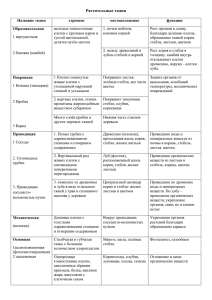
Курс лекций по ботанике
advertisement

Курс лекций по дисциплине «Ботаника» подготовлен д.б.н., профессором Криворотовым С.Б. Содержание: Лекция 1 Введение. Цели и задачи ботаники. Развитие ботаники в XIX и XX вв. 2 Лекция 2 Строение и функции растительной клетки 5 Лекция 3 Морфология корня 12 Лекция 4 Морфология листа 16 Лекция 5 Морфология стебля 20 Лекция 6 Анатомия корня 24 Лекция 7 Анатомия стебля 27 Лекция 8 Анатомия листа 34 Лекция 9 Растительные ткани. Анатомия растительных тканей. 35 Лекция 10 Строение генеративных органов растений 43 Лекция 11 Размножение растений 52 Лекция 12 Систематика растений 55 1 Лекция 1 Введение. Цели и задачи ботаники. Развитие ботаники в XIX и XX вв. Ботаника - наука о растениях. Ботанические знания зародились и быстро накапливались с практической деятельностью человека. Ботаника как наука сформировалась более 2000 лет назад. Основоположниками ее были выдающиеся деятели древнего мира Аристотель (384 - 322 гг. до н. э.) и Феофраст (371 - 286 гг. до н. э.). Они обобщили накопленные сведения о разнообразии растений и их свойствах, приемах возделывания, размножения и использовании, географическом распространении. В наши дни ботаника представляет собой большую многоотраслевую науку. Общая задача ее состоит в изучении отдельно взятых растений и их совокупностей - растительных сообществ. Структура и закономерности роста растений, их отношения с окружающей средой, закономерности распространения и распределения отдельных видов и всего растительного покрова на земном шаре; происхождение и эволюция царства растений, причины его разнообразия и классификация; запасы в природе хозяйственно ценных растений и пути их рационального использования, разработка научных основ введения в культуру (интродукции) новых кормовых, лекарственных, плодовых, овощных, технических и других растений - далеко не полный перечень вопросов, которые рассматриваются ботаническими науками. Значение растений в природе и жизни человека. Континенты нашей планеты, составляющие 150 млн. км2, в основном покрыты растительностью. Только ледяные пространства полюсов и высочайшие вершины гор не имеют растительности. Площади, занятые морями и океанами (акватории), составляют около 360 млн. км2. Здесь широко представлены водные растения. Подавляющее число растений имеет зеленый цвет, обусловленный зеленым пигментом хлорофиллом, сосредоточенным в особых органеллах растительных клеток — хлоропластах. Уникальное свойство хлорофилла - участие в сложнейшем процессе трансформации электромагнитной энергии солнечного луча в химическую энергию органических веществ (фотосинтез). Процесс фотосинтеза разворачивается в поистине колоссальных масштабах. По определению К. А. Тимирязева, хлорофилловое зерно - тот фокус, та точка в мировом пространстве, где солнечный луч, превращаясь в химическую энергию, становится источником всей жизни на Земле. Точно определить объем работы, выполняемой растениями, трудно и даже невозможно. По весьма приблизительным подсчетам, растения в процессе фотосинтеза ежегодно образуют около 400 млрд. т 2 органических веществ, при этом они связывают около 175 млрд. т углерода, но, возможно, гораздо больше. Итак, в ходе эволюции жизни на Земле, зародившейся миллиарды лет назад, растения обособились как носители хлорофилла, как единственные организмы, способные синтезировать органические вещества из неорганических. В ходе фотосинтеза параллельно с образованием органического вещества происходит выделение в атмосферу кислорода. До появления зеленых растений газовая оболочка Земли имела очень мало свободного кислорода. Практически можно считать, что весь кислород атмосферы возник благодаря фотосинтезу. Накопление свободного кислорода привело к появлению кислородного дыхания растений и животных. Возросли энергия жизненных процессов и скорость накопления массы органических веществ. Наличие свободного кислорода усилило также процессы химического выветривания горных пород и накопление в верхних слоях земной коры минеральных соединений, необходимых для питания растений. Растительный покров играет первостепенную регулирующую роль в общем газообмене и в водном балансе Земли, защищает от разрушения почву, обогащает ее элементами питания, создает пищевую и энергетическую базу для всего животного мира. Жизнь человека немыслима без использования растений. Это - пища, строительный материал, сырье для различных отраслей промышленности. Разделы ботаники. Ботанику как часть более общей науки - биологии, в свою очередь, подразделяют на ряд частных наук, в задачи которых входит изучение тех или иных закономерностей строения и жизни растений или растительного покрова. Морфология - один из наиболее крупных и рано сформировавшихся разделов ботаники. Это наука о закономерностях возникновения и развития разнообразных жизненных форм растений и отдельных их органов. Заложение и развитие органов растения рассматривают и в ходе индивидуального развития отдельной особи от прорастания семени до конца жизни (онтогенез), и в ходе исторического развития (эволюции) всего вида или любой другой систематической группы, к которой относят данную особь (филогенез). В процессе развития морфологии в ее недрах обособились еще более специализированные науки: цитология (закономерности строения и развития основной структурной единицы растений — клетки); гистология, или анатомия (заложение, развитие и строение разнообразных тканей, формирующих органы); эмбриология (закономерности развития и строения зародыша); органография (заложение, развитие и структура органов растения); палинология (строение пыльцы и спор). 3 Флорография. В задачу этой науки входит распознавание и описание видов. Виды, описанные флорографами, систематики распределяют в группы по признакам сходства, отражающим родство. Систематика - наука о разнообразии видов и причинах этого разнообразия. Задача систематики - приведение в легко обозримую научную систему всех наших знаний о видах, описанных флорографами. На основании целой серии методов систематик объединяет родственные виды в систематические группы более высокого ранга - роды, семейства и т. д. География растений (фитогеография) - крупнейший раздел ботаники, основная задача которого состоит в изучении закономерностей распространения и распределения растений и их сообществ (ценозов) на суше и в воде. Экология. Жизнь растений зависит от окружающей среды (климата, почвы и др.), но и растения, в свою очередь, влияют на создание этой среды - принимают участие в почвообразовательном процессе, изменяют климат. Задача экологии - изучение строения и жизни растений в связи с окружающей средой. Эта наука имеет первостепенное значение для практического земледелия. Физиология растений - наука о процессах жизнедеятельности растений, преимущественно об обмене веществ, движении, росте, ритмах развития, размножении и т. д. Микробиология - наука об особенностях жизненных процессов, происходящих в микроскопических организмах, преобладающую часть которых составляют бактерии и некоторые грибы. Успехи почвенной микробиологии широко используют в сельскохозяйственной практике. Палеоботаника - наука об ископаемых растениях прошлых геологических периодов. Другие разделы ботаники настолько обособились в связи с решением специальных задач и применяемыми методами работы, что давно уже составляют особые науки, например биофизика, биохимия, радиобиология, генетика и др. Взаимосвязь ботаники и агрономии. Эти науки связаны общим объектом изучения, методами работы и историей развития. Ботаники исследуют закономерности строения и развития, видовой состав дикорастущих растений и их группировок; агрономы имеют дело с возделываемыми растениями. Агрономия возникла как приложение ботаники к растениеводству. Перед агрономами и ботаниками стоит одна цель - возможно более полное использование растений для практических потребностей человека. Комплексное использование растительных ресурсов, как природных, так и возделываемых, - важный показатель общего уровня развития страны и ее земледелия. 4 Цитология - наука о микроскопической и субмикроскопической структуре клетки и ее жизнедеятельности. Лекция 2 Строение и функции растительной клетки Клетка - это элементарная структурная и функциональная единица тела растений и животных, способная к самовоспроизведению. Первым увидел клетку английский естествоиспытатель Р. Гук при изучении покровной ткани бузины - пробки. Он усовершенствовал микроскоп, изобретенный Г. Галилеем в 1609 г., и использовал его для исследования тонких срезов органов растений. Свои наблюдения Р. Гук изложил в сочинении «Микрография», изданном в 1665 г., где он впервые применил термин «клетка». В 1833 г. английский ботаник Р. Броун обнаружил ядро, а в 1839 г. чешский физиолог Я. Пуркинье - цитоплазму. Многочисленные наблюдения клеточного строения растений и животных позволили немецким ученым - ботанику М. Шлейдену и зоологу Т. Шванну - в 1838 - 1839 гг. сформулировать клеточную теорию, суть которой заключается в том, что клетка - это основная элементарная структурная единица всех живых орга¬низмов. Клеточная теория доказывает единство происхождения, строения и эволюции растений и животных. К концу XIX в. цитология окончательно сформировалась в самостоятельную науку. На базе светового микроскопа были изучены основные компоненты клетки, накапливались данные об их функциях. Дальнейший прогресс цитологии связан с изобретением электронного микроскопа. В биологии его начали использовать в середине XX в. Методы исследования клетки. Применяемые для изучения клеток методы очень разнообразны. Основной из них микроскопический. Большую роль продолжает играть световой микроскоп, современные модели которого дают увеличение до 2 тыс. раз. Но возможности светового микроскопа ограничены, частицы менее 0,2 мкм рассмотреть при помощи такого микроскопа невозможно. Электронный микроскоп дает увеличение в 200 - 300 тыс. раз и более. Здесь вместо пучка света используют поток электронов, движущихся с высокой скоростью. Методом культуры тканей изучают структуру и жизнедеятельность живых клеток вне организма. Цитохимический метод позволяет выявить наличие и определить количество различных веществ в клетке. 5 Разделить компоненты клетки с различной плотностью для изолированного изучения их можно с помощью метода центрифугирования. Извлечь из клетки отдельные компоненты (ядро, митохондрии и др.) позволяет метод микроскопической хирургии. Разнообразие клеток. Все разнообразие форм клеток можно свести к двум группам: паренхимные клетки - длина равна ширине или превышает ее не более чем в 2 - 3 раза; прозенхимные клетки - длина превышает ширину во много раз. Средняя длина клеток высших растений 10 - 100 мкм. Наиболее крупные паренхимные клетки достигают в длину несколько миллиметров и видны невооруженным глазом, например клетки плодов арбуза, лимона, клубней картофеля. Но особенно большую длину имеют прозенхимные клетки стеблей льна (40 мм), крапивы (80 мм), рами (200 мм). Компоненты клетки. Рассматривая взрослую растительную клетку при помощи светового микроскопа, можно увидеть следующие компоненты: плотную стенку, одну большую или 2 - 3 небольшие вакуоли, занимающие центральную часть клетки; цитоплазму, расположенную между стенкой клетки и вакуолью; находящееся в цитоплазме ядро. Ядро и цитоплазма - живые части клетки и в совокупности составляют протопласт. Стенка и вакуоли - неживые части клетки, производные протопласта, продукты его жизнедеятельности. Цитоплазма - сложная структурная система, имеет мембранную организацию, состоит из гиалоплпзмы и органелл. Ее структуру образуют тонкие (4 - 10 нм), довольно плотные пленки - биологические мембраны. Основу их составляют липиды. Молекулы липидов расположены упорядоченно - перпендикулярно к поверхности, в два слоя, так, что их части, интенсивно взаимодействующие с водой (гидрофильные), направлены наружу, а части, инертные по отношению к воде (гидрофобные), - внутрь. Молекулы белка расположены несплошным слоем на поверхности липидного каркаса с обеих его сторон. Часть их погружена в липидный слой, а некоторые проходят через него насквозь, образуя участки, проницаемые для воды. Мембраны образуют пограничный слой цитоплазмы, а также внешнюю границу ее органелл и участвуют в создании их внутренней структуры. Они делят цитоплазму на изолированные отсеки, в которых одновременно и независимо друг от друга могут протекать биохимические процессы часто в противоположном направлении (например, синтез и распад). Одно из основных свойств биологических мембран - избирательная проницаемость (полупроницаемость): одни вещества проходят через них с трудом, другие легко и даже в 6 сторону большей концентрации. Мембраны во многом определяют химический состав цитоплазмы и клеточного сока. Плазмодесмы - тончайшие цитоплазматические нити, которые осуществляют связь между клетками. Клеточная пластинка, образующаяся при цитокинезе, пронизана трубочками эндоплазматического ретикулума, которые не разобщаются. На их основе и формируются плазмодесмы. Стенка плазмодесменного канала выстлана плазмалеммой, соединяющей плазмалеммы смежных клеток. В центре канала проходит трубка, сохраняющая непрерывность ретикулума обеих клеток. Между плазмалеммой и трубкой находится гиалоплазма, также непрерывная для обеих клеток. Плазмодесмы чаще всего бывают собраны в группы по нескольку десятков. Поодиночке они располагаются в стенках, не имеющих вторичных утолщений. Плазмалемма и тонопласт. Плазмалемма — это мембрана, отграничивающая цитоплазму от стенки клетки и обычно плотно прилегающая к ней. Иногда плазмалемма бывает волнистой или образует глубокие складки. Она регулирует обмен веществ клетки с окружающей средой, а также участвует в синтезе веществ. Тонопласт отграничивает цитоплазму от вакуоли. Функция его та же, что и плазмалеммы. Гиалоплазма. Это жидкая непрерывная среда, в которую погружены органеллы. Гиалоплазма содержит ферменты и нуклеиновые кислоты. Белки, входящие в состав гиалоплазмы, образуют сеть из тонких фибрилл (диаметром 2 - 3 нм) - трабекулярную систему, которая связывает между собой органеллы. Эта система очень динамична, она может распадаться при изменении внешних условий. Гиалоплазма способна к активному движению, которое может быть вращательным - вдоль стенки клетки, если в центре находится одна большая вакуоль, и струйчатым - по тяжам, пересекающим центральную вакуоль, в разных направлениях. Скорость движения зависит от температуры, интенсивности света, снабжения кислородом и других факторов. При движении гиалоплазма увлекает за собой органеллы. Гиалоплазма осуществляет взаимосвязь органелл, участвует в обмене, транспорте веществ, передаче раздражения и т. д. Эндоплазматический ретикулум (эндоплазматическая сеть). Представляет собой отграниченную, мембранами систему взаимосвязанных субмикроскопических каналов и цистерн, пронизывающих гиалоплазму. Имеются две формы ретикулума: гранулярный (шероховатый) и агранулярный (гладкий). Гранулярный эндоплазматический ретикулум несет на поверхности мелкие гранулы - рибосомы. Агранулярный эндоплазматический ретикулум состоит из ветвящихся трубочек, отходящих от цистерн гранулярного ретикулума, не имеет рибосом. Обычно он развит слабее, чем гранулярный. Участвует в синтезе и транспорте эфирных масел, смол, каучука. 7 Рибосомы. Это гранулы диаметром около 20 нм, расположенные в гиалоплазме или прикрепленные к поверхности мембран эндоплазматического ретикулума. Они обнаружены также в митохондриях и пластидах. Рибосомы состоят из белка и рибонуклеиновой кислоты (РНК) и не имеют мембранной структуры. Функция рибосом синтез белка, самовоспроизводство живой материи. Этот процесс происходит в рибосомах, расположенных группой и связанных между собой нитевидной молекулой иРНК. Такие группы называют полисомами. Считают, что рибосомы формируются в ядре. Аппарат Гольджи. Состоит из диктиосомы и пузырьков Гольджи. Диктиосома представляет собой стопку из 5 - 7 плоских цистерн, ограниченных агранулярной мембраной. Диаметр цистерн около 1 мкм, толщина 20 - 40 нм. Цистерны не соприкасаются друг с другом. Пузырьки Гольджи отчленяются от краев цистерн и распространяются по всей гиалоплазме. В диктиосоме происходят синтез, накопление и выделение полисахаридов. Пузырьки Гольджи транспортируют их, в том числе и к плазмалемме. Мембрана пузырьков встраивается в плазмалемму, а содержимое оказывается снаружи от плазмалеммы и может включаться в стенку. Пузырьки Гольджи могут включаться и в тонопласт. Происхождение диктиосом еще точно не установлено. Считают, что в их образовании принимает участие эндоплазматический ретикулум. В некоторых клетках аппарат Гольджи отсутствует. Сферосомы. Округлые блестящие тельца диаметром 0,5 - 1 мкм. Это центры синтеза и накопления растительных масел. Они отшнуровываются от концов тяжей эндоплазматического ретикулума. Мембрана, расположенная на поверхности сферосомы, по мере накопления масла редуцируется, и от нее остается только наружный слой. Лизосомы. Пузырьки размером 0,5—2 мкм, имеющие на поверхности мембрану. Содержат ферменты, которые могут расщеплять белки, липиды, полисахариды и другие органические соединения. Образуются так же, как и сферосомы, из тяжей эндоплазматического ретикулума. Их функция - разрушение отдельных органелл или участков цитоплазмы (локальный автолиз), необходимое для обновления клетки. Митохондрии. Форма митохондрий чрезвычайно разнообразна - овальная, округлая, цилиндрическая, гантелевидная, ветвистая и т.д. Длина их равна 2 - 5 мкм, диаметр — 0,3 - 1 мкм. На поверхности митохондрии находятся две мембраны. Внутренняя мембрана образует выросты в полость митохондрии в виде гребней или трубочек, называемых кристами. Кристы значительно увеличивают мембранную поверхность митохондрии. Пространство между кристами заполнено жидким веществом - матриксом, в котором находятся рибосомы и содержится дезоксирибонуклеиновая кислота (ДНК). Поверхность 8 внутренней мембраны покрыта мельчайшими тельцами, имеющими шаровидную головку и ножку (АТФ-сомы). Митохондрии - это энергетические лаборатории клетки. Здесь происходят расщепление углеводов, жиров и других органических веществ при участии кислорода (дыхание) и синтез АТФ. Выделяемая при дыхании энергия преобразуется в энергию макроэргических связей молекулы АТФ, которая затем используется для осуществления процессов жизнедеятельности клетки - деления, поглощения и выделения веществ, синтеза и т.д. Пластиды. Бывают только у растений. Эти органеллы имеют на поверхности две мембраны. В зависимости от окраски различают три типа пластид: хлоропласт - зеленого цвета; хромопласты - желтого, оранжевого, красного цветов; лейкопласты - бесцветные. Хлоропласты содержат зеленый пигмент хлорофилл, а также пигменты из группы каротиноидов - каротин (оранжевый) и ксантофилл (желтый). Именно с хлорофиллом связана основная функция хлоропластов - синтез органических веществ из неорганических при участии энергии света (фотосинтез). Поэтому хлоропласты присутствуют только в клетках наземных органов, на которые падает солнечный свет. Хлоропластам растения обязаны зеленой окраской. У высших растений хлоропласты в большинстве случаев имеют линзовидную форму. Диаметр их 4 - 6 мкм, толщина 1 - 3 мкм. У водорослей хлоропласты, называемые также хроматофорами, очень разнообразны по форме и величине. Они могут иметь звездчатую форму, лентовидную, сетчатую и др. Обычно в клетке имеются от 1 до 50 хлоропластов. Располагаются они в постенном слое цитоплазмы. Внутри хлоропластов находится однородное вещество - строма, пронизанная системой параллельно расположенных мембран. Мембраны имеют вид плоских мешков, их называют тилакоидами, или ламеллами. У большинства высших растений часть тилакоидов имеет дисковидную форму. Эти тилакоиды собраны в стопки, называемые гранами. Хлорофилл и каротиноиды находятся в каждой из двух мембран тилакоида граны. Граны связаны между собой тилакоидами стромы. Внутренняя мембрана оболочки хлоропласта иногда образует складки и переходит в тилакоиды стромы. В строме находятся молекулы ДНК, рибосомы, капли липидов, называемые пластоглобулами, крахмальные зерна и другие включения. Крахмал образуется в хлоропластах из продуктов фотосинтеза, его называют фотосинтетическим, или первичным. При помощи ферментов фотосинтетический крахмал осахаривается и в виде глюкозы транспортируется из листа на построение органов или в запас. 9 Лейкопласты не содержат пигментов. По размеру они значительно меньше хлоропластов и не имеют строго определенной формы. Лейкопласты присутствуют чаще всего в клетках тканей и органов, на которые не падает солнечный свет: в корнях, клубнях, семенах и др. Внутренняя мембранная система у лейкопластов развита значительно слабее, чем у хлоропластов. В строме имеются молекулы ДНК, рибосомы, пластоглобулы. Основная функция лейкопластов - синтез и накопление запасных питательных продуктов, в первую очередь крахмала, иногда белков, редко масла. Лейкопласты, накапливающие крахмал, называют амилопластами. В них из сахаров, поступающих из фотосинтезирующих органов, образуются крахмальные зерна различного размера и формы - вторичный крахмал. Запасной белок может откладываться в виде кристаллов или аморфных гранул, масло - в виде пластоглобул. Хромопласты содержат красные, оранжевые, желтые пигменты из группы каротиноидов. По размеру они меньше хлоропластов, форма их очень разнообразна. Внутренняя мембранная система у них чаще всего отсутствует. В зависимости от формы накопления каротиноидов различают хромопласты глобулярного, фибриллярного (трубчатого) и кристаллического типов. У хромопластов наиболее распространенного глобулярного типа пигменты растворены в пластоглобулах. У хромопластов фибриллярного типа каротиноиды не только присутствуют в пластоглобулах, но и образуют группы параллельных нитей или трубок, расположенных в строме. У хромопластов кристаллического типа каротиноиды присутствуют главным образом в виде кристаллов различной формы, определяющих форму самой пластиды (серповидную, ромбовидную, игловидную и т. д.). Хромопласты встречают в клетках лепестков некоторых растений, зрелых плодов, осенних листьев. Их функция в процессе обмена веществ не выяснена. Косвенное биологическое значение хромопластов состоит в привлечении насекомых для перекрестного опыления и животных для распространения семян. В процессе эволюции первыми из пластид появились хлоропласты, из которых при расчленении тела растения на органы образовались лейкопласты и хромопласты. В онтогенезе почти все виды пластид могут переходить друг в друга. Наиболее часто происходит превращение лейкопластов в хлоропласты (например, при образовании зародыша из оплодотворенной яйцеклетки) и хлоропластов в хромопласты (например, при осеннем пожелтении листьев). Лишь хромопласты в природных условиях, как правило, не превращаются в другие виды пластид. Количество пластид в клетке увеличивается за счет деления их путем перетяжки. 10 Ядро может функционировать только в цитоплазматической среде. Это - место хранения и воспроизводства наследственной информации, определяющей признаки данной клетки и всего организма в целом, а также центр управления синтезом белка. Если из клетки удалить ядро, то она вскоре погибнет. Обычно в клетке имеется одно ядро, но у некоторых видов водорослей и у грибов многоядерные клетки. Бактерии и сине-зеленые водоросли не имеют оформленного ядра. Форма ядра разнообразна, но обычно соответствует форме клетки: в паренхимных клетках чаще всего шаровидная, в прозенхимных - линзовидная или веретеновидная. Диаметр ядра клеток вегетативных органов покрытосеменных растений 10 - 25 мкм. У плесневых грибов диаметр ядра всего 1 - 2 мкм, а у харовых водорослей может достигать 2,5 мм. В процессе онтогенеза форма, размер и местоположение ядра в клетке могут изменяться. Под световым микроскопом ядро имеет вид пузырька с 1 -3 темными пятнышками ядрышками. Оно состоит из ядерной оболочки, нуклеоплазмы, хромосом, ядрышек. Ядерная оболочка отграничивает содержимое ядра от цитоплазмы. Состоит из двух мембран с промежутком между ними, называемым перинуклеарным пространством. Толщина мембран 10 нм, а толщина перинуклеарного пространства варьирует. Общая толщина оболочки 40 - 80 нм. Внутренняя мембрана оболочки агранулярная, к наружной мембране прикреплены рибосомы. По структуре и химическому составу ядерная оболочка близка к эндоплазматическому ретикулуму, тем более что ее наружная мембрана образует выросты, переходящие в ретикулум цитоплазмы. Ядерная оболочка имеет особые образования - ядерные поры. Это сложные структуры. По границе поры, образованной в результате слияния двух мембран, расположены гранулы, от которых отходят фибриллы. Часть фибрилл сходится в центре, формируя диафрагму. Диаметр поры 80—90 нм. Через поры макромолекулы проходят из нуклеоплазмы в гиалонлазму и в обратном направлении. Ядерная оболочка контролирует обмен веществ между ядром и цитоплазмой, способна к синтезу белков и липидов. Нуклеоплазма представляет собой коллоидный раствор, в котором размещены хромосомы и ядрышки. В состав нуклеоплазмы входят различные ферменты, нуклеиновые кислоты. Она не только осуществляет связь между органеллами ядра, но и трансформирует вещества, проходящие через нее. 11 Лекция 3 Морфология корня Корень в типичных случаях - осевой полисимметричный подземный орган, который неопределенно долго нарастает в длину верхушкой, защищенной чехликом, и никогда не образует листьев, ветвление и заложение почек происходят эндогенно (из внутренних тканей). Корень выполняет разные функции: поглощает воду, минеральные и органические вещества из почвы и проводит их в стебель, закрепляет растение в субстрате, синтезирует некоторые органические вещества, осуществляет связь с микроорганизмами почвы - грибами, бактериями; накапливает запасные продукты, служит для вегетативного размножения. Разнообразие корней. В зависимости от происхождения различают главный корень, придаточные и боковые. Главный корень образуется только из зародышевого корешка. Придаточные корни берут начало от стебля и листа или их видоизменений. От главного и придаточного корней отходят боковые корни - оси второго и последующих порядков ветвления. По форме корни исключительно многообразны: нитевидные, шнуровидные, конусовидные, веретеновидные, реповидные, клубневидные и др. По отношению к субстрату различают корни: земляные, водяные, воздушные и гаустории (присоски растений-паразитов). Приведенная классификация не охватывает всего разнообразия корней. Так, выделяют сокращающиеся (контрактильные) корни, которые втягивают в почву основания побегов с почками возобновления, и корневые мочки - кратковременные нитевидные корни, образующиеся близко к поверхности почвы в связи с сезонным увлажнением или поливом. Корневая система. Это совокупность всех корней растения, образующихся в результате их нарастания и ветвления. По происхождению корневые системы делят на три основных типа. Корневые системы: главного корня, образующаяся из корешка зародыша и состоящая из главного корня (ось первого порядка) и боковых корней разных порядков; придаточных корней, образующаяся из любой части стебля или листа; смешанная, состоящая из одновременно функционирующих систем главного корня и придаточных корней. По форме корневые системы бывают стержневые, когда ось первого порядка преобладает над остальными по размеру (у бобовых); мочковатые, состоящие из многих одинаковых по размеру корней (у злаков); ветвистые, когда оси второго порядка по мощности едва уступают оси первого порядка (у деревьев), и др. 12 Размеры корневой системы зависят как от жизненной формы растений (дерево, кустарник, трава), так и от внешних условий. У водных и болотных растений корневая система небольшая. Растения засушливых местообитаний, например степных или пустынных, имеют очень мощную корневую систему. Так, у пшеницы, произрастающей на богаре, подземная масса в 140 раз превышает надземную. Главный, боковой и придаточные корни имеют довольно сходное строение. Зоны корня. Корень по длине можно разделить на несколько зон, имеющих различное строение и функции. Выделяют зоны деления клеток, растяжения клеток, всасывания (корневых волосков), проведения (ветвления). Зоны деления и растяжения клеток расположены на самом кончике. Это небольшой гладкий участок с корневым чехликом на верхушке. Корневой чехлик состоит из тонкостенных клеток. Он предохраняет конус нарастания от повреждения о частицы почвы. Поверхностные клетки корневого чехлика отпадают, что облегчает продвижение корня. Чехлик покрывает зону деления клеток, состоящую из первичной меристемы. Выше деление клеток постепенно прекращается, клетки увеличиваются, вытягиваясь в длину, это зона растяжения клеток. Иногда эти две зоны объединяют в одну - зону роста. Зона всасывания примыкает к зоне растяжения. Здесь на поверхности корня появляется множество бугорков, которые вытягиваются и превращаются в корневые волоски. Каждый корневой волосок представляет собой длинный (0,15 - 1 мм) вырост одной из поверхностных клеток. Стенка волоска тонкая, целлюлозная, ядро обычно находится в кончике его. Корневые волоски поглощают из почвы раствор минеральных веществ. Они функционируют 10 - 20 дней. В более старой части зоны они постоянно отмирают, а в молодой - постоянно образуются вновь. Поэтому зона всасывания перемещается и находится вблизи кончика корня. Длина ее обычно составляет несколько миллиметров. Одновременно с образованием корневых волосков происходит дифференциация внутренних тканей этой зоны. Далее расположена зона проведения. Она тянется вплоть до корневой шейки и составляет большую часть корня. Здесь уже нет корневых волосков, на поверхности находится покровная ткань. На этом участке корень ветвится. Первичное строение. Дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы зоны роста. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. При первичном строении в корне различают центральный цилиндр и первичную кору, покрытую одним рядом клеток с корневыми волосками - эпиблемой. 13 Наружный слой первичной коры - экзодерма - состоит из плотно сомкнутых многоугольных или вытянутых клеток, стенки которых у однодольных впоследствии опробковевают и выполняют защитную функцию. За экзодермой расположена основная паренхима, составляющая основную массу первичной коры. Внутренний слой первичной коры - эндодерма - состоит обычно из одного ряда клеток, на антиклинальных стенках которых образуются утолщения, опоясывающие клетку (поясок Каспари). Эти утолщения стенки и срединная пластинка опробковевают. Считают, что пояски Каспари блокируют передвижение поглощенного корневыми волосками почвенного раствора по стенкам клетки, и он вынужден проходить через цитоплазму клеток эндодермы, обладающую избирательной проницаемостью. У однодольных растений утолщение может распространяться не только на всю антиклинальную стенку клетки эндодермы, но и на внутреннюю периклинальную. При этом утолщенные стенки не только опробковевают, но и одревесневают. Однако у клеток эндодермы, расположенных напротив участков ксилемы, стенка может оставаться тонкой. Такие клетки называют пропускными. Эндодерма регулирует поступление веществ в ксилему. Наружный слой центрального цилиндра - перицикл - состоит из одного ряда живых паренхимных клеток, обладающих меристематической активностью. Из перицикла образуются боковые корни и придаточные почки. Центральная часть центрального цилиндра занята радиальным проводящим пучком. Ксилема расположена в центре и образует ряд острых выступов в виде лучей, заканчивающихся снаружи более мелкими сосудами. Между выступами ксилемы располагаются участки флоэмы. У однодольных растений первичное строение сохраняется и в зоне проведения. Здесь лишь отсутствует эпиблема, а защитную функцию выполняет экзодерма. Основные признаки отличия первичного строения корня от первичного строения стебля следующие: у корня никогда не бывает эпидермы, колленхимы; первичная кора занимает большую часть корня; проводящий пучок всегда радиального типа и расположен в центре центрального цилиндра, в центральном цилиндре чаще всего не образуется сердцевина. Вторичное строение. У двудольных и голосеменных растений в центральном цилиндре корня уже в раннем возрасте формируется камбий, деятельность которого приводит к вторичным изменениям и значительному утолщению корня. Камбий образуется между флоэмой и ксилемой из клеток прокамбия. К центру он откладывает клетки, дифференцирующиеся в элементы вторичной ксилемы, а кнаружи - клетки, дифференцирующиеся в элементы вторичной флоэмы. В тех местах, где сосуды первичной ксилемы соприкасаются с перициклом, клетки последнего также превращаются в камбий, который дифференцируется только в паренхиму, образуя 14 радиальные лучи. Первоначально слой камбия на поперечном срезе имеет извилистую форму, а затем округляется и отодвигается от центра. Вместе с увеличением вторичных ксилемы и флоэмы первичная флоэма отодвигается на периферию. Из клеток перицикла возникает пробковый камбий, который образует кнаружи слой пробки. Благодаря этому первичная кора изолируется от центрального цилиндра, отмирает и отпадает. Остается только центральный цилиндр, покрытый пробкой. Таким образом, при вторичном строении в центре корня расположена первичная ксилема. От ее лучей начинаются радиальные лучи - участки тонкостенной живой паренхимы. С радиальными лучами чередуются широкие участки вторичной ксилемы с крупными сосудами и мелкоклеточной древесинной паренхимой. Они окружены камбиальной зоной, состоящей из мелких тонкостенных клеток, расположенных правильными радиальными рядами. Кнаружи от камбиальной зоны, против каждого участка вторичной ксилемы, находится вторичная флоэма. Камбий, производящий паренхиму радиальных лучей, малозаметен. Кнаружи он откладывает также основную паренхиму. Ткани, расположенные кнаружи от камбия (флоэма, основная паренхима, феллодерма и пробковый камбий), называют вторичной корой. Метаморфизированные корни. Наиболее распространены из них следующие. Запасающие корни. Видоизменение возникает в результате разрастания древесинной и лубяной паренхимы в связи с отложением в них запасных продуктов. По происхождению и строению различают два типа запасающих корней: корнеплоды и корневые клубни. Корнеплоды формируются из главного корня. В их образовании принимает участие также гипокотиль. У корнеплода раз¬личают: головку - укороченную стеблевую часть с листьями; шейку - наиболее толстую часть, образовавшуюся за счет разрастания гипокотиля; собственно корень, от которого отходят боковые корни. Длина шейки у корнеплодов различных видов растений может быть разной. Корнеплоды бывают монокамбиальные - с одним слоем камбия, и поликамбиальные - с несколькими слоями камбия. У одних монокамбиальных корнеплодов большую часть корнеплода занимает вторичная кора, где и накапливаются запасные продукты (морковь), у других вторичная кора маленькая, а большую часть корнеплода занимает ксилема, в которой откладываются запасные продукты (репа). Поликамбиальный корнеплод образуется у свеклы. При таком строении в центре находится диархная первичная ксилема, к которой прилегают два участка вторичной ксилемы, разделенные радиальными участками паренхимы. Камбий окружает ксилему. Снаружи к нему прилегают участки вторичной флоэмы. Таким образом, вторичное строение такое же, как и у других 15 корнеплодов. Но вслед за вторичным наступает третичное изменение. Вокруг вторичной флоэмы на периферии корня благодаря делению клеток перицикла образуется слой паренхимных клеток. В этом слое один ряд клеток начинает делиться тангенциальными перегородками и превращается в новый слой камбия, откладывающий внутрь ксилему, а кнаружи - флоэму в виде коллатеральных пучков, отделенных друг от друга прослойкой тонкостенной паренхимы. Одновременно в периферическом слое паренхимы образуется новое кольцо камбиальных клеток и т. д. Корневые клубни (корневые шишки) образуются в том случае, когда запасные продукты откладываются в боковых и придаточных корнях. На корневых клубнях нередко имеются придаточные почки. Они могут служить для вегетативного размножения. Микориза (грибокорень). Так называют кончики корней вместе с живущими с ними в симбиозе гифами гриба. Микориза может быть: эктотрофной, когда гифы расположены только снаружи корня; экто-эндотрофной, когда гифы частично проникают в клетку корня; эндотрофной, когда гифы живут только в клетках корня. Эктотрофная микориза чаще бывает у деревьев и кустарников, эндотрофная - у травянистых растений. Гриб, поселяясь на корне растения, питается органическими веществами из тканей последнего и в то же время доставляет ему из почвы воду с растворенными в ней минеральными солями. Ферменты, имеющиеся в клетках гриба, минерализуют органические вещества почвы и этим способствуют усвоению их растением. Микориза распространена очень широко. Клубеньки. В корнях бобовых поселяются особые бактерии из рода ризобиум, способные усваивать атмосферный азот. Бактерии питаются органическими веществами растения, а растение потребляет азотистые соединения, синтезированные бактериями. Внедрение бактерий в корень вызывает разрастание тканей коры корня в виде опухолей, называемых клубеньками, Клетки, заполненные бактериями (бактероидная ткань), расположены группой, образуя бактериальное гнездо. В клубеньке может быть одно или несколько таких гнезд. Лекция 4 Морфология листа. Это боковой орган ограниченного роста, нарастающий основанием путем вставочного роста (однодольные) или всей поверхностью (двудольные); у деревьев и кустарников это временный орган. 16 Основные функции - фотосинтез, газообмен, транспирация. В листьях могут откладываться запасные продукты, в отдельных случаях листья могут служить для вегетативного размножения. У однолетних растений продолжительность жизни листа примерно равна продолжительности жизни стебля, у многолетних - значительно короче. У большинства растений лист живет не более 1 -1,5 года, но чаще меньше. У вечнозеленых растений лист функционирует 1 - 5 лет, а у некоторых - до 10 - 15 лет (ель, араукария). Исключение составляет растение африканских пустынь - вельвичия, у которой лист - постоянный орган и живет 90 - 100 лет. Листопад - это биологическая защита растений от испарения при физической (летом) или физиологической (зимой) засухе. Вместе с листьями растение освобождается от накопившихся экскреторных веществ. У однодольных и травянистых двудольных растений лист отмирает и разрушается постепенно, оставаясь на стебле. У древесных двудольных растений у основания черешка формируется отделительный слой, клетки претерпевают естественную мацерацию, и достаточно небольшого механического воздействия (ветер, дождь), чтобы лист упал. След, оставшийся от листа на стебле, покрывается пробкой, его называют листовым рубцом. У вечнозеленых растений листья опадают неодновременно. Макроскопическое строение. Размер листьев сильно варьирует, нередко даже в пределах одного растения. В нашей флоре некоторые виды имеют очень мелкие листья - длиной до 1 - 1,5 мм. У растений тропической и субтропической зон листья достигают 20 - 22 м длины (пальмы). Формации листьев, гетерофиллия. Обычно на одном побеге образуются листья, неодинаковые по величине, форме, окраске. Различают три формации листьев: низовую, срединную и верхушечную. Листья низовой формации обычно недоразвитые или видоизмененные в связи с выполнением специализированной функции (защитной, запасающей) - семядольные листья, почечные чешуйки, редуцированные листья корневищ, а иногда и надземных побегов. Листья срединной формации составляют основную массу листьев растения. Это типичные для данного вида листья и в отличие от предыдущих хлорофиллоносные. Листья верхушечной формации расположены на цветоносных побегах (в соцветиях). Это прицветники, обвертки и т. д. Все они, как правило, недоразвиты, не имеют черешков, окрашены или бесцветны. Когда говорят о листьях растения, то подразумевают срединную формацию листьев. Иногда срединные листья одного побега заметно различаются по форме, как, например, 17 подводные, плавающие и надводные листья некоторых водяных растений. Такое явление получило название гетерофиллии. Части листа. У большинства растений лист состоит из более или менее широкой пластинки, прикрепленной к стеблю при помощи черешка (черешковый лист). Пластинка выполняет основные функции листа. Черешок ориентирует пластинку по отношению к источнику света. Если черешок отсутствует, лист называют сидячим. Если пластинка сидячего листа прирастает к стеблю на некотором протяжении, образуется нисбегающий лист. Часто у основания черешка образуются парные боковые выросты - прилистники, зеленые или пленчатые. Обычно они меньше пластинки, но у некоторых растений превосходят ее по величине и функционируют как пластинка (бобовые). Если прилистники срастаются, то образуется раструб (гречишные). Иногда основание черешка расширяется во влагалище, охватывающее стебель (зонтичные). У злаков лист состоит из длинного трубчатого влагалища и узкой пластинки. У основания пластинки имеется пленчатый придаток - язычок, а иногда еще два выроста по бокам - ушки. Жилкование. Различают следующие типы: простое - листовую пластинку пронизывает от основания до верхушки только одна жилка (проводящий пучок); встречается у высших споровых (моховидных, плауновидных), многих голосеменных (хвойных) и некоторых покрытосеменных (элодея); дихотомическое - жилки ветвятся вильчато (дихотомически); из семенных растений известно у гинкго (один из немногих представителей широколиственных голосеменных); сетчатое - одна или несколько крупных жилок дают боковые ответвления, образующие густую сеть; наиболее широко распространенный тип жилкования; различают перистое и пальчатое; параллельное и дуговое - листовую пластинку от основания до верхушки пронизывают несколько неветвящихся одинаковых жилок; в одних случаях они расположены строго параллельно (злаки, осоки), в других - дугообразно (ландыш). Разнообразие листьев. Различают листья простые и сложные. Простые листья имеют одну пластинку, цельную или выемчатую. У древесных растений они опадают осенью, а у травянистых чаще всего отмирают вместе со стеблем. Сложные листья состоят обычно из нескольких (двух или более) листочков, прикрепленных (у тройчатых и пальчатых листьев) к общему черешку или (у перистых листьев) к общей оси - рахису - короткими черешочками, образующими сочленения. Благодаря этому сложный лист опадает по частям - сначала листочки, а затем черешок (или черешок с рахисом). 18 Простые листья свойственны почти всем травянистым растениям и подавляющему большинству деревьев и кустарников. Их классифицируют по целому ряду признаков. Вот выжнейшие из них. Листья с цельной пластинкой: по форме пластинки - яйцевидные, округлые, ланцетные, эллиптические, продолговатые, линейные и др.; по форме верхушки пластинки - тупые, острые, заостренные, остроконечные, выемчатые; по форме основания пластинки - сердцевидные, округлые, клиновидные, стреловидные, копьевидные; по форме края пластинки - цельнокрайние, пильчатые, двоякопильчатые, зубчатые, городчатые, выемчатые. Листья с выемчатой пластинкой: по глубине выемки - лопастные (выемки достигают не более четверти ширины половины пластинки), раздельные (выемки достигают одной трети ширины половины пластинки и более), рассеченные (выемки достигают главной жилки листа), по расположению выемок тройчато-, пальчато-, перисто-. Если у перисторассеченного листа крупные доли чередуются с мелкими, то его называют прерывистоперисторассеченным (картофель). Иногда пластинка бывает рассечена дважды или многократно (тысячелистник, укроп). Сложные листья классифицируют по расположению листочков: пальчатосложные листочки располагаются на верхушке черешка в одной плоскости и расходятся более или менее радиально; перистосложные - листочки располагаются по длине рахиса, причем на верхушке его может располагаться один листочек (непарноперистосложные) или два листочка (парноперистосложные); тройчатые - лист имеет только три листочка. Перистосложные листья иногда бывают более сложной конструкции— дважды- и многократноперистосложные. Наиболее примитивным типом листа у одних групп растений (многоплодниковые, букоцветные и другие, а также голосеменные) считают простой лист, цельный или лопастной; у других (бобовоцветные, розоцветные, рутовые и др.) - сложный лист. Прилистники также рассматривают как признак низкой организации. Наиболее примитивным типом жилкования следует считать простое и дихотомическое. 19 Лекция 5 Морфология стебля Побег. Это орган, который возникает из верхушечной меристемы и расчленяется на раннем этапе морфогенеза на специализированные части: стебель, листья, почки. Основная его функция - фотосинтез. Части побега могут служить также для вегетативного размножения, накопления запасных продуктов, воды. Макроскопическое строение. Части побега. Участок стебля на уровне отхождения листа называют узлом, а участок стебля между двумя узлами - междоузлием. Над узлом в пазухе листа образуется пазушная почка. В случае ясно выраженных междоузлий побег называется удлиненным. Если же узлы сближены и междоузлия практически незаметны, то это укороченный побег (плодушка, розетка). Метамерия. Обычно побег имеет несколько узлов и междоузлий. Такое повторение отрезков побега, имеющих одноименные органы, называют метамерией. Каждый метамер типичного побега состоит из узла с листом и пазушной почкой и нижележащего междоузлия. Почка. Это зачаточный побег. Он состоит из меристематической оси, заканчивающейся конусом нарастания (зачаточный стебель), и листовых примордиев (зачаточных листьев), то есть из серии зачаточных метамеров. Расположенные ниже дифференцированные листья прикрывают конус нарастания и примордии. Так устроена вегетативная почка. У вегетативно-репродуктивной почки конус нарастания превращен в зачаточный цветок или зачаточное соцветие. Репродуктивные (цветочные) почки состоят только из зачаточного цветка или соцветия и не имеют зачатков фотосинтезирующих листьев. Часто наружные листья видоизменяются в почечные чешуйки, предохраняющие почку от высыхания. Такие почки называют защищенными (закрытыми) в отличие от голых (открытых) почек, не имеющих почечных чешуй (калина, живучка, кошачья лапка). Надо помнить, что в голых почках, как и в любых растущих, конус нарастания и зачатки листьев закрыты более взрослыми фотосинтезирующими листьями. По местоположению различают верхушечные и боковые почки. Последние по происхождению могут быть пазушными и придаточными. Пазушные почки закладываются на конусе нарастания экзогенно (наружно) в пазухах листовых примордиев. Пазушные почки, длительное время не дающие побегов, называют спящими. Пазушные почки располагаются или по одной (одиночные), или по нескольку 20 (групповые). Придаточные почки могут возникнуть в любой части стебля эндогенно за счет деятельности меристемы. Иногда придаточные почки образуются на листьях и сразу же дают маленькие побеги с придаточными корнями (бриофиллум) или луковички (лук). Такие придаточные почки называют выводковыми. Листорасположение. Различают три основных варианта листорасположения: спиральное (очередное) - на узле имеется только один лист, на стебле листья располагаются по спирали; супротивное - на узле имеются два листа, расположенные друг против друга; мутовчатое - на узле находятся три или более листьев. Нарастание. Побег растет в длину обычно верхушкой вследствие деятельности расположенной там верхушечной меристемы. Кроме того, побеги многих растений существенно удлиняются благодаря росту вставочной меристемы. Если побег растет неопределенно долго за счет одной и той же верхушечной меристемы, такое нарастание называют моноподиальным. Однако у многих растений верхушечная меристема функционирует ограниченное время, обычно в течение одного вегетационного периода. Тогда в следующий сезон рост побега продолжается за счет ближайшей боковой ночки. Происходит так называемое перевершинивание. Такое нарастание побега называют симподиальным. Способность к замещению отмерших верхушечных почек боковыми (симподиальное нарастание) имеет большое биологическое значение. Это увеличивает жизнеспособность растений. Растения, у которых боковые почки слаборазвиты и неспособны замещать отмершие верхушечные почки, при повреждении верхушек стебля гибнут (например, некоторые пальмы). Поэтому в сухом (аридном) и холодном климатах практически все многолетние растения имеют симподиальное нарастание. Для влажных тропиков характерны растения с моноподиальным нарастанием. Возможность симподиального нарастания широко используют в практике. На этом явлении основаны приемы обрезки плодовых и декоративных растений. Оно лежит в основе отрастания травостоя при покосе и пастьбе скота. Ветвление. Ветвление бывает двух типов: верхушечное и боковое. При верхушечном (дихотомическом) ветвлении происходит разделение конуса нарастания на две или большее число осей. Такое ветвление свойственно низшим растениям (некоторые водоросли) и лишь немногим высшим (плауновидные, некоторые папоротниковидные). При боковом ветвлении новые оси возникают ниже верхушки. В результате одного или нескольких ветвлений образуется система осей. При боковом ветвлении система осей может быть или моноподиальной - при моноподиальном нарастании, или симподиальной - при симподиальном нарастании. 21 Особую форму ветвления представляет кущение, при котором наиболее крупные боковые разветвления образуются только у основания побегов, обычно из приземных и подземных почек. Этот участок побега называют зоной кущения. Кущение свойственно кустарникам, многолетним, а иногда и однолетним травам. У некоторых растений боковые почки на оси первого порядка недоразвиты и не образуют боковых разветвлений. Такие растения имеют неразветвленный стебель (большинство пальм, дынное дерево, агава). Направление роста. Вертикально растущие побеги могут быть прямостоячими, цепляющимися, вьющимися. Побеги, лежащие на земле, называют стелющимися. Если стелющийся побег образует придаточные корни, его называют ползучим. Побеги могут менять направление роста, тогда их называют приподнимающимися, восходящими. Метаморфизированные побеги. Возникновение их часто связано с выполнением функций вместилища запасных продуктов, перенесения неблагоприятных условий года, вегетативного размножения. Корневище - это многолетний подземный побег с горизонтальным, восходящим или вертикальным направлением роста, выполняющий функции накопления запасных продуктов, возобновления, вегетативного размножения. Корневище имеет редуцированные листья в виде чешуек, почки, придаточные корни. Запасные продукты накапливаются в стеблевой части. Нарастание и ветвление происходит так же, как у обычного побега. Корневище отличают от корня по наличию листьев и отсутствию корневого чехлика на верхушке. Корневище может быть длинным и тонким (пырей) или коротким и толстым. Ежегодно из верхушечной и пазушных почек образуются надземные однолетние побеги. Старые части корневища постепенно отмирают. Растения с горизонтальными длинными корневищами, формирующими много надземных побегов, быстро занимают большую площадь, и если это сорняки (пырей), то борьба с ними довольно затруднительна. Такие растения используют для закрепления песков (колосняк, аристида). В луговодстве злаки с длинными горизонтальными корневищами называют корневищными (полевица, мятлик), а с короткими - кустовыми (тимофеевка, белоус). Корневища встречаются в основном у многолетних травянистых растений, но иногда у кустарников (бересклет) и кустарничков (брусника, черника). Клубень - это утолщенная часть побега, вместилище запасных продуктов. Клубни бывают надземными и подземными. Надземный клубень представляет собой утолщение главного (кольраби) или бокового (тропические орхидеи) побега и несет нормальные листья. 22 Подземный клубень - утолщение гипокотиля (цикламен) или недолговечного подземного побега - столона (картофель). Листья на подземном клубне редуцируются, в пазухах их находятся почки, называемые глазками. Надземный столон - это недолговечный ползучий побег, служащий для распространения (захвата территории) и вегетативного размножения. Он имеет длинные междоузлия и зеленые листья. На узлах образуются придаточные корни, а из верхушечной почки укороченный побег (розетка), который после отмирания столона продолжает самостоятельное существование. Нарастает надземный столон симподиалыю. Надземные столоны, утратившие функцию фотосинтеза и выполняющие в основном функцию вегетативного размножения, иногда называют усами (земляника). Луковица - это укороченный стебель (донце), несущий многочисленные, тесно сближенные листья и придаточные корни. На верхушке донца находится почка. У многих растений (лук, тюльпан, гиацинт и др.) из этой почки образуется надземный побег, а из боковой пазушной почки формируется новая луковица. Наружные чешуи в большинстве случаев сухие, пленчатые и выполняют защитную функцию, внутренние - мясистые, заполненные запасными продуктами. По форме луковицы бывают шаровидные, яйцевидные, сплющенные и т. д. Клубнелуковица внешне похожа на луковицу, но все листовые чешуи у нее сухие, а запасные продукты откладываются в стеблевой части (шафран, гладиолус). Колючки имеют различное происхождение - из побега (яблоня, груша, терн, боярышник, гледичия, цитрусовые), листа (барбарис) или его частей: рахиса (астрагал), прилистников (акация белая), участка пластинки (сложноцветные). Колючки характерны для растений жарких сухих местообитаний. Усики образуются из побега (виноград), листа или его частей: рахиса и нескольких листочков (горох), пластинки (чина.), прилистников (сассапариль). Служат для прикрепления к опоре. Филлокладии - это плоские листовидные побеги, расположенные в пазухах редуцированных листьев. На них образуются цветки. Встречаются у растений преимущественно засушливых местообитаний (иглица, филлантус). Ловчие аппараты - видоизмененные листья, свойственные насекомоядным .растениям (росянка, мухоловка). Имеют форму кувшинчиков, урночек, пузырьков или же захлопывающихся и завертывающихся пластинок. Небольшие насекомые, попадая в них, погибают, растворяются при помощи ферментов и потребляются растениями в качестве главным образом дополнительного источника минеральных веществ. 23 В типичных случаях это - осевой полисимметричный орган неограниченного роста, несущий листья и почки; увеличение в длину происходит путем верхушечного и вставочного роста, ветвление и из наружных тканей (экзогенно). Стебель обеспечивает связь между листьями и корнями, обусловливает образование мощной ассимиляционной поверхности листьев и наилучшее размещение их по отношению к свету, служит вместилищем запасных продуктов. Стебли (как и корни) древесных растений могут достигать возраста 4 - 6 тыс. лет (мамонтово и драконово деревья). У некоторых трав возраст стебля ограничен всего 30 - 45 днями (растенияэфемеры). Макроскопическое строение. Форма поперечного сечения стебля чаще всего бывает цилиндрической, но у травянистых растений встречаются стебли трехгранные (осоки), четырехгранные (яснотковые), крылатые (чина лесная), плоские (рдест). Длина стеблей колеблется в больших пределах: от 280 - 300 м (лазящие стебли ротанговых пальм) до 1,5 мм (водное растение вольфия). Лекция 6 Анатомия корня Корень в типичных случаях - осевой полисимметричный подземный орган, который неопределенно долго нарастает в длину верхушкой, защищенной чехликом, и никогда не образует листьев, ветвление и заложение почек происходят эндогенно (из внутренних тканей). Корень выполняет разные функции: поглощает воду, минеральные и органические вещества из почвы и проводит их в стебель, закрепляет растение в субстрате, синтезирует некоторые органические вещества, осуществляет связь с микроорганизмами почвы - грибами, бактериями; накапливает запасные продукты, служит для вегетативного размножения. Макроскопическое строение. Разнообразие корней. В зависимости от происхождения различают главный корень, придаточные и боковые. Главный корень образуется только из зародышевого корешка. Придаточные корни берут начало от стебля и листа или их видоизменений. От главного и придаточного корней отходят боковые корни - оси второго и последующих порядков ветвления. По форме корни исключительно многообразны: нитевидные, шнуровидные, конусовидные, веретеновидные, реповидные, клубневидные и др. 24 По отношению к субстрату различают корни: земляные, водяные, воздушные и гаустории (присоски растений-паразитов). Приведенная классификация не охватывает всего разнообразия корней. Так, выделяют сокращающиеся (контрактильные) корни, которые втягивают в почву основания побегов с почками возобновления, и корневые мочки - кратковременные нитевидные корни, образующиеся близко к поверхности почвы в связи с сезонным увлажнением или поливом. Зоны корня. Корень по длине можно разделить на несколько зон, имеющих различное строение и функции. Выделяют зоны деления клеток, растяжения клеток, всасывания (корневых волосков), проведения (ветвления). Зоны деления и растяжения клеток расположены на самом кончике. Это небольшой гладкий участок с корневым чехликом на верхушке. Корневой чехлик состоит из тонкостенных клеток. Он предохраняет конус нарастания от повреждения о частицы почвы. Поверхностные клетки корневого чехлика отпадают, что облегчает продвижение корня. Чехлик покрывает зону деления клеток, состоящую из первичной меристемы. Выше деление клеток постепенно прекращается, клетки увеличиваются, вытягиваясь в длину, это зона растяжения клеток. Иногда эти две зоны объединяют в одну - зону роста. Зона всасывания примыкает к зоне растяжения. Здесь на поверхности корня появляется множество бугорков, которые вытягиваются и превращаются в корневые волоски. Каждый корневой волосок представляет собой длинный (0,15 - 1 мм) вырост одной из поверхностных клеток. Стенка волоска тонкая, целлюлозная, ядро обычно находится в кончике его. Корневые волоски поглощают из почвы раствор минеральных веществ. Они функционируют 10 - 20 дней. В более старой части зоны они постоянно отмирают, а в молодой - постоянно образуются вновь. Поэтому зона всасывания перемещается и находится вблизи кончика корня. Длина ее обычно составляет несколько миллиметров. Одновременно с образованием корневых волосков происходит дифференциация внутренних тканей этой зоны. Далее расположена зона проведения. Она тянется вплоть до корневой шейки и составляет большую часть корня. Здесь уже нет корневых волосков, на поверхности находится покровная ткань. На этом участке корень ветвится. Первичное строение. Дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы зоны роста. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. При первичном строении в корне различают центральный цилиндр и первичную кору, покрытую одним рядом клеток с корневыми волосками - эпиблемой. 25 Наружный слой первичной коры - экзодерма - состоит из плотно сомкнутых многоугольных или вытянутых клеток, стенки которых у однодольных впоследствии опробковевают и выполняют защитную функцию. За экзодермой расположена основная паренхима, составляющая основную массу первичной коры. Внутренний слой первичной коры - эндодерма - состоит обычно из одного ряда клеток, на антиклинальных стенках которых образуются утолщения, опоясывающие клетку (поясок Каспари). Эти утолщения стенки и срединная пластинка опробковевают. Считают, что пояски Каспари блокируют передвижение поглощенного корневыми волосками почвенного раствора по стенкам клетки, и он вынужден проходить через цитоплазму клеток эндодермы, обладающую избирательной проницаемостью. У однодольных растений утолщение может распространяться не только на всю антиклинальную стенку клетки эндодермы, но и на внутреннюю периклинальную. При этом утолщенные стенки не только опробковевают, но и одревесневают. Однако у клеток эндодермы, расположенных напротив участков ксилемы, стенка может оставаться тонкой. Такие клетки называют пропускными. Эндодерма регулирует поступление веществ в ксилему. Наружный слой центрального цилиндра - перицикл - состоит из одного ряда живых паренхимных клеток, обладающих меристематической активностью. Из перицикла образуются боковые корни и придаточные почки. Центральная часть центрального цилиндра занята радиальным проводящим пучком. Ксилема расположена в центре и образует ряд острых выступов в виде лучей, заканчивающихся снаружи более мелкими сосудами. Между выступами ксилемы располагаются участки флоэмы. У однодольных растений первичное строение сохраняется и в зоне проведения. Здесь лишь отсутствует эпиблема, а защитную функцию выполняет экзодерма. Основные признаки отличия первичного строения корня от первичного строения стебля следующие: у корня никогда не бывает эпидермы, колленхимы; первичная кора занимает большую часть корня; проводящий пучок всегда радиального типа и расположен в центре центрального цилиндра, в центральном цилиндре чаще всего не образуется сердцевина. Вторичное строение. У двудольных и голосеменных растений в центральном цилиндре корня уже в раннем возрасте формируется камбий, деятельность которого приводит к вторичным изменениям и значительному утолщению корня. Камбий образуется между флоэмой и ксилемой из клеток прокамбия. К центру он откладывает клетки, дифференцирующиеся в элементы вторичной ксилемы, а кнаружи - клетки, дифференцирующиеся в элементы вторичной флоэмы. В тех местах, где сосуды первичной ксилемы соприкасаются с перициклом, клетки последнего также превращаются в камбий, который дифференцируется только в паренхиму, образуя 26 радиальные лучи. Первоначально слой камбия на поперечном срезе имеет извилистую форму, а затем округляется и отодвигается от центра. Вместе с увеличением вторичных ксилемы и флоэмы первичная флоэма отодвигается на периферию. Из клеток перицикла возникает пробковый камбий, который образует кнаружи слой пробки. Благодаря этому первичная кора изолируется от центрального цилиндра, отмирает и отпадает. Остается только центральный цилиндр, покрытый пробкой. Таким образом, при вторичном строении в центре корня расположена первичная ксилема. От ее лучей начинаются радиальные лучи - участки тонкостенной живой паренхимы. С радиальными лучами чередуются широкие участки вторичной ксилемы с крупными сосудами и мелкоклеточной древесинной паренхимой. Они окружены камбиальной зоной, состоящей из мелких тонкостенных клеток, расположенных правильными радиальными рядами. Кнаружи от камбиальной зоны, против каждого участка вторичной ксилемы, находится вторичная флоэма. Камбий, производящий паренхиму радиальных лучей, малозаметен. Кнаружи он откладывает также основную паренхиму. Ткани, расположенные кнаружи от камбия (флоэма, основная паренхима, феллодерма и пробковый камбий), называют вторичной корой. Лекция 7 Анатомия стебля Микроскопическое строение. Наземные условия обитания, более разнообразные и контрастные, чем почвенные, обусловили более сложное и многообразное строение стебля по сравнению с корнем. Первичное строение. На верхушке стебля расположена первичная меристема (конус нарастания). На уровне зачаточных листьев закладывается прокамбий, формирующий первичную флоэму и ксилему. Наружу от прокамбия обособляется первичная кора, внутрь - сердцевина. Поверхностный слой клеток конуса нарастания дифференцируется в эпидерму. Так возникает первичное строение. При первичном строении ткани стебля, как и корня, делят на два комплекса: первичную кору, покрытую эпидермой, и центральный цилиндр. Первичную кору составляют следующие ткани: механическая, расположенная под эпидермой; паренхима, значительная часть клеток которой содержит хлоропласта; эндодерма - внутренний слой первичной коры, клетки которого часто содержат крахмальные зерна, и тогда его называют крах мало поеным влагалищем. Наружный слой центрального цилиндра называют перициклом. Он состоит из одного или нескольких рядов паренхимных клеток, из которых могут возникать вторичные 27 меристемы - камбий и феллоген, придаточные корни и почки. Иногда перицикл наряду с паренхимой содержит и склеренхиму (перициклические волокна). Внутрь от перицикла располагаются проводящие ткани, возникающие из прокамбия. В центре расположена сердцевина, состоящая из крупноклеточной тонкостенной паренхимы, где могут откладываться запасные продукты. У голосеменных и двудольных покрытосеменных прокамбий закладывается отдельными тяжами, расположенными по окружности. Следовательно, проводящие ткани при первичном строении также располагаются отдельными пучками по окружности и разделены участками паренхимы, образующими первичные сердцевинные лучи. У однодольных тяжи прокамбия, а также дифференцирующиеся из него сосудистоволокнистые пучки располагаются произвольно по всей толще центрального цилиндра, включая сердцевину. Вторичное строение. Как и у корня, оно связано с возникновением вторичной меристемы - камбия, поэтому бывает только у голосеменных и двудольных покрытосеменных. Камбий закладывается в центральном цилиндре между первичной флоэмой и первичной ксилемой. Он образуется из прокамбия и паренхимы первичных сердцевинных лучей. В центробежном направлении камбий дифференцируется во вторичную кору, состоящую из вторичной флоэмы (вторичного луба) с типичными для нее элементами: ситовидными трубками, сопровождающими клетками, лубяными волокнами, лубяной паренхимой, а также из паренхимы сердцевинных лучей. В центростремительном направлении камбий откладывает вторичную древесину, состоящую из сосудов, трахеид, древесинных волокон, древесинной паренхимы, а также паренхимы сердцевинных лучей. Элементов вторичной древесины обычно откладывается намного больше, чем элементов вторичной коры. В результате деятельности камбия стебель разрастается в толщину. При переходе во вторичное строение первичная кора стебля продолжает функционировать и отмирает не сразу, как у корня. Таким образом, при вторичном строении стебель состоит из эпидермы, или пробки, первичной и вторичной коры, камбия, вторичной и первичной древесины, сердцевины. Структурное разнообразие вторичного строения стеблей обусловлено разнообразием первичной структуры, а также особенностями деятельности камбия. Наиболее широко распространены следующие типы вторичного строения стебля: непучковое, переходное, пучковое. При непучковом строении прокамбий закладывается цилиндром и вскоре преобразуется в камбий. Следовательно, и при первичном, и при вторичном строении элементы флоэмы и ксилемы располагаются также сплошным цилиндром. Этот тип строения свойствен 28 древесным растениям (хвойным и лиственным), а также некоторым травянистым (подмаренник, ипомея, лен и др.). У сосны в центре стебля находится небольшой участок тонкостенных паренхимных клеток - сердцевина. К периферии от нее располагается древесина (ксилема), занимающая большую часть стебля. Она состоит из трахеид, образующих концентрические, слои — годичные кольца. Годичное кольцо возникает из-за периодичности функционирования камбия, который весной дифференцируется в тонкостенные трахеиды с большой полостью, выполняющие проводящую функцию, а летом и осенью - в толстостенные трахеиды с малой полостью, выполняющие в основном механическую функцию. В древесине повсюду, но преимущественно в осенних участках годичных колец, расположены смоляные ходы. Массив трахеид пересекают по радиусу сердцевинные лучи, состоящие из одного ряда живых паренхимных клеток. По ним осуществляется передвижение веществ в горизонтальном направлении. Таким образом, древесина сосны, как и других хвойных, имеет достаточно однородную и поэтому примитивную организацию: ни сосудов, ни древесинных волокон нет, а древесинная паренхима представлена только клетками сердцевинных лучей и эпителиальными клетками смоляных ходов. Между древесиной и вторичной корой расположен камбий. Вторичная кора состоит из вторичной и первичной флоэмы и перициклической зоны. Между ситовидными клетками находятся более крупные округлые клетки лубяной паренхимы. Сердцевинные лучи и во флоэме состоят из одного ряда клеток, однако более крупных, чем в ксилеме. Снаружи от вторичной коры располагаются крупные клетки паренхимы первичной коры, среди которых заметны большие смоляные ходы. Пробку составляют слои клеток с тонкими опробковевающими стенками, чередующиеся со слоями клеток с толстыми одревесневающими стенками. Стебель липы имеет типичное для древесных двудольных растений строение. В центре расположен небольшой участок сердцевины, состоящий из тонкостенной паренхимы. Сердцевина окружена толстым слоем древесины. На границе с сердцевиной древесина образует небольшие выступы. Это участки первичной древесины, состоящие в основном из кольчатых и спиральных сосудов. Вторичная древесина, как и у сосны, имеет годичные кольца, весенние участки которых состоят из сосудов преимущественно с большим диаметром, летне-осенние из сосудов малого диаметра с преобладанием трахеид и древесинных волокон. Вокруг древесины находится камбий, за которым располагаются участки флоэмы в виде трапеций, состоящие из слоев ситовидных трубок с сопровождающими клетками и 29 лубяной паренхимы, чередующихся со слоями лубяных волокон. Между участками флоэмы находятся широкие сердцевинные лучи, сужающиеся в древесине до одного ряда клеток. За флоэмой и сердцевинными лучами - перициклическая зона из чередующихся по кругу групп лубяных волокон (против участков флоэмы) и паренхимы (против сердцевинных лучей). Участки флоэмы, паренхима сердцевинных лучей и перициклическая зона составляют вместе вторичную кору. Снаружи от вторичной коры начинается первичная кора. К перициклической зоне примыкает эндодерма, которая у древесных растений выражена слабо и почти не отличается от следующей за ней паренхимы, состоящей из крупных клеток, в которых нередко имеются друзы оксалата кальция. Снаружи от паренхимы расположены пластинчатая колленхима и перидерма. У некоторых растений (дуб, тис и др.) с возрастом в более старой части древесины накапливаются продукты обмена веществ - дубильные соединения, смолы, камеди, соли, в связи с чем она приобретает темный цвет. Это так называемая ядровая древесина. Светлый же слой древесины, расположенный ближе к коре, называют заболонью. У стебля льна на поверхности расположены крупные клетки эпидермы, покрытые кутикулой. За эпидермой лежит небольшой слой мелких клеток хлорофиллоносной паренхимы коры. Первичная кора заканчивается волнистым рядом более крупных клеток эндодермы. Внутрь от эндодермы находятся плотные группы толстостенных крупных клеток, округлых или многогранных. Это лубяные волокна перициклического происхождения, ради которых и возделывают лен. Далее к центру расположен тонкий слой флоэмы, а за ней - камбий. Вторичная ксилема состоит из крупных пористых сосудов, трахеид и древесинных волокон. На границе с сердцевиной расположены мелкие кольчатые и спиральные сосуды первичной ксилемы. Ксилему пересекают сердцевинные лучи из паренхимных клеток с одревесневающими стенками. Сердцевина состоит из крупноклеточной паренхимы, которая в центре разрушается, образуя полость. При переходном строении прокамбий закладывается отдельными тяжами, первичное строение пучковое. Камбий образуется из прокамбия и паренхимы первичных сердцевинных лучей. Пучковый камбий дифференцируется в элементы вторичной флоэмы и ксилемы. Межпучковый камбий дифференцируется, так же как и пучковый, в элементы флоэмы и ксилемы и образует новые пучки, расположенные между прежними. Постепенно те и другие разрастаются и могут сливаться. В результате деятельности межпучкового камбия в более старой части стебля появляется сплошной слой древесины и луба. Такое строение имеют травянистые растения (подсолнечник, топинамбур, бодяк и др.). 30 У стебля подсолнечника снаружи расположена эпидерма с крупными многоклеточными волосками, под ней находится механическая ткань - колленхима, а под колленхимой небольшой слой паренхимы первичной коры, заканчивающийся извилистым слоем эндодермы. В паренхиме имеются схизогенные смоляные ходы. Таким образом, первичная кора состоит из колленхимы, основной паренхимы, эндодермы. Внутрь от первичной коры расположен центральный цилиндр. Он начинается перициклической зоной, состоящей из участков склеренхимы, чередующихся по кругу с тонкостенной паренхимой. Тяжи склеренхимы расположены не произвольно, а в комплексе с коллатеральными проводящими пучками, они прилегают к флоэмной части пучка. Пучки открытые, расположены равномерно по окружности стебля. Несколько вогнутая зона пучкового камбия, выходя за пределы пучка, формирует выпуклую дугу межпучкового камбия. Широкая зона камбия - свидетельство активной его деятельности. Действительно, за два месяца стебель подсолнечника утолщается в 8 - 10 раз. Межпучковый камбий возникает из паренхимы сердцевинных лучей после того, как из прокамбия сформируются проводящие пучки и начинается деятельность пучкового камбия. Межпучковый камбий образует элементы нового проводящего пучка: ксилему к центру стебля и флоэму к его периферии. Постепенно новые и старые пучки разрастаются и сливаются. В результате деятельности межпучкового камбия образуются сплошной слой ксилемы с острыми выступами, вдающимися в сердцевину, и сплошной слой флоэмы. Крупноклеточная паренхима сердцевины составляет основную массу стебля. При пучковом строении различие заключается в деятельности камбия. Пучковый камбий преобразуется в элементы вторичных флоэмы и ксилемы, межпучковый - только в паренхиму, образующую сердцевинные лучи. Таким образом, и при вторичном строении сохраняется пучковое расположение проводящих тканей. Такое строение можно наблюдать у ряда травянистых растений (кирказон, тыква, чистотел и др.). У стебля кирказона эпидерма покрыта слоем кутикулы. Под ней расположена колленхима, чаще пластинчатая, иногда уголковая. Паренхима состоит из крупных тонкостенных клеток. В некоторых из них имеются кристаллы оксалата кальция в виде друз. Заканчивается первичная кора эндодермой. Клетки наружного слоя центрального цилиндра - склеренхимы перициклического происхождения - на поперечном разрезе многоугольные, плотно прилегают друг к другу, стенки их толстые, пронизаны простыми порами. Внутренняя граница кольца склеренхимы волнистая, над пучками она приподнимается, между пучками опускается. Коллатеральные проводящие пучки расположены в один ряд по кругу. Флоэма отличается от окружающей ее паренхимы более мелкими клетками. Она состоит из ситовидных 31 трубок, сопровождающих клеток и тонкостенной лубяной паренхимы. Первичная флоэма, расположенная в наружной части пучка, деформирована. Вторичная ксилема, образованная камбием, включает сосуды большого диаметра (сетчато-пористые), древесинные волокна и древесинную паренхиму. Первичная ксилема, расположенная на границе с сердцевиной, состоит из небольшого числа кольчатых и спиральных сосудов малого диаметра и трахеид. В пучке между ксилемой и флоэмой лежит камбиальная зона. Пучки разделены первичными сердцевинными лучами. На более поздних фазах роста в паренхиме сердцевинных лучей также образуется камбий. Тяжи межпучкового камбия примыкают к пучковому камбию, образуя сплошной камбиальный цилиндр. Межпучковый камбий дифференцируется только в паренхиму сердцевинных лучей. Сердцевина состоит из рыхло расположенных паренхимных клеток. В некоторых из них также есть друзы. Структура стеблей у травянистых двудольных растений более разнообразная, чем у деревьев и кустарников, что служит показателем их высокой специализации. К особенностям строения стеблей травянистых растений в отличие от древесных надо отнести: отсутствие или слабое развитие феллогена, значительную паренхиматизацию, ослабление деятельности камбия, редукцию механических и отчасти проводящих тканей. Особенности строения стебля однодольных покрытосеменных. Как и корень, стебель имеет лишь первичное строение. Тип строения пучковый. Сосудисто-волокнистые пучки закрытые. На поперечных срезах они как бы беспорядочно рассеяны по всей основной паренхиме. Отчетливая граница между первичной корой и центральным цилиндром часто отсутствует. Для обеспечения механической прочности стебля наряду со склеренхимой пучков служат утолщенные и одревесневающие стенки клеток эпидермы и паренхимы. Хотя у однодольных камбий не образуется, некоторые из них (в основном древесные лилейноцветные) имеют вторичное утолщение за счет возникшего из перицикла кольца утолщения. У стебля ржи под эпидермой расположен слой механической ткани, прерываемый участками хлорофиллоносной паренхимы. Над ней в эпидерме можно заметить устьичные аппараты. В более старых участках стебля стенки клеток хлорофиллоносной паренхимы одревесневают, как и стенки клеток эпидермы. В механической ткани, ближе к границе с основной паренхимой, находятся небольшие проводящие пучки. В основной паренхиме более крупные закрытые коллатеральные проводящие пучки располагаются в шахматном порядке в два, реже в три ряда. Первичная кора не выражена. В центре стебля сердцевина не сохраняется. При увеличении толщины стебля за счет роста клеток она разрывается и 32 образуется полость, свойственная стеблям большинства злаков. Такой стебель называют соломиной. Стебель кукурузы не имеет полости. Он заполнен основной паренхимой, по всей толщине пронизанной проводящими пучками. Под эпидермой расположен тонкий слой механической ткани. Первичная кора, как и у ржи, не выражена. Проводящие пучки закрытые коллатеральные. Во флоэме нет лубяной паренхимы, ситовидные трубки и сопровождающие клетки на поперечном срезе имеют вид сеточки. Ксилема содержит 3 - 5 сосудов, из них два крупных. Под сосудами имеется полость. Ксилема полуобъемлет флоэму. Пучок окружен слоем склеренхимы. 33 Лекция 8 Анатомия листа Микроскопическое строение. Относительно однообразно, определяется основными функциями листа - фотосинтезом, транспирацией, газообменом. Пластинка состоит из эпидермы, мезофилла, проводящих пучков (жилок). У листа бука клетки верхней эпидермы имеют более толстую кутикулу, чем клетки нижней эпидермы. На верхней эпидерме почти полностью отсутствуют устьичные аппараты. Между верхней и нижней эпидермой находится мезофилл, состоящий из ассимиляционной паренхимы. Клетки ее, расположенные у верхней эпидермы, имеют вытянутую форму, плотно сомкнуты, без межклетников. Это столбчатая (палисадная) паренхима. Здесь в основном происходит фотосинтез. У нижней эпидермы расположены более округлые клетки с крупными межклетниками - губчатая паренхима. Главные ее функции: газообмен и транспирация. В мезофилле на некотором расстоянии друг от друга расположены проводящие пучки. Главная жилка занимает почти всю толщу листа от верхней до нижней эпидермы. Ксилема обращена к верхней стороне листа, флоэма - к нижней. Это закрытый коллатеральный пучок. Он укреплен склеренхимой. Выше и ниже пучка расположена колленхима, примыкающая к эпидерме. С увеличением порядка ветвления из пучка постепенно исчезает флоэмная часть, и он становится простым. Таким образом, у листа бука спинная (дорсальная) и брюшная (вентральная) стороны выполняют разные функции и поэтому имеют различное строение. Такие листья называют дорсивентральными. Они свойственны большинству растений. У листа кукурузы клетки верхней эпидермы образуют простые волоски двух видов: короткие шиловидные и длинные нитевидные. У основания длинных волосков клетки эпидермы более крупные, возвышающиеся над поверхностью листа. Эпидерма покрыта кутикулой. Устьичные аппараты имеются и на верхней, и на нижней эпидерме. Проводящие пучки закрытые, коллатеральные, ксилема обращена к верхней стороне листа, флоэма - к нижней. Пучки двух размеров: крупные и мелкие. Каждый пучок окружен округлыми тонкостенными обкладочными клетками. Предполагают, что они играют роль физиологического барьера, регулирующего передвижение веществ подобно эндодерме осевых органов. Мезофилл состоит из более или менее однородных клеток. Вокруг мелких пучков они расположены венцом. В средней утолщенной части пластинки мезофилл имеется лишь у нижней стороны, остальное пространство заполнено крупными клетками, не содержащими хлоропластов. В этой же части листа под эпидермой 34 расположены тяжи одревесневающей склеренхимы, которые на нижней стороне пластинки образуют выступы, доходящие до пучков. В остальной части пластинки субэпидермальные тяжи склеренхимы примыкают с обеих сторон к крупным проводящим пучкам. Таким образом, у листа кукурузы обе стороны выполняют одинаковые функции и имеют одинаковое строение. Это изолатеральный лист. Такая структура свойственна листьям, расположенным более или менее вертикально. Своеобразное строение имеют листья хвойных растений, называемые хвоей. У хвои сосны защитный покров состоит из двух слоев: эпидермы и гиподермы. Эпидерма покрыта толстым слоем кутикулы. Клетки ее в сечении почти квадратной формы, с толстыми стенками. В углублениях на уровне гиподермы на обеих сторонах листа расположены устьичные аппараты, под которыми имеется большая воздушная полость. У старых листьев стенки клеток эпидермы одревесневают. Гиподерма состоит из одного, а в углах - из 2 - 3 рядов клеток с менее утолщенными одревесневающими стенками. Она выполняет также водозапасающую и механическую функции. Под гиподермой находится мезофилл, состоящий из клеток, стенки которых местами образуют складки, заходящие в полость клетки (складчатая паренхима). Это значительно увеличивает площадь прилегающего к стенке слоя цитоплазмы с хлоропластами, а следовательно, и фотосинтезирующую поверхность. Смоляные ходы пронизывают складчатую паренхиму. В центральной части, отделенной от складчатой паренхимы эндодермой, расположены два проводящих пучка коллатерального типа. Ксилемная часть обращена к плоской стороне хвои, флоэмная - к выпуклой. Следовательно, плоская сторона - верхняя, а выпуклая нижняя. Между проводящими пучками расположена механическая ткань. Остальное пространство центральной части занято трансфузионной тканью, вероятно, участвующей в перемещении веществ между проводящими пучками и мезофиллом. Лекция 9 Растительные ткани. Анатомия растительных тканей. Гистология (учение о тканях). Переход растений от сравнительно однообразных условий жизни в водной среде к наземным сопровождался интенсивным процессом расчленения однородного вегетативного тела на органы - стебель, листья и корень. Эти органы состоят из разнообразных по структуре клеток, которые составляют легко различимые группы. Группы однородных по структуре клеток, выполняющие одинаковую функцию и 35 имеющие общее происхождение, называют тканями. Часто несколько тканей, имеющих одинаковое происхождение, образуют комплекс, функционирующий как единое целое. Выделяют шесть основных групп тканей: меристематические (образовательные), покровные, основные, механические, проводящие и выделительные. Меристематические ткани. Растения, в отличие от животных, растут и образуют новые органы на протяжении всей жизни. Это обусловлено наличием меристематических тканей, которые локализованы в определенных местах растения. Меристема состоит из плотно сомкнутых живых клеток. Полость такой клетки заполнена цитоплазмой, в центре располагается крупное ядро, больших вакуолей нет, клеточная стенка очень тонкая, первичная. Клетки меристемы характеризуются двумя основными свойствами: интенсивным делением и дифференциацией, то есть превращением в клетки других тканей. Первичная и вторичная меристема. Первичная меристема возникает в самом начале развития организма. Оплодотворенная яйцеклетка делится и образует зародыш, который состоит из первичной меристемы, вторичная возникает, как правило, позднее из первичной или из клеток уже дифференцированных тканей. Из первичной меристемы образуются первичные ткани, из вторичной - вторичные. По месту расположения различают четыре группы меристем. Верхушечная (апикальная) меристема. Находится на верхушках главных и боковых осей стебля и корня. Она определяет главным образом рост органов в длину. По происхождению она первичная. На верхушке стебля расположена небольшая группа паренхимных клеток (реже - одна клетка), которые довольно быстро делятся. Это инициальные клетки. Ниже лежат производные инициальных клеток, деление которых происходит реже. А еще ниже в меристеме обосабливаются три группы клеток, из которых дифференцируются ткани первичного тела: протодерма - поверхностный слой клеток, дающий начало покровной ткани; прокамбий - удлиненные клетки меристемы с заостренными концами, расположенные вдоль вертикальной оси группами (тяжами), из них образуются проводящие и механические ткани и вторичная меристема (камбий); основная меристема, дающая начало основным тканям. Верхушечная меристема корня имеет несколько иное строение. На верхушке располагаются инициальные клетки, дающие начало трем слоям: дерматогену, дифференцирующемуся в эпиблему; периблеме, дающей начало тканям первичной коры; плероме, дифференцирующейся в ткани центрального цилиндра. 36 Боковая (латеральная) меристема. Располагается цилиндром вдоль осевых органов параллельно их поверхности. Обычно она вторичная. Обусловливает разрастание органов в толщину. Чаще ее называют камбием. Вставочная (интеркалярная) меристема. Закладывается у основания междоузлий побегов, листьев, цветоножек и других органов. Это первичная или вторичная меристема, она определяет рост органов в длину. Раневая (травматическая) меристема. Возникает на любом участке тела растения, где нанесена травма. По происхождению она вторичная. Покровные ткани. Главное назначение покровных тканей - предохранение растения от высыхания и других неблагоприятных воздействий внешней среды. В зависимости от происхождения различают три группы покровных тканей: эпидерму, пробку, корку. Эпидерма. Первичная покровная ткань, которая образуется из протодермы, покрывает листья и молодые стебли. Чаще всего эпидерма состоит из одного слоя живых, плотно сомкнутых клеток. Хлоропластов в них мало или (чаще) нет совсем, и они фотосинтетически малоактивны. Стенки клеток обычно извилистые, благодаря чему достигается прочное соединение их между собой. Толщина стенок неодинакова: наружные, граничащие с внешней средой, более толстые, чем остальные, и покрыты слоем кутикулы. Защитная функция эпидермы усиливается выростами ее клеток (трихомами) - волосками разнообразного строения, чешуйками и др. В эпидерме имеются особые образования для газообмена и транспирации - устьичные аппараты, состоящие из двух замыкающих клеток и межклетника между ними, называемого устьичной щелью. Замыкающие клетки содержат хлоропласты. Стенка их со стороны клеток эпидермы гораздо тоньше, чем со стороны щели. Клетки эпидермы, примыкающие к замыкающим клеткам, часто имеют иную форму, чем остальные. Такие клетки называют побочными. Устьичные аппараты у наземных растений расположены преимущественно на нижней стороне листовой пластинки, а у плавающих листьев водных растений - только на верхней стороне. Пробка. Клетки эпидермы вследствие роста стебля в толщину деформируются и отмирают. К этому времени появляется вторичная покровная ткань - пробка. Ее образование связано с деятельностью вторичной меристемы - пробкового камбия (феллогена), возникающего из субэпидермальных или глубже лежащих клеток, а иногда из клеток эпидермы. Клетки пробкового камбия делятся тангенциально (перегородками, параллельными поверхности стебля) и дифференцируются в центробежном направлении в пробку (феллему), а в центростремительном - в слой живых паренхимных клеток 37 (феллодерму). Комплекс, состоящий из трех тканей: феллогена, феллемы и феллодермы, называют перидермой. Защитную функцию выполняет только пробка. Она состоит из правильных радиальных рядов плотно сомкнутых клеток, на стенках которых откладывается суберин. В результате опробковения стенок содержимое клеток отмирает. Для транспирации и газообмена в пробке имеются особые образования - чечевички, заполненные округлыми клетками, между которыми имеются большие межклетники. Корка (ритидом) образуется у деревьев и кустарников на смену пробке, которая под напором разрастающегося в толщину стебля через 2 - 3 года разрывается. В более глубоколежащих тканях коры закладываются новые участки пробкового камбия, дающие начало новым слоям пробки. Поэтому наружные ткани изолируются от центральной части стебля, деформируются и отмирают. На поверхности стебля образуется комплекс мертвых тканей, состоящий из нескольких слоев пробки и отмерших участков коры. Наружные слои корки постепенно разрушаются. Основные ткани. Под этим названием объединяют ткани, составляющие основную массу различных органов растения. Их называют также выполняющими, основной паренхимой или просто паренхимой. Основная ткань состоит из живых паренхимных клеток с тонкими стенками. Между клетками имеются межклетники. Паренхимные клетки выполняют разнообразные функции: фотосинтез, хранение запасных продуктов, поглощение веществ и др. Выделяют следующие основные ткани. Ассимиляционная, или хлорофиллоносная, паренхима (хлоренхима) расположена в листьях и коре молодых стеблей. Клетки ассимиляционной паренхимы содержат хлоропласты и осуществляют фотосинтез. Запасающая паренхима находится преимущественно в сердцевине стебля и коре корня, а также в органах размножения - семенах, плодах, луковицах, клубнях и др. К запасающей ткани можно отнести также водозапасающую ткань растений засушливых местообитаний (кактусов, алоэ и др.). Поглощающая паренхима наиболее типично представлена во всасывающей зоне корня (зоне корневых волосков). Аэренхима особенно хорошо выражена в подводных органах растений, в воздушных и дыхательных корнях. Она имеет крупные межклетники, соединенные между собой в одну вентиляционную сеть. Механические ткани. Механические ткани в совокупности составляют остов, поддерживающий все органы растения, противодействуя их излому или разрыву. Эти ткани состоят из клеток с 38 толстыми стенками, часто (но не всегда) одревесневающими. Во многих случаях это мертвые клетки. В осевых органах это в основном прозенхимные клетки, в листьях и плодах - паренхимные. В зависимости от формы клеток, химического состава клеточных стенок и способа их утолщения механические ткани подразделяют на три группы: колленхима, склеренхима, склереиды. Колленхима состоит из живых, обычно паренхимных клеток с неравномерно утолщенными целлюлозными стенками. Если утолщения расположены в углах, то такую колленхиму называют уголковой. Если утолщаются две противоположные стенки, а две другие остаются тонкими, колленхиму называют пластинчатой. Стенки клеток колленхимы способны растягиваться, так как имеют тонкие участки, поэтому она служит опорой молодых растущих органов. Колленхима чаще встречается у двудольных растений. Склеренхима состоит из прозенхимных клеток с равномерно утолщенными стенками. Только молодые клетки живые. По мере старения содержимое их отмирает. Это широко распространенная механическая ткань вегетативных органов наземных растений. По химическому составу стенки клетки различают два вида склеренхимы: лубяные волокна стенка целлюлозная или слегка одревесневающая, древесинные волокна (либриформ) стенка всегда одревесневающая. Склереиды. Это мертвые паренхимные клетки с равномерно толстыми одревесневающими стенками. Они обычны в плодах (каменистые клетки), листьях (опорные клетки) и других органах. Проводящие ткани. Растение имеет два полюса питания: листья, осуществляющие воздушное питание, и корни, обеспечивающие почвенное питание. В соответствии с этим существуют два основных пути для транзита питательных веществ: путь, по которому вода и минеральные соли поднимаются от корня по стеблю к листьям, и путь, по которому органические вещества из листьев направляются во все остальные органы растений, где они потребляются или откладываются в запас. Сосуды (трахеи) и трахеиды - проводящие ткани, по которым осуществляется передвижение воды и минеральных солей. Сосуды (трахеи) - трубки, состоящие из члеников. Они дифференцируются из вертикального ряда клеток прокамбия или камбия, у которых утолщаются и одревесневают боковые стенки, отмирает содержимое, а в поперечных стенках образуются одна или несколько перфораций. Средняя длина сосудов 10 см. 39 Трахеиды, как и сосуды - мертвые образования, но в отличие от последних это не трубки, а прозенхимные клетки, в стенках которых имеются окаймленные поры. Длина трахеид в среднем 1 - 10 мм. В зависимости от формы утолщений стенки сосуды и трахеиды бывают кольчатые, спиральные, сетчатые и др.. Кольчатые и спиральные сосуды имеют небольшой диаметр. Они свойственны молодым органам, .так как стенки их имеют неодревесневающие участки и способны растягиваться. Сетчатые и пористые сосуды гораздо большего диаметра, стенки их полностью одревесневают. Они обычно образуются позднее кольчатых и спиральных сосудов из камбия. Сосуды и трахеиды выполняют также и механическую функцию, придавая прочность растению. Они функционируют несколько лет, пока не происходит их закупорка окружающими живыми клетками паренхимы. Выросты последних, проникающие через поры в полость сосуда, называют тиллами. Ситовидные трубки - проводящая ткань, по которой осуществляется передвижение органических веществ, синтезируемых в листьях. Это вертикальный ряд живых клеток (члеников), у которых поперечные стенки пронизаны перфорациями (ситовидные пластинки). Стенка членика ситовидной трубки целлюлозная, ядро разрушается, большинство органелл цитоплазмы деградирует. В протопласте возникают фибриллярные структуры белковой природы (флоэмный белок). Рядом с члеником ситовидной трубки обычно расположены одна или несколько так называемых сопровождающих клеток (клеток-спутниц) имеющих ядро. Наличие большого числа митохондрий в сопровождающих клетках дает основание считать, что они обеспечивают энергией процесс передвижения органических веществ по ситовидным трубкам. Членик ситовидной трубки и прилегающая к нему сопровождающая клетка образуются из одной клетки меристемы вследствие деления ее вертикальной перегородкой. Ситовидные трубки функционируют чаще всего один год. Осенью ситовидные пластинки становятся непроницаемыми для пластических веществ из-за закупоривания перфораций полисахаридом, близким к целлюлозе, - каллозой. По структуре проводящих тканей можно судить об эволюционном уровне растения. Трахеиды - это более примитивные образования, чем сосуды. Среди сосудов более примитивными будут те, у которых концы члеников скошены и имеют несколько перфораций. Одна большая перфорация - прогрессивный признак. Ситовидные трубки с косо поставленными пластинками, имеющими много ситовидных полей, считают примитивными, а с горизонтальными ситовидными пластинками и небольшим числом ситовидных полей - прогрессивными. 40 Сосуды, трахеиды и ситовидные трубки расположены в растениях, как правило, не беспорядочно, а собраны в особые комплексы - ксилему и флоэму. Ксилема (древесина) состоит из сосудов и трахеид, древесинной паренхимы и (не всегда) древесинных волокон (либриформа). По ксилеме передвигаются вода и минеральные вещества. Вторичную ксилему называют древесиной. Флоэма состоит из ситовидных трубок и сопровождающих клеток, лубяной паренхимы и (также не всегда) лубяных волокон. По флоэме передвигаются органические вещества. Вторичную флоэму называют лубом. Ксилема и флоэма, в свою очередь, часто (но не всегда) располагаются внутри органов растения в виде сосудисто-волокнистых, или проводящих, пучков. Если между флоэмой и ксилемой имеется камбий, то такие пучки называют открытыми. Благодаря деятельности камбия образуются новые элементы ксилемы и флоэмы, поэтому пучок со временем разрастается. Открытые пучки свойственны двудольным. В закрытых пучках между флоэмой и ксилемой камбия нет, поэтому разрастания не происходит. Закрытые пучки имеют однодольные и, как исключение, некоторые двудольные, у которых камбий очень рано перестает функционировать (например, у видов рода лютик). Пучки также классифицируют по взаимному расположению флоэмы и ксилемы. Коллатеральный - флоэма и ксилема располагаются бок о бок, причем флоэма обращена к периферии осевого органа, а ксилема - к центру. Биколлатеральный - флоэма прилегает к ксилеме с обеих сторон, наружный участок флоэмы больше, чем внутренний; свойствен тыквенным, пасленовым, вьюнковым. Концентрический бывает двух видов: ксилема окружает флоэму - амфивазальный (в основном у однодольных); флоэма окружает ксилему - амфикрибральный (у папоротников). Радиальный - ксилема расположена в центре, образует к периферии радиальные выступы, чередующиеся с участками флоэмы, бывает только в корнях при первичном строении. По числу выступов ксилемы различают радиальные пучки диархные (2 выступа), триархные (3 выступа), тетрархные (4 выступа) и полиархные (более 4 выступов). Выделительные ткани. Растения не имеют специализированных органов выделения, но все же экскреторные вещества у них так или иначе удаляются из организма или накапливаются в особых вместилищах. Поэтому выделительные ткани можно разделить на две группы: внутренней и внешней секреции. К первым относят млечники, одиночные выделительные клетки, схизогенные и лизигенные вместилища. Продукты внутренней секреции - дубильные вещества, смолы, эфирные масла и др. Ко вторым относят железистые волоски и желѐзки, 41 расположенные на поверхности органов. Продукты внешней секреции - эфирные масла, нектар, вода и др. Млечники - это живые клетки с цитоплазмой, множеством ядер и вакуолью, заполненной млечным соком (латексом). Стенка их состоит из целлюлозы. Различают два вида млечников: членистые и нечленистые. Членистые млечники образуются так же, как и сосуды, в результате разрушения поперечных стенок у вертикального ряда клеток, нечленистые возникают в результате разрастания специальных клеток зародыша. Это гигантские цилиндрические или разветвленные клетки. Млечники располагаются только во флоэме, или пронизывают весь орган (стебель, корень, лист). Они выполняют не только выделительную, но также проводящую и запасающую функции. Проводящая функция осуществляется тогда, когда млечники соединяют места синтеза органических веществ, например листья, с местами их потребления. О выделительной и запасающей функциях млечников можно судить по составу латекса. У разных растений состав латекса очень разнообразен. Из экскреторных веществ он часто содержит каучук, танниды, алкалоиды, смолы и др., из запасных - крахмал, сахара, белки, масло и др. Иногда в латексе имеются ферменты. Цвет латекса белый или оранжево-красный. Млечники присущи лишь некоторым группам растений, например части сложноцветных, маковым, молочайным и др. Выделительные клетки рассеяны среди клеток других тканей. По мере накопления экскрета протопласт их отмирает, а на стенку клетки изнутри откладывается суберин, изолирующий ядовитые вещества от окружающих клеток. Выделительные клетки имеются в листьях чая, лавра, в корневищах бадана и др. Схизогенные и лизигенные вместилища служат для накопления и длительного хранения многих конечных продуктов жизнедеятельности, выключенных из обмена веществ. Они имеют округлую или каналовидную форму и различную величину. Схизогенные вместилища формируются из межклетников, возникающих в результате раздвигания клеток. Прилегающие к вместилищу живые клетки становятся эпителиальными и выделяют в полость вместилища экскреторные вещества. Схизогенные вместилища бывают у весьма разнообразных групп растений, но особенно характерны для голосеменных, у которых в смоляных каналах накапливается бальзам - раствор смол в эфирных маслах. Лизигенные вместилища образуются в результате растворения группы клеток, заполненных экскреторными веществами. Они характерны для цитрусовых. Железистые волоски образуются из клеток эпидермы. В них накапливаются и выводятся из тела растения во внешнюю среду разнообразные экскреторные вещества в 42 газообразном, жидком и твердом виде. По строению они очень разнообразны, но сохраняют постоянство структуры для каждой группы растений. Нектарии, или нектарники - желѐзки, выделяющие на поверхность органа раствор углеводов. Сахаристые выделения нектариев цветка привлекают насекомых-опылителей. Гидатоды - это желѐзки, выделяющие наружу через устьичные аппараты воду. Замыкающие клетки таких аппаратов лишены подвижности, устьичные щели постоянно открыты. Они сосредоточены по краю листа, преимущественно на верхушках зубчиков. Лекция 10 Строение генеративных органов растений Цветок Представляет собой видоизмененный укороченный побег, приспособленный для образования спор и гамет, для полового процесса, в результате которого образуются семена и плод. Стеблевая часть цветка представлена цветоножкой и цветоложем. Цветоложе может иметь разнообразную форму - от конической до плоской и даже вогнутой в виде бокала. .На цветоложе размещены видоизмененные листья (цветолистики): чашелистики, лепестки, тычинки, пестики. Чаще они располагаются кругами. Такие цветки называют циклическими. Обычно бывает пять или четыре круга: один круг чашелистиков, один лепестков, два или один - тычинок, один - пестиков. При спиральном расположении цветолистиков цветок называют ациклическим. Промежуточное положение занимают цветки гемициклические, у которых круговое расположение одних цветолистиков сочетается со спиральным других. Цветки, имеющие и тычинки и пестики, называют обоеполыми (гермафродитными). Раздельнополые цветки содержат или только тычинки, или только пестики. Соответственно различают цветки тычиночные и пестичные. Растения с тычиночными и пестичными цветками на одном экземпляре называют однодомными (кукуруза). Растения, у которых одни экземпляры несут только тычиночные цветки, а другие - только пестичные, называют двудомными (конопля). Околоцветник - это стерильная (бесплодная, не имеющая способности к оплодотворению) часть цветка, выполняющая защитную функцию, а также функцию привлечения опылителей. Околоцветник, дифференцированный на чашечку и венчик, называют двойным, а не дифференцированный на чашечку и венчик - простым. Простой чашвчковидный околоцветник имеет обычно зеленый цвет, простой венчиковидный окрашен ярко. Цветки, не имеющие околоцветника, называют голыми. 43 Чашечка. Обычно состоит из небольших зеленых цветолистиков - чашелистиков. Иногда чашелистики ярко окрашены, тогда они выполняют или усиливают роль лепестков в привлечении насекомых - опылителей. Чаще чашелистики образуют один круг. Иногда же под чашечкой расположен .круг листовидных образований, именуемый подчашием. Подчашие образуется из прицветников или прилистников. Чашелистики бывают свободными (чашечка свободнолистная, или раздельнолистная) или с более или менее сросшимися основаниями (чашечка сростнолистная, или спайнолистная). Венчик обычно состоит из цветолистиков более крупных, чем чашелистики, и ярко окрашенных - лепестков. Лепестки могут срастаться между собой, хотя бы частично, или оставаться свободными. Различают два типа венчиков: свободнолепестные (раздельнолепестные) и сростнолепестные (спайнолепестные). Если в свободнолепестном венчике лепесток сужен к основанию, как лист в черешок, то это ноготковый лепесток. Если основание широкое, лепесток называют сидячим. Чаще встречаются промежуточные формы лепестков. В сростнолепестном венчике различают трубковидно сросшуюся нижнюю часть венчика трубку, верхнюю часть венчика, обычно расширенную и отогнутую, раположенную перпендикулярно к трубке - отгиб и границу между трубкой и отгибом - зев. Венчик, как и чашечка, может быть: актиноморфным (правильным, полисимметричным), когда через него можно провести две или больше плоскостей симметрии, зигоморфным (неправильным, моносимметричным), когда можно провести только одну плоскость симметрии, асимметричным, когда нельзя провести ни одной плоскости симметрии (канна, орхидеи). Актиноморфные сростнолепестные венчики классифицируют по длине трубки, форме и величине отгиба: колесовидный - трубка мала или почти отсутствует, а отгиб развернут в плоскость; воронковидный - трубка крупная, воронковидная, отгиб сравнительно небольшой; колокольчатый - трубка сферическая, чашеобразная, постепенно переходящая в малозаметный отгиб; трубковидный - трубка цилиндрическая, без отгиба или с коротким отгибом; блюдцевидный - трубка цилиндрическая, с широким отгибом; колпачковый - лепестки срастаются верхушками (виноград). Среди зигоморфных сростнолепестных венчиков наиболее часто встречают следующие: двугубый - отгиб состоит из двух неодинаковых губ: верхней и нижней; язычковый - от трубки отходят сросшиеся лепестки, имеющие вид язычка; 44 шпористый - лепестки образуют полый вырост, именуемый шпорцем; бывают и у свободнолепестных зигоморфных и актиноморфных венчиков. Андроцей - это совокупность тычинок одного цветка. Число тычинок в андроцее колеблется в больших пределах - от одной до нескольких сотен. Тычинки могут быть или свободными (свободный андроцей), или в различной степени сросшимися (сросшийся андроцей). Если срастаются все тычинки, то их называют однобратственными, если одна тычинка остается свободной - двубратственными, и если тычинки срастаются в несколько групп - многобратственными. По высоте относительно друг друга тычинки могут быть одинаковыми или разными - двусильными, когда две тычинки выше других, трехсильными и т. д.. Тычинка. В типичном случае тычинка состоит из тычиночной нити и пыльника. Тычиночные нити у большинства растений простые, неветвящиеся. Но иногда они имеют боковые выросты разнообразной формы или ветвятся. Если тычиночная нить отсутствует, то тычинку называют сидячей. Пыльник обычно состоит из двух половин, называемых теками, соединенных между собой связником. Иногда связник рассматривают как самостоятельную третью часть тычинки, однако под микроскопом ткань его неотличима от ткани пыльника. В каждой из тек находится по два гнезда, или пыльцевых мешка. Снаружи пыльника имеется однослойная эпидерма. Это все, что осталось от микроспорофилла. Глубже расположенные ткани принадлежат микроспорангию. Субэпидермальный слой называют эндотецием. Клетки его рано теряют живое содержимое, а стенки на обращенной внутрь стороне клетки несут фиброзные пояски, способствующие вскрытию пыльника. За эндотецием в молодом пыльнике расположены средний слой и выстилающий слой, или тапетум. Клетки тапетума крупные, с густой цитоплазмой и с несколькими ядрами. В центре каждого гнезда имеется спорогенная ткань, из клеток котором образуются микроспоры, а затем пыльца. По мере созревания пыльника клетки тапетума и среднего сдоя исчезают, так как содержимое их идет на питание растущих микроспор и пыльцы. Строение пыльцы довольно однообразно, но структура ее покрова - спородермы - отличается разнообразием. У некоторых видов растений часть тычинок не имеет пыльников и представлена лишь тычиночными нитями. Такие бесплодные тычинки называют стаминодиями (лен). Иногда стаминодии имеют вид окрашенных лепестковидных пластинок (у канны). Гинецей - это совокупность плодолистиков (мегаспорофиллов) одного цветка, образующих один или несколько пестиков. Гинецей, состоящий из одного плодолистика, образующего один пестик, называют одночленным, а из нескольких плодолистиков многочленным. Многочленный гинецей может быть апокарпным, если плодолистики не 45 срастаются между собой и образуют несколько пестиков, и ценокарпным, если плодолистики срастаются в один пестик. Ценокарпный гинецей состоит из стольких плодолистиков, сколько отдельных столбиков несет завязь или лопастей - рыльце и сколько гнезд или швов имеет завязь. При этом надо учитывать всю совокупность перечисленных признаков, так как один из них не всегда точно указывает на число плодолистиков. Пестик. Это закрытое вместилище семязачатков (семяпочек), образовавшееся вследствие срастания краев одного или нескольких плодолистиков. Пестик состоит из завязи, вздутой части, где находятся семязачатки, столбика и рыльца, воспринимающего пыльцу. Иногда столбик отсутствует, тогда рыльце сидячее. Завязь в зависимости от положения по отношению к другим частям цветка бывает верхняя и нижняя. Верхняя завязь располагается свободно на плоском, выпуклом или вогнутом цветоложе и образуется только плодолистиками. Ее можно легко отделить от цветоложа препаровальной иглой. В образовании нижней завязи, кроме плодолистиков, принимают участие и другие части цветка - чаще основания чашелистиков, лепестков и тычинок, реже цветоложе, с которыми она срастается. Поэтому отделить иглой такую завязь нельзя. Выделяют еще полунижнюю завязь, когда, по крайней мере, нижняя половина ее срастается с другими частями цветка. В зависимости от числа не сообщающихся между собой гнезд завязи бывают одногнездными (если гнезда так или иначе сообщаются между собой), двугнездными и многогнездными. Внутри завязи образуются семязачатки. Место прикрепления семязачатка к стенке завязи называют плацентой (семяносцем). Семязачаток прикрепляется к плаценте фуникулусом (семяножкой). Семязачаток снаружи имеет два покрова - интегумента, которые на верхушке не смыкаются, оставляя отверстие - микропиле (пыльцевход). Основание семязачатка, называемое халазой, находится на противоположной микропиле стороне. Под интегументами сформированного семязачатка расположена многоклеточная ткань нуцеллус (мегаспорангий). Одна из его клеток становится археспориальной. Из нее образуются четыре мегаспоры. Одна мегаспора дает начало восьмиядерному зародышевому мешку. Формула и диаграмма цветка. Строение цветка можно выразить в виде формулы. При ее составлении пользуются следующими обозначениями: чашечка (calyx) - Са, венчик (corolla) - Со, андроцей (androeceum) - А, гинецей (gynoeceum) -, простой околоцветник (perigonium) - P. 46 Типы цветков имеют также условные обозначения:, пестичный -♀, тычиночный - ♂ (обозначение обоеполого цветка в формуле обычно опускают), актиноморфный - *, зигоморфный - ↑. Число членов каждой части цветка обозначают цифрами (5-лепестный венчик Co5, 6членный андроцей — А6) , а в том случае, если их число в цветках одного и того же вида непостоянно (обычно больше 12) - значком ∞. В случае срастания между собой цветолистиков число заключают в скобки: сросшийся 5членный венчик - Со(5), двубратственный андроцей - А(9) + 1. Если чашечка, венчик, андроцей расположены несколькими кругами, то цифры, указывающие на число членов в отдельных кругах, соединяют значком +. Формула должна отражать число плодолистиков, образовавших гинецей, и если их несколько, то срослись ли они между собой (ценокарпный гинецей) или каждый из плодолистиков образовал отдельный пестик (апокарпный гинецей), а также какая завязь верхняя или нижняя: ценокарпный гинецей из трех плодолистиков с верхней завязью G (3), ценокарпный гинецей из трех плодолистиков с нижней завязью G(3). Таким образом, строение цветка калужницы можно выразить формулой *Р5А∞G∞ , яблони - *Са(5)Со5А∞G(5), гороха - ↑Са(5)Со3+(2)А(9),1G1. Еще более полное представление о строении цветка дает диаграмма, которая служит проекцией цветка на плоскость, перпендикулярную к его оси. Диаграммы демонстрируют не только наличие частей цветка и число членов, но и расположение их по отношению друг к другу. Для удобства принят единый способ ориентации диаграммы: ось соцветия вверху, а кроющий лист внизу. Члены цветка обозначают всегда строго определенными фигурами. В случае срастания между собой отдельных членов фигуры, обозначающие их на диаграмме, соединяются дугами или прямыми линиями. Соцветие. Соцветие - это побег или система побегов, несущих цветки. На узлах осей соцветия располагаются такие же листья, как на вегетативной части побега, или видоизмененные, утратившие способность к фотосинтезу, - прицветники, а на узлах цветоножки прицветнички. Биологическое преимущество соцветий перед одиночными цветками несомненно. Оно заключается в повышении гарантии опыления, в уменьшении вероятности повреждения цветков неблагоприятными факторами среды, обусловленное распусканием. Соцветия имеют большинство растений. 47 их постепенным Классификация. Различают два типа соцветий: сложные, когда цветки располагаются на разветвлениях главной оси, и простые, когда цветки с цветоножками или без них располагаются непосредственно на главной оси. Сложные соцветия в зависимости от способа нарастания осей делят на: симподиальные (определенные) - ось заканчивается цветком, распускание цветков идет от верхушки к боковым ветвям или центробежно, если цветки расположены в одной плоскости; и моноподиальные (неопределенные) - ось нарастает неопределенно долго, расцветание цветков идет от основания к верхушке или центростремительно, если цветки расположены в одной плоскости. Сложные симподиальные соцветия: монохазий - главная ось заканчивается цветком; под ним образуется ось второго порядка, также увенчанная цветком, и т.д.; если подцветочные оси отходят в.одну сторону, то образуется завиток, если же попеременно то в одну, то в другую сторону - образуется извилина, завиток, у которого боковые разветвления укорочены, называют клубочком; дихазий - под цветком образуются две супротивные оси, каждая из которых также заканчивается цветком и также дает две подцветочные оси, повторяющие такой же способ ветвления, и т.д.; плейохазий - от главной оси, несущей один верхушечный цветок, отходит больше двух подцветочных осей, образующих мутовку из монохазиев или дихазиев; тирс - на главной оси располагаются указанные выше соцветия, чаще имеет пирамидальную форму. Сложные моноподиальные соцветия: метелка - очень разветвленное соцветие, нижние боковые разветвления ветвятся сильнее, чем верхние; щиток - метелка, у которой цветки расположены более или менее в одной плоскости; сложный колос - главная ось образует разветвления, на которых расположены цветки без цветоножек; эти разветвления называют колосками; сложный зонтик - расстояния между осями второго порядка укорочены, и они отходят от верхушки оси первого порядка; расстояния между цветоножками укорочены, и они расположены на верхушке оси второго порядка; нередко листья у основания осей второго порядка образуют общую обвертку, а у основания цветоножек - частную обвертку. 48 Кроме перечисленных, имеются сложные соцветия, называемые агрегатными. Они образованы сочетанием различных типов соцветий. Простые соцветия с удлиненной осью: кисть - главная ось несет цветки с цветоножками, обычно одинаковой длины; колос - главная ось несет цветки без цветоножек; сережка - повислый колос, то есть колос с мягкой осью; после цветения соцветие обычно опадает; початок - колос с сильно утолщенной осью, окружен одним или несколькими листьями, так называемым покрывалом, или крылом. Простые соцветия с укороченной осью: зонтик - соцветие, у которого главная ось укорочена, и цветоножки, имеющие почти одинаковую длину, отходят от верхушки оси; головка - зонтик, у которого цветки или без цветоножек, или очень короткие; корзинка - верхушка главной оси разрастается в виде ложа, на котором расположены плотно сомкнутые цветки; верху¬шечные листья скучены и образуют обвертку. Семя Это орган, предназначенный для размножения и распространения семенных растений. Оно состоит из зародыша и запасающей ткани, покрытых спермодермой. В неблагоприятных для прорастания условиях (холод, засуха) семя может значительное время пребывать в состоянии покоя. С наступлением благоприятных условий температуры и влажности семя поглощает воду и при достаточном доступе воздуха прорастает. Классификация семян. Различают пять типов семян в зависимости от того, где откладываются запасные продукты: в эндосперме, нуцеллусе, зародыше, эндосперме и нуцеллусе, эндосперме и зародыше. Семена с эндоспермом. У зерновки овса снаружи расположен тонкий пленчатый слой, который трудно отделить от внутренней части зерновки. Это околоплодник, сросшийся с спермодермой (зерновка - односемянный плод). Внутри находятся маленький зародыш и эндосперм. Зародыш состоит из первичной меристемы и имеет зачатки вегетативных органов будущего растения: зародышевый корешок с корневым чехликом, корневое влагалище - колеоризу, зародышевый стебелек (гипокотиль) и почечку, в которой хорошо заметен конус нарастания стебля, прикрытый зародышевыми листьями. Наружный зародышевый лист называют колеоптилем. Он защищает почечку при прохождении через почву. Единственная семядоля прилегает к эндосперму, ее называют щитком. На 49 стебельке со стороны, противоположной щитку, расположен эпибласт, представляющий собой вторую редуцированную семядолю (у некоторых злаков эпибласт не образуется). Эндосперм в периферической части имеет ряд клеток, содержащих гранулы белка алейроновые зерна. Это алейроновый слой. Клетки под ним (в центре зерновки) заполнены сложными крахмальными зернами. Семена с запасными продуктами в зародыше. Примером семени с запасными продуктами в зародыше служит семя фасоли. Снаружи имеется довольно толстая спермодерма. На узкой вогнутой стороне семени находится рубчик - место прикрепления семени к семяножке. На одной линии с рубчиком расположено микропиле, через которое вода и газы поступают внутрь семени. Небольшой бугорок над микропиле образован зародышевым корешком. С противоположной от микропиле стороны к рубчику примыкает семенной шов - след от срастания семязачатка с семяножкой. Под спермодермой находится зародыш, состоящий из двух крупных семядолей почковидной формы, зародышевого корешка, зародышевого стебелька и почечки. Эндосперма в семени нет. Питательные продукты эндосперма были поглощены зародышем. Клетки семядолей заполнены крахмальными и алейроновыми зернами. Плод Это орган, предназначенный для защиты семян, а нередко и для их распространения. Плод образуется из цветка в результате изменений, происходящих с ним после оплодотворения. В образовании плода главную роль играет гинецей. У некоторых растений плод образуется без предшествующего оплодотворения. Такие плоды называют партенокарпическими, они обычно не содержат семян (некоторые сорта винограда, груши, цитрусовых). В отличие от плодов соплодия образуются из нескольких цветков, сросшихся между собой (свекла), или из всего соцветия (шелковица, инжир, ананас). Строение. Плод состоит из околоплодника (перикарпа) и семян. Околоплодник - это стенка плода, которая формируется из стенки завязи, а часто и из других частей цветка оснований тычинок, лепестков, чашелистиков, реже цветоложа. Околоплодник состоит из трех слоев: наружного - экзокарпа, среднего - мезокарпа и внутреннего - эндокарпа. Классификация. Разнообразие плодов очень велико. Оно вызвано большим многообразием систематических групп покрытосеменных и приспособлением плодов к агентам их распространения. Это очень усложняет создание общей филогенетической классификации плодов. Существующая филогенетическая классификация основана на типе гинецея. Плоды, образованные из примитивного апокарпного гинецея, называют апокарпиями, а из эволюционно продвинутого ценокарпного - ценокарпиями. К 50 сожалению, дальнейшая классификация этих двух трупп очень сложна и неприменима для определения растений. Поэтому приходится придерживаться морфологической классификации плодов, явно искусственной. Плод называют простым, если в его образовании принимает участие только один пестик. Иногда такой плод распадается по гнездам на части, называемые мерикарпиями (дробный), или разламывается по поперечным (ложным) перегородкам на односемянные членики (членистый). Плод, образованный несколькими пестиками одного цветка, называют сборным (сложным). В основу дальнейшей классификации простых и сборных плодов положены следующие признаки: консистенция околоплодника (сухой или сочный), число семян (много или одно), вскрывание околоплодника (нераскрывающийся или раскрывающийся, способ раскрывания), число плодолистиков, образующих плод. Выделяют следующие группы плодов. Коробочковидные плоды. Это плоды с сухим околоплодником, многосемянные, обычно раскрывающиеся: листовка - одногнездный плод, образованный одним плодолистиком, вскрывается одной щелью по брюшному шву (линии срастания краев плодолистика); из апокарпного гинецея образуется сборная листовка; боб - одногнездный плод, образованный одним плодолистиком, вскрывается двумя щелями: по брюшному шву и по средней жилке плодолистика; бобы могут быть спирально закрученные, односемянные, нераскрывающиеся, членистые; стручок, стручочек - двухгнездный плод, образованный двумя плодолистиками, семена прикрепляются к продольной перегородке, вскрывается двумя швами; стручочек отличается от стручка соотношением длины и ширины: если у стручка длина превышает ширину в четыре и более раз, то у стручочка длина превышает ширину не более чем в 2 3 раза; стручки могут быть членистыми; коробочка - плод, образованный несколькими плодолистиками; существуют различные способы вскрывания коробочки: дырочками, крышечкой, зубчиками, створками и т. д. Ореховидные плоды. Это плоды с сухим околоплодником, односемянные, нераскрывающиеся: орех, орешек - околоплодник жесткий, деревянистый; орешек отличается от ореха меньшим размером; из апокарпного гинецея образуется сборный орешек; желудь - околоплодник менее жесткий, чем у ореха, у основания плод окружен чашевидной плюской (защитным покровом); семянка - околоплодник кожистый, не слипается со спермодермой; 51 крылатка - семянка, околоплодник которой имеет кожистый или перепончатый крыловидный вырост; зерновка - околоплодник кожистый, слипшийся со спермодермой. Ягодовидные плоды. Это плоды с сочным околоплодником, большей частью многосемянные: ягода - околоплодник, за исключением тонкого экзокарпа, сочный, мясистый; яблоко - в образовании его, кроме завязи, принимают участие нижние части тычинок, лепестков, чашелистиков, а также цветоложе; тыквина - образуется из нижней завязи, состоящей из трех плодолистиков; экзокарп жесткий, деревянистый, мякоть плода образована в основном разросшимися плацентами; гесперидий, или померанец - экзокарп с вместилищами эфирного масла; мезокарп сухой, губчатый, белый; эндокарп сочный, мясистый. Костянковидные плоды. К ним относятся плоды с деревянистым эндокарпом, чаще односемянные: костянка - околоплодник дифференцирован на тонкий экзокарп, мясистый мезокарп и более или менее толстый деревянистый эндокарп; из апокарпного гинецея образуется сборная костянка; изредка бывает сухая костянка. Распространение плодов и семян. Плоды и семена многих растений распространяются воздушными течениями (анемохория), водой (гидрохория), животными (зоохория), людьми (антропохория). В процессе эволюции у плодов и семян возникло множество приспособлений, способствующих распространению определенным агентом: крылатые выросты (клен, береза), волоски (тополь, хлопчатник), воздушные полости (лотос), крючки (дурнишник), щетинки (череда), шипы (якорцы) и др. Плоды с сочным околоплодником поедают животные, но семена не перевариваются. У некоторых растений (вика, недотрога) плоды при вскрывании разбрасывают семена. Однако расстояние, на которое отбрасываются семена, обычно не превышает 15 см, поэтому такой способ распространения значительно уступает другим. Лекция 11 Размножение растений Микроспорогенез и образование пыльцы. Центральную часть гнезда пыльника занимают спорогенные клетки. Они делятся путем мейоза, и в результате из каждой клетки образуются четыре гаплоидные микроспоры. Микроспора имеет плотную цитоплазму и крупное ядро. 52 Микроспоры обычно еще внутри пыльника прорастают и образуют пыльцу (пыльцевые зерна). При прорастании микроспора делится путем митоза. В результате образуются маленькая генеративная клетка и большая клетка-трубка. Снаружи пыльцевое зерно покрыто спородермой, состоящей из двух слоев: наружного - экзины и внутреннего интины. Экзина состоит в основном из спорополленина, отличающегося необычайной стойкостью, и имеет одну или несколько апертур (пор) - более тонких мест, служащих для выхода пыльцевой трубки. Интина состоит из пектина и целлюлозы, тонка, эластична и легко разрушается. Мегаспорогенез и образование зародышевого мешка. Одна из клеток нуцеллуса, называемая археспориальной, делится путем мейоза и образует четыре гаплоидные мегаспоры. В дальнейшем три мегаспоры отмирают и только одна, обычно обращенная к халазе, сохраняется. Мегаспора, как и микроспора, всегда одноядерна. Она увеличивается в размере и прорастает. Ядро претерпевает три последовательных митотических деления. После первого деления образуются два ядра, которые перемещаются к противоположным полюсам, где осуществляются два последующих деления. В результате на каждом из полюсов образуется по четыре ядра. Все восемь ядер, как и исходное, гаплоидны, т. е, набор их хромосом равен n. Три ядра у каждого полюса обособляются в клетки. На микропилярном полюсе одна из клеток становится женской гаметой - яйцеклеткой, две другие - синергидами. Три клетки на халазальном полюсе называют антиподами.. Два оставшихся ядра мигрируют к центру и спустя более или менее продолжительное время сливаются, образуя диплоидное вторичное ядро (2n) центральной клетки. Так формируется зародышевый мешок. Опыление. После созревания пыльцы пыльники лопаются и пыльца попадает на рыльце пестика. Этот процесс называют опылением. Различают два способа опыления: самоопыление и перекрестное опыление. Самоопыление может осуществляться только в обоеполых цветках. Оно наиболее хорошо выражено у нераскрывающихся (клейстогамных) цветков. Перекрестное опыление в ходе эволюции оказалось более прогрессивным. При перекрестном опылении потомство может совмещать наследственные свойства обоих родителей, а это открывает более широкие возможности в приспособлении к разным условиям существования. Подавляющее большинство растений имеет перекрестное опыление. По способу переноса пыльцы различают: анемофилию (ветром), гидрофилию (водой), энтомофилию (насекомыми), орнитофилию (птицами), мирмекофилию (муравьями) и др. Хорошо известны приспособления цветков к определенным насекомым53 опылителям. Так, виды клевера имеют венчик, сросшийся в длинную трубку, и только шмели и некоторые длиннохоботковые пчелы способны доставать из нее нектар и осуществлять перекрестное опыление. При этом территории естественного распространения (ареалы) клевера и этих насекомых совпадают. Когда клевер впервые был ввезен в качестве сельскохозяйственной культуры в Австралию и Новую Зеландию, где не было этих насекомых, опыления не происходило и семена почти не образовывались. Для получения нормальных урожаев семян потребовалось завезти шмелей. Различают две формы перекрестного опыления: гейтеногамию (соседнее опыление) , когда оно осуществляется в пределах одного растения, но пыльца с одного цветка попадает на рыльце другого; и ксеногамию (собственно перекрестное), когда пыльца с цветка одной особи переносится на рыльце цветка другой особи. Для предотвращения самоопыления у растений выработались разнообразные приспособления. Наиболее широко распространены дихогамия и гетеростилия. Дихогамия - это разновременное созревание тычинок и пестиков в обоеполом цветке. При гетеростилии у одних экземпляров в обоеполых цветках пестики с длинными столбиками, а тычинки с короткими тычиночными нитями, у других - наоборот. Оплодотворение. Это процесс слияния двух половых клеток - мужской и женской гамет. Мужскую гамету у покрытосеменных называют спермием, женскую - яйцеклеткой. Клетка-трубка пыльцы, попавшей на рыльце пестика, вытягивается через пору экзины в длинную пыльцевую трубку, достигающую иногда нескольких сантиметров. В ней за счет деления генеративной клетки образуются два спермия. Пыльцевая трубка растет сквозь рыхлую ткань рыльца и столбика в направлении завязи пестика, входит в семязачаток через микропиле или сквозь покровы и вступает в контакт с одной из синергид. Затем пыльцевая трубка проникает внутрь синергиды и лопается, освобождая спермин. Один из них сливается с яйцеклеткой, образуя зиготу (2n), второй сливается с центральной клеткой зародышевого мешка, образуя триплоидную клетку (Зn). Так происходит двойное оплодотворение, свойственное только покрытосеменным. Открытие двойного оплодотворения принадлежит русскому ученому академику С.Г. Навашину (1898 г.). После двойного оплодотворения из зиготы образуется зародыш (2n): из триплоидной клетки - эндосперм (запасающая ткань, Зn), из интегументов - спермодерма (семенная кожура), а из всего семязачатка - семя. Синергиды и антиподы обычно разрушаются, а нуцеллус используется как питательный продукт при формировании зародыша, реже он превращается в запасающую ткань - перисперм. 54 Апомиксис. Апомиксисом называют разнообразные случаи развития зародыша без оплодотворения. В этом случае при образовании зародышевого мешка мейоз не происходит и все его клетки диплоидны. При апомиксисе зародыш может образоваться из яйцеклетки (партеногенез), из любой клетки зародышевого мешка, кроме яйцеклетки (апогамия), из клетки нуцеллуса, интегумента, халазы (апоспория). С апоспорией связана полиэмбриония многозародышевость семени. Лекция 12 Систематика растений Жизненная форма (биоморфа) - это внешний облик растения, сложившийся на уровне популяции под влиянием условий среды обитания. Морфофизиологическое обособление жизненной формы происходит под контролем генетического кода. Жизненная форма служит этапом (квантом) формообразовательного процесса - эволюционного морфогенеза. Учение о жизненных формах составляет крупный раздел морфологии. Впервые этот термин появился в качестве научного в работах Йох. Варминга (1884 г.) - основателя экологической географии растений. По внешнему виду (габитусу) и продолжительности жизни выделяют следующие жизненные формы: деревья - многолетние растения с одревесневающими надземными частями, с хорошо выраженным одним стволом, высотой не ниже 2 м; кустарники - многолетние растения с одревесневающими надземными частями, в отличие от деревьев имеют несколько стволов, так как ветвление начинается от основания стебля; кустарнички - сходны с кустарниками, но не выше 50 см; полукустарники - отличаются от кустарников тем, что у них одревесневают только нижние части побегов, а верхние ежегодно отмирают; лианы - растения с вьющимися, цепляющимися, лазающими стеблями; суккуленты - многолетние растения с сочными побегами, содержащими запас воды; травы - надземная часть ежегодно отмирает; у многолетних трав под землей сохраняются корневища, луковицы, клубни, у однолетнихотмирает и подземная часть. В 1903 г. датский ученый К. Раункиер, обобщив имеющиеся знания, создал систему жизненных форм покрытосеменных, широко распространенную до сих пор. Система эта выгодно отличается от других удивительной простотой построения и последовательностью. В основу ее положен не внешний облик растений, а важные 55 морфофизиологические особенности. Физиологический принцип ее - реакции растений на сезон покоя; морфологический - положение почек возобновления по отношению к поверхности почвы и способы их защиты в течение неблагоприятного времени года. В сокращенном виде классификация жизненных форм по Раункиеру выглядит следующим образом: фанерофиты - растения с почками возобновления, находящимися выше 25 см над поверхностью почвы; хамефиты - растения с низко расположенными почками возобновления (не выше 25 см над поверхностью почвы); гемикриптофиты - растения с почками возобновления, находящимися на уровне поверхности почвы, защищенными мертвым покровом или верхним слоем самой почвы; криптофиты - растения, у которых почки возобновления находятся под землей или под водой; терофиты - однолетники, переносящие неблагоприятное время года в виде семян. Система К. Раункиера универсальна, она охватывает все жизненные формы, дислоцированные в разных эколо-географических районах земного шара. Эта система не только классифицирует жизненные формы покрытосеменных, но демонстрирует их эволюцию - от деревьев к травам. Состав жизненных форм может быть показателем (индикатором) климатических условий страны. К. Раункиер разработал и применил метод статистического анализа при изучении закономерностей распространения и распределения жизненных форм. Влияние внешней среды на структуру вегетативных органов. Экологические группы растений. Растения, приспособившиеся к какому-то одному фактору внешней среды, имеющему важное формообразовательное значение, объединяют в экологические группы. По отношению к влаге выделяют следующие экологические группы. Ксерофиты - растения, живущие в условиях значительного недостатка влаги (постоянного или временного) (виды родов: маслина, сорго, полынь и др.). Для ксерофитов характерны различные приспособления, уменьшающие транспирацию: толстые кутикула и наружная стенка эпидермы (виды рода сахарный тростник; семейства пальмовые), густое опушение, расположение устьичных аппаратов в углублениях (виды рода олеандр), мелкоклеточность тканей и одревеснение клеточных стенок, свертывание листьев (виды рода ковыль), редукция листьев (виды родов эфедра, саксаул) и др. Некоторые растения (суккуленты) накапливают в корнях и листьях большой запас воды 56 (виды родов молодило, алоэ; сем. кактусовые), образуют мощную корневую систему (виды рода бодяг). Мезофиты - растения, живущие в условиях достаточно умеренного увлажнения. Структура их корней, стеблей, листьев служит эталоном. Гидрофиты - растения, обитающие в водной среде (виды родов: стрелолист, сусак, кувшинка и др.). Одни из них полностью погружены в воду (виды рода роголистник), другие - частично. Они либо прикрепляются ко дну водоема (виды родов кувшинка, кубышка), либо свободно плавают в воде (виды родов: сальвиния, ряска). Для них характерны большая поверх¬ность органов, тонкие листья, отсутствие кутикулы, наличие больших межклетников, заполненных воздухом, слаборазвитые сосуды. Гигрофиты - растения, живущие при повышенной влажности почвы и воздуха (виды родов калужница, белокрыльник, рогоз, манник, рис и др.). У некоторых из них отсутствуют тонкостенные, приспособления кутикула для тонкая, уменьшения устьичные транспирации: аппараты клетки расположены эпидермы вровень с поверхностью листа, клетки крупные, лежат рыхло. У других имеются приспособления для быстрого скатывания воды с поверхности листа: гладкая кутикула, капельные острия (виды рода фикус). По отношению к свету выделяют следующие экологические группы. Тенелюбивые растения (сциофиты) живут в условиях постоянного затенения. Это преимущественно травы нижнего яруса леса (виды родов кислица, вороний глаз). Они имеют тонкие вытянутые стебли с неразвитыми механическими тканями, содержат мало хлоропластов (этиолированные). Листья тонкие, толщина слоя палисадной паренхимы относительно небольшая, клетки крупные, с крупными хлоропластами, содержащими много хлорофилла. Светолюбивые растения (гелиофиты) не выносят затенения, растут на открытых местах (виды родов картофель, томаты, свекла и др.). Листья у них более толстые, слой палисадной паренхимы мощный, мезофилл пронизан густой сетью проводящих пучков, число устьичных аппаратов на единицу поверхности листа большое. Клетки палисадной паренхимы более мелкие, содержат бедные хлорофиллом хлоропласты. Однако на единицу объема клетки приходится больше хлоропластов, чем у тенелюбивых растений. Теневыносливые растения лучше растут на открытых местах, однако способны выносить и затенение. В пределах кроны одного растения различают световые и теневые листья. Многие признаки световых листьев совпадают с признаками ксерофитов, так как яркому 57 освещению всегда сопутствуют нагревание и повышенная транспирация, а признаки теневых листьев часто совпадают с признаками гигрофитов. В дождевых тропических лесах в условиях глубокого затенения образовались особые жизненные формы растений, выносящие основную массу побегов в верхние ярусы к свету: лианы и эпифиты. Лианы - это быстрорастущие лазящие растения, которые, используя твердую опору, продвигаются к свету (виды родов плющ, лимонник, актинидия, виноград и др.). Эпифиты - травянистые растения, поселяющиеся на стволах других растений высоко над землей, питаются самостоятельно - не паразитируют (виды сем. орхидные и др.). По отношению к почве выделяют следующие экологические группы. Олиготрофы - растения, живущие на очень бедных минеральными солями субстратах (на верховых болотах, песках и др.). Им свойственны ксероморфные признаки, которые сочетаются с наличием больших межклетников, способствующих аэрации и восполнению недостатка кислорода (виды родов белоус, вереск, клюква, багульник и др.). Галофиты - растения, живущие на сильно засоленных субстратах (виды родов солерос, сарсазан, сведа и др.). Они также имеют черты ксероморфизма, хотя избыток солей часто сочетается с избытком влаги (морские побережья, мокрые солонцы). Клетки галофитов имеют высокий осмотический потенциал. Это необходимое условие поглощения воды. Среди них часто встречают суккуленты. У других имеются мелкие жесткие листья, покрытые железками, выделяющими соли, которые накапливаются на поверхности листа в виде кристалликов. Таким образом, в результате взаимодействия растений с окружающей средой и естественного отбора у них возникают новые физиологические свойства и признаки структуры, наиболее целесообразные в условиях конкретной экологической обстановки. Виды и внутривидовые отдельности, обладающие такими целесообразными признаками, оказываются наиболее приспособленными (адаптированными) к данным экологическим условиям. Такие виды и их комплексы - великолепные индикаторы (показатели) тех компонентов среды, к которым они адаптированы. Размножение растений. Одно из обязательных свойств живого организма - воспроизводство потомства, то есть способность одной особи дать начало целой серии себе подобных дочерних особей или, по крайней мере, одной. Существует два принципиально различных способа размножения: половое и бесполое. Бесполое размножение. 58 Различают два вида бесполого размножения: вегетативное и собственно бесполое. У некоторых низших растений четкой границы между этими видами размножения нет. Вегетативное размножение. Осуществляется частями таллома, корня, стебля, листа. Оно основано на способности растений к регенерации - восстановлению целого организма из его части. У одноклеточных растений (водоросли) вегетативное размножение происходит путем деления клетки, у колониальных и низших многоклеточных - путем расчленения таллома на части. У высших растений вегетативное размножение осуществляется частями корня, стебля, листа или их видоизменениями - корневищами, клубнями, луковицами, выводковыми почками. На основе естественного вегетативного размножения в практике сельского хозяйства разработаны разнообразные способы искусственного вегетативного размножения овощных, плодовых и декоративных растений. Для этого наиболее часто используют: клубни (картофель, батат, георгины), корневища (ирис, флокс), усы (земляника), луковицы (лук, тюльпан), корневые отпрыски (малина, вишня). Многие возделываемые растения размножают черенками, отводками, а также путем прививок. Размножение черенками. Черенок - это отрезанный от материнского растения участок побега, корня, листа. При размножении черенками необходимо помнить о законе полярности. Побеговые черенки могут быть зимними - без листьев (но с почками), длиной 20 - 30 см, в возрасте 1 - 3 года, и летними - с листьями, длиной 3 - 4 см, с побегов текущего года. Листовой черенок состоит из листовой пластинки и черешка. Придаточные корни возникают чаще на морфологически нижней стороне листа в местах разветвления крупных жилок, почки, а затем и побеги - на верхней стороне. Размножать листовыми черенками можно лишь немногие растения (лилия, бегония, алоэ и др.). Корневыми черенками размножают виды растений, корни которых легко образуют придаточные почки (малина, вишня, слива, финиковая пальма, роза, флокс и др.). Размножение прививкой. Прививка - это срастание срезанных почек или стеблевых черенков одного растения (размножаемого) с другим (укорененным). В этом случае размножаемое растение называют привоем, а растение, к которому прививают, - подвоем. Известно около 100 разнообразных способов прививок, но их можно свести к трем основным типам: прививка сближением, или аблактировка, - привой не отчленяется от материнского растения до срастания с подвоем; 59 прививка черенком, или копулировка, - на однолетних черенках привоя с 2 - 3 почками и на стебле подвоя делают косые срезы и прикладывают их друг к другу, при этом стебли привоя и подвоя подбирают одинаковой толщины, чтобы их камбиальные слои совпали. Если диаметры привоя и подвоя не совпадают, то прививку делают врасщеп, под кору, вприклад или другими способами; прививка глазком, или окулировка, - привоем служит спящая или растущая почка, отделенная от срединной части побега размножаемого растения, которую вставляют под кору подвоя. Лучше всего срастание происходит, если привой и подвой принадлежат к одному виду или близким видам одного рода, если же они относятся к разным родам, срастание затруднено. Собственно бесполое размножение. Осуществляется специализированными клетками - спорами или зооспорами. Споры имеют твердую стенку и распространяются ветром, зооспоры не имеют твердой стенки и передвигаются при помощи жгутиков. Они образуются внутри органов бесполого размножения - спорангиев или зооспорангиев. У низших растений это одноклеточный орган, у высших - многоклеточный. Разнообразие спор велико. По происхождению и назначению их подразделяют на .две группы. В первую группу входят споры (зооспоры), которые образуются путем митотического деления и способны непосредственно воспроизводить новую особь, сходную с материнской. При созревании они покидают материнский организм и обычно служат для размножения и распространения. Размножение такими спорами свойственно низшим растениям. Во вторую группу входят споры, которые не могут воспроизвести материнскую особь. Они гаплоидны (n), так как образуются в результате мейоза. Такие споры бывают у высших растений, а также у некоторых водорослей. У последних из споры вначале вырастает заросток (проталлий, протонема), на котором происходит половое размножение. Половое размножение. Суть полового размножения или воспроизведения состоит в формировании растением специализированных клеток - гамет (n), в попарном их слиянии (оплодотворении) и образовании зиготы (2n), из которой вырастает новое растение. Гамета - это половая клетка, ядро которой содержит гаплоидное число хромосом. Сливающиеся гаметы могут различаться между собой по структуре (форме, величине, 60 подвижности), но обязательно физиологически, то есть по полу и по наследственности. В результате слияния гамет в новом организме объединяются материнская и отцовская наследственность. Если сливающиеся гаметы одинаковы по форме, величине и подвижности, их называют изогаметами, а половой процесс - изогамным. Если гаметы одинаковы по форме, но женская гамета более крупная и менее подвижная, чем мужская, их называют гетерогаметами, а половой процесс - гетерогамным. Половой процесс называют оогамным, когда женская гамета крупная, шаровидная, неподвижная (яйцеклетка), а мужская очень маленькая и подвижная (сперматозоид) или неподвижная (спермий). Специализация гамет служит показателем уровня эволюции. Наиболее примитивной формой полового процесса является изогамия, Гаметы формируются в специальных органах, называемых гаметангиями: мужские - в антеридиях, женские - у низших растений в оогониях, а у высших - в архегониях. У низших растений гаметангии, как и спорангии, одноклеточные, а у высших многоклеточные. У некоторых групп водорослей органов полового размножения нет, хотя половой процесс осуществляется регулярно. Зигота у них образуется вследствие слияния неспециализированных клеток или члеников. Такой половой процесс называют конъюгацией. Разновидность конъюгации - гологамия (хологамия), когда происходит слияние двух одноклеточных вегетативных особей без предварительного образования гамет. Чередование ядерных фаз в жизненном цикле. В индивидуальном жизненном цикле высших и некоторых низших растений наблюдают последовательность и взаимную связь бесполого и полового размножения. На основе анализа ядерных фаз при размножении установлено, что спорофит, то есть особь, которая производит споры второй группы и их вместилища (спорангии), состоит из клеток, имеющих диплоидный набор хромосом (2n). При образовании спор происходит мейоз, так что они всегда гаплоидны (n). Из споры образуется гаметофит с органами полового размножения, продуцирующими половые клетки - гаметы. Все клетки гаметофита, в том числе и гаметы, гаплоидны (n). В результате полового процесса (оплодотворения, слияния гамет) возникает зигота (2 n), а из зиготы вновь образуется спорофит (2 n). Систематика - это наука, которая занимается объединением растений, населяющих нашу планету, в родственные группы и расположением этих групп в таком порядке, который бы отражал эволюцию царства растений, длившуюся сотни миллионов лет. Вид. Биологический вид более или менее динамичен во времени и подвижен в пространстве. Его изучение сопряжено с большими трудностями, так как вид чаще всего 61 представлен множеством рас и популяций, как-то обособленных в природе и различающихся между собой. При естественноисторическом подходе определение вида представляет чрезвычайные трудности. Во всяком случае, из многих десятков определений нет ни одного, исчерпывающего суть вида и общепризнанного. Одно из наиболее общеизвестных и удовлетворительных определений вида принадлежит В. Л. Комарову: «Вид есть совокупность поколений, происходящих от общего предка и под влиянием среды и борьбы за существование обособленных отбором от остального мира живых существ; вместе с тем вид есть определенный этап в процессе эволюции». При формально-логическом подходе определение вида не представляет трудностей: вид совокупность тождественных особей, практически неразличимых. Конечно, тождество в данном случае понимают условно и относительно, как тождество, например, растений, выросших из зерновок одного колоса или одного растения пшеницы. Но для общей ориентации в природе, в созданных человеком культурных ценозах, в практической работе агронома и ботаника по определению растений мы пользуемся именно формальнологическим понятием. По структуре виды бывают неравноценными. Различают виды молодые (зачинающиеся), угасающие (вымирающие) и процветающие. Вымирающие виды с течением времени явно уменьшаются по числу особей, что обычно сопровождается и уменьшением площади их распространения. У них отсутствует внутривидовой полиморфизм, их легко различают, поскольку между смежными родственными видами большой морфолого-физиологический разрыв, нет переходных форм. Это, как правило, древние виды. Молодые виды явно экспансируют (расселяются), их численность резко увеличивается. Это полиморфные виды, в составе которых выделяют множество внутривидовых комплексов (подвидов, разновидностей, форм). Изобилие внутривидовых единиц свидетельствует о формообразовательном процессе, динамике вида. В настоящее время внутривидовая дифференциация видов достигает 5 - 7-й степени. Приведем наиболее современные модели: по Н.П. Дубинину - полувид, подвид, раса, микротопографическая раса, линия; по К.М. Завадскому - полувид, подвид, местная популяция, экоэлемент, изореагент (морфобиологическая группа, биотип); по Н.И. Вавилову - для возделываемых растений применяют целую серию других внутривидовых таксонов, например группа разновидностей (пролес), сорт. Все еще нет достаточно определенных согласованных данных о численности видов растений на планете. Предположительно общее число ныне живущих видов растений около 500 тыс. Число это весьма приблизительно. Даже для одной из наиболее изученных 62 групп растений - покрытосеменных - количество видов, приводимое разными учеными, неодинаково: от 175 - 200 тыс. до 300 тыс. и более. Бинарная номенклатура. Виды растений обозначают двумя латинскими словами: первое означает род, к которому относится данный вид, второе - видовой эпитет. После второго слова ставят фамилию ученого (сокращенно или полностью), открывшего либо описавшего этот вид и давшего ему название. Например, научное название пшеницы твердой - Triticum durum L. Бинарная номенклатура была введена шведским ученым К. Линнеем (1707 - 1778). Таксоны (систематические единицы). Классификацию растений на отдельные систематические группы осуществляют на основе общепринятых единиц, называемых таксонами: вид (species) - совокупность тождественных в морфологическом отношении особей; род (genus) - совокупность близкородственных видов; семейство (familia) - совокупность родственных родов; порядок, (ordo) - совокупность родственных семейств; класс (classis) - совокупность родственных порядков; отдел (divisio) - совокупность родственных классов. Кроме основных таксонов, существуют таксоны промежуточные, вспомогательные: подрод, подсемейство и т. д. Внутри вида выделяют подвиды (subspecies) и разновидности (varietas), а применительно к культурным растениям - сорта (cultivar). Сорт - это результат селекционной деятельности человека, он характеризуется определенными хозяйственными качествами и передачей их по наследству. Вне рамок официальных таксонов царство растений делят на низшие и высшие; несколько родственных отделов объединяют в подцарство. 63