- Природа против рака
advertisement

Medical Hypothesis (1996) 46: 551-555
Стресс-индуцированное подавление клеточных иммунных реакций.
Роль нейроэндокринного контроля иммунной системы.
A. Hassig, Liang Wen-Xi, K. StampfliI
Реферат
Задача иммунной системы состоит в том, чтобы поддержать
наследственно определенную индивидуальность организма. Эта задача
охватывает две области: во-первых, устранение экзогенных ''не своих''
(''not-self'') структур и во-вторых, уничтожение эндогенных ''своих
изменившихся'' (''altered-self'') структур, образование которых происходит
в больших количествах с постоянным реструктурированием клеточных
элементов организма [1]. Согласно самым последним данным,
устранение экзогенных ''не своих'' структур - первичная задача
гуморальных иммунных реакций, связанных с B-клетками. Уничтожение
эндогенных ''своих изменившихся'' структур, с другой стороны, является
первичной задачей клеточных иммунных реакций, связанных с
цитотоксическими T-клетками и естественными киллерами.
Иммунологическое здоровье зависит от гармоничного соотношения
между гуморальными и клеточными иммунными реакциями.
Иммунная компетентность как состояние равновесия между
гуморальным и клеточным иммунитетом
В поиске документированных свидетельств дуальности этих реакций
одним из первых является исследования Glaser и Kiecolt-Glaser [2]. В
своих исследованиях они показали, что реакции психического стресса
увеличивают титр гуморальных антител против вирусов типа EBV, CMV и
HSV-1, посредством чего они в то же самое время вредят клеточным
иммунным реакциям. Они объяснили эти результаты действием
латентных вирусов из-за ухудшения клеточных защитных механизмов и
вторичного возбуждения производства гуморальных антител. Такая
интерпретация была поддержана наблюдением, что при реакциях на
стресс титр антител к полиомиелиту не изменялся. Вирусы
полиомиелита полностью уничтожаются иммунной системой после их
распространения в организме, что не имеет место с упомянутыми
вирусами герпеса.
Противоположное поведение гуморальных и клеточные иммунных
реакций, наблюдаемых в этих исследованиях получило поддержку в
исследовании Mosmann и Coffmann функционально противоположных
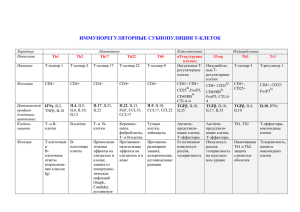
профилях цитокинов CD4 лимфоцитов [3]. Они показали, что среди CD4
клеток могут быть идентифицированы две группы клеток , которые
известны как Th1 и Th2 клетки. Th1 клетки секретируют главным образом
IL-2, IL-12 и IFN-Y, которые стимулируют клеточные реакции. Th2 клетки,
с другой стороны, производят главным образом IL-4, IL-6 и IL-10 и их
посредством стимулируют гуморальные реакции. В последние годы к
этим исследованим проявляется большой интерес и эти результаты
сегодня неоднократно подтверждены.
Нейроэндокринный контроль баланса между гуморальным и
клеточным иммунитетом
В настоящее время среди многих других изучается вопрос, как баланс
производства Th1/Th2 цитокинов CD4 лимфоцитами может быть связан с
нейроэндокринными механизмами контроля иммунной системы в
реакциях стресса. Сущность концепции стресса, выработанной Селье в
1936 г. заключается в том, что организм отвечает на предъявленные ему
соматические и психические требования одинаковым способом. Здесь
центральную роль играет активация оси гипоталамус-гипофизнадпочечники. Увеличение производства катехоламинов и
глюкокортикоидов ведет к метаболизму быстро доступных источников
энергии, особенно глюкозы, ввиду увеличения мышечной активности в
ситуации ''дерись или беги''. Физиологическая функция
глюкокортикоидов, очевидно, состоит в лимитировании активности
эндогенных медиаторов воспаления в угрожающих жизни ситуациях [4].
Увеличение производства кортизола ведет к подавлению клеточных
иммунных реакций, связанных с T-клетками и, таким образом, к
увеличению восприимчивости к условно-патогенным инфекциям.
В отношении соотношения Th1 и Th2 профилей цитокинов CD4
лимфоцитов в нейроэндокринных процессах при реакции на стресс
должно быть заявлено, что еще до того, как Th1 и Th2 профили
цитокинов CD4 лимфоцитов были описаны, уже было известно, что
глюкокортикоиды ингибируют производство IL-2 и IFN-gamma [4]. Мы
должны быть благодарны группе Daynesfor за важный шаг, который они
сделали в разъяснении механизма формирования этих лимфоцитарных
профилей цитокинов [5][6]. Этих исследования показали, что
производство цитокинов активизированными лимфоцитами является
управляемым процессом. Митоген- или антиген- стимулируемые
лимфоциты из лимфоидных органов в области слизистых, производит
главным образом IL-4. Лимфоциты из других органов производят
главным образом IL-2. Решающим для управления типом
лимфоцитарного производства цитокинов - вероятно является
производство стероидных гормонов. В этом процессе, DHEA
произведенный в корой надпочечников играет центральную роль, как
антагонист кортизола. DHEA - гормон коры надпочечников, который из
всех стероидных гормонов содержится в крови в самой высокой
концентрации. Это - сульфатированная инактивная форма (DHEAS).
DHEA десульфатируется стероидной сульфатазой и таким образом
преобразуется в активную форму. В лимфоцитах активный DHEA
ответствен за увеличение производства IL-2 и IFN-gamma, но не IL-4. Эти
результаты показывают, что различное содержание стероидной
сульфатазы в различных тканях имеет большую важность в
преобразовании про-гормона DHEA в активный DHEA при производстве
Th1 и Th2 лимфоцитов. В лимфатической ткани, макрофаги единственные клетки, которые содержат DHEA. Кроме того, DHEA
служит источником производства андрогенов и, в меньшей степени,
эстрогенов.
Все это говорит о том, что баланс Th1/Th2 на участке активации
существенно зависит от баланса стероидных гормонов, кортизола и
DHEA, причем DHEA является антагонистом системного действия
кортизола. При реакциях на стресс изменяется отношение Th1 профиля
CD4 лимфоцитов к Th2 профилю, очевидно вследствие того, что
симпатикотоническое адренергическое - кортикоидное смещение
метаболизма смещает баланс кортизол / DHEA в пользу кортизола, в
результате чего концентрация профилей цитокинов CD4 лимфоцитов
смещается больше в область слизистых. Согласно этой гипотезе, в
ситуациях стресса гуморальные иммунные реакции, которые направлены
против экзогенных ''не своих'' структур, усиливаются, в то время как
клеточные иммунные реакции, направленые против эндогенных ''своих
измененных'' структур, ослабляется.
В отношении прямого эффекта гиперкортицизма на лимфоциты, должно
быть отмечено, что CD4+/CD8+ тимоциты являются наиболее
чуствительными к кортизолу элементами лимфатической ткани.
Увеличение апоптоза действует так, чтобы число иммунологических
клеток-предшественников уменьшилось. Периферические CD4 +/CD8- и
CD4-/CD8 + лимфоциты являются относительно кортизол-резистентными
[7][8]. Уменьшение числа CD4 лимфоцитов в крови при фактически
постоянном числе CD8 лимфоцитов, что является характерным для
реакций стресса, происходит прежде всего благодаря секвестрации CD4
клеток в костном мозге [9].
В отношении эффекта дефицита IL-2 и IFN-gamma, может быть
рассмотретрена следующая гипотеза. IL-2 производится
активизированными T клетками. Это активизирует клональную экспансию
T- и B-клеток а также естественных киллеров. В случае дефицита IL-2
функция идиопатической сети повреждается, поскольку ее
поликлональность меняется на олигоклональность. IFN-gamma
производится активизированными T-клетками. Его главный эффект активация макрофагов в отношении их фагоцитарных функций, особенно
деградации или внутриклеточной иммобилизации бактерий, вирусов и
опухолевых клеток.
Прямая и косвенная активация нейроэндокринной оси
Индуцированное стрессом подавление клеточных иммунных реакций
может быть разделено на две патогенетических группы. Одна группа
включает состояния с прямой активацией нейроэндокринной оси из-за
увеличения производства кортикотропинвысвобождающего гормона
(CRH) гипоталамусом. Психические стрессы и воздействие токсических
веществ принадлежат этой группе. Вторая группа включает состояния с
косвенной активацией нейроэндокринной оси из-за реакций на
повреждения, инфекционные болезни и несбалансированную диету.
Последнее десятилетие стало ''десятилетием психонейроиммунологии''.
Разработанная Робертом Адером область психического влияния на
иммунные реакций превратилась в отдельную, независимую науку
[10][11].
Из токсических стрессовых реакций, наиболее хорошо изучен
иммуносупрессивный эффект опиатов. В экспериментах на мышах было
показано, что хроническое ведение морфия причиняет серьезную
инволюцию тимуса, характеризующиеся увеличенным апоптозом,
особенно CD4+/CD8+ тимоцитов. Это следствие того, что морфий
увеличивает производство CRH гипоталамусом, в результате чего
производство АКТГ гипофизом и кортизола в надпочечниками
активизировано [13][14].
Однако, наиболее важным является косвенная активация
нейроэндокринной оси из-за увеличения производства CRH
гипоталамусом. В результате этого постоянно увеличивается
производство IL-1 и TNF-alpha макрофагами, активизированных
воспалением [15]. Также увеличено производство O2 радикалов [16].
Производство этих радикалов в пределах фагоцитов необходимо для
эффективного разрушения структур, пришедших извне. В
физиологических состояниях, антиокислительные ферменты и
низкомолекулярные антиоксиданты гарантируют, что освобождение
O2 радикалов, не произведет никакого повреждения. Чрезмерное
производство O2 радикалов и/или нарушение антиокислительной защиты
ведет к индуцированному радикалами повреждению нормальных
структур клеток, особенно ДНК, белков и липидов. Наиболее важными
радикалами являются высоко активные гидроксильные радикалы (HO),
чье производство катализирует Fe2+. Накопление радикалов, таким
образом, увеличивает перегрузку железом макрофагов [17]. При любой
воспалительной реакции с острой стадией значительная часть
серологического железа перемещается в макрофаги, где связывается в
ферритин. По этим причинам важно в стрессовых ситуациях,
противодействовать как перегрузке железом так и истощению
антиоксидантной защиты организма.
Другая большая группа индуцированных стрессом супрессии клеточного
иммунитета представлена так называемым белковым дефицитом в
диете у детей с квашиоркор. Beisel описал эту группу болезней как
синдром приобретенного пищевого иммунодефицита (NAIDS 'nutritionally acquired immunodeficiency syndrome). Он привлек внимание к
тому факту, что в мире приблизительно 40,000 детей в возрасте до пяти
лет умирают от этой болезни каждый день [18]. У этих детей
обнаруживают атрофию тимуса и серьезный дефицит T-лимфоцитов в
лимфатических узлах и селезенке. Как и при СПИДЕ, они умирают от
инфекционных болезней, вызванных внутриклеточными патогенами,
такими как Pneumocystis carinii, Herpes simplex или милиарного
туберкулеза [19].
Как и при СПИДЕ, отказ клеточных иммунных реакций - центральный
фактор патологического процесса. В отношении важности вируса
иммунодефицита человека (ВИЧ) в развитии СПИДА, теперь известно,
что пациенты со здоровой иммунной системой с Th1 профилем
цитокинов CD4 лимфоцитов, как правило способны бороться с ВИЧ
инфекцией с помощью клеточных иммунных реакций, без производства
антител к ВИЧ. Обнаружение антител при ВИЧ инфекции - признак
ослабления клеточных иммунных реакций. Возможно это связано со
стрессовыми ситуациями, существующими в группах риска для СПИДА, а
именно гомосексуалистов, наркоманов, при гемофилии и у получателей
донорской крови [20].
Кроме того, необходимо помнить, что синдром сепсиса, который часто
наблюдается при серьезных ожогах и хирургических вмешательствах,
является чрезмерным, постоянным процессом воспаления во всем
организме. В этих случаях массированный выпуск медиаторов
воспаления блокирует антивоспалительное действие глюкокортикоидов
и нарушает работу гуморальной и клеточной иммунной защиты и обычно
безопасные микробы распространяются по всему организму. Чрезмерная
активность макробактериофагов играет центральную роль в этом
процессе.
Итак, Th2 профиль CD4 лимфоцитов соответствует ситуации стресса с
избытком кортизола и дефицитом DHEA. Это вызывает дефицит IL-2, IL12 и IFN-Y в организме и одновременно избыток IL-4, IL-6 и IL-10.
Возможности для диагностирования индуцированной стрессом
иммуносупрессии
Th2 профиль CD4 лимфоцитов может быть определен непосредственно,
если ухудшение клеточного иммунитета проявляется посредством
кожной анергии против инфекционных антигенов. Гиперчувствительность
замедленного типа требует неповрежденного гуморального и клеточного
иммунитета. Косвенные признаки Th2 профиля - увеличение уроовня
белков острой фазы, особенно C-реактивного белка, связано с
увеличением производства IL-6. Самый простой признак для присутствия
реакции в острой стадии - ускорение реакции оседания эритроцитов.
Возможности для предотвращения и лечения индуцированной
стрессом иммуносупрессии
Как увеличение Th2 профиля, с нейроэндокринным подавлением
клеточных иммунных реакций, быть приведено в состояние
гомеостатического равновесия между клеточным и гуморальным
иммунитетом?
Самым важным в этом процессе является восстановление процесса
нейроэндокринной супрессии и адренергического регулирования
метаболизма. Использование цитокинов IL-2, IL-12 и IFN-gamma дает, в
лучшем случае, кратковременный симптоматический эффект. При
прямой активацией HPA оси, устранение ситуаций ментального
конфликта и зависимости от лекарств, кажется, нам решающим. В
случае косвенной активации HPA оси посредством инфекционных и
пищевых механизмов, решающим фактором является устранение
активации фагоцитов и непрерывного производства свободных
радикалов и медиаторов воспаления, через адекватную,
сбалансированную диету.
В отношении фармакологического лечения этих состояний, наиболее
интересная возможность в настоящее время - использование двух
эндогенных гормонов, а именно DHEA и мелатонина [22]-[24]. Поскольку
DHEA является антагонистом кортизола, он в настоящее время предмет обширных фармакологических и клинических исследований.
Мелатонин - наиболее мощный антиоксидант из известных в настоящее
время, и в этом отношении он во много раз мощнее чем витамины E, C и
бета-каротин. Этот гормон также является в настоящее время
предметом серьезного научного интереса.
По нашему мнению, в долгосрочной перспективе решающим для
предотвращения постоянного ухудшения клеточного иммунитета с его
предполагаемой связью с болезнями Западной цивилизации,
артеросклерозом и раком будет являться регулирование жизни и диеты.
Внимание также должно быть состредоточено на балансе между работой
и отдыхом. В отношении диеты, наиболее важным аспектом является
сбалансированное потребление питательных веществ (белки, жиры,
углеводы) и микроэлементов (витамины, минералы). Однако, по
современным представлениям, адекватное потребление растительных
антиоксидантов, а именно полифенолов, флавонидов и танинов, также
является необходимым [25].
В заключение, мы хотели бы заявить следующее: стрессовые ситуации
увеличивают вероятность выживания в ситуациях ''дерись или беги''
(''fight or flight''). Организм толерантен к этому состоянию только в
течение короткого промежутка времени. Если это состояние
сохраняется, оно ведет к разнообразным повреждениям, наиболее
серьезным из которых является ухудшение функции тимус-зависимых Tклеток. По этой причине, при профилактике и лечении болезней
Западной цивилизации, особенно у пожилых людей, необходимо
обратить особое внимание на функцию тимуса [26][27].
Мы благодарим Hans Eggenberger Foundation (Цюрих) за поддержку
этого исследования.
Литература
1. H.U. Lutz: Erythrocyte Clearance Blood Cell Biochemistry. Vol. I. Erythroid
Cells. Chapter 4. Ed. J.R. Harris, Plenum Press, New York, London
2. R. Glaser, J.K. Kiecolt-Glaser: Stress associated Immuno Modulation and
its Implication for Reactivation of latent Herpesviruses. Herpes Virus
Infections R. Glaser, J.E. Jones (Eds) Marcel Dekker Inc. N.Y. (1994)
3. T.R. Mosmann, R.L. Coffman: Thl and Th2 Cells: Different Patterns of
Lymphokine Secretion Lead to Different Functional Properties. Ann.Rev.
Immunol. 7, 145 (1989)
4. A. Munck, P.M. Guyre, N.J. Holbrook: Physiological Funktions of
Glucocorticoids in Stress and their relation to Pharmacological Actions.
Endocrine Reviews 5, 25 (1984)
5. R.A. Daynes, B.A. Araneo et al.: Regulation of Murine Lymphokine
production in vivo III. The Lymphoid Tissue Microenvirement exerts
Regulatory Influence over T Helper Cell functions. J.exp.Med. 171,979 (1990)
6. J.D. Hennebold, R.A. Daynes: Regulation of Macrophage
Dehydroepiandrosteron; Sulfate Metabolism by Inflammatory Cytokins.
Endocrinology 135, 67 (1994)
7. J.J. Cohen: Glucocorticoid-induced apoptosis in the thymus. Sem.
Immunol. 4, 363 (1982)
8. J. Gruber, R. Sgonc et al.: Thymocyte apoptosis induced by elevated
endogenous corticosterone levels. Europ.J. Immunol.24, 1115 (1994)
9. B.F. Haynes, A.S. Fauci: The differential effect of in vivo hydrocortisone on
the kinetics of subpopulations of human peripheral blood thymus-derived
lymphocytes. J .cl in. Invest. 61 703 (1978)
10. R. Ader, D.L. Felten, N. Cohen: Psychoneuroimmunology 2nd ed.
Academic Press San Diego (1991)
11. B.E. Leonard, K. Miller (Eds): Stress, the Immune System and Psychiatry.
J. Wiley, Chichester (1994)
13. H.U. Bryant, E.W. Berton et al.: Role of Adrenal Cortisoh Activation in the
Immunosuppressive Effects of Chronic Morphine Treatment Endocrinology
128, 3253 (1991)
14. B.A. Fuchs, S.B. Pruett: Morphine produces Apoptosis in Murine
Xhymocytes in Vivo but not in Vitro. J. Pharmacol.-expl.Therapeutics 266, 417
(1993)
15. H. Besedovsky, A.E. del Rey, E. Sorkin, C.A. Dinarello: Immunoregulatory
feed back between interleukin-1 and glucocorticoid hormones. Science, 233,
652 (1986)
16. J.M.C. Gutteridge, B. Halliwell: Antioxidants in Nutrition, Health and
Disease. Oxford University Pres (1994)
17. E.D. Weinberg: The Iron-with-holding-Defence-System. Ann.Soc.
Microbiol. News 59, 559 (1993)
18. W.R.Beilel: History of Nutritional Immuology: Introduction an Overview. J.
Nutr. 122, 591 (1992)
19. D.R. Purtilo, D.H. Connor: Fatal infections in protein-calorie malnourished
children with thymolymphatic atrophy. Archives of Diseases in Childhood 50,
149 (1975)
20. A. Hassig, Liang Wen-Xi, K. Stampfli: Can we find a solution of the
HIV/AIDS Controversy, Medital Hypothesis, in print
21. D. Nast-Kolb, M. Jochum et al.: Die klinische Wertigkeit biochemischer
Faktoren beim Polytrauma. Hefte Unfaliheilkd. (Suppl) 215 (1991)
22. A.J. Morales, J.J. Nolan, J.C. Nelson, S.S.C. Yen: Effects of replacement
dose of dehydroepiandrosterone in men and women of advancing age. J. Clin.
Endocrinol. Metab. 78, 1360-1367 (1994)
23. B. Araneo, R. Daynes: Dehydroepiandrosterone Functions as More Than
an Antiglucocorticoid in Preserving Immuncompetence after Thermal Injury.
Endocrinology 136, 393-401 (1995)