, основанных на глубоком и всестороннем исследовании лесных экосистем. Для древостоя устойчивость зависит, прежде всего, от его структуры и взаимного влияния составляющих ее 299 элементов. Воздействие этих показателей на разных стадиях проявляется по-разному, что приводит к неодинаковой устойчивости леса. Важнейшей проблемой лесного хозяйства в республике является улучшение состояния и повышение продуктивности лесов с целью удовлетворения потребностей народного хозяйства в высококачественной древесине. Решение этой проблемы может быть достигнуто путем применения селекции при создании будущих насаждений и, в частности, выведением новых перспективных сортов древесных пород. При формировании сорта сосны обыкновенной должны учитываться лесоводственные особенности и многообразие хозяйственного применения. Улучшенный селекционный материал сосны обыкновенной должен отвечать следующим основным критериям: высокие показатели роста, устойчивость к болезням и вредителям, достаточная репродуктивная воспроизводимость, хорошая форма ствола и высокое качество древесины. Получение новых сортов сосны обыкновенной методами отбора и гибридизации требует оценки их на устойчивость к различным болезням и вредителям. К наиболее опасным вредителям, поражающим молодые растения, относятся хрущи. В Беларуси насчитывается около 20 видов хрущей, способных повреждать корни растений. Наибольший ущерб наносят хрущи сосновым культурам, личинки которых, объедая корни растений, приводят к их гибели. К основным вредителям корней на всей территории республики относятся хрущ майский (Melolontha melolontha) и хрущ июньский (Rhizotrogus solstitialis), которые образуют как самостоятельные, так и совместные очаги. Численность вредителей определяется наличием кормовой базы, а также резистентностью растений к определенному виду вредителей. В качестве объектов исследования для оценки гибридного потомства сосны обыкновенной на устойчивость к пластинчатоусым-ризофагам был проведен лесопатологический мониторинг испытательных культур, созданных на землях, вышедших из сельскохозяйственного пользования (табл. 1) на территории Негорельского лесничества Негорельского учебно-опытного лесхоза (2002, 2004 и 2009 гг. создания) и Ивьевского опытного лесничества ГЛХУ «Ивьевский лесхоз» (2012 г. создания). При осуществлении лесопатологического мониторинга обеспечивается: своевременное выявление участков лесных культур, поврежденных вредителями и болезнями и усыхающих под воздействием природных и антропогенных факторов; получение достоверной информации о лесопатологическом и санитарном состоянии лесов, распространении и развитии вредных организмов; анализ патологических процессов, прогноз их развития, оценка возможных негативных последствий; планиро300 вание и осуществление эффективных лесозащитных и лесохозяйственных мероприятий с учетом экологической и экономической целесообразности. Лесопатологический мониторинг включает общий визуальный надзор за выявлением признаков наличия очагов пластинчатоусыхризофагов; рекогносцировочный надзор на участках насаждений, характерных для возникновения очагов и глазомерной оценки численности пластинчатоусых-ризофагов; детальный надзор на постоянных пунктах учета, на которых ведется детальный учет численности пластинчатоусых-ризофагов. Рекогносцировочный надзор за майскими хрущами проводили на участках испытательных культур в августе путем осмотра надземной части деревьев и устанавливали степень повреждаемости растений. После проведения рекогносцировочного надзора и выявления наличия майских хрущей проводили детальный надзор, основной целью которого является получение данных, позволяющих оценить численность хрущей и определить угрозу повреждения насаждений. Детальный надзор за пластинчатоусыми проводили в сентябре путем почвенных раскопок (размер почвенных ям – 1 × 1 м, количество почвенных площадок на каждом участке – 10 шт.) и определения абсолютной и относительной заселенности почвы хрущами и возрастной структуры личинок. 2002 1,6 Негорельское лесничество 2,0×0,75 НУОЛХ (кв. 70, в. 1) 2004 0,7 Негорельское лесничество НУОЛХ (кв. 72, в. 1) 0,3 Негорельское лесничество НУОЛХ (кв. 94, в. 14) 2009 Схема посадки Место расположения Год создания Площадь, га Таблица 1 – Характеристика испытательных культур сосны обыкновенной, созданных на землях, вышедших из сельхозпользования 3,0×1,0 3,0×1,0 Густота посадки, шт./га Сохранность растений, % Характеристика почвы 6667 96,7 дерново-подзолистая, слабооподзоленная, контактнооглеенная, песчаная, на песке связном, сменяемом песком рыхлым, подстилаемой супесью рыхлой моренной 3333 93,5 дерновая, контактно-оглеенная, песчаная, на песке связном, сменяемом песком рыхлым моренным 72,0 дерновая, контактнооглеенная, супесчаная, на супеси рыхлой, подстилаемой супесью связной моренной 3333 301 В результате проведенных исследований было установлено, что на участках испытательных культур, созданных на землях, вышедших из сельхозпользования, обнаружены отдельные усохшие и ослабленные деревья (табл. 2). Степень поражения растений на исследуемых объектах 2002, 2004 и 2009 гг. создания низкая, а на участке испытательных культур ГЛХУ «Ивьевский лесхоз» пораженных деревьев зафиксировано не было. Таблица 2 – Результаты рекогносцировочного надзора за пластинчатоусыми в испытательных культурах, созданных на землях сельхозпользования Год создания культур здоровых ослабленных усохших 2002 214 4 2004 205 2009 2012 Количество учтенных растений, шт. Степень поражения Намечаемые мероприятия 1 низкая – 1 – низкая – 201 1 1 низкая – 226 – – – – Данные детального надзора за пластинчатоусыми представлены в таблице 3. Таблица 3 – Результаты детального надзора за пластинчатоусыми-ризофагами в испытательных культурах, созданных на землях сельхозпользования Год создания культур Наименование, фаза развития вредителя Абсолютная заселенность, шт./м2 Степень повреждения Намечаемые мероприятия 2002 – – – – 2004 майский хрущ, личинка 1-го года 0,2 низкая – 2009 майский хрущ, личинка 1-го года 0,5 низкая – Исследования почвы в культурах показали, что на участке 2002 г. создания личинок хруща не обнаружено, в то время как в культурах 2004 и 2009 гг. создания в результате почвенных раскопок обнаружены личинки майского хруща 1-го года, однако их абсолютная заселенность в почве невысокая – соответственно 0,2 и 0,5 шт./м2, при этом степень поврежденности корней оказалась незначительной. 302 В результате проведенного лесопатологического мониторинга за пластинчатоусыми-ризофагами в культурах, созданных на землях, вышедших из сельхозпользования, личинки майского хруща обнаружены только на двух участках и их численность является низкой, что дает основания предполагать о незначительных повреждениях культур в предстоящем году и их высокой сохранности. ЗАКОНОМЕРНОСТИ ИЗМЕНЧИВОСТИ У ЭКОТИПОВ СОСНЫ ОБЫКНОВЕННОЙ В ГЕОГРАФИЧЕСКИХ КУЛЬТУРАХ НА СРЕДНЕМ УРАЛЕ Тишечкин А. Н. Ботанический сад УрО РАН, г. Екатеринбург, Tishechkin_A@mail.ru У лесов Урала, имеющих исключительно важное значение, наметилось истощение в силу переруба расчетных лесосек, неполного освоения древесины, пожаров, смены хвойных пород на лиственные, низкой эффективности лесовосстановительных мероприятий и даже их дигрессии. Одним из направлений в повышении продуктивности сосновых лесов является создание испытательных посадок из инорайонных семян и выявление быстрорастущих насаждений (Мамаев, 1973, Шутяев, 2011). Первые опыты по выращиванию сосны из семян различного происхождения были произведены 200 лет назад во Франции Вильмореном. Были испытаны образцы из 30 пунктов Европы. Выявлен замедленный рост северных и восточных происхождений, но они отличались лучшей формой ствола по сравнению с южными и западными. Опыт доказал существование наследственно закрепленных свойств у различных географических экотипов. В подобных опытах (Oleksyn, Giertych, 1984) установлены различия у сосны в длине вегетационного периода и времени цветения в зависимости от происхождения образца. В 1877 г. заложены географические культуры сосны, бука и дуба на территории Мюнхенского ботанического сада (Kienitz, 1911). В 1877–1879 гг. и в 1888–1892 гг. заложил посадки М. К. Турский (Турский, 1899) в Лесной даче бывшей Петровской земледельческой и лесной академии. Происхождения были привлечены из различных губерний Европейской России и Германии. Опыты показали, что в пределах вида у древесных пород существуют заметные различия в интенсивности роста и устойчивости, связанные с географической дифференциацией. Далее в 80-е годы XIX в. А. Цизляр (Cieslar, 1907) провел опытные посевы сосны, ели обыкновенной и лиственницы 303 из семян различного происхождения. Он отметил, что виды в природе дифференцируются на ряд климатических разновидностей или рас, которые адаптированы к длине вегетационного периода соответствующих местообитаний и передают эти свойства потомству. Известно, что иногда перенос растений из мест естественного произрастания в новые условия другого района может дать самые неожиданные результаты. С этой целью в Ревдинском лесхозе в 1976–77 гг. были заложены географические культуры сосны на площади 13,2 га. В опыте задействовано 40 географических экотипов, всего высажено 2-летними сеянцами 70,6 тыс. шт. растений по схеме 2,5×0,75 м в кварталах 68 и 82. Культуры отличаются широким диапазоном географического происхождения (от Кольского полуострова до Красноярского края). Для сравнительного анализа роста экотипов сосны была взята за основу общая методика ВНИИЛМа (Проказин, 1972), утвержденная решением Проблемного совета по лесной генетике, селекции и семеноводству от 5 апреля 1972. В данной работе проведен анализ роста и развития сосны обыкновенной северного происхождения (Ивдельский лесхоз) и южного (Курганский лесхоз). Расстояние между ними более 700 км. Северный образец происходит из подзоны средней тайги, южный – северной лесостепи. Опыт заложен в Ревдинском лесхозе – подзона южной тайги. За все годы наблюдений северный образец показал более замедленный рост по сравнению с южным. Так, например, в 1-летних посевах он отстает по высоте сеянцев на 13 %, массе надземной части на 16 %, общей массе на 10 %. В 2-летнем возрасте его отставание по массе стволика составило 14 %, массе хвои – 11 %. В 3–5–8-летнем возрасте проявилось его отставание и по длине хвои на 21–34–15 %. В посевах у северного образца все растения имели фиолетовую окраску хвои (100 %), и закладка верхушечных почек на 2 октября составила 80 %, а у южного – лишь 40 и 20 %. У двулетних сеянцев ивдельского экотипа меньше величина отношения массы надземной части к массе корней на 23 %. При таком меньшем показателе в более жестких условиях сосне легче адаптироваться и «выживать». Такая же закономерность выявлена у экотипов сосны из Башкортостана. Подобное отношение у зилаирского образца, происходящего из среднегорных областей (более жестких), также меньше на 14 % по сравнению с дюртюлинским образцом, происходящим из равнинных мест с осадочными (песчанистыми) породами, которые более благоприятны для роста сосны. В 8-летнем возрасте у север304 ного образца выявлено меньшее количество заложенных почек в мутовке на 29 %, в 20-летнем – на 5 %. К 20-летнему возрасту сохраняется отставание северного образца по высоте на 11 %, диаметру ствола на 21 %, величине сезонного прироста по высоте на 12 %, по фитомассе надземной части до 38 %. У него ниже охвоенность бокового побега на 31 % и меньше хвои на побеге на 26 %. Но у него выявлена наиболее толстая кора ствола вблизи поверхности почвы (на 34 %). У обоих образцов сосны наиболее крупная хвоя сформировалась в 1989 г. (75–86 мм), короткая – в 1992 г. (49–48 мм). В южных условиях выращивания (на песках в Звериноголовском лесхозе) у северного и южного образцов ускоряется рост по массе сеянцев в 2,6 и 5,0 раза, в том числе массе стволика в 3,8 и 6,6 раза, массе хвои в 2,7 и 5,1 раза, массе корней в 1,7 и 3,6 раза, по диаметру ствола в 17-летнем возрасте северный экотип ускоряет рост на 26 %, а южный на 46 %. У ивдельского (северного) образца в более ранние сроки проходят фенофазы. Так, например, в вегетационный период 1994 года, когда весна была затяжной и с неустойчивым температурным режимом, разверзание почек на 9 июня составило 67–100 %. Тогда как у курганского экотипа разверзания почек еще не наблюдалось. Это обусловлено не одинаковым отношением к температурному режиму наблюдаемых образцов сосны. К 34-летнему возрасту экотипы сосны достигли следующих показателей (табл. 1). Таблица 1 – Сравнительные показатели роста и развития 34-летней сосны обыкновенной двух экотипов в условиях Среднего Урала Экотип сосны Наблюдаемый признак Величина признака в % от ивдельского экотипа ивдельский курганский Высота ствола, м 16,4 18,3 111 Диаметр ствола, см 13,9 17,4 125 175 3 0,1214 0,2124 Количество деревьев, шт./га 2839 1926 68 Запас древесины, м3/га 344,6 409,1 118 Качество ствола, % вильчатость кривизна 6,0 8,6 6,3 15,8 Сохранность, % шт. 53,0 775 36,2 513 Объем ствола, м 305 Мы видим, что у северного экотипа, происходящего из среднетаежной зоны, сохраняется существенное отставание в росте по высоте и диаметру ствола на 11–25 %. Он уступает по объему ствола и запасу древесины на 75 % и 18 %. У него меньше проявилась кривизна ствола, но выше сохранность посадок. Для ивдельского экотипа характерно быстрое раскрывание семенных чешуй шишек, и продолжительность его составляет 6–9 суток (табл. 2). Таблица 2 – Динамика раскрывания шишек у двух экотипов сосны обыкновенной в лабораторных условиях (в % нарастающим итогом) Лесорастительная зона и экотип Дата наблюдения, число (март) 7 8 9 10 11 12 13 14 15 16 Среднетаежная (ивдельский) – 7 54 76 78 83 86 88 94 100 Лесостепная (курганский) – – – 16 36 54 59 66 82 86 17 18 19 20 21 95 98 100 Наблюдается клинальное направление географической изменчивости по линии север – юг с уменьшением величин большинства признаков на север и наоборот увеличение относительной толщины коры. Таким образом, северный образец из Ивдельского лесхоза показал замедленность роста по ряду признаков, обладает меньшей фитомассой, но более высокими показателями коры. По линии север – юг прослеживаются закономерности клинальной географической изменчивости. Эти закономерности необходимо учитывать в селекционных мероприятиях при лесовыращивании на Среднем Урале. Л и т е р а т ур а Мамаев С. А. Географические культуры древесных пород на Урале / С. А. Мамаев, А. Н. Тишечкин // Популяционная экология и интродукция растений: сб. тр. – Екатеринбург: УрО РАН, 2003. – Вып. 2. – С. 95–107. Мамаев С. А. Формы внутривидовой изменчивости древесных растений (на примере семейства Pinaceae на Урале) / С. А. Мамаев. – М.: Наука, 1973. – 284 с. Побединский А. В. Сосна / А. В. Побединский. – М.: Лесная пром-ть, 1979. – 125 с. Правдин Л. Ф. Сосна обыкновенная. Изменчивость, внутривидовая систематика и селекция / Л. Ф. Правдин. – М.: Наука, 1964. – 192 с. Проказин Е. П. Изучение имеющихся и создание новых географических культур (программа и методика работ) / Е. П. Проказин. – Пушкино: ВНИИЛМ, 1972. – 52 с. Турский М. Лесоводство / М. Турский. – М., 1899. Шутяев А. М. Изменчивость хвойных видов в испытательных культурах Центрального Черноземья / А. М. Шутяев. – М., 2007. – 296 с. Шутяев А. М. Каким быть лесному семеноводству в ХХI веке (книга обзор) / А. М. Шутяев. – Воронеж: Истоки, 2011. – 248 с. Cieslar A. Die Bedeutung klimatischer Varietaten unserer Holzwesen fur den Waldbau / A. Cieslar // Cbl. Ges. Forstwesen. – 1907. – Jg. 33. De Cadoll A. Gentative d’experiences sur question de modification das les especes vegetal / A. De Cadoll. – Arch. Sei Genève, 1872. Kienitz М. Formen und Abarten der gemeinen Kiefer (Pinus Sylvestris L.) / 306 М. Kienitz // Zs. Forst und Jagd. – 1911. – H. 1. – P. 13–41. Oleksyn J. Results of a 70 years old Scots pine (Pinus Sylvestris L.) provenance experiment in Pulawy / J. Oleksyn, M. Giertych // Poland Silvae Genetica. – 1984. – Vol. 33, № 1. – P. 22–27. СТРУКТУРА И ДИНАМИКА ЦЕНОПОПУЛЯЦИЙ LILIUM PUMILUM DELILE Челтыгмашева Л. Р. Хакасский государственный университет им. Н. Ф. Катанова, г. Абакан, smilelina2012@ya.ru Изучение структуры ценопопуляций (ЦП) дает представление об общем жизненном состоянии ЦП, ее способности к самовоспроизведению и перспективах развития, позволяет познать историю жизни и динамики ЦП, поэтому является одной из важнейших характеристик при изучении ЦП редких растений (Уранов, 1975; Ценопопуляция растений, 1977; Ценопопуляция растений, 1988). Исследование динамики ЦП дает возможность охарактеризовать развитие ее как системы и предсказать ее поведение в изменчивой среде (Ценопопуляция растений, 1977). Lilium pumilum Delile – многолетнее травянистое, луковичное растение. Вид является реликтом плиоценового степного комплекса (Смирнова, 1973; Положий, Ревердатто, 1976). L. pumilum занесена в Красную книгу Хакасии (2002, 2012), статус 2 – вид, сокращающийся в численности. В качестве лимитирующего фактора приводится деятельность человека. Целью работы является изучение структуры и динамики ЦП L. pumilum. Исследования проводили в вегетационный период 2012–2013 гг. в четырех ЦП. Две ЦП описаны в настоящих каменистых степях, в составе караганового осоково-злакового (ЦП 2) и караганового разнотравнозлакового фитоценозах (ЦП 5). Другие две ЦП были исследованы в луговых степях, в составе караганового разнотравно-злакового (ЦП 3) и кустарникового разнотравно-злакового фитоценозах (ЦП 4). Геоботаническое описание фитоценозов представлено в соответствии методикой Ярошенко (1969). Онтогенетическая структура ЦП вида описана согласно общепринятой методике (Ценопопуляции растений…, 1976; Заугольнова, 1994) и рекомендации по изучению редких видов (Программа…, 1986). Пространственное размещение особей ЦП на территории определялось по методике Н. М. Григорьевой (1977). Для изучения ЦП L. pumilum в пределах исследуемого фитоценоза были заложены четыре стационарные площадки размером 16 м2 каждая. На 307 площадках маркировали все особи данного вида и выделяли их онтогенетические состояния (Ценопопуляция растений, 1977; 1988). Все исследованные ЦП L. pumilum нормальные, неполночленные. В настоящих каменистых степях обнаружены особи имматурного и виргинильного онтогенетического состояния и генеративной фракций. В луговых степях отсутствуют особи постгенеративного периода. На основании полученных результатов были построены онтогенетические спектры. Онтогенетические спектры в оба года исследований для ЦП 2 (настоящая каменистая степь) и ЦП 3 (луговая степь) – центрированные. Резких колебаний изменения численности особей в ЦП не наблюдалось. Тщательные исследования показали отсутствие молодых особей (проростки, ювенильное онтогенетические состояния) в луговой (ЦП 3) и настоящей каменистой степях (ЦП 2). Это объясняется характерной для L. pumilum короткой ювенильной стадией (1 год) и погодными условиями вегетационного сезона. В карагановой разнотравно-злаковой настоящей каменистой степи (ЦП 5) наблюдалось смещение абсолютного максимума с молодого генеративного онтогенетического состояния на особи виргинильного онтогенетического состояния. При повторной маркировке особей в следующий вегетационный период (2013 г.) расположение особей и их число на площадках изменилось. В настоящих каменистых степях (ЦП 2, ЦП 5) наблюдалось увеличение числа особей имматурного онтогенетического состояния. В кустарниковой разнотравно-злаковой луговой степи (ЦП 4) особи из виргинильного онтогенетического состояния перешли в молодое генеративное онтогенетическое состояние. Число особей среднего генеративного онтогенетического состояния резко сократилось. Особи L. pumilum после цветения быстро отмирают и, как правило, редко переходят в старое онтогенетическое состояние, поэтому особи постгенеративной фракции не были обнаружены. Размещение растений на учетных площадках носило неясно выраженный контагиозный характер. В связи с этим скопления были выделены лишь для ЦП 4. В кустарниковой разнотравно-злаковой луговой степи (ЦП 4) были выделены локусы с повышенной плотностью особей (скопления) и локусы с более низкой плотностью (промежутки). Ценопопуляционные локусы развиваются асинхронно, так как онтогенетические изменения происходят в разных частях ЦП не одновременно и с разной интенсивностью. Скопления различаются по размерам, притом крупные скопления включают несколько более мелких скоплений. Данные скопления имеют в своем составе плотный центр, состоящий из особей одного онтогенетического состояния. Так особи молодо308 го (имматурное, виргинильное) онтогенетического состояния образуют плотные скопления, а особи генеративного онтогенетического состояния расположены между собой на большем расстоянии. Именно в связи с этим в кустарниковой разнотравно-злаковой луговой степи (ЦП 4) наблюдается чередование пятен разной плотности и онтогенетической структуры. Таким образом, все исследуемые ЦП нормальные, неполночленные. Резких колебаний в численности особей в ЦП не наблюдалось. Сохранение особей молодой фракции зависит от погодных условий вегетационного периода. Размещение растений на учетных площадках носит неясно выраженный контагиозный характер, реже выделены локусы с повышенной плотностью особей (скопления), образованные особями молодой фракции. Л и т е р а т ур а Григорьева Н. М. Особенности пространственной структуры ценопопуляций некоторых видов растений / Н. М. Григорьева, Л. Б. Заугольнова, О. В. Смирнова // Ценопопуляции растений (развитие и взаимоотношения). – М.: Наука, 1977. – С. 20–36. Заугольнова Л. Б. Ценопопуляция растений (очерки популяционной биологии) / Л. Б. Заугольнова, Л. А. Жукова. – М: Наука, 1988. – 184 с. Заугольнова Л. Б. Структура популяций семенных растений и проблемы их мониторинга: автореф. дис. … д-ра биол. наук / Л. Б. Заугольнова. – СПб., 1994. – 70 с. Положий А. В. Семейство Ranunculaceae. Лютиковые / А. В. Положий, В. В. Ревердатто // Флора Красноярского края. – Томск: ТГУ, 1976. – Вып. 5, ч. 3. – С. 41–114. Программа и методика наблюдений за ценопопуляциями видов растений Красной книги СССР / Л. В. Денисова, Л. Б. Заугольнова, С. В. Никитина. – М.: Наука, 1986. – 34 с. Смирнова В. А. Анализ флоры южных каменистых склонов в долине реки Енисея (от г. Кызыла до реки Подкаменной Тунгуски): автореф. дис. ... канд. биол. наук / В. А. Смирнова. – Томск, 1973. – 19 с. Смирнова О. В. Ценопопуляция растений (основные понятия и структура) / О. В. Смирнова, Л. Б. Заугольнова. – М.: Наука, 1976. – 217 с. Уранов А. А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов / А. А. Уранов // Биол. наук. – 1975. – № 2. – С. 7–34. Ярошенко П. Д. Геоботаника / П. Д. Ярошенко. – М.: Просвещение, 1969. – 200 с. РЕПАТРИАЦИЯ И ИСКУССТВЕННОЕ СОЗДАНИЕ ПОПУЛЯЦИЙ ТУБЕРОИДНЫХ ОРХИДНЫХ КАК ОДИН ИЗ МЕТОДОВ ПОДДЕРЖАНИЯ ИХ ЧИСЛЕННОСТИ Широков А. И., Крюков Л. А., Сырова В. В., Штаркман Н. Н. НИИ БС Нижегородского государственного университета им. Н. И. Лобачевского, г. Нижний Новгород, aishirokov@mail.ru Одним из перспективных путей охраны редких и исчезающих видов растений является их искусственное размножение и введение в культуру. Перспективность этого метода обусловлена возможностью последующего «внедрения редких видов в естественный покров», в место309 обитания, из которых тот или иной вид исчез (Соболевская и др., 1985). До сих пор нет единого мнения для обозначения действий, связанных с «переносом растений в природные экосистемы» (Лубягина, 1990). Наиболее часто употребляются термины «реинтродукция» и «репатриация» (Соболевская и др., 1985). По нашему мнению, наиболее приемлем термин «репатриация», под которым понимается переселение или расселение растений в район их бывшего местообитания (Скворцов, 1971). К настоящему времени накоплен значительный опыт по выращиванию орхидных умеренной зоны in vitro из семян с целью изучения особенностей их развития, охраны и введения в культуру (Rasmussen, 1995; Malmgren, 1996; Куликов, Филиппов, 1998, 2000; Андронова и др., 2000; Батыгина, Васильева, 2002; Широков и др., 2005 и мн. др.). В НИИ Ботанический сад Нижегородского государственного университета им. Н. И. Лобачевского (НИИ БС ННГУ) работы по семенному размножению редких видов орхидных России начались более 10 лет назад. В настоящее время регулярно успешно (80–100 % прорастание семян) выращиваются такие виды, как Dactylorhiza fuchsii (Druce) Soó, D. incarnata (L.) Soó, D maculata (L.) Soó, D. urvilliana (Steud.) H. Baumann et Künkele, D. traunsteinerii (Saut.) Soó, Gymnodenia conopsea (L.) R. Br. ; относительно успешно (с небольшим, но регулярным числом проросших семян) – Platanthera bifolia (L.) Rich, Orchis militaris L., Ophris oestrifera M. Bieb. и др. ; малоуспешно – (единичное прорастание) – Cypripedium calceolus L., C. macranthon Sw. и др. Посев семян осуществляется в биотехнологической лаборатории in vitro. Для посева используются недозрелые коробочки (семена внутри них стерильные и не требуют непосредственной стерилизации). В качестве стерилизующего раствора для коробочек используется спирт. Посев осуществляется на стерильную питательную среду, предложенную С. Мальмгрин (Malmgren, 1996), которая не содержит неорганического азота и включает комплекс из 16 аминокислот. Она идеально моделирует условия микотрофного питания проростков. После посева колбы содержатся в темноте в условиях кондиционируемой камеры, при температуре 18 °С. После прорастания колбы переносятся в холодильник (температура 4 °С) на 3 месяца. В дальнейшем, по мере роста сеянцев и расходования среды, осуществляются пересадки, а также каждые 3 месяца чередуются условия температурного режима (камера-холодильник) для имитации смены условий для вегетации и покоя. В зависимости от вида продолжительность культивирования от 1 до 3 лет. По достижении имматурного онтогенетического состояния (достаточно сформированные запасающие корни, тубероиды, корневища) рас310 тения высаживаются в деревянные ящики (размер 40 × 25 × 15 см), заполненные почвенной смесью, для адаптации и доращивания. Высадка производится в конце мая – начале июня (с наступлением летних температур – не ниже 15 °С), сразу после содержания в холодильнике – для стимуляции ростовых процессов. В качестве почвенного субстрата используется следующая смесь: 1 часть дробленого керамзита (фракция 0,5 см); 1 часть дробленого ракушечника (фракция 0,5 см); 1 часть агроперлита; 2 части полугумифицированной хвойной подстилки (просеянная через крупное сито). Ящики вкапываются в гряды с притенкой и регулярным поливом (по мере необходимости). В таких условиях растения доращиваются на протяжении 2 лет. За это время они, как правило, достигают развитого виргинильного состояния и вполне пригодны для высадки в природные местообитания. Эксперименты по высадке в природные местообитания начаты нами 2 года назад. В качестве модельных выступили виды с различными эколого-ценотическими особенностями – Dactylorhiza fuchsii, D. incarnata и Gymnodenia conopsea. Всего в экспериментах было задействовано около 500 особей указанных видов. Высадка в природные местообитания осуществлялась на охраняемых природных территориях («Пустынский государственный заказник» и памятник природы «Дубрава ботанического сада») в результате тщательного подбора местообитаний и подготовки участка. Большинство орхидных умеренно-холодного климата чутко реагируют на изменение эдафических условий и характеризуются слабой конкурентоспособностью. Поэтому кроме выбора типичных местообитаний для данных видов (оптимально, если хотя бы единично присутствуют особи высаживаемого вида) перед высадкой тщательно готовили участки для посадки. Для этого изготавливали короба из досок 1,5 × 1,5 × 0,2 м. Такого же размера были выкопаны траншеи, и в них установлены короба (верхний край доски по уровню почвы). Далее, в полученные траншеи, обрамленные досками, насыпали почвенный субстрат, состоящий из 2 частей торфа, 1 части песка, 1 части доломитовой крошки (дробленый ракушечник фракция до 1 см), 1 часть местного грунта (выбирался верхний, гумусовый горизонт, но по возможности без дикорастущих растений). Затем высаживали подращенные орхидные с шагом посадки не менее 10 и не более 20 см (для более высокой плотности, чтобы снизить конкурентное воздействие «сорняков»). После посадки растения поливали. Далее 1 раз в месяц (3–4 раза за вегетационный период) осуществляли контрольные проверки с выполнением разовых уходных работ – прополка (внутри контура коробов) от «сорняков», при засухе – полив. В результате – приживаемость растений составила от 70 до 90 %. На второй год единичные особи за311 цвели и образовали полноценные семена. В последующем древесина обрамляющихся досок гумифицируется, но и конкурентное воздействие будет уже не так опасно для растений, так как взрослые особи уже становятся более конкурентоспособными. Однако для восстановления полноценной популяции высадку необходимо осуществлять на протяжении 5–6 лет, высаживая растения поэтапно – раз в 1–2 года. В результате формируется относительно разновозрастная популяция, что характеризует ее более устойчивым состоянием. Л и т е р а т ур а Андронова Е. В. Проблемы и перспективы семенного размножения in vitro орхидных умеренной зоны / Е. В. Андронова, П. В. Куликов, Е. Г. Филиппов, В. Е. Васильева, Т. Б. Батыгина // Эмбриология цветковых растений. Терминология и концепция. Системы репродукции. – СПб.: Мир и семья, 2000. – Т. 3. – С. 513–524. Батыгина Т. Б. Размножение растений / Т. Б. Батыгина, В. Е. Васильева. – СПб.: Изд-во СПб ГУ, 2002. – 232 с. Куликов П. В. О методах размножения орхидных умеренной зоны в культуре in vitro / П. В. Куликов, Е. Г. Филиппов // Бюл. Главного ботан. сада. – М.: Наука, 1998. – С. 125– 131. Куликов П. В. Репродуктивная стратегия орхидных умеренной зоны / П. В. Куликов, Е. Г. Филиппов // Эмбриология цветковых растений. Терминология и концепция. Системы репродукции / под ред. Т. Б. Батыгиной. – СПб.: Мир и семья, 2000. – Т. 3. – С. 442–446. Лубягина Н. П. Формирование популяций охраняемых видов растений в искусственных фитоценозах / Н. П. Лубягина // Бюл. ГБС АН СССР. – М., 1990. – Вып. 155. – С. 55–59. Скворцов А. К. Сущность таксона и внутривидовой систематики растений / А. К. Скворцов // Бюл. МОИП. Отб. биол. – 1971. – Т. 86, № 5. – С. 72–81. Соболевская К. А. Полезные растения Западного участка БАМ / К. А. Соболевская, Г. И. Горохова, Э. М. Гонтарь. – Новосибирск: Наука, 1985. – 230 с. Широков А. И. Культивирование орхидей европейской России / А. И. Широков, Г. Л. Коломейцева, А. В. Буров, Е. В. Каменева. – Н. Новгород, 2005. – 64 с. Malmgren S. Orchid propagation: theory and practice / S. Malmgren // North American Native Terrestrial Orchids «Propagation and Production» Conference proceeding. – Washington, 1996. – P. 63–71. Rasmussen H. N. Terrestrial orchids: from seed to mycotrophic plant / H. N. Rasmussen. – Cambridge University Press, 1995. – 433 р. СТРУКТУРНО-ФУНКЦИОНАЛЬНОЕ ЗОНИРОВАНИЕ МНОГОЛЕТНИХ ПОЛУРОЗЕТОЧНЫХ ЦВЕТОНОСНЫХ ПОБЕГОВ МОНОКАРПИКОВ Щербакова О. Ф., Калистая М. С. Национальный научно-природоведческий музей НАН Украины, г. Киев, botmuseum@ukr.net При выделении последовательности метамеров, представляющих одну структурно-функциональную зону, определяющее значение имеют морфоструктурные, ритмологические и ростовые особенности аксиллярных комплексов. При этом дополнительно учитывается емкость почек, активность их меристем, продолжительность жизни развивающихся боковых побегов, энергия их роста и степень развития. 312 По мере накопления данных о зонировании побегов у представителей различных биоморф возрастает число вариантов (Борисова, Попова 1990) исходной модели строения побега по В. Троллю (Troll, 1964), главным образом, за счет выделения дополнительных структурнофункциональных зон, различных их сочетаний и способов взаиморасположения. Относительно положения на побеге дополнительно стали выделять нижнюю, среднюю и верхнюю зоны торможения (Мусина, 1976; Серебрякова, Петухова, 1978; Серебрякова, Павлова, 1986; Борисова, Попова 1990); в зависимости от степени сформированности аксиллярного комплекса – типичную зону обогащения и скрытогенеративную (Савиных, 1999); в зависимости от положения флоральной зоны – зону ложнотерминального интеркалярного соцветия (Савиных, 2000). Мы, в свою очередь, предлагаем скрытогенеративную зону включать в зону обогащения, выделяя в последней подзону со сниженной ростовой активностью пазушных почек, находящихся на стадии развертывания или бутонизации, а также сформированных, но слаборазвитых или ослабленных побегов обогащения (как цветоносных, так и вегетативных) и подзону нормально функционирующих побегов обогащения: вегетативных, вегетативно-цветоносных (паракладиев) и цветоносных. Анализ структурно-функционального зонирования побегов, проводимый преимущественно для многолетних травянистых поликарпиков, не менее информативен и актуален для монокарпиков. Функциональнозональная структура цветоносных побегов монокарпиков и поликарпиков вполне сопоставима, отличия заключаются, главным образом, в строении зоны возобновления. Монокарпические растения – эволюционно наиболее продвинутая и пластичная жизненная форма (Марков, 1990). Поливариантность зональной организации побегов в большей мере характерна для монокарпиков (одно-, малолетников) и проявляется через разную степень выраженности зон у особей одного вида в зависимости от условий их произрастания (Петухова и др., 2009). Предложенную нами схему зонирования побега отображает рисунок. При проведении зонирования многолетних монокарпических побегов учитывали, что функция одной зоны может меняться в течение вегетационного периода и в зависимости от стадии развития побега (Савиных, Мальцева, 2008). Нижняя зона торможения, или зона ингибирования нижняя (ЗИн), находится в базальной части полициклического монокарпического побега и охватывает серию укороченных метамеров с низкой активностью пазушных меристем. 313 Зона побегов, замещения или супплентативная (СуЗ) (лат. supplentis – замещающий, восполняющий, дополняющий). Как и предыдущая зона включается в подземный участок резида, часть почек которого, имея разный период покоя, реализуется в озимые моно-, олиго-, полициклические латеральные побеги с полным или неполным циклом развития. Активная зона возобновления, или инновативная зона (лат. innovatio – обновление, возобновление) (ИЗа), занимает приземный участок резида, т. е. охватывает метамеры прошлогоднего розеточного годичного побега с формирующимися весной текущего года побегами возобновления. Все рассмотренные выше зоны выделяются в составе многолетней части монокарпического побега – резида. Иногда затруднительно установить их точные границы. Инновативная зона проявляется весной, когда отрастают новые побеги возобновления. Суплентативную зону вы314 деляют по перезимовавшим одно-, многолетним латеральным побегам. Кроме того, не все почки инновативной и суплентативной зон переходят на внепочечную стадию развития. Последующие зоны выделяются в приросте текущего года многолетнего монокарпического побега: вегетативном розеточном (первые две зоны) и цветоносном полурозеточном (все остальные ниже изложенные зоны). Потенциальная инновативная зона (ИЗп) включает метамеры с почками возобновления, пребывающими в состоянии покоя на протяжении одного неблагоприятного сезона, т. е. до весны следующего года. Эта зона охватывает всю розеточную часть прироста монокарпического побега текущего года. Аксиллярный аппарат выделяемого участка цветоносного побега монокарпика либо полностью элиминирует, что связано с полным некрозом особи после завершения единственного репродуктивного цикла, либо сохраняет жизнеспособность и переходит на постинициальную фазу развития в течение текущего вегетационного периода, а в отдельных случаях и в следующем году, т. е. соответствует типичным почкам возобновления. Отмеченная пролонгация жизни пазушных меристем и возможность их последующей реализации в побеги возобновления у некоторых монокарпиков скорее связана с наличием большого запаса питательных веществ в подземном побегово-корневом комплексе. Типичная зона возобновления у монокарпиков также выделяется в случае повреждения верхушечной почки первичного побега (Борисова, Попова, 1990). Характерное для некоторых особей вида осеннее отрастание – результат досрочной активизации почек в составе потенциальной инновативной зоны. Такое ветвление возможно благодаря нерастраченному большому ресурсу пластических веществ подземной сферы и улучшению влагообеспечения в конце вегетационного периода. В этом случае боковые побеги развиваются как пролептические после непродолжительного периода покоя, а их прирост текущего года имеет розеточную структуру и неполный цикл развития. Зону монокарпического побега с позднелетними или осенними пролептическими боковыми побегами предлагаем называть прецелеративной (лат. praecelero – опережать) (ПЗ). Средняя зона торможения, или средняя зона ингибирования (ЗИс), аналогично нижней зоне торможения охватывает серию удлиненных метамеров цветоносного прироста годичного побега с низкой активностью пазушных почек. Зона силлептического обогащения, или симультативная зона (лат. simultaneus – одновременный, совместный) (СЗ), включает мета315 меры побега выше средней зоны торможения. Аксиллярный комплекс этой зоны развивается силлептически, т. е. без периода покоя, одновременно с развертывающейся верхушкой медиального побега. Возможно подразделение данной зоны на пассивную (скрытогенеративную) (СЗп) и активную (СЗа) подзоны. В пассивной зоне обогащения формируются, как правило, ослабленные цветоносные или вегетативные побеги. В зоне активного обогащения развиваются вегетативно-цветоносные (нижние паракладии) и цветоносные боковые побеги. Зона терминальной инфлоресценции, или верхушечного соцветия (цветка) (ЗТИ). Зона терминальной инфлоресценции и симультативная зона в совокупности составляют объединенное соцветие или синфлоресценцию монокарпического побега. Иногда между терминальной флоральной единицей и симультативной зоной можно выделить верхнюю зону ингибирования. Таким образом, полный набор структурно-функциональных зон многолетнего монокарпического побега с полным циклом развития отражает формула: [ЗИн–Суз*–ИЗа*]–[/ИЗп (ПЗ*)/–//ЗИс–СЗп*–СЗа– ЗИв*–ЗТИ//]. В структуре первичного (одноосного, некустящегося) побега, как правило, суплентативная и активная инновативная зоны не выражены. Прецелеративная зона проявляется факультативно в случае формирования пролептических побегов отрастания в конце вегетации осевого годичного побега. В структуре цветоносных монокарпических побегов не всегда выражена и пассивная симультативная зона. Первые три зоны выделяются в составе многолетнего резида, последующие – в составе прироста текущего года. Потенциальная инновативная зона охватывает всю розеточную часть годичного побега. В структуре завершающего удлиненного вегетативно-цветоносного прироста полициклических монокарпических побегов выделяются: средняя зона ингибирования, симультативная зона, зона ингибирования верхняя и зона терминальной инфлоресценции. Последние три зоны составляют единую флоральную зону или синфлоресценцию. Л и т е р а т ур а Борисова И. В. Разнообразие функционально-зональной структуры побегов многолетних трав / И. В. Борисова, Г. А Попова // Ботан. журн. – 1990. – Т. 75, № 10. – С. 1420– 1426. Марков М. В. Популяционная биология розеточных и полурозеточных малолетних растений / М. В. Марков. – Казань: Изд-во Казанского ун-та, 1990. – 178 с. Мусина Л. С. Побегообразование и становление жизненных форм некоторых розеткообразующих трав / Л. С. Мусина // Бюл. МОИП. Отд. биол. – 1976. – Т. 81, вып. 6. – С. 123–132. Петухова Л. В. Варианты структуры побегов однолетних растений / Л. В. Петухова, О. Б. Черноброва, Е. Н. Степанова // Вестник ТвГУ. Сер., Биология и экология. – 2009. – Вып. 5. – С. 151–154. Савиных Н. П. Модуль у растений как структура и категория / Н. П. Савиных, Т. А. Мальцева // Вестник ТвГУ. Сер., Биология и экология. – 2008. – Вып. 9. – С. 227– 233. Савиных Н. П. Биоморфология вероник России и сопредельных государств: авто316 реф. дис. … канд. биол. наук / Н. П. Савиных. – М., 2000. – 32 с. Савиных Н. П. Побегообразование, морфогенез Veronica gentianoides Vahl. (Scrophulariaceae) и происхождение полурозеточных трав / Н. П. Савиных // Ботан. журн. – 1999. – Т. 84, № 6. – С. 20–31. Серебрякова Т. И. Побегообразование, ритм развития и вегетативное размножение в секции Potentilla рода Potentilla (Rosaceae) / Т. И. Серебрякова, Н. Р. Павлова // Ботан. журн. – 1986. – Т. 71, № 2. – С. 154–167. Серебрякова Т. И. Архитектурная модель и жизненные формы некоторых травянистых Розоцветных / Т. И. Серебрякова, Л. В. Петухова // Бюл. МОИП. Отд. биол. – 1978. – Т. 83, вып. 6. – С. 51–66. Troll W. Die Infloreszenzen / W. Troll. – Bd. 1. – Jena: Gustav Fischer-Verlag, 1964. – 615 p. 317 АВТОРСКИЙ УКАЗАТЕЛЬ Абдыева Р. Т. 245 Абрамова Л. М. 107 Агаева П. Н. 221 Азин А. Л. 7 Алейников А. А. 249 Алиева С. А. 240 Анциферов А. Л. 224 Арсланова А. А. 190 Ахметова А. Ш. 227 Байкова И. В. 101 Баранова О. Г. 10 Бахшалиева К. Ф. 280 Безель В. С. 110 Бекмансуров М. В. 88, 91, 230 Беляченко А. А. 95 Бирюкова О. А. 67 Бобровский М. В. 114 Бухарина И. Л. 179 Васильева В. К. 209 Ведерников А. А. 54, 190 Ведерникова О. П. 70, 233, 237 Воскресенский В. С. 206 Гаврилик О. П. 295, 299 Гаврилко Д. Е. 131 Гаранин В. И. 153 Гарбар Ю. И. 295, 299 Гасанова М. Ю. 240 Гетманец И. А. 52, 98, 101 Глаголев Д. 193 Глушко С. Г. 182 Голованов Я. М. 104, 107 Гордеева В. А. 110 Горошко А. И. 295, 299 Грозовская И. С. 114 Гулиев Ф. Б. 240 Гусева А. А. 243 Давиденко О. Н. 284 Дадашева А. Г. 245 Дбар Р. С. 77 Дементьева С. М. 185 Денисова Т. А. 67 Деревенская О. Ю. 80 Дробот В. И. 118 Дробот Г. П. 54, 190 Дубровная С. А. 58 Еникеев А. Г. 287 Ермакова И. М. 61 Ефименко А. С. 249 Ефремова Л. П. 67 Жуйкова Т. В. 110 Жукова Л. А. 14, 70 Жукова О. В. 123 Забродин И. В. 255 Закамская Е. С. 124 Замалетдинов Р. И. 80 Зарипова А. А. 227 Зарипова Н. Р. 80 Злобин Ю. А. 257 Зонтиков Д. Н. 274 Зотина С. Г. 124 Зуева Л. В. 185 Ибрагимова К. К. 35 Иванова В. М. 77 Иванова Н. В. 262 Иванова Р. Р. 193, 197 Иванова С. А. 185 Ившин В. П. 21 Ившина И. Б. 110 Ильина В. Н. 266 Калашников Н. В. 98 Калистая М. С. 312 Карнаухова Н. А. 269 Клименко А. А. 127 Козырева С. В. 70 Колегова Е. Б. 272 Комаревцева Е. К. 73 Копытина Т. В. 287 Костина Л. В. 110 Кохов А. В. 23 Криницын И. Г. 274 318 Крюков Л. А. 309 Крюкова А. В. 107 Кудрявцева А. Е. 209 Кузьмин П. А. 179 Куфтина Н. И. 193 Кучер Е. Н. 277 Кушион М. Т. 200 Латышева Д. Ю. 274 Лебедев В. П. 274 Леонова Т. Ш. 35, 140 Литвинская С. А. 26 Мавлюдова Л. У. 58 Макеев И. С. 131 Максимова Л. А. 287 Малкова Т. 193 Мальцева Н. Л. 54 Мартынов А. В. 134 Мастерова А. В. 274 Мейсурова А. Ф. 185 Мингазова Д. Ю. 77 Мингазова Н. М. 77, 80, 217 Мирсаитов Н. Г. 140 Морова Н. С. 32 Москвина И. В. 52 Мударисов Р. Г. 138, 140 Мустаева Н. В. 88 Мустафаева С. Д. 280 Набеева Э. Г. 77, 80 Невский С. А. 284 Никитин А. В. 217 Нотов А. А. 14, 144, 185 Нургизаринов А. М. 214 Нурминская Ю. В. 287 Османова Г. О. 221, 290 Палагушкина О. В. 80 Пендинен Г. И. 48 Полежаева А. Ю. 224 Полянская Т. А. 150 Поролло А. А. 200 Разгуляева Н. А. 274 Рахимов И. И. 35 Ребко С. В. 295, 299 Ремизова О. С. 54 Решетник Г. В. 277 Романов М. С. 114 Русов В. А. 23 Рыбакова М. А. 91 Савиных Н. П. 38 Сарбаева Е. В. 206 Свинин А. О. 54, 153 Сидоренко М. В. 157, 161 Сиханова Н. С. 214 Смирнов В. Э. 114 Соловьева М. Н. 230 Соловьева О. С. 209 Стаменов М. Н. 83 Сугоркина Н. С. 61 Сырова В. В. 309 Темнухин В. Б. 163 Тишечкин А. Н. 303 Токарева Н. А. 197 Тубаева М. А. 173 Турмухаметова Н. В. 164 Утенкова С. Н. 167 Фархуллин Р. Ш. 170 Фокина М. Е. 134 Халиуллина Л. Р. 233 Ханина Л. Г. 114 Хохлова Л. П. 41 Хульт К. 21 Хусаинов Р. В. 86 Цепкова Н. Л. 173 Челтыгмашева Л. Р. 307 Чернов В. Е. 48 Чернова С. Ю. 290 Чмелева С. И. 277 Широков А. И. 309 Штаркман Н. Н. 309 Шынбергенов Е. А. 214 Щербакова О. Ф. 312 Юнина В. П. 157 Юпина Г. А. 217 Ягодарова О. С. 237 Яковлев В. А. 176 Яковлева А. В. 176 319 СОДЕРЖАНИЕ Предисловие ........................................................................................................................5 Пленарные доклады Азин А. Л. Первичное здоровье человека и вероятность феномена «стопэволюция» на уровне Homo sapiens .........................................................................................7 Баранова О. Г. Охрана видового разнообразия растений на особо охраняемых природных территориях в Удмуртской Республике .......................................10 Жукова Л. А., Нотов А. А. Популяционно-онтогенетические исследования и проблема сохранения биоразнообразия .............................................................................14 Ившин В. П., Хульт К. Перспективы производства пеллет в Республике Марий Эл ..................................................................................................................................21 Кохов А. В., Русов В. А. Проблемы изучения биоразнообразия в средней школе .........................................................................................................................................23 Литвинская С. А. Особенности биоразнообразия Российской части Кавказа в условиях изменения окружающей среды ...........................................................................26 Морова Н. С. Экология детства и новые подходы к социальнопедагогической реабилитации детей-инвалидов ..................................................................32 Рахимов И. И., Ибрагимова К. К., Леонова Т. Ш. Сохранение биоразнообразия природных экосистем Татарстана в условиях интенсивной хозяйственной деятельности ...................................................................................................35 Савиных Н. П. О некоторых особенностях структурной организации растений в условиях переменного увлажнения .....................................................................38 Хохлова Л. П. Генетические основы сортового разнообразия яровой пшеницы при действии повышенных температур и засухи ..................................................................41 Чернов В. Е., Пендинен Г. И. Сохранение и изучение биоразнообразия дикорастущих видов рода Hordeum L. в коллекции ВИР .....................................................48 Секция 1. Биоразнообразие жизненных форм живых организмов в наземных и водных экосистемах Гетманец И. А., Москвина И. В. Классификация и экологическое разнообразие фитоценозов подзоны темнохвойных лесов Челябинской области .............52 Дробот Г. П., Ведерников А. А., Ремизова О. С., Мальцева Н. Л., Свинин А. О. Предварительные материалы о паразитарных инвазиях у представителей герпетофауны Республики Марий Эл........................................................54 Дубровная С. А., Мавлюдова Л. У. Анатомическое строение надземных побегов зверобоя продырявленного (Hypericum perforatum L.) полукустарничковой жизненной формы ................................................................................58 Ермакова И. М., Сугоркина Н. С. Влияние отмены выпаса на биоразнообразие растительности многолетних пастбищ в пойме реки Угры .....................61 Ефремова Л. П., Бирюкова О. А., Денисова Т. А. Сравнительное изучение видов и сортов лапчаток ..........................................................................................................67 320 Козырева С. В., Ведерникова О. П., Жукова Л. А. Онтогенетический гербарий как основа для изучения внутрипопуляционного биоразнообразия ....................70 Комаревцева Е. К. Развитие кустарничка Pentaphylloides Fruticosa (L.) O. Schwarz в Горном Алтае .....................................................................................................73 Мингазова Д. Ю., Мингазова Н. М., Набеева Э. Г., Иванова В. М., Дбар Р. С. Видовой состав макрозообентоса рек Абхазии .....................................................................77 Мингазова Н. М., Деревенская О. Ю., Палагушкина О. В., Замалетдинов Р. И., Набеева Э. Г., Зарипова Н. Р. Оценка биологического разнообразия водных объектов и прибрежных территорий г. Казани ......................................................................80 Стаменов М. Н. Разнообразие систем годичных побегов Quercus robur L. в центре европейской части России ........................................................................................83 Хусаинов Р. В. К экологии нематод рода Aphelenchoides ..............................................86 Секция 2. Таксономическое и структурное биоразнообразие на особо охраняемых и нарушенных территориях Бекмансуров М. В., Мустаева Н. В. О фиторазнообразии лугов Марийского Присурья ...................................................................................................................................88 Бекмансуров М. В., Рыбакова М. А. Характеристика лесного покрова Марийского Присурья .............................................................................................................91 Беляченко А. А. Хозяйственная деятельность на пруду Третий (ГПЗ «Саратовский») и ее влияние на состав гнездового населения птиц .........................95 Гетманец И. А., Калашников Н. В. Экологическая оценка некоторых растительных сообществ Челябинского (городского) бора ..................................................98 Гетманец И. А., Байкова И. В. Таксономическое разнообразие и экотипы гигрогелиофитов озера Увильды ...........................................................................................101 Голованов Я. М. Флора города Мелеуза (Республика Башкортостан) ........................ 104 Голованов Я. М., Крюкова А. В., Абрамова Л. М. К оценке фитоценотической приуроченности редкого вида Iris pumila L. в Башкирском Предуралье .............................................................................................................................107 Гордеева В. А., Жуйкова Т. В., Ившина И. Б., Костина Л. В., Безель В. С. Разнообразие и численность бактерий цикла азота в почвах техногенно нарушенных территорий ........................................................................................................110 Грозовская И. С., Ханина Л. Г., Бобровский М. В., Смирнов В. Э., Романов М. С. Биомасса пространственных микрогруппировок видов растений напочвенного покрова в темнохвойных лесах Костромской области ................................ 114 Дробот В. И. Фауна позвоночных животных памятников природы Республики Марий Эл «Болото Железное» и «Болото Березовое» ................................... 118 Жукова О. В. Ромашка душистая (Chamomilla suaveolens (Pursh) Rydb.) в некоторых местообитаниях на территории Республики Марий Эл ................................. 123 Закамская Е. С., Зотина С. Г. Изменение биоразнообразия растений в ходе сукцессионных процессов на залежах ..................................................................................124 Клименко А. А. Фрагментация популяций редких видов растений как форма их деградации .........................................................................................................................127 Макеев И. С., Гаврилко Д. Е. Видовое разнообразие зоопланктона как показатель экологического состояния малых водотоков Нижнего Новгорода ................ 131 Мартынов А. В., Фокина М. Е. Особенности распространения и учет по норам барсука обыкновенного (Meles мeles Brisson, 1762) на территории Жигулевского государственного заповедника .....................................................................134 321 Мударисов Р. Г. Состояние зеленых насаждений садово-парковых территорий г. Казани ..............................................................................................................138 Мударисов Р. Г., Леонова Т. Ш., Мирсаитов Н. Г. Особенность экологии Passer domesticus L., 1758 и Passer montanus L., 1758 садово-парковых территорий города Казани .....................................................................................................140 Нотов А. А. Состояние индикаторного компонента коренных лесных фитоценозов как показатель природоохранной ценности экосистем ............................... 144 Полянская Т. А. Онтогенетическая структура ценопопуляций растений бореальной эколого-ценотической группы ..........................................................................150 Свинин А. О., Гаранин В. И. Распространение и численность амфибий и рептилий антропогенных ландшафтов города Йошкар-Олы ........................................... 153 Сидоренко М. В., Юнина В. П. Структурное разнообразие экосистем проектируемой зоны затопления Чебоксарской ГЭС (до 68 отметки)................................ 157 Сидоренко М. В. Таксономическое разнообразие орхидных на особо охраняемых территориях и в водоохранных зонах Нижегородской области .................... 161 Темнухин В. Б. Валежный комплекс как фактор сохранения биоразнообразия на городских ООПТ Нижнего Новгорода ............................................................................163 Турмухаметова Н. В. Разнообразие членистоногих консортов некоторых лиственных деревьев в условиях урбанизированной среды ............................................... 164 Утенкова С. Н. К вопросу о флористическом разнообразии района Пятигорья, как части особо охраняемого эколого-курортного региона РФ «Кавказские минеральные воды» ..........................................................................................167 Фархуллин Р. Ш. Динамика изменения структуры лесных насаждений и фитомассы Лубянского участкового лесничества за 1940–2007 годы ............................ 170 Цепкова Н. Л., Тубаева М. А. Эдемики национального парка «Приэльбрусье» (Центральный Кавказ) ............................................................................................................173 Яковлев В. А., Яковлева А. В. Редкие и охраняемые виды беспозвоночных в бассейне р. Цивиль (Чувашская Республика) .................................................................... 176 Секция 3. Мониторинг абиотических и биотических компонентов экосистем Бухарина И. Л., Кузьмин П. А. Динамика содержания аскорбиновой кислоты в листьях древесных видов растений в насаждениях с различной техногенной нагрузкой ................................................................................................................................179 Глушко С. Г. Особенности лесорастительного районирования Среднего Поволжья .................................................................................................................................182 Дементьева С. М., Нотов А. А., Мейсурова А. Ф., Иванова С. А., Зуева Л. В. Основные компоненты региональных моделей комплексного биомониторинга ............. 185 Дробот Г. П., Арсланова А. А., Ведерников А. А. Использование гистоморфометрических показателей печени и миокарда рептилии для оценки качества наземных экосистем ...............................................................................................................190 Иванова Р. Р., Малкова Т., Куфтина Н. И., Глаголев Д. Мониторинг содержания нефтепродуктов в почве разных зон г. Йошкар-Олы .................................... 193 Иванова Р. Р., Токарева Н. А. Оценка воздействия АЗС на состояние окружающей среды в г. Йошкар-Оле ...................................................................................197 Поролло А. А., Кушион М. Т. Сравнительный генетический анализ рода пневмоцистов с точки зрения метаболических путей ......................................................... 200 322 Сарбаева Е. В., Воскресенский В. С. Использование древесных растений в локальном мониторинге состояния окружающей среды г. Йошкар-Олы ........................ 206 Соловьева О. С., Кудрявцева А. Е., Васильева В. К. Состояние насаждений на территориях общего пользования города Йошкар-Олы ................................................ 209 Шынбергенов Е. А., Сиханова Н. С., Нургизаринов А. М. Влияние колебания уровня северного малого арала на экологическую обстановку в регионе ......................... 214 Юпина Г. А., Никитин А. В., Мингазова Н. М. Разнообразие и оценка состояния зеленых насаждений г. Казани по древесной растительности .......................... 217 Секция 4. Популяционное биоразнообразие и устойчивость популяций Агаева П. Н., Османова Г. О. Онтогенетическая структура ценопопуляций посконника конопляного (Eupatoria cannabinum L.) ............................................................ 221 Анциферов А. Л., Полежаева А. Ю. Фаунистическая структура почвообитающих жесткокрылых (Coleoptera) поймы малых рек Костромской области ....................................................................................................................................224 Ахметова А. Ш., Зарипова А. А. Размножение видов рода Hedysarum L. in vitro ......................................................................................................................................227 Бекмансуров М. В., Соловьева М. Н. Онтогенетическая и виталитетная структура ценопопуляций ясеня обыкновенного на границе ареала ................................. 230 Ведерникова О. П., Халиуллина Л. Р. Эколого-морфологические особенности особей и онтогенетическая структура ценопопуляций кровохлебки лекарственной (Sanguisorba officinalis L.) ............................................................................233 Ведерникова О. П., Ягодарова О. С. Онтогенетическая структура ценопопуляций Tanacetum vulgarе L. ..................................................................................237 Гасанова М. Ю., Алиева С. А., Гулиев Ф. Б. Биоэкологические особенности, размножение и значение ксантоцераса рябинолистного (чекалкин орех) – Xanthoceras sorbifolia Bunge на Апшеронском полуострове Азербайджана ..................... 240 Гусева А. А. Онтогенетическая структура Scutellaria galericulata L. ......................... 243 Дадашева А. Г., Абдыева Р. Т. Влияние экологических факторов на биоразнообразие флоры Илисуинского государственного природного заповедника Азербайджана ...................................................................................................245 Ефименко А. С., Алейников А. А. Влияние жизненности на морфометрические и продукционные характеристики подроста Picea obovata Ledeb. ......................................................................................................................................249 Забродин И. В. Состояние смешанных лесных посадок сосны обыкновенной (Pinus sylvestris L.) в национальном парке «Марий Чодра» ............................................... 255 Злобин Ю. А. Биоразнообразие: количественный и качественный подход .............. 257 Иванова Н. В. Онтогенетическая структура субпопуляций лишайника (Lobaria pulmonaria) в лесах северо-востока Костромской области ................................. 262 Ильина В. Н. Структура и динамика популяций некоторых кальцефитов Средней Волги ........................................................................................................................266 Карнаухова Н. А. Популяционное биоразнообразие и оценка состояния популяций южносибирских видов рода Hedysarum L. (Fabaceae) ..................................... 269 Колегова Е. Б. Онтогенетическая структура ценопопуляций Thymus marschallianus Willd. ..............................................................................................................272 323 Криницын И. Г., Зонтиков Д. Н., Лебедев В. П., Латышева Д. Ю., Разгуляева Н. А., Мастерова А. В. Популяционная морфология спорофита Botrychiaceae Nakai и ее влияние на стуктуру популяций .................................................. 274 Кучер Е. Н., Чмелева С. И., Решетник Г. В. Аутэкологические особенности пластичности морфометрических параметров в популяциях Dactylorhiza romana (Seb. et mauri) Soo (Orchidaceae) ............................................................................................277 Мустафаева С. Д., Бахшалиева К. Ф. Антимикотические свойства некоторых видов Anthemis L. флоры Азербайджана .......................................................... 280 Невский С. А., Давиденко О. Н. К вопросу оценки функциональной устойчивости сообществ с участием редких видов растений ............................................ 284 Нурминская Ю. В., Максимова Л. А., Копытина Т. В., Еникеев А. Г. Влияние генетической трансформации на эффективность работы механизмов адаптации у растений табака .................................................................................................287 Османова Г. О., Чернова С. Ю. Онтогенетическая структура ценопопуляций пыльцеголовника красного (Cephalanthera rubra (L.) Rich.) ............................................. 290 Ребко С. В., Гаврилик О. П., Гарбар Ю. И., Горошко А. И. Оценка устойчивости к энтомовредителям наземных органов гибридного потомства сосны обыкновенной (Pinus sylvestris L.) в испытательных культурах ............................. 295 Ребко С. В., Гаврилик О. П., Гарбар Ю. И., Горошко А. И. Оценка устойчивости гибридного потомства сосны обыкновенной (Pinus sylvestris L.) к пластинчатоусым-ризофагам на землях, выведенных из сельскохозяйственного оборота ....................................................................................................................................299 Тишечкин А. Н. Закономерности изменчивости у экотипов сосны обыкновенной в географических культурах на Среднем Урале ........................................ 303 Челтыгмашева Л. Р. Структура и динамика ценопопуляций Lilium pumilum Delile ........................................................................................................................................307 Широков А. И., Крюков Л. А., Сырова В. В., Штаркман Н. Н. Репатриация и искусственное создание популяций тубероидных орхидных как один из методов поддержания их численности ................................................................................................309 Щербакова О. Ф., Калистая М. С. Структурно-функциональное зонирование многолетних полурозеточных цветоносных побегов монокарпиков ........... 312 Авторский указатель .......................................................................................................318 Тем. план 2013 г. № 156. Подписано в печать 25.11.2013 г. Формат 60×84/16. Усл. печ. л. 18,83. Уч.-изд. л. 13,70. Тираж 500. Заказ № 2591. Оригинал-макет подготовлен к печати в РИЦ и отпечатан в ООП ФГБОУ ВПО «Марийский государственный университет» 424000 г. Йошкар-Ола, пл. Ленина, 1. 324