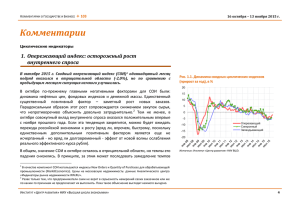
ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ «ДАЛЬНЕВОСТОЧНЫЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ»
advertisement

ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ «ДАЛЬНЕВОСТОЧНЫЙ ГОСУДАРСТВЕННЫЙ АГРАРНЫЙ УНИВЕРСИТЕТ» На правах рукописи ТИТОВА СВЕТЛАНА АНАТОЛЬЕВНА Влияние фитопатогенных микроорганизмов на энзиматическую активность растения-хозяина Glycine max (L.) Merr. и Glycine soja Sieb. et Zucc. 03.02.08 - ЭКОЛОГИЯ Диссертация на соискание ученой степени кандидата биологических наук Научный руководитель: к.б.н., доцент Семенова Е.А. БЛАГОВЕЩЕНСК – 2014 1 Оглавление Введение………………………………………………………………………... 4 1 Аналитический обзор литературы……………………………………… 8 1.1 Вредоносность и распространение болезней сои………………………... 8 1.2 Влияние экологических факторов на развитие инфекции сои………………………………………………………………………………. 27 1.3 Ферменты инфицированных растений…………………………………… 32 2 Условия, объекты и методика проведения исследований ……………. 44 2.1 Агроклиматические и почвенные условия Амурской области ………… 44 2.2 Материалы и методы исследований…………………………………… 53 3 Энзиматическая активность в корнях и листьях сои при поражении почвенными (корневыми) инфекциями……………...…………………… 61 3.1 Видовой состав возбудителей корневой гнили сои и динамика развития болезни………………………………………………………………. 61 3.2 Изменение активности пероксидазы, каталазы и кислой фосфатазы в корнях сои при поражении возбудителями корневой гнили...……………… 66 3.3 Изменение активности пероксидазы, каталазы и кислой фосфатазы в листьях сои при поражении возбудителями корневой гнили ………………. 72 4 Энзиматическая активность в листьях сои при поражении листостеблевыми инфекциями………………………………..…………………... 75 4.1 Динамика развития грибных и бактериальных заболеваний и оценка болезнеустойчивости сои……………………………………………………… 75 4.2 Участие ферментов в ответных реакциях сои на поражение листостеблевыми инфекциями……………………………….……………………… 84 4.2.1 Изменение активности пероксидазы, каталазы и кислой фосфатазы в листьях сои при поражении грибной и бактериальной инфекциями в естественных условиях………………………………………………………… 4.2.2 Изменение активности пероксидазы, каталазы и кислой фосфатазы в 2 85 изолированных листьях сои при искусственном заражении септориозом……………………………………………………………………. 98 5 Энзиматическая активность в семенах сои при поражении фитопатогенами.…………………………........................................................ 105 5.1 Влияние зараженности семян сои болезнями на их биохимический состав и продуктивность растений……………………………………………. 105 5.2 Изменение активности каталазы, пероксидазы и кислой фосфатазы в семенах сои при поражении грибной и бактериальной инфекциями………. 111 Выводы…………………………………………………………………………. 117 Практические рекомендации………………………………………………. 118 Список литературы…………………………………………………………... 119 Приложения…………………………………………………………………… 148 3 Введение Актуальность темы. Растения живут в постоянно изменяющихся условиях внешней среды, подвергаются действию различных факторов абиотической и биотической природы. Им приходится адаптироваться к этим факторам и формировать механизмы противодействия их негативному влиянию. Одними из самых серьезных и универсальных биотических воздействий, которым подвергаются растения, являются патогенные микроорганизмы. Это могут быть вирусы, бактерии, простейшие и грибы (Чесноков, 2007). Соя относится к числу главнейших культурных растений мирового значения. Универсальное значение сои объясняет резко возрастающий интерес к выращиванию этого белково-масличного растения. Основным соесеющим регионом на Дальнем Востоке является Амурская область, где сосредоточено 60% её посевов. Несмотря на сравнительно благоприятные почвенно-климатические условия для возделывания сои, средняя урожайность остается крайне низкой и составляет в среднем 11 ц/га (Прогноз…, 2011; Посевные…, 2012). Урожаи сои в значительной степени зависят от влияния целого ряда различных факторов, в том числе поражения болезнями и вредителями. Особенно вредоносны воздушно-капельно-семенные инфекции: пероноспороз, септориоз, церкоспороз, бактериоз, которые освоили в качестве дополнительной экологической ниши семена и в связи с этим приспособились к передаче инфекций из года в год (Заостровных, Дубовицкая, 2003). Широкое использование пестицидов в борьбе с вредителями и болезнями растений оказывает негативное воздействие не только на патогенного возбудителя, но и на растение-хозяина и окружающую среду. Наиболее экономически выгодным и экологически безопасным методом борьбы с заболеваниями является возделывание устойчивых сортов, что позволит существенно улучшить экологическую ситуацию. 4 Изучение биохимических особенностей инфицированного растения представляет важный этап на пути познания природы устойчивости к патогенным микроорганизмам. Взаимодействие возбудителя и хозяина вызывает значительные изменения в функциональном состоянии растений, что прежде всего отражается на уровне ферментативной активности (Савич, 1989). О влиянии фитопатогенов на изменение активности ферментов сообщается в целом ряде работ 60 – 90-х годов прошлого столетия (Farkas et al, 1960, 1966; Kawashima et al., 1963, 1964; Рубин и др. 1965, 1966, 1969, 1970, 1973, 1974, 1975; Коно, 1961; Гоцуляк, Кондратюк, 1967; Grzelinska, 1970; Naoi, Teruniko, 1977; Андреева, 1973, 1979, 1988; Газарян, 1992). В последнее время большое внимание уделяется изучению роли пероксидазы в формировании устойчивости к возбудителям болезней (Минибаева, Гордон, 2003; Максимов и др., 2006, 2010; Граскова, 2008), однако недостаточно данных об участии каталазы и кислой фосфатазы в защитных реакциях растений, отсутствуют сведения о влиянии патогенных микроорганизмов на активность ферментов в онтогенезе сои. Поэтому данное исследование, направленное на изучение ферментов в системе «растение-хозяин – патоген» как возможных биохимических факторов устойчивости, является актуальным и имеет большое научное и практическое значение. Цель работы: исследовать влияние фитопатогенных микроорганизмов на энзиматическую активность растения-хозяина Glycine max (L.) Merr. и G. soja Sieb. et Zucc., сопоставить изменения этого показателя с устойчивостью к ряду грибных и бактериальных заболеваний. Для достижения поставленной цели необходимо было решить следующие задачи: 1. Изучить динамику развития грибных и бактериальных заболеваний, оценить болезнеустойчивость G. max и G. soja. 2. Определить удельную активность пероксидазы, каталазы и кислой фосфатазы в корнях и листьях сои при инфицировании возбудителями корневой гнили. 5 3. Выявить постинфикционные изменения удельной активности пероксидазы, каталазы и кислой фосфатазы в листьях G. max и G. soja при поражении листо-стеблевыми болезнями в естественных условиях. 4. Установить динамику удельной активности пероксидазы, каталазы и кислой фосфатазы в изолированных листьях G. max и G. soja при искусственном заражении S. glycines. 5. Определить биохимические показатели семян сои при поражении фитопатогенами. Научная новизна. Впервые проведено многостороннее исследование изменения активности ферментов в растениях G. max и G. soja под влиянием грибной и бактериальной инфекций. Установлено, что устойчивость сои к фитопатогенам (S. glycines, P. manshurica, Ps. glycineum) не зависит от изначальной активности пероксидазы, каталазы и кислой фосфатазы в листьях сои, а связана с изменением активности ферментов в ответ на внедрение патогена. Определена динамика активности ферментов в изолированных листьях сои при искусственном инфицировании септориозом. Выявлено, что активность пероксидазы и каталазы на начальном этапе, а кислой фосфатазы на поздних сроках развития заболевания может служить критерием устойчивости к S. glycines. Практическая значимость. Полученные результаты и сделанные на их основе выводы расширяют представление о защитных механизмах растений к действию фитопатогенов и могут быть использованы в практической селекции на устойчивость, позволяющей снизить влияние биотических стрессоров на агроценоз. Результаты исследования могут быть использованы как в лекционных курсах, так и практикумах по сельскохозяйственной экологии и защите растений. Основные положения, выносимые на защиту: – постинфекционные изменения активности ферментов определяются генотипом, фазой развития сои и видом возбудителя; 6 – устойчивость сои к фитопатогенам не зависит от изначальной активности пероксидазы, каталазы и кислой фосфатазы в листьях сои, а связана с изменением активности ферментов в ответ на внедрение патогена; – активность ферментов можно использовать в тестовых системах для селекционного отбора устойчивых сортов сои. Апробация работы. Результаты исследования докладывались на научных конференциях ФГБОУ ВПО ДальГАУ (г. Благовещенск, 2009, 2010, 2012 гг.); на международной научно-практической конференции «Аграрные проблемы соесеющих территорий Азиатско-тихоокеанского региона», на координационном совещании зоны Дальнего Востока и Сибири «Итоги координации НИР по сое за 2006-2010 гг. и направления исследований на 2012-2015 гг.» в ГНУ ВНИИ сои Россельхозакадемии (г. Благовещенск, 2010, 2012 гг.); на II-м «Амурском региональном фестивале науки» (г. Благовещенск, 2012 г.). Связь с планами НИР. Диссертационная работа является частью тематического плана ФГБОУ ВПО ДальГАУ, тема 2.3 «Биохимическая адаптация культурной и дикой сои», № госрегистрации 0120.0503579. Публикации. По материалам диссертации опубликовано 8 печатных работ, в том числе 3 статьи в журналах, рекомендованных ВАК РФ. Объем и структура работы. Диссертация состоит из введения, 5 глав, выводов, списка литературы, 35 приложений. Работа изложена на 185 страницах машинописного текста, иллюстрирована 47 рисунками и 19 таблицами. Список цитируемой литературы включает 293 источник, в том числе 81 на иностранных языках. Благодарности. Автор выражает благодарность научному руководителю к.б.н., доценту Е.А. Семеновой за руководство и помощь при подготовке и написании диссертационной работы; к.с.-х.н., профессору Л.К. Дубовицкой за помощь и консультации при проведении экспериментальной части работы; ректору ФГБОУ ВПО ДальГАУ, д.с.-х.н., профессору П.В. Тихончуку; декану факультета агрономии и экологии, к.с.-х.н., доценту О.А. Селиховой. 7 1 АНАЛИТИЧЕСКИЙ ОБЗОР ЛИТЕРАТУРЫ 1.1 Вредоносность и распространение болезней сои Эпифитотии болезней растений наносили и наносят значительный ущерб сельскому хозяйству, поэтому специалисты выясняют видовой состав возбудителей потенциально опасных инфекций, которые при тех или иных условиях способны снижать урожайность и качество сельскохозяйственной продукции. В связи с этим оценка вредоносности болезней сои для современных сортов приобретает важное значение (Торопова, 2005). Анализируя литературные источники, следует отметить, что большой интерес к изучению болезней сои проявляли прежде всего зарубежные исследователи (Wolf, Lehman, 1924, 1926; Boosalis, 1950; Murakishi, 1951; Tiffany, Gilman, 1954; Macneil, Zalasky, 1957; Kurata, 1960; French, Kennedy, 1963; Ross, 1965, 1982; Mckenzie, Wyllie, 1971; Riggle, Dunleavy, 1974; Riggle, 1977). Начало изучению болезней сои в России было положено А.А. Ячевским, он описал примерно 35 потенциально опасных фитопатогенов, однако степень их изученности была различной (Заостровных, Дубовицкая, 2003). Затем этим вопросом занимались на Северном Кавказе А.И. Лобик и О.В. Катаева, на Дальнем Востоке – И.Н. Абрамов (1931, 1938), в Московской области – С.В. Владимирский, на Украине – М.С. Заянчковская (Заостровных, Дубовицкая, 2003). Диагностические признаки в начале XX века были определены по 2820 болезням (Хохряков и др., 1966). Распространённость, вредоносность патогенов были обобщены М.В. Горленко (1968) по 350 болезням, В.Ф. Пересыпкиным (1989а) – примерно по 400. Впоследствии изучение болезней было проведено в основной зоне соесения – на Дальнем Востоке. Изучали широту распространения заболеваний, видовой состав и биологию возбудителя, а также проводили разработку агротехнических и химических мер борьбы (Иванова, Холопова, 1956; Корецкий, 1967). 8 Изучением болезней сои в дальневосточном регионе в разные годы занимались или продолжают работать: И.Н. Абрамов (1931, 1938), А.Н. Иванова (1956), А.И. Никитина (1962), А.М. Михайленко (1965), М.Ф. Муравьёва (1967, 1969, 1971, 1975, 1981), А.М. Корецкий (1967), Е.С. Нелен (1968), В.Г. Рейфман (1968), С.А. Жуковская (1968, 1977, 1980, 1982, 1990), А.М. Гунина (1968, 1978, 1983), А.М. Овчинникова (1968, 1970, 1971а, 1971б, 1980), Т.А. Поливанова, (1968, 1971, 1980), В.И. Заостровных (2003), Л.К. Дубовицкая (1984, 1987, 2002, 2003), Т.И. Лукьянович (2006), Л.А. Дега (2001, 2012). В 70-х – 80-х годах, в связи с расширением посевов сои в различных зонах страны, возобновились исследования по изучению болезней этой культуры в европейской её части. Появились работы, посвящённые корневым гнилям в Абхазии (Вардания, 1971, 1974) и переноспорозу сои (Простакова, 1981) – в Молдавии; фузариозу – в Краснодарском крае (Дворядкин и др., 1979). Подробно изучены грибные болезни сои в Молдавии А.И. Ганей в 1981 году. Установлено, что наиболее распространёнными на сое являются пероноспороз и вирусные заболевания: морщинистая мозаика, желтая мозаика, кольцевая пятнистость, задержка роста и другие (Поливанова, 1971, 1980; Дьяконов, Голаюда, 1971). Изучено их влияние на показатели симбиотической и фотосинтетической активности посевов, урожай семян и его качество. Под руководством профессора Г.И. Посыпанова в условиях Центрального Нечерноземья впервые (1991 – 1995 гг.) были изучены видовой состав и вредоносность основных патогенов сои (Посыпанов, Сихарулидзе, Демьяненко, 1995). Изучение этиологии наиболее распространенных и вредоносных болезней показало, что подавляющее их большинство (67,2-74,9%) составляют грибные болезни, 10,0-13,2% – бактериальные, 12,6-19,2% –вирусные (Торопова, 2005). В Амурской области, Хабаровском и Приморском краях широко распространены грибные заболевания сои, а также бактериозы, несколько видов вирусных заболеваний и многие другие (Овчинникова, 1980; Заостровных, 9 Дубовицкая, 2003). Из грибных заболеваний в дальневосточном регионе выявлено свыше 30 видов возбудителей. Доминирующими патогенами сои являются возбудители пероноспороза, септориоза, церкоспороза, аскохитоза, фузариозных корневых гнилей и бактериальных заболеваний. В Приамурье наиболее распространены следующие болезни сои: корневая гниль, фузариоз, септориоз, аскохитоз, бактериоз (Рейфман, Поливанова, 1968; Гунина, 1968; Солотчина, 1971; Дубовицкая, Гунина, 1979, 1983). Широкое распространение во всех странах, возделывающих сою, имеет вирус мозаики сои. На Дальнем Востоке первые вирозные растения обнаружены И.Н. Абрамовым в 1931 году, затем их изучала М.Ф. Муравьёва (1967, 1969, 1971). Дальнейшее изучение и описание вирусных болезней сои на Дальнем Востоке проведено Т.А. Поливановой (1971; 1980), а также в соавторстве с А.В. Крыловым (1985). По их сведениям, вирусные заболевания сои могут вызываться более чем 20 возбудителями. Наибольшее распространение и вредоносность вирусных болезней наблюдается при зарожении посевов мозаикой (вирус мозаики сои). В результате исследований А.М. Гуниной (1978) установлено, что в Амурской области наибольший экономический ущерб посевам сои наносят фузариоз, белая гниль или склеротиниоз, септориоз, бактериальные заболевания и вирусная мозаика (вирус жёлтой мозаики фасоли и вирус мозаики люцерны). Грибные болезни. Урожаи сои в значительной степени зависят от влияния целого ряда различных факторов, из них особое значение имеют грибные болезни. Всего в природе существует не менее 10000 видов фитопатогенных грибов. Это наиболее многочисленная группа вредоносных организмов. Среди бактерий возбудителями болезней растений являются лишь 120-200 видов (Метлицкий, Озерецковская, 1985). Первые данные о распространении на сое 35 видов фитопатогенных грибов приводил А.А. Ячевский в 1929 г. (Заостровных, Дубовицкая, 2003). Позже И.Н. Абрамов (1931) сообщал о грибных болезнях сои на Дальнем Востоке. 10 Наиболее полный перечень грибных болезней сои, распространенных в Приморском, Хабаровском краях и Амурской области, приводит A.M Овчинникова (1968). В Московской области микрофлора сои изучалась С.В. Владимирским. Автор выделил 8 видов грибов из рода Fusarium, возбудителей филлостиктоза, аскохитоза, пероноспороза и фомопсиса сои (Заостровных, Дубовицкая, 2003). В европейской части страны первые сообщения о грибных болезнях сои появились в 1930 г. На Северном Кавказе А.И. Лобик и О.Е. Катаева сообщали о паразитировании на сое Peronospora manchyrica (Naum.) Syd., Septoria glycines Hemmi, Sclerotinia sderotiorum (Lib.), Phomopsis sojae Lehman и приводили список грибных патогенов, выделенных из семян (Заостровных, Дубовицкая, 2003). К наиболее вредоносным грибным заболеваниям сои относятся: фузариоз, аскохитоз, пероноспороз, церкоспороз, белая гниль, южная склероциальная гниль, септориоз (Козырева, Примаковская, Скрипка, 1982; Жуковская, Серебренникова, Куликова, 1990; Миронова, 1995). Септориоз является одним из самых вредоносных заболеваний сои на Дальнем Востоке (рис. 1). В Приморском крае, по сведениям В.И. Заостровных, Л.К. Дубовицкой (2003), распространённость септориоза на семядолях и листьях составила от 40 до 100% при степени поражения 39,2-59,2% Возбудитель септориоза – несовершенный гриб Septoria glycines T. Hemmi. Патоген характеризуется широкой органотропностью и легко переходит с одних органов на другие. Он поражает практически все надземные части растения: семядоли, листья, стебли, бобы. Заболевание у сои начинается очень рано и продолжается в течение всего вегетационного периода (Дега, 2012). Возбудитель септориоза впервые был описан японским профессором Takewo Hemmi в 1915 году в Японии (Hemmi, 1940). Затем он был обнаружен в США (Wolf, Lehman, 1926), в Канаде (Macneil, Zalasky, 1957), Китае (Сунь Синдун, 1958) и др. Заболевание также распространено в Бразилии, Италии, КНДР, Югославии и других странах (Заостровных, Дубовицкая, 2003). 11 Сильное распространение септориоза отмечалось на сое в провинции Киото и на севере Японии (Hemmi, 1940). Новая для Канады листовая пятнистость, вызываемая грибом Septoria glycines T. Hemmi, появилась здесь в 1943 году. В России септориоз первоначально был отмечен в виде пятнистости на листьях А.А. Ячевским в 1929 году. Затем заболевание обнаружено на Северном Кавказе (Заостровных, Дубовицкая, 2003). В 70 – 80-е годы отмечалось поражение этим заболеванием семядолей, листьев, стеблей и бобов (Гунина, 1968; Фалеева, Ала, 1976; Гунина, 1978; Корсаков, Овчиникова, Мизева, 1979; Жуковская, Овчинникова, 1980; Муравьёва, 1981; Простакова, Ганя, 1983). Возбудитель поражает все органы, кроме корневой системы. Вредоносность септориоза проявляется в преждевременном пожелтении и опадании поражённых листьев (Wolf, Lehman, 1926; Hemmi, 1940; Овчинникова, 1971; Муравьёва 1981), а также в снижении урожайности на 12-24% (Lim, 1980). В период налива бобов может опадать до 50% поражённых септориозом листьев (Возбудители…, 1980). Рисунок 1 – Септориоз тройчатого листа (фото С.А. Титовой) 12 Особенно вредоносно это заболевание на Дальнем Востоке, где поражает в значительной степени все районированные сорта. Ещё в 1976 году Л.К. Фалеевой отмечалось, что септориоз на листьях в условиях Амурской области достигал развития от 25 до 75% (Фалеева, Ала, 1976). А.М. Гунина также считает, что наибольший экономический ущерб в Амурской области по сравнению с другими болезнями сои приносит септориоз (Гунина, 1978). При заражении в фазе образования бобов масса 1000 семян уменьшалась на 50,6 г, или на 29%. Кроме того, у поражённых септориозом растений наблюдалось снижение качества семян. Наибольшее снижение содержания белка и масла, на 5,82 и 0,76% соотвественно, наблюдалось при инокуляции в фазу образования бобов. При заражении в фазе цветения наиболее сильно снижалась масса 1000 семян, а при заражении в более поздние сроки – качество семян (Заостровных, Дубовицкая, 2003). Пероноспороз, или ложная мучнистая роса (возбудитель Perenospora manshurica (Naum.) Syd. Syn.) – одно из наиболее распространённых и вредоносных заболеваний сои (Овчинникова, 1971а, 1971б; Sinclair, Shurtleff, 1975) (рис. 2). Болезнь широко распространена в большинстве производящих сою стран. В Приморье эпифитотии его возникают практически ежегодно. Количество поражённых семян в посевном материале часто составляет 20-30%. Степень поражения растений достигает 50-75% (Овчинникова, Потлайчук, 1980). В нашей стране пероноспороз впервые был обнаружен в 1914 г. на Дальнем Востоке, к настоящему времени известен в Московской, Воронежской областях, Краснодарском, Ставропольском краях, в Украине, в Грузии, Средней Азии (Корсаков, Овчинникова, Мизева,1979). Позднее это заболевание получило повсеместное распространение в Европе (Дания, Франция, Италия, Польша), Америке (США, Канада, Бразилия), Азии и в некоторых странах Африки. О распространении этого заболевания сообщают многие исследователи: И.Н. Абрамов (1931, 1938), А.М. Михайленко (1965), П.М. Корецкий (1967), А.М. Овчинникова (1970) – на Дальнем Востоке. 13 Поражение растений септориозом может проявляться в двух формах – общей (диффузной) и местной (локальной). На Дальнем Востоке И.Н. Абрамовым (1931, 1938) и другими отмечалось поражение листьев в локальной и диффузной формах. При диффузном поражении наблюдается опадание листьев и засыхание всего растения, при локальной форме пероноспороза происходят лишь постепенная некротизация и разрыв тканей. При локальном поражении листьев масса 1000 семян снижается до 50% (Заостровных, Дубовицкая, 2003). Пероноспороз также поражает бобы и семена (Корецкий, 1967; Овчинникова, 1971а; Жуковская, Серебренникова, Куликова, 1990). В России это заболевание на семенах впервые обнаружено О.Е. Катаевой на Северном Кавказе (Заостровных, Дубовицкая, 2003). Рисунок 2 – Пероноспороз тройчатого листа (фото С.А. Титовой) По данным П.М. Корецкого (1967), A.M. Овчинниковой (1968), З.И. Родиной (1981), основная вредоносность пероноспороза заключается в ухудшении посевных качеств семян сои. Пероноспороз снижает всхожесть семян 14 до 30%. Масса 1000 семян уменьшается на 6%, при посеве таких семян 8% всходов уже имеют зараженные семядоли, и 5-10% растений в дальнейшем поражаются диффузной формой этого заболевания. В связи с поражением бобов происходит снижение качества семян, содержание масла в них уменьшается на 1,07%. Фузариозы - вызывают несовершенные грибы рода Fusarium. В своих работах авторы большое значение придавали фузариозам всходов. Было установлено, что гибель всходов сои происходит при участии нескольких представителей грибов из рода Fusarium, при этом могут поражаться семядоли, точка роста, подсемядольный стебелёк и корень (рис. 3). Рисунок 3 – Фузариоз семядолей (фото С.А. Титовой) При сильном поражении происходит гибель всходов и взрослых растений. Фузариозные растения отстают в росте и развитии. Больные фузариозом семена щуплые, деформированные, тусклые, они теряют всхожесть или дают поражённые всходы. Сильно поражённые семена загнивают, покрываются мицелием и не прорастают (Возбудители…, 1980). 15 По сведениям авторов (Овчинникова, 1971; Жуковская, Овчинникова, 1980; Простакова, Ганя, 1983), всходы сои являются одной из наиболее уязвимых фаз поражения фузариозом. Поражённые всходы плохо растут, их листья желтеют, появляется гниль корешков, перетяжка стебля, поражённая ткань буреет и всходы погибают. Источники инфекции – поражённые растительные остатки в почве, а также семенной материал (Яковлева, 1992). Аскохитоз – (Ascochyta sojaecola Abramoff), возбудитель болезни относится к порядку Sphaeropsidales, классу Coellomycetes (Возбудители…, 1980) (рис. 4). На Дальнем Востоке аскохитоз на сое впервые обнаружен фитопатологом И.Н. Абрамовым в 1931 г. В своих работах А.М. Овчинникова (1971а, 1971б) отмечает значительное распространение аскохитоза в Амурской области и Приморском крае и менее интенсивное – в Хабаровском крае. Аскохитоз встречается также в ряде стран Западной Европы, в Украине, в Московской области (Заостровных, Дубовицкая, 2003). Рисунок 4 – Аскохитоз на тройчатых листьях (фото Л.А. Дега) По данным С.А. Жуковской, Н.И. Серебренниковой, Л.С. Куликовой (1990), патоген поражает все органы растений. В условиях Амурской области в большей 16 степени развивается на стеблях, семенах и в меньшей – на корнях. Наибольшая вредоносность аскохитоза наблюдается в Амурской области, где количество пострадавших всходов в отдельные годы достигает 20% (Дега, 2012). Как и при поражении пероноспорозом, наиболее опасна семенная форма аскохитоза. Проявляется он довольно рано – в момент появления семядолей и поражает последовательно все части растения – корень, семядоли, листья, стебли, ветви, черешки, бобы и семена (Дубовицкая, Кравцова, 2002). Сильно заражённые семена загнивают и теряют всхожесть, слабо заражённые – дают всходы с пораженными семядолями. Церкоспороз, или округлая серая пятнистость (рис. 5). Возбудитель болезни – гриб Cercospora sojina Hara. Заболевание отмечено во всех районах соесения Дальнего Востока. Наиболее широко оно распространено в Приморском крае. Семена районированных сортов в годы возникновения эпифитотий поражаются на 20% и более (Овчинникова, Потлайчук, 1980). Первоначально церкоспороз на листьях был описан в США (Wolf, Lehman, 1926). Подробное описание симптомов поражения всех органов сои этим заболеванием было проведено несколько позднее (Sherwin, Kreitlow, 1952; Сун Син-дун, 1958). В России церкоспороз в виде пятнистости листьев был отмечен А.А. Ячевским в 1929 году (Заостровных, Дубовицкая, 2003); на Дальнем Востоке – И.Н. Абрамовым (1931;1938); в европейской части страны – Н.А. Наумовым (1936), Н.И. Василевским и Б.П. Каракулиным (1937). С.В. Владимирским (1939). Гриб поражает все надземные органы сои: семядоли, листья, стебли, бобы и семена. В Амурской области наблюдалось поражение церкоспорозом листьев, стеблей и бобов сои, на семенах заболевание встречалось крайне редко (Гунина, 1968). В 1972-1973 годах при оценке на устойчивость к болезням генетического материала культурной и дикорастущей сои церкоспороз отсутствовал (Фалеева, Ала, 1976; Гунина 1978). Установлено, что наибольшую вредоносность представляет семенная инфекция церкоспороза, в связи с чем всхожесть заражённых семян снижается на 17 12-15%, в результате этого урожайность уменьшается в 3-6 раз. Семена с различной степенью заражения вызывают различную заболеваемость всходов сои: при посеве слабо заражённых семян развиваются 43-82% поражённых всходов, средне и сильно заражённых – соответственно 67-97 и 77-95%. В дальнейшем из таких всходов развиваются больные растения, на которых формируются неполноценные семена. Масса 1000 заражённых семян снижается на 2-7 г; в сравнении с контрольными здоровыми растениями содержание жира в них уменьшается на 2,1-6,9%, протеина на 4-5% (Заостровных, Дубовицкая, 2003). Рисунок 5 – Церкоспороз тройчатого листа (фото С.А. Титовой) Пурпурный церкоспороз вызывается грибом Cercospora kikuchii (Matsu, et Tomoyasu) Yardn. Первое сообщение о заболевании появилось в Корее в 1921 году, затем в Германии (Deutschmann, 1952), во Франции (Bernaux, 1976). В настоящее время заболевание распространено в Бразилии, Колумбии, Венесуэле, Никарагуа, Индии, Корее, Китае, Японии, Франции и Югославии (Заостровных, Дубовицкая, 2003). 18 Болезнь поражает сою во все фазы роста и проявляется на всех надземных органах. Заражённые семена часто имеют пурпурную или розовую окраску. Даже при сильном поражении семенная инфекция распространяется только в семенной кожуре (Овчинникова, 1971а). Больные семена дают изреженные всходы, у поражённых проростков семядольные листочки часто сморщены, иногда становятся тёмно-пурпурными и преждевременно опадают (Дега, 2012). Вредоносность болезни заключается в снижении посевных качеств семян и уменьшении содержания в них масла. Поражённые семена прорастают только на 15-25% (Овчинникова, 1971а). Белая гниль сои, склеротиниоз (возбудитель Sclerotinia sclerotiorum (Lib.) dBy.) (рис. 6). Заболевание встречается в основных зонах возделывания культуры. Оно было обнаружено ещё в конце XX века и нанесло большой ущерб соевым плантациям на одном из японских островов (Абрамов, 1931). В 1924 году заболевание было впервые обнаружено в США (Sinclair, Shurtleff, 1975). В настоящее время белая гниль является распространённым заболеванием в Аргентине, Франции, Венгрии, на Северном Кавказе и в Украине. Повсеместно белая гниль обнаружена и на территории Молдавии, однако чаще встречается в северных районах республики, где поражение растений достигает 20%. Подробное изучение белой гнили сои в Молдавии было проведено А.И. Ганя (1981) и Ж.Г Простаковой (1983). Были изучены распространение и вредоносность белой гнили сои, симптомы заболевания, анатомо- морфологические особенности строения возбудителя заболевания и другие вопросы. В России первые исследования этого заболевания были проведены на Дальнем Востоке (Абрамов, 1931, 1938). Автор отмечает, что в условиях Приморья обильные осадки в середине или конце августа всегда сопровождались значительным распространением склеротиниоза. В условиях повышенной влажности воздуха и при достаточно высокой температуре заболевание быстро распространялось по посевам и могло за несколько дней уничтожить 19 значительный процент растений. Массовое поражение соевых бобов (4-11%) было зафиксировано И.Н. Абрамовым в 1928 году в Гродековском районе (Заостровных, Дубовицкая, 2003). Рисунок 6 – Склеротиниоз на стебле сои (фото Л.К. Дубовицкой) Из всех грибных болезней сои на Дальнем Востоке наиболее вредоносны корневые гнили (Жуковская 1982; Есипенко, 1984, Дубовицкая, Гунинина, 1979; Дубовицкая, 1984). Вредоносность их приводит к изреженности посевов на 20-25%. По сообщению Т.А. Асеевой и соавторов (2008), в Хабаровском крае за последние годы на посевах с непротравленными семенами поражение растений сои корневыми гнилями (различной этиологии) составляло от 70 до 93% при развитии болезни от 35 до 48%; с протравленными семенами, соответственно, 45-50% и 1319% (Асеева, Золотарева, Паланица, 2008). Болезнь развивается в течение всей вегетации от всходов до созревания. Она вызывается одним или комплексом видов паразитных грибов. Заболевания корневой системы отмечаются также в Молдавии (Ганя, 1981), Абхазии 20 (Вардания, 1972), Грузии (Енукидзе, 1981), Краснодарском крае (Подкина, 1981; Подкина, Лавриченко, 1982). Корневые гнили имеют значительное распространение в Японии, Индии, Корее, Канаде, США (Liu, 1948; French, 1963; Овчинникова, 1970). Авторы подчеркивают высокую вредоносность этого заболевания. Так, в годы с плотной популяцией патогенов в почве потери зерна составляют 2-3 ц/га, а урожай зелёной массы снижается на 30-40%. Оценка вредоносности корневой гнили сои, проведённая в производственных и опытных посевах Л.К. Дубовицкой (1987), показала, что заболевание является одним из наиболее вредоносных, оказывающих отрицательное влияние на рост растений и продуктивность. В результате экспериментальных исследований морфологических структур здоровых и поражённых корневой гнилью растений Л.К. Дубовицкой (1987) установлено, что у больных растений снижалась высота стебля, масса сухого вещества, угнеталось развитие корневой системы и образование клубеньков, уменьшалось число бобов и масса зерна. Исследованиями микрофлоры корней сои в Приморском крае установлено, что в этиологии корневых гнилей ведущее место занимают грибные патогены (Нелен, Жуковская, 1967; Жуковская, 1968; Овчинникова, 1971а, 1971б). В зависимости от вида возбудителя различают: фузариозную, ризоктониозную, аскохитозную корневые гнили. Они широко распространены во всех исследованных почвах соевых полей, а под дикой соей встречаются редко или отсутствуют (Жуковская, 1977). Фузариозная корневая гниль – широко распространённое заболевание сои, встречающееся во всех зонах её возделывания в России, в том числе и на Дальнем Востоке. Поражаются первичные и вторичные корни, прикорневая часть стебелька и семядоли, отчего последние приобретают бурую окраску (рис. 7). Значение видов Fusarium в развитии корневых гнилей сои первым выяснил R.O. Cromwell в 1917 г. (Заостровных, Дубовицкая, 2003). 21 При сильном развитии болезни поражённая часть корней истончается, сморщивается. Заболевание может охватывать основания стебля и проникать в сосудистую систему, вызывая её окрашивание в красно-бурый цвет (Дубовицкая, 1987). Поражённые ткани покрываются белым или бело- розовым налётом гриба. Растения отстают в росте, мало образуется боковых корешков, а семядоли могут не раскрываться. Учёными Дальневосточного НИИ защиты растений определено, что потери урожая сои при поражении грибами рода Fusarium достигают 19,1% (5,1 ц/га). Фитоэкспертиза семян сои выявила, что фузариозом из них заражено 21,4%. Было установлено, что корневые гнили сои распространены во всех зонах края. Вызывались они грибами рода Fusarium, среди которых преобладает вид Fusarium solani. Поражение фузариозом проявилось также на семядольных листьях сои. Его распространение колебалось в пределах от 20 до 27%. Максимальное распространение фузариозной корневой гнили (86%) отмечено в фазе цветения культуры (Дега, 2012). Рисунок 7 – Фузариозная корневая гниль (фото С.А. Титовой) 22 Ризоктониозная корневая гниль – (Rhizoctonia solani Kuehn). Поражённая ткань имеет светло-коричневый цвет, зачастую носит опоясывающий характер, в местах поражения наблюдается утолщение стебля или корня (рис. 8). Корневая гниль сои и увядание взрослых растений, которые возникают при поражении грибом Rhizoctonia solani, отмечены в Иране (Hedjaroude, 1973), США (Boosalis, 1950; Tachibana, 1968; Henis, Ghaffar, Baker, 1979), Индии (Verma, Thapliyal, 1976). Рисунок 8 – Ризоктониозная корневая гниль (фото Ю.В. Положиевой) Развитие болезни приводит к поражению главного и боковых корней, стебелька и семядолей у проростков. При сильном поражении зачастую проростки ненормально развиты, без придаточных корешков, семядоли не раскрываются. Поражённая ткань размягчается, проросток увядает и гибнет, часть семян не прорастает. Поражённые проростки могут не давать всходов; если они прорастают, то имеют поражённый гипокотиль в области изгиба и обычно 23 погибают. Молодой поражённый проросток полупрозрачный и имеет симптомы «мокнущего корня». Верхушечная меристема может сильно замедлять рост, гипокотиль в диаметре может быть в 2 – 3 раза больше, чем нормальный, при этом не иметь признаков поражения или обесцвечивания. При поражении гифы гриба распространяются внутриклеточно и внедряются в стебель, проникают в оболочку и семядоли, не ограничиваясь никакими анатомическими зонами. Растения отстают в росте, листья скручиваются (Дубовицкая, Кравцова, 2002). Аскохитозная гниль – возбудитель Ascochyta sojaecola Abramoff. Syn. В результате поражения возбудителем заболевания наблюдается чаще локальное поражение с побурением и загниванием, затем подсыханием поражённой ткани в области прикрепления семени (рис. 9). Рисунок 9 – Аскохитозная корневая гниль (фото Ю.В. Положиевой) В результате образования на поражённых проростках расплывчатых овальных пятен проростки могут загнивать и погибать. На поражённой ткани 24 корней и основания стебля наблюдается концентричность и хорошо заметные под микроскопом плодовые тела. Из-за образования на поражённых проростках расплывчатых овальных пятен они могут загнивать и погибнуть. На стеблях, бобах и корнях образуются белёсые участки отмирающей ткани, на которой концентрично или беспорядочно формируются пикниды гриба, хорошо заметные под микроскопом (Заостровных, Дубовицкая, 2003). Бактериальные болезни сои. Бактериозы – широко распространённые вредоносные заболевания сои, встречающиеся повсеместно, где возделывается эта культура, и поражающие её во все фазы развития. Эти заболевания зарегистрированы на всей территории России, где культивируется соя: на Дальнем Востоке, в Краснодарском и Ставропольском краях, в Поволжье и др. В настоящее время выявлено около 10 заболеваний сои, вызываемых бактериями (Дега, 2012). Первые сведения о бактериальных болезнях сои приводят F. Coerper(1919), J. Kendrik (1921) и J. Shunk (1921) (Заостровных, Дубовицкая, 2003). В России бактериальные болезни сои изучали на Дальнем Востоке – И.Н. Абрамов (1938), А.Н. Иванова, З.В. Холопова (1956), Г.Ф. Солотчина, Э.Ш. Пехтерева (1966); в Московской, Воронежской областях – С.В. Владимирский (цит. по Заостровных, Дубовицкая, 2003), в Омской области – Е.В. Багаева, Н.И. Виноградова (1964); на Северном Кавказе – А.И. Лобик, О.Е. Катаева, В.И. Взоров (Заостровных, Дубовицкая, 2003), А.П. Клыков (1953), М.В. Краснова (1963). Самым распространённым и вредоносным из бактериальных заболеваний в Амурской области в настоящее время, по данным В.И. Заостровных (2003), является угловатая пятнистость. Бактериальная угловатая пятнистость, бактериальный «ожог», Pseudomonas glycineum Coerp. = P. syringae Hall распространена повсеместно, где возделывается соя: в США, России, Японии, Индии, Китае, Монголии, в странах Западной Европы (Заостровных, Дубовицкая, 2003). 25 Угловатая пятнистость считается одним из опасных заболеваний сои. В отдельные годы гибель всходов от неё составляет 30% (Абрамов, 1931, 1938; Никитина, 1962; Иванова, Холопова, 1956). Патоген поражает все надземные части растения, но чаще всего встречается на примордиальных и настоящих листьях (рис. 10). Сильно поражённые семена ослизняются, загнивают, издавая резкий запах, и не прорастают. На поражённых проростках образуются ослизнённые светло-жёлтые сквозные язвы или пятна с резким гнилостным запахом. Заболевание на листьях сои развивается в умеренно влажные годы. Максимальное развитие болезнь достигает в период цветения сои и продолжается до конца вегетации (Дега, 2012). Рисунок 10 – Бактериальный ожог, или угловатая бактериальная пятнистость тройчатого листа (фото С.А. Титовой) Вредоносность болезни выражается в снижении продуктивности больных растений, которое идёт за счёт уменьшения количества бобов на больных растениях и веса семян в них (Дубовицкая, Кравцова, 2002). В Приморье вредоносность заболевания, по данным А.М. Овчинниковой (1971а, 1971б), заключается в поражении всходов от 5% до 71%, вегетирующих растений и семенного материала – до 40-100%, что приводит к снижению урожая 26 и его качества. При поражении листовой формой бактериального ожога снижение урожая может составлять в среднем 22% (Солотчина, 1980). При семенной инфекции количество бобов на заражённых растениях на 15-20% меньше по сравнению со здоровыми растениями, а при листовой форме – на 26-32%. Урожайность семян с заражённых растений снижается в среднем на 27% (Заостровных, Дубовицкая, 2003). Таким образом, видовой состав болезней на сое в различных экологогеографических зонах её возделывания весьма разнообразен, как различна и их вредоносность. Приведённые данные о распространенности и вредоносности болезней свидетельствуют о том, что, несмотря на длительный период борьбы с ними, человечество так и не научилось сводить потери урожая до экономически приемлемого уровня при экологической безопасности технологий для окружающей среды. 1.2 Влияние экологических факторов на развитие инфекции сои Экологические условия играют важную роль, как в возникновении заболевания, так и его развитии. Интенсивность распространения и развития болезней культурных растений определяется характером взаимоотношений трех компонентов: возбудителя болезни, растения-хозяина и внешней среды. Среди комплекса факторов среды решающая роль принадлежит ее абиотическим элементам – климату и погоде (Гофман, 2007). Абиотические факторы оказывают огромное влияние на взаимоотношения растения-хозяина – патоген, на возникновение и течение патологических процессов в нём (Стэкмен, Харрар, 1959). На заражение патогенами, споры, которых переносятся по воздуху, наибольшее воздействие оказывают температура и влажность, а в некоторых случаях – свет и атмосферное давление. Характер поражения растений 27 возбудителями, передающимися через почву, определяется также величиной рН субстрата, аэрацией и общим содержанием питательных веществ (Эванс, 1971). Температура, влажность воздуха и почвы, кислотность субстрата, аэрация, освещенность – все эти факторы существенно влияют на развитие возбудителей. В то же время в зависимости от этих факторов формируется устойчивость растений к заражению патогенами, их восприимчивость и выносливость к заболеваниям в различные периоды вегетации. Температура среды является одним из основных факторов, определяющих возможность возникновения заболевания растений и степень его вредоносности. Влияние этого фактора начинает проявляться уже на первых этапах инфекционного процесса, обусловливая жизнеспособность возбудителя болезни и возможность его сохранения к началу вегетационного периода (Гофман, 2007). Основные грибные болезни в дальневосточном регионе – септориоз, пероноспороз, церкоспороз, фузариоз, аскохитоз, склеротиниоз. Из бактериальных чаще всего встречается бактериальная угловатая пятнистость (Дега, 2012). Наиболее полно исследовано воздействие экологических условий на Peronospora manshurica (Корецкий, 1967; Mckenzie, Wyllie, 1971; Dunleavy, 1982), Septoria glycines (Hemmi, 1940; Mullens, Hilty, 1979; Peterson, 1982). Влияние условий среды на развитие Ascochyta sojaecola ранее не изучалось (Наумов, 1986). Возбудитель септориоза развивается при температуре от 5 до 36°С (Hemmi, 1940). Спороношение гриба происходит при температуре не ниже 18-20°С и относительной влажности воздуха 95-100%. Частые дожди, обильные росы и высокая температура во второй половине июля и августа способствуют усилению развития септориоза. Условия, способствующие эпифитотийному развитию септориоза, следующие: температура воздуха в пределах 26-28°С и относительная влажность в период заражения 80-90%. Очень благоприятны для распространения заболевания частые дожди, высокая температура во второй половине июля – августа и обильные росы (Возбудители…, 1980). 28 В литературе имеются сведения о поражении сои септориозом в первой половине её вегетации (фаза второго настоящего листа) (Jonson, Kochler, 1943; Kurata, 1960), но для успешного развития патогена в этот период необходима тёплая и влажная погода (Овчинникова, 1971а; Mullens, Hilty, 1979; Young, Ross, 1979; Ross,1982). При обильном количестве осадков, частых росах и туманах происходит интенсивное распространение болезни (Корецкий, 1967). На формирование пикноспор не влияют свет и темнота. Существует тенденция ускоренного их развития при колеблющейся температуре (Hemmi, 1940). Жизнеспособность спор септориоза снижается при pH от 9 до 10 и повышается при pH от 3 до 8 (Заостровных и др., 2000). Возбудитель пероноспороза (Peronospora marshurica (Naum.) Syb), относится к представителям воздушно-капельно-семенной подгруппы инфекций. Они нуждаются в наличии капельно-жидкой влаги или высокой относительной влажности воздуха (80-100%) на всех этапах передачи (Гофман, 2007). Распространение конидий происходит особенно интенсивно после дождей и длительных туманов, когда капли воды сохраняются не менее 5 часов (Возбудители…, 1980). По результатам исследований А.М. Овчинниковой (1971а), Ж.Г. Простаковой, А.И. Гани (1983) развитие болезни приостанавливается не только в сухую жаркую погоду, но и в период сильных ливневых дождей, когда конидии смываются с листьев. Для прорастания конидий пероноспоровых грибов необходимо наличие капельно-жидкой влаги в течение определённого времени (Корецкий, 1967; Mckenzie, Wyllie, 1971; Lim, 1978; Новотельнова и др., 1979). Е.С. Наумовой (1986) было отмечено, что в условиях Воронежской области наибольшего развития пероноспороз достигал в 1978 г. Обильные осадки, составившие 136-198% нормы во все три декады июля, повышенная относительная влажность воздуха, длительное сохранение росы на листьях способствовали быстрому распространению и прорастанию конидий гриба. В 29 июле соя находилась в фазе цветения, для которой характерно быстрое нарастание верхушек растений, что также способствовало развитию гриба, поражающего преимущественно молодые растущие ткани (Wyllie, Williams, 1965; Riggle, 1977). Развитию возбудителей пероноспороза способствуют повышенная влажность воздуха и большое количество осадков в сочетании с пониженной температурой. Оптимальной температурой для развития гриба считается 18-20°С при относительной влажности воздуха 95-100% (Заостровных, Дубовицкая, 2003). Условиями, способствующими интенсивному развитию церкоспороза и образованию спороношения, являются температура в пределах 20-30°С и относительная влажность 90-100%. Сильное развитие заболевания может происходить при частых росах, туманах, высокой с резкими колебаниями относительной влажности воздуха (величина гидротермического коэффициента более 2) (Заостровных, Дубовицкая, 2003). Оптимальные условия для развития возбудителя болезни пурпурного церкоспороза – относительная влажность воздуха выше 90% и температура 2328°С. Способствуют развитию болезни выпадение обильных осадков, появление туманов или рос, запаздывание с уборкой сои. Заражению растений аскохитозом способствуют влажность воздуха более 90% и температура выше 4°С (Заостровных, Дубовицкая, 2003). В Воронежской области в 1980 г. большое количество осадков и повышенная относительная влажность воздуха (до 87%) способствовали нарастанию степени развития аскохитоза на листьях сои до 79,9% в первой половине августа, а в сентябре – до 79,4% на стеблях (Наумова, 1986). Массовому развитию антракноза способствуют высокая относительная влажность воздуха, частые дожди (Жуковская, 1990). Растения, поражённые грибом Sclerotinia sclerotiorum (возбудитель склеротиниоза), погибают в течение 4-5 дней. Белая гниль особенно вредоносна на поливных участках, где условия развития и распространения патогена особенно благоприятны (Соя, 1984). 30 Белая гниль в условиях повышенной влажности воздуха и при достаточно высокой температуре быстро распространяется по посевам и обычно за несколько дней уничтожает значительный процент растений (Кузнецова, 1964). Начальное развитие возбудителя при большинстве инфекционных болезней растений тесно связано с наличием влажности, достаточное количество которой является предварительным условием заражения. Решающим фактором в распространении заболевания являются осадки. Первые этапы развития возбудителей большинства инфекционных болезней растений тесно связаны с наличием капельно-жидкой влаги, достаточное количество которой является предварительным условием заражения, а также зависят от pH среды. Исследования Л.К. Дубовицкой (1987) показали, что возбудители корневых гнилей хорошо развиваются в широких пределах pH (58,5). Максимальный рост грибов, F. solani, G. roseum, C. destructans, наблюдался при рН близкой к 7. Для проявления и распространения фузариозов большое значение имеет температура в период прорастания семян. Так, в результате исследований в центральной и южной лесостепи Кемеровской области (1996-1998 гг.) было установлено, что при посеве сои 15-16 мая, когда температура почвы составляла 10°С, всходы появляются на 13-14-й день, а при посеве сои 26-29 мая (температура почвы 18-19°С) продолжительность появления всходов сокращалась почти в 2 раза и при этом значительно уменьшалось количество болезней, в том числе и фузариозов. При понижении температуры почвы появление всходов замедляется, а гриб при этих условиях может интенсивно развиваться (Заостровных, Бакач, Незнамова, 1991). Развитию бактериальных болезней в значительной степени способствуют высокая влажность, температура и слабощелочная или нейтральная реакция среды. Большинство бактерий хорошо развиваются при температуре 18-25°С. Для фитопатогенных бактерий наиболее благоприятна повышенная влажность воздуха и почвы. Свет неблагоприятно действует на большинство бактерий, поэтому в загущенных посевах сильнее развивается бактериоз (Яковлева, 1992). 31 Максимальное развитие угловатой пятнистости наблюдается во второй половине июля-августе, когда среднесуточная температура воздуха составляет 2227°С и выпадает значительное количество осадков – до 300 мм и выше. Оптимальная температура для развития возбудителя составляет 22-23°С (Дубовицская, Кравцова, 2002). Благодаря теплой погоде и постоянным ветрам болезнь интенсивно развивается и распространяется. Таким образом, различные экологические факторы оказывают существенное влияние на развитие патогенов сои. Развитию возбудителей аскохитоза и пероноспороза способствуют повышенная влажность воздуха и большое количество осадков в сочетании с пониженной температурой. Однако аскохитоз может развиваться в более широком интервале температур (14-28°С, оптимальная 18-24°С), чем пероноспороз (14-20°С, оптимальная 18-20°С). Более высокая температура необходима для развития септориоза (26-28°С) и церкоспороза (20-30°С) в сочетании с повышенной относительной влажностью воздуха (90-100%). Развитию фузариоза способствуют похолодание после посева и избыток влаги весной. Для возбудителей белой гнили сои необходима прохладная и влажная погода во второй половине вегетации растений. Развитию бактериальной инфекции способствуют высокая температура (18-25°С), достаточная относительная влажность и сильные ветра. 1.3 Ферменты инфицированных растений Одним из самых серьёзных и универсальных биотических воздействий, которым подвергаются растения, являются патогенные микроорганизмы. Они представляют собой довольно разнообразные типы организмов: грибы, бактерии, вирусы, вироиды, фитоплазмы, актиномицеты. Насекомые и нематоды также часто вызывают у растений схожие реакции на патогенное воздействие. В процессе эволюции у растений сформировались разнообразные системы (генетические, физиологические, биохимические, 32 морфологические), обеспечивающие их устойчивость к поражению болезнями и вредителями (Жученко, 2001). Механизмы иммунитета растений многообразны (Метлицкий, 1976; Аверьянов, 1991; Чесноков, 2007). Наиболее ранней ответной реакцией растений на воздействие стрессоров биотической и абиотической природы является образование активных форм кислорода (АФК) (Bolwell et al., 1999; Torres et al., 2000; Минибаева, Гордон, 2003). К ним относятся супероксидный (О 2-), гидрооксидный (ОН-) свободные радикалы, перекись водорода (Н2О2), а также оксид азота (Hippeli et al., 1999; Dröge, 2002; Mittler, 2002; Neill et al., 2002; Чеснокова и др., 2006; Полесская, 2007). Взаимодействие патогена и растения-хозяина вызывает значительные изменения в функциональном состоянии растений, что, прежде всего, отражается на активности ферментов (Савич, 1989а). Роль ферментов в жизнедеятельности животных, растений и микроорганизмов колоссальна. Сложнейшая сеть химических превращений, которые идут в живом организме, непрерывно и всецело зависит от действия ферментов. Ферменты не только избирательно ускоряют определенные химические реакции, но и столь же избирательно отвечают глубоким изменением каталитической активности на тонкие сдвиги в состоянии среды, где эти реакции протекают. В 50-е годы ХХ века было установлено, что многие ферменты присутствуют в клетках животных, растений и микроорганизмов в виде множественных молекулярных форм (изоферментов). Исследования 60-70-х гг. показали, что изоферменты имеют как генетическое, так и негенетическое (посттрансляционное) происхождение (Уилкинсон, 1968; Райдер, Тейлор, 1983; Яаска, 1985; Корочкин, 1987; Глазко, Созинов, 1993). Присутствие в клетке изоферментов, различающихся по структурным, физико-химическим и кинетическим свойствам, клеточной компартмен- тализации, а также ткане- и видоспецифичности наряду с другими механизмами, обеспечивает быструю и тонкую регуляцию клеточного метаболизма в условиях 33 приспособления организма к постоянным изменениям внешней среды (Хочачка, Сомеро, 1977; Райдер, Тейлор, 1983). Работы Л.И. Корочкина (1977), Б.П. Плешкова (1980), В.Г. Конарева (1983), Ф. Айала (1984), В. Яаска (1985) и др. внесли значительный вклад в разработку фундаментальных исследований изоферментов. Значительное число работ посвящено исследованию изоферментов растений (Титов, 1975, 1978; Chauhan et al., 1985; Левитес, 1986; Шерепитко и др., 1990; Газарян, 1992). В повышении устойчивости растений к неблагоприятным факторам среды пероксидазе отводится особое место (Рубин и др., 1975; Перепадя, Диканев, 1977; Карташова и др., 2000; Рогожин, 2004; Максимов и др., 2011). В настоящее время это наиболее изученный фермент растений. Пероксидаза представляет собой двухкомпонентный фермент – железопротеин, трехвалентное железо которого не меняет своей валентности в процессе катализа. Фермент локализован в клеточной стенке, митохондриях, хлоропластах, ядре, цитоплазме и других органеллах. Специфичность распределения пероксидазы связана с выполнением ферментом различных физиологических функций (Саундерс, 1972; Левитес, 1986; Садвакасова, 1987; Андреева, 1988; Савич, 1989а; Газарян, 1992). Основная функция пероксидазы – катализировать окисление химических веществ за счет кислорода перекиси водорода. Установлено, что фермент обладает не только пероксидазными, но и оксидазными свойствами, катализируя окисление целого ряда соединений за счет неактивированного молекулярного кислорода. Необходимым условием этой реакции является наличие кофакторов – ионов марганца и фенольных соединений. Субстратами при этом служат ароматические амины, анилин, n-толуидин, бензидин, тирозин, триптофан, индол, индолилуксусная кислота (ИУК), фенолы (катехин, гидрохинон, резорцин, гваякол, ванилин, n-крезол, пирагаллол), ароматические кислоты (бензойная, салициловая, галловая), ферроцитохром С, НАДФ.Н2, аскорбат, натраты (Саундерс, 1972; Droillrd et al., 1987; Андреева, 1988; Газарян, 1992). Регуляция активности пероксидазы может происходить как за счет синтеза фермента de novo (Lagrimini et al., 1997) и освобождения связанных форм 34 (Андреева и др., 1980, 1981; Андреева, 1988), так и её модификации протеазами, низкомолекулярными соединениями, например салициловой кислотой, супрессорами и элиситорами патогена (Christensen et al., 1998). Пероксидаза является ферментом, обладающим повышенной чувствительностью к внешним воздействиям среды на растения. Обладая функциональностью, она способна реагировать на большинство нарушений гомеостаза. Изменения активности пероксидазы и её спектров прослежены при действии низких и высоких температур (Савич и др., 1988; Савич, 1989б, 1989в; 1990; Хайруллина, 2010), фитогормонов (Liman et al., 1998), в условиях дефицита минерального питания, засухи, засоления, в отклике на поранение (Lazar, Farkas, 1970; König, Nienhaus, 1970; Lagrimini et al., 1997), в условиях анаэробиоза и других неблагоприятных факторов (Андреева, 1988; Савич, 1989; Fieldes, Gerhardt, 1998; Kazan et al., 1998). Воздействие биотических стрессоров приводит к ответу растений, в основных чертах сходному с ответом на абиотические стрессоры (Тарчевский, 2002). Роли фермента пероксидазы в метаболизме больного растения посвящено большое количество работ Б.А. Рубина (Рубин и др., 1965; Рубин, Лущинская, 1969; Рубин, Будилова, 1970; Рубин и др., 1975). При заражении растений капусты грибом Botrytis cinerea установлено, что одним из постоянных и отчетливых эффектов, наблюдаемых в инфицированной ткани иммунных форм, является активирование в них пероксидазы, которое может достигать 150-160%, а иногда и большей величины по сравнению с активностью этого фермента в здоровой ткани (Рубин и др., 1973). Образующиеся в результате окисления пероксидазой хиноны и продукты их конденсации обладают фунгитоксическими и бактерицидными свойствами и служат химическим барьером при образовании некротической поверхности (Рубин и др., 1975). Принимая участие в синтезе лигниноподобных веществ, пероксидаза имеет отношение к созданию механического барьера на пути инфекции как при 35 заживлении поранений, так и в случае заражения (Brown, 1965; Гамбург и др., 1990). Б.А. Рубин и др. (1974) в экспериментах с разными сортами пшеницы установили, что все устойчивые к стеблевой и бурой ржавчине образцы отличались от восприимчивых сортов более значительным увеличением активности пероксидазы. Активирование пероксидазы под влиянием инфекции является характерной ответной биохимической реакцией, по которой можно судить об устойчивости растений. Повышение активности фермента пероксидазы идет параллельно с формированием приобретенной устойчивости (Vegetti et al., 1975; Рубин, Ладыгина, 1977; Андреева, 1988). Однако повышенная пероксидазная активность не всегда коррелирует с устойчивостью и не всегда её индуцирует. Об этом свидетельствуют исследования (Чигрин и др., 1968; Seevers, Daly, 1970; Seevers et al., 1971; Daly, 1972; Серова и др., 1982), в которых установлено, что активность пероксидазы в пораженных грибной инфекцией растениях устойчивых сортов не изменяется, а у восприимчивых увеличивается или изменяется одинаково у устойчивых и восприимчивых сортов. Некоторые ржавчинные инфекции вызывают снижение активности пероксидазы. Так, поражение осота Puccinia suaveolens сопровождается падением активности фермента на всех стадиях развития гриба, за исключением периода возникновения спор, когда происходит возрастание пероксидазной активности (Серова и др., 1982). Ряд исследователей пришли к выводу, что активация пероксидазы, наблюдаемая в вирозных растениях, происходит за счет синтеза ферментного белка de novo (Farkas et al., 1966; Solymosy et al., 1967; Chant, Bates, 1970; Ладыгина и др., 1970; Ладыгина, 1971); другие, отмечая повышение активности, не обнаружили качественных различий в спектрах пероксидаз в здоровых и пораженных вирусами растениях (Novacky, Hempton, 1968; Marcinka, 1970; Khatri, Chenulu, 1970; Jones, Stoddort, 1971; Серова и др., 1982). 36 Изучая изменения в изоферментном наборе пероксидазы, сопровождающие вирусный патогенез устойчивого и неустойчивого к вирусу табачной мозаики (ВТМ) видов табака, М.Е. Ладыгина и др. (1970) пришли к выводу о возможной функциональной неравноценности разных изоферментов. Авторы считают, что с устойчивостью к ВТМ связаны в основном медленно движущиеся фракции ферментов, участвующие в превращении фенолов. Инфицирование растений льна Melampsora lini приводило к увеличению количества изопероксидаз (Andreev, Shaw, 1965), часть из которых была, по мнению авторов, патогенного происхождения, а часть – растения-хозяина. В корнях растений сои, устойчивых к Phytophtora megasperma var sojae, после поражения появляется одна дополнительная зона белков с пероксидазной активностью, а у восприимчивого идентифицированы две зоны (Ho, Waever, 1970). Некоторые авторы считают, что изменения в составе пероксидаз не связаны с защитной реакцией растений, а являются следствием нарушения обмена веществ (Grzelinska, 1970). В листьях разных сортов пшеницы, устойчивых и восприимчивых к стеблевой ржавчине, при заражении значительно повышалась активность пероксидазы и изменялся изоэнзимный спектр. Её возрастание всегда было связано с появлением одного (9-го) изоэнзима. При 25оС пшеница теряла устойчивость, при этом активность 9-го изоэнзима исчезла (Seevers et al., 1971). Эти данные ещё раз подчеркивают важную роль пероксидазы в системе защитных реакций растения. В.А. Андреева (1988) считает, что можно использовать пероксидазные маркеры для характеристики защитного механизма растений. По её мнению, таким путем нужно искать подходы к диагностике устойчивости сельскохозяйственных культур к вирусам. Устойчивость растения к патогену определяется способностью быстро распознать его и запустить защитные реакции (Lee, Hwang, 2005). 37 Анджана и соавторы (Анджана и др., 2007) в листьях устойчивого генотипа подсолнечника выявили экспрессию трех изоформ пероксидазы (ПО1, ПО2, ПО3), которые были обнаружены через 2 и 12 ч после заражения Alternaria helianthi. В листьях восприимчивого генотипа изоформа ПО2 присутствовала спустя 24 и 48 ч после инкубации с патогеном, изоформа ПО3 обнаружилась через 24 ч. Simons с соавторами (Simons et al., 1971) было показано, что заражение нижних листьев растений табака вирусом табачной мозаики приводит к повышению активности пероксидазы в листьях, расположенных выше инокулированных. И.А. Граскова (Граскова, 2004; 2008) изучала влияние бактериального патогена вирулентного штамма 5369 возбудителя кольцевой гнили картофеля Clavibacter michiganensis subsp. sepedonicus на активность пероксидазы в различных тканях растений картофеля, выращенных в стерильных условиях. Показано, что при корневом заражении растения активность фермента возрастала в клетках всех типов тканей (корни, листья, стебли). В тканях устойчивых сортов картофеля эта активность была значительно выше, чем у восприимчивого сорта, причем сам патоген пероксидазной активностью не обладал. По возрастанию пероксидазной активности в тканях можно судить об эффективности использования системных фунгицидов и других соединений с целью повышения иммунитета растений. Установлено, что фунгициды (фундазол и байлетон) повышают иммунные свойства растений пшеницы к мучнистой росе, что сопровождается увеличением активности пероксидазы (Юрина и др., 1998). В.А. Аксенова и соавторы (Аксенова и др., 1971) показали, что резкое возрастание оксидазной активности пероксидазы в пораженных тканях устойчивого сорта капусты приводит к накоплению в них продуктов окисления флороглюцина, которые обладают четко выраженным фунгицидным действием и поэтому создают на пути инфекции химический барьер. Активность пероксидазы при поражении возрастает в растениях обоих сортов, но сильнее – у устойчивого. 38 О влиянии вирусной, грибной и бактериальной инфекции на изменение активности пероксидазы сообщается в целом ряде других работ (Минибаева, Гордон, 2003; Юсупова и др., 2006; Максимов и др., 2006, 2010). Таким образом, пероксидаза является одним из компонентов защитной системы, эффективно функционирующей как на начальных стадиях защиты организма от патогенов, так и в процессе индуцированного иммунитета. Каталаза. Как по структуре, так и по функциям каталазы родственны пероксидазам. Они принадлежат к числу гемопротеиновых ферментов. Молекулы каталаз содержат по четыре атома железа, каждый из которых хелатирован с протопорфирином IX и, кроме того, связан с белком (Саундерс, 1972). Самая характерная функция каталазы – высокоэффективный катализ разложения перекиси водорода на воду и кислород. Кроме того, каталаза проявляет умеренную пероксидазную активность, катализирует реакции окисления перекисью водорода (Саундерс, 1972). Перекись водорода образуется в растениях и клетках животных в процессе окисления. В повышенных концентрациях перекись водорода оказывает токсическое действие на цитоплазму клеток. Каталаза играет главную роль в нейтрализации перекиси водорода, поэтому она является важнейшим компонентом антиоксидантной системы (Рубин, Ладыгина, 1977; Скулачев, 1996; Ленинджер, 1999; Чеснокова и др., 2006). Каталаза встречается в клетке вместе с пероксидазой. Между этими ферментами, как указывал А.Н. Бах (1936), существует кажущийся антагонизм: пероксидаза активирует перекись водорода, в то время как каталаза быстро и полностью разлагает её. Однако, действуя одновременно, оба фермента не нарушают своих специфических функций: каталаза разрушает ту часть перекиси водорода, которая не может быть использована пероксидазой для окислительных процессов. В результате изучения физико-химических свойств каталазы установлено, что фермент активен в слабокислой среде, но в нейтральной достигает наибольшей активности. В щелочной среде активность каталазы резко падает. 39 Установлено, что активность каталазы в зависимости от концентрации белка увеличивается пропорционально его качеству. При увеличении количества Н2О2 активность фермента возрастает до определённого значения, после чего идет насыщение субстратом и изменение активности не наблюдается. Установлено, что каталаза высокоактивна при температуре инкубационной среды до + 60 оС (Иваченко и др., 1997). Каталаза обнаружена почти во всех клетках растительного и животного происхождения. Однако в литературе мало сведений о состоянии этого фермента в растительных тканях. Лишь в отдельных работах изучалось состояние каталазы некоторых растений (Акулова, Смолов, 1974; Droillrd et al., 1987; Гигрин, 1988; Filipe et al., 1988). Литературные данные о роли каталазы в формировании взаимоотношений хозяин – паразит противоречивы. А. Коно (1961) установил, что у устойчивых к корончатой ржавчине сортов овса активность каталазы более высокая, чем у восприимчивых. В.П. Нилова с соавторами (1948) считают, что устойчивые сорта пшеницы обладают пониженной активностью каталазы, вследствие чего в тканях накапливается перекись, к которой ржавчина чрезвычайно чувствительна. Возрастание активности каталазы после заражения восприимчивых сортов пшеницы стеблевой и листовой ржавчиной обнаружили В.Д. Гоцуляк, О.К. Кондратюк (1967). Аналогичная зависимость установлена и М.И. Дементьевой (1957) для инфицированных ржавчиной восприимчивых сортов крыжовника. Особенно высокая активность свойственна тканям, в которых непосредственно локализован гриб. В связи с изменением путей окисления метаболитов, происходящим при заболевании, создаются условия для возникновения значительных количеств перекисных соединений (Серова и др., 1982). Влияние факультативных паразитов на активность каталазы питающего растения изучено значительно слабее, чем облигатных. По данным S. Vir, J. 40 Grewal (1975), Ascochyta rabiei индуцирует в тканях нута устойчивого сорта увеличение каталазной активности и снижение её у восприимчивых. Л.А. Ганеева, И.С. Докучаева (2009) изучали влияние соединений марганца на активность каталазы и пероксидазы в клубнях картофеля и на проявление агрессивности фитофторы. Результаты исследования показали, что обработка ткани картофеля соединениями марганца приводит к снижению активности каталазы и пероксидазы и индуцирует устойчивость картофеля к фитофторе. Таким образом, вследствие небольшого количества экспериментальных данных роль каталазы в формировании взаимоотношений патогена и растенияхозяина полностью не определена. Кислые фосфатазы. Кислые фосфатазы найдены во всех изучаемых тканях растений, где, кроме того, часто присутствовали и различные изоформы фермента (Сафонов и др., 1971; Лукашевич и др., 1974). Множественные формы кислой фосфатазы различаются между собой по целому ряду физико-химических параметров и свойств: относительной молекулярной массе, изоэлектрической точке, термостабильности, потребности в катионах металлов, оптимуму рН, Км (Hollander, 1971; Власюк, Тернавский, 1973; Naoi, Teruniko, 1977; Эчкалов, 1988). Внеклеточные кислые фосфатазы присутствуют, например, в корнях растений. Они могут быть локализованы в клеточной стенке и, возможно, секретируются клетками корня в окружающую его сферу. Конкретная физиологическая роль растительных внеклеточных кислых фосфатаз пока ещё не установлена. Но несомненно то, что внеклеточные кислые фосфатазы корней, локализованные преимущественно в апикальной меристеме и клетках наружной поверхности корня, задействованы в гидролизе фосфорсодержащих продуктов, а также в непосредственной мобилизации органического фосфата почвы для питания растения (Hagima, Saulescu, 1982; Галимова, 1987). Внутриклеточные кислые фосфатазы обнаружены в находящихся в покое семенах, развивающихся семенах, прорастающих семенах, листьях, побегах, корнях, цветах, плодах, а также в культивируемых клетках растений. 41 Экспрессия кислых фосфатаз модулируется разнообразием факторов окружающей среды, в частности, стрессовыми воздействиями различного характера. Значительная индукция активности кислых фосфатаз происходит при прорастании семян. Хотя и в покоящихся семенах имеется невысокая активность кислых фосфатаз, сильное её увеличение (до 30-кратного) обычно сопровождается прорастанием семян (Полякова, 1981). Синтез кислых фосфатаз индуцируется гиббереллиновой кислотой (Duff et al., 1994). Другие фазы развития, на которых происходит возрастание активности кислых фосфатаз, – это фазы цветения, созревания плодов и старения. Здесь главную роль играет абсцизовая кислота, которая повышает активность кислых фосфатаз при старении листьев (Гуляй, Макарчиков, 2001). На активность кислых фосфатаз большое влияние оказывают температурный режим, рН среды, условия влагообеспечения (Schidhara, Bhat, 1963; Khan, 1978; Шнырева, 1992). Резкое угнетающее действие на фосфатазную активность оказывает повышение температуры до +65оС. Однако тепловой шок приводит к повышению теплоустойчивости кислых фосфатаз проростков гороха (Schridhara, Bhat, 1963; Hollander, 1971). Кислая фосфатаза является защитным ферментом растений. Она способна нейтрализовать действие широкого спектра инфекций, вызванных различными микроорганизмами (Naoi, Teruniko, 1977; Шнырева, 1992). В целом можно отметить, что кислые фосфатазы принимают участие как в деструктивных процессах, полифункциональными так и в ферментами реакциях благодаря анаболизма, широкой являясь субстратной специфичности и многообразию изоферментов. Они являются существенными компонентами углеводно-фосфорного и липидного обмена. Важность их очевидна из-за большого количества фосфорсодержащих соединений в растении. Однако данных о роли кислых фосфатаз в формировании иммунитета растений недостаточно. 42 Из приведенного анализа литературных источников видно, что наиболее характерная реакция растений на заболевание проявляется в изменении ферментативной активности, контролирующей метаболизм хозяина. Несмотря на большой объем научных исследований по изучению роли пероксидазы в метаболизме больного растения, недостаточно данных об участии каталазы и кислой фосфатазы в защитных реакциях растений, отсутствуют сведения об изменении активности ферментов в растениях G. max и G. soja под влиянием грибной и бактериальной инфекции. Подобные исследования заслуживают особого внимания, так как изучение биохимической природы устойчивости в системе «хозяин-растение (Glycine max (L.) Merrill и G. soja Siebold & Zucc.) – патоген» позволит более объективно подойти к оценке и отбору исходного материала для селекции, понять причины изменчивости степени устойчивости растений в зависимости от экологических факторов, разработать наиболее эффективную систему интегрированной защиты растений. 43 2 УСЛОВИЯ, ОБЪЕКТЫ И МЕТОДИКА ПРОВЕДЕНИЯ ИССЛЕДОВАНИЙ 2.1 Агроклиматические и почвенные условия Амурской области Амурская область расположена на юго-востоке Азиатской части России и занимает 11,7% территории Дальнего Востока. Общая площадь области составляет 363,7 тыс. км2. Протяженность её с севера на юг – 760 км и с запада на восток – 700 км. Рельеф Амурской области горно-равнинный: горы занимают 57,5%, равнины – 42,5% территории. На трёх крупных равнинах (Зейско-Буреинская, Амуро-Зейская, Верхнезейская) расположены южная, центральная, северная, северная таёжная, горная таёжная зоны. Сельским хозяйством занимаются в четырёх зонах, а земледелием – в трёх (южная, центральная, северная) (Зональная…, 1982). Климат Амурской области резко континентальный по температурным признакам и муссонный по характеру формирования. Он формируется под влиянием Азиатского континента и Тихого океана, имеющих различную температуру поверхностей в зимнее и летнее время (Система…, 2003). Влияние материка проявляется главным образом зимой, когда сухой и сильно охлаждённый на континенте воздух проникает на территорию области в виде сильного муссона. Поэтому в Амурской области наблюдаются холодные и малоснежные зимы. Весна и осень являются переходными периодами. Амурская область характеризуется неустойчивым гидротермическим режимом муссонного характера. Это проявляется коротким безморозным периодом, поздним возвратом холодов в весенний период, неравномерным распределением тепла и влаги. Весна в Приамурье сухая, холодная, с сильными ветрами в апреле-мае. Среднемесячные положительные температуры воздуха в южной части наступают в первой декаде, в центральной и северной – во второй декаде апреля. Прогревание почвы на глубине 10 см до + 5 оС наступает в южных районах в начале второй декады, в центральных и северных – в середине- конце второй декады апреля. Количество осадков в марте-мае составляет 56-70 мм, или 11-18 % 44 среднегодового. Весной влажность почвы в Приамурье определяется в основном летне-осенними запасами, обычно они в слое 0-20 см составляют 60-70 мм продуктивной влаги. Отношение осадков к испарению в этот период 0,5-0,6. Лето в Приамурье повсеместно влажное и теплое. С июля по сентябрь выпадает 286-350 мм осадков (66-76 % годового количества), что вызывает периодическое переувлажнение почвы. Среднемесячная температура июля в основных сельскохозяйственных районах 20-21оС. Осень сухая, теплая. Среднесуточные положительные температуры воздуха в южных и центральных районах области удерживаются до конца третьей декады октября, в северной – до конца второй декады. Осадков выпадает 75-90 мм, или 16-19 % годовой нормы. Безморозный период в южной зоне составляет около 130 дней, в центральной зоне 111-113 дней, в северной зоне – 92-100 дней. Высокое напряжение тепла, обилие света и достаточное количество осадков в течение наиболее теплых месяцев благоприятствуют выращиванию сои. Особенности агроклиматических условий и опыт возделывания сои указывают, что в основных сельскохозяйственных районах Амурской области можно возделывать сорта сои преимущественно с коротким периодом вегетации. Основные типы почв в сельскохозяйственном производстве области представлены лугово-чернозёмовидными, луговыми, бурыми лесными, пойменноаллювиальными и лугово-бурыми (Зональная…, 1982). Почва опытного участка ДальГАУ представлена в основном более 60% от площади пашни, наиболее плодородной в Амурской области лугово- черноземовидной различной мощности. (Система…, 1997). По агрохимическим свойствам почва опытного участка характеризуется средне- и слабокислой реакцией среды и в основном не нуждается в известковании. Значительная часть пашни имеет низкое содержание доступных растениям форм фосфора. Содержание доступного растениям калия в почвах опытного участка среднее (8,8 % пашни) и повышенное (91,2 %). 45 Метеорологические условия 2008 – 2011 годов имели некоторые отличия от среднемноголетних по температурному режиму и количеству осадков (приложения 1, 2). Весна в 2008 году наступила рано, была неустойчивой, в начальный период необычно тёплой. Май этого года характеризовался неравномерным ходом температурного режима. Запасы продуктивной влаги в пахотном слое почвы составляли 36-45 мм – это 50-60% от наименьшей полевой влагоёмкости (НПВ), в полуметровом слое – 80-100 мм, это 60-70% от НПВ. Средняя температура воздуха за месяц составила 11,2оС (в пределах климатической нормы). С дождём было 5-7 дней. Сумма их составила 55 мм – это 125% нормы. Сумма эффективных температур выше +5оС составила 290оС, на 80оС больше нормы. Запасы продуктивной влаги на 28 мая составляли в пахотном слое 30-40 мм, в полуметровом слое 70-110 мм, это 60-70% от НПВ. Агроклиматические условия для начального роста и развития сельскохозяйственных культур складывались благополучно. Начало лета 2008 года было тёплым и сухим. В июне средняя температура воздуха превысила среднемноголетнюю норму на 3оС. Июль и август характеризовались тёплой погодой с неустойчивым температурным режимом и неравномерным выпадением осадков. Средняя температура воздуха за летние месяцы оказалась выше на 2-3оС. Сумма положительных температур за период с температурами выше +15оС составила 2150оС – это выше климатической нормы на 350оС. Осадки в летний период распределялись неравномерно. В июне их было мало, сумма составила 38 мм, это 38% многолетней нормы. Погодные условия июня были неблагоприятны для роста и развития сельскохозяйственных культур, так как была низкая влагообеспеченность, что отрицательно сказывалось на состоянии растений. У сои развитие задерживалось – к концу второй декады июня отмечалась фаза «примордиальных листьев – 1-го тройчатого листа». Растения сои увядали, усыхала корневая система. Погодные условия в период вегетации способствовали слабому развитию фузариоза. 46 Заболевание отмечено с фазы всходов, 3 июня, в виде пятен на семядолях, листьях, а с фазы 1-го тройчатого – в виде корневых гнилей (Фитосанитарный…, 2009). В июле месячная норма осадков составила 123 мм, что в пределах нормы. С дождём за весь период было 25-30 дней. Агрометеорологические условия для роста и развития сельскохозяйственных культур были неблагоприятны. Недостаток влаги в почве, особенно в июне, отрицательно повлиял на урожайность всех культур. В августе сумма температур составила 18,8°С, что в пределах среднемноголетних значений. Сумма осадков составила 113 мм – это 97% климатической нормы. Сложившиеся погодные условия во 2 и 3-й декадах августа способствовали развитию септориоза на сое. Единичные пятна на растениях отмечены с 15 июля. К концу августа было поражено 80% растений (Фитосанитарный…, 2009). Осень 2008 года характеризовалась значительными колебаниями температурного режима и неравномерным распределением осадков в течение периода. Среднемесячная температура воздуха в сентябре составила 10,4°С тепла. За месяц сумма осадков составила 48 мм – это 63% многолетней нормы. Повышенный температурный режим был благоприятен для развития сои. Теплая погода и прошедшие дожди способствовали наливу зерна и созреванию бобов. Для октября была характерна погода с повышенными температурами. Все три декады октября были теплыми. Среднемесячная температура воздуха составила 2,4°С тепла, что на 1°С выше среднемноголетней. Сумма осадков составила 37 мм – 168% климатической нормы. Теплая погода способствовала созреванию бобов сои среднего и верхнего ярусов. В целом, агрометеорологические условия 2008 года для роста и развития сои были не совсем благоприятные. Недостаток влаги в почве, особенно в июне, отрицательно повлиял на урожайность данной культуры. Агрометеорологические условия в 2009 году для проведения весенних полевых работ складывались благоприятно. 47 В первой декаде мая сохранялась преимущественно теплая, сухая и ветреная погода. Среднемесячная температура воздуха составила 13,8°С, что на 4°С выше среднемноголетних значений. Осадки составили 52 мм, это 118% климатической нормы. Средняя температура на глубине почвы 10 см составила 10-13°С. Декада была ветреной. Ветры способствовали испарению влаги из почвы. В пахотном слое запасы продуктивной влаги составляли 30-40 мм, в полуметровом 70-120 мм, что составило 50-80% от НПВ. Погодные условия во время посевных работ на соевых полях характеризовались неустойчивым режимом температуры. В начале третьей декады отмечалась умеренно дождливая погода с преобладанием ветров северовосточного направления. Летний сезон 2009 года характеризовался дождливой и прохладной погодой. Сумма положительных температур за летний период с температурами выше +15°С составила 1870°С – это на 110°С больше среднемноголетнего. Дожди летом шли часто, временами они были очень интенсивными. За три летних месяца число дней с дождем более 1 мм составило 45-57 дней. Преобладание облачной погоды и частые дожди сдерживали уход за посевами, обработку полей ядохимикатами и гербицидами. Июнь характеризовался холодной и дождливой погодой. В целом месяц был холоднее нормы на 1°С. Дожди шли часто, в июне количество дней с осадками более 1 мм составило 17-25 дней. Наиболее дождливыми были первая и вторая декады июня, дожди шли ежедневно. Сумма осадков за месяц составила 206 мм, 210% климатической нормы. Агрометеорологические условия для роста и развития сои в первой декаде июня были удовлетворительными. Во второй декаде были напряженными, на полях преобладало сильное увлажнение почвы. В третьей декаде сельскохозяйственные работы велись с перерывами из-за дождей. Однако такие 48 условия способствовали широкому распространению таких болезней, как бактериоз, аскохитоз и др. (Фитосанитарный…, 2010). В июле преобладала преимущественно теплая погода с неустойчивым температурным режимом и неравномерным распределением осадков. Средняя температура воздуха за месяц составила 19,6°С, что на 1°С ниже среднемноголетних значений. Дожди были частыми, в отдельные дни носили ливневый характер и сопровождались грозами. За месяц осадков выпало 128 мм – это 103% климатической нормы. Во второй декаде июля в связи с частыми дождями на полях поддерживалось сильное, а в отдельные дни избыточное увлажнение почвы. Низкие температуры воздуха в первой декаде июля сдерживали развитие сои. Высокая относительная влажность воздуха была благоприятной для появления болезней на сое (корневые гнили, септориоз, пероноспороз, бактериоз) (Фитосанитарный…, 2010). Август был относительно тёплым, особенно его первая декада, которая была теплее на 2-3°С, вторая – в пределах среднемноголетней, третья декада ниже среднемноголетней на 4°С. Среднемесячная температура воздуха составила 18,4°С, что на 1°С ниже климатической нормы. В течение месяца осадки распределялись неравномерно, в отдельных районах носили ливневый характер. Их месячная норма составила 102 мм – это 87% климатической нормы. Погодные условия были благоприятны для развития грибных и бактериальных болезней (Фитосанитарный…, 2010). Осень 2009 года характеризовалась теплой погодой с неустойчивым температурным режимом и неравномерным распределением осадков. Сентябрь этого года характеризовался теплой погодой и неравномерным распределением осадков. Первая и третья декады сентября были теплыми. Осадки выпадали в каждой декаде. Всего за месяц сумма осадков составила 45 мм, что на 32 мм ниже средней многолетней. 49 Октябрь был теплым. Среднемесячная температура воздуха составила 2°С, что на 1°С выше среднемноголетней. Осадки прошли в начале месяца, в конце второй декады и в первой пятидневке третьей декады октября. Сумма осадков за месяц составила 9 мм, что составляет 41% нормы. Весна в 2010 году была поздней, с резкими перепадами температур и сильными ветрами. Первая половина весны была холодной, со второй декады мая очень теплой. Среднемесячная температура за май составила 13,5°С, что на 3°С выше среднемноголетней. Сумма осадков за месяц составила 40 мм – это в пределах климатической нормы. Летний сезон 2010 года характеризовался теплой и дождливой погодой. Средняя температура воздуха за июнь составила 21,8°С, что на 5°С выше среднемноголетней. Дожди летом шли часто, временами они были очень интенсивными. Меньше осадков выпало в первой и третьей декадах июня. Осадков за месяц выпало 40 мм – это 41% от климатической нормы. Июнь характеризовался повышенным температурным фоном. Первая и вторая декады июня были выше нормы на 2-5°С, третья на – 7°С. Были отмечены первые признаки заболевания фузариоза в третьей декаде июня в виде единичных пятен на пирамидальных листьях. Июль характеризовался теплой погодой и неравномерным распределением осадков. Сумма осадков за месяц составила 267 мм –это 215% от многолетней нормы. Во второй декаде июля отмечены единичные пятна септориоза (фаза бутонизации) и пероноспороза. В августе погода была теплой и дождливой. Первая и вторая декады августа были в пределах нормы. Температура за месяц была в пределах среднемноголетних значений и составила 17,8°С. Осадков за август выпало 177 мм – это 151% климатической нормы. За месяц наблюдалось 15-18 дней с дождем. У сои продолжалось образование бобов. Высота растений 80-90 см. Состояние посевов сои хорошее и удовлетворительное. Запасы продуктивной влаги составили 30-60 мм – 60- 90% нормы от наименьшей полевой влагоёмкости. 50 Осень 2010 года характеризовалась преимущественно теплой, сухой погодой с неравномерным распределением осадков. Сентябрь характеризовался теплой погодой. Среднемесячная температура воздуха составила 11,1°С, что в пределах многолетней нормы. Осадки распределялись неравномерно, прошли преимущественно во второй и третьей декадах. Сумма осадков составила 25 мм – это 33% климатической нормы. В октябре преобладала преимущественно теплая погода. Среднемесячная температура воздуха составила 1,4°С, что в пределах среднемноголетних значений. Сумма осадков за месяц составила 16 мм – это 73 % климатической нормы. Агрометеорологические условия для роста и сельскохозяйственных культур были неблагоприятными и результате дождливого продолжительного периода развития сложными. В произошло опасное агрометеорологическое явление «переувлажнение почвы», что сказалось на росте и развитии сельскохозяйственных культур. В третьей декаде августа агрометеорологические условия для развития сельскохозяйственных растений и проведения полевых работ были неблагоприятными. На посевах сои отмечались местами лиманы, листья нижнего и среднего ярусов пожелтели, местами наблюдалась гибель растений. Весна 2011 года была поздней, затяжной, с резкими перепадами температур, неравномерным распределением осадков. Первая половина весны была преимущественно сухой, вторая – дождливой. Осадки за май составили 91мм – это 202%. Среднемесячная температура за май составила 11,7°С, что выше среднемноголетней на 2°С. Посев сои был проведён в оптимальные агротехнические сроки. Влагообеспеченность и теплообеспеченность растений были достаточными. Летний период 2011 года характеризовался необычно тёплой погодой. Лето началось раньше климатических сроков. В течение летнего периода относительная влажность воздуха повышалась до 90%. Из-за частых и интенсивных дождей полевые работы велись с перерывами. 51 Июнь характеризовался тёплой и дождливой погодой. Среднемесячная температура воздуха составила 17,5°С тепла, что выше нормы на 1°С. Осадков за месяц выпало 83 мм – это 85% многолетней нормы. Условия для проведения полевых работ были благоприятные. У сои проходила фаза «3-5 настоящий лист». Высота растений 13-17см. В июле преобладала жаркая погода с ливневыми дождями. Средняя за месяц температура воздуха превысила многолетние значения на 2°С и составила 22,6°С. Месячная сумма осадков составила 71 мм, это 57% климатической нормы. 31 июля 2011 года в связи с происхождением активного атмосферного фронта в Благовещенском районе наблюдалось опасное метеорологическое явление «смерч». В июле у сои наблюдались фазы цветения и образования бобов. Высота растений 45-55 см, состояние сои хорошее и отличное. Тепла для её развития было достаточно. Во второй декаде июля погодные условия способствовали раннему проявлению септориоза на примордиальных листьях сои. Первые признаки пероноспороза проявились в конце 2-й декады июля, аскохитоз отмечался на тройчатых листьях. В августе температура воздуха составила 19,5°С, что выше среднемноголетней на 2°С. Все три декады были с осадками. Сумма осадков за месяц составила 92 мм, это 79% многолетней нормы. У сои продолжалось образование бобов, у ранних сортов началось созревание бобов. Состояние посевов хорошее и отличное. Развитие сои проходило при достаточном количестве тепла и влаги, сумма активных температур на конец декады составила 2299°С. Сухая погода июня сдерживала развитие и распространение бактериоза, существенного влияния на вредоносность болезни не оказала. Фузариоз не получил большого развития, отмечен на бобах и семенах сои. Средний процент поражения бобов 0,2%, семян – 0,1%. Сентябрь 2011 года характеризовался холодной, с осадками погодой. Среднемесячная температура воздуха составила 10,4°С, что в пределах 52 многолетней нормы. Осадки выпадали в каждой декаде и за месяц составила 26 мм – это 34% нормы. Агрометеорологические условия для сои были благоприятными. В третьей декаде сентября после дождей наблюдалось сильное увлажнение почвы. Повышенный температурный режим был благоприятен для созревания сои. Уборку сои начали с 12 сентября. Октябрь характеризовался тёплой, местами рекордно тёплой, с осадками погодой. Среднемесячная температура воздуха составила 3,8°С, что выше климатической нормы на 2,8°С. Сумма осадков за месяц составила 5 мм – это 23% многолетней нормы. Таким образом, метеорологические условия в годы проведения исследований различались по температурному режиму, количеству выпавших осадков, длине безморозного периода, что, безусловно, отразилось на росте и развитии сои, а также оказало влияние на развитие и распространение фитопатогенов. 2.2 Материалы и методы исследований Объектами исследования в 2008-2011 годах были сорта культурной сои (Glycine max (L.) Merr.) селекции ВНИИ сои: Соната, Гармония, Даурия и дикорастущая форма (Glycine soja): КА-1344. Выращиваемые на Дальнем Востоке сорта сои относятся к маньчжурскому подвиду. Они имеют толстый прямой стебель кустовой формы высотой до 1 м, с крупными тройчатыми листьями и сильным опушением всего растения рыжего цвета. Листья узколистной или широкоовальной формы. Цветки фиолетовые или белые, очень мелкие. Семена овальные или шаровидные. Масса 1000 семян – 110 – 250 г (Золотницкий, 1962; Черноголовин, 1971; Лавриненко, 1978; Щегорец, 2002). Сорт сои Соната выведен методом гибридизации с последующим многократным индивидуальным отбором с оценкой по потомству (педигри). 53 Авторы – Л.К. Малыш, Т.П. Рязанцева, Н.Д. Фоменко, Г.Н. Беляева, А.П. Дымова, Е.Н. Мельникова, К.Е. Малышев. Сорт предназначен для возделывания в умеренно холодных соесеющих регионах. Сорт средневысокорослый: в среднем высота растений – 72 см. Куст компактный, слабоветвистый, с хорошо выполненной верхушкой. Листья овально-заострённые, кончик листа заострённый, окраска листа зелёная. Цветок средней величины, фиолетовый, кисти укороченные, 2-8-цветковые. Бобы коричневые, слабоизогнутые, 2-4семенные, не растрескиваются. Опушение рыжее с желтоватым оттенком. Высота прикрепления нижних бобов – 14,5 см. Семена средней величины, округлые и округло-овальные, окраска семян светло-жёлтая, белёсая, без блеска, с матовым оттенком, пигментация не наблюдалась. Рубчик средний, линейный и слабоовальный, цвета семян иногда темнее. У основания рубчика имеется слабовыраженная точка. Масса 1000 семян – 136 г, с колебаниями по годам 120145 г, содержание белка в семенах 39,2%, жира – 20,2%. Продолжительность вегетации – 96-100 дней. Урожайность 21-29 ц/га. Внесен в Государственный реестр селекционных достижений с 1998 года по дальневосточному региону (Щегорец, 2002; Технологии…, 2011). Сорт Соната сочетает в себе как скороспелость, так и высокую продуктивность, что обусловило широкое распространение данного сорта в производстве. Даурия – высокоурожайный сорт, относится к апробационной группе communis Enk. Авторы сорта: Н.С. Слободяник, Г.С. Беляева, А.П. Дымова, Е.Н. Мельникова, Н.Д. Фоменко, Г.Н. Лавриченко. Внесен в Государственный реестр селекционных достижений по дальневосточному региону с 2003 года. Сорт среднеспелый, период вегетации 106 дней. Растение средневысокое (50-100 см), куст компактный. Число междоузлий от 8 до 12, число цветков на цветоносе 4-9, цветок фиолетовый. Форма листа яйцевидная. Бобы серые, слабоизогнутые, заострённые, 2-3-х семенные. Форма семян округлая, почти шаровидная. Окраска семян жёлтая, рубчик цвета семени, линейный. 54 Масса 1000 семян – 195,7 г, с колебаниями по годам 182,5-214,5 г. Содержание в семенах белка 39,0%, жира – 20,7%. Урожайность сорта 28-35 ц/га. Сорт предназначен для зернового и пищевого использования (Тихончук, Оборская, 2010; Технологии…, 2011). Сорт Гармония относится к маньчжурскому (manshurica) подвиду апробационной группы flavida Enk. Авторы сорта: Л.К. Малыш, Т.П. Рязанцев, Г.Н. Беляева, А.П. Дымова, Е.Н. Мельникова, Н.Д. Фоменко, Г.П. Лавриченко. Внесен в Государственный реестр селекционных достижений по дальневосточному региону с 2003 года. Продолжительность периода вегетации 102 дня, с колебаниями по годам от 100 до 108 дней. Растения средней высоты – 51,4-69,7 см. Число междоузлий 7-13, опушение рыжее, редкое. Лист узкий, копьевидный. Окраска цветка белая. Бобы слабоизогнутые с заострённой верхушкой, 2-4-х семенные. Высота прикрепления нижнего боба 15 см. Семена жёлтые, иногда с зеленоватым оттенком, слабо блестящие, средней крупности. Масса 1000 семян 153,7 г. Содержание в семенах белка 37,5-39,6%, жира – 19,3-22,0%. Урожайность семян 27 – 34 ц/га. (Тихончук, Оборская, 2010; Технологии…, 2011). Сорт Гармония, относящийся к среднеспелой группе, превышает по урожайности сорт Соната в среднем на 5 ц/га. Форма G. soja КА 1344 из Архаринского района - однолетнее растение с тонкими, обильно ветвящимися, вьющимися стеблями, покрытыми жесткими, редкими рыжеватыми волосками. Высота растения 124 см. Листья тройчатые, опушенные, с ланцетными заостренными листочками. Окраска венчика цветка фиолетовая. Бобы мелкие (0,7-2,5 см длиной и 4-5 мм шириной), темно-бурые, покрытые жесткими волосками. При созревании они растрескиваются и разбрасывают очень мелкие семена. Окраска кожуры семян темно-умбровая. Масса 1000 семян – 30,4 г. Содержание белка – 45,3%, содержание жира –11,0%. Продолжительность периода вегетации составляет 95 дней (Ала, Гамолин, 1984; Ала и др., 2000; Ала, Тильба, 2005). 55 Форма КА 1344 широко используется в генетико-селекционных исследованиях, это обусловлено высоким содержанием белка в семенах и её повышенной устойчивостью к неблагоприятным экологическим факторам климата. Полевые исследования проводили в 2008 – 2011 гг. на опытном поле ДальГАУ. Сою высевали вручную на глубину 4-5 см, широкорядным способом 3 х 45 см. Такой способ посева способствует созданию естественного инфекционного фона. Повторность 4-кратная, расположение делянок рендомизированное. В процессе роста сои отмечали следующие фазы развития: всходы, появление первого и третьего тройчатого листа, цветение, бобообразование, созревания. Наступление фаз развития растений сои отмечали по методике ГСИ (Методика…, 1985). Растения в фазу учета просматривали непосредственно в поле. Все учёты проводили по методике ВИР (Корсаков и др., 1979). Частоту встречаемости болезни выражали в процентах. Степень поражения или процент развития болезни характеризуется количеством пятен (табл.1). Таблица 1 – Шкала для оценки пораженности растений сои болезнями (Голубев, 1976; Корсаков и др., 1979) Оценка Степень поражения по 5балльной шкале по классификатору ВИР % Иммунологическая характеристика Оч. слабое 1 1 1-10 УУ – высокоустойчивый Слабое 2 3 11-25 У – устойчивый Среднее 3 5 26-50 С – среднеустойчивый Сильное 4 7 51-75 В – восприимчивый Оч. сильное 5 9 75-100 ВВ – сильновосприимчивый 56 Поражение корневой системы оценивали по методике ВИЗР (Котова, Степанова, 1979). Для диагностики устойчивости сортов к корневой гнили пользовались шкалой (Голубев, 1976; Корсаков и др., 1979): УУ – высокоустойчивые формы, развитие болезни в различных условиях не превышает 10% (балл 1); У – устойчивые формы, ценные для использования в селекции на устойчивость, поражаемые не более чем на 25% (балл 2); С – средневосприимчивые формы, использование которых в селекционных целях целесообразно из-за других хозяйственно-ценных признаков, поражаемость не превышает 50% (балл 3); В – восприимчивые формы, поражаемость которых превышает 50% в разных условиях заражения и эпифитотий (балл 4); ВВ – сильновосприимчивые формы, процент развития болезни превышает 75% (балл 5) и отмечается массовая гибель растений или существенное снижение урожая. Степень развития болезни вычисляли по формуле: П=∑ (а*б) 100 / к*В, (Корсаков и др., 1979) где: П – процент развития болезни, ∑ – сумма произведений, а – число пораженных растений, б – балл поражения растений, к – общее количество учтенных растений, В – высший балл шкалы учета. В лабораторных условиях осуществляли инокуляцию возбудителем септориоза изолированных листьев сои (Корсаков и др., 1979). Для выделения возбудителя Septoria glycines Hemmi в чистую культуру использовали картофельно-глюкозный агар (Хохряков и др., 1976). При лабораторной оценке пораженности листьев септориозом использовали шкалу (Корсаков и др., 1979): 57 0 – пятен нет, 1 – некрозы или одиночные пятна без спороношения занимают до 10% поверхности листа, 2 – пятна со спороношением занимают 25% листа, 3 – пятна со спороношением занимают 26-50% листа, 4 – пятна со спороношением занимают 51-75% листа, 5 – пятна со спороношением занимают более 75% листа или сплошь покрывают лист. Определение заражённости семян болезнями проводили по ГОСТ 12044-93. Для проращивания семян использовали два слоя фильтровальной бумаги, которую сворачивали в рулон. Рулоны ставили вертикально в сосуды и помещали в термостат при температуре 22-25°С. Просмотр семян проводили в сроки определения всхожести семян по ГОСТ 12038 – 84. В каждой из трёх проб подсчитывали количество семян, заражённых каждой болезнью, и общее количество заражённых семян. Заражённость семян (Х4) в процентах вычисляли по формуле: Х4= N1 / n *100, (Корсаков и др., 1979) где: N1 – суммарное количество заражённых семян в трёх пробах, шт; n – общее количество семян, взятых для анализа, шт. Содержание белка и масла в семенах определяли на ИК-сканере «Nir-5000» в лаборатории ГНУ Всероссийского научно-исследовательского института сои. Выделение растворимых белков для определения активности ферментов Навеску (1 г) растительного материала (корни, листья, семяна) гомогенизировали в фарфоровой ступке при температуре 4 оС 0,15М раствором NaCl (для определения активности каталазы и кислой фосфатазы) или ацетатным буфером (для определения активности пероксидазы) в течение 15 мин. Полученный экстракт центрифугировали в течение 15 мин. при 3000 g. Осадок отбрасывали, а надосадочную жидкость фильтровали через слой капроновой 58 ткани для удаления липидной пленки и использовали для дальнейшего анализа (Методы…, 1987). Определение общего белка в экстрактах проводили методом Лоури (Lowry et al., 1951). Определение активности ферментов Метод определения активности пероксидазы (1.11.1.7) (Бояркин, 1951; Малый…, 1994) основан на образовании окрашенных продуктов при окислении бензидина под действием фермента, содержащегося в растениях, до образования продукта окисления синего цвета (n,n’-диаминдифенилхинона). Активность фермента определяли на фотоэлектроколориметре (КФК-2). Об активности пероксидазы судили по времени развития окраски до определенного значения оптической плотности (значение D выбирают в зависимости от скорости развития окраски в пределах от 0,25 до 0,5). Расчет активности пероксидазы вели по формуле: А D , (Бояркин, 1951) td где: А – активность фермента, выраженная в относительных единицах на 1 г навески за 1 с; D – зарегистрированная в опыте оптическая плотность; t – время, с; d – толщина слоя жидкости, см; α, β, γ – степень разведения: α – отношение количества жидкости, взятой для приготовления вытяжки, мл, к массе навески, г; β – степень дополнительного разведения вытяжки после центрифугирования (если это требовалось); γ – степень постоянного разведения вытяжки в кювете. Активность каталазы (1.11.1.6) определяли газометрическим методом (Методы…, 1987; Иваченко и др., 1997). Метод основан на определении объема кислорода, выделившегося после прибавления Н2О2 к экстракту из растений, содержащего каталазу. Активность каталазы выражается в миллилитрах кислорода, который выделяется под действием фермента из 1 г растительного материала за 3 минуты (учитывали содержание белка в пробе и разбавление). Определение активности кислой фосфатазы (3.1.3.2) (Филиппович и др., 1982; Шаройко и др., 2002) основано на количественном учете неорганического 59 фосфора, образующегося при расщеплении органических фосфорных соединений в строго определенных условиях под действием этого фермента. Удельную активность выражали в миллиграммах отщепленного неорганического фосфора под действием фермента из 1 мг белка, которую определяли по формуле: А Е ав , (Филиппович и др., 1982) сd t где: А – удельная активность фермента; Е – показания ФЭКа (определенное по калибровочной кривой); а – количество инкубационной смеси; в – количество раствора для фотоколориметрирования; с – аликвота из инкубационной смеси; d – количество белка; t – время инкубации, мин. Построение калибровочного графика для определения содержания фосфора осуществляли по методу Фиске-Суббароу (Методы…, 1987). Все анализы осуществляли в двух биологических и трех аналитических повторностях. Статистическую обработку полученных данных проводили методами дисперсионного и корреляционного анализов (Доспехов, 1979; Плохинский, 1970). Достоверность результатов оценивали, используя t – критерий Стьюдента при уровне вероятности (р) 0,05 (Доспехов, 1979; Короневский, 1985). 60 3 ЭНЗИМАТИЧЕСКАЯ АКТИВНОСТЬ В КОРНЯХ И ЛИСТЬЯХ СОИ ПРИ ПОРАЖЕНИИ ПОЧВЕННЫМИ (КОРНЕВЫМИ) ИНФЕКЦИЯМИ 3.1 Видовой состав возбудителей корневой гнили сои и динамика развития болезни Почва по ряду экологических параметров представляет собой промежуточную среду жизни между наземно-воздушной и водной. Буферные свойства почвы препятствуют резкому колебанию температуры и влажности, поэтому она является достаточно стабильной и благоприятной средой для существования почвенных организмов. Наиболее вредоносным заболеванием на посевах сои является корневая гниль. По данным исследователей, видовой состав возбудителей корневой гнили характеризуется большим разнообразием и неодинаковым соотношением видов как в зонах Российской Федерации (Жуковская, 1977; Подкина, 1981, 1982), так и за рубежом (Hildebrand, Koch, 1974; Sato, Kitazawa, 1980; Reeve, 1983). Микрофлора пораженных корней сои в Приамурье разнообразна и формируется в основном из микрофлоры почвы и семян. Л.К. Дубовицкой (1987) было установлено, что основными доминирующими видами являются грибы рода Fusarium, они составляют до 51,3% от общего количества микрофлоры корней сои. В результате проведенных исследований было установлено, что процентное соотношение возбудителей, выделенных в разные фазы развития сои в среднем за три года, было непостоянным. Наиболее часто изолировали из пораженных корней F. solani (47,3-69,4%), F. oxysporum (20,1-23,2%), значительно реже F. moniliforme (1,5-4,3%), очень редко F. gibbosum и прочие сопутствующие виды: Actinomyces, Penicillium, Mucor, Trichoderma, Sporotrihum (рис. 11). 61 Mycelia sterilia, Alternaria, Всходы Цветение 12,73 24,6 2,3 47,3 4,2 1,5 56,8 5 2,1 0,7 22,3 20,1 Бобообразование 4,3 2,7 2 8,2 23,2 59,4 Рисунок 11 – Количественное соотношение грибов, выделенных из пораженных корней в разные фазы развития сои, %, 2008 – 2010 гг. Экологические факторы среды оказывают огромное влияние на состояние растения-хозяина, на возникновение и течение патологических процессов в нем (Стэкмен, Харрар, 1959; Заостровных, Дубовицкая, 2003). Для выяснения влияния погодных условий на развитие заболевания были проанализированы сумма осадков и величина среднесуточной температуры воздуха за вегетационный период. Погодные условия в 2008 году по температурному показателю за вегетационный период были выше среднемноголетних на 2 оС (приложения 1, 2). Количество осадков в июне составило 38 мм, что на 60 мм ниже среднемноголетних данных. Сложившиеся погодные условия препятствовали 62 заражению растений, так как длительная засуха затрудняет распространение патогенов. Высокие температуры в июле и выпавшие осадки (98% от многолетней нормы) привели к развитию заболевания в фазе цветения, первоначальный уровень повысился в 2 раза. В фазе образования бобов развитие болезни продолжилось и составило 27,5% (рис. 12). Развитие корневой гнили, % 35 33 30 25 21 20 15 10 9 5 0 1-й тройчатый лист цветение бобообразование 25 Средняя температура воздуха, оС 250 20 206 200 15 150 128 10 102 100 52 5 Количество осадков, мм 300 50 0 0 май июнь июль август 2008 2009 2010 среднемноголетнее 2008 2009 2010 среднемноголетнее Средняя температура воздуха и количество осадков Рисунок 12 – Динамика развития корневой гнили сои в зависимости от метеорологических условий, %, 2008 – 2010 гг. 63 Самым благоприятным для развития корневой гнили был 2009 год. В июне сложилась дождливая и прохладная погода, это способствовало ослаблению растений и повышению восприимчивости к патогенам. Теплая погода с неравномерным распределением осадков в июле и обилие осадков в августе способствовали распространению корневых гнилей. Условия, неблагоприятные для интенсивного роста и развития сои, являлись благоприятными для патогена. В фазе цветения развитие болезни составило 21,2%, к периоду уборки – 33,0% (рис. 12). В 2010 году погодные условия по температурному режиму были выше среднемноголетних значений: июнь характеризовался жаркой и сухой погодой, в июле выпало рекордное количество осадков (267 мм), что составило 215% от климатической нормы. Известно, что развитие патогенов находится в зависимости от влажности, слишком высокое содержание влаги в почве для возбудителей корневой гнили вредно, так как затрудняет снабжение их кислородом. В августе установилась дождливая погода с неустойчивым температурным режимом. При таких условиях развитие болезни составило 15,5% в фазе цветения, максимум (28,3%) приходился на фазу образования бобов (рис. 12). Статистическая обработка результатов показала, что наибольшее поражение сои корневыми гнилями наблюдается при низких показателях относительных значений накопленных сумм температур. Коэффициенты корреляции между уровнем температуры и развитием корневых гнилей варьировали от r = -0,39±0,06 (сорт Соната) до r =-0,81±0,04 (сорт Гармония). Корреляционная зависимость между степенью развития болезни и количеством осадков была положительной: сильной для сортов Соната (r = 0,85±0,06) и Даурия (r = 0,70±0,02), средней для сорта Гармония (r = 0,58±0,02). Изучение распространенности и развития корневой гнили в динамике по фазам развития сои показало, что болезнь проявляется на ранних стадиях при формировании примордиальных листьев и достигает высокого уровня к периоду цветения и началу бобообразования. 64 В фазе первого тройчатого листа распространение болезни составило от 34,2 до 48,3%, при развитии болезни 5,7 – 11,0% (среднее по сортам) (рис. 13, 14, приложение 3). К фазе цветения болезнь распространялась быстрее и превысила первоначальный уровень в 1,4 – 1,5 раза. Частота встречаемости в период цветения не превышала 51,7 – 66,7%, при развитии корневой гнили 8,5 – 26,0% (рис. 13, 14, приложение 3). В фазе бобообразования в среднем за три года исследований распространенность болезни составила 64,2 – 81,7%, развитие соответственно 11,5 – 50,6% по сортам (рис. 13, 14, приложение 3). % 90 80 70 60 50 40 30 20 10 0 1-й тройчат.лист цветение Гармония Даурия бобообразование Соната Рисунок 13 – Динамика распространенности корневой гнили сои, %, 2008 – 2010 гг. Анализ результатов исследования за 2008 – 2010 гг. показал, что различные сорта сои поражались корневой гнилью по-разному. Среднеспелый сорт Гармония (2 балл поражения) оказался более устойчивым к поражению корневыми гнилями, в период максимального развития заболевания (фаза бобообразования) степень поражения варьировала по годам в пределах 10,0 – 13,0% (приложение 3). Сорт Даурия (среднеспелый) зарекомендовал себя как среднеустойчивый, в фазе бобообразования он имел 3 балл поражения, процент развития болезни составлял 23,5 – 33,0% (приложение 3). 65 Наиболее восприимчивым к заболеванию оказался раннеспелый сорт Соната (4 балл поражения), в фазе бобообразования развитие болезни доходило до 53,0% при распространенности 90,0% (приложение 3). % 60 50 40 30 20 10 0 1-й тройчат.лист цветение Гармония бобообразование Даурия Соната Рисунок 14 – Динамика развития корневой гнили сои, %, 2008 – 2010 гг. Таким образом, поражению растений корневыми гнилями способствует теплая с достаточным количеством осадков погода. Выявлена положительная корреляционная зависимость между степенью развития болезни и количеством осадков (r = 0,58 – 0,85) и отрицательная с температурой воздуха (r = -0,39 – 0,81). Распространенность и развитие заболевания увеличивается по фазам развития сои, наиболее интенсивно поражаются растения от фазы всходов до цветения, максимум приходится на фазу бобообразования. Наибольшей устойчивостью к корневым гнилям в условиях южной зоны Амурской области обладает сорт Гармония, наименьшей – сорт Соната. 3.2 Изменение активности пероксидазы, каталазы и кислой фосфатазы в корнях сои при поражении возбудителями корневой гнили Подземные части растений живут в гораздо более сложной биологической среде, чем побеги, так как в почве обычно обитает большое количество грибов, бактерий, актиномицетов, простейших, нематод, насекомых и других организмов. 66 Все эти организмы взаимодействуют друг с другом, и, кроме того, на их численность влияют химические и физические условия почвы (Тарр, 1975). Воздействие патогена вызывает значительные изменения в функциональном состоянии растения, а это прежде всего отражается на уровне ферментативной активности. Удельная активность пероксидазы в корнях, пораженных корневой гнилью, как увеличивается, так и уменьшается относительно контроля (здоровые корни). В частности, установлено увеличение удельной активности пероксидазы в пораженных корнях сортов Гармония и Даурия в фазе первого тройчатого листа. Наоборот, у сорта Соната активность пероксидазы снижается по сравнению с контролем (рис. 15, приложение 4). В фазе цветения удельная активность пероксидазы увеличивается в инфицированных корнях сорта Гармония (в 1,5 раза) с 1384 до 2076 ед/мг белка х 10-2. У сортов Даурия и Соната активность фермента соответственно в 1,2 и 1,3 раза ниже, чем в контроле (рис. 15, приложение 4). Результаты исследования показали, что активность пероксидазы в фазе бобообразования у сорта Гармония в 1,7 раза выше, чем в контроле. Вероятно, высокая активность пероксидазы в корнях устойчивых сортов сои наряду с другими механизмами защиты обеспечивает способность этих растений противостоять действию возбудителя. Наименьшая активность пероксидазы в наших исследованиях отмечена в пораженных корнях сорта Соната (454 ед/мг белка х 10-2) (рис. 15, приложение 4). Как свидетельствуют наши наблюдения, активность пероксидазы в пораженных корнях сорта Соната ниже, чем в контроле во все фазы развития сои. Согласно литературным данным (Серова и др., 1982), такой характер изменения активности пероксидазы свойствен восприимчивым к заболеваниям растениям. 67 200 180 160 % к контролю 140 120 100 80 60 40 20 0 1-й тройчат.лист контроль цветение Гармония Даурия бобообразование Соната Рисунок 15 – Удельная активность пероксидазы (% к контролю) в корнях сои при поражении корневой гнилью, 2008 – 2010 гг. На основе полученных данных установлена отрицательная корреляционная зависимость между степенью поражения корней и активностью пероксидазы в пораженных корнях сои: средняя в фазе первого тройчатого листа и сильная в фазы цветения и бобообразования (табл. 2). Чем выше активность фермента, тем ниже был балл поражения корней сои. Таблица 2 – Корреляционная зависимость между степенью поражения корневой гнилью и удельной активностью пероксидазы в пораженных корнях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения корневой 1-й тройчатый лист гнилью – удельная активность цветение пероксидазы в корнях сои бобообразование tтеор при Р0,95=2,01 Коэффициент корреляции - 0,51 ± 0,08 - 0,74 ± 0,06 - 0,86 ± 0,05 Каталаза встречается в клетке вместе с пероксидазой. Её биологическая роль заключается не только в защите клеток от образующейся в ходе метаболизма перекиси водорода, но и в снабжении растения кислородом. 68 Результаты нашего исследования показали, что в фазе первого тройчатого листа удельная активность каталазы в пораженных корнях сорта Гармония снижается в 1,5 раза, но возрастает у сортов Соната (в 1,5 раза) и Даурия (в 1,2 раза) (рис. 16, приложение 5). В фазе цветения активность каталазы в пораженных корнях у сортов Гармония и Даурия несущественно отличается от контроля, а у сорта Соната она в 1,3 раза выше, чем в здоровых корнях (рис. 16, приложение 5). В фазе бобообразования у устойчивого сорта Гармония активность каталазы в инфицированных корнях в 1,4 раза ниже, чем в здоровых (рис. 16, приложение 5). 180 160 % к контролю 140 120 100 80 60 40 20 0 1-й тройчат.лист цветение контроль Гармония бобообразование Даурия Соната Рисунок 16 – Удельная активность каталазы (% к контролю) в корнях сои при поражении корневой гнилью, 2008 – 2010 гг. У сорта Даурия, занимающего промежуточное положение по устойчивости к корневым гнилям, активность фермента достоверно не изменялась по сравнению со здоровыми корнями (рис. 16, приложение 5). У сильно поражаемого корневой гнилью сорта Соната активность фермента в 1,3 раза выше, чем в контроле (рис. 16, приложение 5). Повышение активности каталазы привело к снижению концентрации перекиси водорода, а это лимитирует реакции, катализируемые пероксидазой. Не исключено, 69 что подобные изменения активности каталазы у сорта Соната способствуют формированию совместимых отношений в комбинации хозяин (соя) – патоген. Изучение зависимости поражения сои корневой гнилью от активности каталазы показало, что между указанными параметрами существует положительная корреляция средней силы в фазе первого тройчатого листа и сильная в фазе цветения: чем ниже активность каталазы, тем ниже балл поражения (табл. 3). Однако в фазе бобообразования была выявлена сильная отрицательная взаимосвязь между степенью поражения растений корневой гнилью и активностью каталазы в пораженных корнях сои, при которой с увеличением активности фермента происходит снижение устойчивости (табл. 3). Таблица 3 – Корреляционная зависимость между степенью поражения корневой гнилью и удельной активностью каталазы в пораженных корнях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения корневой 1-й тройчатый лист гнилью – удельная активность цветение каталазы в корнях сои бобообразование tтеор при Р0,95=2,01 Динамика изменения удельной активности Коэффициент корреляции 0,55 ± 0,07 0,78 ± 0,05 - 0,74 ± 0,06 кислой фосфатазы в инфицированных корнях сорта Гармония следующая: в фазе первого тройчатого листа она снижается на 24%, в фазах цветения и бобообразования достоверного отличия активности этого фермента между больными и здоровыми растениями не выявлено (рис. 17, приложение 6). В пораженных корнях сорта Даурия активность кислой фосфатазы во всех фазах развития ниже, чем у здоровых растений (рис. 17, приложение 6). В пораженных корнях сорта Соната активность кислой фосфатазы в фазе первого тройчатого листа снижается, а затем в фазах цветения и бобообразования увеличивается в 2,0 и 1,6 раза соответственно (рис. 17, приложение 6). Рост активности кислой фосфатазы говорит о возможности усиления литических 70 процессов в пораженных корнях, что может негативно отразиться на гомеостазе фосфорных эфиров. На основании литературных данных (Лисицын, Кропотов, 2002) и наших исследований можно полагать, что под действием стрессового фактора в корнях сильнопоражаемых растений активность кислой фосфатазы увеличивается, а у слабопоражаемых незначительно снижается или приближается к контролю. 220 200 180 % к контролю 160 140 120 100 80 60 40 20 0 1-й тройчат.лист цветение контроль Гармония Даурия бобообразование Соната Рисунок 17 – Удельная активность кислой фосфатазы (% к контролю) в корнях сои при поражении корневой гнилью, 2008 – 2010 гг. Корреляционный анализ полученных данных позволил выявить сильную положительную связь между степенью поражения корневой гнилью и активностью кислой фосфатазы в фазе первого тройчатого листа. При этом в фазах цветения и образования бобов устойчивость сои к корневой гнили слабо связана с активностью кислой фосфатазы в пораженных корнях (табл. 4). Проанализированные корреляционные зависимости между активностью пероксидазы, каталазы, кислой фосфатазы и степенью поражения сои корневой гнилью на разных стадиях онтогенеза позволили выявить тесные взаимосвязи, при которых с изменением активности ферментов (убывание или возрастание) возрастает устойчивость. 71 Таблица 4 – Корреляционная зависимость между степенью поражения корневой гнилью и удельной активностью кислой фосфатазы в пораженных корнях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения корневой 1-й тройчатый лист гнилью – удельная активность цветение кислой фосфатазы в корнях сои бобообразование tтеор при Р0,95=2,01 Корреляционная связь несущественна tфакт < tтеор Коэффициент корреляции 0,69 ± 0,06 0,26 ± 0,08 0,13 ± 0,09 Таким образом, инфицирование корней сои корневой гнилью приводит к увеличению активности пероксидазы и снижению каталазы у устойчивого сорта Гармония (активность кислой фосфатазы изменяется несущественно) и, наоборот, к снижению активности пероксидазы и росту активности каталазы и кислой фосфатазы у восприимчивого сорта Соната. 3.3 Изменение активности пероксидазы, каталазы и кислой фосфатазы в листьях сои при поражении возбудителями корневой гнили Известно (Ильинская и др., 1991; Граскова и др., 2004; Яруллина и др., 2005), что заражение вызывает изменение метаболизма в органах, отдаленных от места внедрения и локализации патогена. В связи с этим нами была определена активность ферментов в листьях сои при поражении корней корневыми гнилями. Важнейшим ферментом, задействованным в формировании защитной реакции растений на внедрение патогена и, в частности, в регуляции уровня перекиси водорода в клетках, является пероксидаза. Развитие заболевания приводит к увеличению активности пероксидазы у устойчивого сорта Гармония не только в корнях, но и в листьях пораженных растений. Повышение удельной активности пероксидазы у сорта Гармония свидетельствует о его высоком адаптивном потенциале (табл. 5, приложение 7). 72 У сортов Соната и Даурия активность пероксидазы в листьях пораженных растений, так же как и в корнях, ниже, чем в здоровых (табл. 5, приложение 7). Важно заметить, что инфицирование растений через корни приводит к изменению активности фермента в листьях, которые не контактировали с патогеном. Следовательно, можно сделать предположение об участии пероксидазы в развитии системной устойчивости сои при заражении корневыми гнилями. Это согласуется с существующими представлениями о том, что пероксидазу можно рассматривать в качестве маркера системной устойчивости растений при патогенезе (Серова и др., 1982). Каталаза вместе с другими оксидоредуктазами направляет метаболизм клетки на создание неблагоприятных условий для патогена. Известно о различных ответных реакциях каталазы у устойчивых растений: от сильной активации у овса при заражении корончатой ржавчиной до снижения активности у пшеницы, пораженной стеблевой ржавчиной (Коно, 1961; Нилова, Егорова, 1948). Под влиянием заражения сои корневой гнилью активность каталазы в листьях сорта Гармония в фазе цветения повышается, а в фазе бобообразования снижается, при этом в пораженных корнях активность фермента в данные фенологические фазы была ниже, чем в контроле (табл. 5, приложение 8). Активность каталазы в листьях пораженных растений сортов Соната и Даурия в фазах цветения и бобообразования снижается, в пораженных корнях сорта Соната – увеличивается, а в корнях сорта Даурия – увеличивается в фазе цветения и снижается в фазе бобообразования (табл. 5, приложение 8). В наших исследованиях активность кислой фосфатазы, выделенной из корней и листьев разных сортов сои, под действием патогена изменялась поразному. У сорта Гармония активность кислой фосфатазы в пораженных корнях остается на уровне здоровых, а в листьях снижается; у сорта Даурия – снижается в корнях и в листьях; у сорта Соната – повышается в корнях, а в листьях снижается (табл. 5, приложение 9). 73 Таблица 5 – Удельная активность ферментов в листьях сои при поражении корневой гнилью, ед/мг белка, 2008 – 2010 гг. цветение 142 ±8 116± 10 82±6 60±3 207±20 154±16 бобообразование 204± 52 122± 36 121±12 95±6 302±31 119±24 цветение 134 ±9 239± 22 84±4 120±6 165±18 117±26 бобообразование 198± 45 300 ± 14 180±23 168±56 390±37 300±20 цветение 148± 16 104±5 75±15 64±17 118±17 89±15 бобообразование 182± 30 158± 24 149±17 129±12 325±23 216±14 НСР05 контроль пораженные Кислая фосфатаза х 10-6 контроль Соната Гармония Даурия Каталаза х 10-4 пораженные Пероксидаза х 10-3 пораженные Фенологическая фаза контроль Сорт 0,51 0,29 1,68 Проанализировав полученные результаты, можно сделать заключение, что при патогенезе корневой гнилью только активность пероксидазы, выделенной из разных органов сои, изменяется однонаправленно, изменение активности других изучаемых ферментов носит разнонаправленный характер. Так как удельная активность пероксидазы, выделенной из корней и листьев, изменяется сходным образом, то для установления устойчивости к патогену можно использовать любой из этих органов. 74 4 ЭНЗИМАТИЧЕСКАЯ АКТИВНОСТЬ В ЛИСТЬЯХ СОИ ПРИ ПОРАЖЕНИИ ЛИСТО-СТЕБЛЕВЫМИ ИНФЕКЦИЯМИ 4.1 Динамика развития грибных и бактериальных заболеваний и оценка болезнеустойчивости сои Основной средой обитания возбудителей листо-стеблевых инфекций являются вегетативные органы, дополнительной – генеративные органы растений. Распространяются возбудители из года в год благодаря сохранению их на растительных остатках и семенах. Они нуждаются в наличии капельно-жидкой влаги или высокой относительной влажности воздуха (80 – 100%) на всех этапах передачи: в период выделения пикноспор во внешнюю среду, при их распространении и при внедрении в ткани наземных органов растений. Учеты распространенности и развития болезней сои проводили в течение всего вегетационного периода. В результате учетов и наблюдений в годы исследований наиболее широкое распространение получили следующие болезни: септориоз, пероноспороз и бактериоз. Время появления и степень развития болезни, вызываемой тем или иным патогеном сои, в значительной степени определялись метеорологическими условиями года, а также биологическими особенностями самих возбудителей болезни. Септориоз (возбудитель Septoria glycines Hemmi. Syn.) является одним из самых вредоносных заболеваний этой культуры. На степень развития септориоза большое влияние оказали метеорологические условия 2008 – 2011 гг. (приложения 1, 2). Заражению и развитию болезни способствует влажная и теплая погода, которая вызывает споруляцию патогена на первичных некрозах. Конидии распространяются ветром и брызгами от падающих дождевых капель. Оптимальной для развития возбудителя является температура 24-28оС и влажность воздуха 90%. Очень благоприятны для распространения заболевания 75 частые дожди, высокая температура во второй половине июля-августа и обильные росы (Заостровных, Дубовицкая, 2003). Погодные условия в 2008 году оказались самыми благоприятными для развития гриба, несмотря на то что осадки распределялись неравномерно: в июне их было мало (39% от средних многолетних), в июле и августе количество осадков увеличилось, что способствовало повышению относительной влажности воздуха. Развитие септориоза в фазе бобообразования составило 62,2% (рис. 18, приложение 10). 70 Развитие септориоза, % 62,2 60 50 40 30 26,5 20 10 2 0 3-й тройчатый лист цветение бобообразование 300 250 20 200 15 150 123 113 10 100 5 55 38 50 0 Количество осадков, мм Средняя температура воздуха, оС 25 0 май июнь июль август 2008 2009 2010 2011 среднемноголетнее 2008 2009 2010 2011 среднемноголетнее Средняя температура воздуха и количество осадков Рисунок 18 – Динамика развития септориоза сои в зависимости от метеорологических условий, %, 2008 – 2011 гг. 76 Самые неблагоприятные условия для развития патогена сложились в 2010 году. В период всходов и образования тройчатых листьев температура на поверхности почвы достигла 50-63оС, развитие септориоза находилось в депрессии. Распространение и заражение растений началось в первой декаде июля после выпадения осадков. Развитие заболевания долгое время было невысоким и лишь в конце августа возросло до 24,6% (рис. 18). Септориоз у сои проявляется очень рано, развитие начинается с семядолей и примордиальных листьев и продолжается в течение всего вегетационного периода. Распространенность септориоза на листьях культурной сои в фазе цветения составила 51,2 – 79,5%, развитие болезни – 14,4 – 28% (рис. 19, 20, приложение 10). % 120 100 80 60 40 20 0 3-й тройчат.лист Гармония цветение Даурия бобообразование Соната КА-1344 Рисунок 19 – Динамика распространенности септориоза сои, %, 2008 – 2011 гг. S. glycines особенно сильно поражает старые ткани и достигает наибольшего развития во второй половине вегетации. Развитие септориоза у G. max в фазе бобообразования в среднем по сортам за четыре года составило 22,5 – 60,7% при частоте встречаемости 74,5 – 97,5%, в отдельные годы достигало 100,0% (рис. 19, 20, приложение 10). 77 % 70 60 50 40 30 20 10 0 3-й тройчат.лист Гармония цветение Даурия бобообразование Соната КА-1344 Рисунок 20 – Динамика развития септориоза сои, %, 2008 – 2011 гг. Среди исследованных сортов культурной сои были как высоко, так и среднепоражаемые септориозом. Среднеспелые сорта Гармония и Даурия оказались более устойчивыми к данному заболеванию, чем раннеспелый сорт Соната, степень развития септориоза в фазе бобообразования варьировала в пределах 46,0 – 82,3% при распространенности болезни от 90,0 до 100,0% (рис. 19, 20, приложение 10). Устойчивость сортов сои к S. glycines возрастает с увеличением их периода вегетации, полученные нами данные согласуются с результатами исследования Л.А. Дега (2012). Септориоз очень часто встречается и на растениях дикорастущей сои. Согласно А.К. Лещенко (1978), G. soja может служить резерватором и накопителем инфекции для заражения культурной сои. Степень поражения листьев дикорастущей сои в фазе бобообразования в среднем по годам составляла 31,2% при распространенности 61,7% (рис. 19, 20, приложение 10). Пероноспороз, или ложная мучнистая роса (возбудитель Peronospora manshurica (Naum.) Syd. Syn.) является широко распространенным заболеванием сои на Дальнем Востоке. Известно (Заостровных, Дубовицкая, 2003), что оптимальной температурой для развития возбудителя пероноспороза считается 18-20оС. Распространение конидий происходит особенно интенсивно после 78 дождей и длительных туманов, когда капли воды сохраняются на листьях не менее 5 часов. Развитие болезни может приостанавливаться не только в сухую жаркую погоду, но и в период сильных ливневых дождей, когда конидии смываются с листьев. Наибольшего развития в условиях Амурской области пероноспороз достигал в 2009 году, в 2010 году отмечено самое низкое развитие болезни (рис. 21, приложение 11). В 2008 году погодные условия по температурному режиму за вегетационный период были выше среднемноголетних на 1-4оС, жаркая и сухая погода в начале лета сдерживала развитие пероноспороза. Относительная влажность повысилась лишь в июле, что привело к распространению болезни. Обильные осадки в июне 2009 года (206% от климатической нормы) при температуре воздуха ниже средней многолетней на 1оС создали благоприятные условия для развития патогена. Дождливая погода в июле, повышенная относительная влажность воздуха, длительное сохранение росы на листьях способствовали быстрому распространению и прорастанию конидий гриба. Вегетационный период 2010 года также характеризовался большим количеством осадков, но пероноспороз не получил широкого распространения, так как сильные ливневые дожди, прошедшие в июле, смывали с листьев конидии возбудителя и тем самым приостановили развитие заболевания. Июнь, июль и август 2011 года отличались высокой среднесуточной температурой воздуха, которая на 1-2оС была выше средней многолетней, и незначительным выпадением осадков, что привело к умеренному развитию пероноспороза. Во все годы исследований гриб начинал развиваться на сое в фазе пятого настоящего листа, к фазе цветения болезнь быстро распространялась, так как нарастание верхних листьев способствовало развитию гриба, поражающего преимущественно молодые растущие ткани. Нами не отмечено диффузного поражения растений возбудителем. 79 Развитие пероноспороза, % 50 46,2 45 40 35 28,4 30 25 20 15 10 5,55 5 0 5-й тройчатый лист цветение бобообразование 300 250 20 206 200 15 150 128 10 102 100 52 5 50 0 Количество осадков, мм Средняя температура воздуха, оС 25 0 май июнь июль август 2008 2009 2010 2011 среднемноголетнее 2008 2009 2010 2011 среднемноголетнее Средняя температура воздуха и количество осадков Рисунок 21 – Динамика развития пероноспороза сои в зависимости от метеорологических условий, %, 2008 – 2011 гг. Развитие болезни у сортов культурной сои в фазе цветения составляло 20,7 – 31,4%, при распространенности 39,4 – 64,5% (рис. 22, 23). Наиболее сильно поражались листья культурной сои в фазе бобообразования, развитие болезни составляло 52,9% при распространенности 97,0%, в отдельные годы поражалось до 100,0% растений (рис. 22, 23). 80 % 120 100 80 60 40 20 0 5-й тройчат.лист Гармония цветение Даурия бобообразование Соната КА-1344 Рисунок 22 – Динамика распространенности пероноспороза сои, %, 2008 – 2011 гг. % 60 50 40 30 20 10 0 5-й тройчат.лист Гармония цветение Даурия бобообразование Соната КА-1344 Рисунок 23 – Динамика развития пероноспороза сои, %, 2008 – 2011 гг. У сорта Гармония степень поражения пероноспорозом в фазе бобообразования варьировала в пределах 25,5 – 50,0%, у сорта Соната – 39,3 – 56,2% (рис. 22, 23, приложение 11). У сорта Даурия развитие болезни достигало 64,5% при распространенности 100,0% (рис. 22, 23, приложение 11). 81 За все четыре года исследования дикорастущая соя слабо поражалась пероноспорозом, на листьях были отмечены единичные некротические пятна. В фазе пятого тройчатого листа пораженных растений не наблюдалось, распространенность пероноспороза в фазе цветения не превышала 8,8% при развитии болезни 2,1%, к фазе бобообразования распространенность составила 20,4%, развитие – 11,0% (рис. 22, 23, приложение 11). Как мы отмечали выше, в Амурской области самым вредоносным из бактериальных заболеваний является угловатая пятнистость, или бактериальный «ожог» (возбудитель Pseudomonas glycineum Coerper. Syn.). За время проведения исследований (2008 – 2011 гг.) только в 2009 году данное заболевание было обнаружено на растениях G. max и G. soja. Решающим фактором в распространении бактериоза являются осадки. Осадки (206% к климатической норме), выпавшие в июне 2009 г., способствовали развитию бактериальной инфекции. Распространение бактериоза в фазе цветения на листьях сои колебалось в зависимости от сорта от 18,0 до 32,6%, его развитие – от 11,2 до 14,6%; в фазе бобообразования – соответственно от 40,0 до 65,0% и от 26,0 до 46,0% (рис. 24, 25, приложение 12). % 70 60 50 40 30 20 10 0 3-й тройчат.лист Гармония цветение Даурия бобообразование Соната КА-1344 Рисунок 24 – Динамика распространенности бактериоза сои, %, 2009 г. 82 % 50 45 40 35 30 25 20 15 10 5 0 3-й тройчат.лист Гармония цветение Даурия Соната бобообразование КА-1344 Рисунок 25 – Динамика развития бактериоза сои, %, 2009 г. Согласно нашим наблюдениям все сорта культурной сои и форма дикорастущей сои в фазе бобообразования проявили себя как среднеустойчивые с баллом поражения бактериозом 3. Для учета совместного влияния влажности и температуры на развитие болезней мы применили интегральный показатель – гидротермический коэффициент (ГТК) Г.Т. Селянинова (рис. 26). ГТК 3 70 степень развития болезней, % 60 2,5 50 2 40 1,5 30 1 20 0,5 10 0 0 2008 пероноспороз 2009 2010 септориоз бактериоз 2011 ГТК Рисунок 26 – Зависимость развития болезней от ГТК 83 Гидротермические условия среды определяли все основные аспекты жизнедеятельности возбудителей болезней и степень нанесенного ими вреда. Выявлена положительная зависимость между показателями ГТК и развитием септориоза, пероноспороза и бактериоза. Так, в 2009 году отмечено максимальное значение ГТК (2,5) и степень развития основных заболеваний сои. Низкое значение ГТК (1,2) в 2010 году способствовало снижению развития заболеваний. Однако в 2008 году, несмотря на то что ГТК составил 1,4, развитие септориоза было самым высоким. Обобщая полученные данные, можно сделать следующее заключение: – различные экологические факторы оказывают существенное влияние на развитие патогенов сои, наиболее благоприятные условия для развития грибной и бактериальной инфекций сложились в 2009 году. Развитию возбудителей пероноспороза и бактериоза способствуют повышенная влажность воздуха и большое количество осадков в сочетании с пониженной температурой. Более высокие температуры в сочетании с повышенной относительной влажностью воздуха необходимы для развития S. glycines, наибольшее развитие болезни отмечено в 2008 году; – сорт Соната наиболее сильно поражался септориозом; сорт Даурия – пероноспорозом. Сорт Гармония проявил среднюю устойчивость ко всему комплексу заболеваний. Форма дикорастущей сои КА-1344 оказалась устойчивой к пероноспорозу и среднеустойчивой к септориозу и бактериозу. 4.2 Участие ферментов в ответных реакциях сои на поражение листостеблевыми инфекциями Изучение биохимических и физиологических особенностей больного растения представляет важный этап на пути познания природы устойчивости его к патогенным микроорганизмам. При внедрении патогена в ткани растенияхозяина возникают различные нарушения метаболизма (изменяется интенсивность фотосинтеза, дыхания и транспирации, задерживается отток 84 ассимилянтов и т.д.). Значительные изменения претерпевают белковые компоненты клетки и в первую очередь белки-ферменты (Кожанова, Аксенова, 1976). Известно (Серова и др., 1982), что приспособление организма к условиям окружающей среды сводится к поддержанию на должном уровне скорости и направленности энзиматических реакций, благодаря которым обеспечивается состояние гомеостаза. Изменение ферментативной активности инфицированных растительных тканей свидетельствует о нарушении состояния гомеостаза. Вместе с тем оно указывает на то, что растительный организм активно борется с болезнью. Поэтому важным, на наш взгляд, было провести исследование влияния фитопатогенов на растение-хозяин по изменению активности ферментов, обеспечивающих нейтрализацию неблагоприятных факторов среды и являющихся важнейшими инструментами сохранения гомеостаза обмена веществ. 4.2.1 Изменение активности пероксидазы, каталазы и кислой фосфатазы в листьях сои при поражении грибной и бактериальной инфекциями в естественных условиях На следующем этапе работы испытанные на устойчивость к болезням сорта культурной сои (Гармония, Даурия, Соната) и дикорастущая форма КА-1344 анализировались на активность ферментов. Особый интерес представляет дикорастущая соя как уникальный источник генов, обусловливающих экологическую пластичность, высокий гомеостаз, устойчивость к болезням и вредителям, а также скороспелость и повышенную урожайность с одного растения (Ала и др., 2000). Сорта Гармония и Даурия, а также дикорастущая форма КА-1344 зарекомендовали себя как среднеустойчивые к возбудителю септориоза. Известно, что важную роль в развитии устойчивости растений к патогену играет пероксидаза, которая участвует в детоксикации активных форм кислорода, 85 образующихся в ходе физиологического стресса, вызванного патогенезом (Kawano, 2003). У сорта Гармония активность пероксидазы в пораженных листьях в фазе третьего тройчатого листа увеличивается в 4 раза. В фазах цветения и бобообразования активность пероксидазы также выше в инфицированных листьях, чем в здоровых (рис. 27, приложение 13). В пораженных листьях сорта Даурия в фазе третьего тройчатого листа активность пероксидазы ниже, чем в контроле, на 40%, в фазе бобообразования активность пероксидазы повышается на 56% (рис. 27, приложение 13). 450 400 350 % к контролю 300 250 200 150 100 50 0 3-й тройчат.лист контроль цв етение Гармония Даурия бобообразов ание Соната КА 1344 Рисунок 27 – Удельная активность пероксидазы (% к контролю) в листьях G. max и G. soja при поражении септориозом, 2008 – 2011 гг. После заражения септориозом в фазе третьего тройчатого листа активность пероксидазы в листьях формы КА-1344 увеличивается в 1,8 раза по сравнению с контролем (здоровые листья). В ходе развития патологического процесса активность фермента растёт: в фазе цветения она увеличивается в 1,9 раза, в фазе образования бобов – в 3,8 раза (рис. 27, приложение 13). В листьях восприимчивого сорта Соната в фазе третьего тройчатого листа активность пероксидазы ниже, чем в контроле, на 18%, в фазе бобообразования 86 достоверного отличия между пораженными и здоровыми листьями не выявлено (рис. 27, приложение 13). Проведенные нами исследования по изучению активности ферментов устойчивых и восприимчивых сортов сои к септориозу показали, что такой характер изменения активности пероксидазы свойствен восприимчивым растениям (Семенова, Титова, Дубовицкая, 2012). Активность пероксидазы в пораженных листьях сои достоверно отрицательно коррелировала со степенью поражения сои септориозом, особенно в фазе бобообразования: чем выше активность фермента, тем ниже балл поражения (табл. 6). Таблица 6 – Корреляционная зависимость между степенью поражения септориозом и удельной активностью пероксидазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара параметров степень поражения септориозом – удельная активность пероксидазы в листьях сои tтеор при Р0,95=1,96 Фаза развития сои 3-й тройчатый лист цветение бобообразование Коэффициент корреляции - 0,48 ± 0,05 - 0,44 ± 0,06 - 0,52 ± 0,05 Удельная активность каталазы в листьях сои, пораженных септориозом, в фазах третьего тройчатого листа и цветения ниже, чем в контроле у всех исследованных образцов (рис. 28, приложение 14). Существенное снижение активности каталазы (в 2,2 – 3 раза) наблюдается в фазе образования бобов у всех сортов культурной и дикорастущей сои (рис. 28, приложение 14). Снижение каталазной активности в этот период патогенеза может быть обусловлено разрушением хлорофилла. Известно, что под воздействием большинства фитопатогенных грибов уменьшается количество хлорофилла в листьях, так как микроорганизмы разрушают хлоропласты. Согласно исследованиям Б.А. Рубина и М.Е. Ладыгиной (1966), в листьях, в которых происходит частичное или полное разрушение хлорофилла, всегда наблюдается падение каталазной активности. 87 120 % к контролю 100 80 60 40 20 0 3-й тройчат.лист контроль цветение Гармония Даурия бобообразование Соната КА 1344 Рисунок 28 – Удельная активность каталазы (% к контролю) в листьях G. max и G. soja при поражении септориозом, 2008 – 2011 гг. Устойчивость сои к септориозу значимо и положительно коррелировала с удельной активностью каталазы в пораженных листьях сои только в фазы третьего тройчатого листа и цветения. В то же время в фазе бобообразования взаимосвязь между изучаемыми параметрами была слабой и недостоверной (табл. 7). Таблица 7 – Корреляционная зависимость между степенью поражения септориозом и удельной активностью каталазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения 3-й тройчатый лист септориозом – удельная цветение активность каталазы в листьях бобообразование сои tтеор при Р0,95=1,96 Корреляционная связь несущественна tфакт < tтеор Коэффициент корреляции 0,96 ± 0,02 0,68 ± 0,04 - 0,29 ± 0,15 Удельная активность кислой фосфатазы в пораженных листьях всех изучаемых сортов сои ниже, чем в здоровых. Существенно (на 50 – 60%) 88 снижается активность фермента в пораженных листьях сорта Соната, в меньшей степени – у дикорастущей сои (рис. 29, приложение 15). 120 % к контролю 100 80 60 40 20 0 3-й тройчат.лист контроль цветение Гармония Даурия бобообразование Соната КА 1344 Рисунок 29 – Удельная активность кислой фосфатазы (% к контролю) в листьях G. max и G. soja при поражении септориозом, 2008 – 2011 гг. По данным корреляционного анализа выявлена средняя положительная связь степени поражения сои септориозом с активностью кислой фосфатазы в пораженных листьях в фазе третьего тройчатого листа, слабая отрицательная (недостоверная) в фазе цветения и отрицательная средней силы в фазе бобообразования (табл. 8). Таблица 8 – Корреляционная зависимость между степенью поражения септориозом и удельной активностью кислой фосфатазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения 3-й тройчатый лист септориозом – удельная цветение активность кислой фосфатазы в бобообразование листьях сои tтеор при Р0,95=1,96 Корреляционная связь несущественна tфакт < tтеор 89 Коэффициент корреляции 0,46 ± 0,04 - 0,31 ± 0,15 - 0,45 ± 0,15 Активность ферментов в листьях сои, инфицированных P. manshurica, определяли в фазы цветения и образования бобов. Удельная активность пероксидазы в листьях дикорастущей сои, устойчивой к пероноспорозу, увеличивается в фазе цветения – в 1,4 раза, в фазе бобообразования – в 2 раза (рис. 30, приложение 16). Сорта Гармония и Соната, характеризующиеся средней устойчивостью к пероноспорозу, отличаются высокой активностью пероксидазы в пораженных листьях: в фазе бобообразования она повышается в 1,7 и 1,3 раза соответственно (рис. 30, приложение 16). У сильнопоражаемого сорта Даурия активность фермента в пораженных листьях повышается незначительно по сравнению с контролем (рис. 30, приложение 16). 250 % к контролю 200 150 100 50 0 цветение контроль бобообразование Гармония Даурия Соната КА 1344 Рисунок 30 – Удельная активность пероксидазы (% к контролю) в листьях G. max и G. soja при поражении пероноспорозом, 2008 – 2011 гг. Коэффициенты корреляции между устойчивостью сои к пероноспорозу и удельной активностью пероксидазы в пораженных листьях сои представлены в таблице 9. Взаимосвязь между анализируемыми параметрами была отрицательной средней силы: чем выше активность пероксидазы, тем ниже степень поражения листьев. 90 Таблица 9 – Корреляционная зависимость между степенью поражения пероноспорозом и удельной активностью пероксидазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения цветение пероноспорозом – удельная бобообразование активность пероксидазы в листьях сои tтеор при Р0,95=1,98 Коэффициент корреляции - 0,59 ± 0,05 - 0,54 ± 0,05 Пораженные растения характеризуются низкой активностью каталазы. В фазе цветения она ниже, чем у здоровых растений, в 1,2 – 1,8 раза. К фазе бобообразования активность каталазы продолжает снижаться: у дикорастущей сои она составила 76%, у сорта Гармония – 74 %, у сорта Соната – 38%, у сорта Даурия – 18% от контроля (рис. 31, приложение 17). Снижение каталазной активности в этот период патогенеза может быть обусловлено разрушением хлорофилла. 120 % к контролю 100 80 60 40 20 0 цветение контроль бобообразование Гармония Даурия Соната КА 1344 Рисунок 31 – Удельная активность каталазы (% к контролю) в листьях G. max и G. soja при поражении пероноспорозом, 2008 – 2011 гг. 91 У изучаемых сортов установлена положительная связь средней силы между степенью поражения пероноспорозом и удельной активностью каталазы в пораженных листьях сои в фазе цветения: чем выше активность, тем выше пораженность сои. В фазе бобообразования связь между анализируемыми параметрами была сильной отрицательной: чем выше активность, тем меньше степень поражения сои (табл. 10). Таблица 10 – Корреляционная зависимость между степенью поражения пероноспорозом и удельной активностью каталазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои Коэффициент параметров корреляции степень поражения цветение 0,43 ± 0,06 пероноспорозом – удельная бобообразование - 0,88 ± 0,02 активность каталазы в листьях сои tтеор при Р0,95=1,98 В листьях G. max и G. soja, пораженных пероноспорозом, активность кислой фосфатазы ниже в сравнении со здоровыми листьями. В меньшей степени активность фермента снижается в листьях дикорастущей сои, устойчивой к пероноспорозу, в большей степени – у сортов Даурия и Соната (рис. 32, приложение 18). 120 % к контролю 100 80 60 40 20 0 цветение контроль бобообразование Гармония Даурия Соната КА 1344 Рисунок 32 – Удельная активность кислой фосфатазы (% к контролю) в листьях G. max и G. soja при поражении пероноспорозом, 2008 – 2011 гг. 92 Проанализированная корреляционная зависимость между степенью поражения пероноспорозом и удельной активностью кислой фосфатазы в пораженных листьях сои в фазах цветения и бобообразования оценивается как слабая и недостоверная (табл. 11). Таблица 11 – Корреляционная зависимость между степенью поражения пероноспорозом и удельной активностью кислой фосфатазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения цветение пероноспорозом – удельная бобообразование активность кислой фосфатазы в листьях сои tтеор при Р0,95=1,98 Корреляционная связь несущественна tфакт < tтеор По-видимому, снижение активности кислой Коэффициент корреляции - 0,31 ± 0,52 - 0,37 ± 0,50 фосфатазы в листьях пораженных растений связано не с формированием устойчивости, а с нарушением углеводно-фосфорного и липидного обменов, которое происходит под влиянием патогенов. Исследованные сорта культурной сои (Гармония, Даурия и Соната) и дикорастущая форма КА-1344 обладали средней устойчивостью к бактериозу. Инфицирование растений бактериозом приводит к изменению активности пероксидазы в пораженных листьях всех образцов, она увеличивается относительно контроля, особенно у дикорастущей сои (рис. 33, приложение 19). Как видно из данных таблицы 12, существует средняя отрицательная корреляционная зависимость между удельной активностью пероксидазы в пораженных листьях сои и устойчивостью к бактериозу: чем выше активность пероксидазы, тем ниже балл поражения. 93 300 % к контролю 250 200 150 100 50 0 цветение контроль бобообразование Гармония Даурия Соната КА 1344 Рисунок 33 – Удельная активность пероксидазы (% к контролю) в листьях G. max и G. soja при поражении бактериозом, 2009 г. Таблица 12 – Корреляционная зависимость между степенью поражения бактериозом и удельной активностью пероксидазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения цветение бактериозом – удельная бобообразование активность пероксидазы в листьях сои tтеор при Р0,95=2,04 Коэффициент корреляции - 0,51 ± 0,08 - 0,52 ± 0,07 Под влиянием заражения удельная активность каталазы в фазе цветения в листьях сортов Соната и Даурия увеличивается в 2,7 – 4,5 раза соответственно, а у сорта Гармония и формы КА-1344 снижается. В фазе бобообразования в листьях культурной и дикорастущей сои активность каталазы была ниже, чем в контроле. Минимальная активность фермента (13 х 10-4 ед/мг белка) обнаружена у G. soja (рис. 34, приложение 20). 94 500 450 400 % к контролю 350 300 250 200 150 100 50 0 цветение контроль бобообразование Гармония Даурия Соната КА 1344 Рисунок 34 – Удельная активность каталазы (% к контролю) в листьях G. max и G. soja при поражении бактериозом, 2009 г. На основе полученных данных не установлено достоверной корреляции устойчивости сои к бактериозу с активностью каталазы в фазе цветения (табл. 13). Таблица 13 – Корреляционная зависимость между степенью поражения бактериозом и удельной активностью каталазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения цветение бактериозом – удельная бобообразование активность каталазы в листьях сои tтеор при Р0,95=2,04 Корреляционная связь несущественна tфакт < tтеор Коэффициент корреляции 0,32 ± 0,15 - 0,62 ± 0,06 Однако в фазе бобообразования наблюдалась сильная отрицательная зависимость степени поражения сои бактериозом от активности каталазы в пораженных листьях сои. Заражение растений сои бактериозом вызывает снижение активности кислой фосфатазы на 20 – 40% в листьях исследованных образцов в фазах цветения и бобообразования. В большей степени активность фермента снижается 95 у сорта Даурия (на 55%) в фазе бобообразования. В меньшей степени активность кислой фосфатазы снижается у дикорастущей формы (на 18%) (рис. 35, приложение 21). 120 % к контролю 100 80 60 40 20 0 цветение контроль бобообразование Гармония Даурия Соната КА 1344 Рисунок 35 – Удельная активность кислой фосфатазы (% к контролю) в листьях G. max и G. soja при поражении бактериозом, 2009 г. Степень поражения бактериозом положительно коррелировала с удельной активностью кислой фосфатазы в пораженных листьях сои (фаза цветения). Не установлено достоверной корреляции степени поражения листьев сои бактериозом с активностью кислой фосфатазы в пораженных листьях (фаза бобообразования) (табл. 14). Таблица 14 – Корреляционная зависимость между степенью поражения бактериозом и удельной активностью кислой фосфатазы в пораженных листьях сои (в среднем по сортам) Анализируемая пара Фаза развития сои параметров степень поражения цветение бактериозом – удельная бобообразование активность кислой фосфатазы в листьях сои tтеор при Р0,95=2,04 Корреляционная связь несущественна tфакт < tтеор 96 Коэффициент корреляции 0,48 ± 0,09 - 0,34 ± 0,12 Проанализировав корреляционные зависимости между степенью поражения листьев патогенами листо-стеблевой экологической ниши и активностью ферментов, была выявлена отрицательная средней силы связь активности пероксидазы с устойчивостью к септориозу, пероноспорозу и бактериозу. Корреляция активности каталазы с устойчивостью была положительной в фазы третьего тройчатого листа и цветения, отрицательной в фазе бобообразования. Несмотря на то, что связь активности кислой фосфатазы со степенью поражения листьев фитопатогенами в основном была слабой и недостоверной, нельзя не учитывать тот факт, что активность литических ферментов имеет большое значение в исходе противоборства растения-хозяина и патогена. Поскольку характер взаимодействия патогена и растения-хозяина зависит не только от патогена, но и от растения-хозяина, нами была исследована активность изучаемых ферментов в листьях здоровых растений. При проведении анализа было обнаружено, что удельная активность пероксидазы, каталазы и кислой фосфатазы возрастает в онтогенезе, достигая максимума в фазе образования бобов (приложения 16 – 18), подобные изменения активности ферментов в листьях сои были отмечены в ранее проведенных исследованиях (Семенова, 2006). Корреляционный анализ показал, что зависимости между устойчивостью сои к болезням и активностью ферментов (пероксидазы, каталазы, кислой фосфатазы) в листьях здоровых растений слабые и иногда недостоверные (табл. 15). Таблица 15 – Корреляционная зависимость между степенью поражения болезнями и удельной активностью ферментов в здоровых листьях сои (в среднем по сортам) Анализируемая пара параметров Фаза развития сои степень поражения болезнями – 3-й тройчатый лист удельная активность пероксидазы цветение в листьях сои бобообразование 97 Коэффициент корреляции -0,31 ± 0,06 -0,36 ± 0,05 -0,06 ± 0,06 Продолжение таблицы 15 степень поражения болезнями – 3-й тройчатый лист удельная активность каталазы в цветение листьях сои бобообразование степень поражения болезнями – 3-й тройчатый лист удельная активность кислой цветение фосфатазы в листьях сои бобообразование tтеор при Р0,95=1,98 Корреляционная связь несущественна tфакт < tтеор 0,37 ± 0,06 0,26 ± 0,19 -0,16 ± 0,16 0,26 ± 0,17 0,37 ± 0,05 -0,03 ± 0,06 Таким образом, постинфекционные изменения исследованных ферментов определяются генотипом и фазой развития сои. Формирование совместимых или несовместимых взаимоотношений растение-хозяин – патоген не зависит от изначальной активности пероксидазы, каталазы и кислой фосфатазы в листьях сои, а связано с изменением активности ферментов в ответ на внедрение патогена. 4.2.2 Изменение активности пероксидазы, каталазы и кислой фосфатазы в изолированных листьях сои при искусственном заражении септориозом Данное исследование было предпринято с целью изучения динамики изменения активности ферментов (пероксидазы, каталазы, кислой фосфатазы) в изолированных листьях, искусственно зараженных септориозом в лабораторных условиях, для того, чтобы исключить влияние абиотических стрессоров действующих в естественных условиях. Известно, что абиотический стресс может подавлять защитные реакции растений на биотическое воздействие (Перфильева, Рымарева, 2012). Результаты исследования показали, что степень поражения септориозом культурной и дикорастущей сои различна. Первые проявления болезни были отмечены на 3 сутки у сорта Соната, у сортов Гармония и Даурия - на 4 сутки и на 7 сутки – у формы КА-1344 (рис. 36). 98 % 90 80 70 60 50 40 30 20 10 0 1 3 4 7 9 12 сут ки Гармония Даурия Сонат а КА-1344 Рисунок 36 – Динамика развития септориоза на листьях G. max и G. soja, %, 2008 – 2010 гг. При учете развития септориоза на 9 сутки после инокуляции возбудителем болезни форма КА-1344 (1 балл поражения) была классифицирована как высокоустойчивая, сорта Гармония (2 балл поражения) и Даурия (2 балл поражения) – как устойчивые, и только сорт Соната (4 балл поражения) был отнесен к восприимчивым. На 12 сутки после заражения S. glycines только форма КА-1344 (2 балл поражения) была отнесена к устойчивым, сорта Гармония (3 балл поражения) и Даурия (3 балл поражения) – к среднеустойчивым, сорт Соната (5 балл поражения) – к сильновосприимчивым. Патогенность и конечный результат взаимодействия патогена и растенияхозяина во многом зависят от биологической активности возбудителя септориоза. Поэтому нами была определена активность ферментов в вытяжке из мицелия гриба S. glycines (рис. 37). Обнаружено, что возбудитель септориоза обладает некоторой ферментативной активностью. Удельная активность каталазы составила 20 х 10-4 ед/мг белка, пероксидазы – 12 х 10-4 ед/мг белка, кислой фосфатазы – 56 х 10-6 ед/мг белка. 99 Рисунок 37 – Возбудитель септориоза Septoria glycines Hemmi. Syn. (фото С.А. Титова) В контрольных (неинфицированных) листьях сои в процессе опыта активность каталазы (в 2,3 – 3,5 раза) и кислой фосфатазы (в 1,4 – 3,5 раза) снижается, а пероксидазы, напротив, увеличивается в 3,3 – 6,0 раза (приложения 22 – 24). Известно, что пероксидаза играет существенную роль в процессе старения, деградация листьев приводит к увеличению активности пероксидазы. Аналогичная закономерность отмечалась рядом исследователей ранее (Гусев и др., 1982; Lazar, Farkas, 1970; Омельченко, Андреева, 1976), но на других объектах. Инфицирование листьев септориозом привело к изменению активности ферментов по отношению к контролю. Незначительное увеличение активности пероксидазы у всех изучаемых сортов было зафиксировано на 1 сутки после нанесения инокулюма. Но пик активности приходится на 2 – 3 сутки: в наибольшей степени возрастает активность фермента у дикорастущей сои (в 4 раза), у сортов Гармония и Даурия ~ в 3 раза и 1,9 раза у сорта Соната (рис. 38, приложение 22). По данным корреляционного анализа была установлена взаимосвязь изменения активности пероксидазы в инфицированных листьях (3 100 сутки) с уровнем устойчивости к септориозу: чем выше активность пероксидазы, тем ниже балл поражения (r = - 0,97 ± 0,02) (приложение 25). 700 600 % к контролю 500 400 300 200 100 0 0 Соната 1 2 3 Гармония 4 5 6 Даурия 7 8 9 КА-1344 10 11 12 сутки контроль Рисунок 38 – Изменение удельной активности пероксидазы (% к контролю) в листьях G. max и G. soja при инфицировании септориозом, 2008 – 2010 гг. Второй пик активности пероксидазы в инфицированных листьях сои был зафиксирован на 9 сутки и зависел от степени поражения листа (рис. 38, приложение 22). Наиболее значительное увеличение активности фермента (в 6 раз) было отмечено у сорта Соната, развитие болезни при этом составляло 70%. Наименьшее повышение пероксидазы по сравнению с контролем наблюдалось у дикорастущей сои, некротические пятна занимают 10% поверхности листа. Сорта Гармония и Даурия, развитие болезни – 20 – 25%, по изменению активности пероксидазы занимают промежуточное положение. Согласно литературным данным увеличение активности пероксидазы происходит либо за счет повышения активности имеющихся в клетке молекул энзима, либо за счет синтеза фермента de novo (Mader, Walter, 1986). Полученные нами результаты не исключают вероятности, что увеличение активности фермента, по мере развития заболевания, может происходить и за счет пероксидазы, синтезируемой самим грибом S. glycines. Некоторые соображения в пользу такого предположения имеются в литературе (Wong, Preece, 1978). Б.А. 101 Рубин с соавторами (1973, 1975) считают, что активация пероксидазы в тканях растений, инфицированных грибами, может происходить, во-первых, за счет пероксидазы, синтезируемой самим паразитом, во-вторых, за счет активации имеющихся в клетке изопероксидаз. Активность каталазы в листьях сои повышается на 1 сутки после заражения, у формы КА-1344 – в 2 раза, у сортов культурной сои – в 1,2 – 1,5 раза относительно контроля (рис. 39, приложение 23). 250 % к контролю 200 150 100 50 0 0 1 2 3 4 5 6 7 8 9 10 11 12 сутки Соната Гармония Даурия КА-1344 контроль Рисунок 39 – Изменение удельной активности каталазы (% к контроля) в листьях G. max и G. soja при инфицировании септориозом, 2008 – 2010 гг. Корреляция между активностью каталазы в инфицированных листьях и их поражаемостью септориозом в этот период была сильной отрицательной r = - 0,96 ± 0,04 (приложение 25). По-видимому, высокая активность каталазы подавляет появление видимых некрозов после инокуляции возбудителем септориоза. По мере развития заболевания активность каталазы в листьях, пораженных септориозом, снижается у всех исследованных образцов, в большей степени у дикорастущей сои, что приводит к существенному накоплению перекиси водорода в клетке, тем самым создаются неблагоприятные условия для патогена (рис. 39, приложение 23). 102 Интерес к изучению отдельных групп фосфатаз определяется участием их в обмене углеводов, фосфолипидов, нуклеотидов и других соединений. Эти ферменты обладают гидролитическим и феразным действием, что имеет важное значение в обмене веществ. У восприимчивого сорта Соната пик активности кислой фосфатазы был зафиксирован на 1 сутки после заражения. У сортов, слабо поражаемых септориозом, активность фермента значительно повышалась только на 3 сутки развития болезни (рис. 40, приложение 24). 350 300 % к контролю 250 200 150 100 50 0 0 1 2 3 4 5 6 7 8 9 10 11 12 сутки Соната Гармония Даурия КА-1344 контроль Рисунок 40 – Изменение удельной активности кислой фосфатазы (% к контролю) в листьях G. max и G. soja при инфицировании септориозом, 2008 – 2010 гг. Дальнейшее развитие заболевания приводит к снижению активности кислой фосфатазы: у сорта Соната – в 4 раза, у слабо поражаемых сортов Гармония и Даурия – в 1,7 раза (рис. 40, приложение 24). Наибольшую устойчивость к действию возбудителя проявила форма КА1344, активность кислой фосфатазы в инфицированных листьев близка к контролю. В ходе корреляционного 103 анализа была выявлена сильная отрицательная связь между активностью кислой фосфатазы в инфицированных листьях (12 сутки) и степенью поражения (r = - 0,99 ± 0,02) (приложение 25). Таким образом, инфицирование листьев сои септориозом сопровождается изменением ферментативной активности, которое зависит от устойчивости растений к патогену. Активность пероксидазы значительно возрастает (в 3 – 4 раза) на 2 – 3 сутки в инфицированных листьях у устойчивых сортов культурной и дикорастущей сои и на 9 сутки у восприимчивого сорта Соната; активность каталазы – возрастает в 1,3 – 2 раза на 1 сутки, причем степень активации фермента зависела от восприимчивости к септориозу. Пик активности кислой фосфатазы у восприимчивого сорта Соната приходится на 1 сутки, у устойчивых растений – на 3 сутки. Дальнейшее развитие септориоза в инфицированных листьях G. max и G. soja сопровождается снижением активности каталазы и кислой фосфатазы. Устойчивость сои к возбудителю септориоза коррелируют с активностью пероксидазы (r = - 0,97 ± 0,02) и каталазы (r = - 0,96 ± 0,04) на начальном этапе, а кислой фосфатазы (r = - 0,99 ± 0,02) на поздних сроках развития заболевания в изолированных листьях сои. 104 5 ЭНЗИМАТИЧЕСКАЯ АКТИВНОСТЬ В СЕМЕНАХ СОИ ПРИ ПОРАЖЕНИИ ФИТОПАТОГЕНАМИ 5.1 Влияние зараженности семян сои болезнями на их биохимический состав и продуктивность растений Семена являются уникальной средой обитания для вредных организмов, особенно фитопатогенов. Вредные организмы, которые передаются через семенной материал, первыми проникают в проростки и всходы, вызывая или их гибель, или ингибирование ростовых процессов и развитие растений. Во всех районах возделывания соя поражается различными возбудителями болезней, что приводит к значительному недобору урожая. Пораженные семена имеют низкую полевую всхожесть и жизнеспособность, являются первоисточниками распространения заболеваний сои. Кроме того, больные семена дают всходы с пораженными семядолями и растения с диффузной формой заболевания. В годы исследований семена изучаемых сортов в той или иной степени были заражены возбудителями аскохитоза, пероноспороза, антракноза, церкоспороза, фузариоза и бактериоза (рис. 41). В последние годы широкое распространение на семенах сои получил пурпурный церкоспороз, вызываемый грибом Cercospora kikuchii T. Matsu et Tomoyasu. Анализ семян показал, что все изучаемые сорта несут на семенах инфекцию, которая составляет от 2,40% (Гармония) до 3,92% (Даурия) (рис. 42). Доминировали на семенах возбудители пероноспороза и бактериоза. Заселение возбудителем пероноспороза составляло от 0,87% (Гармония) до 3,06% (Даурия). Наибольшее поражение бактериозом отмечено у сорта Гармония (0,76%). 105 А Б В Г Д Рисунок 41 – Семена сои, пораженные болезнями (А – фузариозом; Б – пероноспорозом; В – пурпурным церкоспорозом; Г – церкоспорозом; Д – бактериозом) (фото С.А. Титовой) 106 На всех сортах отмечено поражение семян аскохитозом, фузариозом, церкоспорозом (округлая серая пятнистость) и пурпурным церкоспорозом. Поражение антракнозом выявлено на семенах сортов Гармония (0,13%) и Даурия (0,06%), сорт Соната не поразился этим заболеванием (рис. 42). Соната Соната Гармония Гармония 0,22 0,42 0,60 0,76 0,16 0,11 0,87 0,12 0,25 1,19 0,09 0,08 0,13 Даурия Даурия 0,14 0,06 0,10 0,30 0,20 0,06 3,06 Рисунок 42 – Поражение семян сои болезнями, %, 2009 – 2011 гг. В семенах сои содержится до 52% белка, до 27% – жира, до 30% – углеводов и около 6% – минеральных солей. В состав семян входят 12 основных витаминов (Кретович, 1979). По биологической ценности белок семян сои занимает первое место среди белков важнейших сельскохозяйственных культур. Биологическая ценность белка определяется сбалансированностью в нём незаменимых аминокислот: лизина, метионина, триптофана, треонина, валина, 107 фенилаланина, лейцина, изолейцина, которые жизненно необходимы для организма, но могут быть синтезированы только растениями. Соя высоко ценится и как масличная культура. Жир, содержащийся в ней, на 85% состоит из ненасыщенных жирных кислот. Результаты нашего исследования показали, что заражение семян пероноспорозом не оказывает существенного влияния на содержание белка и незаменимых аминокислот (рис. 43а, приложения 26, 27). % % 20 45 40 35 15 30 25 10 20 15 5 10 5 0 0 Соната Гармония пораженные Соната Даурия Гармония пораженные здоровые а Даурия здоровые б Рисунок 43 – Содержание белка (а) и масла (б) в семенах сои, пораженных пероноспорозом, %, 2010 – 2011 гг. Наибольшее отклонение между пораженными и здоровыми семенами было выявлено у сорта Даурия, белка в пораженных семенах было на 0,92% меньше, чем в здоровых. Содержание масла незначительно снижается в пораженных семенах у всех исследованных сортов сои: у Сонаты на 1,05%; у Гармонии на 1,50%; у Даурии на 1,34% (рис. 43б, приложение 28, 29). Заражение семян сои P. manshurica не оказывает влияния на содержание насыщенных жирных кислот пальмитиновой и стеариновой в масле (приложения 28, 29). 108 Ценность соевого масла обусловлена содержанием олеиновой и линолевой кислот, линоленовая кислота – нежелательный компонент. Нами были отмечены существенные различия в содержании олеиновой и линоленовой кислот в здоровых и пораженных семенах: количество олеиновой кислоты уменьшается, а Соната пораженные Гармония пораженные Даурия линоленовой, наоборот, увеличивается (рис. 44, приложения 28, 29). пораженные здоровые здоровые здоровые 0% 20% С/18:1 40% С/18:2 60% 80% 100% С/18:3 Рисунок 44 – Содержание ненасыщенных жирных кислот в семенах сои, пораженных пероноспорозом, %, 2010 – 2011 гг. Увеличение содержания линоленовой кислоты в пораженных семенах ухудшает их качество. От содержания линоленовой кислоты зависит быстрое прогоркание масла, поэтому содержание её должно быть минимальным. Анализ основных элементов продуктивности показал, что сорта со средней устойчивостью к септориозу Гармония и Даурия имели урожайность на 15 – 31% выше, чем восприимчивый сорт Соната (рис. 45, приложение 33). У восприимчивого сорта Соната масса 1000 семян была ниже на 14,6 – 51,7 г, масса семян с растения снижается на 1,4 – 1,7 г (приложение 30). Заражение сои пероноспорозом не сказалось на урожайности растений. Восприимчивый к пероноспорозу сорт Даурия в среднем имел самую высокую урожайность. Вероятно, это связано с тем, что заболевание проявляется поздно, 109 максимум развития пероноспороза приходится на фазу налива семян, когда семена уже сформировались. ц/га 22,75 25 20,06 20 17,30 15 10 5 0 Соната Гармония Даурия Рисунок 45 – Урожайность сортов сои с разной устойчивостью к септориозу и пероноспорозу, ц/га, 2009 – 2011 гг Септориозом растения поражаются очень рано (в фазе всходов), максимум приходится на фазы цветения и бобообразования. Поэтому это заболевание оказывает существенное влияние на продуктивность сои. В результате статистической обработки установлена сильная отрицательная связь между урожайностью и степенью поражения сои септориозом: чем ниже балл поражения, тем выше урожайность (табл. 16). Корреляционная зависимость между урожайностью и степенью поражения сои пероноспорозом средняя отрицательная (табл. 16). Таблица 16 – Корреляционная зависимость между урожайностью сортов сои и степенью поражения болезнями Анализируемая пара параметров урожайность – степень поражения септориозом урожайность – степень поражения пероноспорозом tтеор при Р0,95=2,05 110 Коэффициент корреляции - 0,68 ± 0,06 - 0,35 ± 0,09 Анализ полученных данных позволяет сделать следующее заключение: – все изучаемые сорта несут на семенах инфекцию от 2,40% (Гармония) до 3,92% (Даурия). Наибольший процент из комплекса болезней занимал пероноспороз (5,12%), затем бактериоз (1,48%) и пурпурный церкоспороз (1,02%); – в семенах, пораженных пероноспорозом, снижается содержание масла и количество олеиновой кислоты, но увеличивается содержание линоленовой. Заражение семян пероноспорозом не оказывает существенного влияния на содержание белка и его аминокислотный состав; – корреляционная зависимость между урожайностью и степенью поражения сои септориозом сильная отрицательная (r = - 0,68 ± 0,06), между урожайностью и степенью поражения пероноспорозом - средняя отрицательная (r = - 0,35 ± 0,09). 5.2 Изменение активности пероксидазы, каталазы и кислой фосфатазы в семенах сои при поражении грибной и бактериальной инфекциями Результаты исследования показали, что удельная активность ферментов различна у здоровых и пораженных болезнями семян изучаемых сортов сои. Изменение ферментативной активности инфицированных растительных тканей свидетельствует о нарушении состояния гомеостаза (Серова и др., 1982). Способность сохранять необходимый уровень метаболических процессов и скорость их перестройки зависят от степени устойчивости сорта (Удовенко, 1975). В семенах сои, пораженных грибной и бактериальной инфекциями, удельная активность пероксидазы ниже, чем в здоровых (табл. 17, приложение 31). Согласно литературным данным (Рубин и др., 1975; Андреева и др., 1979), снижение активности пероксидазы связано с изменением уровня окислительновосстановительных процессов инфицированных семян, инфекция вызывает снижение метаболической активности и интенсивности дыхания. В наибольшей степени активность фермента снижается в семенах сортов Гармония и Даурия, пораженных фузариозом (в 3,9 – 4,7 раза соответственно). 111 Следует отметить, что активность пероксидазы в семенах всех сортов, пораженных пероноспорозом, снижается незначительно. Противоположно изменению удельной активности пероксидазы в семенах, пораженных разной инфекцией, активность каталазы увеличивается, что соответствует наличию обратной зависимости между удельной активностью каталазы и пероксидазы (Селихова, 2003). Повышенная каталазная активность способствует снижению концентрации перекиси водорода в клетке, что лимитирует реакции, катализируемые пероксидазой. Данные таблицы 18 показывают, что активность каталазы в семенах сои сорта Соната, пораженных пероноспорозом, аскохитозом, фузариозом, церкоспорозом и пурпурным церкоспорозом, выше в 1,2 – 2,4 раза, чем в здоровых. В семенах сорта Гармония, пораженных грибной инфекцией, активность каталазы увеличивается незначительно: в 1,8 раза при заражении пероноспорозом и фузариозом и остается на уровне здоровых в семенах, пораженных аскохитозом, церкоспорозом, пурпурным церкоспорозом и антракнозом (приложение 32). Удельная активность каталазы в пораженных семенах сорта Даурия возрастает в 1,2 – 2,0 раза и в меньшей степени под влиянием возбудителей аскохитоза и церкоспороза. Увеличение активности каталазы выявлено в семенах всех сортов сои, пораженных бактериальной инфекцией. По-видимому, вместе с другими оксидоредуктазами она направляет метаболизм клетки на создание неблагоприятных условий для патогена. Полученные данные показывают, что удельная активность кислой фосфатазы отличается у здоровых и пораженных разными болезнями семян. Активность кислой фосфатазы в пораженных семенах как увеличивается, так и уменьшается по сравнению со здоровыми (табл. 19, приложение 33). 112 Таблица 17 – Удельная активность пероксидазы в семенах сои при поражении разной инфекцией, ед/мг белка, 2009 – 2011 гг. Сорт Пораженные семена Здоровые семена пероноспороз аскохитоз фузариоз церкоспороз 105 ± 13 114 ± 21 127 ± 4 2,62 96 ± 14 107 ± 21 115 ± 29 1,31 42 ± 2 90 ± 6 67 ± 2 3,32 46 ± 1 29 ± 2 27 ± 1 2,74 56 ± 0,5 30 ± 1 86 ± 3 2,40 Соната Гармония Даурия НСР05 пурпурный церкоспороз 75 ± 2 61 ± 2 89 ± 4 2,74 антракноз бактериоз 39 ± 1 68 ± 2 2,22 87 ± 15 95 ± 11 106 ± 11 2,62 Таблица 18 – Удельная активность каталазы в семенах сои при поражении разной инфекцией, ед/мг белка х 10 -3, 2009 – 2011 гг. Сорт Соната Гармония Даурия НСР05 Пораженные семена Здоровые семена пероноспороз аскохитоз фузариоз церкоспороз 54 ± 2 62 ± 2 49 ± 1 3,04 89 ± 3 110 ± 15 90 ± 8 0,75 66 ± 8 65 ± 1 53 ± 3 2,60 130 ± 34 110 ± 12 96 ± 19 0,76 101 ± 15 70 ± 11 57 ± 9 3,32 пурпурный церкоспороз 127 ± 13 70 ± 10 69 ± 8 2,74 антракноз бактериоз 69 ± 0,5 58 ± 0,5 1,61 125 ± 6 95 ± 8 68 ± 3 2,96 Таблица 19 – Удельная активность кислой фосфатазы в семенах сои при пораженных разной инфекцией, ед/мг белка х 10-5, 2009 – 2011 гг. Сорт Соната Гармония Даурия НСР05 Пораженные семена Здоровые семена пероноспороз аскохитоз фузариоз церкоспороз 17 ± 0,5 23 ± 4 20 ± 2 3,61 22 ± 1 31 ± 2 27 ± 1 2,74 18 ± 1 11 ± 1 10 ± 1 3,04 5 ± 0,3 8 ± 0,3 7 ± 0,2 2,22 42 ± 2 37 ± 2 33 ± 1 2,22 113 пурпурный церкоспороз 14 ± 1 17 ± 1 19 ± 4 1,86 антракноз бактериоз 19 ± 1 15 ± 1 1,20 15 ± 0,5 15 ± 0,5 16 ± 3 3,05 Существенное снижение активности фермента (~ в 3 раза) было установлено при поражении семян сои фузариозом. Изменение ферментативной активности, вероятно, связано с нарушением углеводно-фосфорного и липидного обменов, которое происходит под влиянием инфекции. Таким образом, удельная активность пероксидазы в семенах сои под влиянием различной инфекции снижается, а каталазы – повышается или остается на уровне здоровых семян, что определяется сортом сои и может характеризовать его устойчивость к заболеванию. Инфицирование семян фузариозом приводит к резкому снижению активности кислой фосфатазы у всех сортов сои. На следующем этапе работы нами была определена удельная активность пероксидазы, каталазы и кислой фосфатазы в проростках сои, полученных из семян, пораженных разной инфекцией. Из данных, представленных на рисунке 46, видно, что активность пероксидазы проростков сои изменяется относительно контроля (проростков из здоровых семян). По-видимому, под влиянием метаболитов патогена нарушается обмен веществ не только в пораженных семенах, но и в проростках, полученных из них. При этом наблюдались различия в реакции проростков разных сортов сои на инфекцию. Увеличение активности пероксидазы относительно контроля происходит в проростках сорта Соната, полученных из семян, пораженных церкоспорозом (в 1,6 раза) и пероноспорозом (в 1,3 раза). Наоборот, в проростках, полученных из семян, пораженных пурпурным церкоспорозом и бактериозом, активность фермента снижается (приложение 34). В проростках сорта Гармония активность пероксидазы под влиянием инфекции изменяется незначительно (приложение 34). Согласно исследованиям (Серова и др., 1982) стабильная энзиматическая система свойственна устойчивым растениям. К ферментам этой системы принадлежит пероксидаза. 114 200 180 % к контролю 160 140 120 100 80 60 40 20 0 Соната Гармония Даурия контроль пероноспороз церкоспороз пурпурный церкоспороз бактериоз антракноз Рисунок 46 – Удельная активность пероксидазы в проростках, полученных из пораженных разной инфекцией семян, % к контролю, 2010 – 2011 гг. У сорта Даурия самая высокая активность пероксидазы (18 х 10 -4 ед/мг белка) была в проростках из семян, пораженных антракнозом, самая низкая (6 х 10-4 ед/мг белка) – в проростках из семян, пораженных бактериозом (приложение 34). В зависимости от возбудителя реакция на него, проявляемая в активности каталазы, может быть различной. Активность каталазы в проростках из пораженных семян как увеличивается, так и уменьшается по сравнению с контрольными (рис. 47). Повышается активность фермента в проростках сортов Соната и Гармония, полученных из семян, пораженных пероноспорозом и церкоспорозом, в проростках сорта Даурия – из семян, пораженных пероноспорозом (приложение 35). Существенное снижение активности каталазы (в 1,7 – 2,0 раза) наблюдается в проростках сортов Даурия и Гармония, полученных из семян, пораженных антракнозом. В проростках сортов Соната, Гармония и Даурия, полученных из 115 семян, пораженных пурпурным церкоспорозом и бактериозом, активность каталазы остается на уровне контрольных (приложение 35). 160 140 % к контролю 120 100 80 60 40 20 0 Соната Гармония Даурия контроль пероноспороз церкоспороз пурпурный церкоспороз бактериоз антракноз Рисунок 47 – Удельная активность каталазы в проростках, полученных из пораженных разной инфекцией семян, % к контролю, 2010 – 2011 гг. Полученные данные позволяют сделать вывод о том, что изменение активности ферментов в пораженных семенах сои и проростках, полученных из них, зависит от генотипа растения и вида возбудителя. Было выявлено, что наибольшей патогенностью обладает возбудитель фузариоза. Согласно изменению активности пероксидазы и каталазы в проростках, полученных из зараженных семян, сорт Гармония проявил себя как наиболее устойчивый к воздействию фитопатогенов. 116 Выводы 1. Инфицирование корней сои корневой гнилью приводит к увеличению активности пероксидазы и снижению активности каталазы у устойчивого сорта Гармония (активность кислой фосфатазы достоверно не изменилась) и, наоборот, к снижению активности пероксидазы и росту активности каталазы и кислой фосфатазы у восприимчивого сорта Соната. 2. При патогенезе корневой гнили активность пероксидазы, выделенной из корней и листьев сои, изменяется сходным образом, в отличие от активности каталазы и кислой фосфатазы, что указывает на возможность использования любого из этих органов для установления устойчивости к патогену по активности пероксидазы. 3. Обнаружено, что комплексную среднюю устойчивость проявил сорт Гармония при поражении листо-стеблевыми инфекциями, форма дикорастущей сои КА-1344 оказалась устойчивой к пероноспорозу и среднеустойчивой к септориозу и бактериозу. Сорт Соната наиболее сильно поражался септориозом, сорт Даурия – пероноспорозом. 4. Установлено, что формирование совместимых или несовместимых взаимоотношений растение-хозяин – патоген не зависит от изначальной активности пероксидазы, каталазы и кислой фосфатазы в листьях сои, а связано с изменением активности ферментов в ответ на внедрение патогена (S. glycines, P. manshurica, Ps. glycineum). 5. Определена динамика энзиматической активности при искусственном инфицировании изолированных листьев S. glycines. Установлено, что активность пероксидазы (r = - 0,97 ± 0,02) и каталазы (r = - 0,96 ± 0,04) на начальном этапе, а кислой фосфатазы (r = - 0,99 ± 0,02) на поздних сроках развития заболевания может служить критерием устойчивости к септориозу. 6. Показано, что изменение энзиматической активности в пораженных семенах сои и проростках, полученных из них, зависит от генотипа растения и 117 вида возбудителя. Выявлено, что наибольшей патогенностью обладает возбудитель фузариоза. 7. Совокупность полученных экспериментальных данных свидетельствует о том, что система на основе антиоксидантных (пероксидазы и каталазы) и литических (кислая фосфатаза) ферментов может быть использована в качестве биохимического критерия при селекции сои на устойчивость к фитопатогенам. Практические рекомендации 1. При селекции на повышение устойчивости сои к фузариозной корневой гнили рекомендуется использовать в качестве исходного материала сорт Гармония, к септориозу, пероноспорозу и бактериозу – генотипы с комплексной устойчивостью к возбудителям листо-стеблевой экологической ниши: сорт Гармония и дикорастущую форму КА 1344. 2. Для диагностики устойчивости сои к септориозу следует использовать тестовую систему на основе антиоксидантных (пероксидазы и каталазы) и литических (кислая фосфатаза) ферментов в контролируемых лаборатории с последующей оценкой в естественных условиях. 118 условиях Список литературы 1. Абрамов, И.Н. Болезни и вредители соевых бобов на Дальнем Востоке / И.Н. Абрамов. – Владивосток, 1931. – С. 40 – 56. 2. Абрамов, И.Н. Болезни сельскохозяйственных растений на Дальнем Востоке / И.Н. Абрамов. – Хабаровск: Дальневост. изд-во, 1938. – С. 221 –225. 3. Аверьянов, A.A. Влияние света на биохимические свойства мицелия Fusarium oxysporum Schlecht. / А.А. Аверьянов, Н.В. Гужова, В.В. Мочалов // Микология и фитопатология, 1981. – Т. 15, № 5. – С. 361 – 365. 4. Аверьянов, А.А. Активные формы кислорода и иммунитет растений / А.А. Аверьянов // Успехи современной биологии. – 1991. – Т. 111, вып. 5. – С. 722 – 735. 5. Айала, Ф. Введение в популяционную и эволюционную генетику / Ф. Айала. – М.: Мир, 1984. – 230 с. 6. Аксенова, В.А. О некоторых свойствах пероксидазы инфицированных тканей растений / В.А. Аксенова, О.Н. Кожанова, Б.А. Рубин // Физиология растений. – 1971. – Т. 18, № 2. – С. 387 – 391. 7. Акулова, Е.А. О двух формах каталазы в хлоропластах и листьях гороха / Е.А. Акулова, А.П. Смолов // Физиология и биохимия культурных растений. – 1974. – Т. 6, вып. 4. – С. 418 – 422. 8. Ала, А.Я. Дикая соя – источник хозяйственно-ценных признаков / А.Я. Ала, А.А. Гамолин // Физиолого-генетические основы селекции сои: сб. науч. тр. / ВАСХНИЛ. Сиб. отд-ние. – Новосибирск, 1984. – Вып. 27. – С. 38 – 44. 9. Ала, А.Я. Изучение коллекции дикой сои по хозяйственно ценным и морфологическим признакам / А.Я. Ала, В.С. Ала, Ю.А. Супряга // Вопросы биологии и технологии возделывания сои на Дальнем Востоке России. – Благовещенск, 2000. – С. 26 – 30. 10. Ала, А.Я. Соя: генетические методы селекции G. max (L.) Merr. X G. soja / А.Я. Ала, В.А. Тильба. – Благовещенск: ПКИ «Зея», 2005. – 128 с. 119 11. Анджана, Г. Изоформы пероксидаз и кодирующие их РНК в листьях подсолнечника после заражения Alternaria helianthi / Г. Анджана, К. Р. Кайни, Х.С. Шетти [и др.] // Физиология растений. – 2007. – Т. 54, № 4. – С. 579 – 583. 12. Андреева, В.А. Изозимный спектр пероксидазы в растениях, инфицированных вирусом L / В.А. Андреева, С.И. Омельченко, Л.А. Паутова // Труды Биолого-почвенного института. Вирусные болезни растений Дальнего Востока. – Владивосток, 1973. – Т. 14. – С. 46 – 52. 13. Андреева, В.А. Активность, изоферментный спектр, термостабильность и молекулярный вес пероксидазы, выделенной из здоровых и зараженных вирусами растений табака / В.А. Андреева, В.А. Воронова, Н.Н. Угарова // Биохимия. – 1979. – Т. 44, вып. 3. – С. 394 – 399. 14. Адреева, В.А. Индукция активности пероксидазы при вирусном и механическом повреждениях / В.А. Андреева, В.А. Воронова // Вирусные болезни растений и меры борьбы с ними. – Владивосток: ДВНЦ АН СССР, 1980. – С. 26 – 29. 15. Андреева, В.А. Влияние штаммов Х-вируса картофеля на кинетику фермента пероксидазы / В.А. Андреева, В.А. Воронова // Труды ЛСХА, 1981. – Вып. 191. – С. 12 – 14. 16. Андреева, В.А. Фермент пероксидаза: Участие в защитном механизме растений / В.А. Андреева. – М.: Наука, 1988. – 128 с. 17. Асеева, Т.А. Эффективность комплексного применения агрохимических средств и биопрепаратов на посевах сои в условиях Среднего Приамурья / Т.А. Асеева, Е.В. Золотарева, С.Р. Паланица // Экологические функции агрохимии в современном земледелии / ВНИИА. – М.: ВНИИА, 2008. – С 27 – 30. 18. Багаева, Е.В. Заболевания зернобобовых культур в лесостепной зоне Омской области / Е.В. Багаева, Н.И. Виноградова // Тр. Омского с.-х. ин-тa им. С.М. Кирова. – 1964. – Вып. 54. – С. 16. 19. Бах, А.Н. Биологическое и технологическое значение ферментативных процессов / А. Н. Бах. – Изв. АН СССР. Сер. Биол., 1936 – №4 – С. 627 – 638. 120 20. Бояркин, А.Н. Быстрый метод определения активности пероксидазы // Биохимия. – 1951. – Т. 16, вып. 4. – С. 352. 21. Вардания, Л.Я. Болезни сои в Абхазии / Л.Я. Вардания // Защита растений. – 1971. – № 3. – С 40 – 41. 22. Вардания, Л.Я. Устойчивость сои к возбудителям корневых гнилей и увядания в условиях Абхазии: автореф. дис. … канд. биол. наук: 03.02.08 / Вардания Л.Я. – Сухуми, 1972. – 37 с. 23. Вардания, Л.Я. Фузариоз сои в Абхазии / Л.Я. Вардания // Защита растений. – 1974. – № 10. – С 52 – 137. 24. Овчинникова, А.М. Болезни семян на Дальнем Востоке и методика их фитопатологического анализа / А.М. Овчинникова, В.И. Потлайчук // ВИЗР. – Л., 1980. – Вып. 48. – С. 41 – 46. 25. Василевский, Н.И. Паразитные несовершенные грибы. Гифомицеты / Н.И. Василевский, Б.П. Каракулин. – М.-Л.: Изд-во АН СССР, 1937. – 52 с. 26. Владимирский, С.В. Болезни сои в северной зоне ее культуры / С.В. Владимирский // Зап. Ленинград. с.-х. ин-та. – 1939. – Вып. 3. – С. 135 – 149. 27. Власюк, П.Н. Неспецифические фосфатазы растений / П.Н. Власюк, А.И. Тернавский // Сельскохозяйственная биология. – 1973. – Т. 5, № 4. – С. 377 – 383. 28. Возбудители болезней сельскохозяйственных растений Дальнего Востока / Т.А. Барбаянова, Л.Н. Егорова, А.В. Крылов [и др.]; под. ред. З.М. Азбукиной. – М.: Наука, 1980. –370 с. 29. Газарян, И.Г. Пероксидазы растений / И.Г. Газарян // Итоги науки и техники. Сер. Биотехнология. – М., 1992. – Т. 36. – С. 4 – 30. 30. Галимова, И.В. Изучение активности и молекулярной гетерогенности кислой фосфатазы у гетерозисных гибридов тополя / И.В. Галимова // Физиология и биохимия культурных растений. – 1987. – Т. 19, № 2. – С. 119 – 124. 31. Гамбург, К.З. Ауксины в культурах тканей и клеток растений / К.З. Гамбург, Н.И. Рекославская, С.Г. Швецов. – Новосибирск: Наука., 1990. – 241 с. 121 32. Ганеева, Л.А. Влияние соединений марганца на активность каталазы и пероксидазы и на проявление агрессивности фитофторы / Л.А. Ганеева, И.С. Докучаева // Вестник Казанского технологического университета. – 2009. – № 6. – С. 115 – 120. 33. Ганя, А.И. Основные грибные патогенны сои в Молдавии и выявление источников устойчивости к ним: автореф. дис. … канд. биол. наук: 03.02.08 / Ганя Анатолий Иванович – М., 1981a. – 21 с. 34. Ганя, А.И. Изучение белой гнили сои в Молдавии / А.И. Ганя // Фитопатогенные микроорганизмы культурных растений в Молдавии. – Кишинев: Штиинца, 1981б. – С. 131 – 146. 35. Гигрин, В.В. Окислительные, липолитические и протекторные ферменты в листьях устойчивых и восприимчивых к ржавчине растений пшеницы / В.В. Гигрин // Физиология растений. – 1988. – Т. 35, вып. 6. – С. 1198 – 1201. 36. Глазко, В.И. Генетика изоферментов животных и растений / В.И. Глазко, И.А. Созинов; под ред. А.А. Созинова. – Киев: Урожай, 1993. – 528 с. 37. Голубев А.А. Методические указания по изучению устойчивости зерновых бобовых культур к болезням / А.А. Голубев. – Л., 1976. – С. 61 – 76. 38. Горленко, М.В. Сельскохозяйственная фитопатология: учебник / М.В. Горленко. – М.: Высшая школа,1968. – 434с. 39. ГОСТ 12038-84. Семена сельскохозяйственных культур. Методы определения всхожести. Введен 01.07.86. – М: Изд-во стандартов, 2004. – 219 с. 40. ГОСТ 12044-93. Семена сельскохозяйственных культур. Методы определения зараженности болезнями. Введен 01.07.86. – М.: Изд-во стандартов, 2004. – 219 с. 41. Гофман, А.В. Особенности развития болезней на различных сортах сои и применение средств защиты в условиях орошения в зоне неустойчивого увлажнения Ставропольского края: дисс. …канд. биол. наук: 06.01.11 / Гофман Анна Викторовна. – Ставрополь, 2007. – 135 с. 42. Гоцуляк, В.Д. Активность пероксидазы и каталазы у разных по устойчивости пшениц к бурой листовой ржавчине / В.Д. Гоцуляк, О.К. Кондратюк 122 // Респ. научн. конф. молод. исслед. по физиологии и биохимия растений. – 1967. – С. 128 – 129. 43. Граскова, И.А. Изменение активности пероксидазы при патогенезе кольцевой гнили картофеля / И.А. Граскова, А.С. Романенко, С.В. Владимирова, А.В. Колесниченко // Физиология растений. – 2004. – Т. 51, № 4. – С. 529 – 533. 44. Граскова, И.А. Роль слабосвязанных с клеточной стенкой пероксидаз в устойчивости растений к биотическому стрессу: дис. … д-ра биол. наук: 03.00.12 / Граскова Ирина Алексеевна. – Иркутск, 2008. – 392 с. 45. Гуляй, И.Э. Четвертичная структура и регуляторные свойства кислой фосфатазы из листьев петрушки / И.Э. Гуляй, А.Ф. Макарчиков // Сб. статей научно-практич. конф. – Гродно, 2001. – С. 374 – 377. 46. Гунина, А.М. Болезни сои в Амурской области / А.М. Гунина // Тр. Амурской с.-х. опытной станции. – 1968. – Т. 2, вып. 2. – С. 73 – 80. 47. Гунина, А.М. Результаты исследований по защите сои от болезней и вредителей на Дальнем Востоке / А.М. Гунина // Науч. техн. бюл. / ВНИИ сои. – Новосибирск, 1978. – Вып. 13. – С. 3 – 31. 48. Гунина, А.М. Влияние заражённости семян сои болезнями на посевные и урожайные качества / А.М. Гунина, Л.К. Дубовицкая // Науч. техн. бюл. / ВНИИ сои. – Новосибирск, 1983. – Вып. 11. – С. 47 – 51. 49. Гусев, М.В. Пероксидазная активность в молодых и деградирующих листьях Arabidopsis thaliana (L.) / М.В. Гусев, Е.Р. Карташова, Р.А. Афифи // Ферменты, ионы и биоэлектрогенез у растений: сб. научн. тр. – Горький: Изд-во ГГУ, 1982. – С. 93 – 98. 50. Дворядкин, Н.И. Болезни и вредители сои и меры борьбы с ними: рекомендации / Н.И.Дворядкин, О.И Тихонов, Ю.П. Мякушко // ВНИИМК. – Краснодар, 1979. – 19 с. 51. Дега, Л.А. Изучение закономерностей наследуемости устойчивости сои к поражению грибом Septoria glycines Hemmi в различных группах спелости: авторефер. дис. … канд. с.-х. наук: 06.01.05 / Дега Людмила Андреевна. – п. Тимирязевский, 2001. – 25 с. 123 52. Дега, Л.А. Вредители и болезни сои на Дальнем Востоке / Л.А. Дега. – Владивосток: Дальнаука, 2012. – 98 с. 53. Дементьева, М.И. Комплексный иммунитет и физиолого- биохимические особенности сортов крыжовника / М.И. Дементьева // Докл. Моск. с.-х. акад. им. К. А. Тимирязева. – 1957. – вып. 29. – С. 142–146. 54. Доспехов, Б.А. Методика полевого опыта / Б.А. Доспехов. – М.: Агропромиздат, 1979. – 351 с. 55. Дубовицкая, Л.К. Поражаемость сои корневыми гнилями в севообороте и бессменно / Л.К. Дубовицкая, А.М. Гунина // Науч.-техн. бюл. / ВНИИ сои. – Новосибирск, 1979. – Вып. 16/17. – С. 27 – 30. 56. Дубовицкая, Л.К. Корневые гнили сои / Л.К. Дубовицкая // Масличные культуры. –1984. – № 4. – С. 40. 57. Дубовицкая, Л.К. Корневая гниль сои в Приамурье и обоснование мер борьбы с ней: дис. … канд. с.-х. наук: 06.01.11 / Дубовицкая Любовь Кондратьевна. – Л., 1987. – 194 с. 58. Дубовицкая, Л.К. Болезни и вредители сои и меры борьбы с ними в условиях Амурской области: учебное пособие / Л.К. Дубовицкая, Н.Н. Кравцова. – Благовещенск: ДальГАУ, 2002. – 191 с. 59. Дьяконов, К.П. Вирусные болезни сои и их переносчики в условиях Приморского края / К.П. Дьяконов, Е.П. Голаюда // Вирусные болезни сельскохозяйственных растений Дальнего Востока / АН СССР, ДВНЦ, Биол.почв. ин-т. – Владивосток, 1971. – Вып. 3. – С. 171 – 178. 60. Енукидзе, Н.В. Болезни сои в Грузии и обоснование мер борьбы с ними: автореф. дис. … канд. биол. наук: 03.02.08 / Енукидзе Н.В. – Тбилиси, 1981. – 37 с. 61. Есипенко, Л.П. Борьба с корневой гнилью – важный фактор повышения выхода семян и зелённой массы сои / Л.П. Есипенко // Проблемы кормопроизводства на Дальнем Востоке. – Новосибирск, 1984. – С. 95 – 99. 62. Жуковская, С.А. Увядание зернобобовых культур / С.А. Жуковская // Защита растений. – 1968. – №10. – С. 45 – 46. 124 63. Жуковская, С.А. Корневые гнили сои в Приморском крае / С.А. Жуковская // Микология и фитопатология. – 1977. – Т.11, вып. 2. – С.140 –144. 64. Жуковская, С.А. Возбудители грибных болезней сои / С.А. Жуковская, А.М. Овчинникова // Возбудители болезней с.-х. растений Дальнего Востока / АН СССР, ДВНЦ, Биолого-почв. ин-т. – М.: Наука, 1980. – С. 5 – 50. 65. Жуковская, С.А. О вредоносности корневых гнилей сои / С.А. Жуковская // Пути повышения продуктивности растениеводства на Дальнем Востоке. – Владивосток, 1982. – С. 144 – 153. 66. Жуковская, С.А. Болезни сои на Дальнем Востоке / С.А. Жуковская, Н.И. Серебренникова, Л.С. Куликова // Защита растений. – 1990. – №10. – С. 42 – 45. 67. Жученко, А.А. Адаптивная система селекции растений (экологогенетические основы): монография: в 2 т. / А.А. Жученко. – М.: Изд-во РУДН, 2001. – Т. 1. – 780 с. 68. Заостровных, В.И. Изучение некоторых элементов агротехники сои в условиях Кемеровской области / В.И. Заостровных, А.А. Бакач, Г.А. Незнамова // Сб. науч. тр. преподавателей и сотрудников агрономического и экономического факультетов Кемеров. фил. Новосиб. ГАУ. – Кемерово, 1991. – Вып. 4. – С. 67 – 70. 69. Заостровных, В.И. Иммунологическое изучение исходного материала зернобобовых культур / В.И. Заостровных, Т.А. Манакова, С.И. Рудакова [и др.] // Почва, жизнь, благосостояние: сб. материалов Всерос. конф. – Пенза, 2000. – С. 263 – 266. 70. Заостровных, В.И. Вредные организмы сои и система фитосанитарной оптимизации её посевов: монография / В.И. Заостровных, Л.К. Дубовицкая; под ред. В.А. Чулкиной. – Новосибирск, 2003. – 582 с. 71. Золотницкий, В.А. Соя на Дальнем Востоке / В.А. Золотницкий; под ред. Е.А. Старостина. – Хабаровск: кн. изд-во, 1962. – 248 с. 125 72. Зональная система земледелия Амурской области / Кузин В.Ф. [и др.]; отв. ред. В.Ф. Кузин. – Благовещенск: Хабаровское кн. изд-во, Амурское отд-е, 1982. – 228 с. 73. Иванова, А.Н. Вредители и болезни сои, и меры борьбы с ними / А.Н. Иванова, З.В. Холопова. – Владивосток, 1956. – 31 с. 74. Иваченко, Л.Е. Активность и множественные формы ферментов в семенах сои, полученных в разных агроклиматических условиях Амурской области. Часть 6. Каталаза / Л.Е. Иваченко, В.И. Шаройко, Г.П. Ефимова // Проблемы экологии Верхнего Приамурья: сб. научн. тр. / под общ. ред. Л.Г. Колесниковой. – Благовещенск: Изд-во БГПУ, 1997. – Вып. 3. – С. 104 – 108. 75. Ильинская, Л.И. Биохимические аспекты индуцированной устойчивости / Л.И. Ильинская, Н.И. Васюкова, О.Л. Озерецковская // Итоги науки и техники. Сер. Защита растений. – М.: ВИНИТИ, 1991. – Т. 7. – С. 196 – 200. 76. Карташова, Е.Р. Полифункциональность растительных пероксидаз и их практическое использование / Е.Р. Карташова, Г.Н. Руденская, Е.В. Юрина // Сельскохозяйственная биология. – 2000. – № 5. – С. 63 – 70. 77. Клыков, А.П. Пути оздоровления сои от бактериальных заболеваний / А.П. Клыков // Земледелие. – 1953. – № 5. – С. 9 – 10. 78. Кожанова, О.Н. Некоторые свойства пероксидазы здоровых и зараженных Botrytis cinerea тканей капусты / О.Н. Кожанова, В.А. Аксенова // Прикладная биохимия и микробиология. – 1976. – Т. 12, № 5. – С. 753 – 758. 79. Козырева, Е.П. Болезни сои / Е.П. Козырева, М.А. Примаковская, О.В. Скрипка // Защита растений. – 1982. – № 11. – С. 38 – 39. 80. Конарев, В.Г. Белки растений как генетические маркеры / В.Г. Конарев. – М.: Колос, 1983. – 320 с. 81. Коно, А. Взаимоотношения между активностью каталазы, пероксидазы, цитохромоксидазы и устойчивостью овса к корончатой ржавчине / А. Коно // Биология. – 1961. – № 23. – С. 23. 126 82. Корецкий, П.М. Биология возбудителя ложной мучнистой росы сои Peronospora manshurica (Naumov) Sydow и меры борьбы с ними: автореф. дис. … канд. биол. наук. / Корецкий П.М. – Л.-Киев, 1967. – 20 с. 83. Короневский, В.И. К методике статистической обработки данных многолетних полевых опытов / В.И. Короневский // Земледелие. – 1985. – № 11. – С. 56 – 57. 84. Корочкин, Л.И. Генетика изоферментов / Л.И. Корочкин [и др.]. – М.: Наука, 1977. – 278 с. 85. Корочкин, Л.И. Генетика изоферментов и проблемы адаптации / Л.И. Корочкин // Экологическая генетика и эволюция. – Кишинев: Штиинца, 1987. – С. 135 – 146. 86. Корсаков, Н.И. Изучение устойчивости сои к грибным болезням: метод. указ. / Н.И. Корсаков, А.М. Овчинникова, В.И. Мизева // ВИР. – Л., 1979. – 46 с. 87. Котова, В.В. Методические указания по диагностике корневых гнилей зернобобовых культур / В.В. Котова, М.Ю. Степанова // ВИЗР. – Л., 1979. – 28с. 88. Краснова, М.В. Бактериозы сои и борьба с ними на Северном Кавказе / М.В. Краснова // Агробиология. – 1963. – №5. – С. 738 – 743. 89. Кретович, В.Л. Молекулярные механизмы биологической фиксации азота / В.Л. Кретович // Вестник АН СССР. – 1979. – № 7. – С. 23 – 32. 90. Кузнецова, С. Белая гниль сои / С. Кузнецова // Зернобобовые культуры. – 1964. – №2. – С. 36. 91. Лавриненко, Г.Т. Соя / Г.Т. Лавриненко [и др.] – М.: Россельхозиздат, 1978. – 189 с. 92. Ладыгина, М.Е. Особенности изоэнзимного состава пероксидазы и полифенолоксидазы при вирусном патогенезе табака / М.Е. Ладыгина, Э.А. Таймла, Б.А. Рубин // Физиология растений. – 1970. – Т. 17, вып. 5. – С. 928 – 935. 93. Ладыгина, М.Е. Влияние заражения вирусом Y на белково-ферментный комплекс растений картофеля / М.Е. Ладыгина // Труды НИИКХ. – 1971. – Т. 8. – С. 14 – 19. 127 94. Левитес, Е.В. Генетика изоферментов растений / Е.В. Левитес. – Новосибирск: Наука, 1986. – 144 с. 95. Ленинджер, А. Биохимия. Молекулярные основы структуры и функции клетки / А. Ленинджер. – М.: Мир, 1999. – С. 390 – 422. 96. Лещенко, А.К. Культура сои / А.К. Лещенко. – Киев: Наукова думка, 1978. – 236 с. 97. Лисицын, Е.М. Динамика ферментативной активности корней проростков овса при действии кислотного стресса / Е.М. Лисицын, А.В. Кропотов // Доклады РАСХН. – 2002. – № 2. – С. 6 – 8. 98. Лукашевич, И.В. Активность и изоферментный состав кислой фосфатазы гибридов и линий кукурузы / И.В. Лукашевич, И.В. Хамматова, А.И. Левина [и др.] // Физиология растений. – 1974. – Т. 21, вып. 6. – С. 1168 – 1173. 99. Лукьянович, Т.И. Эффективность биосила и бинорама против комплекса грибных болезней сои в условиях Приморского края / Т.И. Лукьянович, А.Ю. Санкин // Энергосберегающие технологии возделывания сельскохозяйственных культур в условиях Дальнего Востока. – Владивосток: Дальнаука, 2006. – С. 346 – 348. 100. Максимов, И.В. Связь устойчивости проростков Aegilops umbellulata zhuk. к Septoria nodorum berk. с изоферментным составом пероксидаз / И.В. Максимов, Е.А. Черепанова, Г.Ф. Муртазина [и др.] // Известия РАН. Серия биологическая. – 2006. – № 5. – С. 575 – 580. 101. Максимов, И.В. Влияние бактерий Bacillus subtilis 26D на содержание пероксида водорода и активность пероксидазы в растениях яровой пшеницы / И.В. Максимов, Р.Р. Абизгильдина, З.Р. Юсупова [и др.] // Агрохимия. – 2010. – № 1. – С. 55 – 60. 102. Максимов, И.В. Структурно-функциональные особенности изопероксидаз растений / И.В. Максимов, Е.А. Черепанова, Г.Ф. Бурханова [и др.] // Биохимия. – 2011. – Т. 76, вып. 6. – С. 749 – 763. 103. Малый практикум по физиологии растений / под ред. А.Т. Мокроносова. – М.: МГУ, 1994. – 184 с. 128 104. Метлицкий, Л.В. Фитоиммунитет. Молекулярные механизмы / Л.В. Метлицкий. – М.: Наука, 1976. – 51 с. 105. Метлицкий, Л.В. Как растения защищаются от болезней / Л.В. Метлицкий, О.Л. Озерецковская. – М.: Наука, 1985. – 192 с. 106. Методика государственного сортоиспытания сельскохозяйственных культур / под ред. М.А. Федина. – М.: Калининская областная типография управления издательств, полиграфии и книжной торговли Калининского облисполкома, 1985. – Вып. 1. – 269 с. 107. Методы биохимических исследований растений / под ред. А.И. Ермакова. – Л.: Агропромиздат, 1987. – 430 с. 108. Мизева, В.И. Результаты исследований по иммунитету сои к грибным заболеваниям в Приамурье / В.И. Мизева, А.П. Ващенко // Пути повышения продуктивности растениеводства на Дальнем Востоке. –Владивосток: ДВНЦ АН СССР, 1981. – С. 118 – 123. 109. Минибаева, Ф.В. Продукция супероксида и активность внеклеточной пероксидазы в растительных тканях при стрессе / Ф.В. Минибаева, Л.Х. Гордон // Физиология растений. – 2003. – Т. 50, № 3. – С. 459 – 464. 110. Миронова, Г.В. Защита сои от инфекционных болезней / Г.В. Миронова // Защита и карантин растений. – 1995. – № 3. – С. 34. 111. Михайленко, А.М. Болезни зернобобовых в Приморском крае / А.М. Михайленко // Защита растений. – 1965. – №2. – С.41 – 43. 112. Муравьёва, М.Ф. Мозаика сои в Приморском крае / М.Ф. Муравьёва // Защита растений. – 1967. - № 2. – С. 20 – 21. 113. Муравьёва, М.Ф. Вирусные болезни зернобобовых культур в Приморском крае / М.Ф. Муравьёва // Вирусные болезни сельскохозяйственных культур Дальнего Востока / АН СССР. Сиб. отд-ние. Биол.-почв. ин-т. – Владивосток, 1969. – Вып. 1. – С. 105 – 111. 114. Муравьева, М.Ф. Вирусные болезни в Хабаровском крае / М.Ф. Муравьёва // Биология и возделывание сои / АН СССР. Дальневост. науч. центр, Биолого-почв. ин-т. – Владивосток, 1971. – С. 191 – 193. 129 115. Муравьёва, М.Ф. Устойчивость к болезням сортообразцов сои / М.Ф. Муравьёва // Защита растений на Дальнем Востоке. – Новосибирск, 1981. – С. 55 – 60. 116. Наумов, Н.А. Болезни технических культур / Н.А. Наумов // Справочник агронома по борьбе с болезнями сельскохозяйственных растений. – М.: Сельхозгиз, 1936. – 27 с. 117. Наумова, Е.С. Влияние метеорологических факторов на развитие патогенных грибов, поражающих сою / Е.С. Наумова // Микология и фитопатология. – 1986. – Т. 20, вып. 3. – С. 227 – 232. 118. Нелен, Е.С. Вертициллез сои / Е.С. Нелен, С.А. Жуковская // Защита растений. – 1967. – № 8. – С. 45 – 46. 119. Нелен, Е.С. Антракноз сои / Е.С. Нелен, С.А. Жуковская // Защита растений. – 1968. – №6. – С. 45. 120. Никитина, А.И. Опасные болезни сои на Дальнем востоке /А.И. Никитина // Защита растений от вредителей и болезней. – 1962. – №7. – С. 37 – 40. 121. Нилова, В.П. Активность каталазы и пероксидазы и иммунитет пшеницы к бурой ржавчине (Puccinia triticina Erikss.) / В.П. Нилова, Г.Н. Егорова // Доклады ВАСХНИЛ. – 1948. – Вып. 1. – С. 34 – 38. 122. Новотельнова, Н.С. Пероноспоровые грибы-патогены культурных растений в СССР: справочник по диагностике и методам исследования / Н.С. Новотельнова, К.А. Пыстина, О.Г. Голубева. – Л.: Наука, 1979. – 151 с. 123. Овчинникова, A.M. Церкоспороз сои в Приморском крае / А.М. Овчинникова // Защита растений. – 1968. – № 1. – С. 11 – 13. 124. Овчинникова, А.М. Патогенная микрофлора сои / А.М. Овчинникова // Тр. / ВИЗР. – 1970. – Вып. 29. – С. 73 – 80. 125. Овчинникова, А.М. Грибные болезни сои / А.М. Овчиникова // Болезни и вредители сои на юге Дальнего Востока и меры борьбы с ними. – Владивосток, 1971а. – С. 5 – 72. 130 126. Овчинникова, А.М. Патогенная микрофлора сои на Дальнем Востоке / А.М. Овчиникова // Биология, экология, география споровых растений Средней Азии. – Ташкент: Изд-во Фан, 1971б. – С. 205 – 206. 127. Овчинникова, А.М. Болезни семян на Дальнем Востоке и методика их фитопатологического анализа / А.М. Овчинникова, В.И. Потлайчук // ВИЗР. – Л., 1980. – Вып. 48. – С. 41 – 46. 128. Омельченко, С.И. Фермент пероксидаза в пораженных растениях / С.И. Омельченко, В.А. Андреева // Метаболизм больного растения. – Владивосток: ДВНЦ АН СССР, 1976. – Т. 40 (143). – С. 21 – 44. 129. Перепадя, Ю.Г. Особенности дыхания и ферментной активности перцев в связи с устойчивостью к неблагоприятным условиям среды / Ю.Г. Перепадя, Г.П. Диканев // Науч.-техн. бюл. / ВАСХНИЛ. – Л., 1977. – Вып. 74. – С. 24 – 27. 130. Пересыпкин, В.Ф. Сельскохозяйственная фитопатология: учеб. для студентов вузов по спец. "Защита растений" / В.Ф. Пересыпкин. – 4 изд., перераб. и доп. – М.: Агропромиздат, 1989а. – 480 с. 131. Перфильева, А.И. Разнонаправленность защитных реакций картофеля при биотическом и абиотическом стрессах / А.И. Перфильева, Е.В. Рымарева // Агро XXI. – 2012. – № 10-12. – С. 24 – 26. 132. Плешков, Б.П. Биохимия сельскохозяйственных растений / Б.П. Плешков. – М.: Колос. – 1980. – 495 с. 133. Плохинский, Н.А. Биометрия / Н.А. Плохинский. – 2-е изд., перераб. и доп. – М.: Изд-во МГУ, 1970. – 560 с. 134. Подкина, Д.В. Патогенная микрофлора в Краснодарском крае / Д.В. Подкина // Науч.- техн. бюл. / ВНИИ масл. культур. – 1981. – №78. – С. 17 – 24. 135. Подкина, Д.В. Болезни сои на Кубани / Д.В. Подкина, О.А. Лавриченко // Масличные культуры. – 1982. – №5. – С. 30 – 32. 136. Полесская, О.Г. Растительная клетка и активные формы кислорода: учебное пособие / О.Г. Полесская; под ред. И.П. Ермакова. – М.: КДУ. – 2007. – 140 с. 131 137. Полякова, Е.Н. Активность ферментов при нарушении покоя и прорастания семян / Е.Н. Полякова // Роль температуры и фитогормонов в нарушении покоя семян. – Л.: Наука, 1981. – С. 137 – 148. 138. Поливанова, Т.А. Вирусные болезни сои на Дальнем Востоке: автореф. дис. … канд. биол. наук / Т.А. Поливанова. – Владивосток, 1968. – 24 с. 139. Поливанова, Т.А. Вирусные болезни сои / Т.А. Поливанова // Болезни и вредители сои на юге Дальнего Востока и меры борьбы с ними / Биолого-почв. ин-т., ДВ СТАЗР. Уссур. МЖК. – Владивосток, 1971. – Гл. 3. – С. 104 – 144. 140. Поливанова, Т.А. Возбудители вирусных болезней сои // Возбудители болезней сельскохозяйственных растений Дальнего Востока / АН СССР, ДВНЦ, Биолого-почв. ин-т. – М.: Наука, 1980. – С. 51 – 70. 141. Поливанова, Т.А. Вирусы, идентифицированные на зернобобовых культурах на Дальнем Востоке / Т.А. Поливанова, А.В. Крылов // Взаимоотношения вирусов с клетками растения-хозяина. – Владивосток: ДВНЦ АН СССР, 1985. – С. 87 – 93. 142. Посевные площади, валовые сборы и урожайность сельскохозяйственных культур по категориям хозяйств Амурской области: Стат. сб. Амурстат. – Благовещенск, 2012. – 258 с. 143. Посыпанов, Г.С. Видовой состав основных патогенов сои и их вредоносность в условиях Центрального Нечерноземья: информ. лист / Г.С. Посыпанов, Т.Д. Сихарулидзе, Е.В. Демьяненко // КМТ ЦНТИ. – Калуга, 1995. – № 57. – 95 с. 144. Прогноз развития сельского хозяйства на Дальнем Востоке на 20152020 гг. / Рос. акад. с.-х. наук, Дальневост. регион. науч. центр Россельхозакадемии, Гос. науч. учреждение Дальневост. науч.-исслед. ин-т экономики, орг. и планирования АПК Россельхозакадемии / А.С. Шелепа и др. – Хабаровск: РИЦ ХГАЭП, 2011. – 100 с. 145. Простакова, Ж.Г. Расовый состав возбудителя пероноспороза сои и эффективность гена устойчивости в условиях центральной зоны Молдавии / Ж.Г. 132 Простакова // 4 съезд генетиков и селекционеров Молдавии. – Кишинев: Штиинца, 1981. – С. 97. 146. Простакова, Ж.Г. Грибные болезни сои и меры борьбы с ними / Ж.Г. Простакова, А.И. Ганя // АН Молдавской СССР. – Кишинев: Штиинца, 1983. – 35 с. 147. Райдер, К. Изоферменты / К. Райдер, К. Тейлор; перевод с англ. М.Д. Гроздовой. – М.: Мир, 1983. – 107 с. 148. Рейфман, В.Г. Вирусные болезни сои в Амурской области / В.Г. Рейфман, Т.А. Поливанова // Проблемы с.-х. Приморья. – Благовещенск, 1968. – С. 177 – 178. 149. Рогожин, В.В. Пероксидаза как компонент антиоксидантной системы живых организмов / В.В. Рогожин. – Спб.: ГИОРД, 2004. – 240 с. 150. Родина, З.И. Вредители и болезни сои в Ставропольском крае / З.И. Родина // Соя в Ставропольском крае. – Ставрополь, 1981. – С. 170 – 179. 151. Рубин, Б.А. О механизме активации пероксидазы в инфицированной ткани иммунных растений / Б.А. Рубин, Т.М. Иванова, М.А. Давыдова // Прикладная биохимия и микробиология. – 1965. – Т. 1(1). – С. 25 – 35. 152. Рубин, Б.А. Энзимология и биология дыхания растений / Б.А. Рубин, М.Е. Ладыгина. – М.: Высшая школа, 1966. – 288 с. 153. Рубин, Б.А. Динамика компонентов пероксидазы листьев кукурузы при поражении пузырчатой головней / Б.А. Рубин, Е.Н. Лущинская // Биохимия. – 1969. – Т. 34, вып. 4. – С. 674 – 678. 154. Рубин, Б.А. Об изоферментах пероксидазы в клубне картофеля / Б.А. Рубин, Е.В. Будилова // ДАН СССР. – M., 1970. – Т. 190, вып. 3. – С. 722 –724. 155. Рубин, Б.А. Новообразование белка пероксидазы в тканях капусты под влиянием заражения Botrytis sinerea / Б.А. Рубин, В.А. Аксенова, О.Н. Кожанова // ДАН СССР. – M., 1973. – Т. 210, №. 2. – С. 485 – 488. 156. Рубин, Б.А. Изменение пероксидазной активности листьев пшеницы под влиянием заражения Puccinia graminis у сортов, различающихся по 133 устойчивости / Б.А. Рубин, Е.В. Юрина // Докл. ВАСХНИЛ. – М., 1974. – №3. – С. 7 – 9. 157. Рубин, Б.А. Биохимия и физиология иммунитета растений / Б.А. Рубин, Е.В. Арциховская, В.А. Аксенова. – М.: Высшая школа, 1975. – 320 с. 158. Рубин, Б.А. Физиология и биохимия дыхания растений: учеб. пособие для биол. спец. ун-тов / Б.А. Рубин, М.Е. Ладыгина. – М.: Высшая школа, 1977. – 512 с. 159. Савич, И.М. Удельная активность изопероксидаз проростков кукурузы и пшеницы в условиях низкотемпературного стресса / И.М. Савич, Т.Л. Таджибаева, Ю.В. Перуанский // Физиология и биохимия культурных растений. – 1988. – Т. 20(2). – С. 128 – 133. 160. Савич, И.М. Пероксидазы – стрессовые белки растений / И.М. Савич // Успехи современной биологии. – 1989а. – Т. 107, вып. 3. – С. 406 – 417. 161. Савич, И.М. Аминокислотный состав щелочных изопероксидаз кукурузы при действии низкотемпературного стресса / И.М. Савич // Прикладная биохимия и микробиология. – 1989б. – Т. 25(5). – С. 634 – 643. 162. Савич, И.М. Изопероксидазы проростков сорго при холодовом стрессе / И.М. Савич // Физиология и биохимия культурных растений. – 1989в. – Т. 20(6). – С. 566 – 572. 163. Савич, И.М. Изменение антигенной структуры основных изопероксидаз проростков кукурузы при криострессе / И.М. Савич // Физиология растений. – 1990. – Т. 37(4). – С. 766 – 773. 164. Садвакасова, Г.Г. Некоторые физико-химические и физиологические свойства пероксидазы растений / Г.Г. Садвакасова, Р.М. Кунаева // Физиология и биохимия культурных растений. – 1987. – Т. 19, № 2. – С. 107 – 119. 165. Саундерс, Б.К. Пероксидазы и каталазы / Б.К. Саундерс; перевод с англ. И.Я. Левитина // Неорганическая биохимия. – М.: Мир, 1972. – Т. 2. – С. 434 – 470. 166. Сафонов, В.И. Об условиях проявления кислых фосфатаз (К.Ф.3.1.3.2) растений на электрофореграммах в полиакриламидном геле / В.И. Сафонов, М.П. 134 Сафонова, В.И. Суворов // Электрофорез в полиакриламидном геле и его применение в биологии, сельском хозяйстве, медицине и пищевой промышленности: Тр. Всесоюзного семинара, 14-18 декабря 1971г. – М., 1971. – С. 59 – 62. 167. Селихова, О.А. Генетические и экологические особенности биохимического состава семян исходного материала для селекции сои: дис. … канд. с.-х. наук: 06.01.05 / Селихова Ольга Александровна. – п. Тимирязевский, 2003. – 171 с. 168. Семенова, Е.А. Активность ферментов у сортов сои с различной степенью устойчивости к септориозу / Е.А. Семенова, С.А. Титова, Л.К. Дубовицкая // Достижения науки и техники АПК, 2012. – № 4. – С. 24 – 26. 169. Семенова, Е.А. Оценка экологической приспособленности Glycine max (L.) Merr. и Glycine soja по энзиматической активности в онтогенезе: дис. …канд. биол. наук: 03.00.16 / Семенова Елена Александровна. – Благовещенск, 2006. – 136 с. 170. Серова, З.Я. Окислительно-восстановительные процессы инфицированного растения / З.Я. Серова, Г.М. Подчуфарова, Д.К. Гесь. – Мн.: Наука и техника, 1982. – 232 с. 171. Система земледелия Амурской области / отв. ред. В.А. Тильба. – Благовещенск: ИПК «Приамурье», 2003. – 304 с. 172. Система земледелия учебно-опытного хозяйства ДальГАУ: методические рекомендации / В.В. Голубев, А.П. Емельянов [и др.]; под общ, ред. П.В. Тихончука и В.Ф. Кузина. – Благовещенск, 1997. – 119 с. 173. Скулачев, В.П. Кислород в живой клетке: добро и зло / В.П. Скулачев // Соросовский образовательный журнал; под ред. Сойорер В.Н. – М.: Изд-во МГУ, 1996. – № 3. – С. 4 – 10. 174. Солотчина, Г.Ф. Видовой состав возбудителей бактериозов сои в Приморском крае / Г.Ф. Солотчина, Э.Ш. Пехтерева. – Киев, 1966. – 78 с. 135 175. Солотчина, Г.Ф. Бактериальные болезни сои / Г.Ф. Солотчина // Болезни и вредители сои на юге Дальнего Востока и меры борьбы с ними. – Владивосток, 1971. – С. 73 – 100. 176. Солотчина, Г.Ф. Бактериальные болезни сои / Г.Ф. Солотчина // Болезни сои на юге Дальнего Востока и меры борьбы с ними. – Л., 1980. – 20 с. 177. Соя / Ю.П. Мякушко [и др.]; под ред. Ю.П. Мякушко, В.Ф. Баранова. – М.: Колос, 1984. – 332 с. 178. Стэкмен, Э. Основы патологии растений / Э. Стэкмен, Д. Харрар. – М., 1959. – 549 с. 179. Сунь Син-Дун. Соя / Сунь Син-Дун; пер. с кит. А.М. Кайгородова. – М.: Сельхозгиз, 1958. – 248 с. 180. Тарр, С. Основы патологии растений / С. Тарр; перевод с англ. Л.М. Дунина, Н.Л. Клячко. – М.: Мир, 1975. – 587 с. 181. Тарчевский, И.А. Сигнальные системы клеток растений / И.А. Тарчевский. – М.: Наука, 2002. – 294 с. 182. Технологии и комплекс машин для производства зерновых культур и сои в Амурской области: коллективная научная монография / В.А. Тильба, В.Т. Синеговская, А.Н. Панасюк, М.М. Присяжный [и др.] – Благовещенск: Изд-во ООО «Агромакс-Информ», 2011. – 134 с. 183. Титов, А.Ф. Изопероксидазы растений / А.Ф. Титов // Успехи современной биологии. – Петрозаводск. – 1975. – Т. 80, вып. 1. – С. 102 – 115. 184. Титов, А.Ф. Полиморфизм ферментных систем и устойчивость к экстремальным (низким) температурам / А.Ф. Титов // Успехи современной биологии. – Петрозаводск, 1978. – Т. 85, вып. 1. – С. 63 – 70. 185. Тихончук, П.В. Соя: морфология, биология, технология возделывания: учебн. пособие / П.В. Тихончук, Ю.В. Оборская. – Благовещенск: ДальГАУ, 2010. – 131 с. 186. Торопава, Е.Ю. Экологические основы защиты растений от болезней в Сибири / Е.Ю. Торопава; под ред. В.А. Чулкиной. – Новосибирск, 2005. – 370 с. 136 187. Удовенко, Г.В. Исследование физиологии устойчивости растений к неблагоприятным условиям среды / Г.В. Удовенко // Труды по прикладной ботанике, генетике и селекции. – 1975. – Т. 56, вып. 1. – С. 154 – 161. 188. Уилкинсон, Дж. Изоферменты / Дж. Уилкинсон; перевод с англ. Б.И. Курганова; под ред. В.О. Шпикитера. – М.: Мир, 1968. – 224 с. 189. Фалеева, Л.К. Болезни сои и оценка на устойчивость генетического материала / Л.К. Фалеева, А.Я. Ала // Вопросы возделывания основных сельскохозяйственных культур в Амурской области. – Новосибирск, 1976. – С. 56 – 58. 190. Филиппович, Ю.Б. Практикум по общей биохимии / Ю.Б. Филиппович, Т.А. Егорова, Г.А. Севастьянова. – М.: Просвещение, 1982. – 311 с. 191. Фитосанитарный прогноз: распространения главнейших вредителей и болезней сельскохозяйственных культур в Амурской области в 2009 году и меры борьбы с ними; прогноз разработан Ю.В. Положиевой; под руководством Н.П. Домчук. – Благовещенск, 2009. – 43 с. 192. Фитосанитарный прогноз: распространения главнейших вредителей и болезней сельскохозяйственных культур в Амурской области в 2010 году и меры борьбы с ними; прогноз разработан Ю.В. Положиевой; под руководством Н.П. Домчук. – Благовещенск, 2010. – 42с. 193. Фитосанитарный прогноз: распространения главнейших вредителей и болезней сельскохозяйственных культур в Амурской области в 2011 году и прогноз развития вредных объектов в 2012 году; прогноз разработан Ю.В. Положиевой; под руководством Н.П. Домчук. – Благовещенск, 2011. – 130 с. 194. Фитосанитарный прогноз: распространения главнейших вредителей и болезней сельскохозяйственных культур в Амурской области в 2012 году и меры борьбы с ними; прогноз разработан Ю.В. Положиевой; под руководством Н.П. Домчук. – Благовещенск 2012. – 123с. 195. Хайрулина, Т.П. Влияние низкой положительной температуры на активность каталазы, пероксидазы и продуктивность сои / Т.П. Хайрулина, П.В. 137 Тихончук, Е.А. Семенова // Достижения науки и техники АПК, 2010. –№ 7. – С. 8 – 10. 196. Хохряков, М.К. Определитель болезней растений / М.К. Хохряков, Т.Л. Доброзракова, К.М. Степанов, М.Ф. Летова. – Л.: Колос, 1976. – 592 с. 197. Хочачка, П. Стратегия биохимической адаптации / П. Хочачка, Дж. Сомеро; перевод с англ. Ю.И. Лашкевича; под ред. и с предис. Е.М. Крепса. – М.: Мир, 1977. – 398 с. 198. Черноголовин, В.П. Соя в восточных районах страны / В.П. Черноголовин [и др.] – Благовещенск: Дальневосточный научн.-исслед. ин-т сельского хозяйства, 1971. – 125 с. 199. Чесноков, Ю.В. Устойчивость растений к патогенам / Ю.В. Чесноков // Сельскохозяйственная биология. – 2007. – № 1. – С. 16 – 33. 200. Чеснокова, Н.П. Общая характеристика источников образования свободных радикалов и антиоксидантных систем / Н.П. Чеснокова, Е.В. Понукалина, М.Н. Бизенкова // Успехи современного естествознания. – 2006. – № 7. – С. 37 – 41. 201. Чигрин, В.В. Активность фенолокисляющих ферментов и динамика редуцирующих соединений у двух сортов яровой пшеницы в норме и при заражении стеблевой ржавчиной / В.В. Чигрин [и др.] // Физиология растений. – 1968. – Т. 15, вып. 2. – С. 336 – 342. 202. Шаройко, В.И. Активность и множественные формы ферментов в семенах сои, полученных в разных агроклиматических условиях Амурской области. Часть 8. Кислая фосфатаза / В.И. Шаройко, Л.Е. Иваченко, О.А. Селихова // Проблемы экологии Верхнего Приамурья: сб. научн. тр. / под общ. ред. Л.Г. Колесниковой. – Благовещенск: Изд-во БГПУ, 2002. – Вып. 6. – С. 109 – 113. 203. Шерепитко, В.В. Соя: Аспекты устойчивости, методы оценки и отбора / В.В. Шерепитко [и др.] – Кишинев: Штиинца, 1990. – 178 с 138 204. Шнырева, М.Г. Кинетика секреции и надмолекулярная организация внеклеточных кислых фосфатаз Saccharomyces cerevisiae: автореф. дис. … канд. биол. наук / Шнырева М.Г. – Москва, 1992. – 18 с. 205. Щегорец, О.В. Соеводство: учебное пособие / О.В. Щегорец. – Благовещенск: ООО Издательская компания РИО, 2002. – 432 с. 206. Эванс, Э. Болезни растений и химическая борьба с ними / Э. Эванс; перевод с англ. Н.А. Емельяновой; под ред. Е.Ф. Гранина. – М.: Колос, 1971. – 228 с. 207. Эчкалов, А.П. Выделение и некоторые свойства кислой фосфатазы шелкоотделительной железы тутового шелкопряда / А.П. Эчкалов, Ю.Б. Филиппович // Биохимия насекомых (ферменты метаболизма): межвузовский сб. научн. тр. – М., 1984. – С. 38 – 50. 208. Юрина, Т.П. О влиянии системных фунгицидов на ферментативную активность и устойчивость пшеницы к мучнистой росе / Т.П. Юрина, Е.В. Юрина, В.А. Караваева и др. / Тр. Междунар. конф., посвященной 80-летию кафедры микологии и альгологии МГУ и 90-летию со дня рождения М.В. Горленко // В сб.: Современные проблемы микологии, альдологии и фитопатологии. – М., 1998. – С. 134 – 135. 209. Юсупова, З.Р. Активность пероксидазы в различных клеточных фракциях при инфицировании пшеницы Septoria nodorum berk. / З.Р. Юсупова, Р.М. Хайруллин, И.В. Максимов // Физиология, растений. – 2006. – Т. 53(6). – С. 910 – 917. 210. Яаска, В. Изоферменты как молекулярно-генетические маркеры в селекции растений / В. Яаска // Молекулярные механизмы генетических процессов. – 1985. – С. 251 – 266. 211. Яковлева, Н.П. Фитопатология / Н.П. Яковлева. – 2-е изд., перераб. и доп. – М.: Колос, 1992. – 384с. 212. Яруллина, Л.Г. Защитная роль оксалатоксидазы при поражении пшеницы возбудителем корневой гнили Bipolaris sorokiniana / Л.Г. Яруллина, 139 И.В. Максимов, Г.Ф. Муртазина // Микология и фитопатология. – 2005. – Т. 39, № 4. – С. 90 – 96. 213. Andreew, L. A note on the effect of rust infection on peroxidase isozymes in flax / L. Andreew, M. Shaw // Canad. J. Botany. – 1965. – Vol. 43, N 11. – P. 1479 – 1483. 214. Bernaux P. Le Cercospora kikuchii Mats et Tom. du soja en France. – Ann. / P. Bernaux // Phytopathol. – 1976. – Vol. 8, N 1. – P. 83 – 86. 215. Bolwell, G.P. Recent advances in understanding the origin of the apoplastic oxidative burst in plant cells / G.P. Bolwell, K.A. Blee, V.S. Butt et al. // Free Radical Res. – 1999. – Vol. 31. – P. 137 – 145. 216. Boosalis, M.G. Studies on the parasitism of Rhizoctonia solani (Kuehn.) on soybeans / M.G. Boosalis // Phytopathology. – 1950. – Vol. 40, N 9. –P. 820 – 831. 217. Brown, A.C. The reversal of tumor growth / A.C. Brown // Soi. Amer. – 1965. – Vol. 213, N 5. – P. 1 – 9. 218. Chant, S.R. The affect of tobacco mosaic virus and potato virus X on peroxidase activity and peroxidase isosymes in Nicotina glutinosa / S.R. Chant, D.C. Bates // Phytochemistry. – 1970. – Vol. 9, N 11. – P. 2323 – 2326. 219. Chauhan, K.P.S. Electrophoretic variations of proteins and enzymes in relation to seed quality / K.P.S. Chauhan, M.C. Gopinathan, C.R. Babu // Seed Science and Technology. – 1985. – Vol. 13. – P. 629 – 641. 220. Christensen, J.H. Purification and characterization of peroxidases correlated with lignifications in poplar xylem / J.H. Christensen, G. Bauw, K.G. Welinder // Plant Physiol. – 1998. – Vol. 118. – P. 125 – 135. 221. Daly, J.M. The use of near-isogenic lines in biochemical studies of the resistance of wheat to stem rust / J.M. Daly // Phytopathology. – 1972. – Vol. 62, N 4. – P. 392 – 400. 222. Deutschmann, F. Über die “Purple stain” Krankheit der sojabohnen und di Farbstoffbildung ihres Erregers Cercosporina kikuchii (Mats.) et Tomoyasu / F. Deutschmann // Phytopathol. L., Bd. – 1952. – N. 3. – P. 297 – 310. 140 223. Droillrd, M.J. Pree radical production, catalase and superoxide dismutase activities and membrane integrity during senescence of petals of cut camations / M.J. Droillrd, A. Paulin, J.C. Massot // Physiol plantarum. – 1987. – Vol. 71, N 2. – P. 197 – 203. 224. Dröge, W. Free Radicals in the Physiological Control of Cell Function / W. Dröge // Physiol Rev. – 2002. – Vol. 82. – P. 47 – 95. 225. Duff, S.M.G. The role of acid phosphotases in plant phosphorus metabolism / S.M.G. Duff, G. Sarath, W.C. Plaxton // Physiol. Plantarum. – 1994. – Vol. 90, N 4. – P. 791 – 800. 226. Dunleavy, J.M. Effects of air temperature on disease severity and peroxidase activity of soybean leaves infected by Peronospora manshurica / J.M. Dunleavy // Crop. Sci. – 1982. – Vol. 22. – P. 623 – 625. 227. Farkas, G.L. Role of oxidative metabolism in the localization of plaut viruses / G.L. Farkas, Kiraly Z., Solymosy F. // Virology. – 1960. – Vol. 12, N 3. – P. 408 – 421. 228. Farkas, G.L. On the nature of changes in peroxidase isoenzymes in bean leaves infected by southern bean mosaic virus / G.L. Farkas, M.A. Stahmann // Phytopathology. – 1966. – Vol. 56, N 6. – P. 669 – 677. 229. Fieldes, M.A. Flax guaiacol peroxidases can be used to illustrate the possibility of misinterpreting the effects of stress on the activity of developmentally regulated enzymes / M.A. Fieldes, K.E. Gerhardt // Plant Science. – 1998. – Vol. 132. – P. 89 – 99. 230. Filipe, M.R. Cytochemical study of catalass and peroxidase in the mesophull of Lolium rigidum plants treated with isoproturon / M.R. Filipe, M.M. Lucas, J.M. Porleno // Plant Physiol. – 1988. – Vol. 132, N 1. – P. 67 – 73. 231. French, E.R., The role of Fusarium in the root rot complex of soybean in Minnesota / E.R. French, B.W. Kennedy // Plant Dis. Rep. – 1963. – Vol. 47, N 7. – P. 672 – 676. 232. Grzelinska, A. Peroxidase isoenzymes in fusarium-infected tomato plants / A. Grzelinska // Phytopathology. – 1970. – Vol. 69, N 3. – P. 212 – 222. 141 233. Hagima, J. Acid phosphatase genetic variants in grains of some Triticum aestivum L. cultivars / J. Hagima, N.N. Saulescu // Rev. roum. biochim. – 1982. – Vol. 19, N 3. – P. 209 – 214. 234. Hedjaroude, G.A. Rot of soybean in Iran and methods of its control / G.A. Hedjaroude // Iranian Plant Pathology. – 1973. – Vol. 9, N 1. – P. 1 – 2. 235. Hemmi, T. Studies of septorioses of plants. VI. Septorioria glycines Hemmi causing the brown spot diseases of soybeans / T. Hemmi // Mem. Coll. Agris. – 1940. – N 47. – 14 p. 236. Henis, Y. Factors affecting suppressiveness to Rhizoctonia solani in soil / Y. Henis, A. Ghaffar, R. Baker // Phytopathology. – 1979. – Vol. 69, N 11. – P. 1164 – 1169. 237. Hildebrand, A.A. Soybean diseases in Ontario and effectiveness of seed treatment / A.A. Hildebrand, L.W. Koch // Phytopathology. – 1974. – Vol. 37, N 2. – P. 111 – 124. 238. Hippeli, S. Activated oxygen and free oxygen radicals in pathology: New insights and analogies between animals and plants / S. Hippeli, I. Heiser, E.F. Elstner // Plant Physiol. Biochem. – 1999. – Vol. 37. – P. 167 – 178. 239. Ho, H.H. Peroxidase isoenzymes of soybean roots from varieties resistant and susceptible to Phytophthora megasperma var. sojae / H.H. Ho, E. Waever // Phytopathology. – 1970. – Vol. 27, N 2. – P. 163 – 167. 240. Hollander, V.P. Acid phosphatases. Enzymes / V.P. Hollander // Plant Physiol. – 1971. – Vol. 4, N 3. – P. 450 – 472. 241. Jones, T.W. Enzyme changes in roots of healthy and phyllody-infected white clover Trifollum repens L. / T.W. Jones, J.L. Stoddort // Physiol. Plant Pathol. – 1971. – Vol. 1, N 4. – P. 385 – 396. 242. Jonson, H.W., Soybean diseases and their control / H.W. Jonson, B. Kochler // Fmrs. Bull. U. S. Dep. Agr. – 1943. – N 1937. – P 24. 243. Kawano, T. Role of the Reactive Oxygen Species-Generating Peroxidase Reactions in Plant Defense and Growth Induction / T. Kawano // Plant Cell Rep. – 2003. – Vol. 21. – P. 829 – 837. 142 244. Kawashima, N. Occurence of peroxidases in sweet potato infected by the black rot fungus / N. Kawashima, H. Hyodo, I. Uritani // Arg. Biol. Chem. – 1963. – Vol. 27, N 6. – P. 409 – 417. 245. Kawashima, N. Investigation of antigenic components produced by sweet potato in response to black rot infection /N. Kawashima, H. Hyodo, I. Uritani // Phytopathology. – 1964. – Vol. 54, N 9. – P. 1086 – 1092. 246. Kazan, K. Expression of a pathogenesis related peroxidase of Stylosanthes humilis in transgenic tobacco and canola and its effect on disease development / K. Kazan, K.C. Goulter, H.M. Way, J.M. Manners // Plant Sci. – 1998. – Vol. 136. – P. 207 – 217. 247. Khan, M.A. Phosphatases activity in the eggs of Dysdercus cingulatus related with embryonic development / M.A. Khan // J. Entomol. Res. – 1978. – Vol. 2, N 2. – P. 211 – 213. 248. Khatri, H.L. Metabolism of resistant and susceptible cowpea varieties infected with cowpea virus. 2. Changes in the peroxidase and catalase enzyme activity / H.L. Khatri, V.V. Chenulu // Ind. Phytopathol. – 1970. – Vol. 23, N 3. – P. 553 – 557. 249. König, D. Stoffwechselphysiologische Veränderung in der Pflanze nach Virusinfektion unter Einfluss von Wundreiz. Polyphenoloxydase-, Perohydase- und Cytochromoxydaseaktivität / D. König, F. Nienhaus // Phytopathol. – 1970. – Vol. 68(3). – P. 193 – 205. 250. Kurata, H. Studies of the fungal diseases of soybean in Japan / H. Kurata // Bull. Natl. Inst. Agric. Sci. Japan. – 1960. – Vol. 12. – P. 127. 251. Lagrimini, M.L. Characterization of antisense – transformed plant deficient in the tobacco anionic peroxidase / M.L. Lagrimini, V. Gingas; T. Finger et al. // Plant Physiol. – 1997. – Vol. 114. – P. 1187 – 1196. 252. Lazar, G. Patterns of enzyme changes during leaf senescence / G. Lazar, G. Farkas // Acta Biol. Acad. Sci. Hung. – 1970. – Vol. 21, N 4. – Р. 389 – 396. 253. Lee, S.C. Induction of Some Defense-related Genes and Oxidative Burst Is Required for the Establishment of Systemic Acquired resistance in Capsicum annuum / S.C. Lee, B.K. Hwang // Planta. – 2005. – Vol. 221. – P. 790 – 800. 143 254. Lim, S.M. Disease severity gradient of soybean downy mildew from a small focus of infection / S.M. Lim // Phytopathology. – 1978. – Vol. 68. – P. 1774 – 1778. 255. Lim, S.M. Brown – spot severity and yield reduction in soybean / S.M. Lim // Phytopathology. – 1980. – Vol. 70, N 10. – P. 974 – 977. 256. Liman, F. Phytohormone regulation of isoperoxidases in catharanthus roseus suspension cultures / F. Liman, K. Chahed, N. Ouelhazi et al. // Phytochemistry. – 1998. – Vol. 49. – P. 1219 – 1225. 257. Liu, S.T. Seed borne diseases of soybean // Bot. Bull. Acad. Sinica. – 1948. – Vol. 2, N 1. – P. 69 – 80. 258. Lowry, O.H. Protein measurement with the Folin phenol reagent / O.H. Lowry et al. // J. Biol. Chem. – 1951. – Vol. 193, N 1. – P. 265 – 275. 259. Macneil, B.H. Histological study of host – parasite relationships between Septorioria glycines Hemmi and soybean leaves and leaves and pods / B.H. Macneil, H. Zalasky // Canad. J. Botany. – 1957. – Vol. 35, 4. – P.501 – 505. 260. Mader, M. De novo Synthesis and Release of Peroxidases in Cell Suspension Cultures of Nicotiana tabacum L. / M. Mader, C. Walter // Planta, 1986. – Vol. 169. – P. 273 – 277. 261. Marcinka, K. Changes in peroxidase isozymes in bean ieaves following infection with red clover mottle virus / K. Marcinka // Biologia (CŠSR). – 1970. – Vol. 25, N 9. – P. 599 – 602. 262. Mckenzie, J.R. The effect of temperature and lesion size on the sporulation of Peronospora manshurica / J.R. Mckenzie, T.D. Wyllie // Phytopathology. – 1971. – Vol. 71. – P. 321 – 326. 263. Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance / R. Mittler // Trends Plant Sci. – 2002. – Vol. 7. – P. 405 – 410. 264. Mullens, T.R. Factors affecting the development of brown sport of soybeans in the field / T.R. Mullens, J. W. Hilty // Tennessee Farm. Home. Sci. Progr. Rep. Knoxville, Tenn. – 1979. – Vol. 111. – P. 24 – 25. 265. Murakishi, H.H. Purple seed stain of soybean / H.H. Murakishi // Phytopathology. – 1951. – Vol. 41, N 4. – P. 229 – 234. 144 266. Naoi, N. Acid phosphatases of aspergillus saitoi: purification and properties / N. Naoi, B. Teruniko // Arel and biochem. – 1977. – V. 4, N 10. – Р. 1835. 267. Neill, S. Hydrogen Peroxide Signaling / S. Neill, R. Desikan, J. Hancock // Curr. Opin. Plant Biol. – 2002. – Vol. 5. – P. 388 – 395. 268. Novacky, A. Peroxidase isozymes in virus inlected plants / A. Novacky, R.E. Hempton // Phytopathology. – 1968. – Vol. 58, N 3. – P. 301 – 305. 269. Peterson, D.J. Effects of temperature and leaf wetness period on brown spot disease of soybean / D.J. Peterson // Plant Dis. – 1982. – Vol. 66. – P. 995 – 998. 270. Reeve, S. Root rot robs yields / S. Reeve // Soybean Digest. – 1983. –Vol. 43, N 4. – P. 34. 271. Riggle, J.N. Histopathology of leaf infection of susceptible and resistant soybeans by Peronospora manshurica / J.N. Riggle, J.M. Dunleavy // Phytopathology. – 1974. – Vol. 64. – P. 522 – 526. 272. Riggle, J.H. Peronospora manshurica on a nonhost and on resistant, susceptible and intezmediate soybeans / J.H. Riggle // Can. J. Bot. – 1977. – Vol. 55. – P. 153 – 157. 273. Ross, J.P. Predisposition of soybeans to Fusarium wilt by Heterodera glycines and meloidogyne incognita / J.P. Ross // Phytopathology. – 1965. – Vol. 55, N 3. – P. 361 – 364. 274. Ross, J.P. Effect of simulated dew and postinoculation moist periods on infection of soybean by Septoria glycines / J.P. Ross // Phytopathology. – 1982. – Vol. 72. – P. 236 – 238. 275. Sato, R. Occurrence of soybean root caused by Corynespora cassiicola (Berk. Et Curt.) Wei in Hokkaido / R. Sato, K. Kitazawa // Ann. Phyt. Soc. Japan. – 1980. – Vol. 46, N 2. – P. 193 – 199. 276. Schridhara, Y Alkaline and acid phosphatases of silkworm Bombyx mori L. / Y Schridhara, Y.V. Bhat // J. Insect Physiol. – 1963. – Vol. 9, N 5. – P. 693 – 701. 277. Seevers, P.M. Studies on wheat stem rust resistance controlled of the Sr. 6 locus. II. Peroxidase activities / P.M. Seevers, J.M. Daly // Phytopathology. –1970. – Vol. 60, N 11. – P. 1642 – 1647. 145 278. Seevers, P.M. The role of peroxidase isozymes in resistance to wheat stem rust disease / P.M. Seevers, J.M. Daly, F.F. Catedral // Plant Physiol. – 1971. – Vol. 48, N 3. – P. 353 – 360. 279. Sherwin, H.S. Discoloration of soybean seeds by the frog-eye fungus Cercospora sojina / H.S. Sherwin, K.W. Kreitlow // Phytopathology. – 1952. – Vol. 43, N 10. – P. 69. 280. Simons, T.J. Metabolic changes associated withsystemic induced resistance to tobacco mosaic virus in Samsun NN tobacco / T.J. Simons, A.F. Ross, A. Frank // Ibid. – 1971. – Vol. 61, N 3. – P. 293 – 300. 281. Sinclair J.B. Compendium of soybean diseases / J.B. Sinclair, M.C. Shurtleff // American Fhytopathological Society, St. Paut, Minnesota. – 1975. – 69 p. 282. Solymosy, Q. Changes in peroxidase-isozyme patterns by virus infection / Q. Solymosy et al. // Virology. – 1967. – Vol. 32, N 2. – P. 117 – 121. 283. Tachibana, H. Rhizoctonia solani root rot epidemic of soybeans in central Iowa 1967 / Н. Tachibana // Plant Dis. Rep. – 1968. – Vol. 52, N 8. – P. 613 – 614. 284. Tiffany, Z.H. Species of Colletotrichum from legumes / Z.H. Tiffany, J.C. Gilman // Mycologia. – 1954. – Vol. 46. – P. 52 – 75. 285. Torres, M.A. Reactive oxygen species signaling in response to pathogens / M.A. Torres, D.G. Jones et al. // Plant Physiol. – 2000. – Vol. 141. – P. 373 – 378. 286. Vegetti, G. Changes in phenylalanine-ammonia-liase, peroxidase during the development of local necrotic leaves infected with alfalfa mosaic virus / G. Vegetti, G.G. Conti, P. Pesci // Phytopathol. Ztschr. – 1975. – Vol. 84, N 2. – P. 153 – 171. 287. Verma, H.S. Rhizoctonia aerial blight of soybean / H.S. Verma, P.N. Thapliyal // Indian Phytopathology. – 1976. – Vol. 29, N 4. – P. 389 – 391. 288. Vir, S. Peroxidase activity associated with Ascochyta blight of gram (Cicer arietinum L.) / S. Vir, J.S. Grewal // Indian Phytopathology. – 1975. – Vol. 28, N 2. – P. 223 – 225. 289. Wolf, F.A. Report of division of plant pathology / F.A. Wolf, // Ann. Rept. N. Carol. Agr. Expt. Sta. – 1924. – Vol. 47. – P. 83 – 85. 146 290. Wolf, F.A. Diseases of soy-beans which occur both in North Carolina and the Orient / F.A. Wolf, S.G. Lehman // J. Agr. Res. – 1926. – Vol. 33. – P. 391 – 396. 291. Wong, W.C. Erwinia salicis in cricket bat willows; peroxidase, polyphenoloxidase, glucosidase, pectinolytic and cellulolytic enzyme activity in diseased wood / W.C. Wong, T.F. Preece // Physiol. Plant. Pathol. – 1978. – Vol. 12, N 3. – P. 333 – 347. 292. Wyllie, T.D. The effects of temperature and leaf age on the development of lesions caused by Peronospora manshurica on soybean / T.D. Wyllie, L.F. Williams // Phytopathology. – 1965. – Vol. 55. – P. 166 – 170. 293. Young, L.D. Brown spot development and yield response of soybean inoculated with Septoria glycines at various growth stages / L.D. Young, J.P. Ross // Phytopathology. – 1979. – Vol. 69. – P. 8 – 11. 147 Приложение 1 Распределение среднесуточных температур воздуха за вегетационный период в годы исследований, °С (ГМС Благовещенск) Показатели I декада II декада III декада Среднемесячная Среднее многолетнее I декада II декада III декада Среднемесячная Среднее многолетнее I декада II декада III декада Среднемесячная Среднее многолетнее I декада II декада III декада Среднемесячная Среднее многолетнее I декада II декада III декада Среднемесячная Среднее многолетнее I декада II декада III декада Среднемесячная Среднее многолетнее 2008 Май 7,6 13,9 12,0 11,2 10,0 Июнь 17,2 21,9 22,3 20,5 16,8 Июль 20,5 21,7 22,8 21,7 20,4 Август 19,8 18,4 18,3 18,8 17,9 Сентябрь 14,0 11,1 6,1 10,4 10,9 Октябрь 5,1 2,1 0,0 2,4 1,0 148 2009 2010 2011 11,4 13,4 16,7 13,8 10,0 9,1 14,0 17,5 13,5 10,0 9,9 10,6 14,5 11,7 10,0 14,0 16,5 16,4 15,6 16,8 19,1 20,5 25,9 21,8 16,8 15,3 19,1 19,0 17,8 16,8 19,6 18,4 20,7 19,6 20,4 21,7 21,0 20,6 21,1 20,4 20,9 22,8 24,0 22,6 20,4 21,4 18,8 15,1 18,4 17,9 17,5 17,7 18,2 17,8 17,9 22,7 16,7 19,0 19,5 17,9 14,7 9,5 8,2 10,8 10,9 15,4 13,6 4,2 11,1 10,9 12,3 10,8 8,0 10,4 10,9 6,5 2,6 -3,1 2,0 1,0 6,0 0,5 -2,2 1,4 1,0 6,5 4,3 0,5 3,8 1,0 Приложение 2 Распределение осадков за вегетационный период в годы исследований, мм (ГСМ Благовещенск) Показатели I декада II декада III декада Среднемесячная Среднемноголетнее I декада II декада III декада Среднемесячная Среднемноголетнее I декада II декада III декада Среднемесячная Среднемноголетнее I декада II декада III декада Среднемесячная Среднемноголетнее I декада II декада III декада Среднемесячная Среднемноголетнее I декада II декада III декада Среднемесячная Среднемноголетнее 2008 Май 15 1 39 55 44 Июнь 15 6 17 38 98 Июль 36 59 28 123 124 Август 82 0 31 113 117 Сентябрь 6 15 27 48 76 Октябрь 11 6 20 37 22 149 2009 2010 2011 4 9 39 52 44 5 14 21 40 44 24 19 48 91 44 87 71 48 206 98 13 26 1 40 98 56 18 9 83 98 36 64 28 128 124 90 121 56 267 124 22 7 42 71 124 53 34 15 102 117 90 52 35 177 117 24 45 23 92 117 26 18 1 45 76 1 19 5 25 76 7 15 4 26 76 5 0 4 9 22 4 9 3 16 22 0 4 1 5 22 Приложение 3 Пораженность сои корневой гнилью, % Год исследований 2009 2010 Даурия Гармония Соната балл поражения развитие болезни распространенность Ср. по годам развитие болезни развитие болезни распространенность развитие болезни Фенологическая фаза распространенность 2008 распространенность Сорт Первый тройчатый лист 50,0 11,0 60,0 15,0 35,0 7,0 48,3 11,0 1 Цветение 60,0 22,5 70,0 30,5 70,0 25,0 66,7 26,0 2 Бобообразование 67,5 49,0 90,0 53,0 87,5 50,0 81,7 50,6 4 Первый тройчатый лист 30,0 5,5 50,0 6,0 22,5 5,5 34,2 5,7 1 Цветение 40,0 8,0 60,0 9,0 55,0 8,5 51,7 8,5 1 Бобообразование 52,5 10,0 70,0 13,0 70,0 11,5 64,2 11,5 2 Первый тройчатый лист 40,0 6,5 55,0 7,0 22,5 9,0 39,2 7,5 1 Цветение 55,0 14,5 60,0 24,0 60,0 13,0 58,3 17,2 2 Бобообразование 60,0 23,5 85,0 33,0 70,0 23,5 71,7 26,7 3 НСР05 1,13 1,19 1,56 1,74 1,00 1,80 1,98 1,14 НСРа НСРв 0,65 0,65 0,69 0,69 0,90 0,90 1,04 1,04 0,58 0,58 1,03 1,03 0,58 0,58 0,65 0,65 НСРа – фаза развития; НСРв – сорт 150 Даурия Гармония Соната пораженные Ср. по годам контроль (здоровые) контроль (здоровые) пораженные контроль (здоровые) Фенологическая фаза контроль (здоровые) Сорт пораженные 2008 пораженные Год исследований 2009 2010 % к контролю Приложение 4 Удельная активность пероксидазы в корнях сои при поражении корневой гнилью, ед/мг белка х 10-2 Первый тройчатый лист 632 ± 27 423 ± 67 234 ± 8 157 ± 4 464 ± 63 311 ± 5 443±115 297±77 67 Цветение 665 ± 5 492 ± 6 561 ± 5 415 ± 3 807 ± 91 597 ± 7 678±71 502±52 74 1464±163 644 ± 121 441 ± 7 194 ± 7 1195 ± 30 526 ± 9 1033±306 454±134 44 555 ± 7 665 ± 8 424 ± 2 509 ± 5 346 ± 8 415 ± 10 442±60 530±72 120 Цветение 1630±211 2378±258 1104 ± 30 1662 ± 47 1418±107 2187 ± 50 1384±152 2076±214 150 Бобообразование 1437±122 2443±475 1434 ± 50 2438±211 1632 ± 47 2774±258 1501±65 2552±111 170 225 ± 2 290 ± 72 119 ± 2 153 ± 11 163 ± 4 210 ± 5 169±30 218±39 129 Цветение 1289±135 1096 ± 88 536 ± 12 456 ± 5 831 ± 14 706 ± 75 885±219 752±186 85 Бобообразование 1252 ± 83 712 ± 7 708 ± 11 559 ± 7 1559±202 947 ± 64 1173±248 739±112 63 Бобообразование Первый тройчатый лист Первый тройчатый лист НСР05 1,45 1,40 0,52 1,18 НСРа 0,59 0,57 0,21 0,48 НСРв 0,59 0,57 0,21 0,48 0,17 0,39 НСРс 0,48 0,47 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 151 Приложение 5 Удельная активность каталазы в корнях сои при поражении корневой гнилью, ед/мг белка х 10 -4 Даурия Гармония Соната пораженные (здоровые) пораженные (здоровые) контроль контроль Ср. по годам 2010 пораженные (здоровые) контроль фаза 2009 пораженные Фенологическая (здоровые) Сорт контроль 2008 % к контролю Год исследований Первый тройчатый лист 104±7 153±5 126±4 185±5 131±9 191±3 120±8 176±11 147 Цветение 74±2 98±2 93±3 121±2 108±5 140±1 92±9 120±12 130 Бобообразование 22±1 29±2 27±1 36±2 30±1 40±3 26±2 35±3 133 120±11 82±3 115±11 78±3 131±10 89±3 122±4 83±3 68 Цветение 80±1 75±5 64±1 59±5 78±2 73±5 74±5 69±5 93 Бобообразование 70±2 52±1 97±2 71±1 119±5 88±2 95±14 70±10 74 Первый тройчатый лист 127±7 150±5 162±3 191±12 154±9 183±4 148±10 175±12 118 Цветение 102±4 108±2 134±3 142±5 99±2 106±4 112±11 119±11 106 Бобообразование 58±2 56±1 80±2 78±2 69±2 67±1 69±6 67±6 97 Первый тройчатый лист НСР05 НСРа НСРв 1,12 0,46 0,46 3,7 1,53 1,53 НСРс 0,37 1,25 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 152 1,50 0,61 0,61 0,38 0,15 0,15 0,50 0,12 Даурия Гармония Соната пораженные контроль (здоровые) Ср. по годам пораженные пораженные контроль (здоровые) Фенологическая фаза пораженные Сорт контроль (здоровые) 2008 контроль (здоровые) Год исследований 2009 2010 % к контролю Приложение 6 Удельная активность кислой фосфатазы в корнях сои при поражении корневой гнилью, ед/мг белка х 10 -6 Первый тройчатый лист 60±5 43±3 205±12 180±9 80±2 63±1 115±45 95±42 83 Цветение 88±2 231±10 123±7 172±11 115±5 250±7 109±10 218±23 200 Бобообразование 40±1 62±3 68±5 106±5 103±7 162±3 70±18 110±28 157 Первый тройчатый лист 53±2 40±1 89±3 69±5 112±7 85±5 85±18 65±13 76 Цветение 120±3 121±7 210±11 218±10 173±11 160±10 168±26 166±28 99 Бобообразование 102±3 100±2 87±5 82±3 123±9 130±5 104±10 104±14 100 Первый тройчатый лист 55±1 42±2 90±2 16±1 80±2 63±2 75±10 40±13 53 Цветение 84±2 73±5 165±10 90±7 130±9 90±4 126±23 84±5 67 Бобообразование 62±1 60±2 98±3 76±1 122±5 99±2 94±17 78±11 83 НСР05 1,79 4,04 2,09 0,91 НСРа НСРв НСРс 0,36 0,36 0,30 0,017 0,017 0,013 0,85 0,85 0,70 0,37 0,37 0,30 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 153 Год исследований 2009 2010 Даурия Гармония Соната Первый тройчатый лист контроль (здоровые) пораженные Ср. по годам пораженные контроль (здоровые) пораженные контроль (здоровые) Фенологическая фаза пораженные Сорт контроль (здоровые) 2008 % к контролю Приложение 7 Удельная активность пероксидазы в листьях сои при поражении корневой гнилью, ед/мг белка х 10 -3 - - - - - - - - - Цветение 147 ± 3 134 ± 3 126 ± 19 117 ± 20 154 ± 5 97 ± 2 142±8 116±10 82 Бобообразование 282 ± 20 167 ± 10 105 ± 8 50 ± 5 224 ± 12 151 ± 7 204±52 122±36 60 - - - - - - - - - Цветение 147 ± 7 217 ± 3 116 ± 1 215 ± 4 140 ± 5 284 ± 20 134±9 239±22 178 Бобообразование 288 ± 15 328 ± 25 156 ± 6 294 ± 21 149 ± 10 277 ± 17 198±45 300±14 151 - - - - - - - - - Цветение 118 ± 7 96 ± 5 173 ± 7 102 ± 2 153 ± 5 114 ± 6 148±16 104±5 70 Бобообразование 145 ± 4 128 ± 9 242 ± 6 201 ± 10 160 ± 5 125 ± 8 182±30 158±24 87 Первый тройчатый лист Первый тройчатый лист НСР05 0,29 0,29 2,57 0,51 НСРа НСРв 0,11 0,14 0,11 0,14 1,05 1,28 0,20 0,25 1,05 0,20 НСРс 0,11 0,11 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 154 Год исследований 2009 2010 Даурия Гармония Соната Первый тройчатый лист контроль (здоровые) пораженные Ср. по годам пораженные контроль (здоровые) пораженные контроль (здоровые) Фенологическая фаза пораженные Сорт контроль (здоровые) 2008 % к контролю Приложение 8 Удельная активность каталазы в листьях сои при поражении корневой гнилью, ед/мг белка х 10-4 - - - - - - - - - Цветение 92 ± 3 66 ± 1 71 ± 2 59 ± 1 84 ± 1 54 ± 1 82±6 60±3 73 Бобообразование 145 ± 5 104 ± 3 106 ± 7 83 ± 2 112 ± 3 98 ± 1 121±12 95±6 78 - - - - - - - - - 75 ± 2 112 ± 3 88 ± 5 132 ± 5 90 ± 2 117 ± 2 84±4 120±6 143 218 ± 11 280 ± 8 137 ± 4 93 ± 2 186 ± 5 132 ± 5 180±23 168±56 93 - - - - - - - - - 81 ± 4 72 ± 2 46 ± 1 31 ± 1 97 ± 2 89 ± 1 75±15 64±17 85 185 ± 12 154 ± 2 128 ± 6 114 ± 2 135 ± 7 120 ± 5 149±17 129±12 87 Первый тройчатый лист Цветение Бобообразование Первый тройчатый лист Цветение Бобообразование НСР05 0,98 1,55 0,28 0,29 НСРа 0,40 0,63 0,11 0,11 НСРв НСРс 0,49 0,40 0,77 0,63 0,14 0,11 0,14 0,11 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 155 Год исследований 2009 2010 Даурия Гармония Соната Первый тройчатый лист контроль (здоровые) пораженные Ср. по годам пораженные контроль (здоровые) пораженные контроль (здоровые) Фенологическая фаза пораженные Сорт контроль (здоровые) 2008 % к контролю Приложение 9 Удельная активность кислой фосфатазы в листьях сои при поражении корневой гнилью, ед/мг белка х 10 -6 - - - - - - - - - Цветение 244 ± 2 157 ± 5 205 ± 2 181 ± 2 172 ± 17 123 ± 1 207±20 154±16 74 Бобообразование 305 ± 2 87 ± 1 356 ± 7 168 ± 5 246 ± 5 102 ± 1 302±31 119±24 39 - - - - - - - - - Цветение 158 ± 5 79 ± 1 136 ± 5 105 ± 2 200 ± 8 167 ± 1 165±18 117±26 71 Бобообразование 430 ± 7 327 ± 5 316 ± 7 259 ± 2 424 ± 3 314 ± 2 390±37 300±20 77 - - - - - - - - - Цветение 100 ± 8 83 ± 2 153 ± 2 118 ± 3 100 ± 5 67 ± 1 118±17 89±15 75 Бобообразование 287 ± 7 198 ± 5 320 ± 2 206 ± 2 368 ± 5 245 ± 4 325±23 216±14 66 Первый тройчатый лист Первый тройчатый лист НСР05 1,79 1,70 1,68 1,68 НСРа 0,73 0,69 0,68 0,68 НСРв 0,89 0,85 0,84 0,84 0,68 0,68 НСРс 0,73 0,69 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 156 Приложение 10 Ср. по годам распростран енность развитие болезни распростран енность развитие болезни 10,0 8,0 38,4 7,8 32,5 6,5 0 0 20,2 5,6 1 100,0 100,0 0 46,4 82,3 0 90,0 100,0 25,5 24,4 56,0 5,0 50,6 90,0 25,0 15,0 46,0 5,3 77,5 100,0 0 26,2 58,5 0 79,5 97,5 12,6 28,0 60,7 2,6 3 4 1 65,0 90,0 20,5 56,0 60,5 70,0 16,2 36,6 31,7 60,0 10,0 16,0 47,5 78,0 11,0 29,5 51,2 74,5 14,4 34,5 2 3 развитие болезни развитие болезни 2011 распростран енность 2009 развитие болезни Третий тройчатый лист Цветение Бобообразование Третий тройчатый лист Цветение Бобообразование 2008 Год исследований 2010 распростран енность Фенологическая фаза распростран енность Гармония Соната Сорт (форма) балл поражения Пораженность сои септориозом, % 0 0 30,5 5,5 22,5 4,5 0 0 13,2 2,5 1 75,0 34,2 70,0 20,0 55,0 12,5 40,0 11,5 60,0 19,5 2 Бобообразование 100,0 62,5 85,0 44,3 75,5 26,5 80,0 40,0 85,1 43,3 3 Третий тройчатый лист 12,5 Цветение 30,0 Бобообразование 80,5 НСР05 1,66 НСРа 0,33 НСРв 0,38 НСРа – фаза развития; НСРв – сорт 5,2 5,8 48,2 1,12 0,56 0,65 0 53,3 60,0 0,68 0,34 0,39 0 18,0 30,7 0,51 0,26 0,30 0 22,9 46,2 0,28 0,14 0,16 0 3,0 10,0 0,49 0,24 0,28 0 30,4 60,0 0,84 0,42 0,48 0 10,7 36,0 0,84 0,42 0,48 3,1 34,1 61,7 0,28 0,14 0,16 1,3 9,4 31,2 0,27 0,13 0,15 1 1 3 КА 1344 Даурия Третий тройчатый лист Цветение 157 Приложение 11 Ср. по годам 2011 развитие болезни распространен ность развитие болезни распространен ность развитие болезни 20,0 7,0 23,5 8,0 15,5 6,8 0 0 14,7 5,45 1 60,0 82,5 15,0 30,0 44,5 5,0 70,0 100,0 22,0 30,0 56,2 7,0 60,0 91,0 0 17,5 39,3 0 68,0 95,5 0 22,1 48,5 0 64,5 92,2 9,2 24,9 47,1 3,0 2 3 1 31,0 70,0 14,2 30,5 42,5 98,0 36,1 50,0 30,5 78,0 12,1 25,5 53,5 87,5 20,6 33,5 39,4 83,4 20,7 34,9 2 3 Пятый тройчатый лист Цветение Бобообразование 0 0 24,0 7,2 10,0 6,3 17,5 6,5 12,9 5,0 1 55,0 97,5 34,2 48,5 65,0 100,0 44,3 64,5 55,0 92,8 20,0 44,2 60,5 97,8 27,2 54,5 58,9 97,0 31,4 52,9 3 4 0 0 0 0 0 0 0 0 0 0 5,3 12,5 0,75 0,37 0,43 18,5 20,0 0,49 0,24 0,28 3,2 14,5 1,06 0,53 0,61 0 19,5 0,77 0,38 0,44 0 8,0 0,77 0,38 0,44 0 22,0 0,84 0,42 0,48 0 9,0 0,84 0,42 0,48 8,8 20,4 0,49 0,24 0,28 2,1 11,0 0,49 0,24 0,28 1 1 158 развитие болезни распространен ность Пятый тройчатый лист Цветение Бобообразование Пятый тройчатый лист Цветение Бобообразование Пятый 0 тройчатый лист Цветение 16,7 Бобообразование 20,0 НСР05 0,66 НСРа 0,33 НСРв 0,38 НСРа – фаза развития; НСРв – сорт КА 1344 2009 развитие болезни 2008 Год исследований 2010 распространен ность Фенологическая фаза распространен ность Даурия Гармония Соната Сорт (форма) балл поражения Пораженность сои пероноспорозом, % Приложение 12 КА 1344 Даурия Гармония Соната развитие болезни развитие болезни распростран енность Ср. по годам 2011 распростран енность развитие болезни развитие болезни распростран енность Год исследований 2010 2009 распростран енность 2008 развитие болезни Фенологическая фаза распростран енность Сорт (форма) балл поражения Пораженность сои бактериозом, % Третий тройчатый лист Цветение Бобообразование Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 32,6 65 - 14,6 46 - - - - - 32,6 65 - 14,6 46 - 2 3 - - - 18 40 11,2 26 - - - - 18 40 11,2 26 2 3 Третий тройчатый лист Цветение Бобообразование Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 22,5 50 - 12,5 44 - - - - - 22,5 50 - 12,5 44 - 2 3 - - - 30,6 60 13,7 32 - - - - 30,6 60 13,7 32 2 3 1,58 0,79 1,12 1,83 0,91 1,29 1,58 0,79 1,12 1,83 0,91 1,29 НСР05 НСРа НСРв НСРа – фаза развития; НСРв – сорт 159 пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) Третий тройчатый лист Цветение Бобообразование Ср. по годам 2011 158 ±10 134 ±14 125 ±13 326 ±14 211 ±14 342 ±20 305 ±25 243±52 200±39 347 ±23 1410±20 339 ±13 745±10 326± 19 523 ±28 419± 22 819±15 354 ±15 1118±12 306 ±13 860±10 419 ±17 893±28 571 ±23 1585±17 361±20 986±187 409±59 1002±195 102 273±14 1092±28 119 ±14 713±15 268 ±15 938±23 247 ±16 988±20 227±36 933±79 411 347 ±17 1438±15 694 ±15 4403±25 216 ±10 779 ± 64 324 ±14 3387±214 340 ±15 745±10 748±12 4296±27 420 ±15 1902±15 630 ±28 3687±17 331±42 1216 599±94 3943 181 324 Третий тройчатый лист Цветение Бобообразование 184 ±12 119±11 164 ±15 112±10 203±12 148±12 254±11 127±13 201±19 126±7 63 318±17 723±14 343±15 908±12 373±17 1209±26 362±12 1829±20 453±15 800±25 380±17 986±28 373±16 1296±15 369±13 2576±23 379±27 1007±143 363±7 1575±393 96 156 Третий тройчатый лист Цветение Бобообразование 363±12 655±15 - - - - - - 363±12 655±15 180 412±11 1108±39 614±16 4102±2 537±33 1296±15 972±27 7547±30 473±12 1357±29 1021±13 3703±2 524±21 987±15 1120±29 2654±28 487±28 1187±85 932±110 4501±1060 191 379 Гармония 172 ±11 Даурия Фенологическая фаза 2009 КА 1344 Сорт (форма) Соната 2008 Год исследований 2010 % к контролю Приложение 13 Удельная активность пероксидазы в листьях сои при поражении септориозом, ед/мг белка х 10-3 Третий тройчатый лист Цветение Бобообразование НСР05 НСРа НСРв НСРс 1,11 0,39 0,45 0,32 1,0 0,35 0,40 0,28 1,11 0,39 0,45 0,32 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 160 1,96 0,69 0,80 0,56 1,27 0,45 0,52 0,36 Приложение 14 Гармония Соната Третий тройчатый лист Цветение Бобообразование Третий тройчатый лист Цветение Бобообразование пораженные пораженные контроль (здоровые) Ср. по годам 2011 контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) Фенологическая фаза 2009 пораженные Сорт (форма) контроль (здоровые) 2008 Год исследований 2010 % к контролю Удельная активность каталазы в листьях сои при поражении септориозом, ед/мг белка х 10 -4 68±1 63±2 81±2 48±1 66±2 52±1 60±2 38±1 69±4 50±5 73 92±3 145±5 33±2 80±2 62±12 11±1 71±2 106±7 35±1 44±1 40±1 20±3 84±1 112±3 43±2 28±1 56±2 21±2 80±2 123±7 32±2 74±3 60±3 18±1 82±4 121±8 36±2 56±12 54±4 17±2 68 45 47 75±2 196±11 33±2 59±2 88±5 115±4 59±3 46±1 90±2 164±5 35±1 62±2 65±4 178±8 42±1 80±3 79±5 163±17 42±5 62±7 53 38 32±6 17±3 53 74±10 138±16 23±2 47±8 44±2 10±1 63 32 43 75±5 41±8 226±27 78±8 1,51 0,53 0,61 0,43 55 34 КА 1344 Даурия Третий 46±2 21±2 18±1 12±1 23±2 11±2 42±2 23±1 тройчатый лист Цветение 81±4 60±1 46±1 30±3 97±2 64±1 71±5 34±4 Бобообразование 185±12 43±5 128±6 52±3 135±7 41±2 106±7 41±2 Третий 23±2 10±1 тройчатый лист Цветение 70±5 46±3 91±2 61±2 65±4 24±2 75±1 33±2 Бобообразование 287±5 70±2 156±3 101±3 217±6 81±2 244±8 61±2 НСР05 1,71 1,58 1,53 1,54 НСРа 0,60 0,56 0,54 0,54 НСРв 0,69 0,64 0,62 0,62 НСРс 0,49 0,45 0,44 0,44 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 161 пораженные пораженные 62±2 24±1 65±1 40±7 82±3 48±1 70±4 35±5 50 244±2 305±2 100±5 162±1 205±2 356±7 80±3 120±3 172±17 246±5 120±6 125±2 264±7 432±24 75±3 158±2 219±20 334±39 94±10 141±10 43 42 61±2 33±1 85±3 77±1 35±5 35±1 112±2 70±3 73±16 54±11 74 158±5 430±27 125±2 259±2 136±5 316±7 113±3 259±5 200±8 424±13 132±4 308±15 220±8 365±3 61±1 217±2 178±19 384±26 107±16 261±18 60 68 87±2 85±1 46±1 24±1 34±1 20±1 80±2 36±3 62±12 41±14 66 100±8 287±17 92±7 134±2 153±2 320±2 85±3 236±9 100±5 368±5 81±5 143±7 172±3 279±3 168±7 200±5 131±18 313±20 106±20 178±24 81 57 90±1 60±2 - - - - - - 90±1 60±2 67 134±7 265±5 105±3 227±8 150±3 290±5 141±2 287±2 132±1 308±5 127±8 279±3 140±3 195±3 134±5 76±2 139±4 264±24 127±7 217±48 91 82 Даурия Третий тройчатый лист Цветение Бобообразование КА 1344 Третий тройчатый лист Цветение Бобообразование Третий тройчатый лист Цветение Бобообразование НСР05 НСРа НСРв НСРс 0,66 0,23 0,27 0,19 1,26 0,45 0,52 0,36 1,45 0,51 0,59 0,42 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 162 1,35 0,48 0,55 0,39 контроль (здоровые) 30±1 контроль (здоровые) 70±1 контроль (здоровые) Третий тройчатый лист Цветение Бобообразование контроль (здоровые) Фенологическая фаза Гармония Сорт (форма) контроль (здоровые) пораженные Ср. по годам 2011 Соната пораженные 2009 пораженные 2008 Год исследований 2010 % к контролю Приложение 15 Удельная активность кислой фосфатазы в листьях сои при поражении септориозом, ед/мг белка х 10-6 1,63 0,57 0,66 0,47 Даурия Третий тройчатый лист Цветение Бобообразование КА 1344 Третий тройчатый лист Цветение Бобообразование Третий тройчатый лист Цветение Бобообразование НСР05 НСРа НСРв НСРс контроль (здоровые) пораженные пораженные Ср. по годам 2011 контроль (здоровые) пораженные контроль (здоровые) контроль (здоровые) пораженные 2009 пораженные Третий тройчатый лист Цветение Бобообразование контроль (здоровые) Фенологическ ая фаза Гармония Сорт (форма) Соната 2008 Год исследований 2010 - - - - - - - - - - 347± 13 1410±20 486±12 1833±15 326±19 523±18 423±12 941±18 354±15 1118±12 396±14 1342±19 419±17 893±28 568±13 1250±21 361±20 986±187 468±38 1341±184 - - - - - - - - - - 347±17 1438±15 556±13 2013±20 216±10 779 ±64 314±12 1246±12 340±15 745±10 650±12 1175±28 420±15 1902±15 830±13 3804±27 331±42 1216±278 587±107 2059±611 - - - - - - - - - - 318±17 723±14 423±12 670±18 373±17 1209±26 419±12 1450±100 453±15 800±25 589±14 896±27 373±16 1296±35 318±15 1426±28 379±27 1007±143 437±56 1112±194 - - - - - - - - - - 412±11 1108±29 556±13 2216±35 537±33 1296±15 805±26 2338±32 1357±19 2984±28 987±15 1974±25 474±62 1187±84 680±124 2378±215 1,03 0,36 0,51 0,36 1,36 0,48 0,68 0,48 1,11 0,39 0,55 0,39 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 163 0,59 0,21 0,29 0,21 1,42 0,50 0,71 0,50 % к контролю Приложение 16 Удельная активность пероксидазы в листьях сои при поражении пероноспорозом, ед/мг белка х 10 -3 130 136 177 169 115 110 143 200 пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) Третий тройчатый лист Цветение Бобообразование Ср. по годам 2011 - - - - - - - - - - - 92±3 145±15 82±7 57±3 71±2 106±17 64±2 26±5 84±1 112±3 70±3 40±1 80±2 123±17 60±2 63±4 82±4 121±8 69±4 46±8 84 38 - - - - - - - - - - - 75±2 196±11 37±2 143±6 88±5 115±4 59±3 78±3 90±2 164±5 41±2 126±9 65±4 178±8 38±2 132±7 79±5 163±17 44±5 120±14 56 74 Третий тройчатый лист Цветение Бобообразование 81±4 185±12 91±2 14±2 46±1 128±6 43±1 21±1 97±2 135±7 65±3 23±2 71±5 106±7 92±3 42±1 74±10 138±16 73±11 25±5 99 18 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - 70±5 287±5 47±3 240±5 91±2 156±3 66±3 149±13 217±16 123±12 244±18 175±12 80±10 226±27 56±9 172±25 70 76 Гармония Третий тройчатый лист Цветение Бобообразование Даурия Фенологическая фаза 2009 КА 1344 Сорт (форма) Соната 2008 Год исследований 2010 % к контролю Приложение 17 Удельная активность каталазы в листьях сои при поражении пероноспорозом, ед/мг белка х 10 -4 НСР05 НСРа НСРв НСРс 1,67 0,59 0,83 0,59 1,67 0,59 0,83 0,59 1,59 0,56 0,80 0,56 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 164 1,40 0,49 0,70 0,49 1,56 0,55 0,78 0,55 Даурия Третий тройчатый лист Цветение Бобообразование КА 1344 Третий тройчатый лист Цветение Бобообразование Третий тройчатый лист Цветение Бобообразование НСР05 НСРа НСРв НСРс контроль (здоровые) пораженные пораженные Ср. по годам 2011 контроль (здоровые) пораженные контроль (здоровые) контроль (здоровые) пораженные 2009 пораженные Третий тройчатый лист Цветение Бобообразование контроль (здоровые) Фенологическая фаза Гармония Сорт (форма) Соната 2008 Год исследований 2010 % к контролю Приложение 18 Удельная активность кислой фосфатазы в листьях сои при поражении пероноспорозом, ед/мг белка х 10 -6 - - - - - - - - - - - 244±2 305±2 152±5 175±2 205±2 356±7 136±1 193±1 172±17 246±5 102±2 144±2 264±7 432±4 176±3 255±3 219±20 334±39 141±15 200±23 64 60 - - - - - - - - - - - 158±5 430±7 126±3 307±5 136±5 316±7 104±2 263±3 200±8 424±3 163±1 420±3 220±8 365±3 183±2 286±2 178±19 384±26 144±17 319±34 81 83 - - - - - - - - - - - 100±8 287±7 83±1 93±5 153±2 320±12 102±2 204±8 100±5 368±15 78±1 230±6 172±3 279±3 132±3 175±5 131±18 313±20 99±12 175±29 76 56 - - - - - - - - - 134±7 265±5 116±2 248±6 150±3 290±5 146±2 259±3 308±5 280±5 195±3 165±2 142±8 264±24 131±15 238±25 92 90 1,67 0,59 0,83 0,59 1,63 0,57 0,81 0,57 1,32 0,46 0,66 0,46 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 165 1,43 0,50 0,72 0,50 1,66 0,58 0,83 0,58 % к контролю пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные Фенологическая фаза Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 326±33 523±18 489±22 575±24 - - - - 326±33 523±18 489±22 575±24 150 110 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 216±11 779 ±64 259±13 1013±20 - - - - 216±11 779±64 259±13 1013±20 120 130 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 373±17 1209±26 410±16 2007±11 - - - - 373±17 1209±26 410±16 2007±11 110 166 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 537±33 1296±15 1235±27 2726±35 - - - - 537±33 1296±15 1235±27 2726±35 230 210 КА 1344 Даурия Гармония Сорт (форма) Соната контроль (здоровые) Приложение 19 Удельная активность пероксидазы в листьях сои при поражении бактериозом, ед/мг белка х10-3 Год исследований 2008 2009 2010 2011 Ср. по годам НСР05 НСРа НСРв НСРс 1,39 0,49 0,69 0,49 1,39 0,49 0,69 0,49 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 166 Приложение 20 пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные Фенологическая фаза Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 71±2 106±7 192±11 81±4 - - - - 71±2 106±7 192±11 81±4 270 76 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - 88±5 115±4 59±2 64±2 - - - - 88±5 115±4 59±2 64±2 67 56 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 46±1 128±6 209±15 107±9 - - - - 46±1 128±6 209±15 107±9 454 84 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 91±2 156±3 41±1 13±1 - - - - 91±2 156±3 41±1 13±1 45 8 Гармония Сорт (форма) Соната контроль (здоровые) пораженные Ср. по годам 2011 Даурия контроль (здоровые) пораженные 2009 КА 1344 контроль (здоровые) 2008 Год исследований 2010 % к контролю Удельная активность каталазы в листьях сои, пораженных бактериозом, ед/мг белка х 10 -4 НСР05 НСРа НСРв НСРс 1,67 0,59 0,83 0,59 1,67 0,59 0,83 0,59 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 167 пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные контроль (здоровые) пораженные Ср. по годам 2011 - - - - - - - - - - - - - 205±2 356±7 128±2 225±4 - - - - 205±2 356±7 128±2 225±4 62 63 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 136±5 316±7 113±1 204±5 - - - - 136±5 316±7 113±1 204±5 83 64 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 153±2 320±2 96±2 144±5 - - - - 153±2 320±2 96±2 144±5 63 45 Третий тройчатый лист Цветение Бобообразование - - - - - - - - - - - - - 150±3 290±5 107±1 238±7 - - - - 150±3 290±5 107±1 238±7 71 82 Гармония Соната Третий тройчатый лист Цветение Бобообразование Даурия Фенологическая фаза 2009 КА 1344 Сорт (форма) контроль (здоровые) 2008 Год исследований 2010 % к контролю Приложение 21 Удельная активность кислой фосфатазы в листьях сои, пораженных бактериозом, ед/мг белка х 10-6 НСР05 НСРа НСРв НСРс 1,63 0,57 0,81 0,57 1,63 0,57 0,81 0,57 НСРа – фаза развития; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 168 Приложение 22 Удельная активность пероксидазы в листьях сои при искусственном заражении септориозом, ед/мг белка х 10 -3 Гармония пораженные (здоровые) пораженные (здоровые) контроль контроль Ср. по годам 2010 пораженные (здоровые) контроль инфицирования 2009 пораженные (форма) (здоровые) Время после Соната Сорт контроль 2008 % к контролю Год исследований 1 сутки 38±2 44±2 68±1 78±3 128±8 147±8 78±26 90±30 115 3 сутки 72±1 108±10 81±3 121±5 207±15 310±8 120±43 180±65 150 4 сутки 144±6 186±8 120±10 155±8 156±6 201±10 140±10 181±13 129 7 сутки 173±5 123±9 145±4 103±5 201±10 143±10 173±16 123±11 71 9 сутки 173±8 1038±30 160±5 960±15 201±12 1206±25 178±12 1068±72 600 12 сутки 316±20 310±9 205±5 201±8 259±18 254±10 260±32 255±31 98 1 сутки 58±1 73±2 86±2 108±5 63±2 79±1 69±8 87±10 126 3 сутки 75±2 235±8 113±6 354±10 262±12 820±26 150±57 469±178 313 4 сутки 166±12 264±8 124±5 197±8 169±5 269±10 153±14 243±23 159 7 сутки 175±9 136±5 194±5 151±4 171±10 133±12 180±7 140±5 78 9 сутки 192±6 639±15 177±10 589±12 156±4 519±5 175±10 583±34 333 12 сутки 359±8 413±21 346±18 398±15 285±10 328±15 330±22 379±26 115 169 КА 1344 Даурия Продолжение приложения 22 1 сутки 46±1 53±2 66±2 77±3 38±1 44±1 50±8 58±9 116 3 сутки 86±2 245±5 90±2 256±10 85±2 242±8 87±1 248±4 285 4 сутки 119±12 118±5 133±8 132±6 123±10 122±8 125±4 124±4 99 7 сутки 225±13 193±8 105±2 90±3 129±10 111±5 153±36 132±31 86 9 сутки 244±10 549±20 112±5 252±8 148±5 333±12 168±39 378±88 225 12 сутки 329±18 296±6 273±10 246±5 298±8 268±10 300±16 270±14 90 1 сутки 76±2 106±2 72±1 101±3 110±2 154±5 86±12 120±16 140 3 сутки 108±2 463±18 87±1 373±15 78±2 335±17 91±8 390±37 429 4 сутки 103±8 60±2 110±8 64±2 111±5 64±1 108±2 63±1 58 7 сутки 186±10 167±8 170±6 153±5 268±10 241±8 208±30 187±27 90 9 сутки 232±6 334±12 250±10 360±8 286±5 412±20 256±15 369±22 144 12 сутки 405±12 498±18 456±15 561±26 465±9 572±12 442±18 544±23 123 НСР05 1,27 1,63 1,62 1,55 НСРа 0,45 0,57 0,57 0,55 НСРв 0,26 0,43 0,43 0,41 НСРс 0,18 0,30 0,30 0,29 НСРа – время после инфицирования; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 170 Гармония Соната пораженные контроль (здоровые) Ср. по годам пораженные пораженные контроль (здоровые) Время после инфицирования пораженные Сорт (форма) контроль (здоровые) 2008 контроль (здоровые) Год исследований 2009 2010 % к контролю Приложение 23 Удельная активность каталазы в листьях сои при искусственном заражении септориозом, ед/мг белка х 10-4 1 сутки 76±1 91±3 80±2 92±7 72±2 83±5 76±2 89±5 117 3 сутки 213±10 149±26 96±2 67±5 189±8 132±10 166±35 116±10 70 4 сутки 84±2 65±1 60±1 47±1 114±5 89±3 86±15 67±12 78 7 сутки 81±2 38±1 79±2 37±1 80±1 38±1 80±0,5 38±0,3 47 9 сутки 61±1 27±1 102±5 45±1 113±5 50±1 92±15 40±6 44 12 сутки 48±3 34±3 28±2 20±1 24±2 17±1 33±7 23±5 70 1 сутки 69±1 105±5 70±1 106±3 143±10 217±12 94±24 143±37 152 3 сутки 191±8 214±10 145±5 162±5 234±12 262±12 190±25 213±28 112 4 сутки 45±1 31±1 46±1 32±1 62±2 43±1 51±5 36±3 70 7 сутки 43±1 52±1 30±1 37±1 47±1 57±2 40±5 49±6 122 9 сутки 42±1 46±1 46±1 42±1 65±2 65±1 51±7 51±7 100 12 сутки 39±5 16±1 33±3 13±1 40±8 16±1 37±1 15±1 40 171 КА 1344 Даурия Продолжение приложения 23 1 сутки 106±3 138±5 60±1 78±2 71±1 92±1 79±13 103±18 130 3 сутки 85±1 93±2 125±5 137±3 216±10 238±10 142±38 156±42 110 4 сутки 65±1 31±1 30±1 14±1 124±5 59±2 73±27 35±1 48 7 сутки 32±1 43±1 32±1 43±1 116±8 155±3 60±28 80±37 134 9 сутки 43±1 49±1 47±1 54±1 51±1 59±1 47±2 54±2 115 12 сутки 36±5 21±2 33±2 20±1 34±2 20±1 34±0,8 20±0,3 60 1 сутки 129±12 258±10 91±2 195±3 119±2 273±5 113±11 242±12 214 3 сутки 64±2 87±2 81±1 110±3 113±5 154±2 86±14 117±19 136 4 сутки 61±1 79±1 51±1 66±1 50±1 65±1 54±3 70±4 130 7 сутки 40±1 43±1 43±1 46±1 55±1 59±1 46±4 49±4 107 9 сутки 56±1 65±2 46±1 53±1 93±2 108±2 65±14 76±16 116 12 сутки 35±5 12±1 29±1 10±1 31±2 11±1 32± 11±0,5 35 НСР05 1,60 1,45 1,26 1,27 НСРа 0,56 0,51 0,44 0,45 НСРв 0,43 0,39 0,33 0,34 НСРс 0,30 0,27 0,23 0,24 НСРа – время после инфицирования; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 172 Приложение 24 Удельная активность кислой фосфатазы в листьях сои при искусственном заражении септориозом, ед/мг белка х 10 -6 Гармония пораженные (здоровые) пораженные (здоровые) контроль контроль Ср. по годам 2010 пораженные (здоровые) контроль инфицирования 2009 пораженные (форма) (здоровые) Время после Соната Сорт контроль 2008 % к контролю Год исследований 1 сутки 104±3 142±5 75±2 103±2 70±1 96±1 83±10 114±14 137 3 сутки 159±5 103±2 126±8 82±2 120±3 78±1 135±12 88±7 65 4 сутки 75±1 28±1 105±3 39±1 216±12 80±2 132±42 49±15 37 7 сутки 79±1 53±1 80±2 54±1 90±1 60±1 83±3 56±2 67 9 сутки 42±1 14±1 82±1 28±1 122±5 41±1 82±23 28±7 34 12 сутки 52±5 12±1 53±8 12±1 54±5 12±1 53±0,5 12±0 22 1 сутки 93±2 107±5 85±2 98±3 59±1 68±1 79±10 69±11 115 3 сутки 110±5 326±20 90±6 266±15 94±2 278±10 98±6 290±18 296 4 сутки 100±3 78±2 65±1 51±1 30±1 23±1 65±20 51±15 78 7 сутки 83±1 56±1 42±1 28±1 10±0,5 7±0,1 45±21 30±14 67 9 сутки 99±2 74±1 54±1 40±1 51±1 38±1 68±15 51±11 75 12 сутки 49±2 24±1 34±12 17±5 29±5 14±2 37±6 19±2 50 173 КА 1344 Даурия Продолжение приложения 24 1 сутки 126±5 101±3 128±5 102±3 112±2 90±2 122±5 98±3 80 3 сутки 105±2 204±8 76±1 147±5 59±1 114±5 80±13 155±26 194 4 сутки 100±3 76±1 122±5 93±2 150±2 114±2 124±14 94±10 76 7 сутки 80±1 73±1 82±1 75±1 78±1 71±1 80±1 73±1 91 9 сутки 126±8 99±2 130±5 103±2 143±5 113±2 133±5 105±4 79 12 сутки 39±3 23±2 29±2 17±1 37±1 22±1 35±3 20±1 58 1 сутки 78±1 62±1 90±2 71±1 129±5 102±3 99±15 78±12 79 3 сутки 95±2 192±5 80±1 162±3 86±1 174±5 87±4 175±8 202 4 сутки 162±5 264±10 200±15 326±12 241±10 393±21 201±22 327±37 163 7 сутки 152±6 251±3 126±2 208±6 112±3 185±2 130±11 215±19 165 9 сутки 74±10 68±8 73±15 67±5 74±10 68±10 74±0,3 67±0,3 91 12 сутки 53±12 31±2 73±5 43±4 91±10 53±5 72±10 42±6 58 НСР05 1,30 1,56 1,63 1,64 НСРа 0,46 0,55 0,57 0,58 НСРв 0,35 0,41 0,43 0,43 НСРс 0,24 0,29 0,30 0,31 НСРа – время после инфицирования; НСРв – сорт; НСРс – вариант опыта (здоровые и пораженные) 174 Приложение 25 Коэффициенты корреляции степени поражения септориозом с удельной активностью ферментов в пораженных листьях сои (в среднем по сортам) при искусственном заражении Анализируемая пара Время после Коэффициент параметров инфицирования корреляции степень поражения 1 сутки -0,65 ± 0,05 септориозом – удельная 3 сутки -0,97 ± 0,02 активность пероксидазы в 4 сутки -0,38 ± 0,10 изолированных листьях сои 7 сутки -0,37 ± 0,10 9 сутки 0,86 ± 0,03 12 сутки -0,63 ± 0,05 степень септориозом активность – поражения 1 сутки -0,96 ± 0,04 удельная 3 сутки -0,46 ± 0,05 каталазы изолированных листьях сои степень в 4 сутки -0,21 ± 0,22 7 сутки -0,27 ± 0,28 9 сутки -0,54 ± 0,06 12 сутки 0,71 ± 0,04 поражения 1 сутки -0,41 ± 0,07 удельная 3 сутки -0,67 ± 0,05 активность кислой фосфатазы в 4 сутки -0,57 ± 0,06 изолированных листьях сои 7 сутки -0,38 ± 0,09 9 сутки -0,73 ± 0,04 12 сутки -0,99 ± 0,02 септориозом – tтеор при Р0,95=1,96 Корреляционная связь несущественна tфакт < tтеор 175 Приложение 26 Содержание белка и незаменимых аминокислот в семенах сои (здоровые и пораженные пероноспорозом), %, 2010 г. Показатель Protein Сорт Контроль (здоровые) 38,26 ± 0,71 36,03 ± 0,53 38,49 ± 0,72 Соната Гармония Даурия НСР05 Lis Соната Гармония Даурия 6,76 ± 0,09 6,81 ± 0,07 6,69 ± 0,07 Соната Гармония Даурия 4,61 ± 0,02 4,60 ± 0,02 4,64 ± 0,03 Соната Гармония Даурия 9,09 ± 0,05 9,25 ± 0,07 8,84 ± 0,02 Соната Гармония Даурия 4,64 ± 0,01 4,86 ± 0,02 4,83 ± 0,05 Соната Гармония Даурия 7,41 ± 0,03 7,43 ± 0,08 7,08 ± 0,05 Соната Гармония Даурия 7,34 ± 0,05 7,29 ± 0,07 7,05 ± 0,07 1,44 3,63 ± 0,01 3,69 ± 0,01 2,36 ± 0,02 НСР05 Met+Cys 4,87 ± 0,02 4,95 ± 0,01 4,96 ± 0,02 1,58 НСР05 Tre 9,20 ± 0,07 9,39 ± 0,08 8,98 ± 0,06 1,54 НСР05 Val 4,65 ± 0,02 4,62 ± 0,03 4,67 ± 0,03 1,88 НСР05 Iley 6,78 ± 0,05 6,83 ± 0,06 6,73 ± 0,08 1,20 НСР05 Ley 38,31 ± 0,83 36,63 ± 0,65 39,46 ± 0,92 2,04 НСР05 Phe Пораженные Соната Гармония Даурия 3,65 ± 0,03 3,72 ± 0,02 3,59 ± 0,02 1,31 1,21 ± 0,01 1,02 ± 0,01 1,09 ± 0,01 НСР05 176 1,05 ± 0,01 0,94 ± 0,01 1,02 ± 0,01 1,54 Приложение 27 Содержание белка и незаменимых аминокислот в семенах сои (здоровые и пораженные пероноспорозом), %, 2011 г. Показатель Protein Сорт Контроль (здоровые) 42,16 ± 0,80 37,77 ± 0,54 39,63 ± 0,66 Соната Гармония Даурия НСР05 Lis Соната Гармония Даурия 6,77 ± 0,05 6,73 ± 0,05 6,82 ± 0,03 Соната Гармония Даурия 4,61 ± 0,02 4,60 ± 0,04 4,64 ± 0,02 Соната Гармония Даурия 8,99 ± 0,07 8,05 ± 0,09 8,27 ± 0,09 Соната Гармония Даурия 5,89 ± 0,05 6,05 ± 0,06 6,04 ± 0,06 Соната Гармония Даурия 6,70 ± 0,07 6,04 ± 0,04 6,53 ± 0,05 6,23 ± 0,02 6,15 ± 0,06 7,27 ± 0,07 1,54 Соната Гармония Даурия 2,91 ± 0,01 2,59 ± 0,01 2,71 ± 0,02 НСР05 Met+Cys 6,28 ± 0,05 6,58 ± 0,04 5,94 ± 0,04 1,64 НСР05 Tre 8,59 ± 0,08 8,58 ± 0,08 8,76 ± 0,07 1,88 НСР05 Val 4,65 ± 0,04 4,62 ± 0,05 4,67 ± 0,03 1,90 НСР05 Iley 6,71 ± 0,06 6,93 ± 0,07 6,98 ± 0,07 1,80 НСР05 Ley 41,55 ± 0,92 38,13 ± 0,70 39,46 ± 0,84 1,82 НСР05 Phe Пораженные 2,70 ± 0,02 2,83 ± 0,02 2,89 ± 0,02 1,72 Соната Гармония Даурия 1,08 ± 0,01 1,23 ± 0,01 1,35 ± 0,01 НСР05 0,99 ± 0,01 1,27 ± 0,01 1,38 ± 0,01 0,89 177 Приложение 28 Содержание масла и жирных кислот в семенах сои (здоровые и пораженные пероноспорозом), %, 2010 г. Показатель Oil Сорт Контроль (здоровые) 18,05 ± 0,20 17,99 ± 0,41 17,83 ± 0,40 Соната Гармония Даурия НСР05 C/16 Соната Гармония Даурия 9,23 ± 0,08 9,32 ± 0,07 9,10 ± 0,07 Соната Гармония Даурия 3,85 ± 0,01 3,90 ± 0,02 3,78 ± 0,02 Соната Гармония Даурия 14,15 ± 0,09 15,10 ± 0,08 16,17 ± 0,07 13,07 ± 0,04 14,76 ± 0,05 14,61 ± 0,04 0,91 Соната Гармония Даурия 52,66 ± 0,25 52,10 ± 0,53 52,34 ± 0,54 НСР05 C/18:3 3,89 ± 0,03 3,94 ± 0,03 3,82 ± 0,02 0,67 НСР05 C/18:2 9,25 ± 0,09 9,35 ± 0,05 9,17 ± 0,08 0,96 НСР05 C/18:1 17,49 ± 0,23 17,59 ± 0,33 16,36 ± 0,30 0,90 НСР05 C/18 Пораженные 52,01 ± 0,61 51,77 ± 0,74 51,96 ± 0,80 0,97 Соната Гармония Даурия 10,87 ± 0,06 9,69 ± 0,07 10,63 ± 0,09 НСР05 10,34 ± 0,09 9,92 ± 0,05 11,21 ± 0,07 1,30 178 Приложение 29 Содержание масла и жирных кислот в семенах сои (здоровые и пораженные пероноспорозом), %, 2011 г. Показатель Oil Сорт Контроль (здоровые) 18,48 ± 0,32 18,98 ± 0,30 18,12 ± 0,25 Соната Гармония Даурия НСР05 C/16 Соната Гармония Даурия 9,42 ± 0,08 9,18 ± 0,09 9,53 ± 0,09 Соната Гармония Даурия 3,69 ± 0,03 3,55 ± 0,04 3,75 ± 0,03 Соната Гармония Даурия 15,20 ± 0,05 16,76 ± 0,05 16,73 ± 0,03 14,04 ± 0,04 14,86 ± 0,06 12,33 ± 0,03 1,15 Соната Гармония Даурия 50,88 ±0,90 52,08 ± 0,82 52,08 ± 0,85 НСР05 C/18:3 3,83 ± 0,01 3,69 ± 0,02 3,87 ± 0,02 0,95 НСР05 C/18:2 9,44 ± 0,07 9,20 ± 0,07 9,63 ± 0,06 0,70 НСР05 C/18:1 16,94 ± 0,24 16,38 ± 0,40 16,90 ± 0,33 0,84 НСР05 C/18 Пораженные 50,80 ± 0,76 51,43 ± 0,74 51,96 ± 0,80 0,81 Соната Гармония Даурия 7,17 ± 0,05 8,30 ± 0,05 7,59 ± 0,05 НСР05 10,46 ± 0,08 10,10 ± 0,09 10,23 ± 0,06 1,05 179 Приложение 30 Показатели продуктивности сои Сорт Высота растений, см Кол-во бобов, шт Соната Гармония Даурия НСР05 52,77 ± 3,82 41,62 ± 2,90 58,24 ± 2,71 2,66 17,76 ± 2,14 18,83 ± 3,57 17,89 ± 1,89 3,36 Соната Гармония Даурия НСР05 93,18 ± 3,14 98,92 ± 2,20 84,49 ± 3,12 2,68 23,96 ± 3,21 30,95 ± 4,57 29,60 ± 4,13 3,02 Соната Гармония Даурия НСР05 90,33 ± 2,75 76,41 ± 3,82 71,33 ± 4,80 2,63 19,89 ± 3,06 18,19 ± 3,45 14,85 ± 2,97 2,63 Соната Гармония Даурия НСР05 78,76±13,02 72,32±16,6 71,35±7,5 8,11 20,54±1,81 22,66±3,98 20,78±4,49 4,68 Кол-во семян, шт 2009 г 34,87 ± 10,34 40,85 ± 11,04 29,69 ± 8,67 1,56 2010 г 49,78 ± 6,56 71,95 ± 8,75 56,06 ± 7,51 2,43 2011 г 41,78 ± 10,18 44,29 ± 8,50 30,68 ± 7,98 2,92 Ср. по годам 42,14±4,30 52,36±9,84 38,81±8,62 11,97 180 Масса семян с растения, г Масса 1000 семян, г Урожайность, ц/га 4,25 ± 1,46 4,91 ± 1,08 5,15 ± 1,52 0,73 120,73 ± 6,75 126,22 ± 5,27 178,03 ± 11,81 2,44 12,37 ± 1,7 16,77 ± 1,2 20,20 ± 1,2 2,19 6,55 ± 1,08 9,51 ± 2,02 10,5 ± 1,43 2,70 121,12 ± 5,51 135,46 ± 7,36 167,81 ± 6,14 2,69 19,30 ± 1,5 20,95 ± 1,8 26,34 ± 1,4 2,91 4,97 ± 1,78 5,57 ± 1,23 5,22 ± 1,17 2,74 119,81 ± 5,30 143,69 ± 7,56 170,90 ± 8,16 2,72 20,22 ± 1,2 22,45 ± 1,4 21,72 ± 1,3 2,66 5,26±0,67 6,66±1,43 6,96±1,77 2,22 120,55±0,38 135,12±5,04 172,25±3,02 13,08 17,30±2,47 20,06±1,69 22,75±1,84 1,11 Приложение 31 Удельная активность пероксидазы в семенах сои при поражении разной инфекцией, ед/мг белка Сорт Контроль (здоровые) пероноспороз Пораженные аскохитоз Соната Гармония Даурия НСР05 92 ± 4 129 ± 7 136 ± 5 2,63 80 ± 2 108 ± 6 116 ± 5 2,17 62 ± 3 150 ± 9 97 ± 2 3,04 Соната Гармония Даурия НСР05 84 ± 8 71 ± 5 125 ± 16 1,31 83 ± 2 79 ± 12 116 ±15 0,76 - Соната Гармония Даурия НСР05 127 ± 10 141 ± 12 122 ± 6 1,78 125 ± 8 134 ± 17 113 ±13 2,90 22 ± 0,5 30 ± 1 38 ± 3 3,81 Соната Гармония Даурия НСР05 105±13 114±21 127±4 2,62 96±14 107±21 115±29 1,31 42±2 90±6 67±2 3,32 фузариоз 2009 г 82 ± 1 13 ± 0,3 0,76 2010 г 18 ± 0,5 10 ± 0,5 33 ± 1 2,08 2011 г 37 ± 1 49 ± 3 35 ± 1 1,31 Ср. значения 46±1 29±2 27±1 2,74 181 церкоспороз пурпурный церкоспороз антракноз бактериоз - - - 77 ± 2 113 ± 8 2,40 56 ± 2 22 ± 1 94 ± 3 3,73 64 ± 2 70 ± 2 83 ± 5 2,64 39 ± 1 68 ± 2 2,01 - 57 ± 1 39 ± 2 79 ± 4 2,28 86 ± 2 52 ± 2 96 ± 3 2,41 - 106 ± 15 117 ± 11 102 ± 14 3,31 56±0,5 30±1 86±3 2,40 75±2 61±2 89±4 2,74 39±1 68±2 2,22 87±15 95±11 106±11 2,62 Приложение 32 Удельная активность каталазы в семенах сои при поражении разной инфекцией, ед/мг белка х 10-3 Сорт Контроль (здоровые) пероноспороз Пораженные аскохитоз Соната Гармония Даурия НСР05 59 ± 2 83 ± 1 49 ± 2 1,31 89 ± 2 170 ± 2 88 ± 2 3,73 74 ± 1 63 ± 1 43 ± 3 1,30 Соната Гармония Даурия НСР05 34 ± 3 32 ± 3 51 ± 2 2,63 47 ± 1 46 ± 0,5 76 ± 2 1,32 - Соната Гармония Даурия НСР05 69 ± 2 72 ± 1 48 ± 1 2,28 131 ± 5 115 ± 1 105 ± 5 3,32 58 ± 1 67 ± 2 64 ± 3 2,01 Соната Гармония Даурия НСР05 54±2 62±2 49±1 3,04 89±3 110±15 90±8 0,75 66±8 65±1 53±3 2,60 фузариоз 2009 г 88 ± 2 73 ± 2 2,28 2010 г 102 ± 5 98 ± 4 80 ± 2 1,31 2011 г 199 ± 7 122 ± 3 134 ± 1 2,95 Ср. значения 130±34 110±12 96±19 0,76 182 церкоспороз пурпурный церкоспороз антракноз бактериоз - - - 100 ± 3 69 ± 1 2,40 86 ± 2 71 ± 1 66 ± 1 3,18 46 ± 1 60 ± 2 78 ± 2 3,04 69 ± 0,5 58 ± 0,5 1,52 - 116 ± 1 69 ± 2 48 ± 2 2,65 209 ± 6 81 ± 3 61 ± 1 2,8 - 150 ± 10 95 ± 8 67 ± 5 2,74 101±15 70±11 57±9 3,32 127±13 70±10 69±8 2,74 69±0,5 58±0,5 1,61 125±6 95±8 68±3 2,96 Приложение 33 Удельная активность кислой фосфатазы в семенах сои, пораженных разной инфекцией, ед/мг белка х 10 -5 Сорт Контроль (здоровые) пероноспороз Пораженные аскохитоз Соната Гармония Даурия НСР05 17 ± 1,0 21 ± 0,5 22 ± 1,0 2,65 24 ± 1,2 36 ± 3,0 36 ± 1,0 2,90 15 ± 1,0 12 ± 0,7 12 ± 0,5 1,52 Соната Гармония Даурия НСР05 16 ± 1,1 30 ± 2,0 22 ± 1,0 0,93 22 ± 0,8 36 ± 2,0 25 ± 1,0 2,08 - Соната Гармония Даурия НСР05 18 ± 0,7 17 ± 0,6 15 ± 1,0 2,62 19 ± 0,5 21 ± 0,2 19 ± 0,9 3,18 22 ± 0,7 10 ± 0,1 9 ± 0,2 1,54 Соната Гармония Даурия НСР05 17±0,5 23±4 20±2 3,61 22±1 31±2 27±1 2,74 18±1 11±1 10±1 3,04 фузариоз 2009 г 4 ± 0,3 5 ± 0,2 0,76 2010 г 5 ± 0,2 7 ± 0,1 9 ± 0,3 2,63 2011 г 5 ± 0,2 9 ± 0,5 7 ± 0,1 1,61 Ср. значения 5±0,3 8±0,3 7±0,2 2,22 183 церкоспороз пурпурный церкоспороз антракноз бактериоз - - - 16 ± 1,0 19 ± 1,0 2,28 39 ± 3,0 52 ± 3,0 34 ± 2,0 2,95 14 ± 1 21 ± 2 22 ± 1 2,40 19 ± 1,0 15 ± 1,0 1,78 - 46 ± 1,5 23 ± 1,1 32 ± 2,4 2,01 13 ± 1 13 ± 2 16 ± 3 2,90 - 15 ± 0,8 15 ± 0,5 13 ± 0,2 2,24 42±2 37±2 33±1 2,22 14±1 17±1 19±4 1,86 19±1 15±1 1,20 15±0,5 15±0,5 16±3 3,05 Приложение 34 Удельная активность пероксидазы в проростках сои полученных из семян, пораженных разной инфекцией, ед/мг белка х 10-2 Сорт Контроль (проростки из здоровых семян) пероноспороз Соната Гармония Даурия НСР05 27 ± 2 20 ± 1 14 ± 1 2,01 5 ± 0,1 18 ± 0,5 9 ± 0,1 1,54 Соната Гармония Даурия НСР05 42 ± 2 16 ± 1 10 ± 1 1,52 84 ± 3 25 ± 2 8 ± 0,5 1,20 Соната Гармония Даурия НСР05 34±7 18±2 12±2 2,63 44±39 21±3 8±0,5 2,08 Проростки из пораженных семян церкоспороз пурпурный антракноз церкоспороз 2010 г 53 ± 2 20 ± 2 12 ± 1 3,32 2011 г 60 ± 2 18 ± 1 9 ± 0,5 2,28 Ср. значения 56±3 19±1 10±1 1,78 184 бактериоз 17 ± 1 18 ± 1 6 ± 0,1 2,22 19 ± 1 18 ± 1 2,44 20 ± 2 16 ± 1 5 ± 0,1 1,31 50 ± 2 18 ± 1 8 ± 0,1 1,31 - 38 ± 3 12 ± 1 7 ± 0,5 2,22 33±16 18±0 7±1 2,40 19±1 18±1 1,56 29±9 14±2 6±1 2,42 Приложение 35 Удельная активность каталазы в проростках сои полученных из семян, пораженных разной инфекцией, ед/мг белка х 10-4 Сорт Контроль (проростки из здоровых семян) пероноспороз Соната Гармония Даурия НСР05 33 ± 1 45 ± 1 53 ± 2 2,46 44 ± 1 51 ± 1 59 ± 1 2,40 Соната Гармония Даурия НСР05 136 ± 16 105 ± 8 108 ± 10 1,86 194 ± 8 120 ± 5 122 ± 3 2,01 Соната Гармония Даурия НСР05 84±51 75±30 80±27 2,63 119±75 85±34 90±31 2,03 Проростки из пораженных семян церкоспороз пурпурный антракноз церкоспороз 2010 г 42 ± 2 50 ± 2 47 ± 1 1,61 2011 г 140 ± 12 110 ± 5 106 ± 5 1,78 Ср. значения 91±49 80±30 76±29 3,04 185 бактериоз 26 ± 1 39 ± 1 50 ± 2 0,76 38 ± 2 47 ± 2 1,87 27 ± 1 39 ± 1 48 ± 2 1,52 130 ± 6 98 ± 2 100 ± 3 1,86 - 126 ± 10 76 ± 2 90 ± 5 1,78 78±52 68±29 75±25 2,01 38±2 47±2 1,87 76±49 57±18 69±21 1,86 186